
In vivo cell penetration and intracellular transport of anti-Sm and anti-La autoantibodies
9
International Immunology, Vol. 12, No. 4, pp. 415–423 © 2000 The Japanese Society for Immunology In vivo cell penetration and intracellular transport of anti-Sm and anti-La autoantibodies Sophie X. Deng, Elaine Hanson and In ˜ aki Sanz Department of Immunology and Microbiology, and Department of Medicine, University of Rochester Medical Center, 601 Elmwood Avenue, Box 695, Rochester, NY 14642, USA Keywords: autoantibody, autoimmune disease, intracellular antibodies, ribonucleoproteins, systemic lupus erythematosus Abstract Anti-nuclear autoantibodies (ANA) are the hallmark of systemic autoimmune diseases. Yet, the in vivo function of ANA remains controversial to a large extent due to the intracellular nature of their antigenic targets. It has been reported that a subset of autoantibodies can penetrate live cells and translocate into the subcellular compartments containing the corresponding antigens. The studies presented herein show that murine anti-Sm and anti-La monoclonal autoantibodies can also enter a variety of cell types from different animal species and that the cell penetration activity is not isotype- restricted. Interestingly, only mAb with cross-reactivity against double-stranded DNA did enter cells. Both these autoantibodies rapidly accumulate in the nucleus of viable cells but display different penetration kinetics. In co-localization experiments, monoclonal autoantibodies did not accumulate significantly within endocytic vesicles containing dextran, suggesting that they are internalized by mechanisms distinct from conventional receptor-mediated endocytosis. This report represents the first evidence that anti-La and anti-Sm autoantibodies are capable of entering live cells. Our observations support the notion that the phenomenon of intracellular autoantibodies may have a larger scope than previously reported and are consistent with a potential pathogenic role for ANA. Introduction Anti-nuclear antibodies (ANA) have been used to define sys- temic autoimmune diseases for decades, often with great spe- cificity for the disease in question. As a result, ANA constitute important diagnostic tools for a number of diseases, e.g. sys- temic lupus erythematosus (anti-Sm and anti-double-stranded DNA antibodies), generalized systemic sclerosis (anti-DNA topoisomerase I antibodies) and localized systemic sclerosis (anti-centromere antibodies) (1–3). A cardinal feature of ANA is that their antigenic targets usually carry out critical intracellular functions such as DNA and RNA processing and modification, protein synthesis, and cell cycle regulation (4–6). It is also of great significance that the recog- nized epitopes are highly conserved in evolution and localized in domains of functional importance. As a consequence, autoantibodies have been shown to be capable of inhibiting antigen function both in vitro and in vivo (7). At the same time, the very nature of the autoantigens raises a number of significant questions regarding the ability of autoantibodies to interfere with in vivo functions in a manner relevant to disease pathogen- Correspondence to: I. Sanz Transmitting editor: C. Martinez-A Received 20 May 1999, accepted 30 November 1999 esis. Indeed, given the subcellular localization of the autoantig- ens and the systemic nature of the diseases in question, pathogenic autoantibodies should be able to enter live cells in multiple tissues, reach the appropriate intracellular compart- ment and interfere with cell function. The ability of polyclonal IgG anti-ribonucleoprotien (RNP) autoantibodies isolated from autoimmune patients to pene- trate live human peripheral blood mononuclear cells via Fc receptors was first proposed 20 years ago (8). Ever since, an increasing number of autoantibodies of different antigenic specificity have been added to the list of intracellular antibodies (9–19). Yet, it should be noted that only a subset of anti-DNA antibodies has been characterized in some detail for their ability to translocate inside the cell nucleus (16). It should be noted, however, that other investigators have disputed this phenomenon and, indeed, it has been argued that nuclear localization of autoantibody represented a fixation artifact, with movement of Ig into cells during fixation (20). In order to clarify this issue, we have investigated the scope and
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of In vivo cell penetration and intracellular transport of anti-Sm and anti-La autoantibodies

International Immunology, Vol. 12, No. 4, pp. 415–423 © 2000 The Japanese Society for Immunology
In vivo cell penetration and intracellulartransport of anti-Sm and anti-Laautoantibodies
Sophie X. Deng, Elaine Hanson and Inaki Sanz
Department of Immunology and Microbiology, and Department of Medicine, University of RochesterMedical Center, 601 Elmwood Avenue, Box 695, Rochester, NY 14642, USA
Keywords: autoantibody, autoimmune disease, intracellular antibodies, ribonucleoproteins, systemic lupuserythematosus
Abstract
Anti-nuclear autoantibodies (ANA) are the hallmark of systemic autoimmune diseases. Yet, the in vivofunction of ANA remains controversial to a large extent due to the intracellular nature of theirantigenic targets. It has been reported that a subset of autoantibodies can penetrate live cells andtranslocate into the subcellular compartments containing the corresponding antigens. The studiespresented herein show that murine anti-Sm and anti-La monoclonal autoantibodies can also enter avariety of cell types from different animal species and that the cell penetration activity is not isotype-restricted. Interestingly, only mAb with cross-reactivity against double-stranded DNA did enter cells.Both these autoantibodies rapidly accumulate in the nucleus of viable cells but display differentpenetration kinetics. In co-localization experiments, monoclonal autoantibodies did not accumulatesignificantly within endocytic vesicles containing dextran, suggesting that they are internalized bymechanisms distinct from conventional receptor-mediated endocytosis. This report represents thefirst evidence that anti-La and anti-Sm autoantibodies are capable of entering live cells. Ourobservations support the notion that the phenomenon of intracellular autoantibodies may have alarger scope than previously reported and are consistent with a potential pathogenic role for ANA.
Introduction
Anti-nuclear antibodies (ANA) have been used to define sys-temic autoimmune diseases for decades, often with great spe-cificity for the disease in question. As a result, ANA constituteimportant diagnostic tools for a number of diseases, e.g. sys-temic lupus erythematosus (anti-Sm and anti-double-strandedDNA antibodies), generalized systemic sclerosis (anti-DNAtopoisomerase I antibodies) and localized systemic sclerosis(anti-centromere antibodies) (1–3).
Acardinal feature ofANA is that their antigenic targetsusuallycarry out critical intracellular functions such as DNA and RNAprocessing and modification, protein synthesis, and cell cycleregulation (4–6). It is also of great significance that the recog-nized epitopes are highly conserved in evolution and localizedin domains of functional importance. As a consequence,autoantibodies have been shown to be capable of inhibitingantigen functionboth invitroand in vivo (7).At thesametime, thevery nature of the autoantigens raises a number of significantquestions regarding the ability of autoantibodies to interferewith in vivo functions in a manner relevant to disease pathogen-
Correspondence to: I. Sanz
Transmitting editor: C. Martinez-A Received 20 May 1999, accepted 30 November 1999
esis. Indeed, given the subcellular localization of the autoantig-ens and the systemic nature of the diseases in question,pathogenic autoantibodies should be able to enter live cells inmultiple tissues, reach the appropriate intracellular compart-ment and interfere with cell function.
The ability of polyclonal IgG anti-ribonucleoprotien (RNP)autoantibodies isolated from autoimmune patients to pene-trate live human peripheral blood mononuclear cells via Fcreceptors was first proposed 20 years ago (8). Ever since,an increasing number of autoantibodies of different antigenicspecificity have been added to the list of intracellular antibodies(9–19). Yet, it should be noted that only a subset of anti-DNAantibodieshasbeencharacterized insomedetail for theirabilityto translocate inside the cell nucleus (16).
It should be noted, however, that other investigators havedisputed this phenomenon and, indeed, it has been arguedthat nuclear localization of autoantibody represented a fixationartifact, with movement of Ig into cells during fixation (20). Inorder to clarify this issue, we have investigated the scope and

416 Intracellular autoantibodies
Fig. 1.
Fig. 2
Intracellular autoantibodies 417
nature of intracellular autoantibodies. Here we describe twomurine monoclonal autoantibodies with anti-Sm (IgA) and anti-La (IgG) primary antigenic reactivity respectively, which are
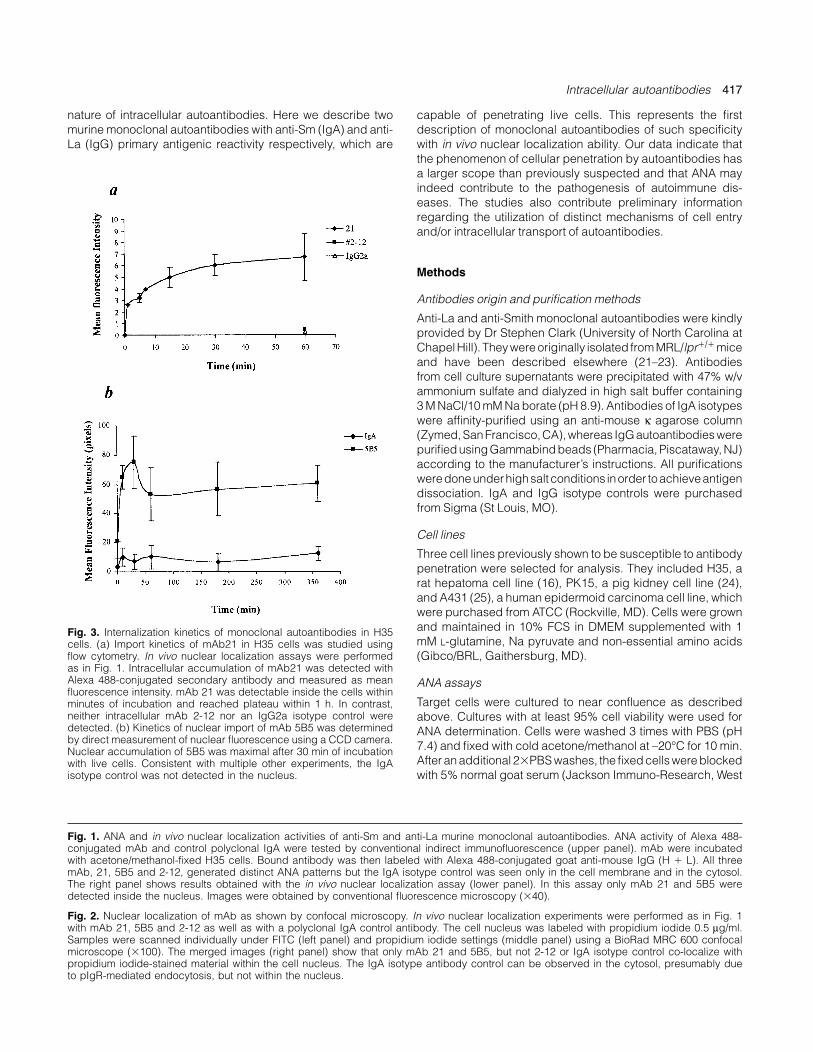
Fig. 3. Internalization kinetics of monoclonal autoantibodies in H35cells. (a) Import kinetics of mAb21 in H35 cells was studied usingflow cytometry. In vivo nuclear localization assays were performedas in Fig. 1. Intracellular accumulation of mAb21 was detected withAlexa 488-conjugated secondary antibody and measured as meanfluorescence intensity. mAb 21 was detectable inside the cells withinminutes of incubation and reached plateau within 1 h. In contrast,neither intracellular mAb 2-12 nor an IgG2a isotype control weredetected. (b) Kinetics of nuclear import of mAb 5B5 was determinedby direct measurement of nuclear fluorescence using a CCD camera.Nuclear accumulation of 5B5 was maximal after 30 min of incubationwith live cells. Consistent with multiple other experiments, the IgAisotype control was not detected in the nucleus.
Fig. 1. ANA and in vivo nuclear localization activities of anti-Sm and anti-La murine monoclonal autoantibodies. ANA activity of Alexa 488-conjugated mAb and control polyclonal IgA were tested by conventional indirect immunofluorescence (upper panel). mAb were incubatedwith acetone/methanol-fixed H35 cells. Bound antibody was then labeled with Alexa 488-conjugated goat anti-mouse IgG (H � L). All threemAb, 21, 5B5 and 2-12, generated distinct ANA patterns but the IgA isotype control was seen only in the cell membrane and in the cytosol.The right panel shows results obtained with the in vivo nuclear localization assay (lower panel). In this assay only mAb 21 and 5B5 weredetected inside the nucleus. Images were obtained by conventional fluorescence microscopy (�40).
Fig. 2. Nuclear localization of mAb as shown by confocal microscopy. In vivo nuclear localization experiments were performed as in Fig. 1with mAb 21, 5B5 and 2-12 as well as with a polyclonal IgA control antibody. The cell nucleus was labeled with propidium iodide 0.5 µg/ml.Samples were scanned individually under FITC (left panel) and propidium iodide settings (middle panel) using a BioRad MRC 600 confocalmicroscope (�100). The merged images (right panel) show that only mAb 21 and 5B5, but not 2-12 or IgA isotype control co-localize withpropidium iodide-stained material within the cell nucleus. The IgA isotype antibody control can be observed in the cytosol, presumably dueto pIgR-mediated endocytosis, but not within the nucleus.
capable of penetrating live cells. This represents the firstdescription of monoclonal autoantibodies of such specificitywith in vivo nuclear localization ability. Our data indicate thatthe phenomenon of cellular penetration by autoantibodies hasa larger scope than previously suspected and that ANA mayindeed contribute to the pathogenesis of autoimmune dis-eases. The studies also contribute preliminary informationregarding the utilization of distinct mechanisms of cell entryand/or intracellular transport of autoantibodies.
Methods
Antibodies origin and purification methods
Anti-La and anti-Smith monoclonal autoantibodies were kindlyprovided by Dr Stephen Clark (University of North Carolina atChapel Hill). They were originally isolated from MRL/lpr�/� miceand have been described elsewhere (21–23). Antibodiesfrom cell culture supernatants were precipitated with 47% w/vammonium sulfate and dialyzed in high salt buffer containing3 M NaCl/10 mM Na borate (pH 8.9). Antibodies of IgA isotypeswere affinity-purified using an anti-mouse κ agarose column(Zymed, SanFrancisco, CA),whereas IgGautoantibodies werepurified using Gammabind beads (Pharmacia, Piscataway, NJ)according to the manufacturer’s instructions. All purificationsweredoneunderhighsalt conditions inorder toachieveantigendissociation. IgA and IgG isotype controls were purchasedfrom Sigma (St Louis, MO).
Cell lines
Three cell lines previously shown to be susceptible to antibodypenetration were selected for analysis. They included H35, arat hepatoma cell line (16), PK15, a pig kidney cell line (24),and A431 (25), a human epidermoid carcinoma cell line, whichwere purchased from ATCC (Rockville, MD). Cells were grownand maintained in 10% FCS in DMEM supplemented with 1mM L-glutamine, Na pyruvate and non-essential amino acids(Gibco/BRL, Gaithersburg, MD).
ANA assays
Target cells were cultured to near confluence as describedabove. Cultures with at least 95% cell viability were used forANA determination. Cells were washed 3 times with PBS (pH7.4) and fixed with cold acetone/methanol at –20°C for 10 min.After an additional 2�PBS washes, the fixed cells were blockedwith 5% normal goat serum (Jackson Immuno-Research, West

418 Intracellular autoantibodies
Fig. 4. Cell lineage susceptibility to autoantibody penetration. Penetration of mAb 21 and 5B5 was demonstrated using cell lines of differenttissue origins derived from several animal species. Both monoclonal autoantibodies were able to penetrate a human epidermoid carcinomacell line (A431) and a pig kidney cell line (PK15) as determined by in vivo nuclear localization assays performed as previously described forH35 cells. Appropriate antibody controls (mAb 2-12, IgA and IgG2a isotype controls) did not enter the cells (not shown). Images were takenusing a conventional fluorescence microscope (�40).
Fig. 5A.

Intracellular autoantibodies 419
Grove, PA) for 10 min. ANA were then detected with Alexa 488-conjugated goat anti-mouse IgG (H � L) (Molecular Probes,Eugene, OR) for 30 min at room temperature and analyzed byconventional indirect immunofluorescence microscope.
In vivo cell penetration assay
Penetration assays were modified from previously publishedprotocols (14,16). mAb H7 and H9, previously shown to pos-sess in vivo intracellular activity, were kindly provided by DrMichael Madaio (University of Pennsylvania, Philadelphia, PA)(16) and used as positive controls in our initial experiments.Briefly, cells were cultured at a concentration of 5�105/ml inLab Tek chamber slides (Nalge Nunc, Naperville, IL) and usedwhen they had reached at least 80% confluence. At that point,cells were washed twice with 1% BSA/PBS and incubated withthe appropriate mAb (30 µg/ml) in 1% BSA/DMEM for a prede-termined period of time. Cells were then extensively washedand their viability was assessed by their ability to exclude vitaldyes such as Trypan blue or propidium iodide. In order toensure that any intracellular antibody observed was indeed theresult of in vivo penetration that occurred prior to fixation, thecells were snap-fixed in pre-chilled acetone/methanol at –20°Cfor 10 min. Alternatively, cells were fixed using different, freshlypreparedcross-linkingagents including4%paraformaldehydefor 1 h at room temperature and 3% formaldehyde on ice for 15min followed by permeabilization with 0.2% Triton X-100 for 5min on ice. After 3 washes with PBS, intracellular antibodieswere detected using Alexa 488-conjugated secondary anti-bodies as described above. IgA antibodies were also labeledusing the same secondary antibody which also recognizes κlight chains. For co-localization experiments, the nucleus wasco-stained with propidium iodide (0.5 µg/ml) for 10 min at roomtemperature. Results were then visualized under an OlympusBX60 fluorescent microscope (Tokyo, Japan) or scanned usinga BioRad MRC 600 confocal microscope (Hercules, CA).
Internalization kinetics
For mAb 21 kinetics, H35 cells were detached from the tissueculture dish by incubation with 10 mM EDTA/PBS (pH 8.0)and cell penetration was assayed at different time points asdescribed above. These experiments were performed either at37°C or, alternatively, at 4°C by keeping the target cells and allreagents on ice previous to the incubation experiments whichwere then carried out in the cold-room at temperatures ranging
Fig. 5. Co-localization of intracellular autoantibodies and endocyticvesicles. (a) Texas Red–dextran (1 mg/ml) and the correspondingmAb were co-incubated with live H35 cells at 37°C for 30 min.Intracellular antibody was labeled with Alexa 488-conjugatedsecondary antibody as before. Samples were scanned using a Leicaconfocal microscope (�100) under the appropriate settings in orderto detect Texas Red (right panel) or Alexa 488 (middle panel).Individual images of the same field were merged to detect co-localization of mAb and dextran (right panel). As compared toendocytosed IgA control, only a small percentage of vesiclescontaining mAb 21 or 5B5 also contained dextran. Quantification ofendocytosis vesicles containing both mAb and dextran. (b) Resultsfrom the experiments shown in (5) are expressed as the percentageof Alexa 488� vesicles also containing Texas Red.
between 0 and 4°C. Cell viability was monitored by Trypan blueexclusion and was consistently �90% at the time of fixation.The amount of internalized antibodies was measured as meanfluorescence intensity using an Elite flow cytometer (Coulter,Fullerton, CA). Nuclear localization kinetics of mAb 5B5 wasdetermined by indirect immunofluorescence as describedabove for in vivo penetration assays. Nuclear fluorescence wasthen quantified from digital images obtained with a Sony DXC-9000 3CCD camera (Tokyo, Japan) using the Image-Pro Plussoftware (Media Cybernetics, Silver Spring, MD). Six fieldswere counted from each sample in at least three independentexperiments
Fluid-phase endocytosis experiments
Intracellular transport of mAb was compared to trafficking ofTexas Red–dextran. To do so, H35 cells were incubated withantibodies at 30 µg/ml and Texas Red–dextran (70,000 kDa) at1 mg/ml (Molecular Probes) in 1% BSA/DMEM for 30 min at37°C. After 3 washes with PBS, cells were fixed with cold acet-one/methanol and intracellular mAb were detected with Alexa488-conjugated goat anti-mouse IgG (H � L) as described forthe penetration assay. Samples were scanned using a Leicaconfocal microscope (Heerbrugg, Switzerland) to obtainimagesofAlexa488andTexas red labeling respectively. Imageprocessing was performed with Adobe Photoshop software(San Jose, CA). Both granules containing only mAb, and thosecontaining dextran and mAb together were counted. Resultswere expressed as the fraction of total mAb granules that alsocontained dextran. A final score was generated from countinga minimum of three fields from three independent experiments.
Antibody sequence analysis
The DNA sequences of the mAb have been previously reportedand were obtained from GenBank (accession nos X67195,L08989, L08980, U26999, L09013, U18595, S71117 andS71116). The sequences were imported into the EditSeq pro-gram of the DNASTAR Lasergene software (Madison, WI) andanalyzed for VH and VL germline homology, and the presenceand pattern of somatic mutations using the Megalign programof DNASTAR and the Blastn search program available throughthe National Center for Biotechnology Information. Analysisof the predicted protein sequences was performed using theProtean program of DNASTAR.

420 Intracellular autoantibodies
Fig. 6. Sequence analysis of the HCDR3 of mAb. The predicted amino acid sequences of both penetrating (mAb21 and 5B5) and non-penetrating mAb (1-12 and 2-12) are aligned. Arrows indicate the areas corresponding to the end of the FR3 as well as conventional HCDR3regions. Also, the extended HCDR3 loop containing the antigen contact residues is indicated in the figure. The last residue shown in thefigure (W) corresponds to the beginning of the FR4 for all sequences. Basic residues are shown in bold italics and the RR doublet unique topenetrating antibodies is encased.
Results
In vivo penetration of H35 cells by mouse monoclonal autoanti-bodies
In order to explore the prevalence of the intracellular autoanti-body phenomenon and given that the preponderance of previ-ousstudieshad focusedonanti-DNAantibodies,wedecided totest a panel of disease-associated monoclonal autoantibodieswhose primary specificity was other than DNA. Moreover, wewanted to determine whether this phenomenon is restricted toany particular isotype. Thus, four anti-Sm and one anti-La mAbderived from autoimmune MRL/lpr�/� mice were selected forour in vivo penetration assay. These mAb were previously char-acterized and have been described elsewhere (21–23). Inselecting these autoantibodies we were mindful of the factthat anti-RNP autoantibodies often display significant cross-reactivity with single-stranded DNA and/or double-strandedDNA. Therefore, we chose to study antibodies with differentdegrees of cross-reactivity with DNA. Furthermore, it has beensuggested that pathogenic anti-DNA antibodies may predomi-nantly cross-react with Sm/RNP-associated polypeptides Aand D (24). Accordingly, we selected anti-Sm antibodies withdifferent degrees of reactivity against either the D or the B/B� polypeptides of the Sm antigen, which represent the mainantigenic targets of the anti-Sm response. Relevant character-istics of these mAb including in vitro ANA pattern and in vivopenetration ability are summarized in Table 1.
As expected, all mAb showed in vitro ANA activity. As shownin Fig. 1, among those five mAb, only two mAb, 21 (anti-La,IgG2a) and 5B5 (anti-Sm, IgA), were detected in the nucleusof live H35 cells. In order to ensure that nuclear localization ofmAb was not due to leakage of cell and nuclear membranes,cell viability was monitored by Trypan blue or propidium iodideexclusion before fixation in all experiments (data not shown).The in vivo nuclear accumulation of mAb was not affectedby different fixatives including cross-linking reagents such asfreshly prepared 4% paraformaldehyde or methanol-free 3%formaldehyde (data not shown). Significant similarities couldbe observed between the ANA patterns and the in vivo stainingprofiles. Thus, mAb21 created a nuclear rim pattern in bothassays while a consistent speckled pattern was seen with 5B5.In addition to the nuclear localization, granules containing mAb21 were also observed in the cytoplasm of most H35 cellsand somewhat finer ones with mAb 5B5. Large cytoplasmicgranules were also observed with the IgA isotype control anti-body, but in the absence of nuclear staining. This observation
might be explained by endocytosis of IgA via the polymericIgA receptor (pIgR) that mediates transcytosis in hepatocytes(26–29).
Taken together, these results strongly suggest that the cellentry and nuclear localization of the mAb is Fc receptor inde-pendent (as demonstrated by lack of activity of other mAb ofthe same isotype and of polyclonal IgG and IgA antibodies).Moreover, the consistently positive results obtained with mAb21 and 5B5 under different fixation protocols along with thenegative controls just discussed provide further proof that thisphenomenon is not the result of fixation artifacts.
The staining profile obtained with mAb21 suggested that thisantibody might simply bind to the nuclear membrane withoutactually entering the nucleus. This issue was resolved by usinglaser scanning confocal microscopy to show that both mAb 21and 5B5 did indeed co-localize with propidium iodide-stainedmaterial within the cell nucleus (Fig. 2). It should be appreci-ated, however, that within the nucleus, there was only partialoverlapbetween the twoprobes,consistentwith the recognitionby the mAb of nuclear antigens other than or in addition todouble-stranded DNA. This is not surprising in light of theirprimary antigenic reactivity against the Sm and La proteinsrespectively.
In vivo penetration kinetics
Both mAb, 21 and 5B5, penetrated live cells in a time-depend-ent manner. Internalization kinetics of mAb 21 was determinedby flow cytometry. Cell aliquots sampled at each time pointwere also observed under fluorescent microscopy to confirmnuclear localization of the antibody. In turn, mAb 5B5 presenteda different challenge based on its IgA isotype. As previouslyshown, IgAantibodiesare internalizedbyH35cellspresumablythrough the pIgR and, therefore, mere cell penetration as indi-cated by flow cytometry would not be indicative of nuclearlocalization. Accordingly, 5B5 kinetics was determined bydirect measurement of nuclear fluorescence intensity from thedigital images obtained by a CCD camera. As shown in Fig. 3,mAb 21 and 5B5 entered live cells in a very rapid manner,and could be detected in the cell nucleus within 15 min ofincubation. Penetration of mAb 21 reached a plateau between30 and 60 min, while maximal nuclear staining with mAb 5B5was seen within 30 min. Furthermore, the maximum nuclearaccumulation of mAb 21 was significantly higher than that ofmAb 5B5 when both antibodies were assayed under fluores-cent microscopy (data not shown). These results may reflectdifferences between both mAb in terms of efficiency in cell

Intracellular autoantibodies 421
penetration or differences in the efficiency of nuclear importand/or export. Also of note, no significant differences in cellpenetration or nuclear accumulation were observed when thesame experiments were performed at temperatures of 0–4°C,which are expected to significantly interfere with active trans-port mechanisms (results not shown).
Cell type susceptibility to autoantibody penetration
Along with H35, two additional cell lines, A431, a human epi-dermoid carcinoma, and PK15, a pig kidney cell line, weretested todetermine their susceptibility toautoantibody penetra-tion. As shown in Fig. 4, mAb 21 and 5B5 were also able toenter both cell types. As with H35, nuclear localization wasfound only with these autoantibodies but not with IgA or IgGisotype controls or with a negative autoantibody control mAb2-12.
Nuclear localizing mAb utilize distinct intracellular transportmechanisms
As a preliminary attempt to characterize the mechanism(s) ofautoantibody uptake and transport, we asked whether the mAbcould be found in endocytic vesicles. We used dextran, anindicator of fluid phase endocytosis (30,31), to track the endo-cytic pathway. As shown in Fig. 5, only a relatively small fractionof vesicles contained both mAb and dextran together (27.4and 18.7% for mAb 5B5 and mAb21 respectively). In contrast,�85% of dextran-positive vesicles also carried the IgA isotypecontrol antibody. The scattering pattern of the cytoplasmic IgA–dextran vesicles is consistent with them being contained byendosomes. The remaining 15% of vesicles likely representstranscytotic vesicles carrying post-endosomal IgA molecules.
Discussion
The actual role and mechanism(s) of action of anti-nuclearautoantibodies in the systemic autoimmune diseases are stillquestionable 50 years after the discovery of the ANA phenom-enon (32). The most widely accepted pathogenic mechanismis the formation of circulating immune complexes with inflam-matory properties and indeed such mechanism has been welldocumented with anti-DNA antibodies. Given the intranuclearnatureofmostof theautoantigens recognized, it seemsobviousthat the ability of autoantibodies to enter subcellular compart-ments would create additional and tantalizing opportunities forthese molecules to generate functional perturbation and tissuedamage. This report sheds light on this poorly understoodaspect of the autoimmune process. Our results demonstratethat, in addition to DNA, other major nuclear autoantigens suchas the Sm and La RNP can also be the target of intracellularautoantibodies.First, it shouldbeemphasized that thispropertywas observed in only a subset of the autoantibodies tested andthat it was independent of the type of fixatives used, eitheracetone or cross-linking agents including 4% paraformal-dehyde. This result indicates that our observations are not theresult of fixation artifacts with disruption of the plasma mem-brane and/or the nuclear envelope as it has been suggestedelsewhere (20). Also of importance, both the absence of Fcreceptors in the cell lines tested as well as the lack of internaliz-ation of isotype control antibodies strongly suggest that thisphenomenon is Fc receptor independent and is most likely
mediated by the antigen binding site of the antibodies tested.Being a liver epithelial cell line, however, H35 cells do expressthe pIgR that normally mediates endocytosis and transcytosisof both dimeric IgA and pentameric IgM. Yet, as demonstratedby using a control polyclonal IgA antibody, internalization ofIgA through the pIgR does not result in nuclear translocation,thus reaffirming the specificity of this phenomenon for the IgAmAb with anti-Sm reactivity (5B5).
Several factors related to the antigenic reactivity of mAb 21and 5B5 might be responsible for their intracellular activity.First, anti-Sm and anti-La antibodies may display significantcross-reactivity with single-stranded and/or double-strandedDNA, and, therefore, it is plausible that such cross-reactivitymight be ultimately responsible for cell penetration. In oursample, only autoantibodies with cross-reactivity with double-stranded DNA but not single-stranded DNA were able to enterlive cells (see Table 1) suggesting that double-stranded DNAbinding may be important for antibody penetration. Secondly,anti-Sm antibodies may react with one or more of the multiplepolypeptides that form the snRNP complex. The mAb tested inthis study were chosen to represent a spectrum of anti-Smreactivity with special emphasis on the main antigenic targets,i.e. the B/B� and/or D polypeptides (21). While mAb 5B5 reactsonly with the Sm-D polypeptide, other anti-Sm-D mAb failed topenetrate live cells, as did mAb 1-12 which binds to both Sm-B/B� and -D polypeptides. Hence, on a superficial analysisit would seem that the global polypeptide reactivity of theseautoantibodies is not essential, or at least not sufficient, for cellentry. It is possible, however, that the intracellular propertiescould be dependent on fine epitope specificity in which casethe broader reactivity against the full-lengthpolypeptides mightnot be informative enough (detailed epitope mapping of theanti-Sm mAb is currently underway). Along similar lines, thefine specificity of anti-La antibodies might also represent animportant factor in their ability to translocate into the cell nuc-leus. The demonstration that the ability of anti-La antibodies torecognize different La–RNA complexes is dependent on theirfine specificity would appear to support this notion (33,34).Alternatively, intracellular autoantibodies might need to cross-react with surface-exposed antigens as it has been suggestedfor some anti-DNA antibodies that recognize myosin I (35).Although our antibodies did not recognize myosin I as deter-mined by conventional ELISA (results not shown), other pos-sible targets include laminin, a main component of basalmembranes that is recognized by some pathogenic anti-Laantibodies capable of inducing congenital heart block (36).Another potential scenario contemplates the possibility that theintracellular antigens themselves could be expressed in thecell membrane, and, indeed, both La and the snRNP have beenshown to translocate to the cell surface under diverse stimuliincluding UV radiation, estrogen stimulation, viral infectionsand exposure to tumor necrosis factor-α (37–41).
Whether the antigen responsible for autoantibody penetra-tion turns out to be the nominal antigen (Sm or La) or a cross-reactive antigen(s) such as DNA, our results suggest that therecognized target must be a widely expressed molecule sinceseveral cell lines including rat liver cells, pig kidney cells andhuman epidermoid carcinoma cells were susceptible to anti-body penetration. This finding is consistent with the observa-tions of Vlahakos et al., that the in vivo pathogenic potential of

422 Intracellular autoantibodies
a subset of anti-double-stranded DNA antibodies administeredto normal mice correlated with their ability to accumulate in thecell nucleus in multiple tissues (16), and it supports the notionthat anti-Sm and anti-La autoantibodies could mediate sys-temic autoimmune tissue damage.
The mechanismresponsible for intracellular transport of mAbremains largely unexplored and may not correspond to thegeneral pathways of protein kinesis. Our results represent asignificant departure from the observations reported with thesubset of anti-DNA autoantibodies that cross-react with myosinwhich are internalized by receptor-mediated endocytosis (35).Thus, in our case nuclear accumulation was faster with signi-ficant nuclear staining observed within minutes. This feature isreminiscent of the rapid cell entry and nuclear translocationobserved with the HIV Tat protein. This translocation propertyhas been assigned to a region of the Tat protein centered on acluster of basic amino acids and recent data have demon-strated that chemical coupling of this Tat-derived peptide toseveral proteins allowed their internalization into several celllines or tissues (42). As with our mAb, the rapid uptake of Tatpeptides appears to be temperature independent, suggestingthat the internalization process does not follow conventionalendocytosis rules.
Multiple studies have shown that the presence of basic res-idues in the antibody hypervariable regions, mostly arginineresidues in the HCDR3, may be responsible for anti-DNA react-ivity (43) and that cationization of otherwise irrelevant antibod-ies also appears to induce cellular uptake (44). Furthermore,these stretches of basic residues may closely resemble con-ventional nuclear localization signals (NLS) such as the oneused by the SV-40 large T antigen (45,46). Close inspection ofthe published hypervariable region sequences of mAb 21 and5B5 did not reveal a typical NLS. However, some distinct fea-tures of potential significance can be found in the HCDR3 ofthese antibodies (Fig. 6). Thus, both HCDR3s display an RRdoublet at the 5� end where the second arginine appears tohave been generated by junctional diversity and presumablyselected by antigen. In the case of mAb21 another amino acidwith positive charge potential (histidine) is located one residueaway from the RR doublet and contributes to create a signific-antly positive charge in the HCDR3 of this antibody. In contrast,these features were absent in the non-penetrating antibodieswhose HCDR3 global charge was significantly less positive, asthe result in some cases of a higher content of acidic residueswhich may promote binding to the basic snRNP polypeptides(47).
In summary, in this paper we demonstrate that autoanti-bodies whose primary antigenic specificity is directed againstthe major RNP targets of systemic autoimmune diseases suchas systemic lupus erythematosus and Sjogren’s syndrome canrapidly traverse the plasma membrane of live cells of differenttissue origin and translocate into the nucleus. Although cross-reactivity with double-stranded DNA may play a role, the actualrequirements for this phenomenon remain to be determined,and, indeed, previous results by other investigators and ourown studies show that anti-DNA reactivity per se is not sufficientfor intracellular activity. The rapid rate of internalization, the lackof significant co-localization with endocytic vesicles and theapparent temperature independence observed in our studiesstronglypoint towardsamechanismdifferent fromconventional
receptor-mediated endocytosis. Experiments aimed at the elu-cidation of these issues are currently underway in ourlaboratory.
The observations presented herein hold significant promisefor the studyof importantbiological functionssuchas intracellu-lar and nucleocytoplasmic transport of proteins as well as forour understanding of systemic autoimmunity. Furthermore,once the requirements for in vivo intracellular activity of (auto)-antibodies are established, this information should proveinvaluable for the design of antibodies as intracellular thera-peutic agents and drug-delivery systems.
Acknowledgements
We are indebted to Drs M. P. Madiao (University of Pennsylvania) andS. Clarke (University of North Carolina at Chapel Hill) for providing theanti-DNA and anti-Sm/La mAb respectively. We are equally indebtedto Dr D. Goldfarb (University of Rochester) for advice and discussionson nuclear transport. We thank Kathy Troughton (University of Ten-nessee at Memphis) and Hiram Lyon (University of Rochester) for theirexcellent assistance with confocal microscopy. We are also grateful toDrs F. Young, E. Schwarz and M. Nahm for reviewing the manuscript.This study was supported partially by NIH grants PHS AG14878 andAG14585. S. X. D. is supported by NIH 2T32AI07285 Training Grantand the MD/PhD program at the University of Rochester.
Abbreviations
ANA anti-nuclear antibody assayNLS nuclear localization signalpIgR polymeric IgA receptorRNP ribonuleoprotein
References
1 Tan, E. M. 1989. Antinuclear antibodies: diagnostic markers forautoimmune diseases and probes for cell biology. Adv. Immunol.44:93.
2 Craft, J. 1992. Antibodies to snRNPs in systemic lupuserythematosus. Rheum. Dis. Clin. N. Am. 18:311.
3 Shero, J. H., Bordwell, B., Rothfield, N. F. and Earnshaw, W. C. 1986.High titers of autoantibodies to topoisomerase I (Scl-70) in serafrom scleroderma patients. Science 231:737.
4 Balczon, R. 1993. Autoantibodies as probes in cell and molecularbiology. Proc. Soc. Exp. Biol. Med. 204:138.
5 Casiano, C. A. and Tan, E. M. 1996. Antinuclear autoantibodies:probes for defining proteolytic events associated with apoptosis.Mol. Biol. Rep. 23:211.
6 Huhn, P., Pruijn, G. J., van Venrooij, W. J. and Bachmann, M. 1997.Characterization of the autoantigen La (SS-B) as a dsRNAunwinding enzyme. Nucleic Acids Res. 25:410.
7 Tan, E. M., Muro, Y. and Pollard, K. M. 1994. Autoantibody-definedepitopes on nuclear antigens are conserved, conformation-dependent and active site regions. Clin. Exp. Rheumatol. 12:S27.
8 Alarcon-Segovia, D., Ruiz-Arguelles, A. and Fishbein, E. 1978.Antibody to nuclear ribonucleoprotein penetrates live humanmononuclear cells through Fc receptors. Nature 271:67.
9 Golan, T. D. 1997. The nuclear compartment of living cells is not anabsolute immunologically sequestered site: evidence forspontaneous intranuclear entry of lupus autoantibodies to livingcells. Leukemia 11:6.
10 Alarcon-Segovia, D., Ruiz-Arguelles, A. and Llorente, L. 1996.Broken dogma: penetration of autoantibodies into living cells.Immunol. Today 17:163.
11 Ma, J., Chapman, G. V., Chen, S. L., Melick, G., Penny, R. and Breit,S. N. 1991. Antibody penetration of viable human cells. I. Increasedpenetration of human lymphocytes by anti-RNP IgG. Clin. Exp.Immunol. 84:83.

Intracellular autoantibodies 423
12 Ma, J., King, N., Chen, S. L., Penny, R. and Breit, S. N. 1993.Antibody penetration of viable human cells. II. Anti-RNP antibodiesbinding to RNP antigen expressed on cell surface, which maymediate the antibody internalization. Clin. Exp. Immunol. 93:396.
13 Hormigo, A., Ball, C. and Lieberman, F. 1996. Internalization of anti-Hu IgG is not Fc gamma receptor mediated. Neurology 47:607.
14 Koscec, M., Koren, E., Wolfson-Reichlin, M., Fugate, R. D., Trieu,E., Targoff, I. N. and Reichlin, M. 1997. Autoantibodies to ribosomalP proteins penetrate into live hepatocytes and cause cellulardysfunction in culture. J. Immunol. 159:2033.
15 Avrameas, A., Ternynck, T., Nato, F., Buttin, G. and Avrameas, S.1998. Polyreactive anti-DNA monoclonal antibodies and a derivedpeptide as vectors for the intracytoplasmic and intranucleartranslocation of macromolecules [in process citation]. Proc. NatlAcad. Sci. USA 95:5601.
16 Vlahakos, D., Foster, M. H., Ucci, A. A., Barrett, K. J., Datta, S. K.and Madaio, M. P. 1992. Murine monoclonal anti-DNA antibodiespenetrate cells, bind to nuclei, and induce glomerular proliferationand proteinuria in vivo. J. Am. Soc. Nephrol. 2:1345.
17 Zack, D. J., Stempniak, M., Wong, A. L., Taylor, C. and Weisbart, R.H. 1996. Mechanisms of cellular penetration and nuclearlocalization of an anti-double strand DNA autoantibody.J. Immunol. 157:2082.
18 Alarcon-Segovia, D., Ruiz-Arguelles, A. and Llorente, L. 1979.Antibody penetration into living cells. II. Anti-ribonucleoprotein IgGpenetrates into Tgamma lymphocytes causing their deletion andthe abrogation of suppressor function. J. Immunol. 122:1855.
19 Alarcon-Segovia, D., Llorente, L. and Ruiz-Arguelles, A. 1996. Thepenetration of autoantibodies into cells may induce tolerance to selfby apoptosis of autoreactive lymphocytes and cause autoimmunediseasebydysregulationand/orcell damage.J.Autoimmun.9:295.
20 Kramers, K., van Bruggen, M. C., Rijke-Schilder, T. P., Dijkman, H.B., Hylkema, M. N., Croes, H. J., Fransen, J. A., Assmann, K. J.,Tax, W. J., Smeenk, R. J. and Berden, J. H. 1996. In vivo ANA is afixation artifact: nucleosome-complexed antinucleosomeautoantibodies bind to the cell surface and are internalized. J. Am.Soc. Nephrol. 7:946.
21 Bloom, D. D., Davignon, J. L., Retter, M. W., Shlomchik, M. J.,Pisetsky, D. S., Cohen, P. L., Eisenberg, R. A. and Clarke, S. H.1993. V region gene analysis of anti-Sm hybridomas from MRL/Mp-lpr/lpr mice. J. Immunol. 150:1591.
22 Bloom, D. D., Davignon, J. L., Cohen, P. L., Eisenberg, R. A. andClarke, S. H. 1993. Overlap of the anti-Sm and anti-DNA responsesof MRL/Mp-lpr/lpr mice. J. Immunol. 150:1579.
23 Bloom, D. D., St. Clair, E. W., Pisetsky, D. S. and Clarke, S. H. 1994.The anti-La response of a single MRL/Mp-lpr/lpr mouse: specificityfor DNA and VH gene usage. Eur. J. Immunol. 24:1332.
24 Koren, E., Koscec, M., Wolfson-Reichlin, M., Ebling, F. M., Tsao, B.,Hahn, B. H. and Reichlin, M. 1995. Murine and human antibodiestonativeDNAthatcross-reactwith theAandDSnRNPpolypeptidescause direct injury of cultured kidney cells. J. Immunol. 154:4857.
25 Golan, T. D., Gharavi, A. E. and Elkon, K. B. 1993. Penetration ofautoantibodies into living epithelial cells. J. Invest. Dermatol.100:316.
26 Takahashi, I., Nakane, P. K. and Brown, W. R. 1982. Ultrastructuralevents in the translocation of polymeric IgA by rat hepatocytes. J.Immunol. 128:1181.
27 Mullock, B. M., Hinton, R. H., Dobrota, M., Peppard, J. and Orlans,E. 1979. Endocytic vesicles in liver carry polymeric IgA from serumto bile. Biochim. Biophys. Acta 587:381.
28 Socken, D. J., Jeejeebhoy, K. N., Bazin, H. and Underdown, B. J.1979. Identification of secretory component as an IgA receptor onrat hepatocytes. J. Exp. Med. 150:1538.
29 Mostov, K. E. 1994. Transepithelial transport of immunoglobulins.Annu. Rev. Immunol. 12:63.
30 Berlin, R.D. andOliver, J. M.1980. Surface functionsduring mitosis.
II. Quantitation of pinocytosis and kinetic characterization of themitoticcyclewithanewfluorescence technique.J.CellBiol.85:660.
31 van Deurs, B., Ropke, C. and Thorball, N. 1984. Kinetics ofpinocytosis studied by flow cytometry. Eur. J. Cell Biol. 34:96.
32 Hargraves, M., Richmond, M. and Morton, R. 1948. Presentation oftwo bone marrow elements: the ‘tart’ cell and ‘LE’ cell. Mayo ClinProc. 23:25.
33 Pruijn, G. J., Thijssen, J. P., Smith, P. R., Williams, D. G. and VanVenrooij, W. J. 1995. Anti-La monoclonal antibodies recognizingepitopes within the RNA-binding domain of the La protein showdifferential capacities to immunoprecipitate RNA-associated Laprotein. Eur. J. Biochem. 232:611.
34 Veldhoven, C. H., Pruijn, G. J., Meilof, J. F., Thijssen, J. P., Vander Kemp, A. W., Van Venrooij, W. J. and Smeenk, R. J. 1995.Characterization of murine monoclonal antibodies against 60-kDRo/SS-A and La/SS-B autoantigens. Clin. Exp. Immunol. 101:45.
35 Yanase, K., Smith, R. M., Puccetti, A., Jarett, L. and Madaio, M. P.1997. Receptor-mediated cellular entry of nuclear localizing anti-DNA antibodies via myosin 1. J. Clin. Invest. 100:25.
36 Li, J. M., Horsfall, A. C. and Maini, R. N. 1995. Anti-La (SS-B) butnot anti-Ro52 (SS-A) antibodies cross-react with laminin–a role inthe pathogenesis of congenital heart block? Clin. Exp. Immunol.99:316.
37 Golan, T. D., Elkon, K. B., Gharavi, A. E. and Krueger, J. G. 1992.Enhanced membrane binding of autoantibodies to culturedkeratinocytes of systemic lupus erythematosus patients afterultraviolet B/ultraviolet A irradiation. J. Clin. Invest. 90:1067.
38 Dorner, T., Hucko, M., Mayet, W. J., Trefzer, U., Burmester, G. R.and Hiepe, F. 1995. Enhanced membrane expression of the 52 kDaRo(SS-A) and La(SS-B) antigens by human keratinocytes inducedby TNF alpha. Ann. Rheum. Dis. 54:904.
39 Furukawa, F., Lyons, M. B., Lee, L. A., Coulter, S. N. and Norris, D.A. 1988. Estradiol enhances binding to cultured humankeratinocytes of antibodies specific for SS-A/Ro and SS-B/La.Another possible mechanism for estradiol influence of lupuserythematosus. J. Immunol. 141:1480.
40 Baboonian, C., Venables, P. J., Booth, J., Williams, D. G., Roffe, L.M. and Maini, R. N. 1989. Virus infection induces redistribution andmembrane localization of the nuclear antigen La (SS-B): a possiblemechanism for autoimmunity. Clin. Exp. Immunol. 78:454.
41 Bachmann, M., Chang, S., Bernd, A., Mayet, W., Meyer zumBuschenfelde, K. H. and Muller, W. E. 1991. Translocation of thenuclear autoantigen La to cell surface: assembly and disassemblywith the extracellular matrix. Autoimmunity 9:99.
42 Vives, E., Brodin, P. and Lebleu, B. 1997. A truncated HIV-1 Tatprotein basic domain rapidly translocates through the plasmamembrane and accumulates in the cell nucleus. J. Biol. Chem.272:16010.
43 Radic, M. Z., Mackle, J., Erikson, J., Mol, C., Anderson, W. F. andWeigert, M. 1993. Residues that mediate DNA binding ofautoimmune antibodies. J. Immunol. 150:4966.
44 Pardridge, W. M., Buciak, J., Yang, J. and Wu, D. 1998. Enhancedendocytosis in cultured human breast carcinoma cells and in vivobiodistribution in rats of a humanized monoclonal antibody aftercationization of the protein. J. Pharmacol. Exp. Ther. 286:548.
45 Michaud, N. and Goldfarb, DS. 1993. Most nuclear proteins areimported by a single pathway. Exp. Cell Res. 208:128.
46 Yoneda, Y., Semba, T., Kaneda, Y., Noble, RL., Matsuoka, Y.,Kurihara, T., Okada, Y. and Imamoto, N. 1992. A long syntheticpeptide containing a nuclear localization signal and its flankingsequences of SV40 T-antigen directs the transport of IgM into thenucleus efficiently. Exp. Cell Res. 201:313.
47 Woppmann, A., Patschinsky, T., Bringmann, P., Godt, F. andLuhrmann, R. 1990. Characterisation of human and murine snRNPproteins by two-dimensional gel electrophoresis andphosphopeptide analysis of U1-specific 70K protein variants.Nucleic Acids Res. 18:4427.




















