
Systems biology of myasthenia gravis, integration of aberrant ...
Upload
independentCategory
view
1download
0
200 (2008) 133–141www.elsevier.com/locate/jneuroim
Journal of Neuroimmunology
Antigen-specific apheresis of human anti-acetylcholine receptorautoantibodies from myasthenia gravis patients' sera using
Escherichia coli-expressed receptor domains
Paraskevi Zisimopoulou a,1, George Lagoumintzis b,1, Konstantinos Poulas b, Socrates J. Tzartos a,b,⁎
a Department of Biochemistry, Hellenic Pasteur Institute, 127, V. Sofias Ave., GR11521, Athens, Greeceb Department of Pharmacy, University of Patras, Rio, GR26500, Patras, Greece
Received 4 March 2008; received in revised form 23 May 2008; accepted 2 June 2008
Abstract
Myasthenia gravis (MG) is an autoimmune disease usually caused by autoantibodies against the muscle nicotinic acetylcholine receptor(nAChR), found at the neuromuscular junction. Current treatments for the disease, including plasmapheresis, are not antigen-specific, and thusoften cause severe side effects. The development and implementation of new therapeutic approaches for the disease attain particular importance,since the number of diagnosed myasthenic patients has increased considerably. In order to develop an antigen-specific approach for the selectivedepletion of anti-nAChR autoantibodies from MG patients' sera, we have expressed the extracellular domains (ECDs) of all human musclenAChR subunits in E. coli. These recombinant proteins were immobilized on CNBr-Sepharose and used in immunoadsorption assays with MGsera. We showed that, despite being purified from E. coli under denaturing conditions, these recombinant ECDs could be successfully used asimmunoadsorbents with a comparable efficiency to the corresponding ECDs produced in water-soluble form in the yeast Pichia pastoris. The highyield of the ECDs, the stability of the constructed ECD resins and the high selectivity for anti-nAChR antibodies increase the therapeutic potentialof the system.© 2008 Elsevier B.V. All rights reserved.
Keywords: Acetylcholine receptor autoantibodies; Extracellular domains; Immunoadsorption; Myasthenia gravis; Therapy; Apheresis
1. Introduction
Myasthenia gravis (MG) is an acquired autoimmune diseaseaffecting 200–400 cases per million (Robertson et al., 1998; Poulaset al., 2001). The typical chronic muscle weakness seen in at least85% of MG patients seems to be caused by autoantibodies againstthe muscle nicotinic acetylcholine receptor (nAChR) (Drachman,1994; Lindstrom, 2000; Vincent, 2002) which result in thedestruction of the postsynaptic membrane at the neuromuscular
Abbreviations: nAChR, nicotinic acetylcholine receptor; MG, myastheniagravis; ECD, extracellular domain; RIA, radioimmunoassay; NHS, normal humanserum; BSA, bovine serum albumin; MuSK, muscle-specific kinase; PBS,phosphate-buffered saline; IVIg, intravenous immunoglobulin.⁎ Corresponding author. Department of Biochemistry, Hellenic Pasteur Institute,
127, V. Sofias Ave., GR11521, Athens, Greece. Tel.: +30 2106478844; fax: +302106478842.
E-mail addresses: [email protected], [email protected] (S.J. Tzartos).1 These authors contributed equally to this work.
0165-5728/$ - see front matter © 2008 Elsevier B.V. All rights reserved.doi:10.1016/j.jneuroim.2008.06.002
junction (NMJ). Despite the fact that the anti-nAChR antibody titredoes not correlate directly with MG severity in the MG population(Lindstrom et al., 1976b; Lindstrom, 1985; Kuks et al., 1993; Romiet al., 2000), there is growing evidence that the anti-nAChRautoantibodies trigger the pathogenesis of the disease, since:a) antibodies are present at the site of the pathology i.e. the NMJ(Drachman et al., 1973; Fambrough et al., 1973); b) immunoglo-bulins (Igs) from MG patients or anti-nAChR antibodies fromexperimental animals cause MG symptoms when injected intorodents (Lindstrom et al., 1976a; Tzartos et al., 1998);c) immunization of animals with the nAChR induces the disease(Patrick and Lindstrom, 1973); d) therapies that remove theautoantibodies decrease the severity of MG symptoms andtransiently relieve the clinical symptoms (Newsom-Davis et al.,1979) and e) isolated anti-nAChR autoantibodies or whole MGsera, but not anti-nAChR antibody-depleted sera, induce nAChRantigenic modulation in vitro (Sideris et al., 2007) and passivetransfer of experimental MG in vivo (Kordas, Sideris, Poulas and

134 P. Zisimopoulou et al. / Journal of Neuroimmunology 200 (2008) 133–141
Tzartos, unpublished data). In approximately one-third of MGpatients without anti-nAChR antibodies (seronegative patients),antibodies against the muscle-specific kinase (MuSK) have beenfound (Hoch et al., 2001; Vincent et al., 2001), while the cause ofMG in a small percentage of patients is still unknown.
Muscle nAChR molecules are transmembrane proteinsformed by 5 subunits: two identical α subunits, which contributeimportant structural elements to the ACh-binding sites, and threedifferent, but homologous, subunits, named β, γ (in the embryo,and replaced by ε in the adult) and δ (Karlin, 2002; Le Novereet al., 2002; Unwin, 2005). The subunits consist of an ∼210amino acid extracellular domain (ECD), four transmembranedomains, a large cytoplasmic loop and a small extracellular C-terminal tail. The ECDs carry most of the epitopes for MGautoantibodies and the α subunit is thought to be immunodo-minant, bearing the main immunogenic region (Tzartos et al.,1982, 1998); the immunogenicity of the non-α subunits has beenless studied, but the ability of their ECDs to bind autoantibodiesfromMG sera has been recently demonstrated (Kostelidou et al.,2007).
Current management of MG includes the use of antic-holinesterase drugs for temporary improvement of neuromusculartransmission, the use of non-specific immunosuppressants orimmunomodulators, such as intravenous Ig (IVIg), to curb theanti-nAChR antibody response, the removal of anti-nAChRautoantibodies by plasma exchange (plasmapheresis) and thy-mectomy (Massey, 1997; Richman and Agius, 2003; Dalakas,2004; Keesey, 2004). However, immunosuppression causesserious side effects, plasmapheresis non-specifically removesimportant plasma constituents (Batocchi et al., 2000) and thepossibility of transmission of as yet unknown infectious agentswith IVIg cannot be excluded. Furthermore, none of the currenttreatments selectively target the MG autoantibodies (Sieb, 2005;Conti-Fine et al., 2006; Fostieri et al., 2006). For this purpose, anaffinity column that selectively removes anti-nAChR antibodieswas introduced by Takamori and co-workers, who used thesynthetic peptide 183–200 of the Torpedo AChR α-subunit as theimmunoadsorbent (Takamori and Maruta, 2001). However, sincethe nAChR has several antigenic epitopes, depletion ofautoantibodies by a single peptide is unlikely to be adequate fortherapy.
In our laboratory, we are developing an antigen-specifictherapeutic approach using the binding of the autoantibodies toimmobilized recombinant ECDs of human nAChR, in order toselectively clear patients' sera of anti-nAChR autoantibodies.These polypeptides have been immobilized on CNBr-Sepharoseand used in immunoadsorption assays with MG sera. Inprevious studies, we used ECDs expressed in water-solubleform in the yeast Pichia pastoris, which proved to be suitableimmunoadsorbents. In an effort to optimize the protein yieldand minimise the difficulty and cost of the procedure, we havenow tested the use of ECDs expressed in E. coli and purifiedunder denaturing conditions as immunoadsorbents. Interest-ingly, in the present study, we showed that these polypeptideswere as capable of specific depletion of MG autoantibodies asthe yeast-expressed ECDs. The stability of the ECD resins andthe high selectivity for the anti-nAChR antibodies increase the
therapeutic potential of the system. We conclude that theseimmunoadsorbents should lead to the development of a newefficient antigen-specific therapy for MG.
2. Materials and methods
2.1. MG sera and measurement of the autoantibody titre
The 41MG serum samples usedwere randomly selected froma relatively large number of anti-nAChR antibody-positive seraheld in our laboratory for diagnostic purposes. Written informedconsent for participation in the study was obtained. All patients'samples were positive for antibody binding to human musclenAChR using a radioimmunoassay (RIA) slightlymodified fromthat described by Lindstrom et al. (1981).
2.2. Human muscle nAChR ECD constructs
All amino acids are numbered according to their position inthe mature protein sequence (ACHB_HUMAN, P11230;ACHG_HUMAN, P07510; ACHD_HUMAN, Q07001;ACHE_HUMAN, Q04844). The cDNA constructs encodingthe entire NH2-terminal extracellular portion of the humanmuscle nAChR subunits, namely β (residues 1–222), γ(residues 1–219), δ (residues 1–225) and ε (residues 1–220),were made by amplification of the corresponding sequence inthe full-length subunit cDNA using PCR (all subunits kindlyprovided by Dr. D. Beeson, University of Oxford, UK). Forconvenience of cloning, an XhoI site (CTCGAG) was added tothe 5′-end of all forward and reverse PCR primers. Addition-ally, a stop codon was added 3′ to the XhoI site of all reverseprimers. Τhe cDNA products of the PCR were digested withXhoI and cloned into the XhoI site of the E. coli expressionvector pET15b (Novagen), downstream of the sequence for the6xHis peptide, and transformed into E. coli TOP10F′ cells(Invitrogen). Plasmids from single clones were checked forinsertion and correct directionality of the insert by restrictiondigest. The sequence of all constructs was confirmed byautomated DNA sequencing (performed by Lark Technologies,Inc., UK). The α subunit ECD (residues 1–207) had alreadybeen cloned in the expression vector pET15b (Tsouloufis et al.,2000).
2.3. Expression and purification of recombinant proteinscorresponding to the extracellular nAChR domains
2.3.1. E. coli-expressed ECDsEach of theE. coli-expressed proteins contained anN-terminal
6xHis tag sequence followed by the residues for the ECD.Recombinant ECDs were produced by growing pET15b/ECD-transformed BL21(pLysS)DE3 cells to an OD590 of 0.6, followedby induction with 1 mM isopropyl-1-thio-b-D-galactopyranosidefor 4 h at 37 °C. Small scale inductions in Luria Bertani broth,followed by cell disruption and centrifugation to separate thesoluble and insoluble protein fractions showed that the proteinswere mainly in the insoluble fraction, so the purificationwas performed under denaturing conditions following the

Table 1Comparison of the yields of recombinant ECDs expressed in E. coli and inP. pastoris
ECD Yield (E. coli) Yield (P. Pastoris)/reference
α (1–210) ∼10 mg/l ∼0.2 mg/l, Psaridi-Linardaki et al., 2005β (1–222) ∼12 mg/l ∼2 mg/l, Kostelidou et al., 2006γ (1–219) ∼12 mg/l ∼0.5 mg/l, Kostelidou et al., 2006δ (1–225) ∼6 mg/l ∼0.2 mg/lε (1–220) ∼8 mg/l ∼0.5 mg/l, Kostelidou et al., 2006
135P. Zisimopoulou et al. / Journal of Neuroimmunology 200 (2008) 133–141
manufacturer's instructions for binding to, and elution from,NiNTAbeads (Qiagen). The proteinswere elutedwith 8Murea in50 mM phosphate buffer containing 150 mM imidazole, pH 8.
2.3.2. P. pastoris-expressed ECDsThe ECD of the α-subunit of the human nAChR (α1–210)
was expressed in P. pastoris strain X-33 as a soluble protein inthe yeast culture supernatant, as described previously (Psaridi-Linardaki et al., 2002). The ECDs of the human β, γ and εsubunits were expressed in a soluble form in the supernatants ofcultures of strain GS115 of P. pastoris (Kostelidou et al., 2006).The human δ-ECD was also expressed as a soluble protein in P.pastoris strain X-33. The purification and elution procedureswere based on the 6xHis tag carried by each protein at its C-terminus and performed under non-denaturing conditions.
2.4. Immobilization of purified recombinant proteins on CNBr-Sepharose beads
One milligram of E. coli-expressed ECD was immobilizedon 0.1 g of CNBr-activated Sepharose 4B according to theprotocol recommended by the manufacturer (GE HealthcareBio-Sciences AB). At the end of the procedure, the Sepharose–protein conjugate was stored in phosphate-buffered saline(PBS), pH 7.4, in a total volume of 12 ml. In parallel, an equalamount of bovine serum albumin (BSA) was immobilized onCNBr-Sepharose using the same experimental procedure for useas a control for the immunoadsorption experiments. The yeast-expressed ECDs were also immobilized under the sameconditions, but, for the same final volume of 12 ml, 0.1 mg ofexpressed protein was mixed with 0.9 mg of BSA andimmobilized on 0.1 g CNBr-activated Sepharose 4B, asdescribed previously (Psaridi-Linardaki et al., 2005).
2.5. Immunoadsorption of anti-nAChR autoantibodies byimmobilized ECDs
Unless otherwise specified, 120 μl of the Sepharose–ECDsuspension (containing 1 μg of yeast-expressed ECD or 10 μg ofE. coli-expressed ECD) or Sepharose–BSA suspension wasincubated for 2 h at 4 °Cwith 40 μl of PBS, 0.2%BSA containing20 fmol of anti-nAChR antibodies, followed by centrifugation. Insome experiments, as indicated in the Results, higher amounts ofanti-nAChR antibodies were used or the duration or temperatureof incubation was altered.
2.6. Radioimmunoassay (RIA)
Supernatants from the immunoadsorption assay describedabove containing unbound anti-nAChR antibodies were testedusing a commercial diagnostic Acetylcholine Receptor Autoanti-body RIA kit (RSR, UK) according to the manufacturer'sinstructions. From the 160 μl immunoadsorption mix, duplicates40 μl samples were used for the assay. Duplicate 40 μl sampleswere also tested when theMuSK autoantibody RIA kit (RSR, UK)was used. The percentage immunoadsorption in both cases wasestimated using the equation 100×{[ΔcpmBSA]− [ΔcpmECD]}/
[ΔcpmBSA], where Δcpm is the cpm of 125I-α-Btx-labellednAChR (or 125I-α-Btx-labelled MuSK) (provided in the RSR kits)precipitated by an MG serum minus that precipitated by a controlnormal human serum (NHS), and ΔcpmBSA and ΔcpmECD arethe Δcpm for samples incubated with immobilized BSA or ECD,respectively.
2.7. Removal of bound anti-nAChR autoantibodies from immobilizedECDs using low pH
The bound anti-nAChR antibodies were removed as describedpreviously (Psaridi-Linardaki et al., 2005). Briefly, Sepharosebeads containing 10 μg of α-ECD pretreated with MG serumwere washed twice with 1 ml of distilled water, resuspended indistilled water in a total volume of 100 μl and incubated for 3 minat room temperature with 100 μl of 0.2 M glycine, pH 2.4 or 2.9,then 20 μl of 1.5 M Tris buffer, pH 8.8, was added. The releasedMG autoantibodies were immediately removed by brief centri-fugation and the ECD/Sepharose immediately washed three timeswith PBS, 2% BSA and resuspended in 120 μl of PBS, 2% BSA,0.05% NaN3 until further use.
3. Results
3.1. Expression and purification of the human muscle nAChRECDs
The recombinant human muscle nAChR ECDs expressed inE. coli were mainly in the insoluble fraction and were thereforepurified under denaturing conditions (8 M urea) by affinitychromatography on a NiNTA column, taking advantage of theNH2-terminal 6xHis tag. In contrast, all the ECDs expressed inP. pastoris were in a soluble form and were purified by affinitychromatography using a C-terminus 6xHis tag, and eluted undernon-denaturing conditions (Psaridi-Linardaki et al., 2005;Kostelidou et al., 2007). As expected, all the E. coli-expressedECDs were produced at a much higher yield than the sameECDs expressed in P. pastoris (Table 1).
3.2. Immunoadsorption of anti-nAChR antibodies from MGsera using E. coli- or P. pastoris-expressed ECDs
The main target of this work was to investigate the ability of theE. coli-expressed ECDs to remove anti-nAChR autoantibodiesfrom MG sera. For this purpose, we immobilized each ECDseparately on activated CNBr-Sepharose. As a control immunoad-sorbent, we used immobilized BSA prepared under the same
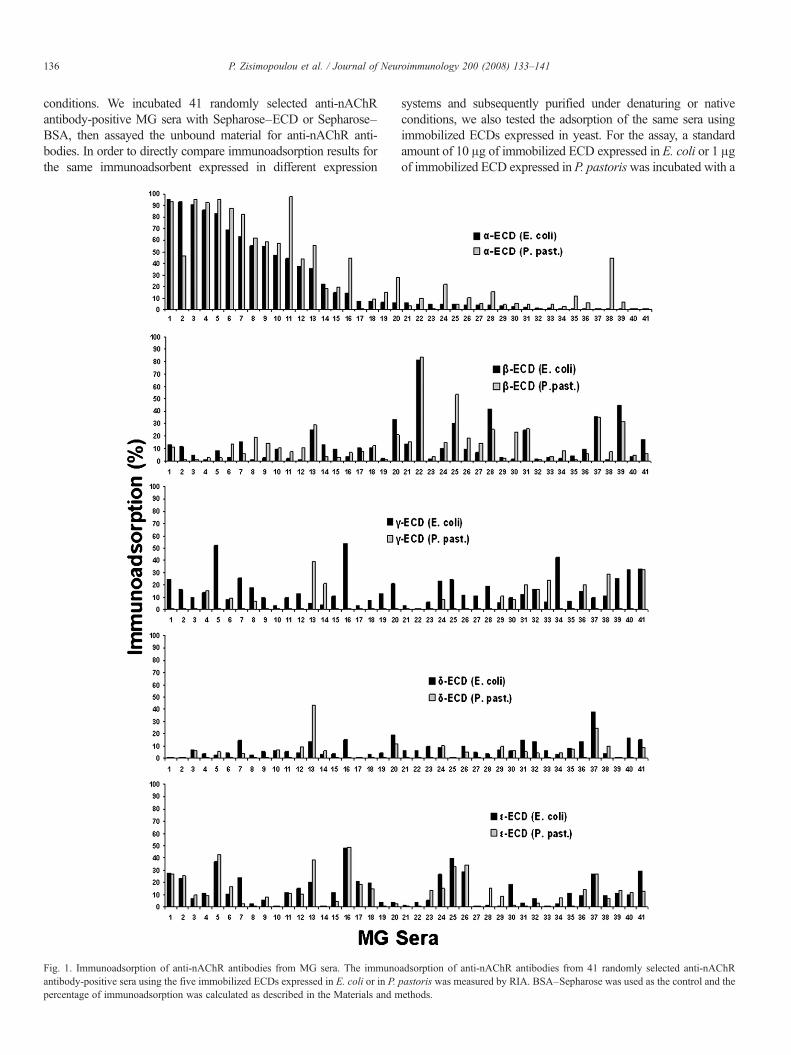
136 P. Zisimopoulou et al. / Journal of Neuroimmunology 200 (2008) 133–141
conditions. We incubated 41 randomly selected anti-nAChRantibody-positive MG sera with Sepharose–ECD or Sepharose–BSA, then assayed the unbound material for anti-nAChR anti-bodies. In order to directly compare immunoadsorption results forthe same immunoadsorbent expressed in different expression
Fig. 1. Immunoadsorption of anti-nAChR antibodies from MG sera. The immunoantibody-positive sera using the five immobilized ECDs expressed in E. coli or in P.percentage of immunoadsorption was calculated as described in the Materials and m
systems and subsequently purified under denaturing or nativeconditions, we also tested the adsorption of the same sera usingimmobilized ECDs expressed in yeast. For the assay, a standardamount of 10 μg of immobilized ECD expressed in E. coli or 1 μgof immobilized ECD expressed in P. pastoriswas incubated with a
adsorption of anti-nAChR antibodies from 41 randomly selected anti-nAChRpastoris was measured by RIA. BSA–Sepharose was used as the control and theethods.

Fig. 3. Specificity of the ECD immunoadsorbent resin. A known anti-α serum(positive control) and two anti-MuSK sera were incubated at 4 °C for 2 h withthe α-ECD resin and the corresponding supernatants were tested by RIA foranti-nAChR and anti-MuSK autoantibodies, respectively.
137P. Zisimopoulou et al. / Journal of Neuroimmunology 200 (2008) 133–141
dilution of MG serum containing 20 fmol of anti-nAChRantibodies. If the anti-nAChR antibodies in the MG serum bindto the immobilized ECDs, therewould be a reduction in the amountof antibodies in the MG sample equivalent to the percentage of thetotal antibodies in the serum reactive with this ECD. The resultsshowed that, on average, the α-, β-, γ-, δ- or ε-ECD expressed inE.coli removed 24.1%, 12.9%, 15.8%, 7.8% or 13.5% of the MGautoantibodies, whereas the corresponding ECDs expressed inyeast removed 31%, 13.9%, 7%, 5.1% and 12.7%, respectively(Fig. 1).
Interestingly, significant immunoadsorption by the δ-ECD wasrare; immunoadsorption above 20%was observed in only two sera(#13 and #37 in Fig. 1) using theP. pastoris-expressed ECD and inonly one (#37) using the E. coli-expressed ECD. In addition, wetested another 100 anti-nAChR antibody-positive MG sera twicewith the yeast-expressed δ-ECD as immunoadsorbent (data notshown), and we did not obtain any serum with immunoadsorptionhigher than 15–20%, i.e. a percentage not high enough to definitelyconfirm the presence of anti-δ antibodies in MG sera and excludethe possibility of cross-reactivity with autoantibodies against otherECDs. In all subsequent experiments, only the E. coli-expressedECDs were used.
3.3. Immunoadsorption of anti-nAChR antibodies from MGsera using a combination of all five ECDs expressed in E. coli
In addition to the immunoadsorptions by individual immobilizedECDs, a mixture of all the 5 ECD–Sepharoses was also used as acombined immunoadsorbent. The experimental procedure usedwasthe same as that for the individual ECD immunoadsorbents. Usingthe mixed ECD resins, 10 μg of each immobilized ECD wasincubated in the same reaction mixture with 20 fmol ofautoantibodies. The 20 selected sera used for this experimentbelonged to different immunoadsorption groups showing high,moderate andnodepletion of autoantibodies selected fromFig. 1.Asshown in Fig. 2, the combination of all five ECDs resulted, ingeneral, in increased immunoadsorption compared to when anindividual ECD column was used alone. For the 20 sera tested, the
Fig. 2. Use of combined immunoadsorbents. A mixture of the 5 E. coli-expressed(6th column) in parallel with individual immobilized ECDs (first 5 columns). The n
average immunoadsorption was 46.1%when the combined columnwas used, while for the α-, β-,γ-, δ- and ε-ECD resins the individualaverages were 24.2%, 11.5%, 10.9%, 6.3% and 8.1% respectively,making a sum of 61%; the difference between this and the 46.1%may be due to partial cross-reactivity of some antibodies withdifferent subunits.
3.4. Specificity of the immunoadsorbents
In order to confirm the specificity of the immunoadsorbentsderived from E. coli expression, we tested the ability of theimmobilized α-ECD to adsorb anti-nAChR or anti-MuSKantibodies. A known anti-α serum and two anti-MuSK serawere incubated with the α-ECD resin and the supernatants testedby RIA assay for anti-nAChR and anti-MuSK antibodies,respectively. If non-specific binding of anti-MuSK autoantibo-dies occurred, a reduction in the amount of anti-MuSKautoantibodies should be seen in the α-ECD-treated MG seracompared to the untreated control. As expected, no immunoad-sorption of anti-MuSK antibodies was seen (Fig. 3), confirmingthat the ECD resin was specific for anti-nAChR autoantibodies.
ECDs immobilized on Sepharose was used as a combined immunoadsorbentumbers of the sera indicate the corresponding sera in Fig. 1.
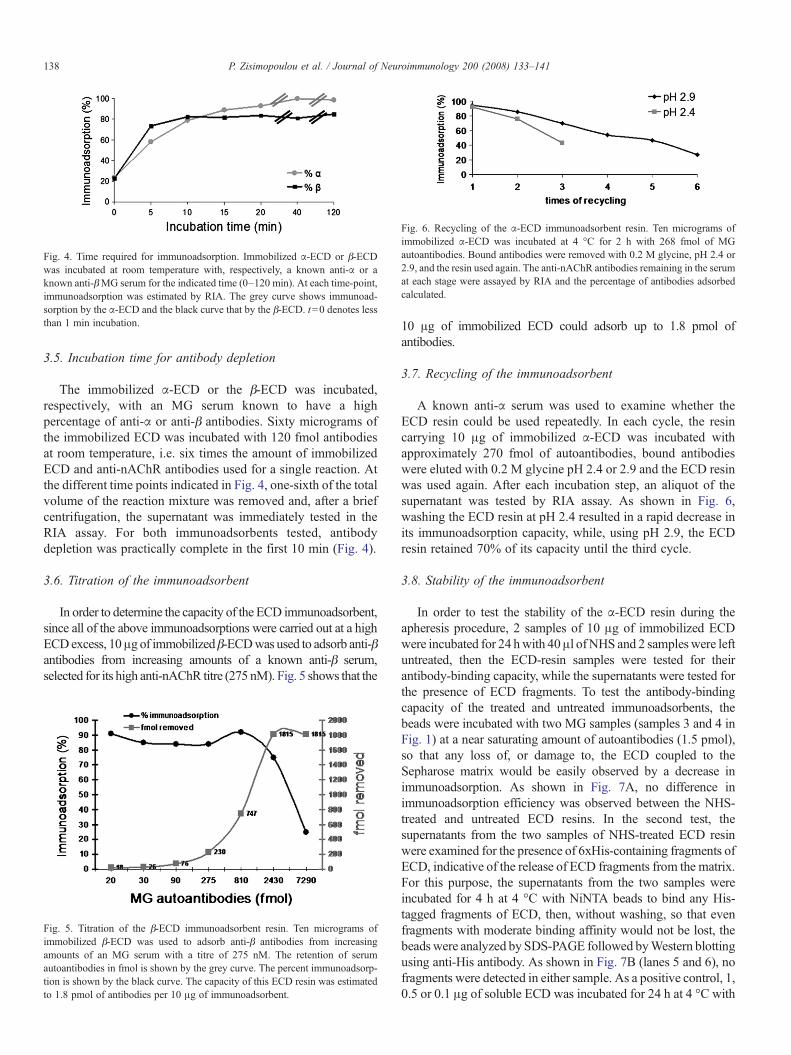
Fig. 4. Time required for immunoadsorption. Immobilized α-ECD or β-ECDwas incubated at room temperature with, respectively, a known anti-α or aknown anti-βMG serum for the indicated time (0–120 min). At each time-point,immunoadsorption was estimated by RIA. The grey curve shows immunoad-sorption by the α-ECD and the black curve that by the β-ECD. t=0 denotes lessthan 1 min incubation.
Fig. 6. Recycling of the α-ECD immunoadsorbent resin. Ten micrograms ofimmobilized α-ECD was incubated at 4 °C for 2 h with 268 fmol of MGautoantibodies. Bound antibodies were removed with 0.2 M glycine, pH 2.4 or2.9, and the resin used again. The anti-nAChR antibodies remaining in the serumat each stage were assayed by RIA and the percentage of antibodies adsorbedcalculated.
138 P. Zisimopoulou et al. / Journal of Neuroimmunology 200 (2008) 133–141
3.5. Incubation time for antibody depletion
The immobilized α-ECD or the β-ECD was incubated,respectively, with an MG serum known to have a highpercentage of anti-α or anti-β antibodies. Sixty micrograms ofthe immobilized ECD was incubated with 120 fmol antibodiesat room temperature, i.e. six times the amount of immobilizedECD and anti-nAChR antibodies used for a single reaction. Atthe different time points indicated in Fig. 4, one-sixth of the totalvolume of the reaction mixture was removed and, after a briefcentrifugation, the supernatant was immediately tested in theRIA assay. For both immunoadsorbents tested, antibodydepletion was practically complete in the first 10 min (Fig. 4).
3.6. Titration of the immunoadsorbent
In order to determine the capacity of the ECD immunoadsorbent,since all of the above immunoadsorptions were carried out at a highECDexcess, 10μg of immobilizedβ-ECDwas used to adsorb anti-βantibodies from increasing amounts of a known anti-β serum,selected for its high anti-nAChR titre (275 nM). Fig. 5 shows that the
Fig. 5. Titration of the β-ECD immunoadsorbent resin. Ten micrograms ofimmobilized β-ECD was used to adsorb anti-β antibodies from increasingamounts of an MG serum with a titre of 275 nM. The retention of serumautoantibodies in fmol is shown by the grey curve. The percent immunoadsorp-tion is shown by the black curve. The capacity of this ECD resin was estimatedto 1.8 pmol of antibodies per 10 μg of immunoadsorbent.
10 μg of immobilized ECD could adsorb up to 1.8 pmol ofantibodies.
3.7. Recycling of the immunoadsorbent
A known anti-α serum was used to examine whether theECD resin could be used repeatedly. In each cycle, the resincarrying 10 μg of immobilized α-ECD was incubated withapproximately 270 fmol of autoantibodies, bound antibodieswere eluted with 0.2 M glycine pH 2.4 or 2.9 and the ECD resinwas used again. After each incubation step, an aliquot of thesupernatant was tested by RIA assay. As shown in Fig. 6,washing the ECD resin at pH 2.4 resulted in a rapid decrease inits immunoadsorption capacity, while, using pH 2.9, the ECDresin retained 70% of its capacity until the third cycle.
3.8. Stability of the immunoadsorbent
In order to test the stability of the α-ECD resin during theapheresis procedure, 2 samples of 10 μg of immobilized ECDwere incubated for 24 hwith 40μl ofNHS and 2 sampleswere leftuntreated, then the ECD-resin samples were tested for theirantibody-binding capacity, while the supernatants were tested forthe presence of ECD fragments. To test the antibody-bindingcapacity of the treated and untreated immunoadsorbents, thebeads were incubated with two MG samples (samples 3 and 4 inFig. 1) at a near saturating amount of autoantibodies (1.5 pmol),so that any loss of, or damage to, the ECD coupled to theSepharose matrix would be easily observed by a decrease inimmunoadsorption. As shown in Fig. 7A, no difference inimmunoadsorption efficiency was observed between the NHS-treated and untreated ECD resins. In the second test, thesupernatants from the two samples of NHS-treated ECD resinwere examined for the presence of 6xHis-containing fragments ofECD, indicative of the release of ECD fragments from the matrix.For this purpose, the supernatants from the two samples wereincubated for 4 h at 4 °C with NiNTA beads to bind any His-tagged fragments of ECD, then, without washing, so that evenfragments with moderate binding affinity would not be lost, thebeads were analyzed by SDS-PAGE followed byWestern blottingusing anti-His antibody. As shown in Fig. 7B (lanes 5 and 6), nofragments were detected in either sample. As a positive control, 1,0.5 or 0.1 μg of soluble ECD was incubated for 24 h at 4 °C with
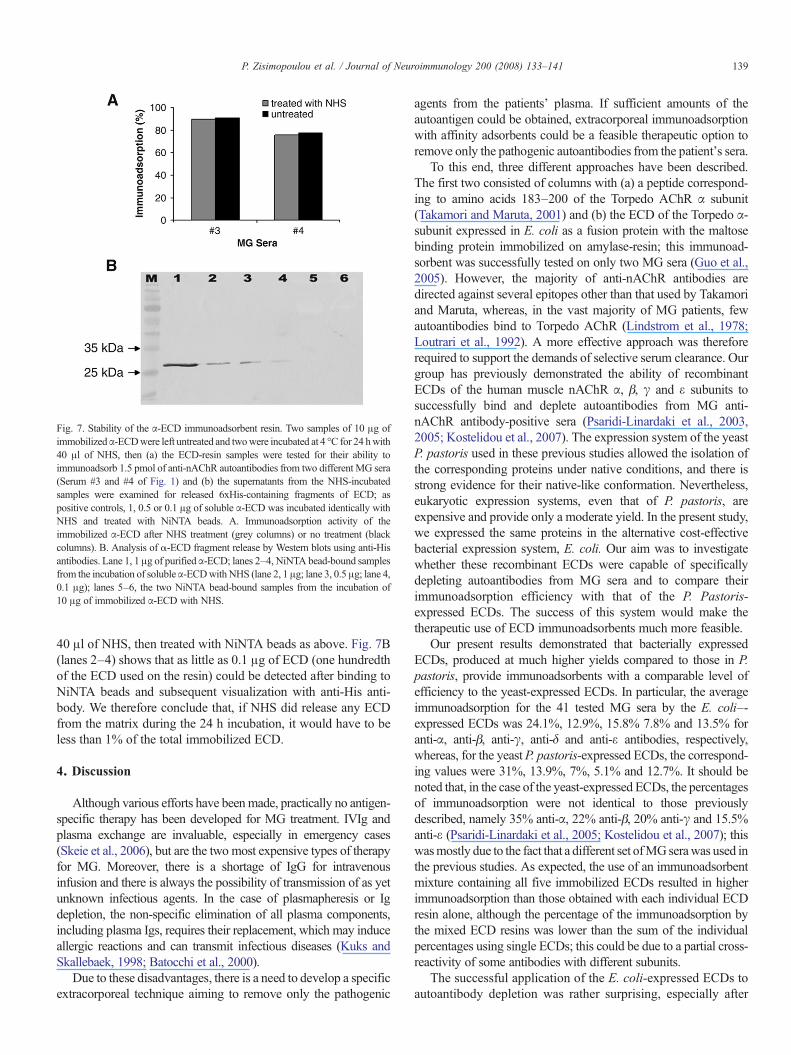
Fig. 7. Stability of the α-ECD immunoadsorbent resin. Two samples of 10 μg ofimmobilizedα-ECDwere left untreated and twowere incubated at 4 °C for 24 hwith40 μl of NHS, then (a) the ECD-resin samples were tested for their ability toimmunoadsorb 1.5 pmol of anti-nAChR autoantibodies from two different MG sera(Serum #3 and #4 of Fig. 1) and (b) the supernatants from the NHS-incubatedsamples were examined for released 6xHis-containing fragments of ECD; aspositive controls, 1, 0.5 or 0.1 μg of soluble α-ECD was incubated identically withNHS and treated with NiNTA beads. A. Immunoadsorption activity of theimmobilized α-ECD after NHS treatment (grey columns) or no treatment (blackcolumns). B. Analysis of α-ECD fragment release by Western blots using anti-Hisantibodies. Lane 1, 1 μg of purifiedα-ECD; lanes 2–4, NiNTA bead-bound samplesfrom the incubation of solubleα-ECDwithNHS (lane 2, 1 μg; lane 3, 0.5 μg; lane 4,0.1 μg); lanes 5–6, the two NiNTA bead-bound samples from the incubation of10 μg of immobilized α-ECD with NHS.
139P. Zisimopoulou et al. / Journal of Neuroimmunology 200 (2008) 133–141
40 μl of NHS, then treated with NiNTA beads as above. Fig. 7B(lanes 2–4) shows that as little as 0.1 μg of ECD (one hundredthof the ECD used on the resin) could be detected after binding toNiNTA beads and subsequent visualization with anti-His anti-body. We therefore conclude that, if NHS did release any ECDfrom the matrix during the 24 h incubation, it would have to beless than 1% of the total immobilized ECD.
4. Discussion
Although various efforts have beenmade, practically no antigen-specific therapy has been developed for MG treatment. IVIg andplasma exchange are invaluable, especially in emergency cases(Skeie et al., 2006), but are the two most expensive types of therapyfor MG. Moreover, there is a shortage of IgG for intravenousinfusion and there is always the possibility of transmission of as yetunknown infectious agents. In the case of plasmapheresis or Igdepletion, the non-specific elimination of all plasma components,including plasma Igs, requires their replacement, which may induceallergic reactions and can transmit infectious diseases (Kuks andSkallebaek, 1998; Batocchi et al., 2000).
Due to these disadvantages, there is a need to develop a specificextracorporeal technique aiming to remove only the pathogenic
agents from the patients' plasma. If sufficient amounts of theautoantigen could be obtained, extracorporeal immunoadsorptionwith affinity adsorbents could be a feasible therapeutic option toremove only the pathogenic autoantibodies from the patient's sera.
To this end, three different approaches have been described.The first two consisted of columns with (a) a peptide correspond-ing to amino acids 183–200 of the Torpedo AChR α subunit(Takamori and Maruta, 2001) and (b) the ECD of the Torpedo α-subunit expressed in E. coli as a fusion protein with the maltosebinding protein immobilized on amylase-resin; this immunoad-sorbent was successfully tested on only two MG sera (Guo et al.,2005). However, the majority of anti-nAChR antibodies aredirected against several epitopes other than that used by Takamoriand Maruta, whereas, in the vast majority of MG patients, fewautoantibodies bind to Torpedo AChR (Lindstrom et al., 1978;Loutrari et al., 1992). A more effective approach was thereforerequired to support the demands of selective serum clearance. Ourgroup has previously demonstrated the ability of recombinantECDs of the human muscle nAChR α, β, γ and ε subunits tosuccessfully bind and deplete autoantibodies from MG anti-nAChR antibody-positive sera (Psaridi-Linardaki et al., 2003,2005; Kostelidou et al., 2007). The expression system of the yeastP. pastoris used in these previous studies allowed the isolation ofthe corresponding proteins under native conditions, and there isstrong evidence for their native-like conformation. Nevertheless,eukaryotic expression systems, even that of P. pastoris, areexpensive and provide only a moderate yield. In the present study,we expressed the same proteins in the alternative cost-effectivebacterial expression system, E. coli. Our aim was to investigatewhether these recombinant ECDs were capable of specificallydepleting autoantibodies from MG sera and to compare theirimmunoadsorption efficiency with that of the P. Pastoris-expressed ECDs. The success of this system would make thetherapeutic use of ECD immunoadsorbents much more feasible.
Our present results demonstrated that bacterially expressedECDs, produced at much higher yields compared to those in P.pastoris, provide immunoadsorbents with a comparable level ofefficiency to the yeast-expressed ECDs. In particular, the averageimmunoadsorption for the 41 tested MG sera by the E. coli–-expressed ECDs was 24.1%, 12.9%, 15.8% 7.8% and 13.5% foranti-α, anti-β, anti-γ, anti-δ and anti-ε antibodies, respectively,whereas, for the yeastP. pastoris-expressed ECDs, the correspond-ing values were 31%, 13.9%, 7%, 5.1% and 12.7%. It should benoted that, in the case of the yeast-expressed ECDs, the percentagesof immunoadsorption were not identical to those previouslydescribed, namely 35% anti-α, 22% anti-β, 20% anti-γ and 15.5%anti-ε (Psaridi-Linardaki et al., 2005; Kostelidou et al., 2007); thiswasmostly due to the fact that a different set ofMGserawas used inthe previous studies. As expected, the use of an immunoadsorbentmixture containing all five immobilized ECDs resulted in higherimmunoadsorption than those obtained with each individual ECDresin alone, although the percentage of the immunoadsorption bythe mixed ECD resins was lower than the sum of the individualpercentages using single ECDs; this could be due to a partial cross-reactivity of some antibodies with different subunits.
The successful application of the E. coli-expressed ECDs toautoantibody depletion was rather surprising, especially after

140 P. Zisimopoulou et al. / Journal of Neuroimmunology 200 (2008) 133–141
comparison of the immunoadsorption results with thoseprovided by yeast-expressed immunoadsorbents. The ECDsexpressed in yeast were isolated under non-denaturing condi-tions, so that a much higher capability of antibody depletionwould be expected compared to denatured proteins. However,even though the E. coli-expressed ECDs were purified underdenaturing conditions, the immobilized ECDs were stored inphosphate buffer, which might have allowed partial refolding ofthe subunits.
The δ-ECD expressed in P. pastoris and purified under nativeconditions was in the form of microaggregates. This ECD did notresult in considerable immunoadsorption of anti-δ autoantibodiesevenwhen 100 additionalMGserawere tested. The presence of onlyone serum showing approximately 40% immunoadsorption with theδ-ECD might have several explanations, two being that (a) anti-δautoantibodiesmaybe strictly conformation-dependent and thereforetheir isolation could not be achieved due to an imperfectconformation of the recombinantδ-ECD (E. coli- or yeast-expressed)and (b) anti-δ antibodies are indeed rare in MG; furthermore, giventhat serawhich seemed to be positive for anti-δ antibodies presented asimilar degree of adsorption with other subunit ECDs, the observedimmunoadsorption might be the result of cross-reactive antibodiesrather than δ-subunit specific antibodies in MG.
Important characteristics of the immobilized ECDs expressed inE. coli that have been tested, i.e. the short time required forimmunoadsorption (less than 10 min), the ability to recycle theimmunoadsorbents, their considerable capacity and their stability tolong-term incubation with human serum, support their potential useas immunoadsorbents for MG therapy. Specifically, the fact that,even after 24 h incubation of the ECD resin with human serum, nodetectable amounts of ECD were removed from the resin (b1%)ensures that no potentially antigenic ECD molecules will bereintroduced into the patient during the much shorter therapeuticapheresis procedure. The capacity of the ECD resin (about 1.8 pmolper 10 μg of ECD used) means that, in the case of a patient with amoderate anti-nAChR antibody titre (3–6 nM), a small columnwithless than 40–80 mg of immobilized ECD would be sufficient toremove the autoantibodies from the patient's blood. Although suchamounts are currently easily produced (∼12 mg/l E. coli culture),the future use of fermentors and the establishment of optimal cultureconditions are expected to multiply the ECD yield and allow thetreatment of patients with almost any anti-nAChR titre.
Acknowledgements
The authors would like to thank Mrs. A. Kokla for excellenttechnical assistance and Dr. K. Lazaridis for the critical readingof the manuscript. This work was supported by grants from theGSRT (PENED 2003 and PEP W. Greece), the MuscularDystrophy Association of the USA (MDA USA), the Associa-tion Francaise contre les Myopathies (AFM), and the EuropeanCommission (Contract LSHM-CT-2006-037833).
References
Batocchi, A.P., Evoli, A., Di Schino, C., Tonali, P., 2000. Therapeutic apheresisin myasthenia gravis. Ther. Apher. 4, 275–279.
Conti-Fine, B.M., Milani, M., Kaminski, H.J., 2006. Myasthenia gravis: past,present, and future. J. Clin. Invest. 116, 2843–2854.
Dalakas, M.C., 2004. Intravenous immunoglobulin in autoimmune neuromus-cular diseases. Jama 291, 2367–2375.
Drachman, D.B., 1994. Myasthenia gravis. N. Engl. J. Med. 330, 1797–1810.Drachman, D.B., Fambrough, D.M., Satyamurti, S., 1973. Reduced acetylcho-
line receptors in myasthenia gravis. Trans. Am. Neurol. Ass. 98, 93–95.Fambrough, D.M., Drachman, D.B., Satyamurti, S., 1973. Neuromuscular
junction in myasthenia gravis: decreased acetylcholine receptors. Science182, 293–295.
Fostieri, E., Kostelidou, K., Poulas, K., Tzartos, S.J., 2006. Recent advances inthe understanding and therapy of myasthenia gravis. Future Neurol. 1,799–801.
Guo, C.Y., Li, Z.Y., Xu, M.Q., Yuan, J.M., 2005. Preparation of an immunoadsor-bent coupled with a recombinant antigen to remove anti-acetylcholine receptorantibodies in abnormal serum. J. Immunol. Methods 303, 142–147.
Hoch, W., McConville, J., Helms, S., Newsom-Davis, J., Melms, A., Vincent, A.,2001. Auto-antibodies to the receptor tyrosine kinase MuSK in patients withmyasthenia gravis without acetylcholine receptor antibodies. Nat. Med. 7,365–368.
Karlin, A., 2002. Emerging structure of the nicotinic acetylcholine receptors.Nat. Rev. Neurosci. 3, 102–114.
Keesey, J.C., 2004. Clinical evaluation and management of myasthenia gravis.Muscle Nerve 29, 484–505.
Kostelidou, K., Trakas, N., Zouridakis, M., Bitzopoulou, K., Sotiriadis, A.,Gavra, I., Tzartos, S.J., 2006. Expression and characterization of solubleforms of the extracellular domains of the beta, gamma and epsilon subunitsof the human muscle acetylcholine receptor. FEBS J. 273, 3557–3568.
Kostelidou, K., Trakas, N., Tzartos, S.J., 2007. Extracellular domains of thebeta, gamma and epsilon subunits of the human acetylcholine receptor asimmunoadsorbents for myasthenic autoantibodies: a combination ofimmunoadsorbents results in increased efficiency. J. Neuroimmunol.
Kuks, J.B., Skallebaek, D., 1998. Plasmapheresis in myasthenia gravis. Asurvey. Transfus. Sci. 19, 129–136.
Kuks, J.B., Limburg, P.C., Horst, G., Dijksterhuis, J., Oosterhuis, H.J., 1993.Antibodies to skeletal muscle in myasthenia gravis. Part 1. Diagnostic valuefor the detection of thymoma. J. Neurol. Sci. 119, 183–188.
Le Novere, N., Corringer, P.J., Changeux, J.P., 2002. The diversity of subunitcomposition in nAChRs: evolutionary origins, physiologic and pharmaco-logic consequences. J. Neurobiol. 53, 447–456.
Lindstrom, J., 1985. Immunobiology of myasthenia gravis, experimentalautoimmune myasthenia gravis, and Lambert-Eaton syndrome. Annu. Rev.Immunol. 3, 109–131.
Lindstrom, J., Campbell, M., Nave, B., 1978. Specificities of antibodies toacetylcholine receptors. Muscle Nerve 1, 140–145.
Lindstrom, J., Einarson, B., Tzartos, S., 1981. Production and assay of antibodiesto acetylcholine receptors. Methods Enzymol. 74 (Pt C), 432–460.
Lindstrom, J.M., 2000. Acetylcholine receptors and myasthenia. Muscle Nerve23, 453–477.
Lindstrom, J.M., Engel, A.G., Seybold, M.E., Lennon, V.A., Lambert, E.H.,1976a. Pathological mechanisms in experimental autoimmune myastheniagravis. II. Passive transfer of experimental autoimmune myasthenia gravis inrats with anti-acetylcholine receptor antibodies. J. Exp. Med. 144, 739–753.
Lindstrom, J.M., Seybold, M.E., Lennon, V.A., Whittingham, S., Duane, D.D.,1976b. Antibody to acetylcholine receptor in myasthenia gravis. Prevalence,clinical correlates, and diagnostic value. Neurology. 26, 1054–1059.
Loutrari, H., Tzartos, S.J., Claudio, T., 1992. Use of Torpedo-mouse hybridacetylcholine receptors reveals immunodominance of the alpha subunit inmyasthenia gravis antisera. Eur. J. Immunol. 22, 2949–2956.
Massey, J.M., 1997. Acquired myasthenia gravis. Neurol. Clin. 15, 577–595.Newsom-Davis, J., Wilson, S.G., Vincent, A., Ward, C.D., 1979. Long-term
effects of repeated plasma exchange in myasthenia gravis. Lancet 1, 464–468.Patrick, J., Lindstrom, J., 1973. Autoimmune response to acetylcholine receptor.
Science 180, 871–872.Poulas, K., Tsibri, E., Kokla, A., Papanastasiou, D., Tsouloufis, T., Marinou, M.,
Tsantili, P., Papapetropoulos, T., Tzartos, S.J., 2001. Epidemiology ofseropositive myasthenia gravis in Greece. J. Neurol. Neurosurg. Psychiatry71, 352–356.

141P. Zisimopoulou et al. / Journal of Neuroimmunology 200 (2008) 133–141
Psaridi-Linardaki, L., Mamalaki, A., Remoundos, M., Tzartos, S.J., 2002.Expression of soluble ligand- and antibody-binding extracellular domain ofhuman muscle acetylcholine receptor alpha subunit in yeast Pichia pastoris.Role of glycosylation in alpha-bungarotoxin binding. J. Biol. Chem. 277,26980–26986.
Psaridi-Linardaki, L., Mamalaki, A., Tzartos, S.J., 2003. Future therapeuticstrategies in autoimmune myasthenia gravis. Ann. N. Y. Acad. Sci. 998,539–548.
Psaridi-Linardaki, L., Trakas, N., Mamalaki, A., Tzartos, S.J., 2005. Specificimmunoadsorption of the autoantibodies from myasthenic patients using theextracellular domain of the human muscle acetylcholine receptor alpha-subunit. Development of an antigen-specific therapeutic strategy. J. Neuroim-munol. 159, 183–191.
Richman, D.P., Agius, M.A., 2003. Treatment of autoimmune myastheniagravis. Neurology 61, 1652–1661.
Robertson, N.P., Deans, J., Compston, D.A., 1998.Myasthenia gravis: a populationbased epidemiological study in Cambridgeshire, England. J. Neurol.Neurosurg. Psychiatry 65, 492–496.
Romi, F., Skeie, G.O., Aarli, J.A., Gilhus, N.E., 2000. Muscle autoantibodies insubgroups of myasthenia gravis patients. J. Neurol. 247, 369–375.
Sideris, S., Lagoumintzis, G., Kordas, G., Kostelidou, K., Sotiriadis, A., Poulas,K., Tzartos, S.J., 2007. Isolation and functional characterization of anti-acetylcholine receptor subunit-specific autoantibodies from myasthenicpatients: receptor loss in cell culture. J. Neuroimmunol. 189, 111–117.
Sieb, J.P., 2005. Myasthenia gravis: emerging new therapy options. Curr. Opin.Pharmacol. 5, 303–307.
Skeie, G.O., Apostolski, S., Evoli, A., Gilhus, N.E., Hart, I.K., Harms, L.,Hilton-Jones, D., Melms, A., Verschuuren, J., Horge, H.W., 2006. Guide-lines for the treatment of autoimmune neuromuscular transmission disorders.Eur. J. Neurol. 13, 691–699.
Takamori, M., Maruta, T., 2001. Immunoadsorption in myasthenia gravis basedon specific ligands mimicking the immunogenic sites of the acetylcholinereceptor. Ther. Apher. 5, 340–350.
Tsouloufis, T., Mamalaki, A., Remoundos, M., Tzartos, S.J., 2000. Reconstitutionof conformationally dependent epitopes on the N-terminal extracellulardomain of the humanmuscle acetylcholine receptor alpha subunit expressed inEscherichia coli: implications for myasthenia gravis therapeutic approaches.Int. Immunol. 12, 1255–1265.
Tzartos, S.J., Seybold, M.E., Lindstrom, J.M., 1982. Specificities of antibodiesto acetylcholine receptors in sera from myasthenia gravis patients measuredby monoclonal antibodies. Proc. Natl. Acad. Sci. U. S. A. 79, 188–192.
Tzartos, S.J., Barkas, T., Cung, M.T., Mamalaki, A., Marraud, M., Orlewski, P.,Papanastasiou, D., Sakarellos, C., Sakarellos-Daitsiotis, M., Tsantili, P.,Tsikaris, V., 1998. Anatomy of the antigenic structure of a large membraneautoantigen, the muscle-type nicotinic acetylcholine receptor. Immunol.Rev. 163, 89–120.
Unwin, N., 2005. Refined structure of the nicotinic acetylcholine receptor at 4Aresolution. J. Mol. Biol. 346, 967–989.
Vincent, A., 2002. Unravelling the pathogenesis of myasthenia gravis. Nat. Rev.Immunol. 2, 797–804.
Vincent, A., Palace, J., Hilton-Jones, D., 2001. Myasthenia gravis. Lancet 357,2122–2128.
Copyright © 2022 FDOKUMEN




















