
Dissociation of Steroid Receptor Coactivator 1 and Nuclear Receptor Corepressor Recruitment to the...
15
Dissociation of Steroid Receptor Coactivator 1 and Nuclear Receptor Corepressor Recruitment to the Human Glucocorticoid Receptor by Modification of the Ligand-Receptor Interface: The Role of Tyrosine 735 ADAM STEVENS, HELEN GARSIDE, ANDREW BERRY, CHARLOTTE WATERS, ANNE WHITE, AND DAVID RAY Endocrine Sciences Research Group, Faculty of Medicine (A.S., H.G., A.B., C.W., A.W., D.R.) and School of Biological Sciences (C.W., A.W.), Stopford Building, University of Manchester, Manchester M13 9PT, United Kingdom Within the human glucocorticoid receptor (GR) ste- roid binding pocket, tyrosine 735 makes hydrophobic contact with the steroid D ring. Substitution of ty- rosine735 selectively impairs glucocorticoid transac- tivation but not transrepression. We now show, using both mammalian two-hybrid and glutathione-S- transferase pull downs, that such substitutions re- duce interaction with steroid receptor coactivator 1, both basally and in response to agonist binding. Us- ing a yeast two-hybrid screen we identified one of the three nuclear receptor interacting domains (NCoR- N1) of nuclear receptor corepressor (NCoR) as inter- acting with the GR C terminus in an RU486-specific manner. This was confirmed in mammalian two- hybrid experiments, and so we used the NCoR-N1 peptide to probe the GR C-terminal conformation. Substitution of Tyr735phe, Tyr735val, and Tyr735 ser, which impaired steroid receptor coactivator 1 (SRC1) interaction, enhanced NCoR-N1 recruitment, basally and after RU486. RU486 did not direct SRC1 recruit- ment to any of the GR constructs, and dexametha- sone did not allow NCoR-N1 recruitment. Using a glutathione-S-transferase pull-down approach, the NCoR-N1 peptide was found to bind the full-length GR constitutively, and no further induction was seen with RU486, but it was reduced by dexamethasone. As both SRC1 and NCoR are predicted to recognize a common hydrophobic cleft in the GR, it seems that changes favorable to one interaction are detrimen- tal to the other, thus identifying a molecular switch. (Molecular Endocrinology 17: 845–859, 2003) T HE GLUCOCORTICOID RECEPTOR (GR) transac- tivates target genes by translocating from the cy- toplasm to the nucleus, dimerizing, and binding to specific glucocorticoid response elements. The GR has two major transactivation domains that bridge to the preinitiation complex by recruiting a family of co- activator proteins including the GR interacting protein 1 and steroid receptor coactivator 1 (SRC1; Refs. 1–5). The two transactivation domains may function inde- pendently, but within the full-length GR may also in- teract in a cooperative manner (6–8). Modified glucocorticoid ligands, which retain either transactivation or transrepression activity (dissociating ligands), provide an interesting insight on receptor function (9–11). The GR is capable of binding these ligands with high affinity, and the activated receptor is capable of nuclear translocation. It is likely that there are signals encoded within the ligand that act through recognition motifs within the ligand binding pocket to generate a new protein interacting surface on the re- ceptor to regulate macromolecule complex formation. Understanding the nature of this switch has major implications for steroid biology and glucocorticoid therapy of inflammatory disease. Nuclear receptor corepressor (NCoR) and silencing mediator of retinoid and thyroid hormone receptor (SMRT) are structurally related proteins originally iden- tified as important mediators of gene repression by unliganded heterodimeric nuclear receptors (12, 13). More recently it was shown that steroid hormone an- tagonists recruit the two corepressors (12, 14, 15). The amount of residual, partial agonist activity of the an- tagonists was shown to be dependent on the expres- sion level of cofactors (15–19). The best characterized GR antagonist is RU486, and RU486 promotes inter- action between the GR and the receptor interacting domains of NCoR (15). These authors identified ligand-independent interaction between the GR N ter- minal and NCoR and also found that both dexameth- asone and RU486 recruited NCoR to the GR C termi- nus. This latter finding is surprising as it might be Abbreviations: AF, Activation function; CMV, Cytomegalo- virus; GR, glucocorticoid receptor; GST, glutathione-S-trans- ferase; LBD, ligand-binding domain; MMTV-luc, mouse mammary tumor virus-luciferase; NCoR, nuclear receptor corepressor; PPAR, peroxisomal proliferator-activated re- ceptor-; SMRT, silencing mediator of retinoid and thyroid hormone receptor; SRC, steroid receptor coactivator; TIF2, transcriptional intermediary factor; wtGR, wild-type GR. 0888-8809/03/$15.00/0 Molecular Endocrinology 17(5):845–859 Printed in U.S.A. Copyright © 2003 by The Endocrine Society doi: 10.1210/me.2002-0320 845
-
Upload
manchester-us -
Category
Documents
-
view
0 -
download
0
Transcript of Dissociation of Steroid Receptor Coactivator 1 and Nuclear Receptor Corepressor Recruitment to the...

Dissociation of Steroid Receptor Coactivator 1 andNuclear Receptor Corepressor Recruitment to theHuman Glucocorticoid Receptor by Modificationof the Ligand-Receptor Interface: The Role ofTyrosine 735
ADAM STEVENS, HELEN GARSIDE, ANDREW BERRY, CHARLOTTE WATERS, ANNE WHITE, AND
DAVID RAY
Endocrine Sciences Research Group, Faculty of Medicine (A.S., H.G., A.B., C.W., A.W., D.R.) andSchool of Biological Sciences (C.W., A.W.), Stopford Building, University of Manchester, ManchesterM13 9PT, United Kingdom
Within the human glucocorticoid receptor (GR) ste-roid binding pocket, tyrosine 735 makes hydrophobiccontact with the steroid D ring. Substitution of ty-rosine735 selectively impairs glucocorticoid transac-tivation but not transrepression. We now show, usingboth mammalian two-hybrid and glutathione-S-transferase pull downs, that such substitutions re-duce interaction with steroid receptor coactivator 1,both basally and in response to agonist binding. Us-ing a yeast two-hybrid screen we identified one of thethree nuclear receptor interacting domains (NCoR-N1) of nuclear receptor corepressor (NCoR) as inter-acting with the GR C terminus in an RU486-specificmanner. This was confirmed in mammalian two-hybrid experiments, and so we used the NCoR-N1peptide to probe the GR C-terminal conformation.
Substitution of Tyr735phe, Tyr735val, and Tyr735 ser,which impaired steroid receptor coactivator 1 (SRC1)interaction, enhanced NCoR-N1 recruitment, basallyand after RU486. RU486 did not direct SRC1 recruit-ment to any of the GR constructs, and dexametha-sone did not allow NCoR-N1 recruitment. Using aglutathione-S-transferase pull-down approach, theNCoR-N1 peptide was found to bind the full-lengthGR constitutively, and no further induction was seenwith RU486, but it was reduced by dexamethasone.As both SRC1 and NCoR are predicted to recognizea common hydrophobic cleft in the GR, it seems thatchanges favorable to one interaction are detrimen-tal to the other, thus identifying a molecular switch.(Molecular Endocrinology 17: 845–859, 2003)
THE GLUCOCORTICOID RECEPTOR (GR) transac-tivates target genes by translocating from the cy-
toplasm to the nucleus, dimerizing, and binding tospecific glucocorticoid response elements. The GRhas two major transactivation domains that bridge tothe preinitiation complex by recruiting a family of co-activator proteins including the GR interacting protein1 and steroid receptor coactivator 1 (SRC1; Refs. 1–5).The two transactivation domains may function inde-pendently, but within the full-length GR may also in-teract in a cooperative manner (6–8).
Modified glucocorticoid ligands, which retain eithertransactivation or transrepression activity (dissociatingligands), provide an interesting insight on receptorfunction (9–11). The GR is capable of binding theseligands with high affinity, and the activated receptor iscapable of nuclear translocation. It is likely that there
are signals encoded within the ligand that act throughrecognition motifs within the ligand binding pocket togenerate a new protein interacting surface on the re-ceptor to regulate macromolecule complex formation.Understanding the nature of this switch has majorimplications for steroid biology and glucocorticoidtherapy of inflammatory disease.
Nuclear receptor corepressor (NCoR) and silencingmediator of retinoid and thyroid hormone receptor(SMRT) are structurally related proteins originally iden-tified as important mediators of gene repression byunliganded heterodimeric nuclear receptors (12, 13).More recently it was shown that steroid hormone an-tagonists recruit the two corepressors (12, 14, 15). Theamount of residual, partial agonist activity of the an-tagonists was shown to be dependent on the expres-sion level of cofactors (15–19). The best characterizedGR antagonist is RU486, and RU486 promotes inter-action between the GR and the receptor interactingdomains of NCoR (15). These authors identifiedligand-independent interaction between the GR N ter-minal and NCoR and also found that both dexameth-asone and RU486 recruited NCoR to the GR C termi-nus. This latter finding is surprising as it might be
Abbreviations: AF, Activation function; CMV, Cytomegalo-virus; GR, glucocorticoid receptor; GST, glutathione-S-trans-ferase; LBD, ligand-binding domain; MMTV-luc, mousemammary tumor virus-luciferase; NCoR, nuclear receptorcorepressor; PPAR�, peroxisomal proliferator-activated re-ceptor-�; SMRT, silencing mediator of retinoid and thyroidhormone receptor; SRC, steroid receptor coactivator; TIF2,transcriptional intermediary factor; wtGR, wild-type GR.
0888-8809/03/$15.00/0 Molecular Endocrinology 17(5):845–859Printed in U.S.A. Copyright © 2003 by The Endocrine Society
doi: 10.1210/me.2002-0320
845

expected that RU486 binding would generate anRU486-specific binding signature. The authors alsoidentified multiple NCoR motifs, N1, N2, and N3, ascapable of interacting with the GR, with evidence ofcooperative binding (15). Interestingly, overexpressionof SMRT has been shown to alter the dexamethasonedose response in transfected cells (17), suggesting adegree of functional interaction between the corepres-sor and the agonist-liganded GR.
By model building we have previously identifiedTyr735 within the human GR as a residue contributingto the ligand binding cleft and forming a hydrophobicinteraction with the C16 group of the steroid D ring ofdexamethasone (20), confirmed by the subsequentlypublished crystal structure (21). Mutation of ty-rosine735, especially to serine, results in selective lossof transactivation with a relative preservation of ligandbinding affinity and RelA repression (20).
We now use a panel of Tyr735-mutated GR mole-cules to dissect the interaction between ligand andreceptor, and also post ligand-binding associationwith coregulatory proteins. Loss of transactivation bymutation to Tyr735 is accompanied by loss of p160coactivator recruitment. However, none of the Tyr735mutations allow recruitment of NCoR-N1 in responseto dexamethasone. Those mutations that impair p160recruitment, Tyr735phe, Tyr735val, and Tyr735 ser,result in enhanced NCoR-N1 recruitment in responseto RU486.
RESULTS
The Effect of D Ring-Substituted Steroids onTransactivation by Tyr735-Mutated GR
The ligand specificity of the Tyr735-mutated GR mol-ecules for transactivation was explored using a tran-sient transfection approach with full-length GR, andthe mouse mammary tumor virus-luciferase (MMTV-luc) reporter gene. We have previously shown thatmutations at position 735 result in reduced transacti-vation in response to dexamethasone, but relativepreservation of transrepression (20). From our model,and the crystal structure, tyrosine at 735 was pre-dicted to interact with the steroid D ring, and espe-cially with the 16-methyl group present on the D ring ofdexamethasone. Hydrocortisone lacks this methylgroup, and therefore we examined its activity on theMMTV template (Fig. 1B). In fact, loss of the terminalhydroxyl group of the tyrosine, by mutation to phenyl-alanine, did not alter the relative potency of hydrocor-tisone compared with dexamethasone (Fig. 1, A andB). Loss of the 735 side-chain aromatic ring, bychange to valine, did reduce the relative potency ofhydrocortisone compared with dexamethasone (Fig.1, A and B) and when the small, polar amino acidserine was introduced at GR735, hydrocortisone hadbarely detectable activity up to 1000 nM (Fig. 1B).RU24858 has been reported to reduce transactivation
activity compared with dexamethasone (11), but wefound it caused significant, GR-dependent transacti-vation on wild-type GR (wtGR) (Fig. 1C). RU24858,with a 16-methyl group, had the same rank order ofpotency across the Tyr735-mutated GR molecules asdexamethasone, in contrast to the effect seen withhydrocortisone (Fig. 1B).
Progesterone is a low affinity competitive antagonistat the GR, and we examined progesterone for illegiti-mate agonist activity, but found none (data notshown).
RU486 Antagonism and Partial Agonist ActivityThrough Tyr735-Substituted GR
RU486 is a high-affinity GR antagonist, and we foundthat it efficiently antagonized dexamethasone transac-tivation on all the GR molecules examined (Fig. 2A).Furthermore, RU486 had significant, partial agonistactivity on all the GR molecules analyzed (Fig. 2B), upto 5% of dexamethasone on the wtGR.
Interactions of Human (h)NCoR-N1 Recruitmentto the wtGR Ligand-Binding Domain (LBD)by RU486
We developed a yeast two-hybrid assay to analyze li-gand-dependent interactions between the GR LBD andcorepressor NCoR. Using this approach we found astrong interaction between the human NCoR first nuclearinteraction domain (N1: amino acids 2263–2272) and thewtGR LBD in the presence of 10 �M RU486 (Fig. 3A).There was no ligand-independent interaction, and nointeraction seen with 10 �M dexamethasone (Fig. 3A).The strength of this interaction was further quantitatedusing liquid cultures (Fig. 3B). We also found a strongdexamethasone-dependent interaction between thesame GR construct and the receptor interacting domain(RID) of human SRC1 (SRC1-RID; amino acids 569-821)in the same assays (data not shown). There was noRU486-dependent interaction between the GR andSRC1 (data not shown).
Recruitment of SRC1 and hNCoR-N1 to the wtGRLBD by Dexamethasone and RU486: MammalianTwo-Hybrid Analysis
We also developed a mammalian two-hybrid assay toanalyze ligand-dependent interactions between the GRLBD and cofactor proteins. We found that RU486 (10 nM)promoted interaction between hNCoR-N1 and the wtGRLBD in COS 7 cells (Fig. 4A). Dexamethasone did notallow the hNCoR-N1/GR interaction (Fig. 4A). In contrast,RU486 did not promote recruitment of SRC-RID to thewtGR LBD, but SRC1-RID was recruited by 10 nM dexa-methasone (Fig. 4B). These effects were specific as therewas no interaction seen with the empty Gal4 vector(Fig. 4C).
846 Mol Endocrinol, May 2003, 17(5):845–859 Stevens et al. • Tyrosine 735 and Cofactor Recruitment to the GR

Recruitment of SRC1 and NCoR to Full-LengthwtGR by Dexamethasone and RU486:Glutathione-S-Transferase (GST)Pull-Down Assay
A GST pull-down approach was used to further charac-terize the interactions between the wtGR and bothSRC1-RID and hNCoR-N1. Initial experiments confirmedexpression of full-length wtGR protein (98 kDa) andshowed that there was negligible interaction with GSTprotein (Fig. 5A). GST-SRC1-RID pulled down little more
GR than the GST alone, but 10 �M dexamethasonecaused a 23-fold induction in the amount of GR pulleddown by GST-SRC1-RID (P � 0.001; Fig. 5B), whereasRU486 caused no increase in GR/SRC1-RID interaction(Fig. 5, A and B). In contrast, there was a significant basalinteraction between GST-hNCoR-N1 and the GR, com-pared with the GST control (Fig. 5A). RU486 caused nofurther increase, but dexamethasone inhibited the GST-NCoR-N1/GR interaction by more than 50% (P � 0.035)(Fig. 5B).
Fig. 1. Steroid Transactivation through Tyr735-Mutated GRFull-length wild-type (TYR), Tyr735phe (PHE), Tyr735val (VAL), and Tyr735ser (SER) GR expression vectors (all in pcDNA3), were
cotransfected with the MMTV-luc reporter gene, and CMV-Renilla into COS 7 cells as described, and were divided post transfectioninto treatment and control groups: dexamethasone (A); hydrocortisone (B); RU24858 (C). All experiments were performed on threeoccasions with similar results. The cells were incubated with 0, 10, 100, 1000, or 10000 nM steroid as indicated for 16 h before harvestand both luciferase and Renilla assay. Luciferase results were corrected for Renilla. Results are expressed as fold induction over thevehicle control. Bars indicate the SEM (n � 3). Steroid structures: dexamethasone (A) and hydrocortisone (B) are full agonists at the GR;RU24858 (C) has been reported to have reduced transactivation, but preserved transrepression activity (11). The components of thebasic ring structure are labeled (A–D) on the dexamethasone structure and carbon atoms C16, C17, and C21 are numbered.
Stevens et al. • Tyrosine 735 and Cofactor Recruitment to the GR Mol Endocrinol, May 2003, 17(5):845–859 847

Effects of GRTyr735 Mutation on SRC1-RIDRecruitment
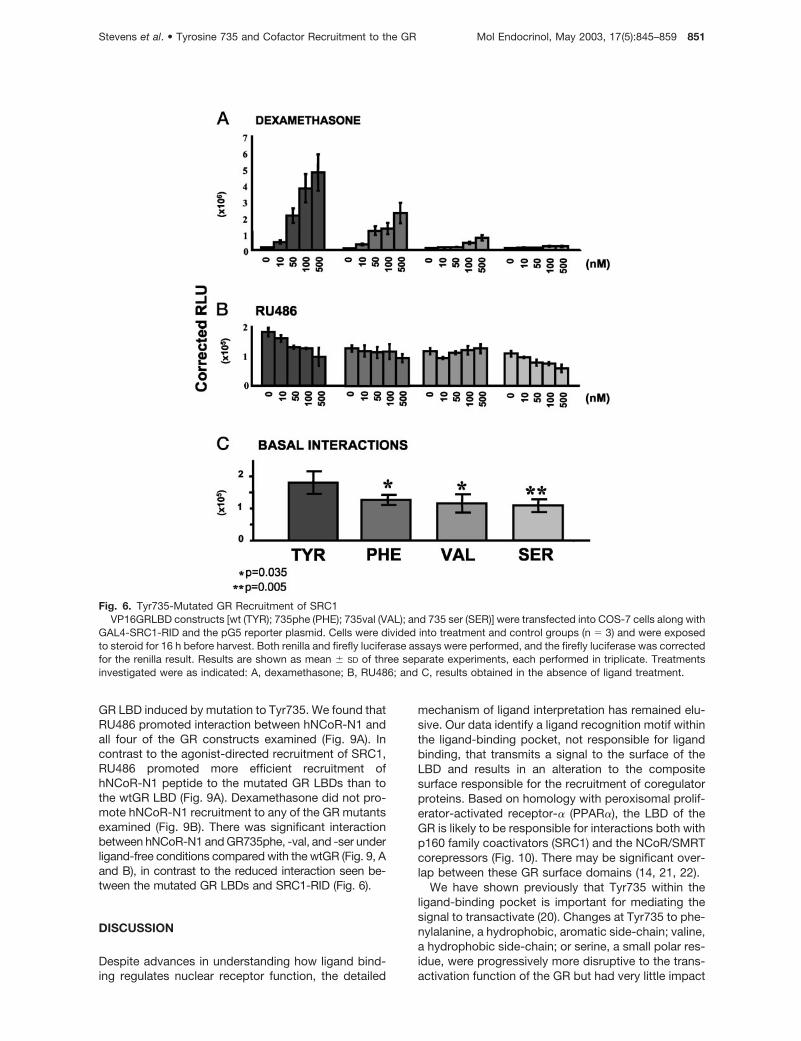
We used the validated mammalian two-hybrid assayto analyze the effects of mutation to the importantTyr735 residue within the GR LBD on recruitment ofthe SRC1-RID. We found that mutation of the Tyr735to phenylalanine reduced the recruitment of SRC1-RID. More disruptive mutation to valine further atten-uated this interaction, and we were unable to showany interaction between the Tyr735 ser-mutated GRLBD and the SRC1-RID (Fig. 6A).
RU486 had partial agonist activity on all the GRmolecules examined (Fig. 2B). Therefore, we exam-ined RU486-dependent recruitment of SRC1-RID tothe mutated GR LBDs. There was no ligand-depen-dent interaction seen with any of the mutated GR LBD
constructs or with the wtGR LBD (Fig. 6B). We alsoexamined the effects of progesterone, but again foundno recruitment of SRC1-RID to the GR LBDs up to 500nM concentration (data not shown).
There was reduced ligand-independent interactionwith SRC1-RID for both the GR735phe (0.75-fold)and GR735val (0.67-fold) mutants (P � 0.035). TheGR735ser had the lowest basal interaction (0.64-fold;P � 0.005; Fig. 6C).
Analysis of Ligand-Dependent Recruitment ofSRC1-RID to the Tyr735-Mutated Full-Length GRMolecules: GST Pull-Down Assay
We also used our validated GST pull-down assay tomeasure changes in GR/SRC1 interaction induced by
Fig. 2. RU486 Antagonism and Partial Agonist Activity through Tyr735-Mutated GRWild-type (TYR), Tyr735phe (PHE), Tyr735val (VAL), and Tyr735ser (SER), were all coexpressed with the MMTV-luc reporter
gene, and CMV-gal in COS 7 cells as described, and were divided into treatment and control groups. All experiments wereperformed on three occasions with similar results. A, Cells were divided and incubated with 100 nM dexamethasone and theindicated concentrations of RU486 for 16 h before harvest and reporter gene assay. Bars indicate the mean and SD (n � 3). Resultsare expressed relative to maximal induction caused by 100 nM dexamethasone. B, Cells were divided and incubated with theindicated concentrations of RU486 for 16 h before harvest and reporter gene assay. Bars indicate the mean and SD (n � 3). Resultsare expressed relative to maximal induction caused by 100 nM dexamethasone.
848 Mol Endocrinol, May 2003, 17(5):845–859 Stevens et al. • Tyrosine 735 and Cofactor Recruitment to the GR

ligand binding. As shown above, the in vitro interactionbetween these two proteins is specific to agonistligands.
We examined the absolute amounts of recruitedfull-length GR to the GST-SRC1-RID, by quantitatingthe radioactivity pulled down relative to input. Usingthis approach, we found there was no significant dif-ference in basal SRC1-RID recruitment between thewtGR and the mutated GR molecules (n � 4; P �0.412; Fig. 7B). Dexamethasone caused a significantincrease in SRC1-RID recruitment to the wild type(TYR; P � 0.005), Tyr735phe (PHE) GR (P � 0.024),and Tyr735val (VAL) GR (P � 0.017). The Tyr735 ser(SER) showed no significant increase over basal inter-action. RU486 did not induce SRC1 recruitment to anyof the GR molecules examined (data not presented).All four GR molecules showed greater binding to theGST-SRC1 than to the GST-only control in the ab-
sence of dexamethasone, but this barely reached sig-nificance (TYR, P � 0.045; PHE, P � 0.048; VAL, P �0.021; SER, P � 0.05).
Mutation of Tyr735 (TYR) in full-length GR to phenyl-alanine (PHE) or to valine (VAL) significantly reduced the
Fig. 3. Yeast Two-Hybrid Analysis of Human NCoR-N1 In-teraction with the GR
A, The p8oplacz, pLexAGR, and hNCoR-AD were sequen-tially transformed into EGY48 S. cereviseae, and cultured onplates lacking histidine, uracil, and tryptophan. Once colonieswere established (3–4 d growth), they were transferred toinduction plates lacking leucine and containing galactose/raffinose, X-Gal, with or without 10 �M ligand, and eitherdexamethasone (DEX) or RU486. B, Overnight cultures ofEGY48 containing the three plasmids were used to inoculateinduction media containing either dimethylsulfoxide vehicle(NL), dexamethasone 50 �m (DEX), or RU486 50 �m (RU486).After a 3-h incubation, the cells were lysed for the �-galassay. �-gal units were calculated as described in Materialsand Methods. Results represent average values from threeindependent yeast transformations. Data are presented asmean and SD (n � 3).
Fig. 4. Mammalian Two-Hybrid Analysis of GR LBD Recruit-ment of SRC1-RID and hNCoR-N1
The GAL4 and VP16 constructs along with a luciferasereporter (pG5luc) were transfected into COS-7 cells. Cellswere divided into treatment and control groups (n � 3) andwere exposed to steroids for 18 h before harvest and fireflyand renilla luciferase assays. Results, as corrected relativelight units, are shown as mean � SD. A, VP16wtGR LBD andGAL4 hNCoR. B, VP16wtGR LBD and GAL4 SRC1. C,VP16wtGR LBD and an empty GAL4 construct.
Stevens et al. • Tyrosine 735 and Cofactor Recruitment to the GR Mol Endocrinol, May 2003, 17(5):845–859 849

induction of SRC1-RID recruitment obtained with 10 �M
dexamethasone. Tyr735 ser (SER) showed no ligand-dependent recruitment of SRC1 (Fig. 7C).
Effects of GR Tyr735 Mutations on Coactivationby SRC1
The mammalian two-hybrid analysis suggests that mu-tation of Tyr735 reduces recruitment of SRC1. In-vitroanalysis also shows a reduced dexamethasone depen-dent recruitment of SRC1 to the mutant Tyr735 mole-cules. Therefore, we measured the effect of over ex-pressing full-length SRC1 on transactivation of the
MMTV-luc reporter by the various GR molecules. wtGR(TYR) shows a marked enhancement of dexamethasonetransactivation when SRC1 is expressed (Fig. 8), and theeffect is also seen with the Tyr735phe (PHE). In contrastthe effect on transactivation by Tyr735val (VAL) is moremodest, and there was minimal impact on transactiva-tion by Tyr735 ser (SER; Fig. 8).
Effects of GR Tyr735 Mutations on NCoR-N1Recruitment
We used our validated mammalian two-hybrid assayto measure changes in hNCoR-N1 interaction with the
Fig. 5. In Vitro Interaction of wtGR and Either SRC1-RID or hNCoR-N1A, Full-length 35S-labeled GR was synthesized in vitro using the pcDNA3GR plasmid as template. The GR was incubated with
GST-SRC1-RID or GST-hNCoR-N1 fusion protein with 10 �M dexamethasone, 10 �M RU486, or no treatment. The GST waspulled down and the complexes resolved on an SDS-PAGE gel. The 35S-labeled GR bound to the GST fusion proteins wasvisualized by phosphoimaging. B, The amount of radioactive GR pulled down by GST-SRC1-RID and GST-hNCoR-N1 fusionswas determined by using a phosphoimager. The mean and SD values of three separate experiments are presented, andcomparisons were performed using a t test.
850 Mol Endocrinol, May 2003, 17(5):845–859 Stevens et al. • Tyrosine 735 and Cofactor Recruitment to the GR
GR LBD induced by mutation to Tyr735. We found thatRU486 promoted interaction between hNCoR-N1 andall four of the GR constructs examined (Fig. 9A). Incontrast to the agonist-directed recruitment of SRC1,RU486 promoted more efficient recruitment ofhNCoR-N1 peptide to the mutated GR LBDs than tothe wtGR LBD (Fig. 9A). Dexamethasone did not pro-mote hNCoR-N1 recruitment to any of the GR mutantsexamined (Fig. 9B). There was significant interactionbetween hNCoR-N1 and GR735phe, -val, and -ser underligand-free conditions compared with the wtGR (Fig. 9, Aand B), in contrast to the reduced interaction seen be-tween the mutated GR LBDs and SRC1-RID (Fig. 6).
DISCUSSION
Despite advances in understanding how ligand bind-ing regulates nuclear receptor function, the detailed
mechanism of ligand interpretation has remained elu-sive. Our data identify a ligand recognition motif withinthe ligand-binding pocket, not responsible for ligandbinding, that transmits a signal to the surface of theLBD and results in an alteration to the compositesurface responsible for the recruitment of coregulatorproteins. Based on homology with peroxisomal prolif-erator-activated receptor-� (PPAR�), the LBD of theGR is likely to be responsible for interactions both withp160 family coactivators (SRC1) and the NCoR/SMRTcorepressors (Fig. 10). There may be significant over-lap between these GR surface domains (14, 21, 22).
We have shown previously that Tyr735 within theligand-binding pocket is important for mediating thesignal to transactivate (20). Changes at Tyr735 to phe-nylalanine, a hydrophobic, aromatic side-chain; valine,a hydrophobic side-chain; or serine, a small polar res-idue, were progressively more disruptive to the trans-activation function of the GR but had very little impact
Fig. 6. Tyr735-Mutated GR Recruitment of SRC1VP16GRLBD constructs [wt (TYR); 735phe (PHE); 735val (VAL); and 735 ser (SER)] were transfected into COS-7 cells along with
GAL4-SRC1-RID and the pG5 reporter plasmid. Cells were divided into treatment and control groups (n � 3) and were exposedto steroid for 16 h before harvest. Both renilla and firefly luciferase assays were performed, and the firefly luciferase was correctedfor the renilla result. Results are shown as mean � SD of three separate experiments, each performed in triplicate. Treatmentsinvestigated were as indicated: A, dexamethasone; B, RU486; and C, results obtained in the absence of ligand treatment.
Stevens et al. • Tyrosine 735 and Cofactor Recruitment to the GR Mol Endocrinol, May 2003, 17(5):845–859 851

on ligand binding affinity or transrepression of nuclearfactor �B (NF�B) RelA (20). Therefore, we concludedthat Tyr735 was important for interpretation of ligand.
Our model of dexamethasone binding to the LBD ofthe GR suggested an interaction with the 16-methylgroup on the steroid D ring (Fig. 1A), and the crystalstructure confirms this (21). Therefore, we comparedthe relative activities of an agonist steroid with themethyl group, dexamethasone, with one that lackedthis group, hydrocortisone (Fig. 1, A and B). We alsoexamined the activity of the partial agonist RU24858
(Fig. 1C), the low-affinity GR antagonist progesterone,and the high-affinity antagonist RU486 (Fig. 2).Tyr735phe did not alter the relative potency of hydro-cortisone compared with dexamethasone, but Tyr735-val and Tyr735ser showed disproportionate reduc-tions in hydrocortisone potency, indicating theimportance of the Tyr735 interaction with the steroid Dring. Previously Lind et al. have noted that substitutionof methionine at 560, which lies close to the Tyr735within the GR ligand-binding pocket (21), has a differ-ential impact on transactivation by synthetic glucocor-
Fig. 7. In Vitro Interaction of Tyr735-Mutated GR and SRC-1A, Full-length 35S-labeled GR, wt (TYR), 735phe (PHE), 735val (VAL), and 735 ser (SER) were incubated with a GST-SRC1-RID
fusion protein with or without 10 �M dexamethasone. The GST was pulled down and the complexes resolved on an SDS-PAGEgel. The 35S-labeled GR bound to the GST-SRC1-RID fusion protein was visualized by phosphoimaging. Staining with Sypro redquantitated GST fusion proteins to confirm equal loading. Image analysis was performed under UV illumination. B, Thepulled-down GR activity was quantitated using the phosphoimager, and the activity present in the GST-alone band wassubtracted from each of the other lanes in each experiment. The GR activity was then expressed as a proportion of input GR, tocontrol for differences in the efficiency of synthesis between experiments. There was no significant difference between theunliganded interactions. The mean and SD of four separate experiments are presented. *, P � 0.017; **, P � 0.024; and ***, P �0.005 comparisons by t test. C, The induction of GR interaction with SRC1 was determined by phosphoimager, and the meanand SD values of five separate experiments are presented. Differences from the induction seen with the wtGR were significant:*, P � 0.007; and **, P � 0.003; comparisons by ANOVA and post hoc Bonferroni t test.
852 Mol Endocrinol, May 2003, 17(5):845–859 Stevens et al. • Tyrosine 735 and Cofactor Recruitment to the GR

ticoids (23), again confirming the functional interactionbetween this portion of the ligand binding pocket andthe steroid D ring. RU24858, which also has a 16-methyl group, had significant agonist activity on all thefour constructs examined and showed the same rankorder of potency as dexamethasone. This again sug-gests the functional interaction between Tyr735 andthe 16-methyl group of some synthetic glucocorti-coids. RU486 had weak agonist activity on all the fourGR molecules, and progesterone had no agonist ac-tivity on any of the GR constructs tested.
To investigate the consequences of modification tothe ligand/receptor interface for cofactor recruitment,we established a series of protein interaction assays.RU486, in common with other steroid receptor antag-onists, leads to recruitment of corepressor proteins(15, 16, 24–26) and, in our assays, is both an efficientantagonist of dexamethasone transactivation and apartial agonist. Interestingly, NCoR appears to interactwith multiple domains of the GR (15). We initiallyscreened the NCoR-N1 domain using a yeast two-hybrid approach for steroid-dependent interactionwith the GR C terminus (Fig. 3). In contrast to the invitro analysis of Schulz et al. (15), we found significantrecruitment of NCoR-N1 to the GR C terminus in anRU486-specific manner. It was previously found that
including multiple copies of the NCoR N domains en-hanced the interaction with the GR C terminus using aGST pull-down approach, but that this effect was seenusing both dexamethasone and RU486 as ligands (15).This suggests that there is a loss of ligand specificityin protein interaction, possibly because of cooperativebinding of multiple GR molecules to the NCoR peptidein a manner similar to that between retinoic acid re-ceptor/retinoic X receptor and NCoR (13). However,we found that the NCoR-N1 peptide can act as aprobe for the RU486-specific conformation of the GRC terminus. This was further examined using a mam-malian two-hybrid approach, and the same pattern ofRU486-specific interaction between GR LBD andNCoR-N1 was seen.
Because the studies in yeast and mammalian cellshad shown a distinct pattern of ligand-dependenthNCoR-N1/GR and SRC1-RID/GR interaction, theproteins were also analyzed in vitro. There was a sig-nificant interaction between the GR and hNCoR-N1 inthe absence of ligand. RU486 did not further enhancethis interaction, but as the proportion of input GRpulled down by hNCoR-N1 was approximately 50% ofthat pulled down by SRC1 in the presence of dexa-methasone, the interaction appears significant. Stud-ies using dexamethasone show greater than 50% re-
Fig. 8. Effect on Overexpression of SRC-1 on the Ability of Tyr735-Mutated GR to TransactivateUsing COS 7 cells the wild-type (TYR), Tyr735phe (PHE), Tyr735val (VAL), and Tyr735 ser (SER) full-length GR expression
vectors were all expressed with the MMTV-luc reporter gene, CMV-renilla, and either the pSG5-SRC1 expression vector (solidline) or the pSG5 empty vector (broken line). The cells were divided into treatment and control groups, and experiments wereperformed on three occasions. Cells were divided and incubated with 0, 1, 3, 10, 30, 100, 300, and 1000 nM dexamethasone for16 h before harvest and reporter gene assay. Bars indicate the mean and SEM (n � 3). Results are expressed as fold inductionover the maximal response to dexamethasone seen in the absence of overexpressed SRC1. Nonlinear regression was performedusing the SIMFIT package (Bardsley, W., University of Manchester).
Stevens et al. • Tyrosine 735 and Cofactor Recruitment to the GR Mol Endocrinol, May 2003, 17(5):845–859 853

duction in hNCoR-N1/GR interactions. This patternof ligand-dependent dissociation of corepressors(NCoR) from nuclear receptors has been well definedfor the heterodimeric nuclear receptor family (12, 13)and, under cell-free conditions, appears to be pre-served for the GR.
The two-hybrid analysis suggests that antagonist isrequired for NCoR recruitment to GR, whereas theGST pull-down assay suggests that interaction is pos-sible in the absence of ligand, and that antagonistadds little to the interaction. In intact cells the unligan-ded GR is sequestered in the cytoplasm, and is, there-fore, in a separate compartment from the nuclear lo-cated corepressors. The GR expression vector usedfor the mammalian two-hybrid analysis directs pro-teins to the nucleus by virtue of a powerful nuclearlocalization signal (27). Indeed, we have shown byimmunohistochemistry that all the GR-VP16 con-structs are expressed and are located in the nucleus inthe absence of ligand (data not shown). Other inves-tigators have also described discrepant results whenthey compared mammalian two-hybrid against GSTpull-down assays when examining steroid receptorinteractions with NCoR proteins (16); as a result, ad-
ditional controls on steroid receptor interactions withcorepressors in living cells have been proposed (16).
Using both mammalian two-hybrid and GST pull-down approaches the recruitment of SRC1 to the GRwas seen to be reduced by the Tyr735phe variant andreduced further by the Tyr735val; there was no signif-icant induction seen when the Tyr735ser variant wasexamined. These results are concordant with thosepresented previously in which the rank order of po-tency for transactivation was wild type � Tyr735phe �Tyr735val � Tyr735 ser (Fig. 1; Ref. 20).
If the Tyr735 mutations reduce SRC1 recruitment tothe GR, then coactivation by SRC1 would be expectedto be impaired also. In fact there was no observeddifference in SRC1 coactivation between the wtGR(Tyr) and the Tyr735phe (PHE). But both Tyr735val(VAL) and Tyr735 ser (SER) showed markedly reducedSRC1 coactivation (Fig. 8). This coactivation assay didnot distinguish between the wtGR and the Tyr735phe,probably because the full-length SRC1 construct usedsynergistically interacted with both GR activation func-tion 1 and 2 (AF1 and AF2) domains (28). More severedisruption of Tyr735 (to Val or Ser) presumably re-duced the AF1/AF2 recruitment of full-length SRC1.
Fig. 9. Interaction of Tyr735-Mutated GR with the Corepressor NCoRThe VP16GR LBD wild-type and mutated constructs were transfected into COS-7 cells along with Gal4-hNCoR-N1 and the
pG5 reporter plasmid. Cells were divided into treatment and control groups (n � 3), and were exposed to RU486 (A) ordexamethasone (B) for 16 h before harvest. Transfection efficiency was corrected for by using renilla luciferase. Results are shownas mean � SD of one experiment performed in triplicate, representative of three separate experiments.
854 Mol Endocrinol, May 2003, 17(5):845–859 Stevens et al. • Tyrosine 735 and Cofactor Recruitment to the GR

This provides additional evidence for a Tyr735 effecton SRC family coactivator recruitment to the agonist-liganded human GR.
RU486 did not promote SRC-1 recruitment to thewtGR, nor to any of the three Tyr735-substituted GRvariants, either in vitro or in vivo. Therefore, it doesnot appear that the partial agonist activity of RU486
could result from aberrant recruitment of SRC pro-teins. Others have shown that RU486 does not leadto recruitment of SRC1 to the GR in mammalian cellsand have suggested that the partial agonist activityof RU486 observed in some cell types results frominteraction of the GR with chromatin remodelingproteins (18).
Fig. 10. Comparison of Nuclear Receptor Cofactor Interaction DomainsA, Sequence alignment of GR LBD helices 3,3� and 4 with PPAR�. These helices make up the hydrophobic coactivator
interaction pocket of the GR (21). This domain also contains residues that are conserved from PPAR� and that have been foundto make contacts with the SMRT-N1 corepressor helix in the presence of antagonist. The PPAR� residue in red is predicted tobe an H bond acceptor for binding to the SMRT-N1 nuclear receptor interacting motif (14), and residues highlighted in blue areH bond donors for interaction with the SMRT motif (14). Homologous residues within the GR are similarly highlighted. B, Sequencealignment of cofactor motifs in NCoR and SRC1 and TIF2. The extended �-helical motif of the NCoR helix is predicted to replacethe binding site of helix 12. The R�6 of the N1 domain corresponds to the R�2 in the N3 domain of TIF2 that forms a hydrogenbond with D590 of the GR. C, Coordination of cofactor motifs by the GR. Upper panel, GR coordination of the coactivator helixN3. The GR helices are shown in green, with helix 12 in red. Charge clamp residues are indicated. The TIF2-N3 coactivator helixis marked in lime green. The GR coordinates the TIF2-N3 helix between E755, K579, and also between R585 and D590 (21). Lowerpanel, Putative GR coordination of corepressor NCoR helix N1. The E�2 of SMRT N1 hydrogen bonds to K310 of PPAR� andR�6 hydrogen bonds to N303 of PPAR�. These residues are homologous in the GR and are indicated Q597 and D590,respectively. K292 of PPAR� coordinates the �-helix of SMRT N1 and is conserved as the K579 in GR that coordinates the �-helixof TIF2-N3.
Stevens et al. • Tyrosine 735 and Cofactor Recruitment to the GR Mol Endocrinol, May 2003, 17(5):845–859 855

There was basal interaction between the wtGR Cterminus and SRC1 seen in the mammalian two-hybrid, but not in the GST pull-down, assays. This wasmade clear by the reduction in interaction seen whenthe Tyr735 was mutated. This implies that there arefunctional interactions between the unliganded GRand SRC1 in living cells, although the compartmental-ization of GR in the cytoplasm in the absence of ligandmay reduce the physiological importance of this phe-nomenon. However, there is evidence of nuclear traf-ficking of the unliganded GR, and even transient inter-actions with coactivators may be sufficient to promotechanges in gene expression (19, 29).
As RU486 is capable of promoting interaction be-tween the GR and NCoR, and as NCoR and SRCfamily coactivators are predicted to recognize a con-served hydrophobic cleft made up of helices 3, 4, and5, we sought to define the effect of changes at Tyr735on corepressor recruitment (13, 14, 21). We used theNCoR-N1 motif both because we found that it inter-acted with the GR C terminus in an antagonist-specificmanner, and also because the interactions of theclosely related SMRT-N1 with PPAR� have been an-alyzed in detail (Fig. 10; Ref. 14). There is conservationof the critical charge clamp residues that in PPAR�anchor the corepressor helix (Fig. 10). Strikingly, mu-tations to Tyr735 that impair SRC1 recruitment in re-sponse to dexamethasone enhance recruitment ofNCoR-N1 in response to RU486 (Fig. 9). We haveshown that the RU486-dependent interaction betweenGR and NCoR-N1 only occurs in living cells, eitheryeast or mammalian, and that GST pull-down ap-proaches show the basal interaction between the twomolecules only. This may indicate an important role forother proteins or factors in mediating or stabilizingprotein interaction. Such discrepancies between invivo and in vitro analyses have been reported before(16). It is also relevant that the unliganded GR interac-tion with NCoR may not readily occur in living cells dueto compartmental segregation. Therefore we did notextend these studies into GST pull-down assays.
It is clear that mutation of Tyr735 increases thebasal, unliganded interaction between NCoR-N1 andthe GR C terminus. This is in contrast to the reductionin SRC1 interaction seen under the same conditions,with the same mutated GR molecules. This supportsthe existence of a functional overlap between the pro-tein motifs responsible for interacting with coactiva-tors in the presence of agonists, and corepressors inthe presence of antagonists, as has been proposed forthe heterodimeric nuclear receptors (14). By homologywith PPAR�, we have mapped candidate residueswithin the GR interacting with the NCoR peptide, andthese overlie the motif found to bind to the transcrip-tional intermediary factor 2 (TIF2) LxxLL peptide (21)(Fig. 10). We also show that mutation to Tyr735 suffi-cient to reduce transactivation does not allow aberrantrecruitment of corepressor to the GR in response todexamethasone binding, but rather enhances binding
to the NCoR-N1 repressor peptide in response toRU486 binding.
Glucocorticoid transactivation is served by recruit-ment of coactivators to both AF1 and AF2 of theligand-bound, DNA-anchored GR. It is interesting thatthe GR C terminus has a cryptic surface capable ofinteracting with the potent corepressor NCoR eitherwhen unmasked by binding to RU486 or under basal,unliganded conditions. Coactivator recruitment to theGR can be selectively reduced by mutations to theLBD that retain high-affinity binding of agonists. Thesetransactivation-defective GR molecules have en-hanced recruitment of corepressors both basally andin response to antagonists, but not agonists. Suchenhanced recruitment of NCoR might contribute toreduced transactivation (17). Therefore, Tyr735 ap-pears to transmit a signal from the ligand to the sur-face of the GR, as measured by alterations in cofactorrecruitment. However, the most conservative changeto the tyrosine (Tyr735phe) did not show the predictedreduction in SRC1 coactivation, which suggests thatrecruitment of SRC1 is not the controlling factor for GRtransactivation. The more severe disruptions to thetyrosine at 735 (Tyr735val, and Tyr735 ser), whichresult in loss of the aromatic ring and introduction of asmall polar side chain, respectively, do result in con-cordant reductions in SRC1 recruitment and the pre-dicted loss of SRC1 coactivation. Therefore, our datasuggest that Tyr735 is an important amino acid fordetermining the final GR conformation after ligandbinding, and contributes to final GR activity, but is notthe sole determining element of cofactor recruitment.As both the SRC coactivators and NCoR/SMRT arepredicted to recognize a common hydrophobic cleft inthe GR, it seems likely that changes favorable to oneinteraction are detrimental to the other, i.e. a molecularswitch.
MATERIALS AND METHODS
Plasmids
The MMTV-luc and pcDNA3GR (both wild-type and Tyr735mutated; Tyr735phe, Tyr735val, and Tyr735 ser) plasmidshave been previously described (20, 30). The cytomegalovi-rus (CMV)-�gal plasmid has been previously described (20,31). The pSG5-SRC1 expression vector has been previouslydescribed and was a kind gift from Dr. Michael Stallcup (32).CMV-Renilla was from Promega Corp. (Southampton, UK).
Gal-4 fusion constructs were made using the pBIND vector(Promega Corp.). The HF2 PCR kit (CLONTECH Laboratories,Inc., Oxford, UK) was used to amplify DNA coding for aminoacid positions 569-821 of the human SRC1 gene, whichincludes the receptor interacting domain, RID), and 2262–2440 of the human NCoR gene, including the N1 motif, togive GAL4 SRC1 and GAL4 hNCoR. The specific primers hadBamHI and NotI restriction enzyme sites added for ligationinto pBIND (SRC1: forward primer, 5�-ggatcccacctagcagatta-aatatacaaccag-3�; reverse primer, 5�-gcggccgcagcgtggg-cagtaactgatc-3�; hNCoR: forward primer, 5�-ggatccttgggct-ggaagacattatc-3�; reverse primer, 5�-gcggccgctcagtcatca-ctatccgac-3�).
856 Mol Endocrinol, May 2003, 17(5):845–859 Stevens et al. • Tyrosine 735 and Cofactor Recruitment to the GR

The B42 activation domain fusion protein expression plas-mids were made by inserting the NCoR and SRC1 genefragments described above into pB42AD (CLONTECHLaboratories, Inc.) to give hNCoR-AD, and SRC1-AD,respectively.
The human GR LBD between codons 523 and 777 wasamplified using the HF2 PCR kit and inserted into the pACTplasmid (Promega Corp.) to give VP16wtGRLBD. TheVP16GR735phe, VP16GR735val, and VP16GR35ser plas-mids were made by amplifying the GR LBD from the appro-priate pcDNA3GR constructs detailed above. The specificprimers had BamHI and NotI restriction enzyme sites addedfor ligation into the vector (forward primer for LBD, 5�-ggatc-ccaacgttaccacaactcacc-3�; forward primer for full-length, 5�-ggatcctggactccaaagaatcattaactc-3�; reverse primer, 5�-gcg-gccgccttttgatgaaacagaagttttttg-3�).
The same GR LBD gene fragment described above wasinserted into pLexA (CLONTECH Laboratories, Inc.) to giveLexAGR.
A GSThNCoR-N1 fusion protein expression plasmid wasmade by inserting DNA coding for amino acid positions2262–2440 of the hNCoR, including the N1 motif, gene intothe pGEX-5X-3 vector (Amersham Pharmacia Biotech, Buck-inghamshire, UK). This fragment was amplified using the HF2PCR kit (CLONTECH Laboratories, Inc.). The specific primershad SalI and NotI restriction enzyme sites added for ligationinto the vector (NCoR: forward primer, 5�-gtcgaccttgggctg-gaagacattatc-3�; reverse primer, 5�-gcggccgctcagtcatcac-tatccgac-3�).
The GST/SRC1-RID plasmid (encoding 569-821) was akind gift from Dr. Malcolm Parker (33). The pG5luc containsfive copies of an upstream activator sequence upstream ofluciferase (Promega Corp.). The yeast reporter gene wasp8OP-lacZ (CLONTECH Laboratories, Inc.).
All plasmid constructs were sequenced to confirm thepresence of the predicted changes and to exclude introduc-tion of errors.
Transfection
COS 7 cells (obtained from ECACC:87021302), were culturedin DMEM with Glutamax (Life Technologies, Inc., Paisley,UK), and 10% fetal calf serum. All transfections were per-formed using lipofectamine plus (Life Technologies, Inc.),according to the manufacturers’ instructions, as previouslydescribed (20). Cells were divided into 24-well plates aftertransfection, and were treated with steroid for 16 h beforeharvest. All experiments were performed in triplicate and onat least three separate occasions with similar results.
Transactivation Assay
COS7 cells were seeded at 5 � 105 per 10-cm tissue cultureplate and were transfected with 2 �g MMTV-luc, and 1 �gpcDNA3-GR (either wild-type or the three Tyr735-mutatedGR cDNAs), and 1 �g CMV-�Gal, to control for transfectionefficiency (20). Cell lysates were subjected to both luciferaseassay and �-galactosidase assay as previously described(20). Luciferase was corrected for �-galactosidase activity aspreviously described (34).
Coactivation Assay
COS7 cells were seeded at 5 � 105 per 10-cm tissue cultureplate and were transfected with 3 �g MMTV-luc, 0.12 �gpcDNA3-GR (wild-type or the three Tyr735-mutated GRcDNAs), either 0.6 �g pSG5-SRC-1 or 0.6 �g of the emptypSG5 vector, and 1 �g CMV-Renilla. Firefly luciferase andRenilla luciferase assays were performed on cell extractsfrom the same experiment using the dual-luciferase reporterassay system (Promega Corp.) according to the manufactur-
er’s instructions. All firefly luciferase results were normalizedusing the Renilla luciferase to control for differences in trans-fection efficiency.
Yeast Two-Hybrid Analysis
Saccharomyces cerevisae (EGY48) were sequentially trans-formed with reporter, bait, and then prey. The bait wasLexAGR, and the prey was SRC1-AD or hNCoR-AD. Trans-formations were performed using a small-scale lithium ace-tate procedure (35).
The transfected yeast cells were replica plated onto syn-thetic dropout/Gal/Raf plates with X-gal, containing the ap-propriate treatment (ligand at 50 �M and dimethylsulfoxidevehicle as the no-ligand control).
Suspension cultures were used to quantitate the interac-tion. Briefly, overnight cultures were used to inoculate liquidSD/Gal/Raf with or without ligands and incubated until mid-log phase (OD600 0.5–0.8). The cells were then washed andresuspended in buffer 1 [100 mM HEPES, 155 mM NaCl, BSA1% (wt/vol), Tween-20, 0.05%]. This mixture was subjectedto three freeze-thaw cycles using liquid nitrogen. Buffer 2(2.23 mM chloro phenol red-�-D-galactopyranoside in buffer1) was added to the mixture, the color was allowed to de-velop, and the OD578 was recorded. The �-Gal units werethen calculated using the following equation: �-Gal units �1000 � OD578/(t � V � OD600) [t � elapsed time (in min) ofincubation, v � 0.1 � concentration factor, OD600 � A600 of1 ml of culture]. One �-Gal unit was defined as the amountthat hydrolyses 1 �mol of chlorophenol red-�-D-galacto-pyranoside to chlorophenol red and D-galactose per min percell (36).
Mammalian Two-Hybrid Analysis
COS 7 cells (5 � 105 per 10-cm plate) were transfected with5 �g of each of the VP16/GR LBD, either Gal4/SRC1, orGal4/NCoR and the pG5luc reporter plasmid. Firefly lucif-erase and Renilla luciferase assays were performed on cellextracts from the same experiment using the dual-luciferasereporter assay system following the manufacturer’s instruc-tions. All firefly luciferase results were normalized using theRenilla luciferase to control for differences in transfectionefficiency.
Interaction of SRC-1 and GRs in Vitro
GST and GST-SRC1 or GST-NCoR-N1 fusion proteins wereexpressed in Escherichia coli strain DH5� (Promega Corp.)and were purified as described (37). Briefly, a 50 ml culture ofthe expression vector was stimulated with 0.5 mM isopropyl-�-D-thiogalactopyranoside (Sigma, Poole, UK) and grown onfor 3 h. The bacteria were then pelleted and resuspended inNETN buffer (20 mM Tris, pH 8.0; 100 mM NaCl; 1 mM EDTA;0.01% Nonidet P-40) supplemented with protease inhibitors(Complete, Roche Molecular Biochemicals, Mannheim, Ger-many). The bacteria were treated with lysozyme (100 �g/ml)for 15 min at 4 C and then disrupted using a 50 W, 20 kHzSonicator (Jencons, Leighton Buzzard, UK) on full power for5 � 30 sec. The sample was then centrifuged to remove thedebris and glutathione-sepharose beads (Amersham Phar-macia Biotech) were added to the supernatant. The fusionproteins were allowed to bind to the beads for 1 h at 4 C. Thebeads were then washed three times in NETN plus proteaseinhibitors and stored at 4 C.
35S-labeled GR was synthesized using the TNT kit (Pro-mega Corp.) following the manufacturer’s instructions withthe full-length, human GR cDNA in pcDNA3 as template,[35S]methionine (Amersham Pharmacia Biotech), and T7polymerase. Labeled GR was incubated with the GST pro-teins bound to glutathione-sepharose beads for 1 h at 4 C.
Stevens et al. • Tyrosine 735 and Cofactor Recruitment to the GR Mol Endocrinol, May 2003, 17(5):845–859 857

The beads were then washed three times in NETN, andboiling in 2� SDS-PAGE sample buffer eluted the boundproteins. Proteins were analyzed by SDS-PAGE, and the gelwas stained in Sypro red to visualize the GST fusion proteins.The 35S-labeled GR was then visualized by exposure of thegel to a phosphoimaging plate (BasIII, Fuji Photo Film Co.,Ltd.).
The Sypro red-stained acrylamide gels were analyzed un-der UV light (Alphaimager 2000, Flowgen, Lichfield, UK). Theamount of radiolabeled protein present on the gel was quan-titated using a phosphoimager (FujiFilm, London, UK;BAS1800) and Aida 2.0 analysis software (Raytest, GMBH,Straubenhardt, Germany).
Acknowledgments
We thank Malcolm Parker for the GST-SRC-1 construct;Michael Stallcup and V. Krishna Chatterjee for helpful dis-cussions; Johnson Liu for the gift of human NCoR cDNA; Ms.Jo Soden for technical help and advice; Hinrich Gronemeyerfor the gift of RU24858; and C.-S. Suen and Andrew Brass forhelp with molecular modeling.
Received September 12, 2002. Accepted January 21,2003.
Address all correspondence and requests for reprints to:Dr. Adam Stevens, Endocrine Sciences Research Group,Stopford Building, University of Manchester, Oxford Road,Manchester M13 9PT, United Kingdom. E-mail: [email protected].
This work was supported by a Glaxo-Wellcome Fellowship(to D.W.R.), Biotechnology and Biological Sciences ResearchCouncil Cooperative Awards in Science and Engineeringawards (to H.G. and C.W.), and Central Manchester HealthCare Trust.
REFERENCES
1. Evans RM 1988 The steroid and thyroid hormone recep-tor superfamily. Science 240:889–895
2. Horwitz KB, Jackson TA, Bain DL, Richer JK, TakimotoGS, Tung L 1996 Nuclear receptor coactivators and core-pressors. Mol Endocrinol 10:1167–1177
3. Mangelsdorf DJ, Thummel C, Beato M, Herrlich P,Schutz G, Umesono K, Blumberg B, Kastner P, Mark M,Chambon P 1995 The nuclear receptor superfamily: thesecond decade. Cell 83:835–839
4. Perlmann T, Evans RM 1997 Nuclear receptors in Sicily:all in the famiglia. Cell 90:391–397
5. Tsai MJ, O’Malley BW 1994 Molecular mechanisms ofaction of steroid/thyroid receptor superfamily members.Annu Rev Biochem 63:451–486
6. Hittelman AB, Burakov D, Iniguez-Lluhi JA, Freedman LP,Garabedian MJ 1999 Differential regulation of glucocor-ticoid receptor transcriptional activation via AF-1-asso-ciated proteins. EMBO J 18:5380–5388
7. Hollenberg SM, Giguere V, Evans RM 1989 Identificationof two regions of the human glucocorticoid receptor hor-mone binding domain that block activation. Cancer Res49:2292s–2294s
8. Hollenberg SM, Evans RM 1988 Multiple and cooperativetrans-activation domains of the human glucocorticoidreceptor. Cell 55:899–906
9. Heck S, Bender K, Kullmann M, Gottlicher M, Herrlich P,Cato AC 1997 I �B �-independent downregulation ofNF-�B activity by glucocorticoid receptor. EMBO J 16:4698–4707
10. Vanden Berghe W, Francesconi E, De Bosscher K,Resche-Rigon M, Haegeman G 1999 Dissociated glu-cocorticoids with anti-inflammatory potential repress in-terleukin-6 gene expression by a nuclear factor-�B-dependent mechanism. Mol Pharmacol 56:797–806
11. Vayssiere BM, Dupont S, Choquart A, Petit F, Garcia T,Marchandeau C, Gronemeyer H, Resche-Rigon M 1997Synthetic glucocorticoids that dissociate transactivationand AP-1 transrepression exhibit antiinflammatory activ-ity in vivo. Mol Endocrinol 11:1245–1255
12. Yen PM 2001 Physiological and molecular basis of thy-roid hormone action. Physiol Rev 81:1097–1142
13. Perissi V, Staszewski LM, McInerney EM, Kurokawa R,Krones A, Rose DW, Lambert MH, Milburn MV, GlassCK, Rosenfeld MG 1999 Molecular determinants of nu-clear receptor-corepressor interaction. Genes Dev 13:3198–3208
14. Xu HE, Stanley TB, Montana VG, Lambert MH, ShearerBG, Cobb JE, McKee DD, Galardi CM, Plunket KD, NolteRT, Parks DJ, Moore JT, Kliewer SA, Willson TM, Stim-mel JB 2002 Structural basis for antagonist-mediatedrecruitment of nuclear co-repressors by PPAR�. Nature415:813–817
15. Schulz M, Eggert M, Baniahmad A, Dostert A, Heinzel T,Renkawitz R 2002 RU486-induced glucocorticoid recep-tor agonism is controlled by the receptor N terminus andby corepressor binding. J Biol Chem 277:26238–26243
16. Lavinsky RM, Jepsen K, Heinzel T, Torchia J, Mullen TM,Schiff R, Del Rio AL, Ricote M, Ngo S, Gemsch J, Hilsen-beck SG, Osborne CK, Glass CK, Rosenfeld MG, RoseDW 1998 Diverse signaling pathways modulate nuclearreceptor recruitment of N-CoR and SMRT complexes.Proc Natl Acad Sci USA 95:2920–2925
17. Szapary D, Huang Y, Simons Jr SS 1999 Opposing ef-fects of corepressor and coactivators in determining thedose-response curve of agonists, and residual agonistactivity of antagonists, for glucocorticoid receptor-regu-lated gene expression. Mol Endocrinol 13:2108–2121
18. Fryer CJ, Kinyamu HK, Rogatsky I, Garabedian MJ, Ar-cher TK 2000 Selective activation of the glucocorticoidreceptor by steroid antagonists in human breast cancerand osteosarcoma cells. J Biol Chem 275:17771–17777
19. Shang Y, Hu X, DiRenzo J, Lazar MA, Brown M 2000Cofactor dynamics and sufficiency in estrogen receptor-regulated transcription. Cell 103:843–852
20. Ray DW, Suen CS, Brass A, Soden J, White A 1999Structure/function of the human glucocorticoid receptor:tyrosine 735 is important for transactivation. Mol Endo-crinol 13:1855–1863
21. Bledsoe RK, Montana VG, Stanley TB, Delves CJ, ApolitoCJ, McKee DD, Consler TG, Parks DJ, Stewart EL, Will-son TM, Lambert MH, Moore JT, Pearce KH, Xu HE 2002Crystal structure of the glucocorticoid receptor ligandbinding domain reveals a novel mode of receptor dimer-ization and coactivator recognition. Cell 110:93–105
22. Darimont BD, Wagner RL, Apriletti JW, Stallcup MR,Kushner PJ, Baxter JD, Fletterick RJ, Yamamoto KR1998 Structure and specificity of nuclear receptor-coac-tivator interactions. Genes Dev 12:3343–3356
23. Lind U, Carlstedt-Duke J, Gustafsson JA, Wright AP1996 Identification of single amino acid substitutions ofCys-736 that affect the steroid-binding affinity and spec-ificity of the glucocorticoid receptor using phenotypicscreening in yeast. Mol Endocrinol 10:1358–1370
24. Wagner BL, Norris JD, Knotts TA, Weigel NL, McDonnellDP 1998 The nuclear corepressors NCoR and SMRT arekey regulators of both ligand- and 8-bromo-cyclic AMP-dependent transcriptional activity of the human proges-terone receptor. Mol Cell Biol 18:1369–1378
25. Smith CL, Nawaz Z, O’Malley BW 1997 Coactivator andcorepressor regulation of the agonist/antagonist activityof the mixed antiestrogen, 4-hydroxytamoxifen. Mol En-docrinol 11:657–666
858 Mol Endocrinol, May 2003, 17(5):845–859 Stevens et al. • Tyrosine 735 and Cofactor Recruitment to the GR

26. Jackson TA, Richer JK, Bain DL, Takimoto GS, Tung L,Horwitz KB 1997 The partial agonist activity of antag-onist-occupied steroid receptors is controlled by anovel hinge domain-binding coactivator L7/SPA andthe corepressors N-CoR or SMRT. Mol Endocrinol11:693–705
27. Schirmbeck R, Deppert W 1989 Nuclear subcompart-mentalization of simian virus 40 large T antigen: evidencefor in vivo regulation of biochemical activities. J Virol63:2308–2316
28. Onate SA, Boonyaratanakornkit V, Spencer TE, Tsai SY,Tsai MJ, Edwards DP, O’Malley BW 1998 The steroidreceptor coactivator-1 contains multiple receptor inter-acting and activation domains that cooperatively en-hance the activation function 1 (AF1) and AF2 domains ofsteroid receptors. J Biol Chem 273:12101–12108
29. McNally JG, Muller WG, Walker D, Wolford R, Hager GL2000 The glucocorticoid receptor: rapid exchange withregulatory sites in living cells. Science 287:1262–1265
30. Grimm SL, Nordeen SK 1999 Luciferase reporter genevectors that lack potential AP-1 sites. Biotechniques 27:220–222
31. Ray DW, Davis JR, White A, Clark AJ 1996 Glucocorti-coid receptor structure and function in glucocorticoid-resistant small cell lung carcinoma cells. Cancer Res56:3276–3280
32. Chen D, Ma H, Hong H, Koh SS, Huang SM, Schurter BT,Aswad DW, Stallcup MR 1999 Regulation of transcriptionby a protein methyltransferase. Science 284:2174–2177
33. Kalkhoven E, Valentine JE, Heery DM, Parker MG 1998Isoforms of steroid receptor co-activator 1 differ in theirability to potentiate transcription by the oestrogen recep-tor. EMBO J 17:232–243
34. Fagart J, Wurtz JM, Souque A, Hellal-Levy C, Moras D,Rafestin-Oblin ME 1998 Antagonism in the human min-eralocorticoid receptor. EMBO J 17:3317–3325
35. Ito H, Fukuda Y, Murata K, Kimura A 1983 Transforma-tion of intact yeast cells treated with alkali cations. JBacteriol 153:163–168
36. Miller JH 1972 Experiments in molecular genetics.Cold Spring Harbor, NY: Cold Spring Harbor Labora-tory Press
37. Heery DM, Kalkhoven E, Hoare S, Parker MG 1997 Asignature motif in transcriptional co-activators mediatesbinding to nuclear receptors. Nature 387:733–736
Stevens et al. • Tyrosine 735 and Cofactor Recruitment to the GR Mol Endocrinol, May 2003, 17(5):845–859 859




















