
Immunomodulatory effect of recombinant RNA-dependent RNA polymerase protein of Macrobrachium...
11
Immunomodulatory effect of recombinant RNA-dependent RNA polymerase protein of Macrobrachium rosenbergii nodavirus in giant freshwater prawn M. rosenbergii Pramoda Kumar Sahoo 1 , Mudagandur S Shekhar 2 , Abhilipsa Das 1 , Manickam Dilli Kumar 2 , Bindu R Pillai 1 & A S Sahul Hameed 3 1 Central Institute of Freshwater Aquaculture, Kausalyaganga, Bhubaneswar, India 2 Genetics and Biotechnology Unit, Central Institute of Brackishwater Aquaculture, R.A. Puram, Chennai, India 3 OIE Reference Laboratory for WTD, C. Abdul Hakeem College, Melvisharam,Vellore, India Correspondence: P K Sahoo, Central Institute of Freshwater Aquaculture, Kausalyaganga, Bhubaneswar 751002, India. Email: [email protected] Abstract The present study evaluated the role of recombinant RNA-dependent RNA polymerase (RdRp) protein of Macrobrachium rosenbergii nodavirus ( MrNV) in mod- ulating the immune response and in reducing MrNV load in infected prawn. In the ¢rst experiment, prawns (25^30 g) were injected with recombinant RdRp protein (RP) at a concentration of 0, 1.0 and 10 mg, and immune parameters and expression of some immune-related genes were measured up to 14 days post injection (p.i.). In the second experiment, early juveniles were injected with a similar dose of RdRp and animals were challenged by immersion with MrNV. The infection status was detected in muscles by nested RT-PCR up to 21 days post chal- lenge. Prawn injected with higher concentration of RP showed signi¢cantly higher total haemocyte count at di¡erent period post injection. Signi¢cant up-regulation of immune-related genes was observed within 24 h in prawn treated with lower dose of RP and after 7 days p.i. at higher level of RP injection compared with adult control. Most of the tested sam- ples (63%) were found to be RT-PCR positive for MrNVat 48 h of post-immersion challenge. After 14 days, MrNV was detected only in control prawn, while both RP-injected groups were MrNV negative. This study elucidated the potential viral load reduc- tion role played by RdRP in MrNV-infected prawn. Keywords: Macrobrachium rosenbergii nodavirus ( MrNV), RNA-dependent RNA-polymerase (RdRp), Immunomodulation, Macrobrachium rosenbergii , Virus clearance Introduction The giant freshwater prawn Macrobrachium rosenber- gii is one of the most important cultured crustacean species with high economic value in global scenario and undoubtedly a major contributor of aquaculture production. Increasing demand of M. rosenbergii in domestic as well as export market has led to a large- scale production through intensi¢cation of culture practices. The poor husbandry condition of the hatcheries as well as high stocking density and trans- port of brood stocks in adverse environmental condi- tions have led to the emergence of new infectious diseases. Since 2001, Macrobrachium hatcheries situ- ated on the south-coast of India are facing a heavy loss because of an emerging disease called white tail disease or white muscle disease. Among the major viral diseases encountered by the giant freshwater prawn M. rosenbergii , white tail disease caused by Macrobrachium rosenbergii nodavirus ( MrNV) is con- sidered as the most devastating one. This virus-borne disease was ¢rst observed in the Island of Guadeloupe © 2011 Blackwell Publishing Ltd 1096 Aquaculture Research, 2012, 43, 1096–1106 doi: 10.1111/j.1365-2109.2011.02913.x
Transcript of Immunomodulatory effect of recombinant RNA-dependent RNA polymerase protein of Macrobrachium...

Immunomodulatory effect of recombinant
RNA-dependent RNA polymerase protein of
Macrobrachium rosenbergii nodavirus in giant
freshwater prawn M. rosenbergii
Pramoda Kumar Sahoo1, Mudagandur S Shekhar2, Abhilipsa Das1, Manickam Dilli Kumar2,Bindu R Pillai1 & A S Sahul Hameed3
1Central Institute of FreshwaterAquaculture, Kausalyaganga, Bhubaneswar, India2Genetics and Biotechnology Unit, Central Institute of BrackishwaterAquaculture, R.A. Puram, Chennai, India3OIE Reference Laboratory forWTD, C. Abdul Hakeem College, Melvisharam,Vellore, India
Correspondence: P K Sahoo, Central Institute of FreshwaterAquaculture, Kausalyaganga, Bhubaneswar 751002, India.
Email: [email protected]
Abstract
The present study evaluated the role of recombinantRNA-dependent RNA polymerase (RdRp) protein ofMacrobrachium rosenbergii nodavirus (MrNV) inmod-ulating the immune response and in reducingMrNVload in infected prawn. In the ¢rst experiment,prawns (25^30 g) were injected with recombinantRdRp protein (RP) at a concentration of 0, 1.0 and10 mg, and immune parameters and expression ofsome immune-related genes were measured up to14days post injection (p.i.). In the second experiment,early juveniles were injected with a similar dose ofRdRp and animals were challenged by immersionwith MrNV. The infection status was detected inmuscles by nested RT-PCR up to 21 days post chal-lenge. Prawn injected with higher concentration ofRP showed signi¢cantly higher total haemocytecount at di¡erent period post injection. Signi¢cantup-regulation of immune-related genes was observedwithin 24 h in prawn treated with lower dose of RPand after 7 days p.i. at higher level of RP injectioncompared with adult control. Most of the tested sam-ples (63%) were found to be RT-PCR positive forMrNVat 48 h of post-immersion challenge. After 14days, MrNV was detected only in control prawn,while both RP-injected groups were MrNV negative.This study elucidated the potential viral load reduc-tion role played by RdRP inMrNV-infected prawn.
Keywords: Macrobrachium rosenbergii nodavirus(MrNV), RNA-dependent RNA-polymerase (RdRp),Immunomodulation, Macrobrachium rosenbergii,Virus clearance
Introduction
The giant freshwater prawnMacrobrachium rosenber-gii is one of the most important cultured crustaceanspecies with high economic value in global scenarioand undoubtedly a major contributor of aquacultureproduction. Increasing demand of M. rosenbergii indomestic as well as export market has led to a large-scale production through intensi¢cation of culturepractices. The poor husbandry condition of thehatcheries as well as high stocking densityand trans-port of brood stocks in adverse environmental condi-tions have led to the emergence of new infectiousdiseases. Since 2001,Macrobrachium hatcheries situ-ated on the south-coast of India are facing a heavyloss because of an emerging disease called white taildisease or white muscle disease. Among the majorviral diseases encountered by the giant freshwaterprawn M. rosenbergii, white tail disease caused byMacrobrachium rosenbergii nodavirus (MrNV) is con-sidered as the most devastating one.This virus-bornediseasewas ¢rst observed in the Island of Guadeloupe
© 2011 Blackwell Publishing Ltd1096
Aquaculture Research, 2012, 43, 1096–1106 doi:10.1111/j.1365-2109.2011.02913.x

in 1995, followed by West Indies, Taiwan, China,Bangkok, India and most recently in Australia (Ar-cier, Herman, Lightner, Redman, Mari & Bonami1999; Sahul Hameed, Yoganandhan, Sri Widada &Bonami 2004a; Shekhar, Azad & Jithendran 2006;Owens, Fauce, Jantunen, Hayakijkosol & Zeng 2009).The causative agent of WTD is identi¢ed as MrNVand is included in Nodaviridae family. The WTD-af-fected larvae and postlarvae show lethargy, milkywhitish coloration of tail region and abdominal mus-cle, degeneration of telson and uropods, muscle ne-crosis and 100% mortality within few days. Thediscoloration starts at the abdominal segments andextends to anterior and posterior parts of the body.Mainly, the postlarvae and early larvae are moreprone to this disease. It may be due to their poor im-munity, whereas the adults are found to be the car-rier of this virus (Bonami, Shi, Qian & Sri Widada2005).Macrobrachium rosenbergii nodavirus is an
icosahedral, non-enveloped particle, 26^27 nm indiameter and has a genome with two pieces ofsingle-stranded RNAs (ssRNA). RNA-1 encodes foran important viral enzyme RNA-dependent RNApolymerase (RdRp) of 2.9 kb and capsid proteintranslated from RNA-2 of 1.3 kb (Qian, Shi, Zhang,Cao, Liu, Li, Xie, Cambournac & Bonami 2003). Dur-ing viral replication, a subgenomic RNA transcribedfrom RNA-1was found to encode B2, a non-structur-al protein that is able to inhibit the cellular RNA in-terference (Pillai, Bonami & Sri Widada 2006).Genomic RNA1of the virus is translated to RdRp thathelps in the replication of virus and in the regulationof its development in the host cell (Bonami & SriWi-dada 2011). RdRp was found to be the major targetprotein, which helps in the stability of the virus, con-taining the machinery for the production of capsidand other subunit RNAs (Wu, Lu & Chi 2010).Although considerable progress has been made in
MrNV and XSV molecular characterization (Pillaiet al. 2006;Tripathy, Sahoo, Kumari, Mishra, Sarangi& Ayyappan 2006), no preventive or control measurehas been worked out for this disease. It was mostlyattributed to the poor understanding of MrNV/XSVreplication mechanism as well as to molecular pro-cesses of the prawn immune responses being com-promised during virus infections. On the otherhand, considerable progress has been made on theuse of using immunostimulants, rendering protec-tion to various crustacean diseases (Balasubrama-nian, Sarathi, Venkatesan,Thomas & Sahul Hameed2008) but these are not adequate to meet the demand
in a fool-proof manner. They may control the diseasefrom the hatcheries but the chances cannot be ex-cluded from the larva and postlarvae from being in-fected in the grow-out stages. Hence alternatively,vaccine and immunization might o¡er an e¡ectiveway for protection against the disease on a long-termbasis, probably even in presence of short immune re-sponse and could be the most potential strategy toovercome the current situation. Recently, manyauthors have described about quasi-immune re-sponse in shrimpwhere the naturally virus-survivedanimals were protected against subsequent viralchallenge (Venegas, Nonaka, Mushiake, Nishizawa &Muroga 2000). Various viral recombinant proteinshave been studied so far as vaccines in di¡erent hostsystems with notable e¡ects against pathogens(Heras, Prieto & Saint-Jean 2009). A high degree ofprotection was induced in Marsupenaeus japonicusagainst WSSV by intramuscular injection of WSSV-recombinant proteins, rVP26 and rVP28 (Namikoshi,Wu, Yamashita, Nishizawa, Nishioka, Arimoto &Muroga 2004; Caipang, Verjan, Ooi, Kondo, Hirono,Aoki, Kiyono & Yuki 2008). Thus, the development ofsubunit ‘vaccines’ based onMrNVreplication-relatedproteins would be feasible and desirable as these pro-teins might interact withvirus inside the host or trig-gering the host defence. RdRp plays crucial role invirus replication. Hence, recombinant RdRp protein(RP) may act as an immune modulator by producingimmune factors against the protein, which will helpin down regulation ofMrNV progeny RNA synthesis(Wu et al. 2010). In order to ¢nd the immune-relevantfactors in prawn modulated after exposure to RdRp,some of the di¡erentially expressed genes wereidenti¢ed and their expression pro¢les in the hepato-pancreas of the M. rosenbergii were studied. Prophe-noloxiadase (PPO) enzyme is a well-known innateimmunity-related enzyme of prophenol oxidase sys-tem, which leads to melanization and production ofsuperoxide radicals, which are bactericidal in nature(Soderhall & Cerenius 1992). Pattern recognitionsproteins like b-glucan binding proteins (BGBP) acti-vates the PO system and are expressed abundantlyin hepatopancreas (Wang, Chang & Chen 2007). a-2macroglobulin (A2M) is a non-speci¢c protease inhi-bitor, which plays a major role in inhibition and re-moval of harmful proteases as well as delivery ofcytokines (Ho, Cheng & Cheng 2009). Cytochromeoxidase (cytox) gene is recognized as a part of respira-tory pathway of host that helps in the production ofsuperoxide radicals. During the process of phagocy-tosis, superoxide radicals are formed, which need to
© 2011 Blackwell Publishing Ltd, Aquaculture Research, 43, 1096–1106 1097
Aquaculture Research, 2012, 43, 1096–1106 Immunomodulation by RdRP protein of MrNV in prawn P K Sahoo et al.

be eliminated promptly. Hence, antioxidant enzymeslike superoxide dismutase (SOD) are used to converttoxic superoxide radicals to molecular oxygen.In our previous study, we have cloned, expressed
and characterized RP of MrNV (Shekhar, Sahoo, Dil-likumar & Das 2011). The present study was carriedout to measure the immunomodulatory role of twodi¡erent dose levels of RP in juveniles ofM. rosenber-gii by looking into alterations invarious haemolymphimmune parameters and innate immune-relatedgene expression in hepatopancreas. The presentstudy also measured the virus clearance e⁄ciency ofRP after challenging RP-exposed early juveniles ofprawnwithMrNV.
Material and methods
Prawn
The experimental animals M. rosenbergii (25^30 g,intermoult stage) were collected from the prawncomplex of CIFA, Bhubaneswar, India, and acclima-tized for 1 week at the wet laboratory before experi-mentation. During the experiment, the watertemperature was maintained at 28 � 2 1C, pH 7.5^8.0, total hardness 80^100mg L�1, dissolved oxygencontent 6^7mg L�1 and ammonia at o0.1mg L�1.The prawns here were fed with a commercial pelletfeed twice a day.
Preparation of recombinant RdRp
Plasmids [pET32a (1) vector, Novagen, Darmstadt,Germanywith RdRp insert] isolated from the positiveclones of DH5a cells were transformed into compe-tent BL21 (DE3) pLysS cells for expression of recombi-nant protein as described previously (Shekhar et al.2011). Brie£y,MrNV RdRpwas expressed as a recom-binant fusion protein (44.5 kDa) in Escherichia coliand was analysed by SDS-PAGE. Puri¢cation of RPwas carried out using an a⁄nity column and puritychecked inWestern blot using anti-His mouse mono-clonal antibodies.
Immunomodulation study
One hundred and twenty-six prawns were randomlydistributed into three groups, Group A, B and C. Ineach group, 42 prawns were equally distributed inthree tanks.The prawns were fedwith standard com-mercial pellet feed during the whole experimental
period at 4% of their bodyweight in two divided doses.In Group A, the prawns were individually injectedwith 100 mL of PBS intramuscularly into the secondabdominal segment and were considered as the con-trol group, whereas Groups B and C were injected in-dividually with a concentration of 1 and 10 mg of RPdissolved in 100 mL of PBS, respectively, for immuno-modulation study. After 0,6,12, 24,72 h, and 7 and14days of injection, various immune parameters werestudied to evaluate the immunomodulatory role ofRP, and hepatopancreas was collected to study the ex-pression of immune genes in immunized prawn.
Immune parameters
Total haemocyte count
Haemolymph (50 mL) was collected from the ventralsinus cavity of six prawns of each group for each timeperiod using a 26-Gauge needle and 1mL syringecontaining 450 mL anticoagulant (sodium chloride0.45M, glucose 0.1M, sodium citrate 30mM, citricacid 26mM, EDTA 20mM, pH 4.5 ) and ¢xative solu-tion (sodium cacodylate 0.1M and 1.5% glutaralde-hyde) in 1:1 ratio at 0, 6, 12, 24, 72 h, and 7 and 14days p.i. for total and di¡erential haemocyte counts.A drop of haemolymph was placed on the haemocyt-ometer slide and di¡erent types of cells were identi-¢ed according to Sierra, Guevara, Lasscurain, Perez,Agundis, Zenteno and Vazquez 2001. For haemo-lymph supernatant collection, haemolymph withoutanticoagulant was collected, allowed to clot and sub-sequently the clot was broken using a sterile needleand left at 4 1C for 2^3 h. The tube was then centri-fuged for 10min at 5000 � g, the supernatant wascollected and kept at ^30 1C for further use. Part ofthe supernatant was used to measure the total pro-tein following the Bradford method (Bradford 1976)and remaining 50 mL was used for haemagglutina-tion (HA) titre measurement using1.5% rabbit RBC.
Preparation of rabbit RBC suspension
A rabbit maintained at the Institute animal housewas bled and the blood was collected in Alsever’s so-lution in a 1:1 ratio. Further, it was centrifuged at4000 � g for 15min and the supernatant was dis-carded. The packed RBCs was washed thrice withsterile PBS (containing Ca21 and Mg21, pH 7.3) andresuspended at1.5% v/v in PBS for the measurementof HA titre (Sahoo, Pillai, Mohanty, Kumari, Mohanty
© 2011 Blackwell Publishing Ltd, Aquaculture Research, 43, 1096–11061098
Immunomodulation by RdRP protein of MrNV in prawn P K Sahoo et al. Aquaculture Research, 2012, 43, 1096–1106

& Mishra 2007). Brie£y, double serial dilution of thehaemolymph supernatant was made in PBS (withCa21 and Mg21), and then 50 mL of1.5% RaRBC wasadded to each well of the microtitre plate and incu-bated for1h at 37 1C. The HA titre was de¢ned as thelast dilution of serum showing minimal positive ag-glutination. Values were expressed as the reciprocalof HA titre.
Expression analysis of immune-related genes
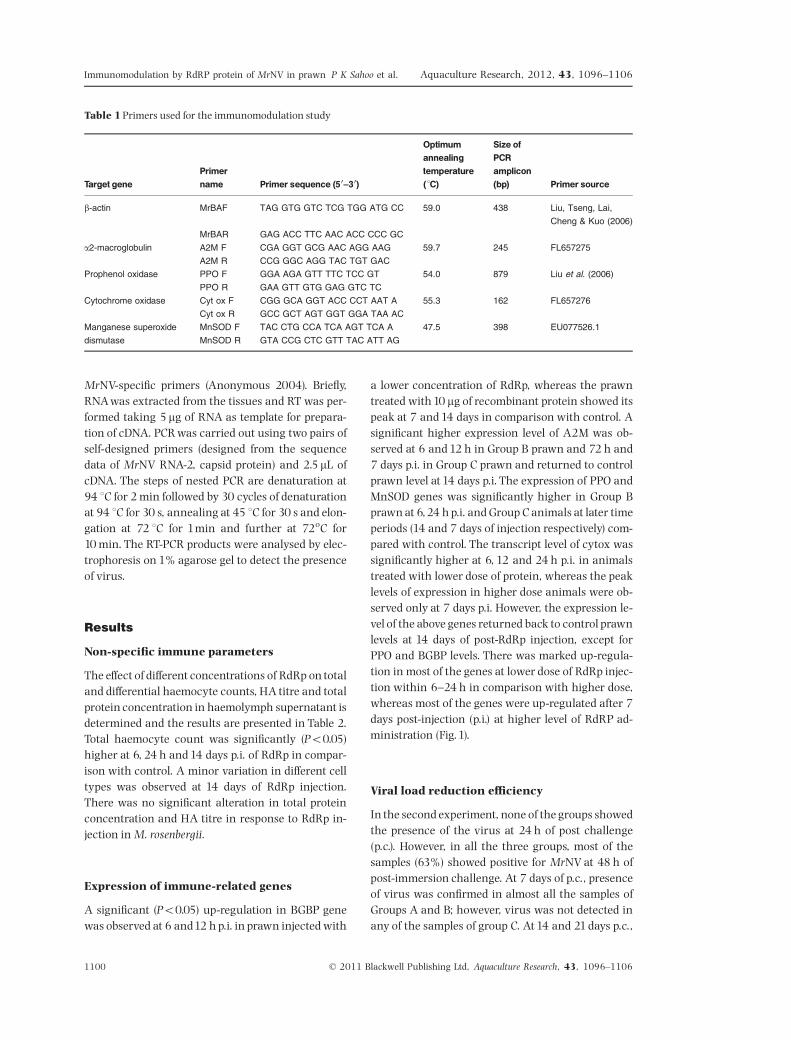
Similarly for gene expression studies, hepatopan-creas from four individual prawns were collected inRNAlater (Sigma, St. Louis, MO, USA) after bleedingfor each time period of a group and RNA extractionwas performed using TRI reagent (Sigma). To mini-mize the chances of genomic DNA contamination,the RNAwas treated with DNAse I (Fermentas, GlenBurnie, MD, USA) and subsequently inactivatedbefore reverse transcription following the instruc-tions of the manufacturer. The purity and quantityof RNA were determined by measuring the ratio ofoptical density at 260/280 nm using a Nanodrop1000 (NanoDrop Technologies Inc.,Wilmington, DE,USA). Total RNAwas reverse-transcribed taking 1 mgof RNA for the preparation of cDNA. Expression of¢ve immune-related genes viz., BGBP, A2M, PPO, cy-tox and manganese superoxide dismutase (MnSOD)and a house-keeping gene b-actin was measured bysemi-quantitative RT-PCR method. Each PCR reactionconsisted of 40.70 mL of dH2O,5 mL of10 � PCRbu¡er,1 mL of 10mM dNTPs , 1 mL (10 pmol) of each forwardand reverse primers, followed by 0.3 mL (1.5 U) of TaqDNA polymerase and 1 mL of cDNA. The ampli¢ca-tion pro¢le was 94 1C for 3min followed by 39 cyclesof denaturation for 45 s at 94 1C, at an appropriateannealing temperature for 45 s and extension at72 1C for1min 30 s, followed by a ¢nal extension for10min at 72 1C.The primers used and the annealingtemperature details are given inTable1.
Agarose gel electrophoresis
The generated PCR products were analysed by elec-trophoresis on1.0% agarose gel. The relative levels ofexpression of each gene were analysed bydensitome-try using ALPHAEASE
s
FC IMAGING software (AlphaInnotech, San Leandro, CA, USA). The ratios of im-mune-related genes/b-actin product were subse-quently calculated after subtraction of thebackground pixel intensity for each gene of interestand mean values ( � SE) were calculated.
Statistical analysis
Statistical di¡erences in gene expression and im-mune parameters between experimental and controlsamples in each group were assessed using one-wayANOVA. Means were comparedwith Duncan’s multiplerange test and the di¡erence was considered signi¢-cant when Po0.05.
Virus load reduction e⁄ciency study
The prawn early juveniles utilized in this experimentwere obtained from CIFA hatchery that had no pre-vious history of MrNV infection. Further, the repre-sentative animal samples were screened for absenceof MrNV using nested RT-PCR as detailed later. Onehundred and twenty-six early juveniles (1^2 g) weredistributed randomly into three groups A, B and C,having forty-two prawns in each group and the ex-periment was performed in triplicate. In Group A,the prawns were individually injected with 20 mL ofPBS intramuscularly into the abdominal segmentand were considered as control, whereas groups Band C were injected with a concentration of 1 and10 mg of RP protein dissolved in 20 mL of PBS respec-tively. After 24 h of injection, an immersion chal-lenge with MrNV suspension was performed. In theimmersion challenge, 14 juveniles from each tripli-cate of a groupwere placed ina 2 L beaker containingfreshwater with continuous aeration. The MrNV in-oculum (obtained from OIE Reference Laboratory forWTD, Melvisharam,Vellore, India ) was actually pre-pared from infected PLs collected from di¡erenthatcheries of Andhra Pradesh, India, and challengedat 0.1% of the total rearing medium (1mL L�1), fol-lowing the protocol of Ravi, Basha,Taju, Ram Kumarand Sahul Hameed (2010). After 3 h of exposure tothe virus suspension, the animals were transferredto the glass jars containing freshwater and fed withartemia. The experimental animals were examinedtwice per day for clinical signs of WTD. The mori-bund/live animals were collected at 24, 48 h and 7,14 and 21days of post-challenge.
Virus detection using nested RT-PCR
In the challenge experiment, RNA from muscle sam-ples were extracted using TRI reagent (Sigma) as permanufacturer’s instruction and a nested RT-PCRwas performed using published primers with CIFA-developed nested RT-PCR-based diagnostics using
© 2011 Blackwell Publishing Ltd, Aquaculture Research, 43, 1096–1106 1099
Aquaculture Research, 2012, 43, 1096–1106 Immunomodulation by RdRP protein of MrNV in prawn P K Sahoo et al.
MrNV-speci¢c primers (Anonymous 2004). Brie£y,RNAwas extracted from the tissues and RT was per-formed taking 5 mg of RNA as template for prepara-tion of cDNA. PCRwas carried out using two pairs ofself-designed primers (designed from the sequencedata of MrNV RNA-2, capsid protein) and 2.5 mL ofcDNA. The steps of nested PCR are denaturation at94 1C for 2min followed by 30 cycles of denaturationat 94 1C for 30 s, annealing at 45 1C for 30 s and elon-gation at 72 1C for 1min and further at 72oC for10min. The RT-PCR products were analysed by elec-trophoresis on1% agarose gel to detect the presenceof virus.
Results
Non-speci¢c immune parameters
The e¡ect of di¡erent concentrations of RdRp on totaland di¡erential haemocyte counts, HA titre and totalprotein concentration in haemolymph supernatant isdetermined and the results are presented in Table 2.Total haemocyte count was signi¢cantly (Po0.05)higher at 6, 24 h and14 days p.i. of RdRp in compar-ison with control. A minor variation in di¡erent celltypes was observed at 14 days of RdRp injection.There was no signi¢cant alteration in total proteinconcentration and HA titre in response to RdRp in-jection inM. rosenbergii.
Expression of immune-related genes
A signi¢cant (Po0.05) up-regulation in BGBP genewas observed at 6 and12 h p.i. in prawn injected with
a lower concentration of RdRp, whereas the prawntreated with10 mg of recombinant protein showed itspeak at 7 and14 days in comparison with control. Asigni¢cant higher expression level of A2M was ob-served at 6 and12 h in Group B prawn and 72 h and7 days p.i. in Group C prawn and returned to controlprawn level at14 days p.i. The expression of PPO andMnSOD genes was signi¢cantly higher in Group Bprawnat 6,24 h p.i. and Group C animals at later timeperiods (14 and 7 days of injection respectively) com-pared with control. The transcript level of cytox wassigni¢cantly higher at 6, 12 and 24 h p.i. in animalstreated with lower dose of protein, whereas the peaklevels of expression in higher dose animals were ob-served only at 7 days p.i. However, the expression le-vel of the above genes returned back to control prawnlevels at 14 days of post-RdRp injection, except forPPO and BGBP levels. There was marked up-regula-tion in most of the genes at lower dose of RdRp injec-tion within 6^24 h in comparison with higher dose,whereas most of the genes were up-regulated after 7days post-injection (p.i.) at higher level of RdRP ad-ministration (Fig.1).
Viral load reduction e⁄ciency
In the second experiment, none of the groups showedthe presence of the virus at 24 h of post challenge(p.c.). However, in all the three groups, most of thesamples (63%) showed positive for MrNVat 48 h ofpost-immersion challenge. At 7 days of p.c., presenceof virus was con¢rmed in almost all the samples ofGroups A and B; however, virus was not detected inany of the samples of group C. At14 and 21days p.c.,
Table 1Primers used for the immunomodulation study
Target genePrimername Primer sequence (5 0^30)
Optimumannealingtemperature(1C)
Size ofPCRamplicon(bp) Primer source
b-actin MrBAF TAG GTG GTC TCG TGG ATG CC 59.0 438 Liu, Tseng, Lai,
Cheng & Kuo (2006)
MrBAR GAG ACC TTC AAC ACC CCC GC
a2-macroglobulin A2M F CGA GGT GCG AAC AGG AAG 59.7 245 FL657275
A2M R CCG GGC AGG TAC TGT GAC
Prophenol oxidase PPO F GGA AGA GTT TTC TCC GT 54.0 879 Liu et al. (2006)
PPO R GAA GTT GTG GAG GTC TC
Cytochrome oxidase Cyt ox F CGG GCA GGT ACC CCT AAT A 55.3 162 FL657276
Cyt ox R GCC GCT AGT GGT GGA TAA AC
Manganese superoxide
dismutase
MnSOD F TAC CTG CCA TCA AGT TCA A 47.5 398 EU077526.1
MnSOD R GTA CCG CTC GTT TAC ATT AG
© 2011 Blackwell Publishing Ltd, Aquaculture Research, 43, 1096–11061100
Immunomodulation by RdRP protein of MrNV in prawn P K Sahoo et al. Aquaculture Research, 2012, 43, 1096–1106

MrNVcould be only detected in PBS-injected group,while all RdRp-injected groups showed negative forthe presence of this virus (Fig. 2).
Discussion
The present study was carried out to ¢nd the e⁄cacyof the recombinant RPofMrNVas an immunomodu-lator in modulating the immune status to inhibit the
viral replication and propagation in infected prawn.In the ¢rst experiment, prawns were used to checkthe immunomodulatory e¡ect of RP and in the sec-ond experiment, early juveniles were taken to studythe virus load reduction. It is well understood thatadult animals act as carriers of infection, whereasearly stages (larvae, postlarvae and juveniles) aremore prone to infection and mortality (Sahul Ha-meed,Yoganandhan, SriWidada, Bonami 2004b).Immune-relevant molecules from crustacean hae-
mocytes play an essential role in defence against in-vasive pathogens (Johansson & S˛derh�ll 1989). THCre£ects the immune status of a shrimp as it is in-volved in many cellular mechanisms such as phago-cytosis, encapsulation, nodule formation and phenoloxidase activity. (Pipe & Coles 1995). Increase in THCis considered as a consequence of rapid proliferationof haematopoietic cells in host in order to respond toany stimulant or foreign pathogens (Liu, Soderhall &Jiravanichpaisal 2009). In this experiment, a signi¢-cant increase in THC was observed at 6 and 24 h inM. rosenbergii treated with RP signifying prolifera-tion of haemocytes in response to a viral protein.The increase in haemocyte count was also well cor-related with an increase in the expression of PPOtranscripts at the same time periods. This increase in
Table 2 E¡ect of di¡erent concentrations of RNA-dependent RNA polymerase on di¡erent immune parameters ofMacrobra-chium rosenbergii
Timeperiod Group
THC(107 cells/mL)
Fusiformcells (%)
Large ovoidcells (%)
Undifferentiatedcells (%)
Haemagglutinationtitre (log 2)
TotalProtein(g/dL)
0h A 0.76 � 0.05 87.66 � 0.82 8.04 � 0.61 4.37 � 0.24 2.75 � 0.14 9.81 � 0.49
B 0.76 � 0.05 87.66 � 0.82 8.04 � 0.61 4.37 � 0.24 2.75 � 0.14 9.81 � 0.49
C 0.76 � 0.05 87.66 � 0.82 8.04 � 0.61 4.37 � 0.24 2.75 � 0.14 9.81 � 0.49
6 h A 0.31 � 0.04a 78.83 � 4.16 14.65 � 3.21 6.63 � 0.79 4.5 � 1.04 8.41 � 1.21
B 0.55 � 0.06 b 75.06 � 3.43 15.9 � 1.03 8.24 � 1.67 3.67 � 0.83 7.09 � 1.06
C 0.51 � 0.03 b 76.55 � 3.7 16.35 � 2.6 7.29 � 1.33 5.5 � 0.29 7.61 � 0.63
12h A 0.56 � 0.16 84.83 � 1.47 9.55 � 1.14 4.61 � 0.16 5.5 � 0.5 6.78 � 0.6
B 0.55 � 0.07 82.11 � 1.79 10.79 � 1.59 5.91 � 0.84 4 � 0.76 6.59 � 0.24
C 0.7 � 0.11 82.72 � 0.71 11.06 � 0 4.86 � 0.06 4.75 � 0.43 7.43 � 0.9
24h A 0.59 � 0.06 a 86.77 � 1.12 b 8.98 � 0.7a 4.14 � 0.44 3.17 � 0.6 6.63 � 0.6
B 0.63 � 0.05 a 85.05 � 1.28ab 10.2 � 0.83a 4.73 � 0.44 4.17 � 1.36 7.92 � 0.81
C 0.9 � 0.05b 80.35 � 1.73 a 13.84 � 1.13b 5.44 � 0.34 6 � 0 6.86 � 0.43
72h A 0.85 � 0.07 81.66 � 1.8 12.41 � 1.67 5.69 � 0.17b 6 � 0 8.19 � 0.95
B 0.69 � 0.08 86.96 � 1.14 8.52 � 1.36 4.85 � 0.18ab 4 � 0.87 7.47 � 0.5
C 0.89 � 0.13 85.77 � 3.48 8.03 � 0.94 3.89 � 0.52a 2.67 � 1.45 9.85 � 1.34a
7 days A 0.67 � 0.04 86.87 � 1.41 8.55 � 0.73 4.56 � 0.69 4.67 � 0.88 7.28 � 1.01
B 0.81 � 0.06 86.09 � 1.99 7.67 � 0.61 5.84 � 1.57 5.75 � 0.14 9.18 � 0.97
C 0.76 � 0.09 88.68 � 1.64 7.25 � 0.76 3.77 � 0.48 4.67 � 0.6 8.12 � 0.74
14 days A 0.42 � 0a 87.46 � 0b 8.28 � 0b 4.25 � 0b 5.17 � 0.44 9.2 � 0.24
B 0.84 � 0b 85.59 � 0.14a 8.68 � 0c 5.97 � 0c 5 � 0 9.26 � 0.42
C 0.91 � 0c 89 � 0.58c 7.06 � 0.01a 3.91 � 0.05a 4.83 � 0.73 8.23 � 0.47
Means bearing di¡erent superscript alphabets at a particular time period in a column di¡er signi¢cantly (Po0.05). Group A: PBS con-trol, Group B: injected with 1 mg of RdRp protein (RP) and Group C: injected with 10 mg of RP.
Figure 1 Bands showing the presence of viral transcriptsat di¡erent interval of post challenge. Group A: injectedwith phosphate bu¡ered saline only; Goup B: injectedwith 1 mg of recombinant RNA-dependent RNA polymer-ase (RdRp) and Group C: injected with 10 mg of recombi-nant RdRp.
© 2011 Blackwell Publishing Ltd, Aquaculture Research, 43, 1096–1106 1101
Aquaculture Research, 2012, 43, 1096–1106 Immunomodulation by RdRP protein of MrNV in prawn P K Sahoo et al.

THC along with PO activity indicates a crucial roleagainst viral infection by host immune system. Ji,Yao andWang (2009) observed a similar increase inTHC at 6 h to 24 h in Litopenaeus vannamei after in-jection of laminarin and up to 12 h when the shrimpwas immunized with poly I:C. However, Braak, Bot-terblom, Huisman, Rombout and van der Knaap(2002) suggested that an exposure toWSSVcan causeloss of circulating haemocytes, which ultimatelyleads to death of animal. The MrNVa¡ects the hae-mocyte proliferation in infected M. rosenbergii. Raviet al. (2010) observed a signi¢cant reduction of THCinM. rosenbergii injected withMrNVand XSVon day1and 3 p.i. Therefore, the immediate regeneration ofhaemocytesmay be a critical factor for the survival ofanimals afterMrNV infection. Hence, the increase inTHC by induction of RdRp as observed in this studyindicates a possible bene¢cial role in protectingprawn fromWTD infection.
Agglutinins in several crustaceans have beencharacterized as heat-labile and calcium-dependentimmune-reactive molecules (Acharya, Mohanty &Sahoo 2004). The production of agglutinins, bacteri-cidin and lysins following exposure to pathogens isrelated to the increase in resistance of the host. Apartfrom this, the total protein content of an animal isalso an indicator of its good physical and nutritionalstatus. However, in the present experiment no signi¢-cant alterationwas observed inagglutininand in totalprotein levels of the host in response to RP injection.The pattern recognition receptors (PRRs) like
BGBP are shown to be a part of innate immune sys-tem and act as an acute phase protein by recognizingthe carbohydrate structures in the microbial cell sur-face (Srityunlucksana, Lee & Soderhall 2002). Cellu-lar mRNAs are capped, while viral RNAs have a 5 0-phosphate that can be recognized by cellular PRRs(Pichlmair, Schulz,Tan, Naslund, Liljestrom,Weber &
8765
Rel
ativ
e m
RN
A e
xpre
ssio
n
43210
0h
Group AGroup BGroup C
aaa a
bb
b
b
a ab
ab
a
aa
a a a aa
a
a
6h 12h 24h
BGBP(a) (b)
(c) (d)
(e)
Time post injection
72h 7d 14d
765
Rel
ativ
e m
RN
A e
xpre
ssio
n
43210
0h
Group A
Group B
Group C
aaa a
bb c
b
a aa
a
a a a baa
aaa
6h 12h 24h
A2M
Time post injection
72h 7d 14d
1.61.8
1.41.2
1
Rel
ativ
e m
RN
A e
xpre
ssio
n
0.80.60.40.2
00h
Group AGroup BGroup C
aaaa
b
a
ba
aaa
b
aa
aaa
aa
aa
6h 12h 24h
PPO
Time post injection
72h 7d 14d
78
65
Rel
ativ
e m
RN
A e
xpre
ssio
n
43210
0h
Group AGroup BGroup C
aaa a
bb
b
b
a aab
ab
a a aaaa
aaa
6h 12h 24h
Cytox
Time post injection
72h 7d 14d
Rel
ativ
e m
RN
A e
xpre
ssio
n
0
2
4
6
8
10
12
14
0h
Group AGroup BGroup C
aaa a
bb
b
aa a
aba a a a
a
aa
aa
a
6h 12h 24h
Mn SOD
Time post injection
72h 7d 14d
Figure 2 E¡ect of di¡erent doses of RNA-dependent RNA polymerase on expression of (a) b-glucan binding proteins(BGBP), (b) a-2-macroglobulin (A2M), (c) prophenoloxiadase (PPO), (d) cytochrome oxidase (ctyox) and (e) manganesesuperoxide dismutase (MnSOD) transcripts measured at di¡erent time periods post injection. GroupA serves as phosphatebu¡ered saline control, Group B injected with1 mg of RdRp protein (RP) and Group C injected with10 mg of RP. Bars show-ing di¡erent letter at a particular time di¡er signi¢cantly (Po0.05).
© 2011 Blackwell Publishing Ltd, Aquaculture Research, 43, 1096–11061102
Immunomodulation by RdRP protein of MrNV in prawn P K Sahoo et al. Aquaculture Research, 2012, 43, 1096–1106

ReiseSouca 2006). Thus, ssRNA, dsRNA and viralglycoproteins constitute the basic structures, where-by PRRs recognize an invading virus and lead to pro-duction of cytokines (Workenhe, Hori, Rise, Kibenge& Kibenge 2009). BGBP expression has also beenshown to be increased during WSSV infection inshrimp (Penaeus stylirostris) (Roux, Pain, Klimpel &Dhar 2002). BGBP is mainly expressed in the hepato-pancreas (Wang, Chang & Chen 2008).We observed asigni¢cant up-regulation of BGBP transcript levels at6 and12 h p.i, in lower dose group animals and at 7and 14 days in higher dose group animals, as com-pared with control. Probably, the higher doseof protein could be more than su⁄cient for early im-mune stimulation. The delayed response of BGBP le-vel in higher dose protein-injected group is wellcorrelated with an earlier study with b-1,3-glucan inL. vannamei. (Wang et al. 2008). In the present study,A2Mwas found to be signi¢cantly up-regulated at 6,12, 72 h and 14 days in RdRp-treated groups. Lu,Sung, Liu and Sung (2007) also reported that ele-vated expression of A2Mwas observed after1hwhenprawn was treated with CpG ODN. A similar ¢ndingwas observed when M. rosenbergii was challengedwith LPS (Lu et al. 2007). Evidence of increasedA2M at 24 and 12 h was also observed when theshrimps were injected with heat-killed Lactococcusgarviae and Vibriyo alganolyticus respectively (Hoet al. 2009).Animal cells generate energy in mitochondria
through oxidative phosphorylation electron trans-port system (ETS). The ETS is a major source ofreactive oxygen species (ROS), which are immuno-genically active molecules that help in destruction ofpathogens. Cytochrome oxidase is an electron trans-port enzyme that plays an important role in stimulat-ing the immune response in invertebrates (Rensburg& Coyne 2009). In this study, cytox was up-regulatedat 6 and 12 h post RdRp injection. Similarly,Lu, Sung, Liu and Sung (2009) found up-regulationof cytox at 6 and 12 h after the administration ofLPS inM. rosenbergii (Lu et al. 2009).As stated earlier, ROSare involved in destruction of
pathogen, and to minimize the e¡ect of ROS, SOD isused, which can catalyse the conversion of the highlytoxic superoxide radicals to molecular oxygen (Cho,Lee, Bang, Kim & Nama 2009). SOD is an antioxidantenzyme, which has been widely used to evaluate thedefence ability of shrimp against pathogens. Resultsof our present study showed the signi¢cant up-regu-lation of SOD activity in RP-treated groups at 6, 24 hand 7 days p.i. Campa-CoŁ rdova, Hernandez-Saavedra
and Ascencio (2002) also observed an increased pro-duction of superoxide anion at 24 h in white shrimpL. vannamei challenged withVibrio parahaemolyticus.Similarly, the up-regulated transcription level ofMnSOD has also been marked in L. vannamei at 3^24 h after the shrimp were administered with lami-narin. (Campa-CoŁ rdova et al. 2002). The level of SODwas also reported to increase signi¢cantly in Panaeusmonodon when treated with DNA construct withVP28 gene of WSSV (encapsulated with chitosan na-noparticle) at 7 days p.i, which agrees to our observa-tion in higher dose-treated animals (Rajesh Kumar,Venkatesan, Sarathi, Sarathbabu, Thomas, Basha &Sahul Hameed 2009). Similarly, when the Fennerope-naeus chinensis was fed with recombinant protein ofWSSV (rVP28) orally, the SOD level was found to beup-regulated at third,14th and 28th days of post-ad-ministration (Fu, Shuai, Xu, Li & Li 2010). It is inter-esting to mention that prawns experimentallyinfected with MrNV and XSV revealed a signi¢cantreduction in SOD activityand an increase in superox-ide ion level up to10 dpi (Ravi et al. 2010). This meansthe host defence might be highly reactive at the initi-al stage by producing free radicals to degrade virus,and in turn host antioxidant defence enzyme SODmight be exhausted duringWTD infection. Thus, theincrease in SOD activity observed in this study inRdRp-injected prawn might be helpful to combatMrNV/XSV load.The PO system is the basis of non-speci¢c immune
system in crustacean and it catalyses the formationof quinine from phenolic compounds, which subse-quently becomes melanin. Melanin is involved inwound healing and repair mechanism and quinineacts as an antifungal agent (Cerenius & Soderhall2004). In the current study, PO expressionwas signif-icantly increased when compared with control at 6and 24 h in animals injected with1 mg concentrationof protein, whereas in Group C, the highest expres-sion of POwas observed at day14 only. Sucha changein PO expression has been well documented in Chi-nese mitten crab Eriocheir sinensis, where PO is seento be activated after 6 and 12 h of dsRNA infection(Dong, Zhao, Song,Wang, Qiu, Zheng, Li, Gai & Yang2009). In general, most of the immune-related genesshowed higher expression level at an early periodafter lower dose of RP injection, whereas at a laterstage it occurred after exposure to higher dose of RPexposure. This could be probably due to the excessamount of protein that might be more than su⁄cientto stimulate the genes at higher dose because of feed-back mechanism. However, once the protein level
© 2011 Blackwell Publishing Ltd, Aquaculture Research, 43, 1096–1106 1103
Aquaculture Research, 2012, 43, 1096–1106 Immunomodulation by RdRP protein of MrNV in prawn P K Sahoo et al.

might be reduced or degraded, then the level of ex-pression gradually increased. In general, all thesegene expressions are well correlated with the in-crease in THC level in prawns and probably actshortly to reduce the viral load as marked in the sec-ond experiment.The challenge study was carried out here to evalu-
ate the e⁄ciency of RdRp in reducing theMrNV loadin the injected prawns. The prawns were challengedafter1day of RdRp treatment and nested RT-PCRwasconducted to check the presence of virus. The viruswas below detectable limit after 24 h of challenge. Si-milar to our ¢nding, earlier studies have also ob-served detection of virus after 2^3 days of challenge(Sudhakaran, Syed Musthaq, Haribabu, Mukherjee,Gopal & Sahul Hameed 2006; Ravi et al. 2010). This isprobably because the virus needs more than 24 h toestablish and replicate inside the host or becausethere was insigni¢cant amount of virus to be de-tected by nested RT-PCR method. After 48 h of im-mersion challenge, virus was detected in all thegroups including control in this study. However at 7days p.c., virus was detected only in control as well asinanimals injectedwith lower dose of RdRp, whereasno viral particles could be detected in higher doseRdRp-injected animals. An earlier study suggestedthat the virus was cleared from the adult host after25 days of infection naturally. However, the virus per-sisted in the system to a detectable level up to 20 days(Ravi et al. 2010). Hence, it is quite probable that thevirus has been cleared from the system after 7 or 14days of infection in RdRp-injected prawns because ofthe immunostimulatoryaction of RPexposure. RdRpmight have played an important role in down regu-lating virus replication that needs further in-depthstudy. Besides, there might be modulation of the hostimmune response, as observed in this study, whichcould probably provide an additive e¡ect in reducingviral load. However, further study in this regard iswarranted to prove these concepts. Interestingly, atday 14 and 21, all samples of RdRp-treated groupshowed as nested RT-PCR negative, whereas the pre-sence of this virus was detected only in PBS-injectedcontrol group. Although the mechanism is not clearlyunderstood, it is con¢rmed that exposure to RP couldbe successful in protecting the host againstMrNV.
Conclusion
The absence of viral particles in protein-injectedprawn indicated the immunomodulatory e¡ect of
the recombinant RdRp. This result correlated withthe bioassay tests indicating RdRp triggering the im-mune responsewithmore productionof haemocytes.The increase in haemocytes might be responsible forthe release of inducible anti-viral substances and im-mune e¡ector molecules in RdRp-injected prawnleading to virus reduction in infected prawns.
Acknowledgements
The authors wish to greatlyacknowledge the Depart-ment of Biotechnology for sponsoring the project andthank DrA. E. Ekanath, Director, Central Institute ofFreshwaterAquaculture, Kausalyaganga, Bhubanes-war, India, for providing necessary facilities to carryout the study.
References
Acharya S., Mohanty J. & Sahoo P.K. (2004) Humoral de-fence factors in Indian river prawn Macrobrachium mal-colmsonii. Fish & Shell¢sh Immunology17,137^147.
Anonymous (2004) RT-PCR based diagnostic for Macrobra-chium rosenbergii nodavirus. In: CIFA News 11 (ed by Sar-angi N., B.C. Mohapatra & B.L. Dhir), pp. 3^4. CentralInstitute of FreshwaterAquaculture, Bhubaneswar, India.
Arcier J.M., Herman F., Lightner D.V., Redman R., Mari J. &Bonami J.R. (1999) Aviral disease associated with mortal-ities in hatchery-reared postlarvae of the giant freshwaterprawn Macrobrachium rosenbergii. Diseases of Aquatic Or-ganisms 38,177^181.
Balasubramanian G., Sarathi M.,Venkatesan C.,Thomas J. &Sahul Hameed A.S. (2008) Studies on the immunomodu-latory e¡ect of extract of Cyanodon dactylon in shrimp, Pe-naeusmonodon, and its e⁄cacy to protect the shrimp fromwhite spot syndromevirus (WSSV). Fish&Shell¢sh Immu-nology 25,820^828.
Bonami J.R. & SriWidada J. (2011) Viral diseases of the giantfreshwater prawn Macrobrachium rosenbergii. Journal ofInvertebrate Pathology106,131^142.
Bonami J.R., Shi Z., Qian D. & SriWidada J. (2005) White taildisease of the giant freshwater prawn,Macrobrachium ro-senbergii: separation of the associated virions and
characterisation of MrNV as a new type of nodavirus.
Journal of Fish Diseases 28, 23–31.Braak C.B., Botterblom M.H., Huisman E.A., Rombout J.H. &van der KnaapW.P. (2002) Preliminary study on haemo-cyte response to white spot syndrome virus infection inblack tiger shrimp Penaeus monodon. Diseases of AquaticOrganisms 51,149^155.
Bradford M.M. (1976) A rapid and sensitive method for thequanti¢cation of microgram quantities of protein. Analy-tical Biochemistry 72, 248–254.
© 2011 Blackwell Publishing Ltd, Aquaculture Research, 43, 1096–11061104
Immunomodulation by RdRP protein of MrNV in prawn P K Sahoo et al. Aquaculture Research, 2012, 43, 1096–1106

Caipang C.M.,Verjan N., Ooi E.L., Kondo H., Hirono I., AokiT., Kiyono H. & Yuki Y. (2008) Enhanced survival ofshrimp, Penaeus (Marsupenaeus) japonicus fromwhite spotsyndrome disease after oral administration of recombi-nant VP28 expressed in Brevibacillus brevis. Fish & Shell-¢sh Immunology 25,315^320.
Campa-CoŁ rdova A.I., HernaŁ ndez-Saavedra N.Y., Philippis R.& Ascencio F. (2002) Generation of superoxide anion andSOD activity in haemocytes and muscle of Americanwhite shrimp (Litopenaeus vannamei) as a response to b-glucan and sulphated polysaccharide. Fish & Shell¢sh Im-munology12,353^366.
Cerenius L. & Soderhall K. (2004) The prophenoloxidase-ac-tivating system in invertebrates. Immunological Reviews198,116^126.
Cho Y.S., Lee S.Y., Bang I.C., Kim D.S. & Nama Y.K. (2009)Genomic organization and mRNA expression of manga-nese superoxide dismutase (Mn-SOD) from Hemibarbusmylodon (Teleostei, Cypriniformes). Fish& Shell¢sh Immu-nology 27,571^576.
Dong C., Zhao J., Song L.,Wang L., Qiu L., Zheng P., Li L., GaiY. & Yang G. (2009) The immune responses in Chinesemitten crab Eriocheir sinensis challenged with double-stranded RNA. Fish& Shell¢sh Immunology 26, 438–442.
Fu L.L., Shuai J.B., Xu Z.R., Li J.R. & Li W.F. (2010) Immuneresponses of Fenneropenaeus chinensis against white spotsyndrome virus after oral delivery of VP28 using Bacillussubtilis as vehicles. Fish & Shell¢sh Immunology 28,49^55.
Heras A., Prieto S.I.P. & Saint-Jean S.R. (2009) In vitro and invivo immune responses induced bya DNAvaccine encod-ing the VP2 gene of the infectious pancreatic necrosisvirus. Fish & Shell¢sh Immunology 27,120^129.
Ho P.Y., Cheng C.H. & Cheng W. (2009) Identi¢cation andcloning of a2-macroglobulin of giant freshwater prawnMacrobrachium rosenbergii and its expression in relationwith molt stage and bacteria injection. Fish & Shell¢shImmunology 26, 459–466.
Ji P.F., Yao C.L. & Wang Z.Y. (2009) Immune response andgene expression in shrimp (Litopenaeus vannamei) hemo-cytes and hepatopancreas against some pathogen-asso-ciated molecular patterns. Fish Shell¢sh Immunology 27,563^570.
Johansson M.W. & S˛derh�ll K. (1989) Cellular immunity incrustaceans and the proPO system. Parasitology Today 5,171^176.
Liu C.H.,Tseng D.Y., Lai C.Y., ChengW. & Kuo C.M. (2006) Mo-lecular cloning and characterization of prophenoloxidasecDNA from haemocytes of the giant freshwater prawn,Macrobrachium rosenbergii, and its transcription in relationwith moult stage. Fish & Shell¢sh Immunology 21,60^69.
Liu H., Soderhall K. & Jiravanichpaisal P. (2009) Antiviralimmunity in crustaceans. Fish & Shell¢sh Immunology27,79^88.
Lu K.Y., Sung H.J., Liu C.L. & Sung H.H. (2007) Genes expres-sion in prawn hemocytes of the giant freshwater (Macro-
brachium rosenbergii) elicited by CpG ODN). Journal of theFisheries Society ofTaiwan 34, 265^280.
Lu K.Y., Sung H.J., Liu C.L. & Sung H.H. (2009) Di¡erentiallyenhanced gene expression in hemocytes from Macrobra-chium rosenbergii challenged in vivowith lipopolysacchar-ide. Journal of Invertebrate Pathology100,9^15.
Namikoshi A.,Wu J.L.,Yamashita T., NishizawaT., NishiokaT., ArimotoM. &Muroga K. (2004) Vaccination trials withPenaeus japonicus to induce resistance to white spot syn-drome virus. Aquaculture 229, 25^35.
Owens L., Fauce K.L., Jantunen K., Hayakijkosol O. & Zeng C.(2009) Macrobrachium rosenbergii nodavirus disease(white tail disease) in Australia. Diseases in Aquatic Organ-isms 85,175^180.
Pichlmair A., Schulz O.,Tan C.P., Naslund T.I., Liljestrom P.,Weber F. & ReiseSousa C. (2006) RIG-I-mediated antiviralresponses to single-stranded RNAbearing 5 0-phosphates.Science 314,997^1001.
Pillai D., Bonami J.R. & SriWidada J. (2006) Rapid detectionofMacrobrachium rosenbergii nodavirus (MrNV) and extrasmall virus (XSV), the pathogenic agents of white tail dis-ease of Macrobrachium rosenbergii (De Man), by loop-mediated isothermal ampli¢cation. Journal of Fish Dis-eases 29, 275^283.
Pipe R.K. & Coles J.A. (1995) Environmental contaminantsin£uencing immune function inmarine bivalve molluscs.Fish & Shell¢sh Immunology 5,581^595.
Qian D., Shi Z., Zhang S., Cao Z., LiuW., Li L., XieY., Cam-bournac I. & Bonami J.R. (2003) Extra small virus-likeparticles (XSV) and nodavirus associated with whitishmuscle disease in the giant freshwater prawn, Macrobra-chium rosenbergii. Journal of Fish Diseases 26,521^527.
Rajesh Kumar S.,Venkatesan C., Sarathi M., SarathbabuV.,Thomas J., Basha K.A. & Sahul Hameed A.S. (2009) Oraldelivery of DNA construct using chitosan nanoparticlesto protect the shrimp from white spot syndrome virus(WSSV). Fish & Shell¢sh Immunology 26, 429^437.
Ravi M., Basha A.,Taju G., Ram Kumar R. & Sahul HameedA.S. (2010) Clearance of Macrobrachium rosenbergii noda-virus (MrNV) and extra small virus (XSV) and immunolo-gical changes in experimentally injected Macrobrachiumrosenbergii. Fish & Shell¢sh Immunology 28, 428^433.
RensburgM.J.V. & CoyneV.E. (2009) The role of electrontrans-port in the defence response of the South African abalone,Haliotis midae. Fish & Shell¢sh Immunology 26,171^176.
Roux M.M., Pain A., Klimpel K.R. & Dhar A.K. (2002)The lipopolysaccharide and beta-1,3-glucan binding proteingene is upregulated inwhite spot virus-infected shrimp (Pe-naeus stylirostris). Journal ofVirology 76,7140^7149.
Sahoo P.K., Pillai B.R., Mohanty J., Kumari J., Mohanty S. &Mishra B.K. (2007) Invivo humoral and cellular reactionsand fate of injected bacteria Aeromonas hydrophila infreshwater prawn Macrobrachium rosenbergii. Fish &Shell¢sh Immunology 23,327^340.
Sahul Hameed A.S., Yoganandhan K., Sri Widada J.& Bonami J.R. (2004a) Studies on the occurrence of
© 2011 Blackwell Publishing Ltd, Aquaculture Research, 43, 1096–1106 1105
Aquaculture Research, 2012, 43, 1096–1106 Immunomodulation by RdRP protein of MrNV in prawn P K Sahoo et al.

Macrobrachium rosenbergii nodavirus (MrNV) and extrasmall virus-like particles (XSV) associated withwhite taildisease (WTD) of Macrobrachium rosenbergii in India byRT-PCR detection. Aquaculture 238,127^133.
Sahul Hameed A.S.,Yoganandhan K., SriWidada J. & Bona-mi J.R. (2004b) Experimental transmission and tissuetropism of Macrobrachium rosenbergii nodavirus (MrNV)and its associated extra small virus (XSV). Diseases inAquatic Organisms 62,191^196.
ShekharM.S., Azad I.S. & Jithendran K.P. (2006) RT-PCRandsequence analysis of Macrobrachium rosenbergii noda-virus: Indian isolate. Aquaculture 252,128^132.
Shekhar M.S., Sahoo P.K., Dillikumar M. & Das A. (2011)Cloning, expression and sequence analysis of Macrobra-chium rosenbergii nodavirus genes: Indian isolate. Aqua-culture Research, doi: 10.1111/j.1365-2109.2010.02775.x.
Sierra C., Guevara J., Lasscurain R., Perez A., Agundis C.,Zenteno E. & Vazquez L. (2001) Sialylation is modulatedthrough maturation in hemocytes from Macrobrachiumrosenbergii. Comparative Biochemistry and Physiology130,179^189.
Soderhall K. & Cerenius L. (1992) Crustacean immunity. An-nual Review of Fish Diseases 2,3^23.
Srityunlucksana K., Lee S.Y. & Soderhall K. (2002) A b-1,3-glucan binding protein from the black tiger shrimp,Panaeus monodon. Developmental and Comparative Immu-nology 26, 237^245.
Sudhakaran R., Syed Musthaq S., Haribabu P., MukherjeeS.C., Gopal C. & Sahul Hameed A.S. (2006) Experimentaltransmission of Macrobrachium rosenbergii nodavirus(MrNV) and extra small virus (XSV) in three species of
marine shrimp (Penaeus indicus, Penaeus japonicus and Pe-naeus monodon). Aquaculture 257,136^141.
Tripathy S., Sahoo P.K., Kumari J., Mishra B.K., Sarangi N. &Ayyappan S. (2006) Multiplex RT-PCR detection and se-quence comparision of virusesMrNVand XSVassociatedwith white tail disease in Macrobrachium rosenbergii.Aquaculture 258,134^139.
Venegas C.A., Nonaka L., Mushiake K., NishizawaT. & Mur-oga K. (2000) Quasi-immune response of Penaeus japoni-cus against penaeid rod-shaped DNA virus (PRDV).Diseases of Aquatic Organisms 42,83^89.
WangY.C., Chang P.S. & Chen H.Y. (2007) Tissue expressionsof nine genes important to immune defence of the Paci¢cwhite shrimp Litopenaeus vannamei. Fish & Shell¢sh Im-munology 23,1161^1177.
WangY.C., Chang P.S. & Chen H.Y. (2008) Di¡erential time-series expression of immune-related genes of Paci¢cwhite shrimp Litopenaeus vannamei in response to dietaryinclusion of b-1, 3-glucan. Fish & Shell¢sh Immunology24,113^121.
WorkenheT., HoriT.S., RiseM.L., KibengeM.J. & Kibenge F.S.(2009) Infectious salmonanaemiavirus (ISAV) isolates in-duce distinct gene expression responses in the Atlanticsalmon (Salmo salar) macrophage/dendritic-like celllineTO, assessed using genomic techniques.Molecular Immu-nology 46, 2955^2974.
Wu Y.C., Lu Y.F. & Chi S.C. (2010) Anti-viral mechanism ofbarramundi Mx against betanodavirus involves the inhi-bition of viral RNA synthesis through the interference ofRdRp. Fish & Shell¢sh Immunology 28, 467^475.
© 2011 Blackwell Publishing Ltd, Aquaculture Research, 43, 1096–11061106
Immunomodulation by RdRP protein of MrNV in prawn P K Sahoo et al. Aquaculture Research, 2012, 43, 1096–1106




















