
Production of a novel probiotic date juice with anti-proliferative ...
Upload
independentCategory
view
2download
0
Probiotic effect of Bacillus NL110 and Vibrio NE17 on
the survival, growth performance and immune
response of Macrobrachium rosenbergii (de Man)
K M Mujeeb Rahiman1,Yousuf Jesmi1, Ambat P Thomas1 & A A Mohamed Hatha2
1School of Environmental Sciences, Mahatma Gandhi University, Kottayam, India2Department of Marine Biology, Microbiology and Biochemistry, School of Marine Sciences, Cochin University of Science and
Technology, Cochin, India
Correspondence: A A Mohamed Hatha, Department of Marine Biology, Microbiology and Biochemistry, School of Marine Sciences,
Cochin University of Science and Technology, Fine Arts Avenue, Cochin 682016, India. E-mails: [email protected],
Abstract
Eight hundred and eighty-¢ve strains of bacterial iso-lates fromvarious samples associated with the natur-al habitat ofMacrobrachiumrosenbergiiwere screenedfor their probiotic potential. Two putative probiontsnamely Bacillus NL110 andVibrio NE17 isolated fromthe larvae and egg samples, respectively, were se-lected for experimental studies and were introducedto the juveniles of M. rosenbergii (0.080 � 0.001g)through di¡erent modes such as through feed, waterand both. The probiotic potential of the above bacter-ia in terms of improvements inwater quality, growth,survival, speci¢c growth rate (SGR), feed conversionratio and immune parameters was evaluated. Thetreatment groups showed a signi¢cant improvementin SGR and weight gain (Po0.001). Survival amongdi¡erent treatment groups was better than that inthe control group. There were also signi¢cant im-provements in the water quality parameters such asthe concentration of nitrate and ammonia in thetreatment groups (Po0.05). Improvements in im-mune parameters such as the total haemocyte count(Po0.05), phenoloxidase activity and respiratoryburst were also signi¢cant (Po0.001). It is concludedthat screening of the natural micro£ora of cultured¢sh and shell¢sh for putative probionts might yieldprobiotic strains of bacteria that could be utilized foran environment-friendly and organic mode of aqua-culture.
Keywords: Bacillus,Vibrio, M. rosenbergii, probio-tics, immune enhancement, water quality
Introduction
During the past three decades, aquaculture has be-come the fastest-growing food-producing sector andis contributing signi¢cantly to national economic de-velopment, global food supply and food security.Freshwater prawns such as Macrobrachium rosenber-gii, which is a preferred species due to several biologi-cal characteristics, can be produced in inlandlocations in closer proximity to large urban markets.M. rosenbergii also appears to be resistant to most ofthe viral diseases that have impacted marine shrimp(Wang, Lo, Chang & Kou1998). Bacterial diseases areconsidered to be a menace in the intensive larval cul-ture and production of aquaculture species owing tothe large-scale mortality they cause in hatcheriesand culture systems (Singh 1990; Grisez & Ollevier1995; Cheng & Chen1998). Although antibiotics haveplayed a major role in combating many diseases ofcultured aquatic organisms, their indiscriminate usein aquaculture has led to undesirable consequencessuch as development of antibiotic-resistant bacteriaand persistence of antibiotic residues in farm-raisedshrimp, etc. (Chaithanya, Nayak & Venugopal1999).Recently, the use of probiotics has become a popu-
lar and an environment-friendly alternative toantibiotics to improve and maintain a healthyenvironment for aquaculture. Because shrimp andprawn possess a non-speci¢c immune response, pro-biotic treatments may provide a broader spectrumand greater non-speci¢c disease protection throughcompetitive exclusion and immune modulation(Rengpipat, Rukpratanporn, Piyatiratitivorakul &
Aquaculture Research, 2010, 41, e120^e134 doi:10.1111/j.1365-2109.2009.02473.x
r 2010 TheAuthorse120 Journal Compilationr 2010 Blackwell Publishing Ltd

Menasaveta 2000). Several genera of bacteria such asBacillus, Lactobacillus,Vibrio, Streptococcus, Alteromo-nas, Aeromonas and Nitrosomonas have been used inthe aquaculture practices (Gatesoupe 1994; Rengpi-pat et al. 2000; Venkat, Sahu & Jain 2004; Balcazar,Rojas-Luna & Cunningham 2007; Aly, Abd-El-Rah-man, John & Mohamed 2008).Bacillus andVibrio sp. are used widely as probiotics
in freshwater aquaculture for improvements in survi-val, growthand development (Ringo & Vadstein1998;Huys, Dhert, Robles, Ollevier, Sorgeloos & Swings2001; Kumar, Mukherjee, Prasad & Pal 2006; Keysa-mi, Saad, Sijam, Daud & Alimon 2007; Apun-Molina,Miranda, Gonzalez, Martinez-Diaz & Rojas-Contreras2009). They are also used for enhancement in inhibi-tion of pathogenic bacteria (Austin, Stuckey, Robert-son, E¡endi & Gri⁄th 1995; Gatesoupe 1997; Huyset al. 2001), enhancing enzyme activity (Kumar et al.2006; Ghosh, Sinha & Sahu 2008), better feed con-version, fecundity and fry production (Wang & Xu2006; Ghosh, Sinha & Sahu 2007) as well as for im-munity enhancement and disease resistance (Irianto& Austin 2002; Aly, Mohamed & John 2008). How-ever, relatively less work has been directed towardsthe selection and development of probiotic bacteriafor the culture of M. rosenbergii. Therefore, the aimof the present research is to screenputative probioticsfrom the native micro£ora associated with M. rosen-bergii and to evaluate their probiotic potential. Twobacterial strains such as Bacillus NL110 and VibrioNE17 were evaluated in terms of their antibacterialactivity to pathogens in a culture system, pathogeni-city to the postlarvae (PL), ability to improve waterquality parameters and to enhance the growth, im-munity and survival of the juveniles of giant fresh-water prawnM. rosenbergii.
Materials and methods
Bacterial isolates used for probiotic screening
Four samples each of water, sediment, intestine ofM.rosenbergii, eggs, larvae, PL and feed samples wereanalysed for isolation of bacteria. A total of 885 iso-lates of bacteria, which included 131 isolates fromwater,114 from sediment,155 from intestine ofM. ro-senbergii,133 from eggs, 206 isolates from larvae andpostlarvae ofM. rosenbergii and146 isolates from feedsamples, were characterized to the genus level andevaluated for probiotic potential. All the sampleswere collected from Vembanad lake, an importantRamsar site in India and the home ground of
M. rosenbergii. The isolates were characterized up tothe generic level using the taxonomic key byMuroga,Higashi and Keetoku (1987); Barrow and Feltham(1993) and Holt, Krieg, Sneath, Staley andWillliams(2000).
Selection of probiotic bacterial strains
All the 885 bacterial isolates were subjected to preli-minary screening for the selection of putative probio-tic strains, based on their antibacterial activityagainst¢sh, prawnand humanpathogens.The source and de-tails of the bacterial pathogens used are given below.
Bacterial pathogens used
In order to determine the antibacterial activity of theprobiotic strains, various ¢sh, prawn and humanpathogens were used. Fish pathogen Aeromonas hy-drophila (MTCC 646) and prawn pathogens such asVibrio parahaemolyticus (MTCC 451) andVibrio vulni-¢cus (MTCC 146) were obtained from the culturecollection of Institute of Microbial Technology,Chandigarh, India.Vibrio harveyi (prawn pathogen)was obtained from the culture collection of Schoolof Environmental Studies, Cochin University ofScience and Technology, India. Human pathogenssuch as Escherichia coli, Salmonella newport and Sal-monella typhi were obtained from the culture collec-tion maintained at the School of EnvironmentalSciences, Mahatma Gandhi University, India.
Antibacterial activity of isolates using the well di¡usionand cross-streak method
In order to determine the antibacterial activity of theprobiotic strains using the well di¡usion method(Chythanya, Karunasagar & Karunasagar 2002),lawn cultures of ¢sh, prawn and human pathogenssuch as A. hydrophila,V. parahaemolyticus,V. harveyi,V. vulni¢cus, E. coli, S. newport and S. typhi were pre-pared by pouring 2mL of a 16^18-h-old tryptic soyabroth (TSB) culture of the above microorganismsover sterile tryptic soya agar (TSA) plates. Excess li-quid was drained o¡ and the plates were air dried inan incubator (30 1C) for 15min. Using a sterile gel,puncher wells (3mm diameter) were punched intothe plates. Thirty microlitres of an18 h culture of thebacterial isolates (885 numbers) in TSB was pipettedinto the wells and plates were incubated for 24 h at30 1C. The presence of antibacterial activity wasnoted as a clear zone around the wells.
Aquaculture Research, 2010, 41, e120^e134 Probiotic e¡ect of bacteria onM. rosenbergii K MMujeeb Rahiman et al.
r 2010 TheAuthorsJournal Compilationr 2010 Blackwell Publishing Ltd, Aquaculture Research, 41, e120^e134 e121

Only those isolates that showed antibacterial ac-tivity to the selected ¢sh, prawn and human patho-gens in the well di¡usion method were selected forthe study of antibacterial activity using the cross-streak method (Chythanya et al. 2002). This includedfour bacterial isolates namely Bacillus NL110,VibrioNE17, AeromonasNE2 andVibrioNL40. For the detec-tion of antibacterial activity by a cross-streak assay,an 18 h culture of the above bacterial strains wasstreaked as a 2 cm thick band across the diameter oftheTSA plate. After incubation at 30 1C for 24 h, thegrowth was scraped with a sterile slide. The remain-ing bacteria were killed by exposure to 5mL chloro-form poured on the glass lid and left for 15min bykeeping the medium inverted over the lid. The plateswere then air dried for about 10min to remove anyresidual chloroform, and A. hydrophila,V. parahaemo-lyticus,V. harveyi,V. vulni¢cus, E. coli, S. newport andS. typhi were streaked perpendicular to the bacterialstrain band using a sterile glass rod dipped in an18-h-old culture. The plates were incubated at 30 1C for24 h.The presence of antibacterial activity was notedas a linear clear zone. Two isolates namely BacillusNL110 andVibrio NE17, which showed good antibac-terial activity against the tested pathogens in boththe methods, were selected for further screening.
Pathogenicity of probiotic strains to PL ofM. rosenbergii
Broth cultures (18 h old) of selected probiotic strains(Bacillus NL110 and Vibrio NE17) were prepared inTSB. The cells were harvested by centrifugation at1509 g for 15min, washed in physiological salineand the cells were resuspended in 10mL sterile phy-siological saline. The cell numbers in the suspensionwere determined using the spread plate method byplating serial 10-fold dilution in TSA. From8.35 � 108 cells mL�1 of Bacillus NL110 and7.90 � 108 cells mL�1 Vibrio NE17 in Physiologicalsaline, 0.1mL was added individually at the begin-ning of the experiment to each of the1L beaker con-taining 500mL ¢lter-sterilized (0.45 mm) freshwaterto obtain105 cells mL�1of potential probiotic bacter-ia (Chythanya et al. 2002; Ravi, Musthafa, Je-gathammbal, Kathiresan & Pandian 2007). To eachbeaker, 25 healthy PL of M. rosenbergii (PL-20,clearedWSSVnegative by polymerase chain reactionand showing normal healthy behaviour) collectedfrom a private-owned hatchery, Rosen Fisheries, Ker-ala, India, were introduced. The pathogenicity of the
probiotic strains to PL of M. rosenbergii were evalu-ated by visual observation, swimmingactivityas wellas microscopically for any signs of infection or mor-tality up to 96 h. The experiment was performed induplicate with each bacterial strain, and the controlwas maintained without any bacterial inoculums.
In£uence of pH, temperature and salinity onthe antibacterial activity of probiotic strains
To determine the e¡ect of pHonantibacterial activity,sterileTSB adjusted to pH 5,6,7 and 8was inoculatedwith 0.1mL of an18-h-old culture of selected probio-tic strains (Bacillus NL110 andVibrioNE17) and incu-bated for 24 h at 30 1C. Similarly, the e¡ects oftemperature on antibacterial activity were deter-mined at various temperatures such as10, 20,30 and37 1C, and the e¡ect of salinity on antibacterial activ-ity was determined at salinities such as 10,15, 20, 30and 35 g L�1. After incubation, the probiotic poten-tial of the bacterial strains was tested againstA. hydrophila, V. parahaemolyticus, V. harveyi andV. vulni¢cus using the well di¡usion method.
Experimental set-up
The PL (PL-30) of M. rosenbergii were grown up to0.080 � 0.001g in a rectangular ¢bre-reinforcedplastic (FRP) tank of 1000 L capacity, supplementedwith arti¢cial aeration. Fifteen juveniles of uniformsize were placed in a 50 L plastic tank containing30 L water and were acclimatized for 1 week beforethe trials began. The juveniles were fed twice daily at10% body weight per day for a period of 60 days.Waste removal was carried out by exchanging water(2 L) from every tank once in a week. Nine experi-mental groups with probiotics and a control, all induplicate, were maintained (Table 1). The controlgroup was fed with normal feed, which was free ofany probiotic bacterial inoculation.
Preparation and addition of probiotics
Broth cultures of Bacillus NL110 and Vibrio NE17were prepared in TSB and Lactobacillus plantarum(MTCC1325) in Lactobacillus MRS broth by inoculat-ing the isolates into respective broths and incubatingat room temperature for 24 h. For the addition of pro-biotics throughwater,5mL broth cultures of BacillusNL110 (6.90 � 0.34 � 109 cells mL�1) were directly
Probiotic e¡ect of bacteria onM. rosenbergii K MMujeeb Rahiman et al. Aquaculture Research, 2010, 41, e120^e134
r 2010 TheAuthorse122 Journal Compilationr 2010 Blackwell Publishing Ltd, Aquaculture Research, 41, e120^e134

inoculated to Tanks T2 & T3, Vibrio NE17(7.52 � 0.74 � 109 cells mL�1) to tanks T5 & T6 andL. plantarum (7.41 � 0.54 � 109 cells mL�1) to tanksT8 & T9 once in a week to obtain an approximateconcentration of the probiotic bacteria up to106 cells mL�1of water in the concerned experimen-tal tank.For preparing the probiotic incorporated feed, the
cells of Bacillus NL110,Vibrio NE17 and L. Plantarum,after a 24-h incubation, were harvested by centrifu-gation at 1509 g for 15min. The cells were washedthrice and resuspended in physiological saline. Thecells were thoroughly mixed with the commercialscampi feed (Higashimaru Feeds, Aleppy, India) toobtain109 probiotic bacteria g�1 in feed (Suralikar &Sahu 2001; Nejad, Rezaei,Takami, Lovett, Mirvaghe¢& Shakouri 2006), and the mixture was spread outand aseptically dried in an oven for 1^2 h at 50oC.The bacterial count was checked periodically ata 15-day interval using the spread plate method,and the count was 4.73 � 2.87 � 109 colony-formingunit (CFU) g�1 in the Bacillus NL110-incorpo-ratedfeed, 4.34 � 1.22 � 109 CFUg�1 in the VibrioNE17-incorporated feed, 3.16 � 2.35 � 109 CFUg�1
in the L. plantarum-incorporated feed and 5.01 �1.78 � 105 CFUg�1 in the normal feed. Feed wasstored in clean, plastic bottles at 4oC and provided tothe respective experimental tanks. Probiotic-incor-porated feeds were prepared twice weekly.
Physico-chemical parameters
Water samples were collected from the experimentaltanks on a weekly basis and analysed for changes in
the water quality parameters such as pH, tempera-ture, dissolved oxygen (DO), ammonia and nitrite.The pH of the water sample was analysed using anelectronic pH meter (Systronics, m pH System 361,Chennai, India), temperature using a temperatureprobe (Systronics, m pH System 361), DO using theWinkler method, ammonia using the phenol^hypo-chlorite method and nitrite using the colorimetricmethod as per the American Public Health Associa-tion (APHA1998).
Bacteriological analysis
Preparation of samples
For bacteriological analysis, water samples from theexperimental tanks were collected at a 15-day inter-val and were serially diluted aseptically up to 10�4
using sterile-distilledwater. For the analysis of the in-testinal micro£ora of juveniles in the initial stage, thespecimens were surfacewashed in a sterile 0.1% ben-zalconium chloride solution and then thoroughlyrinsed in sterile-distilled water. The samples werethen ¢ltered using a sterile ¢lter paper and excesswater was removed using a sterile blotting paper.The PL samples were then homogenized in a sterileall-glass homogenizer and serially diluted up to10�5. For analysis of the intestinal micro£ora of juve-niles of M. rosenbergii at the ¢nal stage, the intestinewas removed aseptically, homogenized ina sterile all-glass homogenizer and serially diluted up to 10�5.For the analysis of feed samples, the feed sampleswere aseptically weighed and homogenized in a ster-ile all-glass homogenizer and serially diluted up to10�7.
Table 1 Details of the experimental groups showing the type of treatment, mode, dose and period of application
Groups Type of treatment Mode of application Doseand period of application
T1 Bacillus NL110 through feed Through feed 4.73 � 2.87 � 109 CFU g�1 fed twice in a day
T2 Bacillus NL110 through water Through water 1.15 � 0.56 � 106 CFU mL� 1 once in a week
T3 Bacillus NL110 through feed and water Through feed and water 4.73 � 2.87 � 109 CFU g�1 fed twice in a day
1.15 � 0.56 � 106 CFU mL� 1 once in a week
T4 Vibrio NE17 through feed Through feed 4.34 � 1.22 � 109 CFU g�1 fed twice in a day
T5 Vibrio NE17 through water Through water 1.25 � 1.23 � 106 CFU mL� 1 once in a week
T6 Vibrio NE17 through feed and water Through feed and water 4.34 � 1.22 � 109 CFU g�1 fed twice in a day
1.25 � 1.23 � 106 CFU mL� 1 once in a week
T7 Lactobacillus plantarum through feed Through feed 3.16 � 2.35 � 109 CFU g�1 fed twice in a day
T8 L. plantarum through water Through water 1.24 � 0.93 � 106 CFU mL� 1 once in a week
T9 L. plantarum through feed and water Through feed and water 3.16 � 2.35 � 109 CFU g�1 fed twice in a day
1.24 � 0.93 � 106 CFU mL� 1 once in a week
Control Without probiotic application – –
CFU, colony-forming unit.
Aquaculture Research, 2010, 41, e120^e134 Probiotic e¡ect of bacteria onM. rosenbergii K MMujeeb Rahiman et al.
r 2010 TheAuthorsJournal Compilationr 2010 Blackwell Publishing Ltd, Aquaculture Research, 41, e120^e134 e123

Estimation of the total viable count
Aliquots of 0.2mL samples from each dilution ofwater samples, intestinal samples and feed sampleswere spread plated in triplicate on TSA for the enu-meration of the total aerobic heterotrophic bacteria,expressed as total viable count (TVC).The plates werethen incubated at 30 1C for 24^48 h. After incuba-tion, plates with 30^300 CFUwere selected for count-ing and expression of TVC of bacteria.
Growth performance of juveniles
Juveniles were weighed initially and at the end of theexperiment to assess the growth performance interms of speci¢c growth rate (SGR) and the feed con-version ratio (FCR). SGR and FCR were calculatedusing the following formula:
SGR ¼ ln final bodywt� ln initial bodywt60 days
� 100
where, ln5natural log
FCR ¼ Quantity of feed consumedTotalweight gain
Immunological parameters of the treatedjuveniles
Total haemocyte count (THC)
Haemolymph (HL) from each prawn was collectedfrom the rostral sinus cavity using a 26 G needle anda1mL syringe containing an anticoagulant. HL wascollected from two randomly selected juveniles fromeach treatment group at 5 p.m. Haemocytes werecounted using a haemocytometer and expressed ascells mL�1of HL.
Phenoloxidase (PO) activity
Phenoloxidase activity was estimated spectrophoto-metrically using L-3, 4-dihydroxyphenyl-alanine(L-DOPA) as a substrate (Soderhall1981). HL in anan-ticoagulant (100 mL) was incubated for 30min atroom temperature after adding100 mL of sodium do-decyl sulphate (SDS) in a small cuvette. After the in-cubation, 1000 mL of enzyme substrate L-DOPA wasadded and the optic density was measured at 490nm. Absorbance measurements were performedagainst a blank consisting of SDS and L-DOPA. Oneunit of enzyme activity was de¢ned as an increasein absorbance min�1100 mLHL�1.
Respiratory burst
The respiratory burst or super oxide generation (O2� )
of haemocytes was quanti¢ed using a nitroblue tetra-zolium (NBT) assay (Song & Hsieh 1994). HL in theanticoagulant (100 mL) was incubated for 30min at10oC after adding 100 mL of NBT salt solution. Afterincubation, the mixture was centrifuged at 1610 gfor 10min. After removing the supernatant solution,the cells were washed with phosphate-bu¡ered sal-ine. Then, 100 mL of 100% methanol was added andincubated at room temperature for 10min. The solu-tionwas then centrifuged again at 3622 g for10min.The supernatant was removed and the tubes were airdried for 30min at room temperature. The pellet wasthen rinsed with 50% methanol three to four timesand coated with 120 mL KOH (2M) and 140 mL di-methyl sulphoxide (DMSO) solution to dissolve theformazan. The optical absorbances of the mixtureswere read at 620 nm. The results were expressed asan increase in absorbance100 mLHL�1.
Statistical analysis
The datawere analysed by two-factor analysis of var-iance (ANOVA) using the statistical tool package of MI-
CROSOFT OFFICE EXCEL 2007 software. Wherever thetreatments were found to be signi¢cant, least signi¢-cance was calculated and signi¢cant treatmentswere identi¢ed.Whenever necessary, the results arepresented as the average and standard deviation.
Results and Discussion
Selection of bacterial isolates with probioticpotential
From a total of 885 bacterial isolates from a naturalenvironment and di¡erent life stages of M. rosenber-gii, four were selected initially based on their antibac-terial activity against ¢sh, prawn and humanpathogens. The pathogenicity of the selected patho-gens was well documented by several researchers.Even though Aeromonas spp. are considered to be op-portunistic pathogen in the hatchery and culture en-vironment of M. rosenbergii (New 1995), they havebeen linked to outbreaks of disease like burn spot dis-ease (El-Gamal, Alderman, Rodgers, Polglase &Macintosh 1986), black-spot, bacterial necrosis andgill obstruction in larvae (Lombardi & Labao1991a, b) and septicaemia under adverse conditions(Sung, Hwang & Tasi 2000). Vibrio sp. including
Probiotic e¡ect of bacteria onM. rosenbergii K MMujeeb Rahiman et al. Aquaculture Research, 2010, 41, e120^e134
r 2010 TheAuthorse124 Journal Compilationr 2010 Blackwell Publishing Ltd, Aquaculture Research, 41, e120^e134

V. parahaemolyticus,V. harveyi andV. vulni¢cus are themost important bacterial pathogens of culturedshrimp and prawn responsible for a number of dis-eases and mortalities (Lightner 1983; Chen, Hanna,Altman, Smith, Moon & Hammond 1992; Lavilla-Pi-togo, Leano & Paner1998; Martin, Rubin & Swanson2004; Khuntia, Das, Samantaray, Samal & Mishra2008). The ability of human pathogens such as E. coliand Salmonella to cause gastrointestinal symptomssuch as fever, diarrhoea, abdominal pain, nauseaand vomiting in human is also well documented(Bhan, Bahl & Bhatnagar 2005; Ministry of Health &Welfare 2006; Hamner, Broadway, Mishra, Tripathi,Mishra, Pulcini, Pyle & Ford 2007; Chugh 2008;Woc-Colburn & Bobak 2009).The results of the antibacterial activity of the bac-
terial strains isolated from the natural environmentofM. rosenbergii against the pathogenic bacteria (Ta-ble 2) revealed that the two bacterial strains, BacillusNL110 isolated from larvae andVibrio NE17 from eggof M. rosenbergii, had better probiotic potential thanAeromonas NE2 from the eggs andVibrio NL40 fromthe larvae of M. rosenbergii. Bacillus NL110 showedantibacterial activity against all the tested ¢sh andprawn pathogens, whileVibrioNE17 showed antibac-terial activity against 3 of them. Our observations onthe antibacterial activity of Bacillus to pathogens of¢sh and prawn were similar to those previously re-ported by Balcazar and Rojas-Luna (2007). Aquaticcandidate probionts for larviculture isolated fromadults (Rengpipat et al. 2000; Riquelme, Araya &Escribano 2000; Gullian, Thompson & Rodriguez2004) and healthy larvae (Gatesoupe 1997; Ringo &Vadstein 1998) have been reported previously. It hasbeen suggested that the e⁄cacy of probiotics is likelyto be highest in the host species from where theywere isolated (Verschuere, Rombaut, Sorgeloos &
Verstraete 2000). It is also reported that the non-pathogenic strains of knownpathogenic bacteria likeV. alginolyticus, A. media and A. hydrophila alsoshowed antibacterial activity and have been used asa probiotic inalgal production, shrimp culture, oysterculture and ¢sh culture (Austin et al. 1995; Gibson,Woodworth & George 1998; Gomez-Gil, Roque & Ve-lasco-Blanco 2002; Irianto & Austin 2002). It is possi-ble that when pathogenicity is suppressed or lost,other factors such as growth rate or attachment abil-ity, which are factors that contribute to their successas pathogens, may in£uence the micro£ora to thebene¢t of its host.
Pathogenicity of probiotic strains toM. rosenbergii PL
Figure 1 shows the pathogenicity e¡ect of BacillusNL110 andVibrio NE17 on M. rosenbergii PL. The re-sults showed that Bacillus NL110 and Vibrio NE17have no pathogenic e¡ect on the PL ofM. rosenbergii.The survival of PL was higher in both experimentalgroups when compared with the control (Po0.01).One of the most important criteria for a candidate tobe used in biocontrol is that the organism should benon pathogenic to the host. Because the probioticstrainBacillusNL110was isolated fromM. rosenbergiilarvae andVibrio NE17 from the eggs collected fromthe natural habitat, these bacterial strains mighthave already been selected by the host larvae becauseof their positive e¡ects on them. Similar ¢ndings onprobiotic strains were reported earlier (Chythanyaet al. 2002; Vijayan, Bright Singh, Jayaprakash, Ala-vandi, Somnath Pai, Preetha, Rajan & Santiago2006).
Table 2 Results of antibacterial activity by potential probiotic bacteria against di¡erent pathogenic bacteria
Pathogens tested
Antibacterial activity of potential probiotic strains
Aeromonas NE2 Vibrio NE17 Vibrio NL40 Bacillus NL110
WDM CSM WDM CSM WDM CSM WDM CSM
Aeromonas hydrophila X X X X X X X X
Vibrio parahaemolyticus X X X X X X X X
Vibrio harveyi – – – – – – X X
Vibrio vulnificus – – X X – – X X
Escherichia coli X X X X – – X X
Salmonella newport – – X X X X – –
Salmonella typhi – – – – – – X X
WDM, well di¡usion method; CSM, cross-streak method, X, positive for antibacterial activity.
Aquaculture Research, 2010, 41, e120^e134 Probiotic e¡ect of bacteria onM. rosenbergii K MMujeeb Rahiman et al.
r 2010 TheAuthorsJournal Compilationr 2010 Blackwell Publishing Ltd, Aquaculture Research, 41, e120^e134 e125

In£uence of growth conditions on theantibacterial activity of probiotic strains
The growth and antibacterial activities of probioticstrains Bacillus NL110 and Vibrio NE17 against 4pathogenic bacteria at di¡erent pH, temperature andsalinities are presented in Table 3. Bacterial isolatesdid not show growth at pH 5 and at 10 1C. However,the selected strains did not show antibacterial activ-ity againstV. harveyi andV. vulni¢cus at pH 6. At pH 7and 8, Bacillus NL110 showed antibacterial activityagainst all the tested pathogens, while Vibrio NE17showed antibacterial activity only against A. hydro-phila,V. parahaemolyticus andV. vulni¢cus.Bacillus NL110 showed antibacterial activity
against all the 4 pathogens in a wide range of tem-peratures such as 20, 30 and 37 1C, except againstV. harveyi at 20 1C. Unlike Bacillus NL110,VibrioNE17exhibited antibacterial activity against A. hydrophila,V. parahaemolyticus and V. vulni¢cus only at 30 and37 1C. Bacillus NL110 and Vibrio NE17 showed goodgrowth and antibacterial activity at all the salinitiestested in the present study. Because the probioticstrains have to be used in the ¢eld under di¡erent en-vironmental conditions, the ability of the strains togrow over awide salinity and temperature range hashuge signi¢cance. The study revealed that both theprobiotic strains can be used in freshwater as well asin marine aquaculture under a wide temperatureand salinity range. The range of environmental con-ditions over which the organisms in the presentstudy could grow and show activity was wider thanthose reported by Chythanya et al. (2002), who stu-died the production of the antibacterial componentof the Pseudomonas I-2 strainunder di¡erent environ-mental conditions. Our ¢ndings were also supportedby the observations of Jayaprakash, Pai, Anas, Pre-etha, Philip and Singh (2005) and Vijayan et al. Ta
ble
3Growth
andan
tibacterialactivityof
Bacillus
NL110
andVibrioNE17at
di¡erent
pH,tem
peraturesan
dsalin
ities
Potential
prob
iotic
strains
pHGrowth
Antibac
teria
lactivity
against
Tempe
rature
(1C)
Growth
Antibac
teria
lactivity
against
Salinity
(mgL�1 )
Growth
Antibac
teria
lactivity
against
A.hydroph
ilaV.pa
raha
emolyticus
V.ha
rveyi
V.vulnificus
A.hydroph
ilaV.pa
raha
emolyticus
V.ha
rveyi
V.vulnificus
A.hydroph
ilaV.pa
raha
emolyticus
V.ha
rveyi
V.vulnificus
Bacill
us
NL110
5�
––
––
10
�–
––
–10
1X
XX
X
61
XX
–X
20
1X
X–
X15
1X
XX
X
71
XX
XX
30
1X
XX
X20
1X
XX
X
81
XX
XX
37
1X
XX
X30
1X
XX
X
35
1X
XX
X
Vib
rio
NE
17
5�
––
––
10
�–
––
–10
1X
X–
X
61
XX
––
20
1X
X–
–15
1X
X–
X
71
XX
–X
30
1X
X–
X20
1X
X–
X
81
XX
–X
37
1X
X–
X30
1X
X–
X
35
1X
X–
X
X,p
ositiveforan
tiba
cteriala
ctivity,
1,indicatesgrow
th.
A.h
ydrophila,Aerom
onas
hydrophila;V.p
arahaemolyticus,Vibrioparahaem
olyticus;V.h
arveyi,Vibrioharveyi;V.
vulni¢cus,Vibriovulni¢cus.
70
75
80
85
90
95
100
105
0 12 24 36 48 72 96
% s
urvu
val
Time in hourControl Bacillus NL110 Vibrio NE17
Figure 1 E¡ects of Bacillus NL110 andVibrioNE17 on thesurvival of PL ofMacrobrachium rosenbergii.
Probiotic e¡ect of bacteria onM. rosenbergii K MMujeeb Rahiman et al. Aquaculture Research, 2010, 41, e120^e134
r 2010 TheAuthorse126 Journal Compilationr 2010 Blackwell Publishing Ltd, Aquaculture Research, 41, e120^e134

(2006), who suggested the possibility of developing abiological method of suppressing vibrios associatedwith prawn larval rearing systems.
Physico-chemical parameters of water
As all the treatment and control tanks were main-tained at room temperature at controlled aeration;there was no signi¢cant variation in the averagetemperature and DO (29.5^29.8 1C and 6.57^6.84mg L�1) among the di¡erent treatment groupsand control. The water pH of all the treatment groupswas slightly lower than that of the control (7.78), andthe lowest average pH was observed from the T8group (7.60). The observed physico-chemical para-meters of the water such as temperature, DO and pHof the water from the treatment tanks were withinthe optima for these factors forM. rosenbergii culture.BecauseM. rosenbergii could tolerate a wide range oftemperature (14^35 1C), pH (7.0^8.5) and salinity le-vels (0^25 g L�1), the minor variations in the treat-ment groups did not seem to a¡ect the survival ofthe organism. The results of the present study werecomparable to those of Venkat et al. (2004), who hadreported on the growth and survival of PL of M. ro-senbergiiwith a Lactobacillus-based probiotic feed.The ammonia concentration of the water from
the treatment tanks ranged from 0.0125 to0.0985mg L�1 and the nitrite concentration from0.0205 to 0.1200mg L�1 during the experimentalperiod, and there were signi¢cant di¡erences in theaverage ammonia (0.042^0.056mg L�1) and nitrite(0.054^0.075mg L�1) concentration among di¡erenttreatments (Fig.2, Po0.05).The ammonia concentra-tion of water was the lowest in theT4 group, followedby theT1andT2 groups. Signi¢cantly lower values ofnitrite were observed in theT5,T6,T2 and T3 groupsrespectively.The reduction in ammonia and nitrite inthe probiotic-treated tanks may be because of the ac-tion of probiotic strains Bacillus NL110 and VibrioNE17. Bioremediation of water and biocontrol ofpathogens (Queiroz & Boyd 1998; Gatesoupe 1999;Skjermo & Vadstein 1999) are the 2 major modes ofaction of administering bene¢cial bacteria in the cul-ture water.When commercial microbial products areused to treat culture wastewater, heterotrophic bac-terial assimilation could be the main and powerfulmechanism for organic matter removal and conver-sion of potentially toxic inorganic nitrogen into rela-tively stable organic nitrogen. It has been reportedthat the use of Bacillus spp. improved the water qual-
ity, survival and growth rates and increased thehealth status of juvenile P. monodon (Dalmin, Kathir-esan & Purushothaman 2001). Decamp, Moriartyand Lavens (2008) reported that improved waterquality was as a major factor for better performancein biomass increase and concluded that probioticbacteria such as Bacillus spp. have the potential to in-duce growth enhancement comparable to that ob-tained with antimicrobials, and importantly, in acost-e¡ective manner.
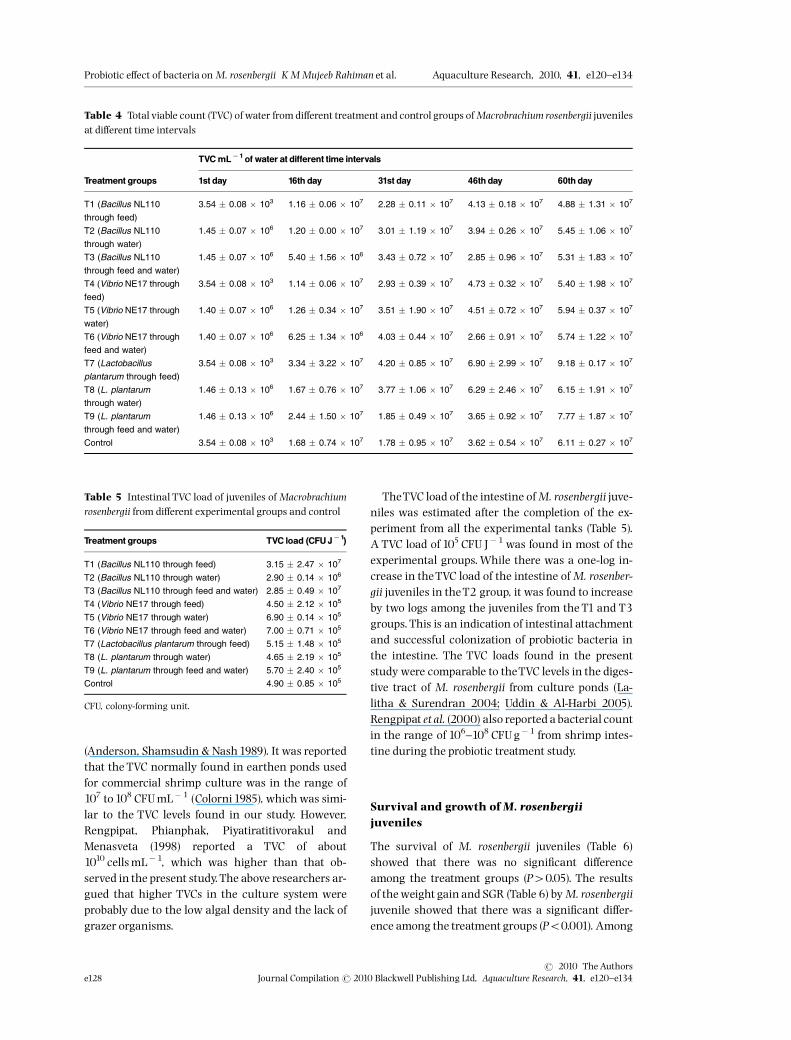
TVC from various samples
The initial TVC load of culture water, commercialscampi feed and the intestine of M. rosenbergiiwere 3.54 � 0.08 � 103 CFUmL�1, 4.95 � 0.07 �105 CFUg�1 and 1.26 � 0.34 � 103 CFU Juvenile�1
respectively. After incorporating the probiotic, theTVC of culture water was 106 CFUmL�1 and that ofthe feed was 109 CFUg�1. There were no signi¢cantdi¡erences between the bacterial counts among theprobiotic-incorporated culture water and prawn feed(P40.05). Thereafter, the TVC load of the rearingwater was analysed at a 15-days interval, up to 60days, in the di¡erent experimental groups (Table 4).There were four-log increases in theTVC of the waterfrom the control tank and the rearing tank waterwhere probiotics were introduced through feed.There was an increase of one log in the TVC of therearing tank water in which probiotics were intro-duced throughwater.There was a signi¢cant di¡erence in TVC during
the culture period (Po0.001). The lowest TVC levelswere observed on the ¢rst and the 16th day, whilethe highest TVC load was found on the 60th day. The£uctuation in the range of the THB load was depen-dent on the quality of feed, removal of unconsumedfeed and the method used in the culture system
0.000.010.020.030.040.050.060.070.080.090.10
Cont. T1 T2 T3 T4 T5 T6 T7 T8 T9Experimental set up
Ammonia Nitrite
mg
L–1
Figure 2 Average ammonia and nitrite values of Macro-brachium rosenbergii juvenile rearing water where probio-tic strains were introduced through di¡erent routes.
Aquaculture Research, 2010, 41, e120^e134 Probiotic e¡ect of bacteria onM. rosenbergii K MMujeeb Rahiman et al.
r 2010 TheAuthorsJournal Compilationr 2010 Blackwell Publishing Ltd, Aquaculture Research, 41, e120^e134 e127
(Anderson, Shamsudin & Nash1989). It was reportedthat the TVC normally found in earthen ponds usedfor commercial shrimp culture was in the range of107 to108 CFUmL�1 (Colorni1985), whichwas simi-lar to the TVC levels found in our study. However,Rengpipat, Phianphak, Piyatiratitivorakul andMenasveta (1998) reported a TVC of about1010 cells mL�1, which was higher than that ob-served in the present study.The above researchers ar-gued that higher TVCs in the culture system wereprobably due to the low algal density and the lack ofgrazer organisms.
TheTVC load of the intestine ofM. rosenbergii juve-niles was estimated after the completion of the ex-periment from all the experimental tanks (Table 5).A TVC load of 105 CFU J�1 was found in most of theexperimental groups.While there was a one-log in-crease in theTVC load of the intestine ofM. rosenber-gii juveniles in theT2 group, it was found to increaseby two logs among the juveniles from the T1 and T3groups. This is an indication of intestinal attachmentand successful colonization of probiotic bacteria inthe intestine. The TVC loads found in the presentstudy were comparable to theTVC levels in the diges-tive tract of M. rosenbergii from culture ponds (La-litha & Surendran 2004; Uddin & Al-Harbi 2005).Rengpipat et al. (2000) also reported a bacterial countin the range of 106^108 CFUg�1 from shrimp intes-tine during the probiotic treatment study.
Survival and growth of M. rosenbergiijuveniles
The survival of M. rosenbergii juveniles (Table 6)showed that there was no signi¢cant di¡erenceamong the treatment groups (P40.05). The resultsof theweight gain and SGR (Table 6) byM. rosenbergiijuvenile showed that there was a signi¢cant di¡er-ence among the treatment groups (Po0.001). Among
Table 4 Total viable count (TVC) of water from di¡erent treatment and control groups ofMacrobrachium rosenbergii juvenilesat di¡erent time intervals
Treatment groups
TVCmL� 1of water at different time intervals
1st day 16th day 31st day 46th day 60th day
T1 (Bacillus NL110
through feed)
3.54 � 0.08 � 103 1.16 � 0.06 � 107 2.28 � 0.11 � 107 4.13 � 0.18 � 107 4.88 � 1.31 � 107
T2 (Bacillus NL110
through water)
1.45 � 0.07 � 106 1.20 � 0.00 � 107 3.01 � 1.19 � 107 3.94 � 0.26 � 107 5.45 � 1.06 � 107
T3 (Bacillus NL110
through feed and water)
1.45 � 0.07 � 106 5.40 � 1.56 � 106 3.43 � 0.72 � 107 2.85 � 0.96 � 107 5.31 � 1.83 � 107
T4 (Vibrio NE17 through
feed)
3.54 � 0.08 � 103 1.14 � 0.06 � 107 2.93 � 0.39 � 107 4.73 � 0.32 � 107 5.40 � 1.98 � 107
T5 (Vibrio NE17 through
water)
1.40 � 0.07 � 106 1.26 � 0.34 � 107 3.51 � 1.90 � 107 4.51 � 0.72 � 107 5.94 � 0.37 � 107
T6 (Vibrio NE17 through
feed and water)
1.40 � 0.07 � 106 6.25 � 1.34 � 106 4.03 � 0.44 � 107 2.66 � 0.91 � 107 5.74 � 1.22 � 107
T7 (Lactobacillus
plantarum through feed)
3.54 � 0.08 � 103 3.34 � 3.22 � 107 4.20 � 0.85 � 107 6.90 � 2.99 � 107 9.18 � 0.17 � 107
T8 (L. plantarum
through water)
1.46 � 0.13 � 106 1.67 � 0.76 � 107 3.77 � 1.06 � 107 6.29 � 2.46 � 107 6.15 � 1.91 � 107
T9 (L. plantarum
through feed and water)
1.46 � 0.13 � 106 2.44 � 1.50 � 107 1.85 � 0.49 � 107 3.65 � 0.92 � 107 7.77 � 1.87 � 107
Control 3.54 � 0.08 � 103 1.68 � 0.74 � 107 1.78 � 0.95 � 107 3.62 � 0.54 � 107 6.11 � 0.27 � 107
Table 5 Intestinal TVC load of juveniles of Macrobrachiumrosenbergii from di¡erent experimental groups and control
Treatment groups TVC load (CFUJ� 1)
T1 (Bacillus NL110 through feed) 3.15 � 2.47 � 107
T2 (Bacillus NL110 through water) 2.90 � 0.14 � 106
T3 (Bacillus NL110 through feed and water) 2.85 � 0.49 � 107
T4 (Vibrio NE17 through feed) 4.50 � 2.12 � 105
T5 (Vibrio NE17 through water) 6.90 � 0.14 � 105
T6 (Vibrio NE17 through feed and water) 7.00 � 0.71 � 105
T7 (Lactobacillus plantarum through feed) 5.15 � 1.48 � 105
T8 (L. plantarum through water) 4.65 � 2.19 � 105
T9 (L. plantarum through feed and water) 5.70 � 2.40 � 105
Control 4.90 � 0.85 � 105
CFU, colony-forming unit.
Probiotic e¡ect of bacteria onM. rosenbergii K MMujeeb Rahiman et al. Aquaculture Research, 2010, 41, e120^e134
r 2010 TheAuthorse128 Journal Compilationr 2010 Blackwell Publishing Ltd, Aquaculture Research, 41, e120^e134

the di¡erent treatment groups that showed a positivee¡ect, M. rosenbergii juveniles from the T2 and T6groups showed the highest weight gain, SGR andthe lowest FCR. Signi¢cant increases in the growthof P. monodon and P. vannamei with di¡erent strainsof Bacillus spp. andVibrio spp. incorporated into thefeed were reported previously (Rengpipat et al. 1998;Gullian et al. 2004). Garriques and Arevalo (1995) re-ported better shrimp growth in commercial P. vanna-mei hatcheries, treated with V. alginolyticus, whileWang (2007) observed a signi¢cantly higher meanweight in probiotic-treated groups of P. vannameithan that of the control.Venkat et al. (2004) and Nejadet al. (2006) reported higher percentage weight gain,SGRand FCRof PL ofM. rosenbergii and Indianwhiteshrimp using Lactobacillus spp. and Bacillus respec-tively.The results of our studies also recon¢rm the ef-¢cacy of probiotics in increasing the SGR of culturedshrimp.It was reported by Wang (2007) and Zhou,Wang
and Li (2009) that application of probiotics induceddigestive enzyme activity and enhanced the survivalof P. vannamei. Furthermore, bacteria, particularlymembers of the genus Bacillus, secrete a wide rangeof exo-enzymes (Moriarty 1996, 1998), which mighthelp them compete with bacterial pathogens for nu-trients and thereby inhibit the growth of pathogens.Such probiotic strains were suggested as avalid alter-native to the prophylactic application of chemicals(Decamp et al. 2008). Microorganisms are capable ofproducing complex molecules, either directly, as partof their metabolic activities, or indirectly, when theydie.The compounds that may bene¢t the host includepigments (Holmstrom, Egan, Franks, McCloy & Kjel-leberg 2002), proteins (Klein, Pack, Bonaparte & Reu-ter 1998), fatty acids (Shirasaka, Nishi & Shimizu
1995), vitamins (Sugita, Miyajima & Deguchi 1991)and digestive enzymes (Cahill 1990; Hansen & Olaf-sen 1999; Ramirez & Dixon 2003). A problem withmany cultured species is the assimilation of arti¢cialdiets during the early stages of larval development. Ithas been suggested that this is due to the low enzymelevels capable of digesting the food (Govoni, Boehlert&Watanabe1986). Probiotic strains with good exoen-zyme potential could help overcome this problem.Nejad et al. (2006) observed increases in the speci¢cactivities of digestive enzymes in probiotic treat-ments that led to enhanced digestion and increasedabsorption of food. Therefore, the addition of probio-tic bacteria capable of producing bene¢cial enzymesmayaid in the digestion of arti¢cial foods, thereby re-ducing the live food feeding period and the subse-quent associated costs. However, further studies areneeded to determine the exact role of any metaboliteproduced by the proposed probiotics Bacillus NL110andVibrio NE17.
Immunological parameters of M. rosenbergiijuveniles treated with probiotics
THC
The THC of M. rosenbergii juveniles after the experi-mental period (Table 7) showed a signi¢cantly higherTHC than that of the T4 and T6 treatment groups(Po0.05). The increase in the haemocyte count ob-served among M. rosenbergii juveniles in the presentstudy was comparable to the observations of Chengand Chen (2001) inM. rosenbergii and Chand and Sa-hoo (2006) in M. malcomsonii. Rodriguez, Espinosa,Echeverria, Cardenas, Roman and Stern (2007)reported increased THC in P. vannamei larvae and
Table 6 Percentage survival, ¢nal weight, SGR and FCR of Macrobrachium rosenbergii juveniles from control and di¡erenttreatment groups with probiotic bacterial strains incorporated through di¡erent routes
Treatment groups Survival (%) Finalweight (g) SGR FCR
T1 (Bacillus NL110 through feed) 93.3 � 0.00 0.703 � 0.004 3.60 � 0.008 2.17 � 0.01
T2 (Bacillus NL110 through water) 90.0 � 4.67 0.805 � 0.007 3.87 � 0.015 1.97 � 0.12
T3 (Bacillus NL110 through feed and water) 93.3 � 0.00 0.630 � 0.014 3.46 � 0.037 2.42 � 0.05
T4 (Vibrio NE17 through feed) 86.7 � 0.00 0.754 � 0.063 3.73 � 0.139 2.18 � 0.18
T5 (Vibrio NE17 through water) 86.7 � 0.00 0.763 � 0.025 3.78 � 0.054 2.16 � 0.07
T6 (Vibrio NE17 through feed and water) 96.7 � 4.74 0.820 � 0.006 3.90 � 0.013 1.80 � 0.07
T7 (Lactobacillus plantarum through feed) 86.7 � 0.00 0.739 � 0.011 3.70 � 0.024 2.23 � 0.03
T8 (L. plantarum through water) 90.0 � 4.67 0.780 � 0.009 3.82 � 0.020 2.03 � 0.08
T9 (L. plantarum through feed and water) 90.0 � 4.67 0.756 � 0.011 3.76 � 0.025 2.10 � 0.08
Control 86.7 � 0.00 0.629 � 0.030 3.44 � 0.079 2.62 � 0.12
SGR, speci¢c growth rate; FCR, feed conversion ratio.
Aquaculture Research, 2010, 41, e120^e134 Probiotic e¡ect of bacteria onM. rosenbergii K MMujeeb Rahiman et al.
r 2010 TheAuthorsJournal Compilationr 2010 Blackwell Publishing Ltd, Aquaculture Research, 41, e120^e134 e129
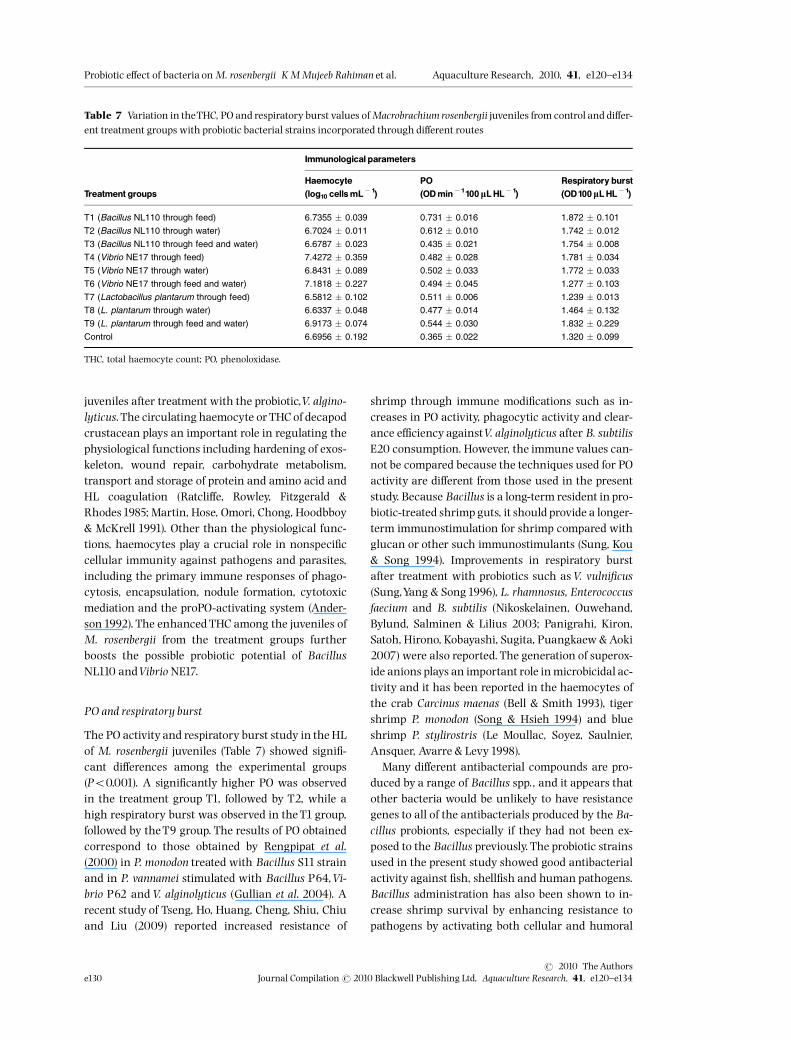
juveniles after treatment with the probiotic,V. algino-lyticus.The circulating haemocyte or THC of decapodcrustacean plays an important role in regulating thephysiological functions including hardening of exos-keleton, wound repair, carbohydrate metabolism,transport and storage of protein and amino acid andHL coagulation (Ratcli¡e, Rowley, Fitzgerald &Rhodes1985; Martin, Hose, Omori, Chong, Hoodbboy& McKrell 1991). Other than the physiological func-tions, haemocytes play a crucial role in nonspeci¢ccellular immunity against pathogens and parasites,including the primary immune responses of phago-cytosis, encapsulation, nodule formation, cytotoxicmediation and the proPO-activating system (Ander-son1992). The enhanced THC among the juveniles ofM. rosenbergii from the treatment groups furtherboosts the possible probiotic potential of BacillusNL110 andVibrio NE17.
PO and respiratory burst
The PO activity and respiratory burst study in the HLof M. rosenbergii juveniles (Table 7) showed signi¢-cant di¡erences among the experimental groups(Po0.001). A signi¢cantly higher PO was observedin the treatment group T1, followed by T2, while ahigh respiratory burst was observed in theT1 group,followed by theT9 group. The results of PO obtainedcorrespond to those obtained by Rengpipat et al.(2000) in P. monodon treated with Bacillus S11 strainand in P. vannamei stimulated with Bacillus P64,Vi-brio P62 and V. alginolyticus (Gullian et al. 2004). Arecent study of Tseng, Ho, Huang, Cheng, Shiu, Chiuand Liu (2009) reported increased resistance of
shrimp through immune modi¢cations such as in-creases in PO activity, phagocytic activity and clear-ance e⁄ciency againstV. alginolyticus after B. subtilisE20 consumption. However, the immune values can-not be compared because the techniques used for POactivity are di¡erent from those used in the presentstudy. Because Bacillus is a long-term resident in pro-biotic-treated shrimp guts, it should provide a longer-term immunostimulation for shrimp compared withglucan or other such immunostimulants (Sung, Kou& Song 1994). Improvements in respiratory burstafter treatment with probiotics such asV. vulni¢cus(Sung,Yang & Song1996), L. rhamnosus, Enterococcusfaecium and B. subtilis (Nikoskelainen, Ouwehand,Bylund, Salminen & Lilius 2003; Panigrahi, Kiron,Satoh, Hirono, Kobayashi, Sugita, Puangkaew &Aoki2007) were also reported. The generation of superox-ide anions plays an important role inmicrobicidal ac-tivity and it has been reported in the haemocytes ofthe crab Carcinus maenas (Bell & Smith 1993), tigershrimp P. monodon (Song & Hsieh 1994) and blueshrimp P. stylirostris (Le Moullac, Soyez, Saulnier,Ansquer, Avarre & Levy1998).Many di¡erent antibacterial compounds are pro-
duced by a range of Bacillus spp., and it appears thatother bacteria would be unlikely to have resistancegenes to all of the antibacterials produced by the Ba-cillus probionts, especially if they had not been ex-posed to the Bacillus previously. The probiotic strainsused in the present study showed good antibacterialactivity against ¢sh, shell¢sh and human pathogens.Bacillus administration has also been shown to in-crease shrimp survival by enhancing resistance topathogens by activating both cellular and humoral
Table 7 Variation in theTHC, PO and respiratory burst values ofMacrobrachium rosenbergii juveniles from control and di¡er-ent treatment groups with probiotic bacterial strains incorporated through di¡erent routes
Treatment groups
Immunological parameters
Haemocyte(log10 cellsmL� 1)
PO(ODmin� 1100 lLHL� 1)
Respiratory burst(OD100 lLHL� 1)
T1 (Bacillus NL110 through feed) 6.7355 � 0.039 0.731 � 0.016 1.872 � 0.101
T2 (Bacillus NL110 through water) 6.7024 � 0.011 0.612 � 0.010 1.742 � 0.012
T3 (Bacillus NL110 through feed and water) 6.6787 � 0.023 0.435 � 0.021 1.754 � 0.008
T4 (Vibrio NE17 through feed) 7.4272 � 0.359 0.482 � 0.028 1.781 � 0.034
T5 (Vibrio NE17 through water) 6.8431 � 0.089 0.502 � 0.033 1.772 � 0.033
T6 (Vibrio NE17 through feed and water) 7.1818 � 0.227 0.494 � 0.045 1.277 � 0.103
T7 (Lactobacillus plantarum through feed) 6.5812 � 0.102 0.511 � 0.006 1.239 � 0.013
T8 (L. plantarum through water) 6.6337 � 0.048 0.477 � 0.014 1.464 � 0.132
T9 (L. plantarum through feed and water) 6.9173 � 0.074 0.544 � 0.030 1.832 � 0.229
Control 6.6956 � 0.192 0.365 � 0.022 1.320 � 0.099
THC, total haemocyte count; PO, phenoloxidase.
Probiotic e¡ect of bacteria onM. rosenbergii K MMujeeb Rahiman et al. Aquaculture Research, 2010, 41, e120^e134
r 2010 TheAuthorse130 Journal Compilationr 2010 Blackwell Publishing Ltd, Aquaculture Research, 41, e120^e134

immune defences in shrimp (Rengpipat et al. 2000).Bacillus surface antigens or their metabolites werealso reported to act as immunogens for shrimp by sti-mulating the phagocytic activity of granulocytes(Itami, Kubono, Asano, Tokushige, Takeno, Nishi-mura, Kondo & Takahashi1998).The various mechanisms of probiotic bacterial ac-
tionmight prevent the emergence of resistant strains.The immune systems defend and collectively conferbacterial disease protection. Probiotics are more de-sirable and environmentally benign compared withthe use of antibiotics and chemicals. It is clear fromthe experiment that the probiotic strains BacillusNL110 introduced throughwater and feed separatelyandVibrio NE17 introduced through feed and watersigni¢cantly enhance the weight gain, SGR, FCR andimmunity of M. rosenbergii juveniles. It is concludedthat the natural bacterial £ora associated with thecultured ¢sh and shell¢sh might have bacterialstrains with good probiotic potential, which may bescreened and exploited for an environment-friendlyand organic mode of aquaculture.
Acknowledgments
The authors wish to express their gratitude to theUniversity Grants Commission (UGC), Govt. of India,for the ¢nancial assistance (F.3-37/2002).
References
Aly S.M., Abd-El-Rahman A.M., John G. & Mohamed M.F.(2008) Characterization of some bacteria isolated fromOreochromis niloticus and their potential use as probiotics.Aquaculture 277,1^6.
Aly S.M., Mohamed M.F. & JohnG. (2008) E¡ect of probioticson the survival, growth and challenge infection inTilapianilotica (Oreochromis niloticus).Aquaculture Research39,647^656.
Anderson D.P. (1992) Immunostimulant, adjuvants and vac-cine carriers in ¢sh: application to aquaculture. AnnualReview of Fish Diseases 2, 281^307.
Anderson I.G., Shamsudin M.N. & Nash G. (1989) A preli-minary study on the aerobic heterotrophic bacterial £orain giant freshwater prawn, Macrobrachium rosenbergii,hatcheries in Malaysia. Aquaculture 81, 213^223.
APHA. (1998) Standard Methods for the Examination ofWaterandWastewater. American Public Health Association,Wa-shington, DC.
Apun-Molina J.P., Miranda A.S., Gonzalez A.L., Martinez-Diaz S.F. & Rojas-Contreras M. (2009) E¡ect of potentialprobiotic bacteria on growth and survival of tilapia Oreo-
chromis niloticus L., cultured in the laboratory under highdensity and sub-optimum temperature. Aquaculture Re-search 40,887^894.
Austin B., Stuckey L.F., Robertson P.A.W., E¡endi I. & Gri⁄thD.R.W. (1995) A probiotic strain of Vibrio alginolyticus ef-fective in reducing diseases caused byAeromonas salmoni-cida,Vibrio anguillarum and Vibrio ordalii. Journal of FishDiseases18,93^96.
Balcazar J.L. & Rojas-Luna T. (2007) Inhibitory activity ofprobiotic Bacillus subtilis UTM 126 against Vibrio spe-cies confers protection against vibriosis in juvenileshrimp (Litopenaeus vannamei). Current Microbiology 55,409^412.
Balcazar J.L., Rojas-LunaT. & CunninghamD.P. (2007) E¡ectof the addition of four potential probiotic strains on thesurvival of paci¢c white shrimp (Litopenaeus vannamei)following immersion challengewithVibrio parahaemolyti-cus. Journal of Invertebrate Pathology 96,147^150.
Barrow G.I. & Feltham R.K.A. (1993) Cowan and Steel’s Man-ual for the Identi¢cation of Medical Bacteria. CambridgeUniversity press, NewYork, USA.
Bell K.L. & SmithV.J. (1993) Invitro superoxide productionbyhyaline cells of the shore crab Carcinus maenas (L.). Devel-opmental & Comparative Immunology17, 211^219.
Bhan M.K., Bahl R. & Bhatnagar S. (2005) Typhoid andparatyphoid fever. Lancet 366,749^762.
Cahill M.M. (1990) Bacterial £ora of ¢shes: a review. Micro-bial Ecology19, 21^41.
Chaithanya R., Nayak D.K. & Venugopal M.N. (1999) Anti-bacterial resistance in aquaculture. INFOFISH Interna-tional 6,30^32.
Chand R.K. & Sahoo P.K. (2006) E¡ect of nitrite on the im-mune response of freshwater prawnMacrobrachium mal-comsonii and its susceptibility to Aeromonas hydrophila.Aquaculture 258,150^156.
Chen D., Hanna P.J., Altman K., Smith A., Moon P. & Ham-mond L.S. (1992) Development of monoclonal antibodiesthat identifyVibrio species commonly isolated from infec-tion of human, ¢sh and shell¢sh. Applied EnvironmentalMicrobiology 58,3694^3700.
ChengW. & Chen J.C. (1998) Isolation and characterizationof Enterococcus like bacterium causing muscle necrosisand mortality with Macrobrachium rosenbergii in Taiwan.Diseases of Aquatic Organisms 34,93^101.
Cheng W. & Chen J.C. (2001) E¡ects of intrinsic andextrinsic factors on the haemocyte pro¢le of the prawn,Macrobrachium rosenbergii. Fish & Shell¢sh Immunology11,53^63.
ChughT.D. (2008) Emerging and re-emerging bacterial dis-eases in India. Journal of Bioscience 33,549^555.
Chythanya R., Karunasagar I. & Karunasagar I. (2002) Inhi-bition of shrimp pathogenic vibrios by a marine Pseudo-monas I-2 strain. Aquaculture 208,1^10.
Colorni A. (1985) A study on the bacteria £ora of giantprawn,Macrobrachrium rosenbergii larva fedwithArtemiasalina nauplii. Aquaculture 49,1^10.
Aquaculture Research, 2010, 41, e120^e134 Probiotic e¡ect of bacteria onM. rosenbergii K MMujeeb Rahiman et al.
r 2010 TheAuthorsJournal Compilationr 2010 Blackwell Publishing Ltd, Aquaculture Research, 41, e120^e134 e131

DalminG., Kathiresan K. & PurushothamanA. (2001) E¡ectof probiotics on bacterial population and health status ofshrimp in culture pond ecosystem. Indian Journal ofExperimental Biology 39,939^942.
Decamp O., Moriarty D.J.W. & Lavens P. (2008) Probiotics forshrimp larviculture: review of ¢eld data from Asia andLatin America. Aquaculture Research 39,334^338.
El-Gamal A.A., Alderman D.J., Rodgers C.J., Polglase J.L. &Macintosh D. (1986) A scanning electron microscopestudy of oxolinic acid treatment of burn spot lesions ofMacrobrachium rosenbergii. Aquaculture 52,157^171.
Garriques D. & Arevalo G. (1995) An evaluation of the pro-duction and use of a live bacterial isolate to manipulatethe microbial £ora in the commercial production ofPenaeus vannamei postlarvae in Ecuador. In: SwimmingThrough Troubled Water. Proceedings of Special Session onShrimp Farming, San Diego, CA, USA (ed. by C.L.
Browdy & J.S. Hopkins), pp. 53–59. World Aquaculture
Society, Baton Rouge, LA, USA.
Gatesoupe F.J. (1994) Lactic acid bacteria increase the resis-tance of turbot larvae, Scophthalmus maximus, againstpathogenicVibrio. Aquatic Living Resources 7, 277^282.
Gatesoupe F.J. (1997) Siderophore production and probiotice¡ect ofVibrio sp. associated with turbot larvae, Scophthal-mus maximus. Aquatic Living Resources10, 239^246.
Gatesoupe F.J. (1999) The use of probiotics in aquaculture.Aquaculture180,147^165.
Ghosh S., Sinha A. & Sahu C. (2007) E¡ect of probiotic onreproductive performance in female live bearing orna-mental ¢sh. Aquaculture Research 38,518^526.
Ghosh S., Sinha A. & Sahu C. (2008) Dietary probiotic sup-plementation in growth and health of live-bearing orna-mental ¢shes. Aquaculture Nutrition14, 289^299.
Gibson L.F.,Woodworth J. & GeorgeA.M. (1998) Probiotic ac-tivity of Aeromonas media on the Paci¢c oyster, Crassostreagigas, when challenged withVibrio tubiashii. Aquaculture169,111^120.
Gomez-Gil B., Roque A. & Velasco-Blanco G. (2002) Cultureof Vibrio alginolyticus C7b, a potential probiotic bacter-ium, with themicroalga Chaetoceros muelleri. Aquaculture211, 43^48.
Govoni J.J., Boehlert G.W. & WatanabeY. (1986) The physiol-ogy of digestion in ¢sh larvae. Environmental Biology ofFishes 61,187^201.
Grisez L. & Ollevier F. (1995) Vibrio anguillarum infectionsin marine ¢sh larviculture. In: Larvi’ 95 - Fish andShell¢sh Larviculture Symposium. Gent, Special PublicationNo. 24 (ed. by P. Lavens, E. Jaspers & I. Roelands),
pp. 497. European Aquaculture Society, Oostende,
Belgium.
Gullian M.,Thompson F. & Rodriguez J. (2004) Selection ofprobiotic bacteria and study of their immunostimulatorye¡ect in Penaeus vannamei. Aquaculture 233,1^14.
Hamner S., Broadway S.C., MishraV.B.,Tripathi A., MishraR.K, Pulcini E., Pyle B.H & FordT.E. (2007) Isolation of po-tentially pathogenic Escherichia coli O157: H7 from the
Ganges River. Applied Environmental Microbiology 73,2369^2372.
Hansen G.H. & Olafsen J.A. (1999) Bacterial interactions inearly life stages of marine cold water ¢sh. Microbial Ecol-ogy 38,1^26.
Holmstrom C., Egan S., Franks A., McCloy S. & Kjelleberg S.(2002) Antifouling activities expressed by marine surfaceassociated Pseudoalteromonas species. FEMSMicrobiologyEcology 41, 47^58.
Holt J.G., Krieg N.R., Sneath P.H.A., Staley J.T. & WillliamsS.T. (2000) Bergy’s Manual of Determinative Bacteriology.LipponcottWilliams &Wilkins, Philadelphia, USA.
Huys L., Dhert P., Robles R., Ollevier F., Sorgeloos P. & SwingsJ. (2001) Search for bene¢cial bacterial strains for turbot(Scophthalmus maximus L.) larviculture. Aquaculture193,25^37.
IriantoA. & Austin B. (2002) Use of probiotics to control fur-unculosis in rainbow trout, Oncorhynchus mykiss (Wal-baum). Journal of Fish Diseases 25,333^342.
Itami T., Kubono K., Asano M., Tokushige K., Takeno N.,Nishimura H., Kondo M. & TakahashiY. (1998) Enhance-ment of disease resistance of kuruma shrimp, Penaeus ja-ponicus, after oral administration of peptidoglycanderived from Bi¢dobacterium thermophilum. Aquaculture164, 277^288.
Jayaprakash N.S., Pai S.S., Anas A., Preetha R., Philip R. &Singh I.S.B. (2005) A marine bacterium, MicrococcusMCCB104, antagonistic to vibrios in prawn larval rearingsystems. Diseases of Aquatic Organisms 68,39^45.
Keysami M.A., Saad C.R., Sijam K., Daud H.M. & AlimonA.R. (2007) E¡ect of Bacillus subtilis on growth develop-ment and survival of larvae Macrobrachium rosenbergii(de Man). Aquaculture Nutrition13,131^136.
Khuntia C.P., Das B.K., Samantaray B.R., Samal S.K. & Mis-hra B.K (2008) Characterization and pathogenicity stu-dies of Vibrio parahaemolyticus isolated from diseasedfreshwater prawn, Macrobrachium rosenbergii (de Man).Aquaculture Research 39,301^310.
KleinG., PackA., Bonaparte C. &Reuter G. (1998) Taxonomyand physiology of probiotic lactic acid bacteria. Interna-tional Journal of Food Microbiology 26,103^125.
Kumar R., Mukherjee S.C., Prasad K.P. & Pal A.K. (2006)Evaluation of Bacillus subtilis as a probiotic to Indian ma-jor carp Labeo rohita (Ham.). Aquaculture Research 37,1215^1221.
Lalitha K.V. & Surendran P.K. (2004) Bacterial micro£ora as-sociated with farmed freshwater prawn Macrobrachiumrosenbergii (de Man) and the aquaculture environment.Aquaculture Research 35,629^635.
Lavilla-Pitogo C.R., Leano E.M. & Paner M.G. (1998) Mortal-ities of pond-cultured juvenile shrimp, Penaeus monodonassociated with dominance of luminescent Vibrios in therearing environment. Aquaculture164,337^349.
Le Moullac G., Soyez C., Saulnier D., Ansquer D., Avarre J.C.& Levy P. (1998) E¡ect of hypoxia stress on the immuneresponse and the resistance to vibriosis of the shrimp
Probiotic e¡ect of bacteria onM. rosenbergii K MMujeeb Rahiman et al. Aquaculture Research, 2010, 41, e120^e134
r 2010 TheAuthorse132 Journal Compilationr 2010 Blackwell Publishing Ltd, Aquaculture Research, 41, e120^e134

Penaeus stylirostris. Fish and Shell¢sh Immunology 8,621^629.
Lightner D.V. (1983) Diseases of cultured Penaeid shrimps.In: CRC Handbook of Mariculture (ed. by J.P. McVey), pp.289^320. CRC Press, Boca Raton, FL, USA.
Lombardi J.V. & LabaoV.L. (1991a) Disease and conditioningfactors of mortality in larval culture of prawn of thegenusMacrobrachium. In: Proceedings of the 3rd BrazilianSymposium on Shrimp Culture, pp. 401–408. Joao Pessoa,
Paraiba, Brazil.
Lombardi J.V. & LabaoV.L. (1991b) Diseases and other factorsleading to mortality in juveniles and adults belonging tothe genus Macrobrachium. In: Proceedings of the 3rd Bra-zilian Symposium on Shrimp Culture, pp. 409–419. JoaoPessoa, Paraiba, Brazil.
Martin G.G., Hose J.E., Omori S., Chong C., Hoodbboy T. &McKrell N. (1991) Localization and roles of coagulogenand transglutaminase in hemolymph coagulation in dec-apod crustaceans.Comparative Biochemistry&Physiology100B,517^522.
Martin G., Rubin N. & Swanson E. (2004) Vibrio parahaemo-lyticus and V. harveyi cause detachment of epitheliumfrom the midgut trunk of the penaid shrimp Sicyonia in-gentis. Diseases of Aquatic Organism 60, 21^29.
Ministry of Health & FamilyWelfare. (2006) Food safety In-dia: Escherichia coli O157:H7. Available at http://foodsafetyindia.nic.in/ecolitaq.htm (accessed January 2007).
Moriarty D.J.W. (1996) Microbial biotechnology: a key ingre-dient for sustainable aquaculture. INFOFISH Interna-tional 4, 29^33.
Moriarty D.J.W. (1998) Control of luminousVibrio species inpenaeid aquaculture ponds. Aquaculture164,351^358.
Muroga K., Higashi M. & Keetoku H. (1987) The isolation ofintestinal micro£ora of farmed red seabream (Pagrus ma-jor) and black seabream (Acanthopagrus schlegeli) at larvaland juvenile stages. Aquaculture 65,79^88.
Nejad S.Z., Rezaei M.H.,Takami G.A., Lovett D.L., Mirvaghe¢A.R. & Shakouri M. (2006) The e¡ect of Bacillus spp. bac-teria used as probiotics on digestive enzyme activity,survival and growth in the Indian white shrimp Fennero-penaeus indicus. Aquaculture 252,516^524.
New M.B. (1995) Status of freshwater prawn farming: a re-view. Aquaculture Research 26,1^54.
Nikoskelainen S., Ouwehand A.C., Bylund G., Salminen S. &Lilius E.M. (2003) Immune enhancement in rainbowtrout (Oncorhynchus mykiss) by potential probiotic bacter-ia (Lactobacillus rhamnosus). Fish and Shell¢sh Immunology15, 443^452.
Panigrahi A., KironV., Satoh S., Hirono I., Kobayashi T., Su-gita H., Puangkaew J. & Aoki T. (2007) Immune modula-tion and expression of cytokine genes in rainbow troutOncorhynchusmykiss uponprobiotic feeding. Developmen-tal and Comparative Immunology 31,372^382.
Queiroz J. & Boyd C. (1998) E¡ects of a bacterial inoculum inchannel cat¢sh ponds. Journal of World Aquaculture So-ciety 29,67^73.
Ramirez R.F. & Dixon B.A. (2003) Enzyme production by ob-ligate intestinal anaerobic bacteria isolated from oscars(Astronotus ocellatus), angel¢sh (Pterophyllum scalare) andsouthern £ounder (Paralichthys lethostigma). Aquaculture227, 417^426.
Ravi A.V., Musthafa K.S., Jegathammbal G., Kathiresan K. &Pandian S.K. (2007) Screening and evaluation of probio-tics as a biocontrol agent against pathogenic vibrios inmarine aquaculture. Letters in Applied Microbiology 45,219^223.
Ratcli¡e N.A., Rowley A.F., Fitzgerald S.W. & Rhodes C.P.(1985) Invertebrate immunity: basic concepts and recentadvances. International Review of Cytology 97,183^350.
Rengpipat S., PhianphakW., Piyatiratitivorakul S. & Menas-veta P. (1998) E¡ects of a probiotic bacterium on black ti-ger shrimp Penaeus monodon survival and growth.Aquaculture167,301^313.
Rengpipat S., Rukpratanporn S., Piyatiratitivorakul S. & Me-nasaveta P. (2000) Immunity enhancement in black tigershrimp (Penaeus monodon) bya probiont bacterium (Bacil-lus S11). Aquaculture191, 271^288.
Ringo E. & Vadstein O. (1998) Colonization ofVibrio pelagicusand Aeromonas caviae in early developing turbot(Scopthalmus maximus L.) larvae. Journal of Applied Micro-biology 84, 227^233.
Riquelme C., Araya R. & Escribano R. (2000) Selective incor-poration of bacteria byArgopecten purpuratus larvae: im-plications for the use of probiotics in culturing systemsof the Chilean scallop. Aquaculture181, 25^36.
Rodriguez J., EspinosaY., Echeverria F., Cardenas G., RomanR. & Stern S. (2007) Exposure to probiotics and b-1,3/1,6-glucans in larviculture modi¢es the immune response ofPenaeus vannamei juveniles and both the survival toWhite Spot SyndromeVirus challenge and pond culture.Aquaculture 273, 405^415.
Shirasaka N., Nishi K. & Shimizu S. (1995) Occurrence of afuran fatty acid in marine bacteria. Biochimica et Biophy-sica Acta1258, 225^227.
Singh I.S.B. (1990) Bacterial £ora of larvae and larval rear-ing system of giant freshwater prawn (M. rosenbergii). In:Proceedings of 2nd Kerala Science Congress, Trivandrum,
India (ed. by N. Balakrishnan Nair), pp. 190–194. State
Committee on Science, Technology and Environment,
Kerala.
Skjermo J. & Vadstein O. (1999) Techniques for microbialcontrol in the intensive rearing of marine larvae.Aquacul-ture177,333^343.
Soderhall K. (1981) Fungal cell wall b-1,3 glucan induce clot-ting and phenoloxidase attachment ato foreign surface ofcray¢sh haemocyte lysate.Developmental and ComparativeImmunology 5,565^573.
Song Y.L. & Hsieh Y.T. (1994) Immunostimulation of tigershrimp Penaeus monodon haemocytes for generation ofmicrobicidal substances: analysis of reactive oxygenspecies. Developmental and Comparative Immunology 18,201^209.
Aquaculture Research, 2010, 41, e120^e134 Probiotic e¡ect of bacteria onM. rosenbergii K MMujeeb Rahiman et al.
r 2010 TheAuthorsJournal Compilationr 2010 Blackwell Publishing Ltd, Aquaculture Research, 41, e120^e134 e133

Sugita H., Miyajima C. & DeguchiY. (1991) The vitamin B12-producing ability of the intestinal micro£ora of fresh-water ¢sh. Aquaculture 92, 267^276.
Sung H.H., Kou G.H. & SongY.L. (1994) Vibriosis resistanceinduced by glucan treatment in tiger shrimp Penaeusmonodon. Fish Pathology 29,11^17.
Sung H.H.,YangY.L. & SongY.L. (1996) Enhancement of mi-crobicidal activity in the tiger shrimp Penaeus monodonvia immunostimulation. Journal of Crustacean Biology16,278^284.
Sung H.H., Hwang S.F. & Tasi F.M. (2000) Responses of giantfreshwater prawn (Macrobrachium rosenbergii) to chal-lenge by two strains of Aeromonas spp. Journal of Inverte-brate Pathology 76, 278^284.
Suralikar V. & Sahu N.P. (2001) E¡ect of feeding probiotic(Lactobacillus cremoris) on growth and survival ofMacro-brachium rosenbergii postlarvae. Journal of Applied AnimalResearch 20,117^124.
Tseng D.Y., Ho P.L., Huang S.Y., Cheng S.C., Shiu Y.L., ChiuC.S. & Liu C.H. (2009) Enhancement of immunity anddisease resistance in thewhite shrimp, Litopenaeus vanna-mei, by the probiotic, Bacillus subtilis E20. Fish & Shell¢shImmunology 26,339^344.
UddinM.N. & Al-Harbi A.H. (2005) Quantitative and qualita-tive bacterial £ora of giant freshwater prawn Macrobra-chium rosenbergii cultured in earthen ponds in SaudiArabia. Journal of Aquatic Animal Health17, 244^250.
Venkat H.K., Sahu N.P. & Jain K.K. (2004) E¡ect of feedingLactobacillus based probiotics on the gut micro£ora,
growth and survival of postlarvae of Macrobrachiumrosenbergii (de Man). Aquaculture Research 35,501^507.
Verschuere L., Rombaut G., Sorgeloos P. & Verstraete W.(2000) Probiotic bacteria as biological control agents inaquaculture. Microbiology and Molecular Biology Reviews64,655^671.
Vijayan K.K., Bright Singh I.S., Jayaprakash N.S., AlavandiS.V., Somnath Pai S., Preetha R., Rajan J.J.S. & SantiagoT.C. (2006) A brackishwater isolate of Pseudomonas PS-102, a potential antagonistic bacterium against patho-genic vibrios in penaeid and non-penaeid rearing sys-tems. Aquaculture 251,192^200.
WangY.B. (2007) E¡ect of probiotics ongrowth performanceand digestive enzyme activity of the shrimp Penaeus van-namei. Aquaculture 269, 259^264.
WangY.B. & Xu Z.R. (2006) E¡ect of probiotics for commoncarp (Cyprinus carpio) based on growth performance anddigestive enzyme activities. Animal Feed Science and Tech-nology127, 283^292.
WangY.C., Lo C.F., Chang P.S. & Kou G.H. (1998) Experimen-tal infection of white spot baculovirus in some culturedand wild decapods inTaiwan. Aquaculture164, 221^231.
Woc-Colburn L. & Bobak D.A. (2009) The expanding spec-trum of disease due to Salmonella: an international per-spective. Current Infectious Disease Reports11,120^124.
Zhou X.,WangY. & LiW. (2009) E¡ect of probiotic on larvaeshrimp (Penaeus vannamei) based on water quality, survi-val rate and digestive enzyme activities. Aquaculture 287,349^353.
Probiotic e¡ect of bacteria onM. rosenbergii K MMujeeb Rahiman et al. Aquaculture Research, 2010, 41, e120^e134
r 2010 TheAuthorse134 Journal Compilationr 2010 Blackwell Publishing Ltd, Aquaculture Research, 41, e120^e134
Copyright © 2022 FDOKUMEN




















