
Identification and expression of cellobiohydrolase (CBHI) gene from an endophytic fungus, Fusicoccum...
6
Identification and expression of cellobiohydrolase (CBHI) gene from an endophytic fungus, Fusicoccum sp. (BCC4124) in Pichia pastoris Pattanop Kanokratana a , Duriya Chantasingh a , Verawat Champreda a , Sutipa Tanapongpipat a , Kusol Pootanakit b , Lily Eurwilaichitr a, * a Molecular and Enzyme Screening Laboratory, BIOTEC Central Research Unit, National Center for Genetic Engineering and Biotechnology, 113 Thailand Science Park, Paholyothin Road, Klong 1, Klong Luang, Pathumthani 12120, Thailand b Institute of Molecular Biology and Genetics, Mahidol University, Salaya Campus, Nakhon Pathom 73170, Thailand Received 2 August 2007, and in revised form 6 September 2007 Available online 19 September 2007 Abstract A gene encoding a cellobiohydrolase (CBHI) was isolated from Fusicoccum sp. (BCC4124), an endophytic fungus belongs in phylum Ascomycota, using 5 0 and 3 0 rapid amplification of cDNA end (RACE) technique. This CBHI gene contains 1395 nucleotides and encodes a 465-amino acid protein with a molecular weight of approximately 50 kDa. The deduced amino acid sequence showed signif- icant similarity to those of other fungal CBHI belonging to family 7 of glycosyl hydrolase. Interestingly, the result from the amino acid alignment revealed that this CBHI does not contain the cellulose binding domain nor the linker region. The CBHI gene was successfully expressed in Pichia pastoris KM71. The purified recombinant CBHI has ability to hydrolyze Avicel, filter paper and 4-methylumbelliferyl b-D-cellobioside (MUC) but not carboxymethylcellulose (CMC). It showed an optimal working condition at 40 °C, pH 5 with K m and V max toward MUC of 0.57 mM and 3.086 nmol/min/mg protein, respectively. The purified enzyme was stable at pH range of 3–11. The enzyme retained approximately 50% of its maximal activity after incubating at 70–90 °C for 30 min. Due to its stability through wide range of pH, and moderately stable at high temperature, this enzyme has potential in various biotechnology applications. Ó 2007 Elsevier Inc. All rights reserved. Keywords: Cellobiohydrolase; Cellulose binding domain; Endophytic fungus; Fusicoccum; Pichia pastoris Cellulolytic enzymes can be divided into three types: 1,4- b-D-endoglucanase (EC 3.1.2.4), 1,4-b-D-cellobiohydrolase (CBH; EC 3.1.2.91), and b-glucosidase (EC 3.2.1.21). CBHs hydrolyze crystalline cellulose by initiating their action from the ends of the cellulose chains and producing primarily cellobiose. Endoglucanases catalyze hydrolysis internally in the cellulose chain and attack the amorphous regions in cellulose, providing new chain ends for CBHs. b- glucosidases hydrolyze cellobiose to glucose. During degra- dation of cellulose, these different types of enzymes act syn- ergistically and efficiently hydrolyze cellulose polymer into glucose. Application of cellulases in many industries and for research proposes has been reviewed [1]. CBHs are key components in the multi-enzyme cellulase complexes [2]. Most belong to family 6 or 7 of glycosyl hydrolase [3–8]. Potent cellulolytic fungi generally produce two different CBHs, CBHI and CBHII. These two types of enzymes which are classified based on sequence identity, have extended tunnel-shaped active sites and can achieve complete, although slow, solubilization of cellulose crystals even without help of endoglucanases [2]. Fungal CBHs usually composed of a catalytic domain, a cellulose binding domain (CBD), and a flexible peptide linker that links the two domains together [9,10]. CBHs are especially proven to be useful in specific fiber modifications and various surface treatments of paper and textiles since, unlike endoglucan- ases, they do not significantly affect fiber strength [2]. In the present study, we reported the isolation of a gene encoding CBHI from Fusicoccum sp. (BCC4124), an 1046-5928/$ - see front matter Ó 2007 Elsevier Inc. All rights reserved. doi:10.1016/j.pep.2007.09.008 * Corresponding author. Fax: +66 2 564 6707. E-mail address: [email protected] (L. Eurwilaichitr). www.elsevier.com/locate/yprep Available online at www.sciencedirect.com Protein Expression and Purification 58 (2008) 148–153
Transcript of Identification and expression of cellobiohydrolase (CBHI) gene from an endophytic fungus, Fusicoccum...

Available online at www.sciencedirect.com
www.elsevier.com/locate/yprep
Protein Expression and Purification 58 (2008) 148–153
Identification and expression of cellobiohydrolase (CBHI) gene froman endophytic fungus, Fusicoccum sp. (BCC4124) in Pichia pastoris
Pattanop Kanokratana a, Duriya Chantasingh a, Verawat Champreda a,Sutipa Tanapongpipat a, Kusol Pootanakit b, Lily Eurwilaichitr a,*
a Molecular and Enzyme Screening Laboratory, BIOTEC Central Research Unit, National Center for Genetic Engineering and Biotechnology,
113 Thailand Science Park, Paholyothin Road, Klong 1, Klong Luang, Pathumthani 12120, Thailandb Institute of Molecular Biology and Genetics, Mahidol University, Salaya Campus, Nakhon Pathom 73170, Thailand
Received 2 August 2007, and in revised form 6 September 2007Available online 19 September 2007
Abstract
A gene encoding a cellobiohydrolase (CBHI) was isolated from Fusicoccum sp. (BCC4124), an endophytic fungus belongs in phylumAscomycota, using 5 0 and 3 0 rapid amplification of cDNA end (RACE) technique. This CBHI gene contains 1395 nucleotides andencodes a 465-amino acid protein with a molecular weight of approximately 50 kDa. The deduced amino acid sequence showed signif-icant similarity to those of other fungal CBHI belonging to family 7 of glycosyl hydrolase. Interestingly, the result from the amino acidalignment revealed that this CBHI does not contain the cellulose binding domain nor the linker region. The CBHI gene was successfullyexpressed in Pichia pastoris KM71. The purified recombinant CBHI has ability to hydrolyze Avicel, filter paper and 4-methylumbelliferylb-D-cellobioside (MUC) but not carboxymethylcellulose (CMC). It showed an optimal working condition at 40 �C, pH 5 with Km andVmax toward MUC of 0.57 mM and 3.086 nmol/min/mg protein, respectively. The purified enzyme was stable at pH range of 3–11. Theenzyme retained approximately 50% of its maximal activity after incubating at 70–90 �C for 30 min. Due to its stability through widerange of pH, and moderately stable at high temperature, this enzyme has potential in various biotechnology applications.� 2007 Elsevier Inc. All rights reserved.
Keywords: Cellobiohydrolase; Cellulose binding domain; Endophytic fungus; Fusicoccum; Pichia pastoris
Cellulolytic enzymes can be divided into three types: 1,4-b-D-endoglucanase (EC 3.1.2.4), 1,4-b-D-cellobiohydrolase(CBH; EC 3.1.2.91), and b-glucosidase (EC 3.2.1.21).CBHs hydrolyze crystalline cellulose by initiating theiraction from the ends of the cellulose chains and producingprimarily cellobiose. Endoglucanases catalyze hydrolysisinternally in the cellulose chain and attack the amorphousregions in cellulose, providing new chain ends for CBHs. b-glucosidases hydrolyze cellobiose to glucose. During degra-dation of cellulose, these different types of enzymes act syn-ergistically and efficiently hydrolyze cellulose polymer intoglucose. Application of cellulases in many industries andfor research proposes has been reviewed [1].
1046-5928/$ - see front matter � 2007 Elsevier Inc. All rights reserved.
doi:10.1016/j.pep.2007.09.008
* Corresponding author. Fax: +66 2 564 6707.E-mail address: [email protected] (L. Eurwilaichitr).
CBHs are key components in the multi-enzyme cellulasecomplexes [2]. Most belong to family 6 or 7 of glycosylhydrolase [3–8]. Potent cellulolytic fungi generally producetwo different CBHs, CBHI and CBHII. These two types ofenzymes which are classified based on sequence identity,have extended tunnel-shaped active sites and can achievecomplete, although slow, solubilization of cellulose crystalseven without help of endoglucanases [2]. Fungal CBHsusually composed of a catalytic domain, a cellulose bindingdomain (CBD), and a flexible peptide linker that links thetwo domains together [9,10]. CBHs are especially proven tobe useful in specific fiber modifications and various surfacetreatments of paper and textiles since, unlike endoglucan-ases, they do not significantly affect fiber strength [2].
In the present study, we reported the isolation of agene encoding CBHI from Fusicoccum sp. (BCC4124), an

1 Abbreviations used: RACE, rapid amplification of cDNA end; MUC,4-methylumbelliferyl b-D-cellobioside; CMC, carboxymethylcellulose;CBD, cellulose binding domain; BCC, BIOTEC Culture Collection;BMGY, buffered minimal glycerol complex medium; BMMY, bufferedminimal methanol medium; DNS, dinitrosalicylic acid.
P. Kanokratana et al. / Protein Expression and Purification 58 (2008) 148–153 149
endophytic fungus belongs in the phylum Ascomycota thathas been collected at BIOTEC Culture Collection, Thai-land. A preliminary study demonstrated that BCC4124produced cellobiohydrolases that were active over a broadpH range (pH 3, 7 and 10), which suggested that theenzymes could be used in various conditions in many typesof industrial processes. To increase the production of theenzyme, a cellobiohydrolase was isolated and heterolo-gously expressed in the yeast Pichia pastoris. Biochemicalcharacteristics of the recombinant CBHI showed that therecombinant enzyme was also highly stable at the extremealkaline pH (9–11). This study provides another step intounderstanding the relationship between enzyme structureand its stability.
Materials and methods
Strains, culture conditions and plasmid
Fusicoccum sp. (BCC4124) was obtained from BIOTECCulture Collection (BCC), Thailand. It was cultured aero-bically in 5% wheat bran liquid media at 30 �C for 5 days.Fungal mycelia were harvested by filtration through a0.5 mm diameter mesh. Escherichia coli DH5a was usedas a host for plasmid propagation. P. pastoris KM71(Invitrogen, USA) was used as a host for expression ofthe recombinant CBHI. It was cultured in YEPD [2% (w/v) peptone, 2% (w/v) glucose and 1% (w/v) yeast extract].pPICZaA (Invitrogen) was used as a vector to carry CBHI
into P. pastoris. The yeast transformants were grown underselective condition in YEPD containing 0.1 mg/ml zeocinas a selectable marker.
Total RNA isolation and RT-PCR
The BCC4124 mycelia were harvested and then groundinto fine powder in liquid nitrogen with a mortar and pes-tle. The mycelia were then lysed and total RNAs were iso-lated using TRI-Reagent (Molecular Research Center,USA), according to the manufacturer’s instruction.
First-strand cDNAs were generated from the totalRNAs using RevertAid H Minus First-Strand cDNA Syn-thesis kit (Fermentas, Lithuania). The cDNAs obtainedwere used as template in semi-nested PCR reactions usingdegenerate primers which were designed from internal con-served region of fungal CBHI genes. The first round ofPCR reactions containing 0.8 lM of each of degenerateprimers, CelF1 and CelR1 (Table 1) were performed. ThePCR condition was as follows: one cycle of 3 min at94 �C, 35 cycles of 30 s at 94 �C, 1 min at 45 �C and1 min at 72 �C, and finally one cycle of 10 min at 72 �C.The second round of PCR was done with 0.4 lM of eachof degenerate primers, Fbasi and CelR1 (Table 1). ThePCR condition was similar to the first round of amplifica-tion except an annealing temperature of 50 �C was used.The PCR reaction product, consisting of partial sequenceof the putative CBHI gene, was cloned into pTZ57R/T
(Fermentas, Lithuania) and sequenced (Macrogen, Korea).This sequence was then used to design another set of inter-nal primers against the putative CBHI gene for rapidamplification of cDNA ends [11].
Rapid amplification of cDNA ends (RACE)1
For 3 0RACE, first-strand cDNAs were synthesized fromtotal RNAs of BCC4124 using poly-dT adaptor (PM1 pri-mer) (Table 1). PCR was performed with 3RaceCBH andPM2 primers (Table 1), using an annealing temperatureof 55 �C to obtain the 3 0end of the CBHI gene.
To obtain the 5 0end of CBHI gene, first-strand cDNAswere generated with an internal primer, 5RaceCBH2(Table 1), derived from the obtained partial sequence ofthe gene. Poly(A) tails were then added to the 3 0end ofthe obtained first-strand cDNA by terminal deoxynucleoti-dyl transferase (Fermentas, Lithuania). Semi-nested PCRwas performed using 5RaceCBH2, PM1 and PM2 primersin the first round of PCR, and 5RaceCBH3 (Table 1) andPM2 in the second round of PCR. The PCR condition inthe first round was as follows: one cycle of 3 min at94 �C, 3 min at 45 �C and 5 min at 72 �C, 32 cycles of30 s at 94 �C, 45 s at 50 �C and 1 min at 72 �C, and finallyone cycle of 10 min at 72 �C. In the second round of PCR,the condition was the same as described for the 3 0RACE.
Cloning and expression of BCC4124 CBHI in P. pastoris
The mature BCC4124 CBHI gene was obtained by RT-PCR with CBH412F-mat-Xho and CBH4124R-full-Xbaprimers (Table 1). The recognition sites for XhoI or XbaIwithin the primers were underlined. The nucleotidesequence of the full-length BCC4124 CBHI cDNA wassubmitted to GenBank (Accession No. EU038070). Theamplified fragment was treated with XhoI and XbaI andligated into the same cleavage sites of an expression vectorpPICZaA, placing the CBHI gene under the influence of amethanol-inducible promoter (AOXI) and directly down-stream of an a-factor secretion signal from yeast. The liga-tion product was transformed into E. coli DH5a. Insertionof the PCR fragment into the correct translational readingframe was verified by sequencing prior to the introductionof the plasmid into P. pastoris. Yeast transformation wasthen performed by electroporation as described in theInvitrogen’s instruction manual. The integration of thefusion gene into the yeast genome was confirmed by geno-mic PCR using 5 0AOX and 3 0AOX primers (Invitrogen).
To induce the production of the recombinant CBHIwith methanol, P. pastoris transformants were cultured inbuffered minimal glycerol complex medium (BMGY con-

Table 1Primers used in this study
Name Nucleotide sequences
CelF1 50 ACNAAYTGYTAYACN GGN 3 0
CelR1 50 RTTNGCYTGNCCRTTDATRAA 3 0
Fbasi 50 TGYGCNYTNGA YGGNGCNGAYTAY 3 0
PM1 50 CCGGAATTCAAGCTTCTAGAGGATCCTTTTTTTTTTTTTTTT 3 0
PM2 50 CCGGAATTCAAGCTTCTAGAGGATCC 30
3RaceCBH 50 CCTCACGATGAACTTCGTGACC 3 0
5RaceCBH2 50 CGAGTCCATCTCGGAGAAGTAC 3 0
5RaceCBH3 50 TAGACACGGCCACCGATGTTGG 3 0
CBH412F-mat-Xho 50 TATCTCTCGAGAAAAGACAGCAGGTCGGCACGCAAAC 3 0
CBH4124R-full-Xba 50 TTTGTTCTAGACTAAGCGGTCTTGTTGAAGGTGGAG 30
150 P. Kanokratana et al. / Protein Expression and Purification 58 (2008) 148–153
taining 100 mM phosphate buffer (pH 6.0), 0.67% (w/v)yeast nitrogen base, 1% (v/v) glycerol) at 30 �C with shak-ing until the culture reached an OD600 of 5–6. The cell pel-lets were then harvested by centrifugation and resuspendedin buffered minimal methanol medium (BMMY containing0.67% (w/v) yeast nitrogen base, 1% (w/v) yeast extract, 2%(w/v) peptone, 100 mM potassium phosphate, pH 6.0,0.00004% (w/v) biotin) using 1/5th volume of the originalculture in BMGY. Absolute methanol was added every24 h to a final concentration of 0.5–3% (v/v) to maintaininduction. Culture supernatant was collected bycentrifugation.
Purification and characterization of BCC4124 CBHI from
P. pastoris
A large-scale production of the recombinant protein wasperformed in 200 ml of BMMY containing 3% methanol.The cell-free medium containing secreted CBHI was col-lected by centrifugation and concentrated prior to purifyby gel-filtration (Superdex 200 in XK 16 column) usingAKTA Explorer (Amersham Biosciences, Sweden). Elutionwas performed with 50 mM MOPS (3-morpholinopro-panesulfonic acid) buffer (pH 6.5) containing 50 mM NaCl,at flow rate of 0.5 ml min�1. All fractions were tested forCBH activity and the fractions containing the recombinantprotein were determined by SDS–PAGE and then concen-trated with Amicon Ultra-15 (Millipore, USA).
Specific activities of the purified recombinant CBHIwere determined at 40 �C, pH 5.0 for 1 h for hydrolysisof Avicel, filter paper [12], carboxymethylcellulose (CMC)and 4-methylumbelliferyl b-D-cellobioside (MUC) (Sigma,USA). To determine enzymatic activity against Avicel, fil-ter paper and CMC, CBHI was diluted to appropriate con-centrations in 0.1 M sodium acetate buffer, pH 5.0 with 1%of Avicel, 2% of CMC or 1 piece (1 · 1.5 cm) of filter paper(Whatman No. 1). Enzymatic activity was measured bydinitrosalicylic acid (DNS) method [13]. When MUC wasused as a substrate, fluorescence of 4-methylumbelliferylgroup released from MUC was determined at excitationand emission wavelengths of 355 and 460 nm, respectively.Assays were performed in 100 ll reaction volume in 0.1 Msodium acetate buffer (pH 5.0) containing 1 mM of MUC.
The reaction was terminated by the addition of 25 ll of1 M Na2CO3. One unit of enzyme activity was defined asthe amount of the enzyme that produced the equivalenceof 1 nmol of product (reducing sugar or 4-methylumbellife-ryl) at the optimal conditions in 1 min.
The pH and temperature profiles of the enzyme weredetermined by using MUC as a substrate at different pHsand temperatures as indicated in the figure legends. Thekinetic parameters (Km and Vmax) were determined byKaleidaGraph version 3.51 data analysis software (SynergySoftware, USA)
Results
Isolation of a gene encoding BCC4124 CBHI
A partial sequence of the CBHI gene was isolated byRT-PCR with three degenerate primers (CelF1, CelR1and Fbasi; Table 1), which were designed from the con-served amino acid sequence of CBHI from various isolatesof Ascomycetes and Basidiomycetes. A single DNA bandof approximately 360 bp was obtained after two roundsof PCR (data not shown). Sequence analysis showed thatit is 78% amino acid identity to cellulase CEL7A of Lentin-
ula edodes (Accession No. AAK95563) [7]. Sequence infor-mation of the amplified fragment was further used todesign another set of gene-specific primers for 3 0RACEand 5 0RACE. The full-length sequence of a 1395-bp geneencoding for BCC4124 CBHI was then obtained. Itsdeduced amino acid sequence of 464 amino acids showedhigh homology to those of other fungal CBHI enzymesbelonging to glycosyl hydrolases family 7, 68% identity tocellobiohydrolase I of Schizophyllum commune (AccessionNo. AAX55505), 66% identity to cellobiohydrolase I–IIof Volvariella volvacea (AAT64007) and 63% identity tocellulase of Irpex lacteus (BAA76365). However, the resultfrom the amino acid alignment revealed that BCC4124CBHI does not contain cellulose binding domain (CBD)and a linker region typical of other cellulases. A signal pep-tide was predicted to contain 19 amino acids at the N-ter-minal as determined by SignalP program [14]. Therefore,the mature CBHI consisted of 445 amino acids residueswith a predicted molecular mass of 48 kDa.
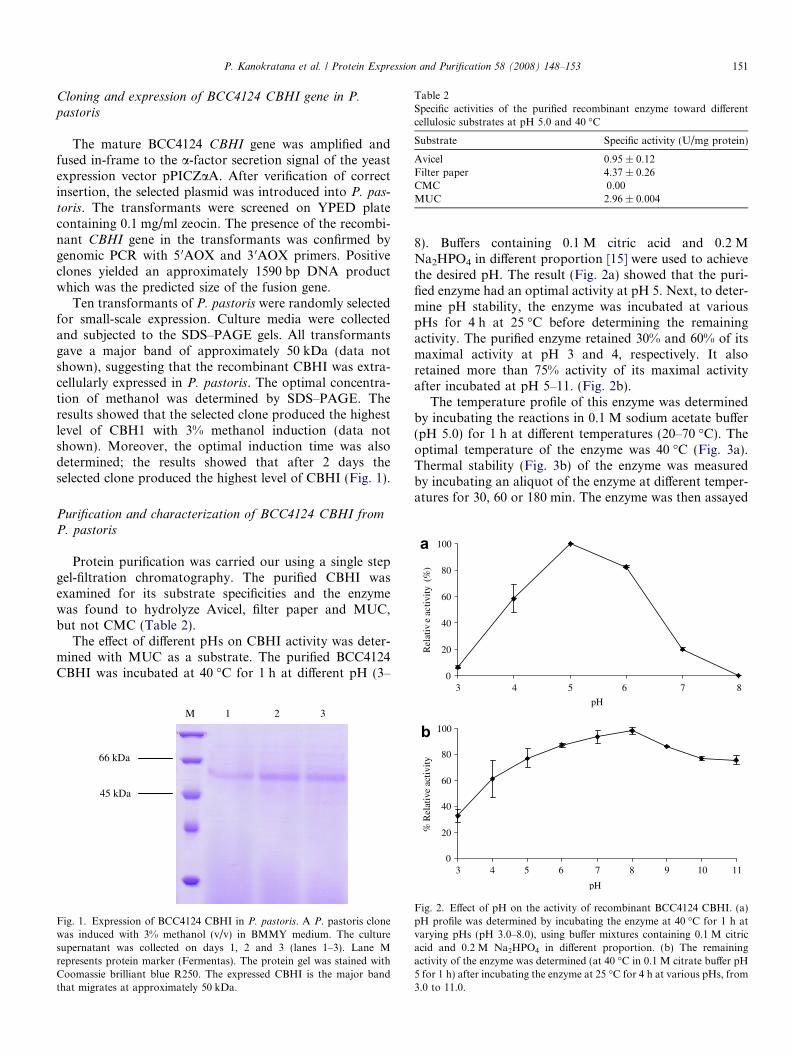
Table 2Specific activities of the purified recombinant enzyme toward differentcellulosic substrates at pH 5.0 and 40 �C
Substrate Specific activity (U/mg protein)
Avicel 0.95 ± 0.12Filter paper 4.37 ± 0.26CMC 0.00MUC 2.96 ± 0.004
P. Kanokratana et al. / Protein Expression and Purification 58 (2008) 148–153 151
Cloning and expression of BCC4124 CBHI gene in P.
pastoris
The mature BCC4124 CBHI gene was amplified andfused in-frame to the a-factor secretion signal of the yeastexpression vector pPICZaA. After verification of correctinsertion, the selected plasmid was introduced into P. pas-
toris. The transformants were screened on YPED platecontaining 0.1 mg/ml zeocin. The presence of the recombi-nant CBHI gene in the transformants was confirmed bygenomic PCR with 5 0AOX and 3 0AOX primers. Positiveclones yielded an approximately 1590 bp DNA productwhich was the predicted size of the fusion gene.
Ten transformants of P. pastoris were randomly selectedfor small-scale expression. Culture media were collectedand subjected to the SDS–PAGE gels. All transformantsgave a major band of approximately 50 kDa (data notshown), suggesting that the recombinant CBHI was extra-cellularly expressed in P. pastoris. The optimal concentra-tion of methanol was determined by SDS–PAGE. Theresults showed that the selected clone produced the highestlevel of CBH1 with 3% methanol induction (data notshown). Moreover, the optimal induction time was alsodetermined; the results showed that after 2 days theselected clone produced the highest level of CBHI (Fig. 1).
0
20
40
60
80
100
Rel
ativ
e ac
tivity
(%
)
Purification and characterization of BCC4124 CBHI fromP. pastoris
Protein purification was carried our using a single stepgel-filtration chromatography. The purified CBHI wasexamined for its substrate specificities and the enzymewas found to hydrolyze Avicel, filter paper and MUC,but not CMC (Table 2).
The effect of different pHs on CBHI activity was deter-mined with MUC as a substrate. The purified BCC4124CBHI was incubated at 40 �C for 1 h at different pH (3–
M 1 2 3
66 kDa
45 kDa
Fig. 1. Expression of BCC4124 CBHI in P. pastoris. A P. pastoris clonewas induced with 3% methanol (v/v) in BMMY medium. The culturesupernatant was collected on days 1, 2 and 3 (lanes 1–3). Lane Mrepresents protein marker (Fermentas). The protein gel was stained withCoomassie brilliant blue R250. The expressed CBHI is the major bandthat migrates at approximately 50 kDa.
8). Buffers containing 0.1 M citric acid and 0.2 MNa2HPO4 in different proportion [15] were used to achievethe desired pH. The result (Fig. 2a) showed that the puri-fied enzyme had an optimal activity at pH 5. Next, to deter-mine pH stability, the enzyme was incubated at variouspHs for 4 h at 25 �C before determining the remainingactivity. The purified enzyme retained 30% and 60% of itsmaximal activity at pH 3 and 4, respectively. It alsoretained more than 75% activity of its maximal activityafter incubated at pH 5–11. (Fig. 2b).
The temperature profile of this enzyme was determinedby incubating the reactions in 0.1 M sodium acetate buffer(pH 5.0) for 1 h at different temperatures (20–70 �C). Theoptimal temperature of the enzyme was 40 �C (Fig. 3a).Thermal stability (Fig. 3b) of the enzyme was measuredby incubating an aliquot of the enzyme at different temper-atures for 30, 60 or 180 min. The enzyme was then assayed
3 4 5 6 7 8
pH
0
20
40
60
80
100
3 4 5 6 7 8 9 10 11
pH
% R
elat
ive
activ
ity
Fig. 2. Effect of pH on the activity of recombinant BCC4124 CBHI. (a)pH profile was determined by incubating the enzyme at 40 �C for 1 h atvarying pHs (pH 3.0–8.0), using buffer mixtures containing 0.1 M citricacid and 0.2 M Na2HPO4 in different proportion. (b) The remainingactivity of the enzyme was determined (at 40 �C in 0.1 M citrate buffer pH5 for 1 h) after incubating the enzyme at 25 �C for 4 h at various pHs, from3.0 to 11.0.

0
20
40
60
80
100
30 40 50 60 70Temperature (ºC)
% R
elat
ive
activ
ity
0
20
40
60
80
100
0 60 120 180
Incubation time (min)
% R
elat
ive
activ
ity
30
40
50
60
70
80
90
Fig. 3. Effect of temperature on the activity of recombinant BCC4124CBHI. (a) Temperature profile was determined by incubating the enzymein 0.1 M citrate buffer pH 5 for 1 h at different temperatures. (b) Thermalstability was carried out by incubating the enzyme in 0.1 M citrate bufferpH 5 at different temperatures (as indicated in the graph legends) for 30,60 and 180 min before the remaining activity was assayed (at 40 �C in0.1 M citrate buffer pH 5 for 1 h).
152 P. Kanokratana et al. / Protein Expression and Purification 58 (2008) 148–153
for the remaining activity at its optimal pH and tempera-ture. The enzyme was found to retain more than 90% ofits maximal activity for at least 3 h at 30 and 40 �C. Theremaining activity of the enzyme incubating at 50 �C and60 �C was decreased to 10% and 15%, respectively, after30 min of incubation. Interestingly, it was found that theremaining activities of the enzyme incubating at 70, 80and 90 �C for 30 min were 57%, 58% and 43%, respectively,which were 4–5 times higher than that incubating at 50 �C(Fig. 3b).
The kinetic parameter of recombinant BCC4124 CBHIon MUC was determined using different concentration ofMUC ranging from 0.025 to 0.5 mM. Km and Vmax ofthe enzyme toward MUC were calculated to be 0.57 mMand 3.086 nmol/min/mg protein, respectively.
Discussion
A gene encoding cellobiohydrolase was isolated fromFusicoccum sp. (BCC4124) and successfully expressed inyeast P. pastoris. SDS–PAGE analysis showed that CBHIconstitutes a major secreted protein to the media by P. pas-
toris, with a molecular weight of 50 kDa, which agrees withthe predicted size from the deduced amino acid sequence.Sequence analysis using Blastx indicated that the enzymecontained putative conserved domains of glycosyl hydro-
lase family 7. Amino acid alignment revealed that theenzyme does not contain a CBD and a linker region.Although cellulases with no CBD are unusual, they havebeen reported previously to work well in biostoning (anenzymatic process of finishing fiber or fabric to give astone-washed appearance) with low backstaining (redepo-sition of released dye in the frabric) in textile manufacture[6]. BCC4124 CBHI contains three conserve carboxylateresidues (Glu233, Asp235 and Glu238) that are implicatedas the catalytic residues, based on the studies of Tricho-
derma reesei cellobiohydrolase I [16].The substrate specificities assay indicated that BCC4124
CBHI contains cellobiohydrolase activity since it couldhydrolyze crystalline cellulose, but not CMC. The specificactivity toward Avicel was not high compared to other fun-gal CBHI [3,17], probably due to the lack of CBD. Thisresult correlates well with a previous report of two cellobio-hydrolases secreted by Humicola grisea [3]. One cellobiohy-drolase, called EXO1, lacks a CBD and has lower substratespecificity toward Avicel than the second CBHI which con-tains a CBD [3].
The purified enzyme showed a broad pH stability, espe-cially at alkaline pH, as it retained more than 75% activityof its maximal activity after incubated at pH 5–11 for 4 h at25 �C (Fig. 3a). Comparing to CBHs from Thermoascus
aurantiacus [5] and Trametes versicolor [18], which arefound to be stable up to only pH 9, our recombinant CBHIshowed greater stability through wide range of pH, includ-ing high alkali condition. This property makes it useful forbiostoning in the textile and detergents industries [6].
The recombinant enzyme also showed interesting prop-erty of thermal stability since its remaining activities afterincubating at 70, 80 and 90 �C were significantly higherthan those incubating at 50 or 60 �C (Fig. 3b). The reasonfor the decrease in thermostability of recombinant CBHIincubated at 50–60 �C is most likely due to the irreversibil-ity of the intermediate conformation or molten globule-likestate of protein at 50–60 �C. The enzyme might be in a fullyunfolded state at temperature of 70, 80 and 90 �C and afterthe incubated enzymes were subject to an assay condition(40 �C), the fully unfolded protein (treated at 70, 80 and90 �C) could be readily refolded to its active form. Whileat 50 and 60 �C, the enzyme which is likely to be in anintermediate conformation or molten globule-like statewhich commonly exposes more hydrophobic surfaces toexternal environment and thus are suscepted to aggrega-tion [19], has very low ability to recover to its active formduring an assay at 40 �C. Therefore, very low amount ofthe enzyme activity could be recovered.
In summary, a gene encoding CBHI was successfullyisolated from Fusicoccum sp. (BCC4124) using RT-PCR.This gene contains the conserved catalytic amino acid res-idues that have been reported in other fungal cellulases.However, it does not contain a CBD. The enzyme was suc-cessfully expressed and secreted in yeast P. pastoris and itsecreted in an active form. Enzymatic properties ofBCC4124 CBHI were also determined. Due to its high tem-

P. Kanokratana et al. / Protein Expression and Purification 58 (2008) 148–153 153
perature (up to 70 �C) and high alkaline stabilities (pH 9–11), our cellobiohydrolase might be useful for textiles, pulpand paper industries.
Acknowledgments
The authors thank Dr. Piyanun Harnpicharnchai forproofreading this article. This study was supported bythe National Center for Genetic Engineering and Biotech-nology (BIOTEC), National Science and TechnologyDevelopment Agency (NSTDA), Thailand (Grant No.B21FJ0017).
References
[1] M.K. Bhat, Cellulases and related enzymes in biotechnology,Biotechnol. Adv. 18 (2000) 355–383.
[2] T.T. Teeri, Crystalline cellulose degradation: new insight into thefunction of cellobiohydrolases, Trends Biotechnol. 15 (1997) 160–167.
[3] S. Takashima, H. Iikura, A. Nakamura, M. Hidaka, H. Masaki, T.Uozumi, Isolation of the gene and characterization of the enzymaticproperties of a major exoglucanase of Humicola grisea without acellulose-binding domain, J. Biochem. 124 (1998) 717–725.
[4] B. Henrissat, A classification of glycosyl hydrolases based on aminoacid sequence similarities, Biochem. J. 280 (1991) 309–316.
[5] J. Hong, H. Tamaki, K. Yamamoto, H. Kumagai, Cloning of a geneencoding thermostable cellobiohydrolase from Thermoascus auran-
tiacus and its expression in yeast, Appl. Microbiol. Biotechnol. 63(2003) 42–50.
[6] H. Haakana, A. Miettinen-Oinonen, V. Joutsjoki, A. Mantyla, P.Suominen, J. Vehmaanpera, Cloning of cellulase genes from Mela-
nocarpus albomyces and their efficient expression in Tricoderma reesei,Enzyme Microb. Technol. 34 (2004) 159–167.
[7] C.C. Lee, D.W. Wong, G.H. Robertson, Cloning and characteriza-tion of two cellulase genes from Lentinula edodes, FEMS Microbiol.Lett. 205 (2001) 355–360.
[8] J. Jia, P.S. Dyer, J.A. Buswell, J.F. Peberdy, Cloning of the cbhIand cbhII genes involved in cellulose utilisation by the straw
mushroom Volvariella volvacea, Mol. Gen. Genet. 261 (1999)985–993.
[9] N. Gilkes, B. Henrissat, D. Kilburn, R. Miller, R. Warren, Domainsin microbial b-1,4-glycanases sequence conservation, function, andenzyme families, Microbiol. Rev. 55 (1991) 303–315.
[10] H. Van Tilbeurgh, P. Tomme, M. Claeyssens, R. Bhikhabhai, G.Pettersson, Limited proteolysis of the cellobiohydrolase I fromTrichoderma reesei. Separation of the functional domains, FEBSLett. 204 (1986) 223–227.
[11] M. Frohman, M. Dush, G. Martin, Rapid production of full-lengthcDNAs from rare transcripts: amplification using a single gene-specific oligonucleotide primer, Proc. Natl. Acad. Sci. USA 85 (1988)8998–9002.
[12] T.K. Ghose, Measurement of cellulase activities, Pure Appl. Chem. 59(1987) 257–268.
[13] G.L. Miller, Using dinitrosalicylic acid for determination of reducingsugar, Anal. Chem. 31 (1959) 426–428.
[14] J.D. Bendtsen, H. Nielsen, G. von Heijne, S. Brunak, Improvedprediction of signal peptides: SignalP 3.0, J. Mol. Biol. 340 (2004)783–795.
[15] R. Dawson, D. Elliott, W. Elliott, K. Jones, Data for BiochemicalResearch, third ed., Clarendon Press, Oxford, 1986.
[16] J. Stahlberg, C. Divne, A. Koivula, K. Piens, M. Claeyssens, T.T.Teeri, T.A. Jones, Activity studies and crystal structures of catalyt-ically deficient mutants of cellobiohydrolase I from Trichoderma
reesei, J. Mol. Biol. 264 (1996) 337–349.[17] A.V. Gusakov, A.P. Sinitsyn, T.N. Salanovich, F.E. Bukhtojarov,
A.V. Markov, B.B. Ustinov, C. van Zeijl, P. Punt, R. Burlingame,Purification, cloning and characterisation of two forms of thermo-stable and highly active cellobiohydrolase I (Cel7A) produced by theindustrial strain of Chrysosporium lucknowense, Enzyme Microb.Technol. 36 (2005) 57–69.
[18] K. Lahjouji, R. Storms, Z. Xiao, K.B. Joung, Y. Zheng, J.Powlowski, A. Tsang, L. Varin, Biochemical and molecular charac-terization of a cellobiohydrolase from Trametes versicolor, Appl.Microb. Biotechnol. 75 (2007) 337–346.
[19] S.M. Asghari, K. Khajeh, F. Moradian, B. Ranjbar, H. Naderi-Manesh, Acid-induced conformational changes in Bacillus amyloliq-
uefaciens alpha-amylase: appearance of a molten globule like state,Enzyme Microb. Technol. 35 (2004) 51–57.




















