
Diversity of endophytic bacterial communities in poplar grown under field conditions
12
RESEARCH ARTICLE Diversity of endophytic bacterial communities in poplar grown under ¢eld conditions Kristina Ulrich 1 , Andreas Ulrich 2 & Dietrich Ewald 1 1 Bundesforschungsanstalt f ¨ ur Forst- und Holzwirtschaft, Institut f ¨ ur Forstgenetik- und Forstpflanzenz ¨ uchtung, Waldsieversdorf, Germany; and 2 Leibniz-Zentrum f ¨ ur Agrarlandschaftsforschung (ZALF), Institut f ¨ ur Landschaftsstoffdynamik, M ¨ uncheberg, Germany Correspondence: Kristina Ulrich, Bundesforschungsanstalt f ¨ ur Forst- und Holzwirtschaft, Institut f ¨ ur Forstgenetik- und Forstpflanzenz ¨ uchtung, Eberswalder Chaussee 3a, D-15377 Waldsieversdorf, Germany. Tel.: 149 33433 157 175; fax: 149 33433 157 199; e-mail: [email protected] Received 31 January 2007; revised 12 October 2007; accepted 16 October 2007. First published online January 2008. DOI:10.1111/j.1574-6941.2007.00419.x Editor: Kornelia Smalla Keywords endophytic bacteria; bacterial community composition; poplar; 16S rRNA gene. Abstract Bacterial endophytes may be important for plant health and other ecologically relevant functions of poplar trees. The composition of endophytic bacteria colonizing the aerial parts of poplar was studied using a multiphasic approach. The terminal restriction fragment length polymorphism analysis of 16S rRNA genes demonstrated the impact of different hybrid poplar clones on the endophytic community structure. Detailed analysis of endophytic bacteria using cultivation methods in combination with cloning of 16S rRNA genes amplified from plant tissue revealed a high phylogenetic diversity of endophytic bacteria with a total of 53 taxa at the genus level that included Proteobacteria, Actinobacteria, Firmicutes and Bacteroidetes. The community structure displayed clear differences in terms of the presence and relative proportions of bacterial taxa between the four poplar clones studied. The results showed that the genetic background of the hybrid poplar clones corresponded well with the endophytic community structure. Out of the 513 isolates and 209 clones identified, Actinobacteria, in particular the family Microbacteriaceae, made up the largest fraction of the isolates, whereas the clone library was dominated by Alpha- and Betaproteobacteria. The most abundant genera among the isolates were Pseudomonas and Curtobacterium, while Sphingo- monas prevailed among the clones. Introduction Endophytic bacteria reside within the inner parts of plant hosts without causing disease symptoms and can be isolated from surface-disinfested plants or extracted from internal plant tissue (Hallmann et al., 1997). Various reports indi- cated that endophytic bacteria exist in a variety of tissue types within a broad range of plants, suggesting an ubiqui- tous existence in nearly all higher plants. After penetration, endophytes may colonize the plant systematically with bacterial colonies and biofilms, residing latently in the intercellular spaces, inside the vascular tissue or within cells (Jacobs et al., 1985; Hurek et al., 1994; Di Fiori & Del Gallo, 1995). Although the interaction between endophytic bacter- ia and their host plants is not fully understood, many isolates showed beneficial effects on their hosts and may play an important role in the physiology of the plant. Several bacterial endophytes have been reported to support plant growth by providing phytohormones, low-molecular compounds or enzymes (Lambert & Joos, 1989; Frommel et al., 1991; Glick et al., 1998). The colonization of ecological niches is similar to that of phytopathogens. The release of antimicrobial substances such as antibiotics or HCN (Ban- gera & Thomashow, 1996; Blumer & Haas, 2000), the production of siderophores (O’Sullivan & O’Gara, 1992) or the induction of systemic resistance to pathogens (Liu et al., 1995; Madhaiyan et al., 2004) favour them as candidates for biological control. On the other hand, many well-known plant pathogens could also be identified as typical endophy- tic bacteria that cause no disease symptoms (Kobayashi & Palumbo, 2000), but could become pathogenic under cer- tain conditions or within different host genotypes (Misaghi & Donndelinger, 1990). Plants can be colonized simultaneously by a large variety of endophytic bacteria. The variety of bacteria that has been reported as endophytes spans a significant range of Gram- positive and Gram-negative bacteria to include genera of Alpha-, Beta- and Gammaproteobacteria, Actinobacteria, FEMS Microbiol Ecol 63 (2008) 169–180 c 2007 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Diversity of endophytic bacterial communities in poplar grown under field conditions

R E S E A R C H A R T I C L E
Diversityof endophytic bacterial communities in poplargrownunder¢eld conditionsKristina Ulrich1, Andreas Ulrich2 & Dietrich Ewald1
1Bundesforschungsanstalt fur Forst- und Holzwirtschaft, Institut fur Forstgenetik- und Forstpflanzenzuchtung, Waldsieversdorf, Germany; and2Leibniz-Zentrum fur Agrarlandschaftsforschung (ZALF), Institut fur Landschaftsstoffdynamik, Muncheberg, Germany
Correspondence: Kristina Ulrich,
Bundesforschungsanstalt fur Forst- und
Holzwirtschaft, Institut fur Forstgenetik- und
Forstpflanzenzuchtung, Eberswalder
Chaussee 3a, D-15377 Waldsieversdorf,
Germany. Tel.: 149 33433 157 175; fax: 149
33433 157 199; e-mail:
Received 31 January 2007; revised 12 October
2007; accepted 16 October 2007.
First published online January 2008.
DOI:10.1111/j.1574-6941.2007.00419.x
Editor: Kornelia Smalla
Keywords
endophytic bacteria; bacterial community
composition; poplar; 16S rRNA gene.
Abstract
Bacterial endophytes may be important for plant health and other ecologically
relevant functions of poplar trees. The composition of endophytic bacteria
colonizing the aerial parts of poplar was studied using a multiphasic approach.
The terminal restriction fragment length polymorphism analysis of 16S rRNA
genes demonstrated the impact of different hybrid poplar clones on the endophytic
community structure. Detailed analysis of endophytic bacteria using cultivation
methods in combination with cloning of 16S rRNA genes amplified from plant
tissue revealed a high phylogenetic diversity of endophytic bacteria with a total of
53 taxa at the genus level that included Proteobacteria, Actinobacteria, Firmicutes
and Bacteroidetes. The community structure displayed clear differences in terms of
the presence and relative proportions of bacterial taxa between the four poplar
clones studied. The results showed that the genetic background of the hybrid
poplar clones corresponded well with the endophytic community structure. Out of
the 513 isolates and 209 clones identified, Actinobacteria, in particular the family
Microbacteriaceae, made up the largest fraction of the isolates, whereas the clone
library was dominated by Alpha- and Betaproteobacteria. The most abundant
genera among the isolates were Pseudomonas and Curtobacterium, while Sphingo-
monas prevailed among the clones.
Introduction
Endophytic bacteria reside within the inner parts of plant
hosts without causing disease symptoms and can be isolated
from surface-disinfested plants or extracted from internal
plant tissue (Hallmann et al., 1997). Various reports indi-
cated that endophytic bacteria exist in a variety of tissue
types within a broad range of plants, suggesting an ubiqui-
tous existence in nearly all higher plants. After penetration,
endophytes may colonize the plant systematically with
bacterial colonies and biofilms, residing latently in the
intercellular spaces, inside the vascular tissue or within cells
(Jacobs et al., 1985; Hurek et al., 1994; Di Fiori & Del Gallo,
1995). Although the interaction between endophytic bacter-
ia and their host plants is not fully understood, many
isolates showed beneficial effects on their hosts and may
play an important role in the physiology of the plant. Several
bacterial endophytes have been reported to support
plant growth by providing phytohormones, low-molecular
compounds or enzymes (Lambert & Joos, 1989; Frommel
et al., 1991; Glick et al., 1998). The colonization of ecological
niches is similar to that of phytopathogens. The release of
antimicrobial substances such as antibiotics or HCN (Ban-
gera & Thomashow, 1996; Blumer & Haas, 2000), the
production of siderophores (O’Sullivan & O’Gara, 1992) or
the induction of systemic resistance to pathogens (Liu et al.,
1995; Madhaiyan et al., 2004) favour them as candidates for
biological control. On the other hand, many well-known
plant pathogens could also be identified as typical endophy-
tic bacteria that cause no disease symptoms (Kobayashi &
Palumbo, 2000), but could become pathogenic under cer-
tain conditions or within different host genotypes (Misaghi
& Donndelinger, 1990).
Plants can be colonized simultaneously by a large variety
of endophytic bacteria. The variety of bacteria that has been
reported as endophytes spans a significant range of Gram-
positive and Gram-negative bacteria to include genera of
Alpha-, Beta- and Gammaproteobacteria, Actinobacteria,
FEMS Microbiol Ecol 63 (2008) 169–180 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved

Firmicutes and Bacteroidetes (Lodewyckx et al., 2002; Bacon
& Hinton, 2006). Most studies on the occurrence of
endophytic bacteria have been achieved using culture-de-
pendent approaches. However, due to the unknown growth
requirements of many bacteria and the presence of bacterial
cells that are in a viable but noncultivable state (Tholozan
et al., 1999), culture-dependent biodiversity studies of the
endophytic community are somewhat limited. Various
reports concerning endophytic bacteria in agricultural
plants demonstrated that the use of fingerprinting techni-
ques and clone analysis can provide additional information
for analysing the community composition of endophytic
bacteria (Chelius & Triplett, 2001; Garbeva et al., 2001;
Seghers et al., 2004; Sessitsch et al., 2004).
Although most of the studies concerning bacterial en-
dophytes were focused on agricultural and horticultural
plant species, endophytic bacteria were also detected in trees
like elm, pine, oak, citrus and coffee (Bacon & Mead, 1971;
Gardner et al., 1982; Araujo et al., 2002; Bent & Chanway,
2002; Mocali et al., 2003; Vega et al., 2005). For poplar trees,
which become more and more important not only as a
future energy source but also as ‘the trees of choice’ for
phytoremediation purposes and as target species for genetic
transformations (Van Aken et al., 2004b; Fladung & Ewald,
2006), most studies described only specific endophytic
strains (Germaine et al., 2004; Van Aken et al., 2004a; Doty
et al., 2005). In contrast, little is known about the commu-
nity composition of endophytic bacteria in poplar. Recently,
Moore et al. (2006) analysed the composition of culturable
endophytic bacteria in poplar grown on a benzene, toluene,
ethylbenzene and xylene (BTEX)-contaminated site and
reported a high diversity of isolates dominated by Gamma-
proteobacteria, especially Pseudomonas spp.
In this paper, the diversity of endophytic bacteria in aerial
parts of hybrid poplars grown in field trials under natural
conditions was systematically analysed using a combination
of culturing methods and direct amplification of 16S rRNA
genes from plant tissue. In addition, the community compo-
sition was compared with respect to different plant clones.
Materials and methods
Plant material and study sites
Samples were collected from four hybrid poplar clones
in summer 2005. The intersectional hybrid poplar clone
741 , [Populus alba L.� (Populus davidiana Dode1Populus
simonii Carr.)� Populus tomentosa Carr.] (Prof. Han, Beij-
ing, China) is derived from the species of two different
sections (Leuce and Tacamahaca) of the genus Populus,
while the hybrid clones Esch5 , (P. tremula L.�Populus
tremuloides Michx.), Brauna11 , (P. tremula L.) and W52 <(P. tremula L.) belong to the section Leuce. The plants
originated from cuttings from several trees and were grown
in experimental fields. The samples were obtained from 2-
year-old shoots in a clonal archive based on 10-year-old
stools that were cut down every second year. The clones 741
and Esch5/W were sampled at Waldsieversdorf, while the
clones Brauna 11, W52 and Esch5/G at Großhansdorf. The
field site Waldsieversdorf is characterized by a continental
climate with an average rainfall of 530 mm and nutrient-
poor sandy soils. In contrast, the Großhansdorf site has an
Atlantic climate with rainfalls of about 780 mm and is
located on a loamy sand.
Sample preparation
For terminal restriction fragment length polymorphism
(T-RFLP) analysis, 4 g of leaves and 4 g of branch sections
of about 2 cm (Ø = 5 mm) were sampled from four indivi-
dual trees as independent replicates for each plant clone. For
the screening of isolates and cloning, pooled samples from
five to 10 plants per poplar clone were used. The samples
were washed with tap water and surface-disinfected with
HgCl2. Leaves were disinfected in 0.1% HgCl2 containing a
drop of Tween 20 for 10 min. A higher concentration of
HgCl2 (0.25%) was used for branch sections. After three
rinses in sterile tap water, the bark of surface-disinfected
branches was removed with a sterilized razor blade, and the
branches were cut into 3–6 mm long pieces. To confirm that
the disinfection process was successful, the plant material
(pieces of the bark and leaves) was placed on nutrient agar
and the plates were examined for growth after incubation at
28 1C for 6 days. Nonsterile samples were discarded. The
samples were ground in 4 mL of sterile 0.85% NaCl with a
sterile mortar and pestle. Branch and leaf tissue extracts
were pooled to analyse the aerial endophytic bacterial
community.
T-RFLP
Bacterial pellets from ground plant material were obtained
by differential centrifugation and stored at � 80 1C. The
pellets were homogenized in Eppendorf tubes containing a
lysing matrix (Fast DNA Spin Kit for Soil; QBIOgene,
Carlsbad), using a shaking mill (Retsch GmbH, Haan,
Germany) for 4 min at maximum speed and the total DNA
was extracted following a miniaturized CTAB-based proto-
col (Dumolin et al., 1995). The bacterial primers 799f
(Chelius & Triplett, 2001) labelled at the 50-end with
6-carboxyfluorescein (6-FAM) and 1525r (Lane, 1991) were
used to amplify a fragment of c. 760 bp of the 16S rRNA
gene. PCR reaction (50mL) was performed using AccuPrime
polymerase with buffer II (Invitrogen, Carlsbad) to which
10 ng of template DNA were added. Thermal cycling condi-
tions were: 2 min denaturation at 94 1C followed by 35 cycles
of 20 s at 94 1C, 40 s at 53 1C and 60 s at 68 1C, and a final
FEMS Microbiol Ecol 63 (2008) 169–180c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
170 K. Ulrich et al.

extension for 7 min at 68 1C. The PCR products were
examined in a 1% agarose gel and subsequently purified
with an Invisorbs Spin PCRapid Kit (Invitek, Berlin,
Germany). T-RFLP analysis was performed as described by
Ulrich & Becker (2006) with HhaI-digested PCR products.
The samples were separated with GeneScan 1000 Rox
(Applied Biosystems) as an internal size standard on an
ABI 310 DNA sequencer (Applied Biosystems) using POP6
polymer. Data analysis of the T-RFLP profiles was per-
formed using the GELCOMPAR II software v. 2.5 (Applied
Maths, Saint-Martens-Latem, Belgium) and the ABI files
were converted into the GELCOMPAR curve format. To consider
both the presence and the relative abundance of terminal
restriction fragments (T-RFs), densitometric curves of
whole profiles were analysed through Pearson’s correlation.
The resulting similarity matrix was the basis for clustering
by the Ward algorithm (Ward, 1963). T-RFLP profiles in the
range of 35–740 bp were used for the cluster analysis. To test
for significant differences between T-RFLP profiles, the
Pearson’s similarity matrix was used for a permutation test
as described by Smalla et al. (2007). The significance of
effects was indicated by the difference of mean within-group
and between-group similarities (Kropf et al., 2004).
Screening of isolates
For the isolation of endophytic bacteria, ground tissue
extracts were serially diluted in 0.85% NaCl and plated on
R2A, a nutrient-poor medium suitable for the growth of
diverse plant-associated bacteria (Difco, Detroit) to recover
bacterial endophytes. Plates were incubated for 3–6 days at
28 1C. CFU were counted and expressed as CFU per gram of
fresh weight. In total, at least 80 bacterial colonies per poplar
clone were randomly selected and subcultured on R2A or
tryptic soy agar (TSA, Sigma-Aldrich Co., St Louis).
Identification of endophytic bacteria
For DNA extraction, single colonies were resuspended in
20 mL 25 mM NaOH/0.25% sodium dodecyl sulphate and
heated for 15 min at 95 1C. Aliquots (0.1mL) of the resulting
lysate were directly used for PCR without further purifica-
tion. The 16S rRNA gene fragments of the bacterial isolates
were amplified using primers 799f and 1525r. For routine
assays, a 50 mL reaction mixture containing 2.5 U Taq-
Polymerase (AppliChem, Darmstadt, Germany), 2.5 mM
MgCl2, 200 mM of each dNTP and 15 pmol of each primer,
was used. The amplifications were performed with the
following protocol: initial denaturation at 95 1C for 3 min;
25 cycles of 30 s at 94 1C, 40 s at 53 1C, 1.5 min at 72 1C,
followed by a single final extension at 72 1C for 8 min and
a final soak at 4 1C. PCR products were examined electro-
phoretically in a 1% agarose gel. The PCR products were
purified with an Invisorbs Spin PCRapid Kit (Invitek,
Berlin, Germany). DNA sequencing was performed using a
BigDye terminator cycle sequencing Kit (Applied Biosys-
tems, Foster City) and the primer 1492r. As suggested by
Stackebrandt & Rainey (1996), single strand sequencing of
about 400–600 bp was applied, which was sufficient for a
precise taxonomic placement of the 513 isolates in commu-
nity analysis. The assignment of the isolates at genus or
family level was based on the BLAST analysis and on the
taxonomic assignment by the CLASSIFIER program of the RDP.
The composition of the identified genera of culturable
endophytic bacteria was analysed by UPGMA clustering
using the percent disagreement method (STATISTICA 6.1,
StatSoft, Tulsa). For phylogenetic analysis, 30 isolates (one
for each identified taxum) were chosen for sequencing both
complementary strands with the primers 1492r and 799f.
Cloning of bacterial 16S rRNA genes
Isolation of total DNA from the endophytic community of
the poplar clones Esch5/W and 741 was performed as
described for T-RFLP analysis. The 16S rRNA gene frag-
ments were amplified using the same PCR primers as above,
purified using the Invisorbs Spin PCRapid Kit (Invitek)
and cloned into pCR2.1 (TA Clonings Kit, Invitrogen,
Carlsbad). About 100 positive clones per sample were
selected. Sequencing was performed as described for the
isolates. 209 clones were identified by single strand sequen-
cing of about 400–600 bp and 51 clones were chosen for
phylogenetic analysis. The partial 16S rRNA gene sequences
were evaluated by the program CHIMERA CHECK of the
Ribosomal Database Project (RDP, http://rdp8.cme.msu.
edu/cgis/chimera.cgi?su=SSU; Cole et al., 2005) to detect
chimeric artefacts. Clones harbouring a sequence with a
potential chimeric structure were excluded from further
analyses.
Phylogenetic analysis
The DNA sequences of representative isolates and clones, as
well as various 16S rRNA gene sequences for selected
reference strains were aligned by the CLUSTALW algorithm
(programme version 1.74; Thompson et al., 1994). The
phylogenetic trees were constructed using the Neighbor-
Joining and Maximum-Likelihood algorithms (PHYLIP com-
puter program package, version 3.6a2; Felsenstein, 1993).
The Neighbor-Joining algorithm (Saitou & Nei, 1987) was
based on a matrix of pair-wise distances corrected for
multiple base substitutions (Kimura, 1980) with a transi-
tion/transversion ratio of 2.0. The Maximum-Likelihood
method (Felsenstein, 1981) was applied with three jumbles
of the data set (randomized input order) and without global
rearrangement. The unrooted trees were calculated using the
16S rRNA gene sequence of Escherichia coli (J01695) or
Microbacterium lacticum (D21343) as an outgroup. The 16S
FEMS Microbiol Ecol 63 (2008) 169–180 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
171Endophytic bacteria in poplar

rRNA gene sequences obtained in this study (one isolate or
clone per taxum identified) were deposited in the EMBL
nucleotide sequence database under the accession nos.
AM489612–AM489640, AM489641–AM489691 and
AM698087.
Results
Molecular fingerprinting of endophyticbacterial communities
T-RFLP analysis of 16S rRNA genes resulted in comparable
profiles for the aerial endophytic bacterial community of the
four poplar clones studied. The profiles showed eight to 14
dominant T-RFs and several small T-RFs. The profiles of the
individual trees of poplar clone 741 displayed the highest
number of T-RFs and two dominant T-RF’s (440 and
664 bp), which were not identified in the profiles of the
endophytic community of the other poplar clones. As a
result of analysing the densitometric curves of the profiles,
the similarity of the bacterial communities is shown in
Fig. 1. The four replicates (individual trees) of three poplar
clones formed separate branches. In the case of Esch5/W,
three replicates clustered together. The test showed signifi-
cant results in the global comparison (Po 0.001) as well as
in all pair-wise comparisons (P = 0.029) except for the
comparison of Esch5 with Brauna11 and with 741. This
demonstrated a higher similarity within a poplar clone than
between the different poplar clones. The first branch of the
dendrogram clearly differentiates the intersectional hybrid
poplar 741 from the other clones indicating the most
evident differences in the community structure of that
poplar clone.
Culturable endophytic bacteria
To describe the diversity of endophytic bacteria in poplar, as
well as differences between the poplar clones in more detail,
culturable bacteria from pooled samples were analysed
from hybrid poplar clones (Esch5 was sampled from
two locations). Population densities of endophytic bacteria
recovered from R2A medium ranged from 8� 104 to
6� 105 CFU g�1 fresh weight. A total of 513 bacterial isolates
was identified at the genus level by partial sequencing of the
16S rRNA genes. The endophytic isolates obtained from
all poplar clones could be assigned to 27 different genera
(in one case to unclassified Microbacteriaceae). The majority
of isolates were related to common plant-associated,
endophytic or soil bacteria. Actinobacteria dominated the
collection of isolates, comprising 49% of the total isolates,
which includes Microbacteriaceae (47% of total isolates),
Sanguibacteriaceae (2%) and one isolate of Nocardiaceae.
The genera Curtobacterium (19%) and Plantibacter (9%)
represented the majority of the Microbacteriaceae. Gamma-
proteobacteria were the second most abundant class (28%
of total isolates) dominated by Pseudomonas (19%) and
Xanthomonas (7%) species. Alphaproteobacteria comprised
17% of the total isolates, represented largely by Sphingomo-
nas spp. (10%) and Methylobacterium spp. (5%). Bacteria
belonging to Bacteroidetes were represented mainly by
the genus Pedobacter and formed only 4% of the total
isolates. Firmicutes (Paenibacillus) comprised only 1.4% of
the total isolates.
The composition of the culturable endophytic bacteria
found in the single poplar clones is demonstrated in Fig. 2a.
The poplar clones showed clear differences in terms of the
relative proportions of Alpha- and Gammaproteobacteria as
well as Actinobacteria. Bacteroidetes isolates could only be
found in the Brauna11 and W52 clones. The seven isolates
belonging to Firmicutes were obtained from W52. Moreover,
differences in presence and relative proportions of the
identified genera were obvious between the poplar clones
(Table 1). The poplar clone 741 showed the highest diversity
with 17 identified genera, followed by Esch5/G and W52
with 14 and 12 genera. Esch5/W and Brauna11 comprised
10 and 9 genera, respectively. In Esch5 from both sites, the
same genera of Microbacteriaceae could be detected and the
composition of Gammaproteobacteria was similar, too.
The poplar clones Brauna11 and W52 displayed similarities
in their endophytic isolates with high proportions of Pseu-
domonas (33% and 46%) and Curtobacterium (40% and
28%). Moreover, isolates belonging to the genera Chryseo-
bacterium and Pedobacter could only be obtained from the
P. tremula clones Brauna11 and W52. The similarity of the
Fig. 1. Dendrogram showing the similarity between the T-RFLP profiles
of the endophytic bacterial community derived from the four hybrid
poplar clones (four replicates).
FEMS Microbiol Ecol 63 (2008) 169–180c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
172 K. Ulrich et al.
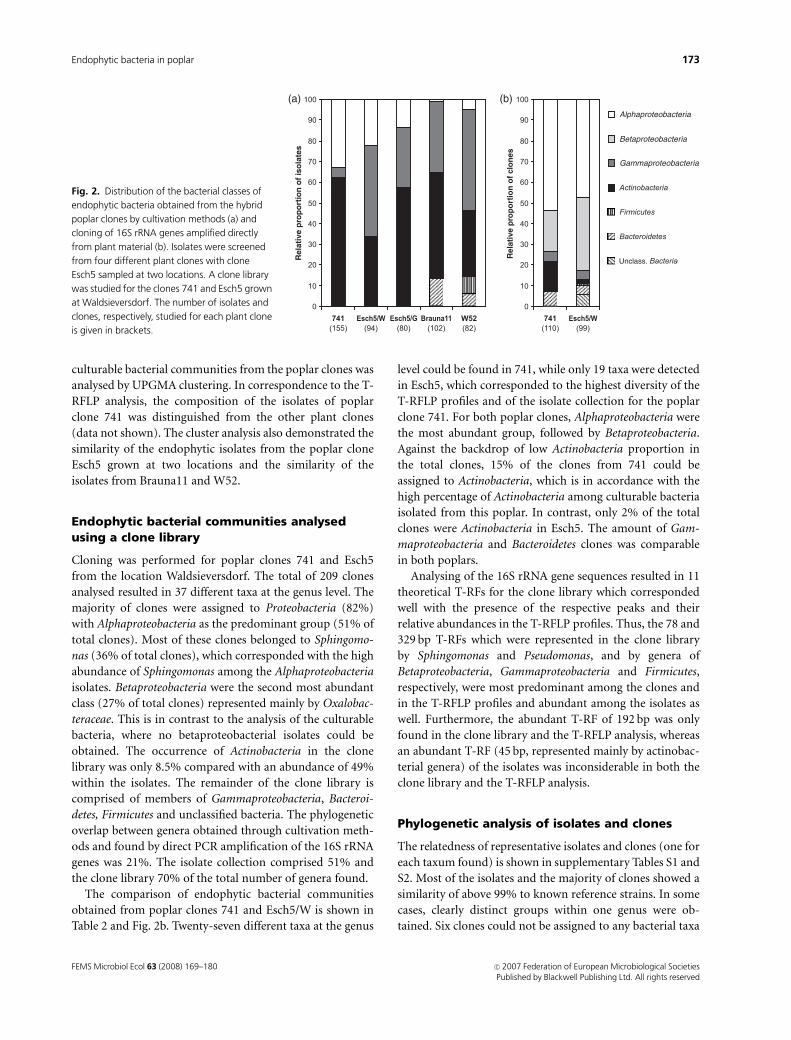
culturable bacterial communities from the poplar clones was
analysed by UPGMA clustering. In correspondence to the T-
RFLP analysis, the composition of the isolates of poplar
clone 741 was distinguished from the other plant clones
(data not shown). The cluster analysis also demonstrated the
similarity of the endophytic isolates from the poplar clone
Esch5 grown at two locations and the similarity of the
isolates from Brauna11 and W52.
Endophytic bacterial communities analysedusing a clone library
Cloning was performed for poplar clones 741 and Esch5
from the location Waldsieversdorf. The total of 209 clones
analysed resulted in 37 different taxa at the genus level. The
majority of clones were assigned to Proteobacteria (82%)
with Alphaproteobacteria as the predominant group (51% of
total clones). Most of these clones belonged to Sphingomo-
nas (36% of total clones), which corresponded with the high
abundance of Sphingomonas among the Alphaproteobacteria
isolates. Betaproteobacteria were the second most abundant
class (27% of total clones) represented mainly by Oxalobac-
teraceae. This is in contrast to the analysis of the culturable
bacteria, where no betaproteobacterial isolates could be
obtained. The occurrence of Actinobacteria in the clone
library was only 8.5% compared with an abundance of 49%
within the isolates. The remainder of the clone library is
comprised of members of Gammaproteobacteria, Bacteroi-
detes, Firmicutes and unclassified bacteria. The phylogenetic
overlap between genera obtained through cultivation meth-
ods and found by direct PCR amplification of the 16S rRNA
genes was 21%. The isolate collection comprised 51% and
the clone library 70% of the total number of genera found.
The comparison of endophytic bacterial communities
obtained from poplar clones 741 and Esch5/W is shown in
Table 2 and Fig. 2b. Twenty-seven different taxa at the genus
level could be found in 741, while only 19 taxa were detected
in Esch5, which corresponded to the highest diversity of the
T-RFLP profiles and of the isolate collection for the poplar
clone 741. For both poplar clones, Alphaproteobacteria were
the most abundant group, followed by Betaproteobacteria.
Against the backdrop of low Actinobacteria proportion in
the total clones, 15% of the clones from 741 could be
assigned to Actinobacteria, which is in accordance with the
high percentage of Actinobacteria among culturable bacteria
isolated from this poplar. In contrast, only 2% of the total
clones were Actinobacteria in Esch5. The amount of Gam-
maproteobacteria and Bacteroidetes clones was comparable
in both poplars.
Analysing of the 16S rRNA gene sequences resulted in 11
theoretical T-RFs for the clone library which corresponded
well with the presence of the respective peaks and their
relative abundances in the T-RFLP profiles. Thus, the 78 and
329 bp T-RFs which were represented in the clone library
by Sphingomonas and Pseudomonas, and by genera of
Betaproteobacteria, Gammaproteobacteria and Firmicutes,
respectively, were most predominant among the clones and
in the T-RFLP profiles and abundant among the isolates as
well. Furthermore, the abundant T-RF of 192 bp was only
found in the clone library and the T-RFLP analysis, whereas
an abundant T-RF (45 bp, represented mainly by actinobac-
terial genera) of the isolates was inconsiderable in both the
clone library and the T-RFLP analysis.
Phylogenetic analysis of isolates and clones
The relatedness of representative isolates and clones (one for
each taxum found) is shown in supplementary Tables S1 and
S2. Most of the isolates and the majority of clones showed a
similarity of above 99% to known reference strains. In some
cases, clearly distinct groups within one genus were ob-
tained. Six clones could not be assigned to any bacterial taxa
(a) (b)
Fig. 2. Distribution of the bacterial classes of
endophytic bacteria obtained from the hybrid
poplar clones by cultivation methods (a) and
cloning of 16S rRNA genes amplified directly
from plant material (b). Isolates were screened
from four different plant clones with clone
Esch5 sampled at two locations. A clone library
was studied for the clones 741 and Esch5 grown
at Waldsieversdorf. The number of isolates and
clones, respectively, studied for each plant clone
is given in brackets.
FEMS Microbiol Ecol 63 (2008) 169–180 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
173Endophytic bacteria in poplar

based on the 16S rRNA gene sequence similarity. Five of
these clones, represented by Esch5-32 in supplementary
Table S2, were closely related with an uncultured bacterium
clone from the cryptoendolithic communities from McMur-
do Dry Valleys of Antarctica (De la Torre et al., 2003). The
clone Esch5-52 showed a strong similarity to an unclassified
clone obtained from volcanic deposits in Hawaii (supple-
mentary Table S2).
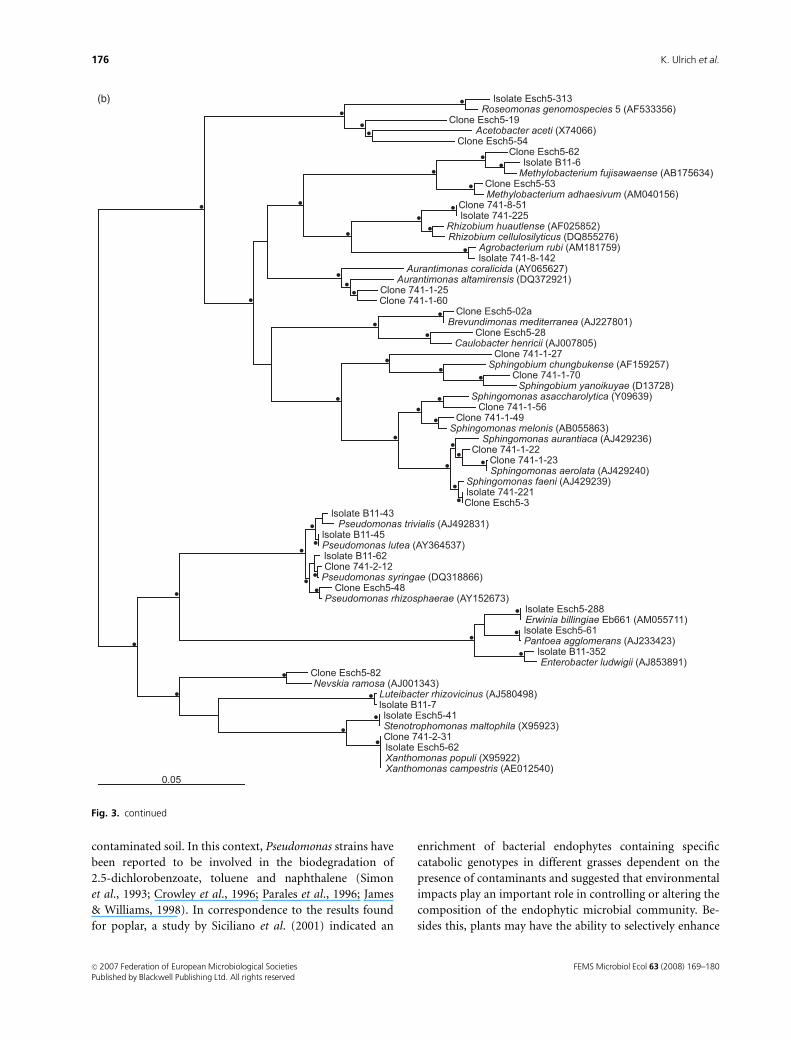
The phylogenetic relationship of isolates and clones in
comparison to the type strains of related species is demon-
strated for three selected bacterial classes (Fig. 3). The
Actinobacteria as the most abundant group of isolates
displayed a high diversity of phylogenetically related genera.
Actinobacterial isolates often showed a close relationship to
the species of the family Microbacteriaceae, while the clones
belonged to species of other families of Actinobacteria, as
well as Microbacteriaceae (Fig. 3a). Thus, more families of
the Actinobacteria could be found by cloning, even though
only a minor part of the total clones was assigned to
Table 1. Composition of endophytic isolates obtained from the four
hybrid poplar clones
Identified taxum
Waldsieversdorf� Großhansdorf
741 Esch5/W Esch5/G Brauna11 W52
Alphaproteobacteria
Agrobacterium 3w
Rhizobium 7 2 1
Methylobacterium 9 7 6 1
Roseomonas 1
Sphingomonas 32 14 2 3
Gammaproteobacteria
Enterobacter 1
Erwinia 1 1
Pantoea 3
Pseudomonas 1 23 3 34 38
Luteibacter 1
Stenotrophomonas 3 1
Xanthomonas 2 15 19
Actinobacteria
Curtobacterium 3 19 10 41 23
Clavibacter 5 1 17 3
Plantibacter 39 3 1 1
Okibacterium 18
Rathayibacter 4 11
Agreia 1
Microbacterium 17 3 1
Frigoribacterium 1 5 4
Cryocola 5
Unclassified
Microbacteriaceae
2 2 1
Sanguibacter 11
Nocardioides 1
Firmicutes
Paenibacillus 7
Bacteroidetes
Chryseobacterium 2 3
Pedobacter 12 2
Total isolates 155 94 80 102 82
�The clones 741 and Esch5/W were sampled at Waldsieversdorf, while
the clones Brauna 11, W52 and Esch5/G at Großhansdorf.wThe number of isolates obtained from the respective poplar clone is
presented for each identified taxum.
Table 2. Composition of endophytic bacteria obtained from two hybrid
poplar clones by cloning of 16S rRNA genes amplified directly from plant
material
Identified taxum 741� Esch5/W
Alphaproteobacteria
Caulobacter 3w
Brevundimonas 1
Sphingomonas 43 32
Sphingobium 2
Methylobacterium 5 9
Rhizobium 2
Acetobacteraceae 2
Aurantimonadaceae 7
Betaproteobacteria
Acidovorax 1 1
Variovorax 4 1
Comamonadaceae 3
Oxalobacter 10
Massilia 5 3
Oxalobacteraceae 3 14
Xylophilus 3
Leptothrix 1
Burkholderiales genera incertae sedis 5 1 2
Alcaligenaceae 1
Burkholderia 3
Neisseriales 1
Gammaproteobacteria
Pseudomonas 3 2
Xanthomonas 2
Xanthomonadaceae 2
Actinobacteria
Frigoribacterium 8 2
Curtobacterium 1
Clavibacter 1
Microbacterium 2
Beutenbergiaceae 1
Kineococcus 1
Frankineae 1
Friedmanniella 1
Firmicutes
Anaerococcus 1
Bacteroidetes
Flexibacteraceae 2
Chryseobacterium 2
Pedobacter 4
Sphingobacteriales 4
Unclassified Bacteria 6
Total clones 110 99
�The clones 741 and Esch5 were grown at Waldsieversdorf.wThe number of clones obtained from the respective poplar clone is
presented for each identified taxum.
FEMS Microbiol Ecol 63 (2008) 169–180c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
174 K. Ulrich et al.

Actinobacteria. As displayed in the phylogenetic tree, endo-
phytic bacteria related to Kineococcus, Solicoccus, Salana, and
Friedmanniella were only found in the clone library.
Alpha- and Gammaproteobacteria contained isolates or
clones belonging to distinct groups of the same genus.
Within Sphingomonas, which was found to be one of the
most abundant genera, isolates and clones related to several
species could be revealed (Fig. 3b). In total, endophytic
isolates and clones belonging to at least seven different
species of sphingomonads were found. Similar results were
obtained for the genus Pseudomonas with close relationships
to Pseudomonas trivialis, Pseudomonas lutea, Pseudomonas
syringae and Pseudomonas rhizosphaerae. Several clones
showed an identical sequence to Xanthomonas populi, which
is known as the cause of bacterial stem canker of poplar
(Ride, 1993). However, other Xanthomonas species like
Xanthomonas campestris and Xanthomonas gardneri also
had the same 16S rRNA gene sequences.
Discussion
The objective of this study was to examine the community
composition of endophytic bacteria from different poplar
clones growing under field conditions. A high phylogenetic
diversity of endophytic bacteria could be revealed with a
total of 53 taxa at the genus level that included Proteobacter-
ia, Actinobacteria, Firmicutes and Bacteroidetes. Only little
is known about the endophytic bacterial community har-
boured by poplar plants from other studies. Recently, Moore
et al. (2006) described the diversity of endophytic bacteria in
two cultivars of Populus trichocarpa� Populus deltoides
grown in a phytoremediation field trial on a site contami-
nated with BTEX compounds. High numbers of Gamma-
proteobacteria (61%) were found in the isolate collection,
which was dominated by Pseudomonas spp. that reached up
to 53% of the total isolates in one cultivar. In contrast,
Gammaproteobacteria made up only 28% of the total isolates
and 4% of the clones, with Pseudomonas species comprising
19% of the isolates and only 2% of clones in this study. The
highest percentage of Pseudomonas in a single poplar clone
was found in the W52 with about 46%. The abundance of
Pseudomonas within endophytic bacteria was found to be
highly variable and seems to be dependent on the plant
clone or environmental conditions. The high abundance of
Pseudomonas isolates in poplar grown in the phytoremedia-
tion field may be due to their relatively high amount in the
C
C
C
C
C
C
C
C
I
(a)
Fig. 3. Phylogenetic trees showing the rela-
tionship of the 16S rRNA gene sequences of
representative isolates and clones belonging to
Actinobacteria (a), as well as Alpha- and Gam-
maproteobacteria (b). The trees were generated
using the Neighbor-Joining algorithm. Branches
also found by the Maximum-Likelihood method
are indicated by dots. Type strains of each of the
shown species are used for the calculation. The
16S rRNA gene sequence of Erwinia billingiae
Eb661 (AM055711) was used instead of that of
the type strain (Y13249) to improve sequence
quality. Scale bar = 0.05 relative sequence diver-
gences.
FEMS Microbiol Ecol 63 (2008) 169–180 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
175Endophytic bacteria in poplar
contaminated soil. In this context, Pseudomonas strains have
been reported to be involved in the biodegradation of
2.5-dichlorobenzoate, toluene and naphthalene (Simon
et al., 1993; Crowley et al., 1996; Parales et al., 1996; James
& Williams, 1998). In correspondence to the results found
for poplar, a study by Siciliano et al. (2001) indicated an
enrichment of bacterial endophytes containing specific
catabolic genotypes in different grasses dependent on the
presence of contaminants and suggested that environmental
impacts play an important role in controlling or altering the
composition of the endophytic microbial community. Be-
sides this, plants may have the ability to selectively enhance
C
C
C
C
CC
C
C
C
C
CC
CC
C
C
C
C
C
C
(b)
Fig. 3. continued
FEMS Microbiol Ecol 63 (2008) 169–180c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
176 K. Ulrich et al.

the prevalence of endophytes containing specific catabolic
genes or other properties that benefit plant growth under
specific environmental conditions. Thus, the finding of a
different endophytic community in poplars grown under
normal field conditions and in BTEX-polluted soils pro-
vided further evidence that the mechanism by which plants
control their endophytic bacteria and/or environmental
conditions strongly influence the diversity and composition
of the endophytic bacterial community.
Several studies on agricultural plants revealed the endo-
phytic community as a subset of the soil community
(Germida et al., 1998; Berg et al., 2005; Gotz et al., 2006).
This is also suggested by the results of this study, because
several phylogenetically diverse bacteria, which are also
known as common soil bacteria could be found. On the
other hand, various species found in poplar are typical
phyllosphere bacteria, for example Plantibacter and Curto-
bacterium (Behrendt et al., 2002; Beattie, 2006). Thus,
penetration of the host plant via leaf stomata and stem
lenticels (Kluepfel, 1993), as well as through folia damages
(Leben et al., 1968) may also play an important role.
The T-RFLP analysis revealed significant differences be-
tween the studied poplar clones, which demonstrated the
impact of the plant clone on the endophytic community
structure. All approaches used in this study showed consis-
tently strong differences, as well as the highest diversity of
endophytic bacteria for the intersectional Chinese hybrid
poplar 741 derived from four different Populus species. On
the other hand, the P. tremula clones W52 and Brauna11
displayed a high similarity in their endophytic isolates.
Thus, the genetic background of the hybrid poplar clones
corresponded well with the endophytic community struc-
ture. The impact of plant clones was also observed for
various tree species by other authors. Cambours et al.
(2005) found differences in the endophytic pathogenic and
ice nucleation active bacteria in four Salix clones. The
diversity of endophytic bacteria was also influenced by
different plant genotypes of citrus and poplar (Araujo
et al., 2002; Moore et al., 2006). Thus, endophytic bacterial
communities seem to be dependent on the genotype of
long-living plants, suggesting that each tree species or clone
has an association to specific bacterial endophytes.
Cultivation as well as culture-independent methods were
used to analyse the endophytic community composition of
poplar clones. Both approaches revealed differences between
the poplar clones in terms of the diversity and abundance of
their bacterial classes and genera, whereas the culturable
component of the bacterial community was different from
that obtained by clone analysis. While Actinobacteria, espe-
cially the family Microbacteriaceae represented the majority
of the isolates, the clone library was dominated by Alpha-
proteobacteria and Betaproteobacteria. Some Actinobacteria
are recalcitrant to cell lysis and therefore, they might be
underrepresented in a clone library. However, the selectivity
of cultivation as well as a preferential amplification of
certain bacterial groups with the eubacterial primers could
also cause the different abundance. A disparity in the
representation of different bacterial classes, genera and
species between isolate collection and clone library had also
been observed in several other studies (Dunbar et al., 1999;
Hengstmann et al., 1999; Chelius & Triplett, 2001; Idris
et al., 2004). Therefore, the combination of culturing
methods and cloning analysis is needed for the study of the
endophytic communities of woody plants.
Curtobacterium strains have been isolated as typical
endophytes from several woody plants like sweet-orange,
coffee and grapevine (Bell et al., 1995; Araujo et al., 2002;
Vega et al., 2005). In this study, Curtobacterium could be
obtained from all poplar clones and comprised 19% of total
isolates, whereas the majority were assigned to the species
Curtobacterium flaccumfaciens. Several subspecies or strains
of C. flaccumfaciens are known as causal agents of wilt and
necrotic symptoms on horticultural and ornamental plants
(Vidaver, 1982). However, C. flaccumfaciens had also been
reported as a biocontrol agent for cucumber (Raupach &
Kloepper, 2000) and to play a role in triggering induced
systemic resistance (Raupach & Kloepper, 1998). In citrus, a
higher frequency of C. flaccumfaciens was found in asymp-
tomatic plants compared with plants infected with Xylella
fastidiosa, further suggesting the role of this bacterium in the
resistance of plants to chlorosis (Araujo et al., 2002). In this
context, it remains open whether identical Curtobacterium
strains act as pathogens or antagonistic strains in depen-
dence on the genetic background of the host plant and
environmental conditions, or if different strains of the same
species mediate such different properties.
Some isolates and clones were highly similar to Clavibac-
ter michiganensis, Pseudomonas syringae and Xanthomonas
populi, which have been historically thought to be patho-
genic (Lindow et al., 1978; Shirakawa et al., 1991; Ride, 1993;
Ramstedt et al., 1994). However, no visible symptoms of
disease had been detected on the poplar plants analysed.
In both poplars studied by cloning of 16S rRNA genes, the
highest abundance at the genus level could be found for
Sphingomonas. Among the culturable bacteria, varying
proportions (up to 21%) were detected from the studied
poplars. The high abundance found in some of the poplar
clones combined with the high phylogenetic variability of
sphingomonads suggests a putative function within the
bacterial endophytes.
Besides the discussed relation of endophytic and patho-
genic bacteria, bacterial endophytes may also be important
in forest ecosystems for increasing the phenotypic plasticity
of their long-living tree hosts under variable or deleterious
environmental conditions, e.g. during periods of drought or
nutrient deprivation (Chanway, 1998). However, in contrast
FEMS Microbiol Ecol 63 (2008) 169–180 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
177Endophytic bacteria in poplar

to agricultural plants, where many effects had been de-
scribed (Bacon & Hinton, 2006), little was known about
their interaction and influence on plant growth until now.
There is only some evidence of plant growth-promoting
activities by endophytic bacteria in trees, such as, stimula-
tory effects on seedling growth caused by endophytic
pseudomonads, Bacillus strains and actinomycetes (Gardner
et al., 1984; Chanway & Holl, 1994). However, further
studies are needed to ascertain the association between
woody plants and the wide range of different bacteria living
there as endophytes.
In conclusion, this study revealed a high phylogenetic
diversity of bacterial endophytes in poplar and showed an
influence of the plant genotype on the endophytic bacterial
community. Knowledge on the abundance and composition
of bacterial endophytes is an indispensable precondition for
future applications in areas such as plant health promotion,
phytoremediation, and studies on horizontal gene transfer
between endophytic bacteria and during Agrobacterium-
mediated plant transformation.
Acknowledgements
This work was supported by grant 0313285I from the
Federal Ministry of Education and Research. The authors
are grateful to Mrs Hannelore Enkisch and Mrs Sigune
Weinert for their excellent technical assistance, and to Dr
Siegfried Kropf for helping with the statistical analyses. The
authors also thank Prof. Yang from the Agricultural Uni-
versity Hebei, Baoding for providing the poplar clone 741
and Dr Matthias Fladung for providing plant material from
poplar clones.
References
Araujo WL, Marcon J, Maccheroni W Jr, van Elsas JD, van Vuurde
JWL & Azevedo JL (2002) Diversity of endophytic bacterial
populations and their interaction with Xylella fastidiosa in
citrus plants. Appl Environ Microbiol 68: 4906–4914.
Bacon CW & Hinton DM (2006) Bacterial endophytes: the
endophytic niche, its occupants, and its utility. Plant-
Associated Bacteria (Gnanamanickam SS, ed), pp. 155–194.
Springer, Dordrecht.
Bacon M & Mead CE (1971) Bacteria in the wood of living aspen,
pine and alder. Northwest Sci 45: 270–275.
Bangera MG & Thomashow LS (1996) Characterization of a
genomic locus required for synthesis of the antibiotic 2,4-
diacetylphloroglucinol by the biological control agent
Pseudomonas fluorescens Q2–87. Mol Plant-Microbe Interact 9:
83–90.
Beattie GA (2006) Plant-associated bacteria: survey, molecular
phylogeny, genomics and recent advances. Plant-Associated
Bacteria (Gnanamanickam SS, ed), pp. 1–56. Springer,
Dordrecht.
Behrendt U, Ulrich A, Schumann P, Naumann D & Suzuki K
(2002) Diversity of grass-associated Microbacteriaceae isolated
from the phyllosphere and litter layer after mulching the
sward; a polyphasic characterization of Subtercola pratensis sp.
nov., Curtobacterium herbarum sp. nov. and Plantibacter flavus
gen. nov., sp. nov. Int J Syst Evol Microbiol 52: 1441–1454.
Bell CR, Dickie GA, Harvey WLG & Chan JWYF (1995)
Endophytic bacteria in grapevine. Can J Microbiol 41: 46–53.
Bent E & Chanway CP (2002) Potential for misidentification of a
spore-forming Paenibacillus polymyxa isolate as an endophyte
by using culture-based methods. Appl Environ Microbiol 68:
4650–4652.
Berg G, Krechel A, Ditz M, Sikora R, Ulrich A & Hallmann J
(2005) Endophytic and ectophytic potato-associated bacterial
communities differ in structure and antagonistic function
against plant pathogenic fungi. FEMS Microbiol Ecol 51:
215–229.
Blumer C & Haas D (2000) Mechanism, regulation, and
ecological role of bacterial cyanide biosynthesis. Arch Microbiol
173: 170–177.
Cambours MA, Nejad P, Granhall U & Ramstedt M (2005) Frost-
related dieback of willows. Comparison of epiphytically and
endophytically isolated bacteria from different Salix clones, with
emphasis on ice nucleation activity, pathogenic properties and
seasonal variation. Biomass Bioenergy 28: 15–27.
Chanway CP (1998) Bacterial endophytes: ecological and
practical implications. Sydowia 50: 149–170.
Chanway CP & Holl FB (1994) Growth of outplanted lodgepole
pine seedlings one year after inoculation with plant growth
promoting rhizobacteria. For Sci 40: 238–246.
Chelius MK & Triplett EW (2001) The diversity of archaea and
bacteria in association with the roots of Zea mays L. Microbial
Ecol 41: 252–263.
Cole JR, Chai B, Farris RJ, Wang Q, Kulam SA, McGarrell DM,
Garrity GM & Tiedje JM (2005) The ribosomal database
project (RDP-II): sequences and tools for high-throughput
rRNA analysis. Nucl Acids Res 1 (Database Issue): D294–D296.
doi: 10.1093/nar/gki038.
Crowley DE, Brennerova MV, Irwin C, Brenner V & Focht DD
(1996) Rhizosphere effects on biodegradation of 2,5-dichloro-
benzoate by a bioluminescent strain of root-colonizing
Pseudomonas fluorescens. FEMS Microbiol Ecol 20: 79–89.
De la Torre JR, Goebel BM, Friedmann EM & Pace NR (2003)
Microbial diversity of cryptoendolithic communities from the
McMurdo Dry Valleys, Antarctica. Appl Environ Microbiol 69:
3858–3867.
Di Fiori S & Del Gallo M (1995) Endophytic bacteria: their
possible role in the host plants. Azospirillum VI and Related
Microorganisms (Fendrik I, Del Gallo M, Vanderleyden J & De
Zamaroczy M, eds), pp. 169–187. Springer, Berlin.
Doty SL, Dosher MR, Singleton GL, Moore AL, Van Aken B,
Stettler RF, Strand SE & Gordon MP (2005) Identification of
an endophytic Rhizobium in stems of Populus. Symbiosis 39:
27–35.
FEMS Microbiol Ecol 63 (2008) 169–180c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
178 K. Ulrich et al.

Dumolin S, Demesure B & Petit RJ (1995) Inheritance of
chloroplast and mitochondrial genomes in pedunculate oak
investigated with an efficient PCR method. Theor Appl Genet
91: 1253–1256.
Dunbar J, Takala S, Barns SM, Davis JA & Kuske CR (1999) Levels
of bacterial community diversity in four arid soils compared
by cultivation and 16S rRNA gene cloning. Appl Environ
Microbiol 65: 1662–1669.
Felsenstein J (1981) Evolutionary tree from DNA sequences: a
maximum likelihood approach. J Mol Evol 17: 368–376.
Felsenstein J (1993) PHYLIP (Phylogeny Interference Package),
3.5c edn. University of Washington, Seattle.
Fladung M & Ewald D (2006) Tree Transgenesis. Recent
Developments. Springer, Berlin Heidelberg.
Frommel MI, Nowak J & Lazarovits G (1991) Growth
enhancement and developmental modifications of in vitro
grown potato (Solanum tuberosum ssp. tuberosum) as affected
by nonfluorescent Pseudomonas sp. Plant Physiol 96: 928–936.
Garbeva P, van Overbeek LS, van Vuurde JWL & van Elsas JD
(2001) Analysis of endophytic bacterial communities of potato
by plating and denaturing gradient gel electrophoresis (DGGE)
of 16S rDNA based PCR fragments. Microb Ecol 41: 369–383.
Gardner JM, Feldman AW & Zablotowicz RM (1982) Identity
and behaviour of xylem-residing bacteria in rough lemon
roots of Florida citrus trees. Appl Environ Microbiol 43:
1335–1342.
Gardner JM, Chandler JL & Feldman AW (1984) Growth
promotion and inhibition by antibiotic-producing fluorescent
pseudomonads on citrus roots. Plant Soil 77: 103–113.
Germaine K, Keogh E, Garcia-Cabellos G et al. (2004)
Colonization of poplar trees by gfp expressing bacterial
endophytes. FEMS Microbiol Ecol 48: 109–118.
Germida JJ, Siciliano SD, de Freitas JR & Seib AM (1998)
Diversity of root associated bacteria associated with held-
grown canola (Brassica napus L.) and wheat (Triticum
aestivum L.). FEMS Microbiol Ecol 26: 43–50.
Glick BR, Penrose DM & Li J (1998) A model for the lowering of
plant ethylene concentrations by plant growth promoting
bacteria. J Theor Biol 190: 63–68.
Gotz M, Nirenberg H, Krause S, Wolters H, Draeger S, Buchner
A, Lottmann J, Berg G & Smalla K (2006) Fungal endophytes
in potato roots studied by traditional isolation and
cultivation-independent DNA-based methods. FEMS
Microbiol Ecol 58: 404–413.
Hallmann J, Quadt-Hallmann A, Mahaffee WF & Kloepper JW
(1997) Bacterial endophytes in agricultural crops. Can J
Microbiol 43: 895–914.
Hengstmann U, Chin KJ, Janssen PH & Liesack W (1999)
Comparative phylogenetic assignment of environmental
sequences of genes encoding 16S rRNA and numerically
abundant culturable bacteria from an anoxic rice paddy soil.
Appl Environ Microbiol 65: 5050–5058.
Hurek T, Reinhold-Hurek B, Van Montagu M & Kellenberger E
(1994) Root colonization and systemic spreeding of Azoarcus
sp. strain BH72 in grasses. J Bacteriol 176: 1913–1923.
Idris R, Trifonova R, Puschenreiter M, Wenzel WW & Sessitsch A
(2004) Bacterial communities associated with flowering plants
of the Ni hyperaccumulator Thlaspi goesingense. Appl Environ
Microbiol 70: 2667–2677.
Jacobs MJ, Bugbee WM & Gabrielson DA (1985) Enumeration,
location, and characterization of endophytic bacteria within
sugar beet roots. Can J Bot 63: 1262–1265.
James KD & Williams PA (1998) Ntn genes determining the early
steps in the divergent catabolism of 4-nitrotoluene and toluene
in Pseudomonas sp. strain TW3. J Bacteriol 180: 2043–2049.
Kimura M (1980) A simple method for estimating evolutionary
rates of base substitutions through comparative studies of
nucleotide sequences. J Mol Evol 16: 111–120.
Kluepfel DA (1993) The behavior and tracking of bacteria in the
rhizosphere. Ann Rev Phytopathol 31: 441–472.
Kobayashi DY & Palumbo JD (2000) Bacterial endophytes and
their effects on plants and uses in agriculture. Microbial
Endophytes (Bacon CW & White JF, eds), pp. 199–233. Marcel
Dekker, New York.
Kropf S, Heuer H, Gruning M & Smalla K (2004) Significance test
for comparing complex microbial community fingerprints
using pairwise similarity measures. J Microbiol Methods 57:
187–195.
Lambert B & Joos H (1989) Fundamental aspects of
rhizobacterial plant growth promotion research. Trends
Biotechnol 7: 215–219.
Lane DJ (1991) 16S/23S sequencing. Nucleic Acids Techniques
in Bacterial Systematics (Stackebrandt E & Goodfellow M, eds),
pp. 115–175. John Wiley & Sons, New York.
Leben C, Daft GC & Schmitthenner AF (1968) Bacterial blight of
soybeans: population levels of Pseudomonas glycinea in
relation to symptom development. Phytopathology 58:
1143–1146.
Lindow SE, Arny DC & Upper CD (1978) Distribution of ice-
nucleation active bacteria on plants in nature. Appl Environ
Microbiol 36: 831–838.
Liu L, Kloepper JW & Tuzun S (1995) Induction of systemic
resistance in cucumber against bacterial angular leaf spot by
plant growth-promoting rhizobacteria. Phytopathology 85:
843–847.
Lodewyckx C, Vangronsveld J, Porteous F, Moore ERB, Taghavi S
& Van der Lelie D (2002) Endophytic bacteria and their
potential applications. Crit Rev Plant Sci 21: 583–606.
Madhaiyan M, Poonguzhali S, Senthilkumar M, Seshadri S,
Chung H, Yang J, Sundaram S & Sa T (2004) Growth
promotion and induction of systemic resistance in rice cultivar
Co-47 (Oryza sativa L.) by Methylobacterium spp. Bot Bull
Acad Sin 45: 315–324.
Misaghi IJ & Donndelinger CR (1990) Endophytic bacteria in
symptom-free cotton plants. Phytopathology 80: 808–811.
Mocali S, Bertelli E, Di Cello F, Mengoni A, Sfalanga A, Viliani F,
Caciotti A, Tegli S, Surico G & Fani R (2003) Fluctuation of
bacteria isolated from elm tissue during different seasons and
from different plant organs. Res Microbiol 154: 105–114.
FEMS Microbiol Ecol 63 (2008) 169–180 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
179Endophytic bacteria in poplar

Moore PF, Barac T, Borremans B, Oeyen L, Vangronsveld J, Van
der Lelie D, Campbell CD & Moore ERB (2006) Endophytic
bacterial diversity in poplar trees growing on a BTEX-
contaminated site: the characterization of isolates with
potential to enhance phytoremediation. Syst Appl Microbiol 29:
539–556.
O’Sullivan DJ & O’Gara F (1992) Traits of fluorescent
Pseudomonas spp. involved in suppression of plant root
pathogens. Microbial Rev 56: 662–676.
Parales JW, Kumar A, Parales RE & Gibson DT (1996) Cloning
and sequencing of the genes encoding 2-nitrotoluene
dioxygenase from Pseudomonas sp. JS42. Gene 181: 57–61.
Ramstedt M, Astrom B & Von Fircks HA (1994) Dieback of
poplar and willow caused by Pseudomonas syringae in
combination with freezing stress. Eur J Forest Pathol 24:
305–315.
Raupach GS & Kloepper JW (1998) Mixtures of plant-growth
promoting rhizobacteria enhance biological control of
multiple cucumber pathogens. Phytopathology 88: 1158–1164.
Raupach GS & Kloepper JW (2000) Biocontrol of cucumber
diseases in the field by plant growth-promoting rhizobacteria
with and without methyl bromide fumigation. Plant Dis 84:
1073–1075.
Ride M (1993) Xanthomonas populi: cause of bacterial canker of
poplar. Xanthomonas (Swing JG & Civerol EL, eds), pp. 64–69.
Chapman and Hall, London.
Saitou N & Nei M (1987) The neighbor-joining method: a new
method for reconstructing phylogenetic trees. Mol Biol Evol 4:
406–425.
Seghers D, Wittebolle L, Top EM, Verstraete W & Siciliano SD
(2004) Impact of agricultural practices on the Zea mays L.
endophytic community. Appl Environ Microbiol 70:
1475–1482.
Sessitsch A, Reiter B & Berg G (2004) Endophytic bacterial
communities of field-grown potato plants and their plant
growth-promoting abilities. Can J Microbiol 50: 239–249.
Shirakawa T, Sasaki T & Ozaki K (1991) Ecology and control of
tomato bacterial canker and detection methods of this
pathogen. Jpn Agric Res Quart 25: 27–32.
Siciliano SD, Fortin N, Mihoc A et al. (2001) Selection of specific
endophytic bacterial genotypes by plants in response to soil
contamination. Appl Environ Microbiol 67: 2469–2475.
Simon MJ, Osslund TD, Saunders R, Ensley BD, Suggs S,
Harcourt A, Suen W, Cruden DL, Givson DT & Zylstra GL
(1993) Sequences of genes encoding naphthalene dioxygenase
in Pseudomonas putida strains G7 and NCIB 9816-4. Gene 127:
31–37.
Smalla K, Oros-Sichler M, Milling A, Heuer H, Baumgarte S,
Becker R, Neuber G, Kropf S, Ulrich A & Tebbe CC (2007)
Bacterial diversity of soils assessed by DGGE, T-RFLP and
SSCP fingerprints of PCR-amplified 16S rRNA gene
fragments: do the different methods provide similar results? J
Microbiol Methods 69: 470–479.
Stackebrandt E & Rainey F (1996) Partial and complete 16S
rDNA sequences, their use in generation of 16S rDNA
phylogenetic trees and their implications in molecular
ecological studies. Molecular Microbiology and Ecology Manual
3.1.1 (Akkermans ADL, van Elsas JD & de Bruijn FJ, eds), pp.
1–17. Kluwer Academic Publishers, Dordrecht.
Tholozan JL, Cappelier JM, Tissier G, Delattre G & Federighi M
(1999) Physiological characterization of viable-but-
nonculturable Campylobacter jejuni cells. Appl Environ
Microbiol 65: 1110–1116.
Thompson JD, Higgins DG & Gibson TJ (1994) CLUSTAL W:
improving the sensitivity of progressive multiple sequence
alignment through sequence weighting, position-specific gap
penalties and weight matrix choice. Nucl Acids Res 22:
4673–4680.
Ulrich A & Becker R (2006) Soil parent material is a key
determinant of the bacterial community structure in arable
soils. FEMS Microbiol Ecol 56: 430–443.
Van Aken B, Peres CM, Doty SL, Yoon JM & Schnoor JL (2004a)
Methylobacterium populi sp. nov., a novel aerobic, pink-
pigmented, facultatively methylotrophic, methane-utilizing
bacterium isolated from poplar trees (Populus deltoides� nigra
DN34). Int J Syst Evol Microbiol 54: 1191–1196.
Van Aken B, Yoon JM & Schnoor JL (2004b) Biodegradation of
nitro-substituted explosives 2,4,6-Trinitrotoluene,
Hexahydro-1,3,5-Trinitro-1,3,5-Triazine, and Octahydro-
1,3,5,7-Tetranitro-1,3,5-Tetrazocine by a phytosymbiotic
Methylobacterium sp. associated with poplar tissues (Populus
deltoides� nigra DN34). Appl Environ Microbiol 70: 508–517.
Vega FE, Pava-Ripoll M, Posada F & Buyer YS (2005) Endophytic
bacteria in Coffea arabica L. J Basic Microbiol 45: 371–380.
Vidaver AK (1982) The plant pathogenic corynebacteria. Annu
Rev Microbiol 36: 495–517.
Ward JH (1963) Hierarchical grouping to optimize an objective
function. J Am Stat Ass 58: 236.
Supplementarymaterial
The following supplementary material is available for this
article online:
Table S1. 16S rRNA gene sequence similarities of
representative endophytic bacterial isolates from the identi-
fied taxa.
Table S2. 16S rRNA gene sequence similarities of
representative bacterial clones of the identified taxa ob-
tained from endophytic communities of poplar clones 741
and Esch5/W.
This material is available as part of the online article
from: http://www.blackwell-synergy.com/doi/abs/10.1111/
j.1574-6941.2007.00419.x (This link will take you to the
article abstract).
Please note: Blackwell Publishing is not responsible for
the content or functionality of any supplementary materials
supplied by the authors. Any queries (other than missing
material) should be directed to the corresponding author for
the article.
FEMS Microbiol Ecol 63 (2008) 169–180c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
180 K. Ulrich et al.




















