
Identification of the dipeptidyl aminopeptidase responsible for N-terminal clipping of recombinant...
7
Identification of the dipeptidyl aminopeptidase responsible for N-terminal clipping of recombinant Exendin-4 precursor expressed in Pichia pastoris Lakshmi Prabha, Nagaraj Govindappa, Laxmi Adhikary, Ramakrishnan Melarkode, Kedarnath Sastry * Biocon Limited, Research and Development, 20th KM, Hosur Road, Electronic City P.O., Bangalore 560 100, India article info Article history: Received 5 September 2008 and in revised form 30 October 2008 Available online 8 November 2008 Keywords: Dipeptidyl aminopeptidase Pichia pastoris Saccharomyces cerevisiae STE13 DAP2 Exendin-4 abstract Exendin-4 is a naturally occurring 39 amino acid peptide that is useful for the control of Type 2 diabetes. Recombinant Exendin-4, with an extra glycine at the carboxy-terminus (Exdgly), was expressed in the methylotropic yeast Pichia pastoris. A high proportion of the Exdgly molecules secreted into medium were found to be clipped, lacking the first two amino acids (His–Gly) from the N-terminus. Disruption of the P. pastoris homolog of the Saccharomyces cerevisiae dipeptidyl aminopeptidase (STE13) gene in Pichia gen- ome resulted in a clone that expressed N-terminally intact Exdgly. Elimination of N-terminal clipping enhanced the yield and simplified the purification of Exdgly from P. pastoris culture supernatant. Ó 2008 Elsevier Inc. All rights reserved. Introduction Exendin-4 is a peptide hormone found in saliva of the Gila mon- ster Heloderma suspectum [1]. Its amino acid sequence has 53% identity with that of human glucagon-like peptide-1 (GLP-1) [2], an incretin that has glucoregulatory effects. Like GLP-1, Exendin- 4 also has glucose-lowering effects and at least some of these bio- logical activities can be attributed to its ability to bind and activate the GLP-1 receptor [2]. Exendin-4, however, scores over GLP-1 in that it has much longer biological half-life [3]. This stability is attributed to its resistance to DPP-IV, a circulating dipeptidyl ami- nopeptidase responsible for the degradation of GLP-1 in vivo. Exendin-4 enhances secretion of insulin [4] and suppresses inappropriate glucagon release [5], both in a glucose-dependent fashion. It also improves peripheral sensitivity to insulin [6], slows gastric emptying [5] and reduces food intake [6]. Perhaps, the most interesting observation is that Exendin-4 can improve pancreatic b-cell health by suppression of b-cell apoptosis and stimulation of b-cell neogenesis and proliferation [7]. This raises the possibility that progression of diabetes can be checked, perhaps even re- versed, by the use of Exendin-4 [8]. There has consequently been a lot of interest in this molecule for treatment of diabetes. Exena- tide (synthetic Exendin-4) has already been approved as a drug and is available under the trade name Byetta Ò . Although Exen- din-4 has been expressed in Escherichia coli as a GST fusion protein, the yield appears to be low (3.15 mg/10 g bacteria) [9]. We describe here the expression of Exendin-4 precursor, Exdgly, (with an extra glycine at the Carboxy terminus) in the methylotropic yeast, Pichia pastoris. Analysis of the amino acid sequence of Exdgly by NetOGlyc server (http://www.cbs.dtu.dk/services/NetOGlyc/) indicated five potential O-linked glycosylation sites at T5, T7, S32, S33 and S39. However, Exdgly was secreted into the medium non- glycosylated, but with the deletion of first two amino acids (His– Gly) from the N-terminus of >50% of the molecules. Exendin-4 lack- ing first two amino acids from N-terminus is known to function as an antagonist as opposed to agonistic action by full length molecule [10]. We disrupted two dipeptidyl aminopeptidase genes in Pichia pastoris to determine if this could prevent clipping. Inactivation of the Pichia homolog of S.cerevisiae STE13 gene abolished N-terminal clipping eliminating impurities and improving the yield of Exen- din-4. STE13 is also known as dipeptidyl aminopeptidase A (DPAP A or YSCIV peptidase) is a single-pass type II vacuole membrane serine protease Among 13 substrates studied so far, the second amino acid at N-terminal end was found to be either proline (in 7 cases) or alanine (6 cases) (http://merops.sanger.ac.uk/). Thus the consen- sus substrate cleavage site for Saccharomyces STE13 is Xaa-Pro/ Ala. The main function of STE13 identified so far is in maturation of a-factor precursor and kexin precursor. Materials and methods Strains, plasmids and media The Pichia pastoris strain GS115 (his4) was used for expression of Exendin-4. The expression vector used was pPIC9K (Invitrogen, 1046-5928/$ - see front matter Ó 2008 Elsevier Inc. All rights reserved. doi:10.1016/j.pep.2008.10.021 * Corresponding author. Fax: +91 80 2808 2303. E-mail address: [email protected] (K. Sastry). Protein Expression and Purification 64 (2009) 155–161 Contents lists available at ScienceDirect Protein Expression and Purification journal homepage: www.elsevier.com/locate/yprep
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Identification of the dipeptidyl aminopeptidase responsible for N-terminal clipping of recombinant...

Protein Expression and Purification 64 (2009) 155–161
Contents lists available at ScienceDirect
Protein Expression and Purification
journal homepage: www.elsevier .com/locate /yprep
Identification of the dipeptidyl aminopeptidase responsible for N-terminalclipping of recombinant Exendin-4 precursor expressed in Pichia pastoris
Lakshmi Prabha, Nagaraj Govindappa, Laxmi Adhikary, Ramakrishnan Melarkode, Kedarnath Sastry *
Biocon Limited, Research and Development, 20th KM, Hosur Road, Electronic City P.O., Bangalore 560 100, India
a r t i c l e i n f o a b s t r a c t
Article history:Received 5 September 2008and in revised form 30 October 2008Available online 8 November 2008
Keywords:Dipeptidyl aminopeptidasePichia pastorisSaccharomyces cerevisiaeSTE13DAP2Exendin-4
1046-5928/$ - see front matter � 2008 Elsevier Inc. Adoi:10.1016/j.pep.2008.10.021
* Corresponding author. Fax: +91 80 2808 2303.E-mail address: [email protected] (K.
Exendin-4 is a naturally occurring 39 amino acid peptide that is useful for the control of Type 2 diabetes.Recombinant Exendin-4, with an extra glycine at the carboxy-terminus (Exdgly), was expressed in themethylotropic yeast Pichia pastoris. A high proportion of the Exdgly molecules secreted into medium werefound to be clipped, lacking the first two amino acids (His–Gly) from the N-terminus. Disruption of the P.pastoris homolog of the Saccharomyces cerevisiae dipeptidyl aminopeptidase (STE13) gene in Pichia gen-ome resulted in a clone that expressed N-terminally intact Exdgly. Elimination of N-terminal clippingenhanced the yield and simplified the purification of Exdgly from P. pastoris culture supernatant.
� 2008 Elsevier Inc. All rights reserved.
Introduction We describe here the expression of Exendin-4 precursor, Exdgly,
Exendin-4 is a peptide hormone found in saliva of the Gila mon-ster Heloderma suspectum [1]. Its amino acid sequence has 53%identity with that of human glucagon-like peptide-1 (GLP-1) [2],an incretin that has glucoregulatory effects. Like GLP-1, Exendin-4 also has glucose-lowering effects and at least some of these bio-logical activities can be attributed to its ability to bind and activatethe GLP-1 receptor [2]. Exendin-4, however, scores over GLP-1 inthat it has much longer biological half-life [3]. This stability isattributed to its resistance to DPP-IV, a circulating dipeptidyl ami-nopeptidase responsible for the degradation of GLP-1 in vivo.
Exendin-4 enhances secretion of insulin [4] and suppressesinappropriate glucagon release [5], both in a glucose-dependentfashion. It also improves peripheral sensitivity to insulin [6], slowsgastric emptying [5] and reduces food intake [6]. Perhaps, the mostinteresting observation is that Exendin-4 can improve pancreaticb-cell health by suppression of b-cell apoptosis and stimulationof b-cell neogenesis and proliferation [7]. This raises the possibilitythat progression of diabetes can be checked, perhaps even re-versed, by the use of Exendin-4 [8]. There has consequently beena lot of interest in this molecule for treatment of diabetes. Exena-tide (synthetic Exendin-4) has already been approved as a drugand is available under the trade name Byetta�. Although Exen-din-4 has been expressed in Escherichia coli as a GST fusion protein,the yield appears to be low (3.15 mg/10 g bacteria) [9].
ll rights reserved.
Sastry).
(with an extra glycine at the Carboxy terminus) in the methylotropicyeast, Pichia pastoris. Analysis of the amino acid sequence of Exdglyby NetOGlyc server (http://www.cbs.dtu.dk/services/NetOGlyc/)indicated five potential O-linked glycosylation sites at T5, T7, S32,S33 and S39. However, Exdgly was secreted into the medium non-glycosylated, but with the deletion of first two amino acids (His–Gly) from the N-terminus of >50% of the molecules. Exendin-4 lack-ing first two amino acids from N-terminus is known to function as anantagonist as opposed to agonistic action by full length molecule[10]. We disrupted two dipeptidyl aminopeptidase genes in Pichiapastoris to determine if this could prevent clipping. Inactivation ofthe Pichia homolog of S.cerevisiae STE13 gene abolished N-terminalclipping eliminating impurities and improving the yield of Exen-din-4.
STE13 is also known as dipeptidyl aminopeptidase A (DPAP A orYSCIV peptidase) is a single-pass type II vacuole membrane serineprotease Among 13 substrates studied so far, the second aminoacid at N-terminal end was found to be either proline (in 7 cases)or alanine (6 cases) (http://merops.sanger.ac.uk/). Thus the consen-sus substrate cleavage site for Saccharomyces STE13 is Xaa-Pro/Ala. The main function of STE13 identified so far is in maturationof a-factor precursor and kexin precursor.
Materials and methods
Strains, plasmids and media
The Pichia pastoris strain GS115 (his4) was used for expressionof Exendin-4. The expression vector used was pPIC9K (Invitrogen,

156 L. Prabha et al. / Protein Expression and Purification 64 (2009) 155–161
USA) that provides the promoter and terminator sequences ofAOX1 gene and the Saccharomyces cerevisiae Mat-a signal sequencefor secretion. The E. coli strain DH5a was used for routine cloningand propagation of plasmids.
Yeast extract–peptone–dextrose (YPD) medium containing10 g/L yeast extract, 20 g/L peptone, and 20 g/L dextrose was usedfor routine growth and subculturing of Pichia strains. YNBD med-ium used for selection contained Minimal dextrose medium with13.4 g/L yeast nitrogen base without amino acids, 20 g/L dextrose.Luria broth/agar was used for culturing E. coli. Media componentsused were either from Himedia (Mumbai, India) or Difco (USA).
Synthesis of Exdgly coding sequence
The Exdgly coding sequence was optimized for expression in P.pastoris employing codons that are most frequently used in theorganism. The gene was constructed by PCR from overlappingprimers that span the length of the sequence on both strands. Eachprimer was first individually phosphorylated. The primers werethen annealed together and ligated. The desired fragment wasamplified from the ligation mix and cloned into pTZ57R vector(Fermentas) for sequencing (MWG, Bangalore). After the sequenceintegrity was verified, the synthetic Exdgly coding sequence wassubcloned into the expression vector pPIC9K at XhoI/EcoRI sitesto give Exdgly/ pPIC9K. This cloning step fused the Exdgly codingsequence to the Mata signal sequence for secretion and placed itunder the control of AOX1 promoter and terminator.
Transformation of Pichia pastoris
The construct, Exdgly/pPIC9K was transformed by electropora-tion into P. pastoris using his marker for selection. Transformationwas done by electroporation of freshly grown cells in 0.2 cm cuv-ettes. The pulse was delivered by Gene Pulser (BioRad) at 1500 V,25 lF, and 200 X. The electroporated cells were allowed to recoverfor 1 h in 1 M sorbitol at 30 �C and then plated on YNBD agar.Transformants obtained were screened for integration of multiplecopies of Exendin-4 by their resistance to successively higher con-centrations of G418 and clones chosen on this basis were taken upfor expression. Standard protocols recommended by the manufac-turer (Invitrogen) were followed for screening and expression.
STE13 gene disruption
The STE13 targeting fragment was assembled in two PCR stepsto introduce a unique NdeI site within the region of homology.Restriction sites were introduced at the 50 and 30 ends of the frag-ments to facilitate cloning. In the first PCR two short fragments of�200 bp were amplified from the genomic DNA of Pichia pastorisGS115 strain using the following primers (Table 1): IISTEFP1/IIS-TERP1 and IISTEFP2/IISTERP2. The second PCR step was an overlapPCR, with the two amplified fragments as template to generate the450 bp fragment using IISTEFP1 and IISTERP2 primers. PCR reac-tions were carried out using XT Taq polymerase (Bangalore Genei,India). The 450 bp fragment was first cloned into pTZ57R vector(Fermantas) and its identity was confirmed by sequencing. It wasthen subcloned into pPICZA (Invitrogen) at the BamHI/BglII sites
Table 1Primers used for amplification of Pichia pastoris STE13 gene targeting fragment.
Primer Sequence
IISTEFP1 50 GGA TCC GCA GTT CAA CAT TGG GTT TGA GCA 30
IISTERP1 50 CCA GAA TCA TAT GCC AAT GTC TTA AGC G 30
IISTEFP2 50 CGC TTA AGA CAT TGG CAT ATG ATT CTG G 30
IISTERP2 50 AGA TCT GTC CCG TGA CAA ACC AAG AAT CGG 30
to get Stefrg/Zeo. After linearization of Stefrg/Zeo by NdeI, the con-struct was transformed into the P. pastoris strain producing Exdgly.Transformation was done by electroporation as described above,except that after allowing cells to recover in 1 M sorbitol, one vol-ume of YPD medium was added, and the cells were grown for 1 h at30 �C. Cells were then plated on YPD medium supplemented with1 M sorbitol and 100 lg/ml Zeocin (Invitrogen). The insertion ofthe disruption vector into Pichia genome into STE13 locus was ver-ified by Southern blotting using genomic DNA of GS115 (parent)and STE13 disrupted clone by using a non-radioactive DIG HighPrime DNA Labelling and Detection Starter kit, following manufac-turers instructions (Roche Applied Science, Germany).
Plate assay
The assay has been described previously for Saccharomyces cere-visiae [11]. Briefly, the colonies were first lysed by flooding theplates with chloroform. After the chloroform evaporated com-pletely, a mixture of the substrate, Ala–Pro–4MbNA and Fast gar-net GBC in 1%Tris agar was poured to form an overlay. Theoverlaid plates were incubated at room temperature for about10–15 min. Colonies that did not stain red were picked up as po-tential STE13 disruptants. Pichia pastoris GS115 strain with intactSTE13 gene was included as a control to check the progress of stain-ing. All colonies eventually stain red when left for an extended per-iod of time. This is because DAP2p, another dipeptidylaminopeptidase also acts on the same substrate.
Induction of the Exdgly expression in P. pastoris
P. pastoris transformant which grew on 2 mg/ml of G418 platewas grown on 10 ml YNB medium (1.34% yeast nitrogen base w/o amino acids and 2% dextrose) overnight. It was sub cultured into50 ml BMGY (1% yeast extract, 2% peptone, 1.34% YNB, 100 mMpotassium phosphate at pH 6.0, and 1% glycerol) with 0.5 OD600.Grown this culture at 30 �C for 48 h with shaking at 220 rpm.Thecells were harvested by centrifugation at 5000 rpm for 5 min atroom temperature and resuspended in induction medium (1%yeast extract, 2% peptone,1.34% YNB and 100 mM potassium phos-phate at pH 6.0). 20% methanol was added to the culture mediumto give a final concentration of 0.5% methanol as an inducer. Sam-ples were collected everyday and analyzed by tricine SDS–PAGE aswell as HPLC.
HPLC assay
HPLC assay for measuring the levels of Exdgly employed a C18column (2.4 � 250; 5 u from Waters) and a gradient run. The mo-bile phase consisted of A, 0.1% TFA (75%) and B, Acetonitrile solvent(25%). The runtime was 25 min where the component B composi-tion rises from 25% to 40% in 15 min at a flow rate of 1.2 ml/min.Ten microliter of sample was injected into the column maintainedat 40 �C and the sample was monitored at 220 nm. Mass character-ization of Exdgly was carried out in Q-Star-Excel (ABI-Sciex) withMALDI-front end. Exdgly sample was digested with Trypsin in20:1 (protein to enzyme) ratio, in Tris buffer at pH 8.2–8.5. Thesample was incubated at 37 �C for 3 h and analyzed in MALDI-TOF using Sinapic acid as matrix.
Results
Expression of Glycine extended Exendin-4
The Exdgly coding nucleic acid sequence optimized for expres-sion in Pichia was cloned into the expression vector pPIC9K and

L. Prabha et al. / Protein Expression and Purification 64 (2009) 155–161 157
transformed into the P. pastoris strain GS115. The clones growingat 0.5 mg/ml of G418 were stamped on plates with 1 and 2 mg/ml of G418. Colonies which grew on 2 mg/ml of G418 were se-lected for further study after four subcultures to eliminate unstableintegrants. Following these subcultures, clones were checked forexpression of Exdgly upon induction with methanol. Cell-freesupernatants of induced cultures when analyzed by RP-HPLCshowed the presence of two Exdgly-related species (Fig. 1A). Char-acterization of these by LC-MS indicated that Exdgly was being de-graded at its N-terminus—the two peaks corresponded to intactExdgly and Exdgly with two aminoacids clipped at its N-terminus([N-2] Exdgly) (Fig. 1B). Small levels of other forms that seemed tohave arisen from successive clipping of two aminoacids from N-terminus were also observed with the progress of fermentation(data not shown). This suggested the activity of a dipeptidyl ami-nopeptidase that has reasonable but not a strong affinity for Exdglyas substrate could be responsible for clipping. A bioinformatics ap-proach was taken to determine the Pichia homologs of Saccahromy-ces cerevisiae dipeptidyl peptidase STE13. The STE13 sequence wasused to search the translated database of commercially availablePichia pastoris genome (Integrated Genomics, Chicago). This searchrevealed two Pichia ORFs with homology to STE13 (see Fig. 2 andTable 2).
S. cerevisiae STE13 and DAP2 show 33% identity and 53% similar-ity to each other while Pichia pastoris homologs RPPA08049 andRPPA10441 show 39% identity and 57% similarity to each other.Based on the percentage of identity it was concluded that ORFRPPA08049 (contig number IG-66) is putative STE13 and ORFRPPA10441 (contig IG-66) is most likely to be DAP2. To determineif STE13 or DAP2 are responsible for N-terminal clipping observedin Exdgly, we disrupted these two genes and determined their ef-fect on Exdgly clipping. The S. cerevisiae STE13 and DAP2 gene singlegene knock outs are viable and show no obvious defect in theirphenotypes (www.yeastgenome.org).
Disruption of the S. cerevisiae STE13 homolog in Pichia
Dipeptidyl amino peptidases have not been characterized inPichia. By its homology with Saccharomyces cerevisiae, the PichiaSTE13 protease (STE13p) seemed to be the most likely candidateresponsible for the N-terminal clipping of Exdgly. In Saccharomy-
A.
2 4 6 8
mA
U
0
500
1000
1500
2000
2500
3000
3500
VWD1 A, Wavelength=220 nm (EXENDIN\25050602.D)
B. HG EGTFTSDLSKQMEEEAVRLFIEWLKNGGPSSGAPPPSG
CLEAVGE SITE
Fig. 1. (A) HPLC chromatogram of cell-free supernatant of P. pastoris strain expressing Exof Exdgly, respectively. (B) Amino acid sequence of Exdgly protein with arrow indicatin
ces, STE13p is found in trans-golgi whereas the other two DPPs—DAP2 and DPPIII, are localized to vacuolar or cytoplasmic compart-ment, respectively. As Exdgly is secreted, it is most likely toencounter STE13 protease. The nucleotide sequence for the Pichiahomolog of STE13 and DAP2 genes were obtained from Pichia gen-ome database. The region encoding catalytic domain of PichiaSTE13p was demarcated by homology with the S. cerevisiae STE13pand a fragment of about 450 bp nucleotides was chosen to providehomology for chromosomal gene targeting such that the insertionwould disrupt the functional activity of the catalytic domain. Thetargeting sequences chosen for disruption of Pichia homolog of S.cerevisiae STE13 and DAP2 gene fragments were amplified fromgenome of Pichia pastoris GS115 strain and completely sequencedfor verification. These sequences have been deposited in GenBankunder Accession Nos. EU921448 and EU921449.
STE13 gene was disrupted in the P. pastoris strain expressingExdgly by transformation of the targeted disruption vector, Stef-rg/pPICZA (described in materials and methods). Colonies obtainedon transformation with the linearized plasmid were screened bymeans of a plate assay to screen for STE13 disruptants. Putativesidentified by the assay were verified for correct disruption of thelocus by Southern blotting (Fig. 3) and PCR (not shown). Southernanalysis clearly showed the integration of a single copy of the plas-mid at the STE13 locus without the integration of additional copieselsewhere in the genome. No obvious changes either in growth rateor phenotype could be observed in the STE13 disrupted Pichiastrain as compared to the parent strain.
Exdgly expression in STE13 disruptants
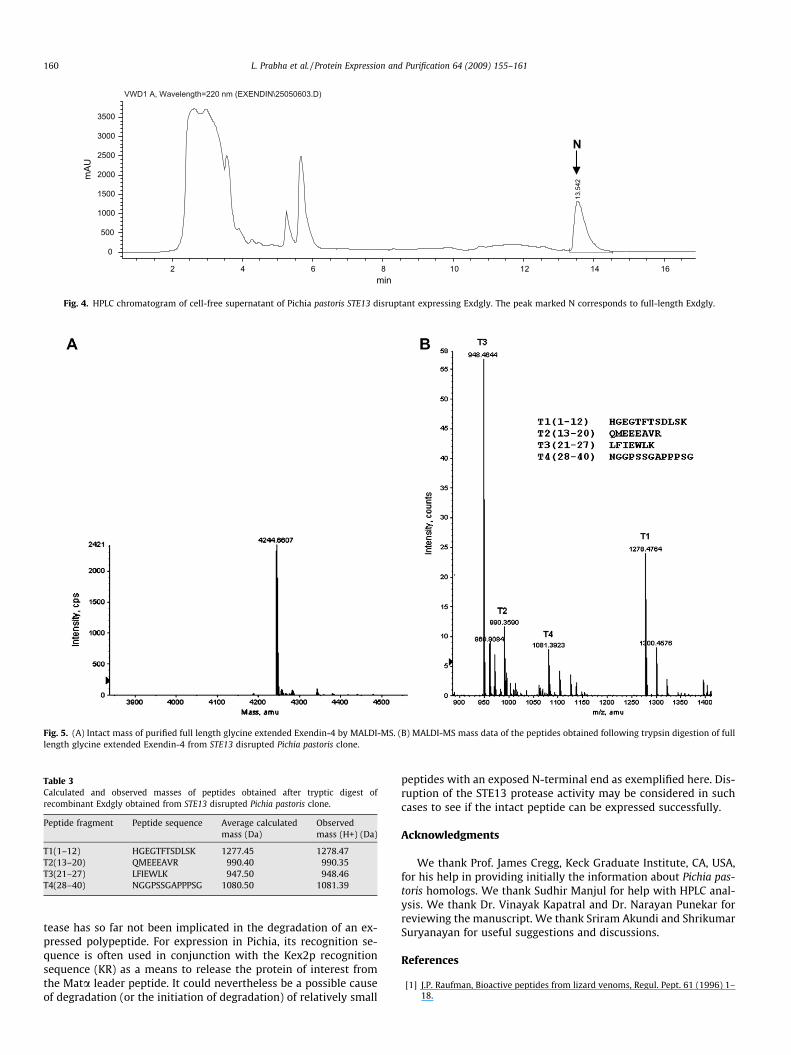
The STE13 disruptant was induced with methanol and the ex-pressed Exdgly was analyzed by HPLC. Only the peak correspond-ing to full-length Exdgly was observed (Fig. 4) indicating that N-terminal degradation had been abolished. This result suggests thatSTE13p is responsible for the N-terminal clipping of Exdgly ob-served in the parent strain GS115. The production level of Exdglyin fermentation broth supernatant increased from 0.3 g/L to0.5 g/L in the STE13 disrupted clone.
The full length Exdgly was purified and completely character-ized by intact mass, peptide mass fingerprinting. The average in-tact molecular mass in MALDI-MS analysis of Exdgly was found
min10 12 14 16
13.
838
14.
547
N
(N-2)
dgly. The peaks marked N and (N�2) correspond to full-length and truncated formsg protease cleavage site.

Fig. 2. Alignment of amino acid sequences of Saccharomyces cerevisiae STE13 and Pichia pastoris homologs.
158 L. Prabha et al. / Protein Expression and Purification 64 (2009) 155–161
to be 4244.66 (C186H284N50O62S; Calculated molecularmass = 4244.62 Da) (Fig. 5A). The peptide mass fingerprinting ofExdgly with trypsin gave rise to four peptide fragments (Fig. 5B)with the masses depicted in Table 3.
Discussion
We expressed glycine-extended Exendin-4 in the Pichia pastorisstrain GS115. Along with the full-length peptide, an additional

Table 2Homology between dipeptidyl peptidases STE13 & DAP2 from Pichia pastoris and S.cerevisiae.
Pichia ORF Homology to S. cerevisiae
STE13 DAP2
Identity(%)
Similarity(%)
Gaps Identity(%)
Similarity(%)
Gaps
RPPA08049 40 57 8 35 53 9RPPA10441 34 52 7 45 62 7
L. Prabha et al. / Protein Expression and Purification 64 (2009) 155–161 159
form with two amino acids missing from the N-terminus was alsoproduced. Previous studies have shown that Exendin is a GLP-1receptor agonist and Exendin lacking two N-terminal amino acids(His–Gly) is an antagonist [10]. Although five amino acids (T5,T7, S32, S33 and S39), which could be potentially O-glycosylatedare found in Exdgly primary amino acid sequence, no O-linked gly-cosylation was observed as revealed by LC-MS characterization of
Fig. 3. Schematic representation of the Pichia pastoris STE13 locus and its disruption. Abbthe catalytic domain; ZeoR, Gene for Zeocin resistance; ori, E. coli origin of replication. (Abeen indicated. (B) The plasmid Stefrg/pPICZA used for disruption. (C) Disrupted STE13 lpastoris STE13 disruptant (Lane 1) and GS115 parent (Lane 2) was digested with restrictishown in (A). The hybridization pattern was as expected—5kb band in GS115 and two b
products from the two observed peaks. There are no consensussites for N-linked glycosylation in Exdgly. As the activity of adipeptidyl aminopeptidase was indicated to be responsible for N-terminal clipping, the Pichia homolog of the S. cerevisiae STE13gene was disrupted by insertional inactivation. The STE13 disrup-tant expressed only the full-length Exdgly showing that STE13pis responsible for the N-terminal clipping of the peptide.
The other dipeptidyl aminopeptidase that Exdgly may be ex-pected to encounter in the secretory pathway is the DAP2 protease.The Pichia homolog of the DAP2 gene was also disrupted, but withno effect seen on the N-terminal degradation of Exdgly (Data notshown).
The STE13 protease is known to cleave on the carboxyl side ofXaa-Ala and Xaa-Pro sequences in Saccharomyces Mata peptide[12]. Exdgly with N-terminal sequence of His–Gly is not expectedto be a substrate for STE13. This work reveals the ability of theputative Pichia homolog of S. cerevisae STE13 aminopeptidase toact on His–Gly sequence, albeit with a lower efficiency. STE13 pro-
reviations used: FR, flanking region; CDS, coding sequence; Cat, sequence encoding) Intact STE13 genomic locus. The fragment used as probe for Southern analysis hasocus. (D) Southern hybridization to verify STE13 disruption. Genomic DNA of Pichiaon enzyme BglII and hybridized with the 450 bp nucleic acid non-radioactive probeands of 3.8 and 3.5 kb in the disruptant.
min2 4 6 8 10 12 14 16
mA
U
0
500
1000
1500
2000
2500
3000
3500
VWD1 A, Wavelength=220 nm (EXENDIN\25050603.D)
13.
542
N
Fig. 4. HPLC chromatogram of cell-free supernatant of Pichia pastoris STE13 disruptant expressing Exdgly. The peak marked N corresponds to full-length Exdgly.
Fig. 5. (A) Intact mass of purified full length glycine extended Exendin-4 by MALDI-MS. (B) MALDI-MS mass data of the peptides obtained following trypsin digestion of fulllength glycine extended Exendin-4 from STE13 disrupted Pichia pastoris clone.
Table 3Calculated and observed masses of peptides obtained after tryptic digest ofrecombinant Exdgly obtained from STE13 disrupted Pichia pastoris clone.
Peptide fragment Peptide sequence Average calculatedmass (Da)
Observedmass (H+) (Da)
T1(1–12) HGEGTFTSDLSK 1277.45 1278.47T2(13–20) QMEEEAVR 990.40 990.35T3(21–27) LFIEWLK 947.50 948.46T4(28–40) NGGPSSGAPPPSG 1080.50 1081.39
160 L. Prabha et al. / Protein Expression and Purification 64 (2009) 155–161
tease has so far not been implicated in the degradation of an ex-pressed polypeptide. For expression in Pichia, its recognition se-quence is often used in conjunction with the Kex2p recognitionsequence (KR) as a means to release the protein of interest fromthe Mata leader peptide. It could nevertheless be a possible causeof degradation (or the initiation of degradation) of relatively small
peptides with an exposed N-terminal end as exemplified here. Dis-ruption of the STE13 protease activity may be considered in suchcases to see if the intact peptide can be expressed successfully.
Acknowledgments
We thank Prof. James Cregg, Keck Graduate Institute, CA, USA,for his help in providing initially the information about Pichia pas-toris homologs. We thank Sudhir Manjul for help with HPLC anal-ysis. We thank Dr. Vinayak Kapatral and Dr. Narayan Punekar forreviewing the manuscript. We thank Sriram Akundi and ShrikumarSuryanayan for useful suggestions and discussions.
References
[1] J.P. Raufman, Bioactive peptides from lizard venoms, Regul. Pept. 61 (1996) 1–18.

L. Prabha et al. / Protein Expression and Purification 64 (2009) 155–161 161
[2] R. Goke, H.C. Fehmann, T. Linn, H. Schmidt, M. Krause, J. Eng, B. Goke, Exendin-4 is a high potency agonist and truncated exendin-(9-39)-amide an antagonistat the glucagon-like peptide 1-(7-36)-amide receptor of insulin-secreting beta-cells, J. Biol. Chem. 268 (1993) 19650–19655.
[3] A.A. Young, B.R. Gedulin, S. Bhavsar, N. Bodkin, C. Jodka, B. Hansen, M. Denaro,Glucose-lowering and insulin-sensitizing actions of exendin-4: studies inobese diabetic (ob/ob, db/db) mice, diabetic fatty Zucker rats, and diabeticrhesus monkeys (Macaca mulatta), Diabetes 48 (1999) 1026–1034.
[4] D.G. Parkes, R. Pittner, C. Jodka, P. Smith, A. Young, Insulinotropic actions ofexendin-4 and glucagon-like peptide-1 in vivo and in vitro, Metabolism 50(2001) 583–589.
[5] O.G. Kolterman, J.B. Buse, M.S. Fineman, E. Gaines, S. Heintz, T.A. Bicsak, K.Taylor, D. Kim, M. Aisporna, Y. Wang, A.D. Baron, Synthetic exendin-4(exenatide) significantly reduces postprandial and fasting plasma glucose insubjects with type 2 diabetes, J. Clin. Endocrinol. Metab. 88 (2003) 3082–3089.
[6] B.R. Gedulin, S.E. Nikoulina, P.A. Smith, G. Gedulin, L.L. Nielsen, A.D. Baron, D.Parkes, A.A. Young, Exenatide (exendin-4) improves insulin sensitivity and b-cell mass in insulin-resistant obese fa/fa Zucker rats independent of glycemiaand body weight, Endocrinology 146 (2005) 2069–2076.
[7] G. Xu, D.A. Stoffers, J.F. Habener, S. Bonner-Weir, Exendin-4 stimulates bothbeta-cell replication and neogenesis, resulting in increased beta-cell massand improved glucose tolerance in diabetic rats, Diabetes 48 (1999) 2270–2276.
[8] L.L. Nielsen, A.A. Young, D.G. Parkes, Pharmacology of exenatide (syntheticexendin-4): a potential therapeutic for improved glycemic control of type 2diabetes, Regul. Pept. 117 (2004) 77–88.
[9] X. Yin, D. Wei, L. Yi, X. Tao, Y. Ma, Expression and purification of exendin-4, aGLP-1 receptor agonist, in Escherichia coli, Protein Expr. Purif. 41 (2005) 259–265.
[10] C. Montrose-Rafizadeh, H. Yang, B.D. Rodgers, A. Beday, L.A. Pritchette, J. Eng,High potency antagonists of the pancreatic glucagon-like peptide-1 receptor, J.Biol. Chem. 272 (1997) 21201–21206.
[11] P. Suarez Rendueles, D.H. Wolf, Identification of the structural gene fordipeptidyl aminopeptidase yscV (DAP2) of Saccharomyces cerevisiae, J.Bacteriol. 169 (1987) 4041–4048.
[12] D. Julius, L. Blair, A. Brake, G. Sprague, J. Thorner, Yeast alpha factor isprocessed from a larger precursor polypeptide: the essential role of amembrane-bound dipeptidyl aminopeptidase, Cell 32 (1983) 839–852.




















