
Dipeptidyl-peptidase IV-beta. Further characterization and comparison to dipeptidyl-peptidase IV...
10
Eur. J. Biochem. 256, 3692378 (1998) FEBS 1998 Dipeptidyl-peptidase IV-β Further characterization and comparison to dipeptidyl-peptidase IV activity of CD26 Julia ` BLANCO 1 , Coralie NGUYEN 2 , Christian CALLEBAUT 1 , Etienne JACOTOT 1 , Bernard KRUST 1 , Jean-Paul MAZALEYRAT 2 , Michel WAKSELMAN 2 and Ara G. HOVANESSIAN 1 1 Unite ´ Virologie et Immunologie Cellulaire, ERS 572 CNRS, Departement SIDA/Retrovirus, Institut Pasteur, Paris, France 2 SIRCOB, Universite ´ de Versailles, Ba ˆtiment Lavoisier, Versailles, France (Received 27 April/22 June 1998) 2 EJB 98 0566/4 Dipeptidyl peptidase IV-β (DPP IV-β) is a novel protein which shows a peptidase activity similar to the T-cell-activation antigen CD26. To further characterize this DPP IV-β and confirm its cell surface expression, we have developed a purification strategy using the CD26 2 cell line C8166. The purification process includes biotinylation of cell surface proteins before preparation of cell extracts and processing by gel-filtration, ion-exchange and lectin chromatographies. Consistent with the molecular mass of DPP IV-β estimated by gel-filtration chromatography, the final purified fraction, manifesting a typical DPP IV activity, showed a major biotinylated 75280-kDa band in SDS/PAGE, thus suggesting the monomeric nature of this enzyme. Kinetic parameters of DPP IV-β and the sensitivity to a new family of irreversible DPP IV inhibitors, were studied in comparison to CD26. Both enzymes followed a Michaelis kinetics with different K m values for Gly-Pro-NH-Np (NH-Np, para-nitroanilide) hydrolysis (0.28 6 0.05 mM and 0.12 60.02 mM). More significant differences were observed in the sensitivity to inhibitors, which exerted a much higher activity on CD26 than on DPP IV-β. These differences permitted us to study DPP IV-β expression in CD26-expressing cells, showing the expression of this new enzyme in all lymphoid cells tested, and a rapid enhancement in phytohemagglutinin-stimulated or protein-A-stimulated peripheral blood mononuclear cells. Our results indicate that, although DPP IV-β and CD26 are coexpressed and manifest a typical DPP IV activity, there are distinct features in their catalytic activities that may confer to each enzyme a complementary role in peptide processing. Keywords: CD26; dipeptidyl peptidase IV; human peripheral blood mononuclear cells; HIV. The dipeptidyl peptidas IV (DPP IV) activity belongs to the family of dipeptidyl aminopeptidases (DPP), which includes four activities, associated, at least, to four different serine prote- ases [1], all characterized by their ability to release N-terminal dipeptides from a variety of small proteins. DPP IV cleaves pref- erentially substrates containing the sequence X-P- at the amino terminus, provided the third residue is neither proline or hydro- xyproline. Thus, DPP IV can also be classified as a postproline- cleaving enzyme, which represents an important family of regu- Correspondence to A. G. Hovanessian, Unite ´ Virologie et Immuno- logie Ce ´llulaire, Departe ´ment SIDA/Retrovirus, Institut Pasteur, 28 rue Dr Roux, F-75724 Paris Cedex 15, France Fax: 133 1 40 61 30 12. E-mail : [email protected] URL: http://www.pasteur.fr/units/vicel Abbreviations. DPP IV, dipeptidyl peptidase IV; CD26, an 110-kDa protein referred to as T-cell-activation antigen or as DPP IV; DPP IV-β, an 82-kDa protein with DPP IV activity; DPP T-L, an 175-kDa protein with DPP IV activity ; PhMeSO 2 F, phenylmethylsulfonyl fluoride; NH- Np, para-nitroanilide ; PHA, phytohemagglutinin ; WGA, wheat germ agglutinin ; Z(NO 2 ), 4-nitrobenzyloxycarbonyl; Pefabloc-SC, 4-(2-ami- noethyl)-benzenesulfonyl fluoride ; Aba, 3-aminobenzoic acid ; cyclo- [ ε Lys-Pro-Aba-(6-CH 2 S 1 Me 2 )-Gly-Gly-] 2 CF 3 COO 2 , ICI-11a; PBMC, peripheral blood mononuclear cells ; ADA, adenosine deaminase ; IC 50 , concentration causing 50% inhibition of activity. Enzymes. DPP IV (EC 3.4.14.5); DPP (EC 3.4.14.2); ADA (EC 3.5.4.4); proline endopeptidase (EC 3.4.21.26); alanine aminopeptidase (EC 3.4.11.2) ; proline aminopeptidase (EC 3.4.11.5) ; arginine amino- peptidase (EC 3.4.11.6). latory proteases [2]. In addition, among dipeptidyl peptidase activities, DPP IV is the only activity known to be located on the outer side of the plasma membrane, thus acting as an ectoen- zyme [3]. In man, DPP IV activity has been associated to a 1052110- kDa cell surface protein, known to be identical to the adenosine- deaminase-binding protein, and to the T-cell-activation antigen CD26 [426]. CD26/DPP IV is constitutively expressed on many cell types, showing high levels of expression in brush-border epithelia from placenta, intestine and kidney [7], where it has been implicated in peptide absorption [8]. In contrast, its expres- sion is highly regulated in peripheral blood T lymphocytes, in which CD26 expression is enhanced upon T-cell activation [9]. In these cells, CD26 is able to deliver intracellular signals, prob- ably by means of its physical association with the membrane- linked tyrosine phosphatase CD45 [10]. Consistently, modula- tion of CD26 with some anti-CD26 mAbs or with its natural ligand, adenosine deaminase (ADA) leads to a comitogenic ef- fect on anti-CD3 T-cell activation [11, 12]. CD26 has been also reported to play two controversed roles in HIV infection. CD26 positively modulates both viral entry into permissive cells and its cytopathic effect [13215]. Although this latter effect seems to be independent of DPP IV activity [16], it may be related to the ability of the envelope protein of HIV to inhibit the binding of ADA to CD26 [17]. In contrast, the weak response of lym- phocytes from HIV infected individuals to recall antigens has been proposed to be the consequence of the potential interaction of HIV-1 Tat with CD26 that might lead to the inhibition of
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Dipeptidyl-peptidase IV-beta. Further characterization and comparison to dipeptidyl-peptidase IV...

Eur. J. Biochem.256, 3692378 (1998) FEBS1998
Dipeptidyl-peptidase IV-βFurther characterization and comparison to dipeptidyl-peptidase IV activity of CD26
Julia BLANCO 1, Coralie NGUYEN2, Christian CALLEBAUT1, Etienne JACOTOT1, Bernard KRUST1, Jean-Paul MAZALEYRAT2,Michel WAKSELMAN 2 and Ara G. HOVANESSIAN1
1 Unite Virologie et Immunologie Cellulaire, ERS 572 CNRS, Departement SIDA/Retrovirus, Institut Pasteur, Paris, France2 SIRCOB, Universite´ de Versailles, Baˆtiment Lavoisier, Versailles, France
(Received 27 April/22 June1998) 2 EJB 98 0566/4
Dipeptidyl peptidase IV-β (DPP IV-β) is a novel protein which shows a peptidase activity similar tothe T-cell-activation antigen CD26. To further characterize this DPP IV-β and confirm its cell surfaceexpression, we have developed a purification strategy using the CD262 cell line C8166. The purificationprocess includes biotinylation of cell surface proteins before preparation of cell extracts and processingby gel-filtration, ion-exchange and lectin chromatographies. Consistent with the molecular mass of DPPIV-β estimated by gel-filtration chromatography, the final purified fraction, manifesting a typical DPP IVactivity, showed a major biotinylated 75280-kDa band in SDS/PAGE, thus suggesting the monomericnature of this enzyme. Kinetic parameters of DPP IV-β and the sensitivity to a new family of irreversibleDPP IV inhibitors, were studied in comparison to CD26. Both enzymes followed a Michaelis kineticswith differentKm values for Gly-Pro-NH-Np (NH-Np,para-nitroanilide) hydrolysis (0.2860.05 mM and0.1260.02 mM). More significant differences were observed in the sensitivity to inhibitors, which exerteda much higher activity on CD26 than on DPP IV-β. These differences permitted us to study DPP IV-βexpression in CD26-expressing cells, showing the expression of this new enzyme in all lymphoid cellstested, and a rapid enhancement in phytohemagglutinin-stimulated or protein-A-stimulated peripheralblood mononuclear cells. Our results indicate that, although DPP IV-β and CD26 are coexpressed andmanifest a typical DPP IV activity, there are distinct features in their catalytic activities that may conferto each enzyme a complementary role in peptide processing.
Keywords:CD26; dipeptidyl peptidase IV; human peripheral blood mononuclear cells; HIV.
The dipeptidyl peptidas IV (DPP IV) activity belongs to thefamily of dipeptidyl aminopeptidases (DPP), which includesfour activities, associated, at least, to four different serine prote-ases [1], all characterized by their ability to release N-terminaldipeptides from a variety of small proteins. DPP IV cleaves pref-erentially substrates containing the sequence X-P- at the aminoterminus, provided the third residue is neither proline or hydro-xyproline. Thus, DPP IV can also be classified as a postproline-cleaving enzyme, which represents an important family of regu-
Correspondence toA. G. Hovanessian, Unite´ Virologie et Immuno-logie Cellulaire, Departe´ment SIDA/Retrovirus, Institut Pasteur, 28 rueDr Roux, F-75724 Paris Cedex15, France
Fax: 133 1 40 61 30 12.E-mail : [email protected]: http://www.pasteur.fr/units/vicelAbbreviations.DPP IV, dipeptidyl peptidase IV; CD26, an110-kDa
protein referred to as T-cell-activation antigen or as DPP IV; DPP IV-β,an 82-kDa protein with DPP IV activity; DPP T-L, an175-kDa proteinwith DPP IV activity ; PhMeSO2F, phenylmethylsulfonyl fluoride ; NH-Np, para-nitroanilide; PHA, phytohemagglutinin; WGA, wheat germagglutinin ; Z(NO2), 4-nitrobenzyloxycarbonyl; Pefabloc-SC, 4-(2-ami-noethyl)-benzenesulfonyl fluoride ; Aba, 3-aminobenzoic acid; cyclo-[ εLys-Pro-Aba-(6-CH2S1Me2)-Gly-Gly-] 2 CF3COO2, ICI-11a; PBMC,peripheral blood mononuclear cells ; ADA, adenosine deaminase ; IC50,concentration causing 50% inhibition of activity.
Enzymes.DPP IV (EC 3.4.14.5); DPP (EC 3.4.14.2) ; ADA (EC3.5.4.4); proline endopeptidase (EC 3.4.21.26); alanine aminopeptidase(EC 3.4.11.2) ; proline aminopeptidase (EC 3.4.11.5) ; arginine amino-peptidase (EC 3.4.11.6).
latory proteases [2]. In addition, among dipeptidyl peptidaseactivities, DPP IV is the only activity known to be located onthe outer side of the plasma membrane, thus acting as an ectoen-zyme [3].
In man, DPP IV activity has been associated to a1052110-kDa cell surface protein, known to be identical to the adenosine-deaminase-binding protein, and to the T-cell-activation antigenCD26 [426]. CD26/DPP IV is constitutively expressed on manycell types, showing high levels of expression in brush-borderepithelia from placenta, intestine and kidney [7], where it hasbeen implicated in peptide absorption [8]. In contrast, its expres-sion is highly regulated in peripheral blood T lymphocytes, inwhich CD26 expression is enhanced upon T-cell activation [9].In these cells, CD26 is able to deliver intracellular signals, prob-ably by means of its physical association with the membrane-linked tyrosine phosphatase CD45 [10]. Consistently, modula-tion of CD26 with some anti-CD26 mAbs or with its naturalligand, adenosine deaminase (ADA) leads to a comitogenic ef-fect on anti-CD3 T-cell activation [11, 12]. CD26 has been alsoreported to play two controversed roles in HIV infection. CD26positively modulates both viral entry into permissive cells andits cytopathic effect [13215]. Although this latter effect seemsto be independent of DPP IV activity [16], it may be related tothe ability of the envelope protein of HIV to inhibit the bindingof ADA to CD26 [17]. In contrast, the weak response of lym-phocytes from HIV infected individuals to recall antigens hasbeen proposed to be the consequence of the potential interactionof HIV-1 Tat with CD26 that might lead to the inhibition of

370 Blanco et al. (Eur. J. Biochem. 256)
its DPP IV activity [18]. However, the significance of such aninteraction under physiological conditions is unclear [19]. Morerecently, Oravecz et al. [20] have described that DPP IV activityof CD26 cleaves RANTES, a chemokine that inhibits HIV infec-tion by blocking HIV interaction with the chemokine receptorCCR5 [21].
The role of DPP IV activity of CD26 in T-cell activationevents, has also been the source of controversy. Some authorshave described that small synthetic inhibitors of DPP IV impairlectin and antigen stimulation of peripheral blood mononuclearcells (PBMC) and other lymphocyte cell types [22]. A possiblemechanism for this effect might be the inhibition of thehyperphosphorylation of the tyrosine kinase p56lck, a substrateof CD45, induced by DPP IV inhibitors [23], but the high con-centrations of inhibitor required make such results questionable.The use of cell clones expressing a mutated, catalytically in-active, form of CD26 also gave contradictory results in two dif-ferent cell lines, Jurkat [24] and BW2 [25]. The failure to un-ravel the role of DPP IV activity is, in part, due to the fact thatno immunologically active substrates have been clearly iden-tified. Recently, evidence was provided for the regulatory roleof DPP IV activity of CD26 in chemokine function [20], how-ever the effect of these chemokines on T-cell proliferation is notwell understood [26]. In addition to CD26, two novel proteinsbearing DPP IV activity have been recently described. Duke-Cohan and co-workers [27, 28] have reported a175-kDa mem-brane bound and soluble form of DPP IV, which is expressed bydifferent epithelial and lymphoblastoid cell lines, and which canbe secreted from activated peripheral blood T cells (referred toas DPP T-L). Our laboratory has recently reported the existenceof a CD26-like 82-kDa protein expressed by the lymphocyticcell line C8166 [29], referred to as DPP IV-β. Both activitiesshow the same substrate specificity and optimal pH as the DPPIV activity of CD26.
The aim of this work was to further characterize this newDPP IV-β activity, and to differentiate it from CD26. We showthat DPP IV-β is a glycoprotein expressed on the cell surface,and demonstrate some catalytic differences in comparison toDPP IV activity of CD26; these differences enabled us to studyDPP IV-β expression on different CD26 expressing cells linesand stimulated PBMC.
MATERIALS AND METHODS
Materials. Peptidase substrates Gly-Pro-, Ala-Ala-, Gly-Arg-, Arg-NH-Np (-NH-Np; p-nitroanilide) were purchasedfrom Sigma, whereas Ala-Pro-, Ala-, Pro- and succinyl-Ala-Pro-NH-Np were generously provided by A.G.M.V. (Halle, Ger-many). 4-(2-Aminoethyl)benzenesulfonyl fluoride (Pefabloc-SC) was from Interchim. Phenylmethylsulfonyl fluoride, (PhMe-SO2F), aprotinin, diprotin A, andN-acetyl-D-glucosamine werefrom Sigma.
Cells and preparation of extracts.The lymphoblastoid cellline C8166 (from Dr G. Farrar) and the clone M8166 from thesecells (from Dr P. Clapham), were generously provided by theMRC AIDS Directed Programme Reagent Project, UK. MOLT4,Jurkat (JBK) and CEM cells are routinely used in our laboratory[29]. CEM clones expressing different levels of CD26 obtainedby transfection, were prepared in our laboratory; the cloneCEM/SH (also referred to as CEM H01 cells) expressing veryhigh levels of CD26 was used as a source of CD26 [30]. Allcells were cultured in suspension medium RPMI-1640 (Bio-Whittaker) supplemented with heat-inactivated (56°C, 30 min)10% (by vol.) fetal calf serum and 2 mM glutamine. HumanPBMC from healthy donors were isolated on a Ficoll-Hypaque
gradient (Pharmacia, Biotechnology) as described [31] and cul-tured in supplemented RPMI1640 medium containing either10 µg/ml phytohemagglutinin (PHA, Difco) or10 µg/ml proteinA (Sigma).
For the preparation of cell extracts, cells were collected 24hours after passaging and washed extensively in137 mM NaCl,6.5 mM sodium phosphate, (NaCl/Pi) before lysis in 20 mMTris/HCl, pH 7.6,150 mM NaCl, 5 mM MgCl2, 0.2 mM PhMe-SO2F, 5 mM 2-mercaptoethanol, aprotinin (100 U/ml) and 0.5%Triton X-100 (75µl/107 cells) (buffer E) and the nuclei werepelleted by centrifugation (350g for 8 min). The supernatantwas diluted with one volume 20 mM Tris/HCl, pH 7.6, 400 mMNaCl, 50 mM KCl,1mM EDTA, 0.2 mM PhMeSO2F, aprotinin(100 U/ml), 5 mM 2-mercaptoethanol,1% Triton X-100, and20% glycerol (buffer I), and centrifuged at12000g for 10 min.The supernatants (cell extracts) were stored at280°C.
Biotinylation and detection of cell surface proteins.Cellsurface proteins were biotinylated as described [32]. Briefly,cells were washed twice in NaCl/Pi, resuspended in ice-coldNaCl/Pi, pH 7.6 (108 cells/ml), and incubated for1 hour at 4°Cwith 10 mM immunopure Sulfo-NHS-biotin (Pierce). Incubationwas stopped by washing cells twice in ice-cold NaCl/Pi contain-ing 10 mM glycine. Extracts from biotinylated cells were pre-pared as described above. Detection of biotinylated proteins wascarried out after electrophoresis of samples in10% acrylamideslab gels, according to the method of Laemmli [33]. Proteinswere then electroblotted onto a poly(vinylidene difluoride)membrane and revealed by using horseradish peroxidase-cou-pled streptavidin and chemiluminescent detection (ECL, Amers-ham).
Inhibitors of DPP IV activity. Irreversible cyclic inhibitorsof DPP IV activity were synthesized as described elsewhere[34]. Briefly, taking into account the substrate specificity of DPPIV for P22P1 . 2P′1 cleavage, we designed and synthesizedcyclopeptides cyclo[εLys-Pro-Aba-(6-CH2S1R2)-Gly-Gly] 2CF3COO2 (Aba5 3-aminobenzoic acid, R5 alkyl), possessinga proline residue at the P1 position, and a lysine residue in theP2 position, which allows the closing of the cycle on its sidechain. These molecules have a free N-terminus, necessary forbinding to the CD26 catalytic site, and a latent quinoniminiummethide electrophile, responsible for irreversible inhibition. Themolecule cyclo[εLys-Pro-3-aminobenzoic acid-(6-CH2S1Me2)-Gly-Gly-] 2 CF3COO2, referred to as ICI-11a, was used here.The reversible DPP IV inhibitor Lys-[Z(NO2)]-pyrrolidide([Z(NO2)], 4-nitrobenzyloxycarbonyl) was kindly provided byDr A. Barth (Halle, Germany).
DPP IV and DPP II activities. The DPP IV activity wasassayed by measuring the cleavage of Gly-Pro-NH-Np at pH 7.6[35], by either cell extracts or intact cells. For determination ofdipeptidyl peptidase IV activity using cell extracts or partiallypurified preparations, 50µl appropriately diluted samples wereincubated in a total volume of 200µl NaCl/Pi containing1mMGly-Pro-NH-Np at 37°C for 120 min in flat-bottom 96-wellmicroplates; activity was determined by measuring absorptionat 405 nm.
Dipeptidyl peptidase II activity in cell extracts and partiallypurified fractions was also measured in flat-bottom 96 wellmicroplates, but using1 mM Ala-Ala-NH-Np in 100 mM so-dium phosphate buffer, pH 5.7, which is the reported optimumpH for DPP II activity [2].
The Km of DPP IV activity of CD26 and DPP IV-β for Gly-Pro-NH-Np hydrolysis was determined by incubating partiallypurified preparations of both enzymes with different concentra-tions of Gly-Pro-NH-Np (ranging from1 µM to 10 mM) inNaCl/Pi. The effect of inhibitors on DPP IV activity was assayedby incubating DPP IV-β and CD26 preparations with different
371Blanco et al. (Eur. J. Biochem. 256)
concentrations of each inhibitor ranging from1 nM to 10 µM,for 15 min, before adding the substrate as indicated.Km and con-centration causing 50% inhibition (IC50) values were calculatedby non-linear regression from activity data as described [36].
When intact cells were used, 23106 cells in 0.5 ml100 mMHepes, pH 7.6,120 mM NaCl, 5 mM KCl, 1.2 mM MgSO4,8 mM glucose and1% BSA, were incubated with 0.5 mM Gly-Pro-NH-Np at 37°C for 90 min. The reactions were stopped bythe addition of1M sodium acetate, pH 4.5 (1 ml). After centrif-ugation at12000g, the production of NH-Np in the supernatantwas determined by measuring absorption at 405 nm.
Chromatography. A FPLC system (Pharmacia) was usedin the chromatographic procedure of DPP IV-β purification. ASuperose-6 column (1.6 cm350 cm, Pharmacia) was used as de-scribed [29]. Briefly, before loading samples, the column wasequilibrated with 20 mM Tris/HCl, pH 7.6, 50 mM NaCl and0.1% Triton X-100 (buffer GF). Elution was performed withthe same buffer, collecting1-ml fractions/2 min. Aliquots (50µl)from each fraction were assayed for DPP IV activity by cleavageof the Gly-Pro-NH-Np at pH 7.6 and for DPP II activity bycleavage of the DPP-II-specific substrate Ala-Ala-NH-Np atpH 5.5.
A Mono Q HR 5/5 column (1 ml, Pharmacia) was also used,after equilibration with GF buffer. Loading and elution were per-formed at1ml/min. Bound proteins were eluted using a linearNaCl gradient of 502500 mM in GF buffer. Fractions of 0.5 mlwere collected and assayed for DPP IV and DPP II activities asdescribed.
Binding of DPP IV-β to different lectins. Fractions froman ion-exchange column containing DPP IV-β activity were di-luted to obtain a NaCl concentration of150 mM, and the neces-sary volumes of1 M stock solutions of CaCl2 and MnCl2 wereadded to adjust the Ca21 and Mn21 concentrations to1 mM.These preparations were then assayed for their ability to bind todifferent lectins. Aliquots were incubated with one volume aga-rose-coupled lectins previously equilibrated with10 mM Tris,150 mM NaCl,1 mM CaCl2 and 1 mM MnCl2, pH 7.4 (bufferLCB), for 3 h at room temperature. Lectins used were Jacalin,concanavalin A, Helix Pomatia, PHA-L, PHA-E and wheat germagglutinin (WGA); all from Sigma. Agarose slurry was thenpoured into a 0.5 cm310 cm column and washed with 5 vol.LCB. DPP II and DPP IV activities were determined in the dif-ferent fractions: unbound, washed and bound. The bound activ-ity was monitored by directly incubating washed agarose withGly-Pro-NH-Np as the soluble fractions. The DPP IV-β boundto the WGA-Sepharose was eluted using a linearN-acetyl-D-glucosamine gradient of 0.120.5 M in buffer LCB. Fractionsof 0.5 ml were collected and assayed for DPP IV activity asdescribed.
RESULTS
DPP IV-β has properties similar to the serine exopeptidaseCD26. We have recently described the characterization of a newprotein with dipeptidyl peptidase IV activity, referred to as DPPIV-β. This protein, with an apparent molecular mass of 82 kDa,is expressed on the cell surface as is the case with CD26 [29].The discovery of DPP IV-β was carried out by the use of theCD26-negative cell line, C8166 cells. Accordingly, extracts ofC8166 cells were used here as a suitable source of this enzyme,whereas extracts of CEM cells transfected to express high levelsof CD26 were used as a source of CD26 [29, 30]. It should benoted that C8166 and CEM cells do not express the recentlydescribed175-kDa form of DPP IV, referred to as DPP T-L.Consequently, assay of DPP IV activity using extracts of C8166
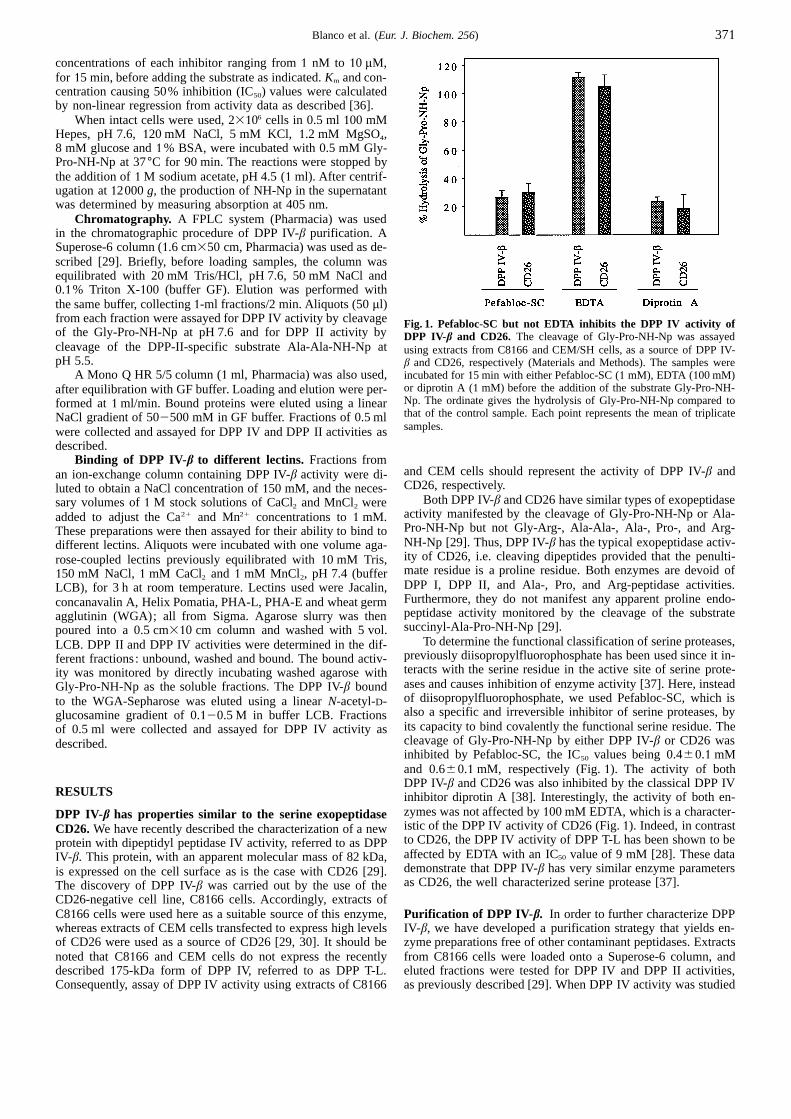
Fig. 1. Pefabloc-SC but not EDTA inhibits the DPP IV activity ofDPP IV-β and CD26. The cleavage of Gly-Pro-NH-Np was assayedusing extracts from C8166 and CEM/SH cells, as a source of DPP IV-β and CD26, respectively (Materials and Methods). The samples wereincubated for15 min with either Pefabloc-SC (1 mM), EDTA (100 mM)or diprotin A (1 mM) before the addition of the substrate Gly-Pro-NH-Np. The ordinate gives the hydrolysis of Gly-Pro-NH-Np compared tothat of the control sample. Each point represents the mean of triplicatesamples.
and CEM cells should represent the activity of DPP IV-β andCD26, respectively.
Both DPP IV-β and CD26 have similar types of exopeptidaseactivity manifested by the cleavage of Gly-Pro-NH-Np or Ala-Pro-NH-Np but not Gly-Arg-, Ala-Ala-, Ala-, Pro-, and Arg-NH-Np [29]. Thus, DPP IV-β has the typical exopeptidase activ-ity of CD26, i.e. cleaving dipeptides provided that the penulti-mate residue is a proline residue. Both enzymes are devoid ofDPP I, DPP II, and Ala-, Pro, and Arg-peptidase activities.Furthermore, they do not manifest any apparent proline endo-peptidase activity monitored by the cleavage of the substratesuccinyl-Ala-Pro-NH-Np [29].
To determine the functional classification of serine proteases,previously diisopropylfluorophosphate has been used since it in-teracts with the serine residue in the active site of serine prote-ases and causes inhibition of enzyme activity [37]. Here, insteadof diisopropylfluorophosphate, we used Pefabloc-SC, which isalso a specific and irreversible inhibitor of serine proteases, byits capacity to bind covalently the functional serine residue. Thecleavage of Gly-Pro-NH-Np by either DPP IV-β or CD26 wasinhibited by Pefabloc-SC, the IC50 values being 0.460.1 mMand 0.660.1 mM, respectively (Fig.1). The activity of bothDPP IV-β and CD26 was also inhibited by the classical DPP IVinhibitor diprotin A [38]. Interestingly, the activity of both en-zymes was not affected by100 mM EDTA, which is a character-istic of the DPP IV activity of CD26 (Fig.1). Indeed, in contrastto CD26, the DPP IV activity of DPP T-L has been shown to beaffected by EDTA with an IC50 value of 9 mM [28]. These datademonstrate that DPP IV-β has very similar enzyme parametersas CD26, the well characterized serine protease [37].
Purification of DPP IV- β. In order to further characterize DPPIV-β, we have developed a purification strategy that yields en-zyme preparations free of other contaminant peptidases. Extractsfrom C8166 cells were loaded onto a Superose-6 column, andeluted fractions were tested for DPP IV and DPP II activities,as previously described [29]. When DPP IV activity was studied
372 Blanco et al. (Eur. J. Biochem. 256)
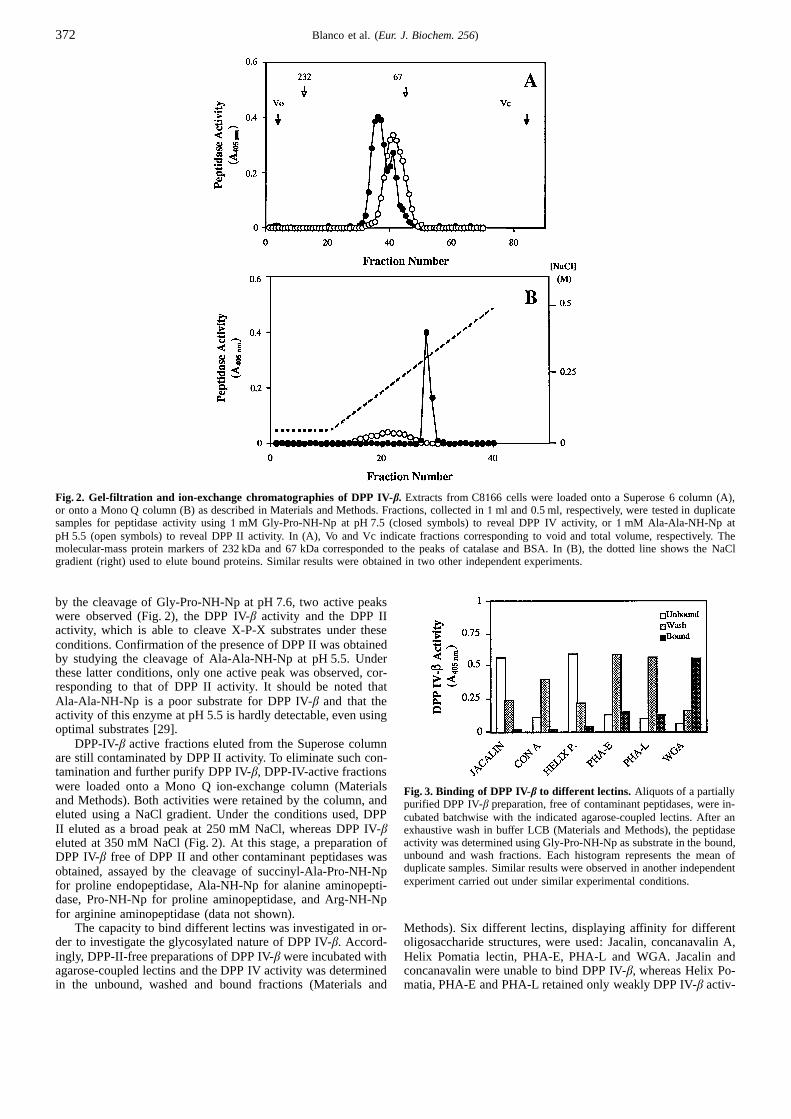
Fig. 2. Gel-filtration and ion-exchange chromatographies of DPP IV-β. Extracts from C8166 cells were loaded onto a Superose 6 column (A),or onto a Mono Q column (B) as described in Materials and Methods. Fractions, collected in1 ml and 0.5 ml, respectively, were tested in duplicatesamples for peptidase activity using1 mM Gly-Pro-NH-Np at pH 7.5 (closed symbols) to reveal DPP IV activity, or1 mM Ala-Ala-NH-Np atpH 5.5 (open symbols) to reveal DPP II activity. In (A), Vo and Vc indicate fractions corresponding to void and total volume, respectively. Themolecular-mass protein markers of 232 kDa and 67 kDa corresponded to the peaks of catalase and BSA. In (B), the dotted line shows the NaClgradient (right) used to elute bound proteins. Similar results were obtained in two other independent experiments.
by the cleavage of Gly-Pro-NH-Np at pH 7.6, two active peakswere observed (Fig. 2), the DPP IV-β activity and the DPP IIactivity, which is able to cleave X-P-X substrates under theseconditions. Confirmation of the presence of DPP II was obtainedby studying the cleavage of Ala-Ala-NH-Np at pH 5.5. Underthese latter conditions, only one active peak was observed, cor-responding to that of DPP II activity. It should be noted thatAla-Ala-NH-Np is a poor substrate for DPP IV-β and that theactivity of this enzyme at pH 5.5 is hardly detectable, even usingoptimal substrates [29].
DPP-IV-β active fractions eluted from the Superose columnare still contaminated by DPP II activity. To eliminate such con-tamination and further purify DPP IV-β, DPP-IV-active fractionswere loaded onto a Mono Q ion-exchange column (Materialsand Methods). Both activities were retained by the column, andeluted using a NaCl gradient. Under the conditions used, DPPII eluted as a broad peak at 250 mM NaCl, whereas DPP IV-βeluted at 350 mM NaCl (Fig. 2). At this stage, a preparation ofDPP IV-β free of DPP II and other contaminant peptidases wasobtained, assayed by the cleavage of succinyl-Ala-Pro-NH-Npfor proline endopeptidase, Ala-NH-Np for alanine aminopepti-dase, Pro-NH-Np for proline aminopeptidase, and Arg-NH-Npfor arginine aminopeptidase (data not shown).
The capacity to bind different lectins was investigated in or-der to investigate the glycosylated nature of DPP IV-β. Accord-ingly, DPP-II-free preparations of DPP IV-β were incubated withagarose-coupled lectins and the DPP IV activity was determinedin the unbound, washed and bound fractions (Materials and
Fig. 3. Binding of DPP IV-β to different lectins. Aliquots of a partiallypurified DPP IV-β preparation, free of contaminant peptidases, were in-cubated batchwise with the indicated agarose-coupled lectins. After anexhaustive wash in buffer LCB (Materials and Methods), the peptidaseactivity was determined using Gly-Pro-NH-Np as substrate in the bound,unbound and wash fractions. Each histogram represents the mean ofduplicate samples. Similar results were observed in another independentexperiment carried out under similar experimental conditions.
Methods). Six different lectins, displaying affinity for differentoligosaccharide structures, were used: Jacalin, concanavalin A,Helix Pomatia lectin, PHA-E, PHA-L and WGA. Jacalin andconcanavalin were unable to bind DPP IV-β, whereas Helix Po-matia, PHA-E and PHA-L retained only weakly DPP IV-β activ-

373Blanco et al. (Eur. J. Biochem. 256)
Fig. 4. Isolation of cell-surface-expressed DPP IV-β. Extracts frombiotinylated and nonbiotinylated C8166 cells (1 :2 ratio) were subjectedto different chromatographic steps in order to purify DPP IV-β. Aftereach purification step, an aliquot was analyzed by SDS/PAGE and bio-tinylated proteins were detected by using horseradish peroxidase-cou-pled streptavidin. The figure shows the autoradiogram obtained afterchemoluminiscent detection of total extracts (1), active fractions pooledafter superose 6 chromatography (2), active fractions after Mono Q chro-matography (3), and final preparation after WGA chromatography (4).Molecular-mass markers are given on the left. Bottom numbers refer tothe amount of recovered DPP IV-β activity after each step in comparisonto total initial peptidase activity, which includes DPP IV-β and DPP IIactivities.
Fig. 5. Determination of Michaelis constants for Gly-Pro-NH-Np hy-drolysis by CD26 and DPP IV-β. Michaelis constant for Gly-Pro-NH-Np hydrolysis was determined in triplicate samples in PB buffer usingincreasing substrate concentrations (ranging over 0.01210 µM). The fig-ure shows a Scatchard plot of activity values of CD26 (closed symbols)and DPP IV-β (open symbols).Km values determined by non-linear re-gression were 0.1260.02 mM for CD26 and 0.2860.05 mM for DPPIV-β (value6SD given by the program).
ity (Fig. 3), which was recovered mainly in the washing process.In contrast, WGA strongly bound DPP IV-β ; more than 75% ofthe loaded activity was still bound after the washing process.These results suggested that DPP IV-β behaves as a glycosylatedprotein. Consistent with this, the WGA-bound material was re-covered by elution at 0.3 MN-acetyl-D-glucosamine (describedin Materials and Methods).
In order to identify the purified DPP IV-β, biotin-labeledproteins were used. For this purpose, C8166 cells were first la-beled with biotin in order to label cell surface proteins beforepreparation of cytoplasmic extracts. DPP IV-β was then purifiedsuccessively by Superose 6, Mono Q and WGA chromatogra-phy. Purification was monitored by measuring DPP IV activityand by determining the content of biotinylated proteins in puri-fied fractions. The final purified fraction obtained after elutionfrom the WGA column contained a major 75280-kDa biotiny-lated protein (Fig. 4) which retained12% of the initial DPP IVactivity (that corresponds to a 20% of initial dipeptidase activ-ity). The presence of biotin in this purified protein band con-
Table 1. Michaelis constants for Gly-Pro-NH-Np hydrolysis by DPPIV/CD26 and DPP IV-β. Purified fractions of each enzyme, obtainedafter Superose and mono Q chromatography, were used forKm determi-nation (each point was performed in triplicate samples). Activity datawere fitted using a non-linear regression program. Results areKm values6SD given by the program.
Enzyme source Gly-Pro-NH-Np hydrolysisKM
CD26 DPP IV-β
mM
C8166 2 0.3 60.1MOLT4 0.1160.01 0.2760.05CEM 0.1360.03 0.2560.04
Table 2. The effect of different inhibitors on the DPP IV activity ofCD26 and DPP IV-β. Partially purified fractions of each enzyme, ob-tained after Superose and mono Q chromatography, were used for IC50
determinations (each point was performed in triplicate samples). Activitydata were fitted using a non-linear regression program. Results are IC50
values6SD given by the program.
Inhibitor Enzyme IC50
sourceCD26 DPP IV-β
µM
Lys [Z(NO2)]-pyrrolidide C8166 2 0.960.1MOLT4 1.86 0.2 1.360.2CEM 1.66 0.1 1.260.1
ICI-11a C8166 2 2.960.1MOLT4 0.01160.001 2.660.2CEM 0.01760.003 2.660.1
firms the extracellular localization of DPP IV-β, as suggestedpreviously [29]. Furthermore, the molecular mass calculated inSDS/PAGE experiments was similar to that obtained in gel-filtration chromatography (82 kDa), thus suggesting that DPPIV-β is solubilized as a monomer from the cell surface in ourextraction conditions.
Kinetic properties of DPP IV-β and CD26. In our previousreport [29], in addition to the CD26-negative C8166 cell line,we studied the DPP IV activities expressed by the CD26-posi-tive MOLT-4 cell line by gel-filtration chromatography. In thiscell line, two DPP-IV-active peaks were observed, one corre-sponding to CD26 and a second peak corresponding to a molec-ular mass of 84 kDa that was not immunoprecipitated by theCD26-specific antibody1F7. This second peak was also foundin extracts from other CD26 expressing cells, as the differentCD26-transfected CEM clones obtained in our laboratory (notshown). In order to reveal that this second peak was DPP IV-β,both peaks were characterized in parallel. For this reason, westudied the kinetic parameters and the effect of specific inhibi-tors on partially purified preparations of each peak, obtainedafter Superose and Mono Q column chromatography of MOLTextracts. Firstly, theKm determined in DPP IV-β preparationsshowed a significant higher value (0.2860.05 mM) than thosefound in CD26 preparations (0.1260.02 mM; Fig. 5). Similarvalues were obtained when theKm was calculated using prepara-tions of the 82-kDa peak and CD26 obtained from different cells(Table1). Secondly, more important differences were found

374 Blanco et al. (Eur. J. Biochem. 256)
Fig. 6. Kinetics of substrate hydrolysis by CD26 and DPP IV-β in the presence of the irreversible inhibitor ICI-2a. The DPP IV activity wasmonitored in duplicate samples by the cleavage of Gly-Pro-NH-Np (0.5 mM) using extracts of CEM/SH (A) and C8166 (B) cells as the source ofCD26 and DPP IV-β, respectively, in the absence or presence of increasing concentrations (0.1, 1 and10 µM) of the irreversible cyclic inhibitorICI-11a. The reactions were stopped at different times (ordinate) and hydrolysis was monitored by measuring the absorbance at 405 nm (abscissa).Similar results were observed in another independent experiment.
when the sensitivity to different inhibitors was studied. As re-ported recently [29], classical DPP IV or general peptidase in-hibitors were unable to differentiate DPP IV-β and DPP IV activ-ity of CD26 (Fig.1), i.e. the inhibitor Lys [Z(NO2)]-pyrrolidideshowed only a slightly higher activity on DPP IV-β preparations(IC50 0.921.3 µM) than in CD26 preparations (IC50 1.621.8 µM; Table 2). However, the newly described irreversible cy-clic inhibitor [34] ICI-11a was about1002200-fold more activeon the CD26 peptidase activity (IC50 0.01120.017 µM) than onDPP IV-β activity (IC50 2.622.9µM; Table 2). These results fur-ther confirm that the 82-kDa peak found in the CD26-expressingcells, MOLT-4 and CEM is DPP IV-β. To further characterizethe significant difference observed in the inhibitory effect of ICI-11a on the activity of DPP IV-β compared to that of CD26, westudied the kinetics of the cleavage of Gly-Pro-NH-Np in thepresence of increasing concentrations of ICI-11a. The resultsdemonstrated, once again, that DPP IV-β responds poorly to theinhibitory effect of ICI-11a compared to CD26 (Fig. 6). Thecomplete inhibition of DPP IV-β by ICI-11a required concentra-tions higher than 30µM (not shown).
In order to demonstrate the irreversible nature of ICI-11a,we previously used a clone of CEM cells expressing very highlevels of recombinant CD26 [34]. In these latter experiments,intact cells were incubated with 5µM ICI-11a before washingto remove unbound inhibitor ICI-11a. Such cells were then cul-tured at 37°C and were monitored for the cell-surface DPP IVactivity of CD26 at different times. The results demonstratedthat10215 min incubation with ICI-11a was sufficient to inhibitcompletely the DPP IV activity of CD26 for a period of several
hours. The cell surface activity of CD26 became detectable at24 h coinciding with the doubling time of cells, which results inthe production of cells that express newly synthesized CD26molecules (Figs 2 and 3, and Table 2 in [34]). As it has beenreported for the irreversible inhibitors of trypsin-like proteases[39], we have suggested that the inhibitory process of ICI-11acould involve two steps: firstly, the reversible binding of theinhibitor to the enzyme, and secondly, the cleavage and subse-quent irreversible covalent modification of the enzyme leadingto its inactivation [34]. By calculating the equilibrium constantof the first step as well as the kinetic constant of the secondstep, we have concluded that the higher potency of the inhibitorICI-11a on the DPP IV activity of CD26 is the consequence ofa higher affinity for this enzyme, along with a faster inactivationrate for CD26 [34].
Expression of DPP IV-β in lymphoblastoid cell lines. Theabove results pointed out that, in spite of a similar catalytic ac-tivity, CD26 and DPP IV-β should have different structures toaccount for the lower inhibitory effect of the molecule ICI-11a.When the inhibitor was tested on the DPP IV activity of cell-surface-expressed DPP IV-β and CD26, ICI-11a completely in-hibited the activity of CD26 on the surface of CEM cellstransfected to express high levels of CD26, whereas it had noeffect on the DPP IV-β activity expressed on the cell surface ofC8166 cells [34]. This may be due to an inefficient covalentinteraction between the inhibitor and the catalytic site of DPPIV-β when expressed on the cell surface. Whatever the case, theinhibitor ICI-11a enabled us to easily evaluate both activities on

375Blanco et al. (Eur. J. Biochem. 256)
Fig. 7. Cell surface expression of CD26 and DPP IV-β in differentlymphoblastoid cell lines.(A) The expression of CD26 or DPP IV-β indifferent lymphoid cell lines. Cell-surface DPP IV activity was measuredin the absence or the presence of 25µM inhibitor ICI-11a. DPP IV-βactivity (white columns) was defined as the activity measured in thepresence of the inhibitor, whereas CD26 peptidase (black columns) ac-tivity was calculated by subtracting DPP IV-β activity from total activity.(B) Increasing cell-surface expression of CD26 does not modify the ex-pression of DPP IV-β. DPP IV activities of different CEM cell clonesexpressing either low levels (clone CEM/L), high levels (clone CEM/H)and very high levels of CD26 (clone CEM/SH), were assayed as de-scribed in (A) Similar results were obtained by quantifying DPP IV ac-tivity of CD26 and DPP IV-β after gel-filtration chromatography of ex-tracts from these cells. Each histogram represents the mean of triplicatesamples. The Jurkat cells used were the JBK cells which express verylow levels of CD26, detectable when cultured at low density. The expres-sion of CD26 on the surface of these cells could also be shown by FACSanalysis [16].
the cell surface of CD26-expressing cells. Thus, the cell-surfaceDPP IV activity associated with CD26 can be defined as theICI-11a-sensitive activity, whereas that associated to DPP IV-βrepresents the DPP IV activity which is resistant to ICI-11a.Using this latter approach, we could determine the expression ofboth proteases on the cell surface of different lymphoblastoidcell lines. The results obtained were comparable to those calcu-lated from quantification of each activity after gel-filtrationchromatography (not shown).
We observed that, in the different lymphoid cell lines tested,DPP IV-β activity was higher than CD26 peptidase activity, evenin the MOLT-4 cell line, known to express important levels ofCD26 (Fig. 7A). In order to investigate the relationship betweenthe expression of CD26 and DPP IV-β, we determined the DPPIV-β activity on CEM clones expressing, by transfection, dif-ferent levels of CD26; clones expressing either, low levels ofendogenous CD26 (CEM/L), clones expressing high (CEM/H)or very high (CEM/SH) levels of recombinant CD26. The ex-pression of CD26 did not modify the levels of DPP IV-β expres-sion, thus confirming that both enzymes are independent entities(Fig. 7B).
Expression of DPP IV-β in activated PBMC. In order to studythe expression and regulation of DPP IV-β in activated humanPBMC, extracts were prepared from non-stimulated PBMC, and
Fig. 8. Dipeptidyl peptidase activities in PBMC.Gel filtration of ex-tracts from human PBMC either before activation (A) and after 4 daysactivation by PHA10 µg/ml (B) or 10 µg/ml protein A (C) was carriedout at experimental conditions as described in Fig. 2A. Aliquots of eachfraction were assayed in duplicate samples for the cleavage of Gly-Pro-NH-Np (ordinates). The position for the peaks of CD26, DPP IV-β andDPP II is indicated in each panel. The dotted line in (B) and (C) showsthe DPP IV activity corresponding to the nonstimulated PBMC (i.e. be-fore activation). Reproducible results were obtained using PBMC fromanother donor. No DPP IV activity was observed in fractions correspond-ing to the175-kDa form of DPP IV (DPP T-L).
from PBMC stimulated for 2 days or 4 days with either PHA orprotein A, respectively. Extracts were analyzed by gel-filtrationchromatography and fractions obtained were assayed for DPPIV activity to monitor the peaks corresponding to CD26 andDPP IV-β (Fig. 8). Non-stimulated PBMC showed more than80% total DPP IV activity associated with DPP IV-β, whereasthe CD26 peptidase activity was a minor contribution to the totalactivity. After PHA stimulation, there was an enhancement ofexpression of both enzymes. The ratio between both activitieswas maintained at 2 days but was diminished at 4 days; at thistime DPP IV-β activity represented 60% of the total DPP IVactivity (Figs 8 and 9). In contrast, protein A stimulation ofPBMC led to a higher enhancement of CD26 expression in com-parison to DPP IV-β. After 4 days stimulation, protein-A-treated

376 Blanco et al. (Eur. J. Biochem. 256)
Fig. 9. Expression of DPP IV-β and CD26 by activated PBMC.Hu-man PBMC were activated either by10 µg/ml PHA (upper panel) or10 µg/ml protein A (lower panel) and extracts were prepared before(time 0) or at 2 and 4 days (ordinate). Extracts were analyzed by gel-filtration chromatography (as in Fig. 8) and the fractions were assayedfor DPP IV activity (Materials and Methods), the activity of CD26(closed symbols), the activity of DPP IV-β (open symbols). Each pointrepresents the mean of duplicate samples. Reproducible results were ob-tained using PBMC from another donor.
PBMC showed the highest values of CD26 peptidase activity,representing 55% of the total DPP IV activity (Figs 8 and 9).
It should be emphasized that, under our experimental condi-tions, the175-kDa form of DPP IV, referred to as DPP T-L [27,28], appeared not to be present in the PBMC. Due to its sizein gel-filtration chromatography (5002600 kDa), the DPP T-Lshould been well separated from CD26 and DPP IV-β. However,in the gel-filtration experiments using extracts from PBMC be-fore and after activation, no DPP IV activity was detectable infractions close to the void volume (Fig. 8), corresponding to theapparent molecular mass of DPP T-L.
DISCUSSION
Using CD262 T lymphoblastoid C8166 cells, we have char-acterized a CD26-like ectopeptidase manifesting DPP IV activ-ity, the DPP IV-β. Previously reported studies [29] showed thatthe profiles of small-substrate specificities and inhibitor mole-cules were almost indistinguishable for both, CD26 peptidaseactivity and DPP IV-β. In addition, both proteins manifested op-timum dipeptidyl peptidase activity at pH 7.5, and resisted pro-teinase K digestion. However, two distinct properties differenti-ate both proteins ; the selective binding of ADA to CD26, andthe molecular mass of110 kDa for CD26 and 82 kDa for DPPIV-β. We also reported that the CD26 mAb 4H12 stained C8166cells, thus suggesting that it cross reacts with a cell surface anti-gen which shares a common epitope with CD26, most likelyDPP IV-β. However, this mAb does not immunoprecipitate DPPIV-β, and so, it cannot be used in the immunopurification of thisenzyme. For this reason, we have developed a classical chroma-tographic procedure in order to purify DPP IV-β. The biotinyla-
tion of cell surface proteins before the purification provided evi-dence that DPP IV-β is a glycoprotein expressed on the cellsurface. The molecular mass of 75280 kDa estimated after SDS/PAGE analysis of the final purified biotin-labeled protein wasin accord with the value obtained by gel-filtration chromatogra-phy, thus favoring the suggestion that DPP IV-β is solubilizedfrom the plasma membrane as a monomer.
Several CD26-related proteins from different sources havebeen recently described by different groups. On the basis of se-quence homology, different cell surface proteins have been iden-tified; FAPA [40], DPPX proteins [41] and BSPL [42], whichshow 48, 40 and 30% identity to CD26, respectively. Neverthe-less, these three proteins lack dipeptidyl peptidase activity. Twoother proteins have been reported to have an intrinsic DPP IVactivity similar to that of CD26; a175-kDa protein called DPPT-L [27, 28] and the 82 kDa protein DPP IV-β [29]. The newlydescribed proteins seem to share common epitopes with CD26,since they can be recognised by CD26-specific monoclonal anti-bodies [27, 29]. Other common feature of these proteins andCD26 is the resistance of large regions to proteolytic cleavage,and their expression on the immune system. Interestingly, thecapacity to bind ADA is a specific feature of CD26. AlthoughCD26, DPP T-L and DPP IV-β are related, because of their cata-lytic activity, their differential expression suggests that they areindependent entities. Accordingly, here we show that there is nostrict correlation between the expression of DPP IV-β and CD26(Fig. 7), and Duke-Cohan and co-workers [28] found no correla-tion between DPP T-L and CD26 expression in different lym-phoid and non-lymphoid cell lines. Moreover, Jurkat andMOLT4 cells, which express important levels of DPP IV-β(Fig. 7), have been reported not to express DPP T-L [28]. Itshould be noted that, in contrast to the results of Duke-Cohan etal. [27, 28], we did not detect DPP T-L in extracts from activatedPBMC. This apparent discordance can be accounted for by con-sidering that the DPP T-L levels are a minor form of cell-associ-ated DPP IV activity or, alternatively, by the rapid release ofDPP T-L, or even due to our experimental conditions. In thisrespect, it should be noted that we cultured PBMC in mediumsupplemented with10% fetal calf serum in contrast to the se-rum-free culture conditions used by Duke-Cohan et al. [27, 28].The mechanism of the DPP T-L release by the activated PBMCis not known. Thus, it remains of interest to determine whetherserum could regulate in a negative manner the production ofDPP T-L.
The DPP IV activity has been suggested to be essential forT-cell activation, although its role in the different stimulationmechanisms has not been completely elucidated. It seems clearthat this activity plays an important role in antigen-induced T-cell proliferation, as addition of recombinant human solubleCD26 was shown to lead to an enhancement of tetanus-toxoid-induced T-cell proliferation [24]. Furthermore, the inhibitor ofDPP IV activity Proboro-Pro, was reported to cause an impair-ment of this proliferation, a phenomenon reversed by the addi-tion of purified porcine CD26 [43]. It has been also reportedthat addition of DPP T-L has a similar effect on tetanus-toxoid-stimulated T cells [28], thus suggesting that a similar mechanismexists for the costimulatory effect of both proteins, most likelydependent on its enzymatic activity, which might act upon oneor more potential substrates. In fact, DPP IV activity may cleavesome bioactive peptides, such as substance P, chorionic gonado-tropin hormone, monomeric fibrin and glucose-dependent insu-linotropic polypeptide. Other N-terminus Xaa-Pro potential DPPIV substrates are some cytokines, such as interleukin-1β, in-terleukin-2, tumour necrosis factorβ, granulocyte colony-stimu-lating factor, and chemokines that inhibit HIV infection (SDF-1,RANTES, MIP-1A and MIP-1β) [44, 45]. More recently it has

377Blanco et al. (Eur. J. Biochem. 256)
been shown that the DPP IV activity of CD26 cleaves RANTESand other chemokines, such as Eotaxin and monocyte chemotac-tic protein-2. In the case of RANTES, this cleavage results in astill active peptide showing a restricted receptor selectivity ifcompared with the whole RANTES molecule [20]. Since post-proline cleavage is a limiting step in peptide degradation [2],cleavage of chemokines may affect, not only selectivity, but alsohalf-life of active peptides.
The observation that different cells express two isoforms ofDPP IV activity on their surfaces (Fig. 7) raises the questionabout the complementary or supplementary role of each isoformin peptide processing. The different isoforms of DPP IV activitymay share different substrates, as suggested by the costimulatoryeffect of CD26 and DPP T-L on antigen-induced T-cell activa-tion [24, 27]. However, it is also possible that some substratesare cleaved specifically or preferentially by one of these iso-forms. The results onKm values of CD26 and DPP IV-β forsubstrate hydrolysis (Table1) and the use of the irreversible cy-clic inhibitor of DPP IV activity, ICI-11a (Table 2), demonstratethat there are important differences between the affinity ofDPPIV/CD26 and DPP IV-β for some small substrates (Gly-Pro-NH-Np) or inhibitors (ICI-11a). These differences may probablyreflect divergences in the spatial structure of the catalytic site ofthese enzymes. Of special interest is the fact that specific DPPIV inhibitors of CD26 does not affect DPP IV-β activity whenthe enzyme is expressed on the cell surface. Such inhibitorscould be useful in different studies to understand the physiologi-cal significance of DPP IV-β in respect to CD26.
We thank Miche`le Fevrier for providing PBMC and Nadine Robertand Josette Svab for excellent technical assistance.
REFERENCES
1. Alba, F., Arenas, J. C. & Lopez, M. A. (1995) Properties of rat braindipeptidyl aminopeptidases in the presence of detergents,Peptides16, 3252329.
2. Yaron, A. & Naider, F. (1993) Proline-dependent structural and bio-logical properties of peptides and proteins,Crit. Rev. Biochem.Molec. Biol. 28, 31281.
3. Fukasawa, K., Fukasawa, K. & Sahara, N. (1981) Immunohisto-chemical localization of dipeptidyl aminopeptidase IV in rat kid-ney, liver, and salivary glands,J. Histochem. Cytochem. 29, 3372343.
4. Morrison, M. E., Vijayasaradhi, S., Engelstein, D., Albino, A. P. &Houghton, A. N. (1993) A marker for neoplastic progression ofhuman melanocytes is a cell surface ectopeptidase,J. Exp. Med.177, 113521143.
5. Kameoka, J., Tanaka, T., Nojima, Y., Schlossman, S. F. & Morimoto,C. (1993) Direct association of adenosine deaminase with a T cellactivation antigen, CD26,Science 261, 1662469.
6. De Meester, I., Vanham, G., Kestens, L., Vanhoof, G., Bosmans, E.,Gigase, P. & Scharpe, S. (1994) Binding of adenosine deaminaseto the lymphocyte surface via CD26,Eur. J. Immunol. 24, 5662570.
7. Fleischer, B. (1994) CD26: a surface protease involved in T-cellactivation,Immunol. Today 15, 1802184.
8. Tiruppathi, C., Miyamoto, Y., Ganapathy, V., Roesel, R. A., Whit-ford, G. M. & Leibach, F. H. (1990) Hydrolysis and transportof proline-containing peptides in renal brush-border membranevesicles from dipeptidyl peptidase IV-positive and dipeptidyl pep-tidase IV-negative rat strains,J. Biol. Chem. 26, 147621483.
9. Fox, D. A., Hussey, R. E., Fitzgerald, K. A., Acuto, O., Poole, C.,Palley, L., Daley, J. F., Schlossman, S. F. & Reinherz, E. L. (1984)Ta1, a novel105 KD human T cell activation antigen defined bya monoclonal antibody,J. Immunol. 133, 125021255.
10. Torimoto, Y., Dang, N. H., Vivier, E., Tanaka, T., Schlossman, S.F. & Morimoto, C. (1991) Coassociation of CD26 (dipeptidylpeptidase IV) with CD45 on the surface of human T lymphocytes,J. Immunol. 147, 251422517.
11. Dang, N. H., Torimoto, Y., Sugita, K., Daley, J. F., Schow, P., Prado,C., Schlossman, S. F. & Morimoto, C. (1990) Cell surface modu-lation of CD26 by anti-1F7 monoclonal antibody. Analysis of sur-face expression and human T cell activation,J. Immunol. 145,396323971.
12. Martin, M., Huguet, J., Centelles, J. J. & Franco, R. (1995) Expres-sion of ecto-adenosine deaminase and CD26 in human T cellstriggered by the TCR-CD3 complex. Possible role of adenosinedeaminase as costimulatory molecule,J. Immunol. 155, 463024643.
13. Callebaut, C., Krust, B., Jacotot, E. & Hovanessian, A. G. (1993) Tcell activation antigen, CD26, as a cofactor for entry of HIV inCD41 cells,Science 262, 204522050.
14. Morimoto, C., Lord, C. I., Zhang, C., Duke-Cohan, J. S., Letvin, N.L. & Schlossman, S. F. (1994) Role of CD26/dipeptidyl peptidaseIV in human immunodeficiency virus type1 infection andapoptosis,Proc. Natl Acad. Sci. USA 91, 996029964.
15. Oravezc, T., Roderiquez, G., Koffi, J., Wang, J., Ditto, M., Bou-Habib, D. C., Lusso, P. & Norcross, M. A. (1995) CD26 expres-sion correlates with entry, replication and cytopathicity of mono-cytotropic HIV-1 strains in a T-cell line,Nat. Med. 1, 9192926.
16. Jacotot, E., Callebaut, C., Blanco, J., Rivie`re, Y., Krust, B. & Ho-vanessian, A. G. (1996a) HIV envelope glycoprotein-induced cellkilling by apoptosis is enhanced with increased expression ofCD26 in CD41 T cells,Virology 223, 3182330.
17. Valenzuela, A., Blanco, J., Callebaut, C., Jacotot, E., Lluı´s, C.,Franco, R. & Hovanessian, A. G. (1997) Adenosine deaminasebinding to CD26 is inhibited by HIV-1 glycoprotein gp120 andviral particles,J. Immunol. 158, 372123729.
18. Gutheil, W. G., Subramanyam, M., Flentke, G. R., Sanford, D. G.,Munoz, E., Huber, B. T. & Bachovchin, W. W. (1994) Humanimmunodeficiency virus1 Tat binds to dipeptidyl aminopeptidaseIV (CD26) : a possible mechanism for Tat’s immunosuppressiveactivity, Proc. Natl Acad. Sci. USA 91, 659426598.
19. Blanco, J., Marie´, I., Callebaut, C., Jacotot, E., Krust, B. & Ho-vanessian, A. G. (1996) Specific binding of adenosine deaminasebut not HIV-1 transactivator protein Tat to human CD26,Exp.Cell Res. 225, 1022111.
20. Oravecz, T., Pall, M. Roderiquez, R., Gorrell, M. D., Ditto, M.,Nguyen, N., Boykins, R., Unsworth, E. & Norcross, M. A. (1997)Regulation of the receptor specificity and function of the chemo-kine RANTES (Regulated on Activation, Normal T cell Ex-pressed and Secreted) by Dipeptidyl Peptidase IV (CD26)-medi-ated cleavage,J. Exp. Med. 186, 186521872.
21. D’Souza, M. P. & Harden, V. A. (1996) Chemokines and HIV-1second coreceptors,Nat. Med.2, 129321300.
22. Ansorge, S., Bühling, F., Hoffmann, T., Kähne, T., Neubert, K. &Reinhold, D. (1995) DP IV/CD26 on human lymphocytes : Func-tional roles in cell growth and cytokine regulation, in: DipeptidylPeptidase IV (CD26) in metabolism and the immune response(Fleischer, B., ed.) pp.1632184, R. G. Landes Company, Austin.
23. Kahne, T., Neubert, K. & Ansorge, S. (1995) Enzymatic activity ofDPIV/CD26 is involved in PMA-induced hyperphosphorylationof p56lck, Immunol. Lett. 46, 1892193.
24. Tanaka, T., Kameoka, J., Yaron, A., Schlossman, S. F. & Morimoto,C. (1993) The costimulatory activity of the CD26 antigen requiresdipeptidyl peptidase IV enzymatic activity,Proc. Natl Acad. Sci.USA 90, 458624590.
25. Steeg, C., Hartwig, U. & Fleischer, B. (1995) Unchanged signalingcapacity of mutant CD26/dipeptidylpeptidase IV molecules de-void of enzymatic activity,Cell Immunol. 164, 3112315.
26. Hedrick, J. A. & Zlotnik, A. (1996) Chemokines and lymphocytebiology, Curr. Opinion Immunol. 8, 3432347.
27. Duke-Cohan, J. S., Morimoto, C., Rocker, J. A. & Schlossman, S.F. (1995) A novel form of dipeptidylpeptidase IV found in humanserum. Isolation, characterization, and comparison with T lym-phocyte membrane dipeptidylpeptidase IV (CD26),J. Biol. Chem.270, 14107214114.
28. Duke-Cohan, J. S., Morimoto, C., Rocker, J. A. & Schlossman, S.F. (1996) Serum high molecular mass dipeptidyl peptidase IV(CD26) is similar to a novel antigen DPPT-L released from acti-vated T cells,J. Immunol. 156, 171421721.
29. Jacotot, E., Callebaut, C., Blanco, J., Krust, B., Neubert, K., Barth,A. & Hovanessian, A. G. (1996b) Dipeptidyl-peptidase IV-β, a

378 Blanco et al. (Eur. J. Biochem. 256)
novel form of cell-surface-expressed protein with dipeptidyl-pep-tidase IV activity,Eur. J. Biochem. 239, 2482258.
30. Callebaut, C., Jacotot, E., Blanco, J., Krust, B. & Hovanessian, A.G. (1998) Increased rate of HIV-1 and its cytopathic effect inCD41/CXCR41T cells expressing relatively high levels ofCD26, Exp. Cell Res. 241, 3522362.
31. Boyum, A. (1968) Isolation of mononuclear cells and granulocytesfrom human blood. Isolation of monuclear cells by one centrifu-gation, and of granulocytes by combining centrifugation and sedi-mentation at1 g, Scand. J. Lab. Clin. Invest. 97, 77289.
32. Cole, S. R., Ashman, L. K. & Ely, P. L. (1987) Biotinylation: analternative to radioiodination for the identification of cell surfaceantigens in immunoprecipitates,Mol. Immunol. 24, 6992705.
33. Laemmli, U. K. (1970) Cleavage of structural proteins during theassembly of the head of bacteriophage T4,Nature 227, 6802685.
34. Nguyen, C., Blanco, J., Mazaleyrat, J. P., Krust, B., Callebaut, C.,Jacotot, E., Hovanessian, A. G. & Wakselman, M. (1998) Specificand irreversible cyclopeptide inhibitors of the dipeptidyl peptidaseactivity of the T-cell activation antigen CD26,J. Med Chem. 41,210022110.
35. Bauvois, B., Sanc¸eau, J. & Wietzerbin, J. (1992) Human U937 cellsurface peptidase activities: characterization and degradative ef-fect on tumor necrosis factor-alpha,Eur. J. Immunol. 22, 9232930.
36. Blanco, J., Canela, E. I., Mallol, J., Lluı´s, C. & Franco, R. (1992)Characterization of adenosine receptors in brush-border mem-branes from pig kidney,Br. J. Pharmacol. 107, 6712678.
37. Fleischer, B. (1995) Dipeptidyl peptidase IV (CD26) in metabolismand the immune response, inMolecular biology intelligence unit,RG Landes Company, Austin, Texas, USA.
38. Umezawa, H., Aoyagi, T., Ogawa, K., Naganawa, H., Hamada,M. & Takeuchi, T. (1984) Diprotins A and B, inhibitors of dipep-tidyl aminopeptidase IV, produced by bacteria,J. Antibiotics. 37,4222425.
39. Wakselman, M., Xie, J., Mazaleyrat, J. P., Bogetto, N., Vilain, A.C., Montagne, J. J. & Reboud-Ravaux, M. (1993) New mecha-nism-based inactivators of trypsin-like proteinases. Selective inac-tivation of urokinase by functionalized cyclopeptides incorporat-ing a sulfoniomethyl-substituted m-aminobenzoic acid residue,J.Med. Chem. 36, 153921547.
40. Scanlan, M. J., Mohan Raj, B. K., Calvo, B., Garin-Chesa, P., Sanz-Moncasi, M. P., Healey, J. H., Old, L. J. & Rettig, W. J. (1994)Molecular cloning of fibroblast activation proteinA, a member ofthe serine protease family selectively expressed in stromal fibro-blasts of epithelial cancers,Proc. Natl Acad. Sci. USA 91, 565725661.
41. Wada, K., Yokotani, N., Hunter, C., Doi, K., Wenthold, R. J. &Shimasaki, S. (1992) Differential expression of two distinct formsof mRNA encoding members of a dipeptidyl aminopeptidase fam-ily, Proc. Natl Acad. Sci. USA 89, 1972201.
42. De Lecea, L., Soriano, E., Criado, J. R., Steffensen, S. C., Henrik-sen, S. J. & Sutcliffe, J. G. (1994) Transcripts encoding a neuralmembrane CD26 peptidase-like protein are stimulated by synapticactivity, Mol. Brain Res. 25, 2862296.
43. Subramanyam, M., Gutheil, W. G., Bachovchin, W. W. & Huber, B.T. (1993) Mechanism of HIV induced inhibition of antigen-spe-cific T-cell responsiveness,J. Immunol. 150, 254422553.
44. Oberlin, E., Amara, A., Bachelerie, F., Bessia, C., Virelizier, J. L.,Arenzana-Seisdedos, F., Schwartz, O., Heard, J. M., Clarck-Lewis, I., Legler, D. F., Loetscher, M., Baggiolini, M. & Moser,B. (1996) The CXC chemokine, stromal cell derived factor1(SDF-1), is the ligand for LESTR/fusin and prevents infection bylymphocyte-tropic HIV-1 syncytium-inducing strains,Nature 382,8332837.
45. Dragic, T., Litwin, V., Allaway, G. P., Martin, S. R., Huang, Y.,Nagashima, K. A., Cayanan, C., Maddon, P. J., Koup, R. A.,Moore, J. P. & Paxton, W. A. (1996) HIV-1 entry into CD41cells is mediated by the chemokine receptor CC-CKR-5,Nature381, 6672671.




















