
Identification and expansion of highly suppressive CD8(+)FoxP3(+) regulatory T cells after...
12
doi:10.1182/blood-2011-12-396119 Prepublished online April 26, 2012; 2012 119: 5898-5908 MacDonald Markey, Motoko Koyama, Andrew D. Clouston, Christian R. Engwerda, Geoffrey R. Hill and Kelli P. A. Antiopi Varelias, Kylie A. Alexander, Bianca E. Teal, Tim Sparwasser, Gunter J. Hammerling, Kate A. Renee J. Robb, Katie E. Lineburg, Rachel D. Kuns, Yana A. Wilson, Neil C. Raffelt, Stuart D. Olver, transplantation regulatory T cells after experimental allogeneic bone marrow + FoxP3 + Identification and expansion of highly suppressive CD8 http://bloodjournal.hematologylibrary.org/content/119/24/5898.full.html Updated information and services can be found at: (1914 articles) Transplantation Articles on similar topics can be found in the following Blood collections http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requests Information about reproducing this article in parts or in its entirety may be found online at: http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprints Information about ordering reprints may be found online at: http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtml Information about subscriptions and ASH membership may be found online at: Copyright 2011 by The American Society of Hematology; all rights reserved. Washington DC 20036. by the American Society of Hematology, 2021 L St, NW, Suite 900, Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.org From For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.org From For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.org From For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.org From For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.org From For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.org From For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.org From For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.org From For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.org From For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.org From For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.org From For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.org From
-
Upload
qimrberghofer -
Category
Documents
-
view
1 -
download
0
Transcript of Identification and expansion of highly suppressive CD8(+)FoxP3(+) regulatory T cells after...

doi:10.1182/blood-2011-12-396119Prepublished online April 26, 2012;2012 119: 5898-5908
MacDonaldMarkey, Motoko Koyama, Andrew D. Clouston, Christian R. Engwerda, Geoffrey R. Hill and Kelli P. A.Antiopi Varelias, Kylie A. Alexander, Bianca E. Teal, Tim Sparwasser, Gunter J. Hammerling, Kate A. Renee J. Robb, Katie E. Lineburg, Rachel D. Kuns, Yana A. Wilson, Neil C. Raffelt, Stuart D. Olver, transplantationregulatory T cells after experimental allogeneic bone marrow
+FoxP3+Identification and expansion of highly suppressive CD8
http://bloodjournal.hematologylibrary.org/content/119/24/5898.full.htmlUpdated information and services can be found at:
(1914 articles)Transplantation �Articles on similar topics can be found in the following Blood collections
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.Washington DC 20036.by the American Society of Hematology, 2021 L St, NW, Suite 900, Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly
For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.orgFrom For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.orgFrom For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.orgFrom For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.orgFrom For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.orgFrom For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.orgFrom For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.orgFrom For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.orgFrom For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.orgFrom For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.orgFrom For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.orgFrom For personal use only. at MHH-Bibliothek on October 17, 2013. bloodjournal.hematologylibrary.orgFrom

TRANSPLANTATION
Identification and expansion of highly suppressive CD8�FoxP3� regulatoryT cells after experimental allogeneic bone marrow transplantationRenee J. Robb,1,2 Katie E. Lineburg,1 Rachel D. Kuns,1 Yana A. Wilson,1 Neil C. Raffelt,1 Stuart D. Olver,1 Antiopi Varelias,1
Kylie A. Alexander,1 Bianca E. Teal,1 Tim Sparwasser,3 Gunter J. Hammerling,4 Kate A. Markey,1 Motoko Koyama,1
Andrew D. Clouston,5 Christian R. Engwerda,6 *Geoffrey R. Hill,1,7 and *Kelli P. A. MacDonald8
1Bone Marrow Transplantation Laboratory, Queensland Institute of Medical Research, Brisbane, Australia; 2School of Medicine, University of Queensland,Brisbane, Australia; 3Institute for Medical Microbiology, Immunology and Hygiene, Technical University of Munich, Munich, Germany; 4Division of MolecularImmunology, Deutsches Krebsforschungszentrum, Heidelberg, Germany; 5Envoi Pathology, Brisbane, Australia; 6Immunology and Infection Laboratory,Queensland Institute of Medical Research, Brisbane, Australia; 7Department of Bone Marrow Transplantation, Royal Brisbane and Women’s Hospital, Brisbane,Australia; and 8Antigen Presentation and Immunoregulation Laboratory, Queensland Institute of Medical Research, Brisbane, Australia
FoxP3� confers suppressive propertiesand is confined to regulatory T cells (Treg)that potently inhibit autoreactive immuneresponses. In the transplant setting, natu-ral CD4� Treg are critical in controllingalloreactivity and the establishment oftolerance. We now identify an importantCD8� population of FoxP3� Treg that con-vert from CD8� conventional donor T cellsafter allogeneic but not syngeneic bonemarrow transplantation. These CD8� Treg
undergo conversion in the mesentericlymph nodes under the influence of recipi-ent dendritic cells and TGF-�. Impor-tantly, this population is as important forprotection from GVHD as the well-studiednatural CD4�FoxP3� population and ismore potent in exerting class I–restrictedand antigen-specific suppression in vitroand in vivo. Critically, CD8�FoxP3� Treg
are exquisitely sensitive to inhibition bycyclosporine but can be massively and
specifically expanded in vivo to preventGVHD by coadministering rapamycin andIL-2 antibody complexes. CD8�FoxP3�
Treg thus represent a new regulatory popu-lation with considerable potential to pref-erentially subvert MHC class I–restrictedT-cell responses after bone marrowtransplantation. (Blood. 2012;119(24):5898-5908)
Introduction
The development of GVHD remains a major limitation to thetherapeutic potential of bone marrow transplantation (BMT).GVHD occurs as a result of immunologic damage to the host tissueby conventional T and NK cells in the transplanted donor graft.Natural regulatory T cells (Treg) are defined by the expression ofFoxP31 and are dedicated to suppression of immune responses. Theimportance of CD4�FoxP3�Treg in peripheral tolerance is wellestablished, and the presence of naturally occurring CD4�Treg
makes them an attractive target for adoptive transfer to controlpathologic immune responses. Consequently, studies in GVHDusing freshly isolated or ex vivo expanded donor CD4�Treg havedemonstrated a delay or prevention of GVHD.2-5 Conversely,depletion of CD4�Treg from the donor graft results in an increasedseverity of GVHD.2 A limitation to using naive Treg in the adoptivetransfer setting is the high ratio required compared with FoxP3�
cells in the ingoing graft (1:1); thus, the number of Treg required isrestrictive. This has resulted in difficulties transferring adoptivecell therapy into the clinic with multiple groups now focusing onex vivo expansion protocols in an endeavor to overcome thislimitation.6-8
In contrast to the extensive research being completed on naiveCD4�Treg, studies investigating the role of Treg undergoing conver-sion in the periphery after BMT are lacking. The identification ofFoxP3 as a Treg marker1 and subsequent development of reagentsthat allow for identification (B6.FoxP3-GFP),9 tracking
(B6.FoxP3.LuciDTR)10 and depletion (B6.DEREG)11 of these cellshas allowed for a more thorough investigation. We have used thesereagents to characterize Treg populations after transplantation withthe aim of identifying more potent and antigen-specific Treg thatsuppress GVHD at low numbers after BMT. In conjunction withthe classic CD4�FoxP3�Treg population, we identified a novelCD8�FoxP3�Treg population that expands in the presence of analloreactive response and is previously unrecognized after BMT.Importantly, this population displays preferential tropism for thegastrointestinal (GI) tract and inhibits class I–restricted immuneresponses. Furthermore, rapamycin (RAPA) and IL-2 antibody(Ab) complex treatment after transplantation increases the percent-age of FoxP3-expressing cells within the CD8� population to morethan 30%. These cells thus represent a novel regulatory populationhighly suited to manipulation for the control of deleteriousalloimmune responses after transplantation.
Methods
Mice
Female C57BL/6 (B6.WT, H-2b,CD45.2�), B6.SJL-Ptprca (PTPrca,H-2b,CD45.1�), BALB/c (H-2d, CD45.2�), or B6D2F1 (H-2b/d,CD45.2�)mice were purchased from the Animal Resource Center. B6.FoxP3-GFP,9
B6.DEREG,11 B6.C-H2bm1 (bm1), FoxP3.LuciDTR-410 (B6.FoxP3-luc�),
Submitted December 1, 2011; accepted April 16, 2012. Prepublished online asBlood First Edition paper, April 26, 2012; DOI 10.1182/blood-2011-12-396119.
*G.R.H. and K.P.A.M. contributed equally to this study.
The publication costs of this article were defrayed in part by page chargepayment. Therefore, and solely to indicate this fact, this article is herebymarked ‘‘advertisement’’ in accordance with 18 USC section 1734.
© 2012 by The American Society of Hematology
5898 BLOOD, 14 JUNE 2012 � VOLUME 119, NUMBER 24

B6.MHC class II-GFPxDBA2F1, B6.�2m�/�, B6.�-actin-luc�, C57BL/6J.VavP-Bcl212 (B6.Bcl-2), and B6.CD11c-DOGxDBA2F113 mice weresupplied by the Queensland Institute of Medical Research animal facility.C3H.SW were purchased from The Jackson Laboratory and subsequentlybred at Queensland Institute of Medical Research animal facility. The age ofmice used ranged between 8 and 14 weeks. Mice were housed inmicroisolator cages and received acidified autoclaved water (pH 2.5) aftertransplantation.
Antibodies
The following antibodies were purchased from BioLegend: phycoerythrin(PE)–conjugated anti-H2Db(KH95), CD80(16/10A1), CD86(GL-1),CD40(1C10), CD45.1(A20), �4�7 (DATK32), CD127(SB/199),CD62L(MEL-14), anti–IL-10(JES5-16E3), anti–IFN-�(XM61.2), allophy-cocyanin-conjugated CD8�(53-6.7), CD4(RM4-5), anti-GITR(YGITR765),PE-Cy7–conjugated CD45.1(A20), biotin-conjugated anti-H2Dd(34-2-12),anti–CTLA-4(UC10-4B9), Pacific blue–conjugated anti-helios(22F6),Hamster anti-IgG(HTK888), and Alexa Flour-647 anti-FoxP3(150D). Thefollowing antibodies were purchased from BD Biosciences: PE-conjugatedCD4(GK1.5), CD25(7D4), CD103(M290), CXCR5(2G8), CD54(3E2),PE-Cy7–conjugated CD8(53-6.7) and biotin-conjugated CD28(37.51). PE-conjugated anti-GARP(YG1C86) and IL-17(18F10) were purchased fromeBioscience, and allophycocyanin-conjugated anti-Granzyme B(MHGB05)was purchased from Invitrogen. Anti–TGF-�14(1D11), anti–IL-10R(1B1.3a),CD3(2c11), and anti-NK1.115(PK136) were produced in house. Briefly,hybridoma cell culture supernatants were harvested and products precipi-tated using ammonium sulfate fractionation.16 Antibody was selected usingGammaBind G Sepharose (Amersham Biosciences), dialysed with PBSusing Membra-cel MD25 membrane, and concentrated using Amicon Ultracentrifugal filters (Millipore). Intracellular cytokine staining was performedusing the FoxP3 cytofix/cytoperm kit (BioLegend) as per the manufactur-er’s instructions.
Cell preparation
T-cell depletion of BM was performed as previously described,17 andresulting cell suspensions contained less than 1% contamination of CD3�
T cells. T cells were purified using magnetic bead depletion as previouslydescribed,17 and subsequent CD3� T-cell purities were greater than 80%.For in vitro assays, cell populations were purified using magnetic activatedcells sorting (MACS) beads (CD4, CD8, or CD11c) and positive selectioncolumns according to the manufacturer’s instructions (Miltenyi Biotec) andthen sorted on the basis of CD4�, CD8�, or MHC class II GFP and CD11cfor dendritic cell (DC) isolation on Moflo cell sorter (Beckman Coulter) tomore than 95% purity.
BMT
On day �1, mice received 1100 cGy (B6D2F1), 1000 cGy (B6 and bm1),or 900 cGy (BALB/c and C3H.SW) total body irradiation (137Cs source at108 cGy/min) split into 2 doses separated by 3 hours. On day 0, recipientswere transplanted with 107 BM cells with or without purified T cells fromB6.FoxP3-GFP donors (2 � 106 T cells for B6D2F1 recipients, 5 � 106
T cells for C3H.SW recipients, or 0.5 � 106 T cells for BALB/c recipients).To examine the effect of irradiation on CD8�FoxP3� Treg development,irradiated B6D2F1 recipients received 107 unfractionated B6.FoxP3-GFPsplenocytes on day 0, and a nonirradiated B6D2F1 cohort received 1 mganti-NK1.1 at day �2 and then 108 unfractionated B6.FoxP3-GFP spleno-cytes on day 0. For in vivo Treg tracking and depletion experiments,irradiated B6D2F1 recipients received B6.WT BM supplemented with0.5 � 106 each of sort purified CD4 and CD8 donor T cells from B6.WT,B6.FoxP3-luc� (tracking), or DEREG (depletion) donors, or combinationsthereof, as described in the figure legends. For depletion experiments, DT(100 �g intraperitoneally; Sigma-Aldrich) commenced on day 4 and wasadministered thrice weekly for the duration of the experiment. To determinethe cellular origin of CD8 Treg, irradiated B6D2F1 recipients receivedB6.WT BM and 1 � 106 unfractionated CD3� T cells from CD45.1�
B6.Ptprca donors supplemented with 1 � 106 unfractionated CD3� T,
1 � 106 sort purified CD4�GFP�, 2 � 105 sort purified CD4�GFP�, or1 � 106 sort purified CD8�GFP� T cells from B6.FoxP3-GFP donors. Fordepletion of host DCs, irradiated B6D2F1 and B6.CD11c.DOGxDBA2F1transplant recipients received daily injections (day �2 to day 3) of saline orDT (160 �g intraperitoneally; Sigma-Aldrich), respectively. For TGF-� andIL-10R blocking, recipients received control mAb (Mac49; 500 �g),anti-TGF (1D11; 500 �g), or anti–IL-10R (1B1.3a; 500 �g) on day 0 andday 2 of transplantation and donor FoxP3� Treg populations analyzed at day4. For in vivo expansion experiments, intraperitoneal injections of RAPA(1.5 mg/kg; Wyeth) were administered daily starting from day 0 aspreviously described.18 Recombinant murine IL-2 (rmIL-2, 1.5 �g; eBiosci-ence) was incubated for 30 minutes with the antibody JES6-1A12 (50 �g)to form IL-2 Ab complexes and injected intraperitoneally on day 0 andday 4 of transplantation. For experiments that compared cyclosporine toRAPA treatment, cyclosporine (50 mg/kg; Novartis) was administeredintraperitoneally daily as previously described.19
Assessment of GVHD
The degree of systemic GVHD was assessed using a cumulative scoringsystem that measures changes in 5 clinical parameters: weight loss, posture(hunching), activity, fur texture, and skin integrity (maximum index 10).Mice were monitored daily, and those with GVHD clinical scores20 greaterthan or equal to 6 were culled and the date of death recorded as the next dayin accordance with institutional animal ethics guidelines.
In vivo luminescence imaging
Recipients were injected with luciferin (0.5 mg) subcutaneously. Fiveminutes later, organs were harvested and imaged using the Xenogenimaging system (Xenogen IVIS 100; Caliper Life Sciences) to determineTreg trafficking. Data were analyzed with Living Image Version 4 software(Xenogen).
Cell labeling for proliferation analysis
For T-cell proliferation assays, purified T cells were suspended at3 � 107 cells/mL in serum-free media and CFSE (Sigma-Aldrich) or CellTrace violet proliferation dye (Invitrogen) added at 1 or 5�M finalconcentration, respectively. After incubation at 37°C for 10 to 20 minutes,cells were washed in media containing 2% FCS. Dye dilution was analyzedusing a FacsCanto or FACSCantoII (BD Biosciences).
T-cell suppression assays
For nonspecific suppression assays, CFSE labeled BioMag purifiedB6.PTPrca T cells were cultured at 50 000/well with either MACS-purifiedB6D2F1 or WT.B6 DCs (5000/well) and CD3 (1 �g/mL). For antigen-specific suppression assays, CFSE labeled sort-purified B6.Ptprca CD4� orCD8� T cells were cultured at 50 000/well with either MACS purifiedDBA/2 or C3H.Hej DC (5000/well). B6.FoxP3-GFP� Treg cells (10 000/well or as indicated in the figure legends) were sort purified from spleen,peripheral lymph node (pLN), and mesenteric lymph node (mLN) fromB6D2F1 recipients of B6.Foxp3-GFP BM and T cell grafts at day 7 aftertransplantation. Recipients received RAPA and IL-2 Ab complex treatmentin some experiments. Tissue culture supernatants were collected forcytokine analysis, and CFSE dilution of the B6.Ptprca T cells was assessedat 72 hours of culture for BioMag T responders, between 72 and 96 hoursfor purified CD8� responders and 120 hours for CD4� responders. Fortranswell suppression assays, transwell permeable supports (Corning LifeSciences) were used according to the manufacturer’s protocol. Briefly,B6.Ptprca CD4 T cells (300 000) and DBA/2 DC (30 000) were cultured inthe bottom of the plates, regulatory T cells (30 000) were added to either thebottom or the top of the wells, and the cultures processed as describedearlier in this paragraph.
To investigate DC costimulatory molecule expression, B6.MHC classII-GFPxDBA2F1 mice were used and DCs sorted based on GFP and CD11cexpression. B6.Ptprca sort purified CD4� T cells (300 000) were culturedwith 30 000 CD11c�GFP� DCs, with or without 30 000 sort purified aftertransplantation FoxP3�Treg. After 72-hour culture, CD11c�GFP� DCs were
CD8�FOXP3� ALLEVIATE GVHD 5899BLOOD, 14 JUNE 2012 � VOLUME 119, NUMBER 24

gated and assessed for costimulatory marker expression. To assess theability of CD8�FoxP3� Treg to kill DCs or responding effector T cells, CD4T cells (50 000; sort purified to 98%) and DCs (5000; MACS purified)from B6.WT, B6.b2m�/�, or B6.Bcl-2 mice were cultured with CD3(1 �g/mL) and 25 000 CD8�FoxP3� or CD4�FoxP3� Treg that wereisolated as described for nonspecific suppression assays.
Histopathology of GVHD target organs
At day 7 after transplantation, small bowel was harvested, formalin-preserved, embedded in paraffin, and processed to generate 5-�m-thicksections. H&E-stained sections of colon, small intestine, lung, and liverwere examined in a blinded fashion (by A.D.C.) using a semiquantitativescoring system for GVHD as previously published.21 Images were acquiredusing an Olympus BX51 microscope (Olympus) and Evolution MP Version5.0 Camera and QCapture software (QImaging).
Statistical analysis
Column graphs shown represent mean with error bars demonstrating theSEM. Statistical significance was determined using 2-tailed Mann-WhitneyU tests with a P value cut-off of .05. Survival curves were generated as
Kaplan-Meier estimates and compared by log-rank analysis. All statisticalanalyses were performed using Prism Version 5 software (GraphPad).
Results
Induction of a CD8�FoxP3� population after allogeneic BMT
To investigate Treg populations after transplantation, weanalyzed FoxP3.GFP expression in the spleen, mLN, and pLN atday 12 after BMT. We examined multiple different models ofGVHD to MHC and/or multiple minor histocompatibilityantigens (B6.FoxP3-GFP3 B6D2F1, B6.FoxP3-GFP3 BALB/c,B6.FoxP3-GFP3 C3H.SW) and compared this with that in synge-neic recipients (B6.FoxP3-GFP3 B6.WT). After allogeneic trans-plantation, a CD4�Foxp3� Treg population was evident in all sitesexamined. Surprisingly, we also observed a CD8�Foxp3� popula-tion in these BMT recipients (Figure 1A). Importantly, whereasCD4�Foxp3� Treg were also evident after syngeneic transplanta-tion, the CD8�Foxp3� population was only seen after allogeneic
Figure 1. CD8�FoxP3� differentiate after allogeneic transplantation from CD8�FoxP3� in the mLN. (A) After lethal irradiation at day �1, BM and T cells fromB6.FoxP3-GFP donors were transplanted into B6D2F1, BALB/c, C3H.SW, or C57Bl6 recipients. At day 12 after transplantation, CD4�FoxP3� and CD8�FoxP3� wereanalyzed in the spleen, pLN, and mLN. Representative plots of total live cells in the pLN are shown. Experiment was performed 3 times (n 10, 7, 7, 6). (B) After lethalirradiation at day �1, BM and T cells from B6.FoxP3-GFP donors were transplanted into B6D2F1 recipients. Spleens were stained for intracellular FoxP3 at day 5 aftertransplantation. (C) After lethal irradiation at day �1, BM and T cells were transplanted from PTPrca donors at day 0. Sorted cell populations (CD3�, CD4�FoxP3�,CD4�FoxP3�, or CD8�FoxP3�) from B6.FoxP3-GFP donors were also transplanted with the donor graft. At day 12 after transplantation, CD8�FoxP3� conversion wasanalyzed in the spleen and pLN. Representative plots of total live cells from peripheral LN are shown from 2 experiments; n 6 per group. (D) Recipients were transplantedand analyzed as in panel C; however, CD8�FoxP3� and CD4�FoxP3� were matched to that within the whole CD3� graft. Experiment was performed twice with resultsdisplayed as mean � SEM; n 6 per group. (E) At day 0, BM and T cells from B6.FoxP3-GFP donors were transplanted into lethally irradiated B6D2F1 recipients. At timepoints shown, spleen, and mLN were analyzed for CD4�FoxP3� and CD8�FoxP3�cells. Combined data from 2 experiments; n 6 per group. (F) After lethal irradiation on day�1, recipients were transplanted with the following grafts: (1) WT CD4� T cells and WT CD8� T cells, (2) WT CD4� T cells and B6.FoxP3-luc� CD8� T cells, (3) B6.FoxP3-luc�
CD4� T cells and WT CD8� T cells, or (4) B6.FoxP3-luc� CD4� T cells and B6.FoxP3-luc� CD8� T cells. Recipients were culled at day 4 after transplantation, 5 minutes afterluciferin injection, and the spleen, inguinal LN, liver, thymus, lung, and gut removed and imaged individually. Representative images from 2 experiments are shown (n 6 pergroup). (G) Mice were transplanted as in panel E; however, donor T cells were labeled with Cell Trace violet proliferation dye and mLN analyzed for CD8 Treg conversion.Representative plots gated on live donor CD8� T cells are shown, with numbers in graph showing mean � SEM.
5900 ROBB et al BLOOD, 14 JUNE 2012 � VOLUME 119, NUMBER 24
BMT (Figure 1A). Thus, an alloreactive T-cell response appearedto be required for the generation of this cell population. To confirmGFP expression as a true representation of FoxP3 expression,intracellular staining of FoxP3 was assessed on splenocytesisolated at day 5 after BMT. Results indicated that all GFP-expressing CD4� and CD8� T cells coexpressed FoxP3 (Figure1B). To determine the origin of the CD8�FoxP3�population,transplantations were performed where all recipients received abaseline graft containing B6 BM and T cells to induce GVHD, andin addition, sort-purified T-cell populations of B6.FoxP3-GFPtransgenic cells (whole CD3�, CD4�FoxP3�, CD4�FoxP3�, orCD8�FoxP3� only). When FoxP3� populations were analyzedafter transplantation, CD8�FoxP3� cells were observed only inthose groups that received whole CD3� or CD8�FoxP3� cellsoriginating from transgenic B6.FoxP3-GFP donors. Thus, a popula-tion of T cells covert to FoxP3� from the ingoing CD8�FoxP3�
population (Figure 1C). To specifically compare the contribution ofconversion of CD4�FoxP3�, recipients received either wholeCD3� T cells containing CD8�FoxP3�, CD4�FoxP3�, andCD4�FoxP3� fractions, or either CD4�FoxP3� or CD8�FoxP3�
cells only with numbers matched to that within the whole CD3�
graft. Results indicated that, within the spleen, conversion contrib-utes to approximately 30% of the total CD4�FoxP3� Treg pool atday 5 compared with 100% for CD8�FoxP3� (Figure 1D).
To determine the anatomic sites where CD8�FoxP3� cells wereemerging, time-course experiments were performed. As shown inFigure 1E, CD8�FoxP3� cells emerge by day 4 after transplanta-tion in the mLN. In contrast, the CD4�FoxP3� cells predominatelyemerge within the spleen after BMT (Figure 1E). Importantly,CD8�FoxP3� remained in higher numbers compared with
CD4�FoxP3� in the mLN during the first 10 days after transplanta-tion. To confirm the mLN as a key site for CD8�FoxP3� expansion,B6.FoxP3-luc� donors were used, in which luciferase expression isdriven off the FoxP3 promoter. Based on our previous dataindicating that all CD8�FoxP3� cells underwent conversion fromCD8�FoxP3� cells in the ingoing graft, we transplanted specificsubsets of T cells that resulted in all FoxP3�, CD8�FoxP3�, orCD4�FoxP3� cells only expressing the luciferase gene. Whenorgans were imaged at day 4 after transplantation, CD8�FoxP3�
were indeed present only in the mLN early after BMT, whereasCD4�FoxP3� were predominately in the spleen (Figure 1F). Todetermine which donor CD8�FoxP3� T cells were undergoingconversion into CD8�FoxP3� T cells, violet dye was used to trackproliferation. Interestingly, it was only the most actively proliferat-ing CD8� T cells that underwent conversion (Figure 1G). Thisconversion occurred very rapidly, with a 50-fold increase in theabsolute number of CD8�FoxP3� cells in the mLN within a24-hour period between day 3 and day 4 after transplantation(Figure 1G). This was not simply because of overall T-cellexpansion, as the CD8�FoxP3�:CD8�FoxP3� ratio decreased overtime (Figure 1G).
CD8�FoxP3� cells display a distinct phenotype and potentlysuppress antigen-specific responses
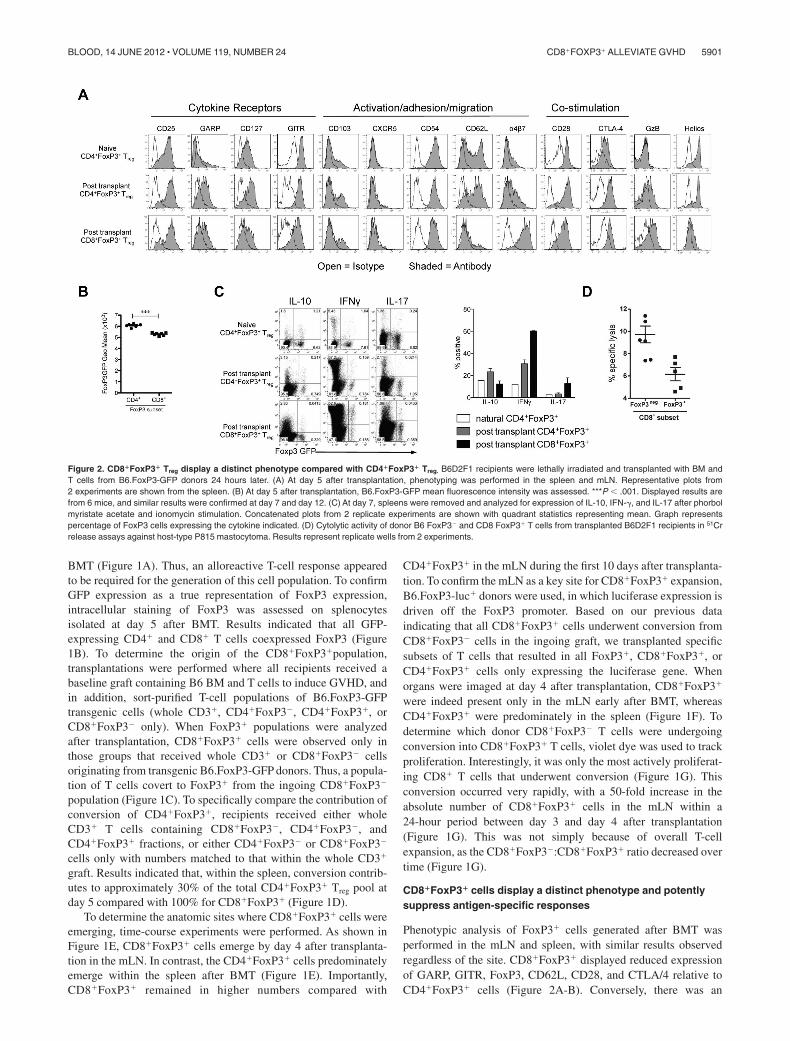
Phenotypic analysis of FoxP3� cells generated after BMT wasperformed in the mLN and spleen, with similar results observedregardless of the site. CD8�FoxP3� displayed reduced expressionof GARP, GITR, FoxP3, CD62L, CD28, and CTLA/4 relative toCD4�FoxP3� cells (Figure 2A-B). Conversely, there was an
Figure 2. CD8�FoxP3� Treg display a distinct phenotype compared with CD4�FoxP3� Treg. B6D2F1 recipients were lethally irradiated and transplanted with BM andT cells from B6.FoxP3-GFP donors 24 hours later. (A) At day 5 after transplantation, phenotyping was performed in the spleen and mLN. Representative plots from2 experiments are shown from the spleen. (B) At day 5 after transplantation, B6.FoxP3-GFP mean fluorescence intensity was assessed. ***P � .001. Displayed results arefrom 6 mice, and similar results were confirmed at day 7 and day 12. (C) At day 7, spleens were removed and analyzed for expression of IL-10, IFN-�, and IL-17 after phorbolmyristate acetate and ionomycin stimulation. Concatenated plots from 2 replicate experiments are shown with quadrant statistics representing mean. Graph representspercentage of FoxP3 cells expressing the cytokine indicated. (D) Cytolytic activity of donor B6 FoxP3� and CD8 FoxP3� T cells from transplanted B6D2F1 recipients in 51Crrelease assays against host-type P815 mastocytoma. Results represent replicate wells from 2 experiments.
CD8�FOXP3� ALLEVIATE GVHD 5901BLOOD, 14 JUNE 2012 � VOLUME 119, NUMBER 24

increase in expression of the integrin �4�7, a molecule critical inhoming to the GI tract. As expected, CD8�FoxP3� Treg did notexpress helios, a marker of thymic-derived CD4� Treg (Figure2A).22 In addition, the CD8�FoxP3� Treg did not express �- TCRor NK1.1 (data not shown), excluding the possibility they were anunusual gamma-delta, NK, or NKT cell. When cytokine productionwas investigated, CD8�FoxP3� Treg produced low levels of IL-10and IL-17, similar to CD4�FoxP3� Treg at the same time point(Figure 2C). They produced higher levels of IFN-� compared withCD4�FoxP3� Treg, but the ratio of cells producing IFN-� wassimilar to that seen within the CD8�FoxP3� compartment (Figure2C). Donor CD8 T cells are known to be highly cytolytic duringGVHD.23-29 However, the CD8�FoxP3� Treg did not exhibit highlevels of cytolytic activity against host targets relative toCD8�FoxP3� effectors (Figure 2D), suggesting that FoxP3 expres-sion and suppression are independent of cytolysis.
We next compared the capacity of the CD4�FoxP3�andCD8�FoxP3� cells to suppress proliferation in vitro. TheCD8�FoxP3� cells suppressed CD4� and CD8� T-cell prolifera-tion in response to Fc-bound CD3 to a greater extent thanCD4�FoxP3� Treg (Figure 3A). Whereas both FoxP3� populationssuppressed the generation of IFN-� by proliferating T cells, IL-10
was only produced in the presence of CD4�FoxP3� Treg (Figure3A). We next analyzed whether the suppressive activity ofCD8�FoxP3� Treg was antigen-specific. CD4� and CD8� Treg weresort purified after transplantation and cultured with CFSE-labeledB6.CD45.1� CD4� or CD8� responder T cells and either DBA/2DCs, which express host MHC (H-2dd) or C3H/Hej DC whichexpress a third party MHC (H-2dk). The CD4�FoxP3� Treg
suppressed proliferation similarly regardless of the stimulatingDCs, whereas CD8�FoxP3� Treg suppressed T-cell proliferation inan antigen-specific manner, preferentially in response to the DBA/2DC, with limited activity against third party C3H/Hej DC (Figure3B). Importantly, CD8�FoxP3� Treg displayed superior suppres-sive capacity against DBA/2 DCs compared with CD4�FoxP3�
Treg and both were superior to freshly isolated naive CD4�FoxP3�
(Figure 3B). Previous studies investigating naive CD4�FoxP3�
commonly use a 1:1 ratio of Treg compared with responding T cells.We have used a 1:5 ratio in these studies; thus, suppression by naiveCD4�FoxP3� Treg is limited. Importantly, suppressive capacitywithin donor CD8� cells was restricted to those expressing FoxP3(hereafter referred to as CD8�FoxP3� Treg), suggesting thatsuppression in the FoxP3� CD8 T cells was not merely aconsequence of cytotoxicity against DCs or responders mediated
Figure 3. CD8 Treg that undergo conversion after BMT exert contact-dependent, antigen-specific suppression and are more potent than CD4 Treg. B6D2F1 recipientswere lethally irradiated and transplanted with BM and T cells from B6.FoxP3-GFP donors 24 hours later. At day 7 after transplantation, LN and spleens were removed, andCD4�Foxp3�, CD8�Foxp3�, and CD8�Foxp3� cells sort purified. These cells were cultured with the following responding cells: (A) CFSE-labeled CD45.1�CD3� T cells,CD45.2� DCs, and 1 �g/mL CD3 with CFSE dilution quantified in CD45.1� cells 3 days later. IL-10 and IFN-� in the tissue culture supernatant were assessed. (B) SortedCFSE-labeled CD45.1�CD4 or CD45.1�CD8 T cells and CD45.2� recipient (DBA/2) or third party (C3H/Hej) DCs with CFSE dilution analyzed in CD45.1� CD8 or CD4responders. (C) CFSE-labeled CD45.1�CD3� T cells, CD45.2� DCs, and 1 �g/mL CD3 with CFSE dilution quantified in CD45.1� cells 3 days later. (D) Sorted CellTrace-labeled responder CD4� T cells and DCs from B6.WT, B6.�2m�/�, or B6.bcl-2 Tg mice with or without CD8�FoxP3� Treg as shown and violet dilution quantified 72 to96 hours later. (E) Sorted CFSE-labeled CD45.1�CD4� T cells and DBA/2 DC in transwell plates with CFSE dilution quantified in CD45.1� cells 3 days later. (F) FoxP3�
populations were sort purified and cultured with CD45.1�CD4� T cells and B6.MHC class II-GFPxDBA2.F1 DC. Costimulatory molecules were analyzed on GFP� DCs after72 hours of culture with histograms shown (open represents isotype; and tinted, costimulatory marker). For all experiments, representative plots are shown from 2 duplicate experiments.
5902 ROBB et al BLOOD, 14 JUNE 2012 � VOLUME 119, NUMBER 24

by effector CD8 T cells (Figure 3C). We confirmed this byperforming suppression assays where both the stimulating antigen-presenting cells and responding T cells were either lacking MHCclass I and so unable to be killed in an MHC class I–restrictedfashion (�2m�/�)30 or overexpressed bcl-2, making them resistantto death.31,32 There was no reduction in the ability of CD8�FoxP3�
Treg to suppress proliferation in either of these conditions (Figure3D), providing further evidence of true regulation by these cells. Tofurther investigate the mechanism of suppression used by Treg
generated after BMT, transwell assays were performed in which theDC-stimulated effector T cells were separated from theCD8�FoxP3� Treg, which suggested that, like CD4�FoxP3�,CD8�FoxP3� Treg require cell contact with the stimulating DCs tomediate their suppression (Figure 3E). In support of this, whenexpression of costimulatory markers on DCs were investigated, thepresence of either CD4�FoxP3� or CD8�FoxP3� Treg inhibitedup-regulation of CD40, CD80, and CD86 (Figure 3F).
Recipient DCs and TGF-� facilitate conversion of CD8 Treg
Because alloreactivity is required to drive CD8�FoxP3�conversion,we hypothesized that recipient antigen-presenting cells may playan important role in this process. To investigate this, DCs weredepleted by administering DT to transgenic B6.CD11c-
DOGxDBA2F1 recipients (in which the DT receptor is driven offthe CD11c promoter). After the depletion of recipient DCs, asignificant reduction in CD8�FoxP3� Treg conversion was evident(Figure 4A). However, no significant changes were observed inCD4�FoxP3� Treg in either the spleen or mLN at this early timepoint after transplantation (Figure 4A). To investigate whether CD4T-cell help is required for CD8�FoxP3� conversion, we used theB6.FoxP3-GFP3 bm1 system, where GVHD is directed towardan isolated MHC class I disparity. This allowed us to investigate theeffects of CD4 T cells in the donor graft, without having theirpresence or absence alter the development of GVHD. The absenceof donor CD4 T cells did not impact on the generation ofCD8�FoxP3� Treg (Figure 4B), suggesting that they are notrequired for conversion of this Treg population after transplantation.
To determine whether the proinflammatory environment associ-ated with myeloablative conditioning was contributing toCD8�FoxP3� Treg conversion, transplantations were performed inthe absence of conditioning. In this setting, CD8�FoxP3� Treg
conversion was still evident (Figure 4C). However, the condition-ing regimen did alter the contribution of CD8� or CD4� subsets tothe total FoxP3� pool, with the CD8�FoxP3� proportion increasedafter irradiation. To further elucidate which cytokines were in-volved in this conversion process, TGF-� and IL-10 were blocked
Figure 4. Host DCs and TGF-� are required for CD8 Treg conversion. (A) T cells from B6.FoxP3-GFP donors were labeled with Cell Trace violet dye and transplanted withBM into lethally irradiated B6.CD11c-DOGxDBA2.F1 recipients or B6D2F1 recipients. Recipients were treated with either DT or saline to deplete host DCs (160 ng/d,day �23 day 3). At day 4 after transplantation, CD4�Foxp3� and CD8�Foxp3� conversion and proliferation were analyzed in the spleen and mLN. Representative plots of themLN are shown, with numbers quantitated and displayed as mean � SEM. ** P � .01. Combined data from 2 experiments; n 6 per group. (B) BM and T cells (either CD8�
only or CD4� � CD8�) from B6.FoxP3-GFP doors were transplanted into lethally irradiated bm1 recipients. At day 12 after transplantation, CD8�FoxP3� Treg cells wereanalyzed in the spleen and mLN. Representative plots gated on total live cells are shown with combined data represented in the bar graphs displaying mean � SEM; n 3 pergroup. (C) B6D2F1 recipients received either irradiation at day �1 or NK1.1 at day �2 and splenocytes from B6.FoxP3-GFP donors at day 0. At day 5 after transplantation,CD8�FoxP3� populations were analyzed in the spleen and mLN. Representative plots gated on total live cells are shown, and percent contribution displayed as mean � SEMwith 3 per group. (D) Lethally irradiated B6D2F1 recipients were transplanted with BM and T cells from B6.FoxP3-GFP donors. Recipients were treated with antibodies againsteither TGF-� or IL-10R. At day 4 after transplantation, CD4�Foxp3� and CD8�Foxp3� conversion and proliferation were analyzed in the spleen and mLN. Representative plotsgated on CD8� T cells in the mLN are shown, with numbers quantitated and displayed as mean � SEM. Combined data from 2 experiments; n 6 per group. **P � .01,control antibody versus anti–TGF-�
CD8�FOXP3� ALLEVIATE GVHD 5903BLOOD, 14 JUNE 2012 � VOLUME 119, NUMBER 24

after transplantation (because of their known importance in CD4� Treg
conversion and function). These studies confirmed that TGF-�, but notIL-10, was required for the conversion of CD8�FoxP3� Treg (Figure4D). The conversion of CD4�FoxP3� Treg was not significantly reducedin the absence of either TGF-� or IL-10 signaling at this time point (datanot shown), consistent with the origin of these cells predominantly fromnatural Treg in the transplanted graft.
CD8�FoxP3� Treg inhibit GVHD
To determine the contribution of CD8�FoxP3�conversion tosurvival and GVHD severity after BMT, B6.DEREG mice wereused as donors. In these animals, GFP expression and the DTreceptor are driven of the FoxP3 promoter, allowing for specificdepletion of FoxP3 expressing cells after DT administration. Basedon previous data demonstrating that CD8�FoxP3� Treg undergoconversion from CD8�FoxP3� cells, combinations of donor cellswere transplanted as described in the figure legends that resulted indepletion of either CD4�FoxP3� or CD8�FoxP3� cells only. Asignificant increase in survival was noted in the presence of eitherCD4�FoxP3� Treg or CD8�FoxP3� Treg compared with recipientsin which all FoxP3� Treg were depleted (Figure 5A). In the DEREGmice, a rebound population of FoxP3-expressing cells that do notexpress GFP has been identified, although the suppressive capacityof this population has not been established.33 We thus undertookadoptive transfer experiments to confirm the suppressive functionof CD8�FoxP3� Treg and their ability to alleviate GVHD. In these
studies, CD8�FoxP3�Treg were transferred at day 0 with the donorBM in the B6.FoxP3-GFP3 BALB/c model to allow for expan-sion in the lymphopenic environment. Conventional donor T cellswere transferred 2 days later.34 GVHD was significantly alleviatedin recipients of CD8�FoxP3� Treg relative to recipients thatreceived no Treg or natural CD4�FoxP3� Treg (Figure 5B). NaturalCD4�FoxP3� Treg were not effective at suppressing GVHD inthese studies because of the low ratio of CD4�FoxP3� Treg
compared with conventional T cells (1:20). Importantly,CD8�FoxP3� Treg were still detected 14 days after transfer whentracked using luciferase-expressing Treg, indicating sustained FoxP3expression (Figure 5C). Thus, CD8�FoxP3� Treg are an importantand highly suppressive population generated after BMT.
RAPA and IL-2 Ab complex coadministration massivelyexpands CD8�FoxP3� Treg after BMT
Previous reports have demonstrated that the mTOR inhibitorrapamycin (RAPA) differentially suppresses CD4�FoxP3� andCD4�FoxP3� T cells after BMT.18 We thus investigated whetherFoxP3 expression by CD8� T cells would also render themrelatively resistant to suppression by RAPA. This was indeed thecase, with the proportion of CD8� T cells expressing FoxP3significantly increasing after RAPA because of the preferentialinhibition of CD8�FoxP3� T-cell expansion (Figure 6A). Further-more, RAPA administration in conjunction with recombinanthuman IL-2 expands CD4�FoxP3� Treg after transplantation35;
Figure 5. CD8�FoxP3� Treg attenuate GVHD mortality after BMT. (A) Recipient B6D2F1 mice were lethally irradiated and 24 hours later transplanted with BM fromB6.FoxP3-GFP mice and T cells from both B6.FoxP3-GFP and DEREG mice in the following combinations: (1) WT CD4� T cells and DEREG CD8� T cells, (2) DEREG CD4�
T cells and WT CD8� T cells, or (3) DEREG CD4� T cells and DEREG CD8� T cells. *P � .05, B6 (CD8� WT � CD4� DEREG) versus B6 (CD8� DEREG � CD4� DEREG).**P � .01, B6 (CD8� DEREG � CD4� WT) versus B6 (CD8� DEREG � CD4� DEREG). Survival data are shown and are combined from 4 duplicate experiments(CD4�FoxP3� deplete, n 42; CD8�FoxP3� deplete, n 39; CD4�FoxP3� and CD8�FoxP3� deplete, n 39; TCD no depletion, n 20). (B) BALB/c recipients werelethally irradiated and transplanted with BM and T cells from B6.FoxP3-GFP donors 24 hours later. At day 7 after transplantation, LN and spleens were removed, andCD4�Foxp3� and CD8�Foxp3� cells sorted on B6.FoxP3-GFP and CD4 or CD8. Sorted FoxP3� populations (3.5 � 104) were transplanted with BM from B6.WT donors intolethally irradiated BALB/c recipients. Whole CD3� T cells (7.5 � 105) were injected 2 days later. *P � .05, BALB/c (� CD8�FoxP3�) versus BALB/c (� CD4�FoxP3�).***P � .001, BALB/c versus BALB/c (� CD8�FoxP3�). Survival data are shown and are combined from 2 duplicate experiments (no FoxP3�, n 13; CD8�FoxP3�, n 17;CD4�FoxP3�, n 12; no FoxP3� TCD, n 6). (C) Recipients were transplanted as in panel B; however, CD8�Foxp3� were sorted from FoxP3.LuciDTR-4 donors (luciferase,GFP, and the DT receptor-driven of the FoxP3 promoter, labeled as B6.FoxP3-luc�). Recipients were imaged at days 2, 7, and 14 after transfer, with images shown contrastingrecipients that did or did not receive Treg.
5904 ROBB et al BLOOD, 14 JUNE 2012 � VOLUME 119, NUMBER 24

however, we found this strategy ineffective at expandingCD8�FoxP3� Treg (data not shown). We thus used IL-2 antibodycomplexes because this dramatically increases the biologic activityIL-2 in vivo.36 After treatment with RAPA and IL-2 complexesafter transplantation, the absolute numbers of CD8�FoxP3�
Treg were expanded 6- to 7-fold, with the percentage of CD8�
T cells expressing FoxP3 within the mLN increasing 30-fold to30% (Figure 6A). In contrast, the treatment of recipients withRAPA and IL-2 complexes only increased the percentage of CD4�
T cells expressing FoxP3 4-fold (1.2% � 0.25% vs 4.2% � 1.0%,P � .001). The CD8 Treg expansion was confirmed using CD8�
T cells from donors with luciferase, GFP, and the DT receptor-
driven of the FoxP3 promoter (FoxP3.LuciDTR-4, labeled asB6.FoxP3-luc�), allowing tracking of CD8�FoxP3� Treg. Atday 7 after transplantation, there was a significant increase inbioluminescence signal after RAPA and IL-2 complex treatment inthe GI tract, mLN, and spleen (Figure 6B). Importantly, thisoccurred in conjunction with a concurrent decrease in the effectorCD8� T-cell expansion in the same organs as assessed bytransferring CD8� T cells with luciferase expressed in all cells(Figure 6C). It is important to note that the bioluminescence signalis approximately 10-fold brighter in the B6.FoxP3-luc� donorsrelative to the B6.�-actin-luc� mice, meaning that luminescencecan only be compared when the cells of the same donor origin are
Figure 6. CD8�FoxP3� Treg are expanded after administration of RAPA and IL-2 Ab complexes after BMT. (A) Recipient B6D2F1 mice were lethally irradiated and24 hours later transplanted with BM and T cells from B6.FoxP3-GFP mice. Recipients received RAPA daily (1.5 mg/kg intraperitoneally) and/or IL-2 Ab complexes at day 0 andday 4 after transplantation in the combinations shown. At day 7, mLN were removed and analyzed for CD8�FoxP3� Treg conversion. Representative plots are shown for graphsin the lower panel displaying combined data from 2 duplicate experiments; n 8 per group. **P � .01. ***P � .005. B6D2F1 recipients were lethally irradiated and received BMand T cells from B6.WT donors combined with sorted CD8� T cells from either (B) FoxP3.LuciDTR-4 donors (luciferase, GFP, and the DT receptor-driven of the FoxP3promoter, labeled as B6.FoxP3-luc�; n 8 per group) or (C) B6.�-actin-luc� (n 3 or 4 per group) mice 24 hours later. RAPA and IL-2 Ab complex was administered as inpanel A. At day 7, recipients were killed 5 minutes after luciferin injection, and spleen, inguinal LN, liver, thymus, lung, and gut removed and imaged individually. Representativeimages from 2 duplicate experiments are shown with bioluminescence imaging quantitated and shown as mean � SEM. *P � .05. **P � .01. (D) Recipients were transplantedas in panel B with only the CD8� component from B6.FoxP3-luc� donors and imaged weekly from day 12. Images are shown with graph displaying mean � SEM. **P � .01,saline versus RAPA � IL-2 Ab complexes at each time point.
CD8�FOXP3� ALLEVIATE GVHD 5905BLOOD, 14 JUNE 2012 � VOLUME 119, NUMBER 24

transplanted. The long-term expansion of CD8�FoxP3� Treg wasconfirmed (Figure 6D) despite administration of only 2 doses ofIL-2 Ab complexes (the last on day 4). We confirmed that theRAPA and IL-2 complex expanded CD8�FoxP3� Treg retainedsuppressive function (Figure 7A). Finally, we undertook experi-ments whereby we could expand CD8�FoxP3� Treg with RAPAand IL-2 complexes and study the physiologic consequence of theirspecific deletion by DT administration in vivo. Depletion wasconfirmed by bioluminescence imaging as only the CD8�FoxP3�
in the ingoing graft were from B6.FoxP3-luc� donors (Figure 7B).As shown, GVHD within the GI tract in the animals receivingRAPA and IL-2 complexes was minimal, and specific deletion ofthe expanded CD8�FoxP3� Treg resulted in significant GVHD inthat organ (Figure 7C). Therefore, the combination of RAPA andIL-2 antibody complex treatment significantly expands theCD8�FoxP3� regulatory population and provides a potent therapeu-tic strategy to inhibit GVHD.
Lastly, an obvious and important question is whether conversion ofCD8�FoxP3� Treg occurs in clinical patients after BMT. We examinedthis in the peripheral blood of patients in the first 6 months after BMTbut disappointingly could not find convincing evidence of CD8�FoxP3�
Treg (data not shown). All these patients were receiving standard GVHDprophylaxis with cyclosporine or tacrolimus. We thus investigatedconversion of CD8�FoxP3� Treg in our murine systems in the context ofcyclosporine administration after BMT. Surprisingly, although a distinctpopulation of CD4�FoxP3� Treg remained in cyclosporine-treatedrecipients, CD8�FoxP3� Treg were absent (Figure 7D). Thus,CD8�FoxP3� Treg appear exquisitely sensitive to inhibition by cyclospor-ine, and the presence of this cell population will need to be examined in
patients receiving calcineurin-free (and ideally RAPA-based) immunesuppression early after BMT.
Discussion
In the current study, we identify and characterize a highlysuppressive CD8�FoxP3� regulatory T-cell population that isgenerated by a process of conversion after allogeneic BMT. Thisrepresents the first time that CD8�FoxP3� cells have been inducedafter physiologic uptake and presentation of endogenous antigen,although similar populations have been identified using artificialantigen presentation in transgenic models, strong adjuvants, ormodified CD3 antibody.37-39 CD8�FoxP3� Treg occur in negligiblenumbers in naive mice, which makes isolation and subsequentcharacterization of viable cells impossible.9
Naturally occurring CD8 regulatory T-cell populations havebeen identified on the basis of surface markers and have beenshown to be regulatory in several autoimmune diseases, includingsystemic lupus erythematosus,40 experimental autoimmune encepha-litis,41,42 and myasthenia gravis43 and to induce tolerance after solidorgan transplantation.44 However, these cells have not been demon-strated to express FoxP3 in vivo and are sometimes present withouthaving suppressive capacity. Furthermore, they have a somewhatcontroversial history, and interstudy comparisons are difficultbecause of the use of varied cell surface markers or properties(CD28�,44,45 CD122�,46 CD103�47 and Qa-1 restriction45,48,49) fortheir identification.
Figure 7. RAPA and IL-2 Ab complex expandedCD8�FoxP3� Treg are functional and protect from gutGVHD. (A) Recipient B6D2F1 mice were lethally irradi-ated and 24 hours later transplanted with BM and T cellsfrom B6.FoxP3-GFP mice. Recipients received RAPAdaily (1.5 mg/kg intraperitoneally) and IL-2 Ab complexesat day 0 and day 4 after transplantation. At day 7 aftertransplantation, LN and spleens were removed,CD8�Foxp3� cells sorted and cultured with CFSE-labeled CD45.1�CD3� T cells, CD45.2� DCs, and1 �g/mL CD3 with CFSE dilution quantified in CD45.1�
cells 3 days later. (B) B6D2F1 recipients were lethallyirradiated and received BM and 0.5 � 106 sortedCD4�FoxP3� T cells from B6.FoxP3-GFP donors inconjunction with 3 � 106 sorted CD8�FoxP3� cells fromFoxP3.LuciDTR-4 donors (luciferase, GFP, and the DTreceptor-driven of the FoxP3 promoter). All recipientsreceived RAPA (daily) and IL-2 Ab complexes (day 0 andday 4). DT (160 ng intraperitoneally) or saline wasadministered daily from day 3 until day 6. Bioluminescentimaging was performed and quantified at day 7; n 6per group. **P � .01. (C) Representative images of smallintestine taken at day 7 after transplantation (originalmagnification �200) and GVHD histopathology scores;n 6 per group. **P � .01. (D) B6D2F1 recipients werelethally irradiated and 24 hours later transplanted withBM and T cells from B6.FoxP3-GFP mice. Recipientsreceived RAPA or cyclosporine daily. Spleen and mLNwere removed at day 7 after transplantation and FoxP3�
populations analyzed. Representative plots are shown ofthe mLN for CD8� enumeration or the spleen for CD4�
enumeration. Absolute numbers were quantitated, andgraphs represent mean � SEM; n 7 or 8 per group.*P � .05. ***P � .005.
5906 ROBB et al BLOOD, 14 JUNE 2012 � VOLUME 119, NUMBER 24

Given that CD8�FoxP3� Treg are highly suppressive, it issurprising that they express lower levels of CTLA/4, GARP, andFoxP3, as these molecules contribute to CD4�FoxP3� Treg func-tion.50-53 Thus, these molecules appear less important in mediatingCD8�FoxP3� Treg suppression, suggesting differing pathways ofeffector function. The other phenotypic differences betweenCD8�FoxP3� Treg and both posttransplantation and naiveCD4�FoxP3� Treg were CD62L, �4�7, CD28, and GITR. The highexpression of �4�7 on CD8�FoxP3� Treg might be anticipatedgiven that �4�7 is involved in migration to the GI tract. Similarly,reduced expression of CD62L is consistent with the fact that thesecells are undergoing conversion from actively proliferatingCD8�FoxP3� T cells. The lower CD28 and GITR expression maybe advantageous for maintenance of suppressive function, as it hasbeen shown that TCR stimulation and CD28 ligation can breakanergy in CD4 Treg,54 whereas signaling of CD4�CD25� cellsthrough GITR also breaks tolerance.55
The use of CD4 Treg to suppress deleterious immune responsesresponsible for GVHD has been extensively investigated over theprevious decade. This initially occurred in preclinical models2,5 andhas now progressed to clinical trials involving CD4 Treg sortedstraight from the donor56 or alternatively ex vivo expanded CD4Treg.
57 These studies have yielded promising results with reducedincidence of acute GVHD and no effects on relapse or infection. Inthis study, we have clearly identified a regulatory T-cell populationthat is greater than 8 times more suppressive than the natural CD4Treg used in these trials. Because clinical GVHD is classicallyCD8� T cell and MHC class I dependent, the ability of theseCD8�FoxP3� Treg to suppress CD8� responses far more effectivelythan naive CD4�FoxP3� Treg would be expected to correlate withenhanced suppression of GVHD in vivo after adoptive transfer.
Despite the increased suppressive capacity of these cells, invitro or in vivo methods will still probably be necessary to furtherexpand CD8�FoxP3� Treg. Although CD8�FoxP3� cells have beenexpanded in vitro in an artificial system using ovalbumin asantigen, in the presence of IL-2, TGF-� and retinoic acid, resultsindicated that, in contrast to our results, these cells were not assuppressive as CD4�Foxp3� Treg.
58 A probable reason for this isthat the in vitro systems used to generate and expand these cells arenot optimal. In support of this, we have expanded these cells invitro under similar conditions and found that they lose suppressivecapacity (data not shown). Clearly, expansion protocols that areoptimal for CD4�Foxp3� Treg expansion are not effective forCD8�FoxP3� Treg, and the specific requirements for expansion ofthese cells in vitro needs further investigation.
When we investigated what was required for CD8�FoxP3� Treg
conversion in vivo, the process of BMT itself was not sufficient, asdemonstrated by lack of conversion after syngeneic transplanta-tion. Thus, because of the requirement for alloreactivity, it is notsurprising that recipient DCs significantly contribute to theirconversion. It has previously been demonstrated in models ofcolitis that specialized DCs, which express CD103 and produceTGF-� and retinoic acid, are involved in peripheral conversion ofCD4 Treg.
59 Because the mLN is the dominant site of conversion of
CD8�FoxP3� Treg, it is probable that this DC subset is involved atthis site. In conjunction with adoptive transfer of Treg populations toalleviate GVHD, using immune suppressant drugs that do notinhibit Treg expansion is also advantageous. Therefore, the ability ofRAPA to inhibit CD8�FoxP3� expansion while allowing conver-sion of FoxP3� to FoxP3� CD8 T cells after BMT provides furtherrationale for its use in the clinic. When IL-2 antibody complexeswere administered in conjunction with RAPA, CD8�FoxP3� Treg
were not only maintained but significantly increased after BMT.This in vivo manipulation of CD8�FoxP3� would appear prefer-able to in vitro expansion, as it does not require the expensive andtime-intensive processes required for the latter. Therefore, it islikely that, in conjunction with known mechanisms via whichRAPA alleviates GVHD, the maintenance of FoxP3 expression inCD8�FoxP3� Treg also contributes. The addition of IL-2 Abcomplexes to RAPA would be expected to further improve survivalbecause of the massive and preferential expansion of thisCD8�FoxP3� regulatory population.
In conclusion, we have identified a new regulatory population,provided a therapeutic approach to their expansion, and demon-strated their functional importance in defining transplant outcome.Importantly, this population appears as important for protectionfrom GVHD as the well-studied CD4�FoxP3� population. Giventhe potent MHC class I dominant suppressive function,CD8�FoxP3� cells represent a new regulatory population highlysuited to manipulation for the control of deleterious alloimmuneresponses after transplantation.
Acknowledgments
G.R.H. is a National Health and Medical Research CouncilAustralia Fellow and Queensland Health Senior Clinical ResearchFellow. K.A.M. is a National Health and Medical Research CouncilClinical Training Fellow. K.P.A.M. is a Cancer Council Queens-land Senior Research Fellow. R.J.R. is an LFQ PhD ScholarshipRecipient.
This work was supported by the Cancer Council Queenslandand the National Health and Medical Research Council Australia.
Authorship
Contribution: R.J.R. performed and designed experiments andwrote the manuscript; K.E.L., R.D.K., Y.A.W., N.C.R., S.D.O.,A.V., K.A.A., B.E.T., K.A.M., and M.K. performed experiments;G.J.H. provided FoxP3.LuciDTR mice; T.S. provided DEREGmice; A.D.C. performed histologic analysis; C.R.E. providedessential reagents; and G.R.H. and K.P.A.M. designed studies andhelped write the manuscript.
Conflict-of-interest disclosure: The authors declare no compet-ing financial interests.
Correspondence: Kelli P. A. MacDonald, Antigen Presentationand Immunoregulation Laboratory, 300 Herston Road, Herston,QLD, 4006, Australia; e-mail: [email protected].
References
1. Fontenot JD, Gavin MA, Rudensky AY. Foxp3programs the development and function ofCD4�CD25� regulatory T cells. Nat Immunol.2003;4(4):330-336.
2. Cohen JL, Trenado A, Vasey D, Klatzmann D,Salomon BL. CD4(�)CD25(�) immunoregulatoryT cells: new therapeutics for graft-versus-hostdisease. J Exp Med. 2002;196(3):401-406.
3. Hoffmann P, Ermann J, Edinger M, Fathman CG,Strober S. Donor-type CD4(�)CD25(�) regula-tory T cells suppress lethal acute graft-versus-host disease after allogeneic bone marrow trans-plantation. J Exp Med. 2002;196(3):389-399.
4. Taylor PA, Lees CJ, Blazar BR. The infusion of exvivo activated and expanded CD4(�)CD25(�)immune regulatory cells inhibits graft-versus-
host disease lethality. Blood. 2002;99(10):3493-3499.
5. Edinger M, Hoffmann P, Ermann J, et al.CD4�CD25� regulatory T cells preserve graft-versus-tumor activity while inhibiting graft-versus-host disease after bone marrow transplantation.Nat Med. 2003;9(9):1144-1150.
6. Feng G, Nadig SN, Backdahl L, et al. Functional
CD8�FOXP3� ALLEVIATE GVHD 5907BLOOD, 14 JUNE 2012 � VOLUME 119, NUMBER 24

regulatory T cells produced by inhibiting cyclicnucleotide phosphodiesterase type 3 prevent al-lograft rejection. Sci Transl Med. 2011;3(83):83ra40.
7. Hippen KL, Merkel SC, Schirm DK, et al. Massiveex vivo expansion of human natural regulatory Tcells (T(regs)) with minimal loss of in vivo func-tional activity. Sci Transl Med. 2011;3(83):83ra41.
8. Sagoo P, Ali N, Garg G, Nestle FO, Lechler RI,Lombardi G. Human regulatory T cells with al-loantigen specificity are more potent inhibitors ofalloimmune skin graft damage than polyclonalregulatory T cells. Sci Transl Med. 2011;3(83):83ra42.
9. Fontenot JD, Rasmussen JP, Williams LM,Dooley JL, Farr AG, Rudensky AY. Regulatory Tcell lineage specification by the forkhead tran-scription factor foxp3. Immunity. 2005;22(3):329-341.
10. Suffner J, Hochweller K, Kuhnle MC, et al. Den-dritic cells support homeostatic expansion ofFoxp3� regulatory T cells in Foxp3.LuciDTRmice. J Immunol. 2010;184(4):1810-1820.
11. Lahl K, Loddenkemper C, Drouin C, et al. Selec-tive depletion of Foxp3� regulatory T cells in-duces a scurfy-like disease. J Exp Med. 2007;204(1):57-63.
12. Ogilvy S, Metcalf D, Print CG, Bath ML,Harris AW, Adams JM. Constitutive Bcl-2 expres-sion throughout the hematopoietic compartmentaffects multiple lineages and enhances progenitorcell survival. Proc Natl Acad Sci U S A. 1999;96(26):14943-14948.
13. Hochweller K, Striegler J, Hammerling GJ,Garbi N. A novel CD11c.DTR transgenic mousefor depletion of dendritic cells reveals their re-quirement for homeostatic proliferation of naturalkiller cells. Eur J Immunol. 2008;38(10):2776-2783.
14. Banovic T, MacDonald KP, Morris ES, et al. TGF-beta in allogeneic stem cell transplantation: friendor foe? Blood. 2005;106(6):2206-2214.
15. Kuns RD, Morris ES, Macdonald KP, et al. Invari-ant natural killer T cell-natural killer cell interac-tions dictate transplantation outcome after alpha-galactosylceramide administration. Blood. 2009;113(23):5999-6010.
16. Grodzki AC, Berenstein E. Antibody purification:ammonium sulfate fractionation or gel filtration.Methods Mol Biol. 2010;588:15-26.
17. Burman AC, Banovic T, Kuns RD, et al. IF-Ngamma differentially controls the developmentof idiopathic pneumonia syndrome and GVHD ofthe gastrointestinal tract. Blood. 2007;110(3):1064-1072.
18. Zeiser R, Leveson-Gower DB, Zambricki EA,et al. Differential impact of mammalian target ofrapamycin inhibition on CD4�CD25�Foxp3�regulatory T cells compared with conventionalCD4� T cells. Blood. 2008;111(1):453-462.
19. Zeiser R, Nguyen VH, Beilhack A, et al. Inhibitionof CD4�CD25� regulatory T-cell function by cal-cineurin-dependent interleukin-2 production.Blood. 2006;108(1):390-399.
20. Cooke KR, Kobzik L, Martin TR, et al. An experi-mental model of idiopathic pneumonia syndromeafter bone marrow transplantation: I. The roles ofminor H antigens and endotoxin. Blood. 1996;88:3230-3239.
21. Hill GR, Crawford JM, Cooke KJ, Brinson YS,Pan L, Ferrara JLM. Total body irradiation andacute graft versus host disease: the role of gas-trointestinal damage and inflammatory cytokines.Blood. 1997;90:3204-3213.
22. Thornton AM, Korty PE, Tran DQ, et al. Expres-sion of Helios, an Ikaros transcription factor fam-ily member, differentiates thymic-derived fromperipherally induced Foxp3� T regulatory cells.J Immunol. 2010;184(7):3433-3441.
23. Hill GR, Teshima T, Gerbitz A, et al. Differentialroles of IL-1 and TNF-alpha on graft-versus-host
disease and graft versus leukemia. J Clin Invest.1999;104(4):459-467.
24. Hill GR, Teshima T, Rebel VI, et al. The p55 TNF-alpha receptor plays a critical role in T cell allore-activity. J Immunol. 2000;164(2):656-663.
25. Markey KA, Burman AC, Banovic T, et al. Solublelymphotoxin is an important effector molecule inGVHD and GVL. Blood. 2010;115(1):122-132.
26. Morris ES, MacDonald KP, Kuns RD, et al. Induc-tion of natural killer T cell-dependent alloreactivityby administration of granulocyte colony-stimulat-ing factor after bone marrow transplantation. NatMed. 2009;15(4):436-441.
27. Morris ES, MacDonald KP, Rowe V, et al. NKTcell-dependent leukemia eradication followingstem cell mobilization with potent G-CSF ana-logs. J Clin Invest. 2005;115(11):3093-3103.
28. Robb RJ, Kreijveld E, Kuns RD, et al. Type I-IFNscontrol GVHD and GVL responses after trans-plantation. Blood. 2011;118(12):3399-3409.
29. Teshima T, Hill GR, Pan L, et al. IL-11 separatesgraft-versus-leukemia effects from graft-versus-host disease after bone marrow transplantation.J Clin Invest. 1999;104(3):317-325.
30. Liao NS, Bix M, Zijlstra M, Jaenisch R, Raulet D.MHC class I deficiency: susceptibility to naturalkiller (NK) cells and impaired NK activity. Science.1991;253(5016):199-202.
31. Strasser A, Harris AW, Cory S. bcl-2 transgeneinhibits T cell death and perturbs thymic self-cen-sorship. Cell. 1991;67(5):889-899.
32. Schroter M, Lowin B, Borner C, Tschopp J. Regu-lation of Fas(Apo-1/CD95)- and perforin-medi-ated lytic pathways of primary cytotoxic T lympho-cytes by the protooncogene bcl-2. Eur J Immunol.1995;25(12):3509-3513.
33. Lahl K, Sparwasser T. In vivo depletion ofFoxP3� Tregs using the DEREG mouse model.Methods Mol Biol. 2011;707:157-172.
34. Nguyen VH, Zeiser R, Dasilva DL, et al. In vivodynamics of regulatory T-cell trafficking and sur-vival predict effective strategies to control graft-versus-host disease following allogeneic trans-plantation. Blood. 2007;109(6):2649-2656.
35. Shin HJ, Baker J, Leveson-Gower DB, Smith AT,Sega EI, Negrin RS. Rapamycin and IL-2 reducelethal acute graft-versus-host disease associatedwith increased expansion of donor typeCD4�CD25�Foxp3� regulatory T cells. Blood.2011;118(8):2342-2350.
36. Boyman O, Kovar M, Rubinstein MP, Surh CD,Sprent J. Selective stimulation of T cell subsetswith antibody-cytokine immune complexes. Sci-ence. 2006;311(5769):1924-1927.
37. Haribhai D, Lin W, Relland LM, Truong N,Williams CB, Chatila TA. Regulatory T cells dy-namically control the primary immune responseto foreign antigen. J Immunol. 2007;178(5):2961-2972.
38. Bisikirska B, Colgan J, Luban J, Bluestone JA,Herold KC. TCR stimulation with modified anti–CD3 mAb expands CD8� T-cell population andinduces CD8�CD25� Tregs. J Clin Invest. 2005;115(10):2904-2913.
39. Fleissner D, Hansen W, Geffers R, Buer J,Westendorf AM. Local induction of immunosup-pressive CD8� T cells in the gut-associated lym-phoid tissues. PLoS One. 2010;5(10):e15373.
40. Sharabi A, Mozes E. The suppression of murinelupus by a tolerogenic peptide involves foxp3-expressing CD8 cells that are required for the op-timal induction and function of foxp3-expressingCD4 cells. J Immunol. 2008;181(5):3243-3251.
41. Koh DR, Fung-Leung WP, Ho A, Gray D,Acha-Orbea H, Mak TW. Less mortality but morerelapses in experimental allergic encephalomyeli-tis in CD8�/� mice. Science. 1992;256(5060):1210-1213.
42. Jiang H, Zhang SI, Pernis B. Role of CD8� T
cells in murine experimental allergic encephalo-myelitis. Science. 1992;256(5060):1213-1215.
43. Ben-David H, Sharabi A, Dayan M, Sela M,Mozes E. The role of CD8�CD28 regulatory cellsin suppressing myasthenia gravis-associated re-sponses by a dual altered peptide ligand. ProcNatl Acad Sci U S A. 2007;104(44):17459-17464.
44. Chang CC, Ciubotariu R, Manavalan JS, et al.Tolerization of dendritic cells by T(S) cells: thecrucial role of inhibitory receptors ILT3 and ILT4.Nat Immunol. 2002;3(3):237-243.
45. Najafian N, Chitnis T, Salama AD, et al. Regula-tory functions of CD8�CD28� T cells in an auto-immune disease model. J Clin Invest. 2003;112(7):1037-1048.
46. Rifa’i M, Kawamoto Y, Nakashima I, Suzuki H.Essential roles of CD8�CD122� regulatory Tcells in the maintenance of T cell homeostasis.J Exp Med. 2004;200(9):1123-1134.
47. Uss E, Rowshani AT, Hooibrink B, Lardy NM,van Lier RA, ten Berge IJ. CD103 is a marker foralloantigen-induced regulatory CD8� T cells.J Immunol. 2006;177(5):2775-2783.
48. Hu D, Ikizawa K, Lu L, Sanchirico ME,Shinohara ML, Cantor H. Analysis of regulatoryCD8 T cells in Qa-1-deficient mice. Nat Immunol.2004;5(5):516-523.
49. Tang X, Maricic I, Purohit N, et al. Regulation ofimmunity by a novel population of Qa-1-restrictedCD8alphaalpha�TCRalphabeta� T cells. J Im-munol. 2006;177(11):7645-7655.
50. Friedline RH, Brown DS, Nguyen H, et al. CD4�regulatory T cells require CTLA-4 for the mainte-nance of systemic tolerance. J Exp Med. 2009;206(2):421-434.
51. Wang R, Kozhaya L, Mercer F, Khaitan A, Fujii H,Unutmaz D. Expression of GARP selectivelyidentifies activated human FOXP3� regulatory Tcells. Proc Natl Acad Sci U S A. 2009;106(32):13439-13444.
52. Probst-Kepper M, Geffers R, Kroger A, et al.GARP: a key receptor controlling FOXP3 in hu-man regulatory T cells. J Cell Mol Med. 2009;13(9B):3343-3357.
53. Wing K, Onishi Y, Prieto-Martin P, et al. CTLA-4control over Foxp3� regulatory T cell function.Science. 2008;322(5899):271-275.
54. Itoh M, Takahashi T, Sakaguchi N, et al. Thymusand autoimmunity: production of CD25�CD4�naturally anergic and suppressive T cells as a keyfunction of the thymus in maintaining immuno-logic self-tolerance. J Immunol. 1999;162(9):5317-5326.
55. Shimizu J, Yamazaki S, Takahashi T, Ishida Y,Sakaguchi S. Stimulation of CD25(�)CD4(�)regulatory T cells through GITR breaks immuno-logical self-tolerance. Nat Immunol. 2002;3(2):135-142.
56. Di Ianni M, Falzetti F, Carotti A, et al. Tregs pre-vent GVHD and promote immune reconstitutionin HLA-haploidentical transplantation. Blood.2011;117(14):3921-3928.
57. Brunstein CG, Miller JS, Cao Q, et al. Infusion ofex vivo expanded T regulatory cells in adultstransplanted with umbilical cord blood: safety pro-file and detection kinetics. Blood. 2011;117(3):1061-1070.
58. Mayer CT, Floess S, Baru AM, Lahl K, Huehn J,Sparwasser T. CD8� Foxp3� T cells share de-velopmental and phenotypic features with classi-cal CD4� Foxp3� regulatory T cells but lack po-tent suppressive activity. Eur J Immunol. 2011;41(3):716-725.
59. Coombes JL, Siddiqui KR, Arancibia-Carcamo CV,et al. A functionally specialized population of mucosalCD103� DCs induces Foxp3� regulatory T cells viaa TGF-beta and retinoic acid-dependent mechanism.J Exp Med. 2007;204(8):1757-1764.
5908 ROBB et al BLOOD, 14 JUNE 2012 � VOLUME 119, NUMBER 24




















