
Growth patterns of Candida albicans in relation to radicular dentin
Upload
independentCategory
view
0download
0
Hyphal Development in Candida albicans Requires TwoTemporally Linked Changes in Promoter Chromatin forInitiation and MaintenanceYang Lu, Chang Su, Allen Wang, Haoping Liu*
Department of Biological Chemistry, University of California, Irvine, California, United States of America
Abstract
Phenotypic plasticity is common in development. For Candida albicans, the most common cause of invasive fungalinfections in humans, morphological plasticity is its defining feature and is critical for its pathogenesis. Unlike other fungalpathogens that exist primarily in either yeast or hyphal forms, C. albicans is able to switch reversibly between yeast andhyphal growth forms in response to environmental cues. Although many regulators have been found involved in hyphaldevelopment, the mechanisms of regulating hyphal development and plasticity of dimorphism remain unclear. Here weshow that hyphal development involves two sequential regulations of the promoter chromatin of hypha-specific genes.Initiation requires a rapid but temporary disappearance of the Nrg1 transcriptional repressor of hyphal morphogenesis viaactivation of the cAMP-PKA pathway. Maintenance requires promoter recruitment of Hda1 histone deacetylase underreduced Tor1 (target of rapamycin) signaling. Hda1 deacetylates a subunit of the NuA4 histone acetyltransferase module,leading to eviction of the NuA4 acetyltransferase module and blockage of Nrg1 access to promoters of hypha-specificgenes. Promoter recruitment of Hda1 for hyphal maintenance happens only during the period when Nrg1 is gone. Thesequential regulation of hyphal development by the activation of the cAMP-PKA pathway and reduced Tor1 signalingprovides a molecular mechanism for plasticity of dimorphism and how C. albicans adapts to the varied host environments inpathogenesis. Such temporally linked regulation of promoter chromatin by different signaling pathways provides a uniquemechanism for integrating multiple signals during development and cell fate specification.
Citation: Lu Y, Su C, Wang A, Liu H (2011) Hyphal Development in Candida albicans Requires Two Temporally Linked Changes in Promoter Chromatin forInitiation and Maintenance. PLoS Biol 9(7): e1001105. doi:10.1371/journal.pbio.1001105
Academic Editor: Joseph Heitman, Duke University Medical Center, United States of America
Received December 7, 2010; Accepted May 31, 2011; Published July 19, 2011
Copyright: � 2011 Lu et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricteduse, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding: This work was supported by the US National Institutes of Health grant R01GM/AI55155 to HL. HL was also supported in part by the NIH NationalInstitute of General Medical Sciences Center grant GM76516. The funders had no role in study design, data collection and analysis, decision to publish, orpreparation of the manuscript.
Competing Interests: The authors have declared that no competing interests exist.
Abbreviations: ChIP, chromatin immunoprecipitation; HAT, histone acetyltransferase; HDAC, histone deacetylase; PKA, protein kinase A; TOR, target ofrapamycin
* E-mail: [email protected]
Introduction
Many organisms or cells are able to alter their phenotype or
developmental fate in response to changes in their environment, a
phenomenon referred to as plasticity. Plasticity is seen in a broad
range of biological systems from embryo development to memory
formation where long-lasting effects on gene expression outlive an
initial transient signal. The dynamic process of cell fate
specification is determined by a network of regulatory genes.
The architecture of the network defines the temporal order of
specification events. Therefore, understanding temporal dynamic
regulation of gene expression in response to extracellular signals is
critical for our comprehension of cell fate specification and
plasticity.
Morphological plasticity is a defining feature of Candida albicans,
a major opportunistic fungal pathogen of humans [1]. C. albicans
resides as harmless commensal flora in the gastrointestinal tract
and mucosal membranes of healthy individuals, but when the host
immune system is suppressed, the fungus can disseminate and
cause systemic infections. Unlike many other pathogenic fungi that
exist primarily in either yeast or hyphal forms and infect a specific
organ, C. albicans is able to undergo reversible morphological
changes between yeast, pseudohyphal, and hyphal forms of growth
in response to environmental cues and can successfully infect many
different anatomical sites of the human host. Its morphological
plasticity is the most important virulence attribute of C. albicans [2].
Hyphae have invasive properties that can promote tissue
penetration and escape from immune cells, whereas yeast cells
are suited for dissemination in the bloodstream. Several of the
genes that are specifically expressed in hyphae encode virulence
factors. For example, HWP1, ALS3, and RBT5 encode cell wall
proteins that are important for adhesion to host cells and iron
acquisition from the host [3–6].
Several signal transduction pathways are involved in the
regulation of hyphal development. Among them, the cAMP-
dependent protein kinase A (PKA) pathway plays an essential role
in hyphal morphogenesis and virulence [7]. The adenylate cyclase
Cyr1 and its associated protein are indispensable for hyphal
growth under all conditions [8–10]. Tpk1 and Tpk2 are catalytic
subunits of PKA; each plays distinct functions in hyphal
development [11–13]. Efg1 and Flo8, two transcription regulators
essential for hyphal development and virulence [14–16], are
PLoS Biology | www.plosbiology.org 1 July 2011 | Volume 9 | Issue 7 | e1001105

implicated to function downstream of the cAMP-PKA pathway
[17,18]. The hyphal transcriptional program is repressed by Tup1
through sequence-specific DNA-binding proteins [19–27], of
which Nrg1 plays a major role. nrg1 mutant cells are constitutively
filamentous under all conditions, similar to tup1 cells [25,26].
Ectopic expression of NRG1 inhibits hyphal filamentation in all
growth conditions [28,29]. Although molecular genetic analyses
have identified a number of key transcriptional regulators of
hyphal morphogenesis, our understanding of the transcriptional
regulation that governs the yeast-to-hypha transition remains
rudimentary.
The yeast-to-hypha transition can be induced by a wide range
of media and environmental conditions in vitro [1]. Serum in
combination with a rise in temperature to 37uC gives the most
robust induction of hyphae. Simple inoculation of stationary cells
into fresh medium at 37uC is also a powerful but transient inducer
of hyphae [30]. Several induction signals are transmitted through
Cyr1, including CO2/HCO32 and peptidoglycan found in serum,
as well as a rise in temperature [31–33]. Cyr1 is also regulated by
Ras1, Ras2, and Gpa2 in response to nutrients [34–38]. Farnesol,
a quorum-sensing molecule secreted to the medium by C. albicans
cells as a cell density signal [39], exerts its inhibitory effects on
germ-tube formation through Ras1-Cyr1 [40]. However, many
widely used hyphal-inducing media are poor in nitrogen and
carbon sources that are not favorable for the activation of the
cAMP-PKA pathway, raising the question of how C. albicans can
undergo hyphal development in both rich and poor media. The
target of rapamycin (TOR) protein kinase pathway is another
major nutrient-sensing pathway conserved in C. albicans [41].
Rapamycin can both inhibit hyphal formation on solid medium
and promote cell aggregation in liquid medium in C. albicans
[41,42], but it does not induce yeast-to-hyphal development.
Transcriptional activation or repression in response to external
stimuli is often mediated through dynamic changes in promoter
chromatin structure. This regulation occurs at many different
levels, including posttranslational modification of histones [43],
chromatin remodeling [44], incorporation of histone variants [45],
and cotranscriptional chromatin disassembly and assembly
[46,47]. All of these dynamic processes work in concert to
establish or alter the local properties of chromatin, although the
relative importance and order of these processes vary at each
individual promoter. Histone acetyltransferases (HATs) and
deacetylases (HDACs) play important roles in regulating chroma-
tin structure and transcription [48]. Histone acetylation is a
dynamic reversible process, and the balance of histone acetylation
is important for proper cellular functions [49]. We have previously
shown a dynamic nucleosomal H4 acetylation at the promoters of
hypha-specific genes during hyphal induction [50]. H4 histone
acetyltransferase complex NuA4 is recruited to promoters in both
yeast and hyphal forms and is required for the induction of hyphal
genes [50]. Here we show that the dynamic increase and decrease
in H4 acetylation at the promoters during hyphal induction
correlates with the sequential dissociation and association of two
different HDACs, Rpd3 and Hda1, respectively. Concomitant
with the dissociation of Rpd3 upon hyphal initiation, Nrg1 protein
disappears from cells in response to the activation of the cAMP-
PKA pathway, a step essential for hyphal initiation. Although
Nrg1 protein returns after hyphal initiation, promoter recruitment
of Hda1 under reduced Tor1 signaling leads to deacetylation and
eviction of the NuA4 HAT module, nucleosome reassembly, and
inhibition of Nrg1 access to the promoters of hypha-specific genes,
a step essential for sustained hyphal development. Importantly,
disappearance of Nrg1 during hyphal initiation is a prerequisite for
promoter recruitment of Hda1. The sequential regulation of
hyphal development by the activation of the cAMP-PKA pathway
and reduced Tor1 signaling provides a molecular mechanism for
the plasticity of dimorphism and how C. albicans adapts to the
varied host environments in pathogenesis.
Results
Yeast-to-Hyphal Development Involves Two TemporallyRelated Phases of Removing Nrg1 Inhibition
The transcription factor Nrg1 plays an essential role in
repressing hyphal development [22,25,26,51]. The significance
of Nrg1 is underscored by recent phenotypic profiling of 143
transcriptional regulator knockout mutants, where only nrg1 and
tup1 mutants are filamentous under all conditions examined [52].
Relief of the transcriptional repression by Nrg1-Tup1 may be the
essential and regulated step for the activation of the hyphal
transcriptional program under all conditions. Indeed, NRG1
transcript level has been shown to be reduced in hyphae
[22,25,26,53]. However, how Nrg1 is regulated at the protein
level and whether Nrg1 directly binds to the promoters of hypha-
specific genes is not clear. By chromatin immunoprecipitation
(ChIP) of C-terminal Myc-tagged Nrg1, we found that Nrg1 was at
the promoters of hypha-specific genes HWP1, ALS3, and ECE1
during yeast growth (Figure 1). We next analyzed the levels of
promoter-bound Nrg1 during yeast to hyphal morphological
development under four related conditions in YPD medium that
induce three distinctive developmental responses (Figure 1A). Cells
inoculated to YPD with or without serum at 30uC remain in the
yeast growth form. Cells inoculated into YPD at 37uC in the
absence of serum show normal germ tube formation, but are
unable to maintain hyphal development. Serum, in combination
with a shift in temperature to 37uC, induces robust and sustained
hyphal growth. By a time course ChIP, we found that, upon
hyphal induction, Nrg1 dissociated rapidly from the promoters
and remained at low levels during hyphal elongation in YPD with
serum (Figure 1B). In contrast, levels of promoter-associated Nrg1
Author Summary
Many organisms are able to change their phenotype inresponse to changes in the environment, a phenomenonreferred to as plasticity. Candida albicans, a majoropportunistic fungal pathogen of humans, can undergoreversible morphological changes between yeast (spheri-cal) and hyphal (filamentous) forms of growth in responseto environmental cues. This morphological plasticity isessential for its pathogenesis and survival in its hosts. Inthis study, we show that hyphal development is initiatedand maintained by two major nutrient-sensing cellulargrowth pathways that act by removing the inhibitionprovided by the transcriptional repressor Nrg1. Whileinitiation requires a rapid but temporary disappearance ofNrg1 via activation of the cAMP-dependent protein kinaseA pathway, maintenance requires the recruitment topromoters of the Hda1 histone deacetylase under condi-tions of reduced signaling by the target of rapamycin(TOR) kinase, leading to chromatin remodeling that blocksNrg1 access to the promoters of hypha-specific genes. Weobserved that recruitment of Hda1 to promoters happensonly during the time window when Nrg1 is absent. Suchtemporally linked regulation of promoter chromatin bydifferent signaling pathways provides a unique mechanismfor integrating multiple signals in the regulation of geneexpression and phenotypic plasticity during developmentand cell fate specification.
Plasticity of Dimorphism
PLoS Biology | www.plosbiology.org 2 July 2011 | Volume 9 | Issue 7 | e1001105

Figure 1. Initiation and maintenance of hyphal development by removing Nrg1 inhibition. An overnight culture of wild-type cellscarrying Nrg1-Myc (HLY3922) were diluted 1:50 into the indicated medium at 30uC or 37uC in the presence or absence of 10% serum, and cells werecollected at 0 min, 15 min, 30 min, 1 h, 2 h, 3 h, and 5 h for cell morphology (A), ChIP (B), and Western blot (C) experiments. ChIP DNA werequantitated by qPCR with primers at the UAS regions of HWP1, ALS3, and ECE1, as described [50]. The ADE2 coding region was used as a control. TheNrg1-Myc enrichment is presented as a ratio of HWP1, ALS3, or ECE1 IP (bound/input) versus the control IP (bound/input). The 0 h values on HWP1were set to 1.00. The ChIP data show the average of three independent qPCR data with error bars representing the SEM. Western analysis was carriedout using a peroxidase-conjugated anti-c-myc (Roche Diagnostics) antibody to assess levels of Nrg1-myc, and with an anti-PSTAIRE (Cdc28, SantaCruz) antibody for loading control.doi:10.1371/journal.pbio.1001105.g001
Plasticity of Dimorphism
PLoS Biology | www.plosbiology.org 3 July 2011 | Volume 9 | Issue 7 | e1001105

recovered after the initial decrease under the condition of YPD
without serum, which correlated with the conversion to the yeast
growth phase around 3 h after the induction (Figure 1A and 1B).
Unexpectedly, Western analyses indicated that Nrg1 protein levels
decreased sharply at 15 and 30 min upon hyphal induction at 37uCcoinciding with germ tube formation. However, the Nrg1 level
recovered after 1 h of hyphal induction (Figure 1C). Serum was not
required for the disappearance of Nrg1, but was critical for
sustained hyphal development and excluding Nrg1 from promoters
during hyphal elongation (Figure 1). The Western analysis of cells
after 3 h of hyphal induction could not discriminate Nrg1 protein
from apical cells and subapical cells. It is possible that Nrg1 protein
was absent in the apical cell of each hypha and, therefore, allowed
sustained hyphal growth from the apical cells. We determined Nrg1
localization by immunofluorescence in hyphal cells after 5 h of
growth in YPD with serum at 37uC and found Nrg1 in the nucleus
of both apical and subapical cells (Figure S1), excluding this
possibility. Our data suggest that hyphal development involves two
phases of releasing Nrg1 inhibition: the first for initiation and the
second for maintenance. We suggest that initiation requires a
transient down-regulation of Nrg1 protein level, while maintenance
requires a regulation that prevents Nrg1 from binding at the
promoters of hypha-specific genes.
The cAMP-PKA Pathway Is Required for theDown-Regulation of Nrg1 for Hyphal Initiation
Several lines of evidence suggest that the rapid and temporary
disappearance of Nrg1 protein upon hyphal induction is regulated
by the cAMP-PKA pathway. First, we found that Nrg1
disappearance required the cAMP-PKA pathway as Nrg1 protein
level did not show an obvious reduction in cyr1 and tpk2 mutants
(Figure 2A). Similarly, no obvious change in Nrg1 level was
observed in an efg1 or flo8 mutant (Figure 2A). Second, the shift to
37uC during hyphal induction was critical for the observed Nrg1
disappearance (Figure 1C). This is consistent with the report that
Hsp90 regulates hyphal development via Cyr1 in response to a
shift in temperature [33]. Third, the major quorum-sensing
molecule farnesol, at a physiological concentration, completely
blocked the down-regulation of Nrg1 (Figure 2A). Farnesol is
reported to exert its inhibitory effects on hyphal initiation through
Cyr1 [39,40]. Consistent with the inhibitory effect of farnesol on
the down-regulation of Nrg1, different folds of dilution during
inoculation could lead to different durations of Nrg1 down-
regulation (Figure 1C versus Figure 2A). We also observed that
hyphal initiation correlated with Nrg1 disappearance in all liquid
media, including nutrient-poor media (Figure 2B). However,
hyphal initiation in a poor medium required inoculation of cells
from a saturated overnight culture. This result is consistent with
the report that release from quorum-sensing molecules, such as
farnesol, triggers the yeast-to-hypha transition at 37uC indepen-
dent of the inoculation media [30]. The timing of hyphal initiation
was slower in medium with mannitol than that with glucose
(Figure 2B), consistent with the activation of the cAMP pathway by
glucose in addition to the release of inhibition from farnesol.
Together, our observations suggest that hyphal initiation involves
a down-regulation of Nrg1 protein in response to the activation of
the cAMP-PKA pathway by a combination of temperature shift,
release from inhibition by farnesol, and nutrients in rich media.
The NRG1 transcript has been reported to be reduced during
hyphal induction in serum containing medium [22,25,26,53]. We
found that the decrease in NRG1 expression during hyphal growth
was dependent on the cAMP-PKA pathway, as Cyr1 (unpublished
data) or Tpk2 was required for the reduction in NRG1 expression
(Figure 2C), consistent with the observation that NRG1 transcript
level was not down-regulated in the efg1 mutant during hyphal
induction in serum [26]. The transcriptional down-regulation was
essential for the initiation of hyphal development, as ectopic
expression of NRG1 under the MAL2 promoter in wild-type cells
could not initiate hyphal development even under robust
induction conditions (Figure 2D). However, the decrease in
NRG1 transcript alone is not sufficient to provide the temporal
dynamic change in Nrg1 protein level during hyphal development.
We observed that, by promoter shut-down experiments, Nrg1 was
unstable when cells were inoculated into fresh medium at 30uCeither from an overnight culture (Figure 2E) or a logarithmic
growing culture (unpublished data). Adding farnesol inhibited
Nrg1 degradation, suggesting that release from farnesol inhibition
during inoculation is important in triggering the degradation of
Nrg1. Therefore, the rapid and temporary disappearance of Nrg1
protein is the combined result of a cAMP-PKA dependent down-
regulation of NRG1 expression and a burst of Nrg1 degradation
upon release from farnesol during inoculation. This temporary
removal of Nrg1 is essential for hyphal induction.
Promoter-Recruitment of Hda1 Is Required for SustainedHyphal Development by Limiting Promoter Access toNrg1
One possible mechanism for the reduced promoter access by
Nrg1 during hyphal elongation is a change in promoter
chromatin. C. albicans has one class II HDAC Hda1 that functions
as a repressor for phenotypic switching [54]. We found that cells
deleted of HDA1 were unable to maintain hyphal growth. They
were impaired in sustained hyphal development and transcription
of hypha-specific genes, but had no detectable defects during
initial germ-tube formation (Figures 3AB, S2A and Table S1A).
Furthermore, levels of promoter-bound Nrg1 increased dramat-
ically in the hda1 mutant after hyphal initiation, suggesting that
Hda1 is required to prevent Nrg1 binding to the promoters for
sustained hyphal growth (Figures 3C and S2B). These observations
of the hda1 mutant are similar to that of wild-type cells inoculated
into rich medium at 37uC without serum (Figure 1). To determine
whether Hda1 functions directly on the promoters of hypha-
specific genes, we performed a time course ChIP of Hda1. We
found that Hda1 was recruited to the promoters during hyphal
induction in a serum-dependent manner (Figures 3D and S2C).
We suggest that serum-induced Hda1 recruitment to the
promoters of hypha-specific genes leads to a change in promoter
chromatin that is no longer accessible to Nrg1, leading to sustained
hyphal development.
In contrast to Hda1, we found that the Rpd3 HDAC
preferentially associated with hypha-specific promoters in yeast
cells and dissociated rapidly from the promoters upon hyphal
induction, similar to the dynamic dissociation of Nrg1 (Figure S3).
The sequential dissociation and recruitment of two different
HDACs to the hypha-specific promoters likely account for the
dynamic change in nucleosomal H4 acetylation levels during
hyphal induction with the peak H4 acetylation at 30 min [50].
The Promoter-Associated Hda1 Deacetylates the Yng2Subunit of NuA4 Histone Acetyltransferase (HAT)module, Leading to the Reduction of Yng2 and NuA4HAT at the Promoters of Hypha-Specific Genes
How does promoter-associated Hda1 prevent Nrg1 from
binding to the promoters of hypha-specific genes during hyphal
elongation? One potential mechanism is chromatin remodeling.
We have previously shown nucleosome reassembly and a decrease
in H4 acetylation at the UAS regions of hypha-specific genes
Plasticity of Dimorphism
PLoS Biology | www.plosbiology.org 4 July 2011 | Volume 9 | Issue 7 | e1001105

Figure 2. Hyphal initiation requires cAMP-PKA dependent down-regulation of Nrg1 protein. (A) Components of the cAMP-PKA pathwayand release from farnesol inhibition are required for down-regulation of Nrg1 protein during hyphal initiation. Western analysis of Nrg1-Myc duringhyphal induction in wild-type, cyr1, tpk2, efg1, and flo8 mutants. Cells of wild type and mutants carrying Nrg1-Myc were diluted 1:100 into freshYPD+10% serum medium at 37uC with or without 30 mM farnesol as indicated. (B) Cell morphology and Western blot analysis of wild-type cellscarrying Nrg1-myc. Cells from overnight cultures of the indicated OD600 were diluted 1:100 into Lee’s media with either mannitol or glucose. Adding30 mM farnesol into Lee’s glucose medium blocked Nrg1 down-regulation and hyphal development. (C) Tpk2 is essential for transcriptional down-regulation of NRG1 during hyphal induction. NRG1 mRNA levels were determined by qRT-PCR in WT and tpk2/tpk2 strains over a time course ofhyphal induction (YPD+10% Serum). The signal obtained from ACT1 mRNA was used as a loading control for normalization. The data show theaverage of three independent qRT-PCR experiments with error bars representing the SEM. (D) Constitutive expression of Nrg1 blocks germ tubeformation. Cells of wild-type and nrg1/nrg1 carrying Nrg1-Myc under the MAL2 promoter were grown in YPMaltose medium at 30uC overnight andwere diluted at 1:20 into pre-warmed YPMaltose medium at 37uC. (E) Nrg1 protein stability by promoter-shutdown. Western or wild-type cellscarrying Nrg1-Myc under the MAL2 promoter inoculated from overnight culture into fresh YPD medium at 30uC with and without 30 mM farnesol.doi:10.1371/journal.pbio.1001105.g002
Plasticity of Dimorphism
PLoS Biology | www.plosbiology.org 5 July 2011 | Volume 9 | Issue 7 | e1001105

during hyphal elongation [50]. The decrease in H4 acetylation
could be a result of H4 deacetylation by promoter-associated
Hda1 or eviction of NuA4 from the promoters. S. cerevisiae Hda1 is
a histone deacetylase specific for H3 and H2B [55]. Therefore, H4
may not be a substrate of the C. albicans Hda1. Yng2, a subunit of
the NuA4 histone acetyltransferase HAT complex essential for
HAT activity, is acetylated at lysine 170 by NuA4 and
deacetylated by Rpd3 in S. cerevisiae [56]. Deacetylation of Yng2
leads to its degradation and eviction of chromatin-bound Yng2
with Esa1, the catalytic subunit of NuA4. In C. albicans, the NuA4
HAT complex is recruited to the promoters of hypha-specific
genes [50]. By immunoprecipitation with anti-acetylated-lysine
antibodies, we detected acetylated Yng2 in wild-type cells, but
observed a dramatic increase in the level of acetylated Yng2 in the
hda1 mutant (Figure 4A). This suggests that deacetylation of Yng2
in vivo depends on Hda1 activity. K175 was identified as a
candidate lysine residue for acetylation by sequence alignment
with S. cerevisiae Yng2. This was confirmed by the loss of
acetylation of Yng2 when we substituted K175 with arginine
(K175R), a mutation that blocks acetylation (Figure 4B). The loss
of acetylation of Yng2K175R also suggests that K175 is the major
acetylated lysine residue of Yng2 in C. albicans. The K175R
mutation led to a reduced level of Yng2, whereas substituting
K175 with glutamine (K175Q, a mutation mimicking constitutive
acetylation) did not cause a detectable change of protein
abundance relative to wild type (Figure 4C). Unlike the yng2
deletion mutant, neither yng2K175R nor yng2K175Q has any
detectable growth defect (unpublished data). We then examined
whether Yng2 acetylation affects hyphal development. Both wild-
type and yng2K175R cells developed long hyphae, whereas yng2K175Q
cells were defective in sustained hyphal development and
transcription (Figures 4D,E and S4A), a phenotype similar to that
of the hda1 mutant. Therefore, Yng2 deacetylation at K175 is
important for sustained hyphal transcription. Conversely, consti-
tutively acetylated Yng2 blocks hyphal filament extension.
If serum-induced Hda1 promoter association leads to Yng2
deacetylation and degradation, the level of promoter-associated
Yng2 is expected to decrease during hyphal induction. Indeed, a
time course ChIP of Yng2-Myc showed that Yng2 level decreased
after 30 min of hyphal induction (Figures 4F and S4B). The level
of promoter-associated Yng2K175R showed a similar decrease,
whereas the levels of Yng2-Myc in the hda1 mutant and
Yng2K175Q-Myc stayed unchanged (Figures 4F, 5C, and S4C).
The level of Esa1, the enzymatic subunit of NuA4, showed a
similar decrease at the promoters during hyphal induction
(unpublished data).
To determine whether the deacetylation of Yng2 is the major
function of Hda1 in hyphal development, we generated an hda1
Figure 3. Promoter recruitment of Hda1 is required for hyphal maintenance by inhibiting Nrg1 access to the promoters of hypha-specific genes. (A, B) Hda1 is required for maintenance of hyphal development as shown by morphology and expression levels of hypha-specificgenes during hyphal induction. Cells of wild-type (TS3.3+pBES116) and hda1/hda1 (HLY4032+pBES116) strains were diluted into YPD with 10% serumat 37uC. HWP1, ALS3, and ECE1 mRNA levels were determined by qRT-PCR. The signal obtained from ACT1 mRNA was used as a loading control fornormalization. (C) Nrg1-Myc binds the promoters of hypha-specific genes in hda1 cells during hyphal elongation in YPD with 10% serum. (D) Serum-dependent binding of Hda1-Myc to the hyphal promoters in wild-type cells during hyphal induction. Cells of wild-type strain containing Hda1-Myc(HLY4033) were diluted into pre-warmed YPD medium at 37uC in the presence or absence of 10% serum. ChIP DNA in (C, D) were quantitated byqPCR with primers at the UAS regions of hypha-specific genes as described in Figure 1. Data for ALS3 and ECE1 from (B, C, D) are shown in Figure S2.All data show the average of three independent qRT-PCR or qPCR experiments with error bars representing the SEM.doi:10.1371/journal.pbio.1001105.g003
Plasticity of Dimorphism
PLoS Biology | www.plosbiology.org 6 July 2011 | Volume 9 | Issue 7 | e1001105

Figure 4. The function of Hda1 in sustained hyphal transcription is mediated through Yng2 deacetylation (A) Yng2p is deacetylatedthrough an Hda1-dependent mechanism in vivo. Cells were grown at 30uC to OD600 0.8, then WCEs were collected, immunoprecipitated with anti-Ac-K, and probed with anti-Myc. (B) K175 is the major acetylated lysine residue of Yng2 in vivo. Substitution of K175 with arginine (K175R) diminishesacetylation of Yng2. (C) Effects of K175 substitutions on Yng2 stability. K175R mutation causes decreased protein abundance of Yng2, while K175Qcauses no detectable change. (D, E) Morphology and expression levels of hypha-specific genes in YNG2 (HLY4035), yng2K175R (HLY4036), and yng2K175Q
(HLY4037) cells during hyphal development in YPD with 10% serum at 37uC. HWP1, ALS3, and ECE1 mRNA levels were determined by qRT-PCR. Thesignal obtained from ACT1 mRNA was used as a loading control for normalization. (F) Dynamic dissociation of Myc-tagged Yng2 and yng2K175R butnot Yng2K175Q from promoters during hyphal development by ChIP with anti-Myc antibodies. (G) ChIP with anti-H3 and anti-acetylated H4 antibodies
Plasticity of Dimorphism
PLoS Biology | www.plosbiology.org 7 July 2011 | Volume 9 | Issue 7 | e1001105

yng2 double mutant and introduced the yng2K175R mutation into
the double mutant. The hda1 yng2K175R mutant behaved like
yng2K175R or wild-type cells with no defect in hyphal filament
extension and transcription (Figure 5A and 5B). Furthermore,
Hda1 is required for the reduction of promoter-associated Yng2,
and the yng2K175R mutant completely bypassed this requirement of
Hda1 (Figures 5C and S5). Therefore, the function of Hda1 in
sustained hyphal transcription is mediated completely through the
deacetylation of Yng2 at K175. In addition, the level of promoter-
associated Yng2K175R decreased in the hda1 mutant as did Yng2 in
wild-type cells during hyphal elongation (Figures 5C and S5),
suggesting that Hda1 deacetylation of Yng2 at the promoters led
to the eviction of Yng2 and Esa1 from the promoters of hypha-
specific genes.
To understand the effect of Yng2 K175 deacetylation on
promoter chromatin, we determined temporal changes in histone
levels and H4 acetylation at the UAS regions of hypha-specific
promoters during hyphal induction (Figures 4G and S4D).
Changes in H3 levels indicated rapid nucleosome disassembly
during hyphal initiation and nucleosome reassembly during
hyphal maintenance. The observed decrease in H3 occupancy
and transcriptional induction within 30 min of hyphal induction is
consistent with the notion that transcriptional activation correlates
with reduced nucleosome occupancy [57]. In contrast, nucleosome
reassembly during hyphal elongation seemed important for
sustained hyphal transcription, as yng2K175Q cells showed a specific
defect in nucleosome reassembly without any detectable impair-
ment in initial nucleosome disassembly. Our data suggest that
nucleosome reassembly during hyphal maintenance may function
in preventing Nrg1 from binding to the promoters.
Reduced TOR Signalling Led to Hda1 PromoterRecruitment and Sustained Hyphal Development
yng2K175Q cells exhibited normal hyphal initiation in several
hyphal growth media in addition to serum (Table S1B) and were
defective in sustained hyphal development in all media that can
support prolonged hyphal development (Figure 6A). This
suggested that the Hda1-mediated deacetylation of Yng2 was
Figure 5. yng2K175R mutation suppresses the deficiency of hda1/hda1 in sustained expression of hypha-specific genes. Morphology (A)and Northern analysis (B) of wild type and hda1/hda1 yng2/yng2 transformed with YNG2 or yng2K175R during hyphal induction as in Figure 3. Northernanalysis was carried out using 12.5 mg RNA and probes to the indicated filament-specific transcripts. The ACT1 transcript is included as a loadingcontrol. (C). Kinetics of Yng2-Myc and Yng2-Myc-K175R promoter binding in hda1/hda1 mutants by ChIP with anti-Myc. Data on ALS3 and ECE1 for (C)are in Figure S5. The ChIP data show the average of three independent qPCR experiments with error bars representing the SEM.doi:10.1371/journal.pbio.1001105.g005
show temporal dynamics in relative levels of H3 and H4 acetylation at the promoters of hypha-specific genes during hyphal induction in the YNG2(HLY4035) and yng2K175Q mutant (HLY4037) strains. Levels of H3 (relative H3 occupancy; bound/input) are normalized to the respective control DNAat the ADE2 coding sequence region (bound/input). The 0-h values in the wild-type strain are set to be 1.00. Acetylation levels normalized withrespect to H4 levels (H4 acetylation/H3 occupancy) are calculated by dividing the values for acetylated H4 with H3 occupancy values. Data on ALS3and ECE1 for (E, F, G) are in Figure S4. All data show the average of three independent qPCR experiments with error bars representing the SEM.doi:10.1371/journal.pbio.1001105.g004
Plasticity of Dimorphism
PLoS Biology | www.plosbiology.org 8 July 2011 | Volume 9 | Issue 7 | e1001105

required for hyphal maintenance in those media. In addition,
Hda1 was found to be associated with the promoters of hypha-
specific genes, whereas Nrg1 promoter-association was inhibited
after 6 h of growth in those conditions (Figure 6B). Interestingly,
those media tend to be nutrient-poor. One commonly used
synthetic nutrient-poor medium for hyphal growth is Lee’s
medium [58]. It contains glucose, ammonium sulfate, and many
amino acids, but lacks glutamine, a preferred nitrogen source. The
rapamycin-sensitive Tor1 kinase is a central regulator of cell
growth in response to nitrogen and amino acid availability in yeast
[59] and is conserved in C. albicans [41]. We hypothesized that a
nutrient-poor medium, such as Lee’s, is sensed by C. albicans cells
as nitrogen limiting, and therefore leads to reduced Tor1 activity.
If so, treatment of C. abicans cells with rapamycin in a rich medium
should mimic a nutrient-poor medium for sustained hyphal
development. As predicted, treatment of C. albicans cells with a
sublethal concentration of rapamycin after hyphal initiation at
37uC led to sustained hyphal filamentation in rich medium
(Figure 7A). A higher concentration of rapamycin, however,
slowed cell growth and hyphal elongation (unpublished data),
consistent with the finding that rapamycin inhibits filamentation
on nutrient-poor media [41]. Sustained hyphal filamentation was
also observed at 30uC with sublethal levels of rapamycin after cells
were first grown at 37uC for hyphal initiation (Figure 7A). The
effect of rapamycin on hyphal maintenance was only observed
after hyphal initiation. Cells treated with rapamycin at 30uC did
not form hyphae (Figure 7B). We further showed that the effect of
Tor1 on hyphal maintenance was mediated through the promoter-
recruitment of Hda1, as yng2K175Q cells were defective in sustained
hyphal filamentation in rapamycin (Figure 7C). In addition, Hda1
was recruited to the promoters of hypha-specific genes in cells
treated with rapamycin in rich medium, whereas Nrg1 was
excluded from the promoters (Figure 7D). The phenotypes of the
yng2K175Q cells and patterns of promoter association for Hda1 and
Nrg1 are similar between poor medium and rapamycin treatment
(Figures 6 and 7). Furthermore, a mutant with a hyperactive TOR
Figure 6. yng2K175Q mutant is defective in hyphal elongation in nutrient-poor media. (A) Cell morphology of wild-type (HLY4035) andyng2K175Q (HLY4037) cells grown at 37uC in YEP+GlcNAc, SCAA, M199 pH 8.0, or Lee’s with mannitol for indicated times. (B) ChIP with anti-Mycantibodies in wild-type cells carrying Hda1-Myc (HLY4033) or Nrg1-Myc (HLY3922). qPCR was used to quantitate ChIP DNA with primers at the UASregion of HWP1 under the indicated conditions. The ChIP data show the average of three independent qPCR experiments with error barsrepresenting the SEM.doi:10.1371/journal.pbio.1001105.g006
Plasticity of Dimorphism
PLoS Biology | www.plosbiology.org 9 July 2011 | Volume 9 | Issue 7 | e1001105
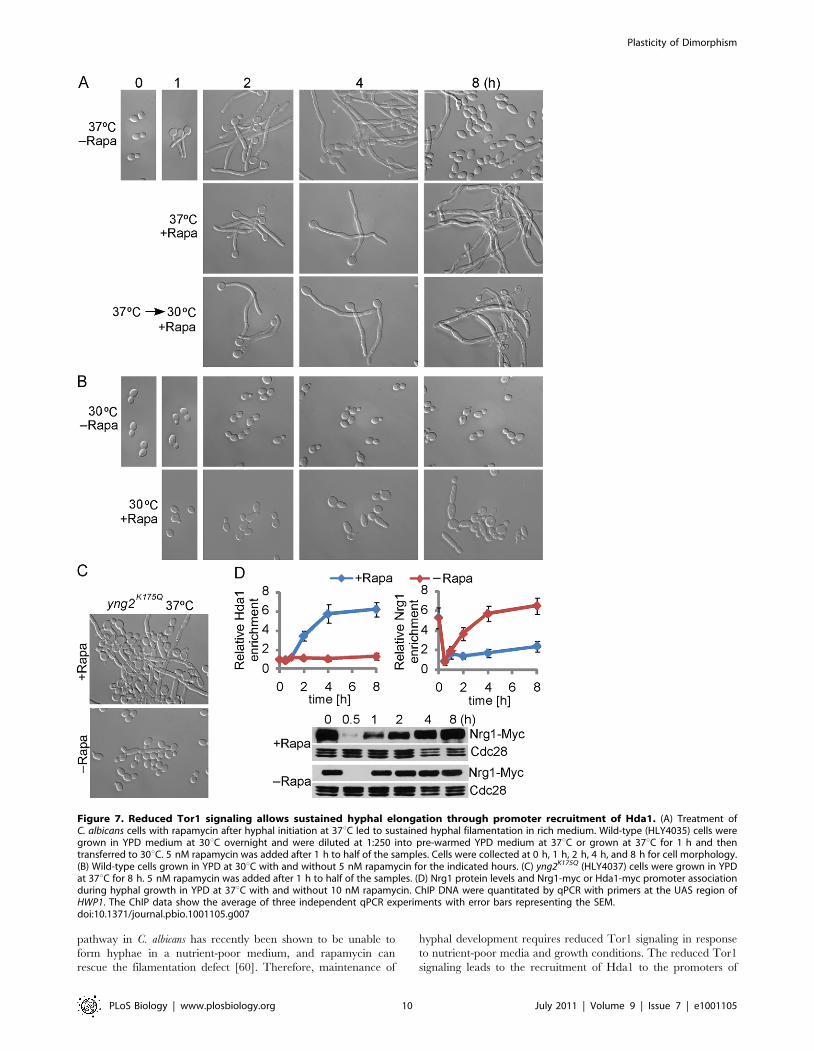
pathway in C. albicans has recently been shown to be unable to
form hyphae in a nutrient-poor medium, and rapamycin can
rescue the filamentation defect [60]. Therefore, maintenance of
hyphal development requires reduced Tor1 signaling in response
to nutrient-poor media and growth conditions. The reduced Tor1
signaling leads to the recruitment of Hda1 to the promoters of
Figure 7. Reduced Tor1 signaling allows sustained hyphal elongation through promoter recruitment of Hda1. (A) Treatment ofC. albicans cells with rapamycin after hyphal initiation at 37uC led to sustained hyphal filamentation in rich medium. Wild-type (HLY4035) cells weregrown in YPD medium at 30uC overnight and were diluted at 1:250 into pre-warmed YPD medium at 37uC or grown at 37uC for 1 h and thentransferred to 30uC. 5 nM rapamycin was added after 1 h to half of the samples. Cells were collected at 0 h, 1 h, 2 h, 4 h, and 8 h for cell morphology.(B) Wild-type cells grown in YPD at 30uC with and without 5 nM rapamycin for the indicated hours. (C) yng2K175Q (HLY4037) cells were grown in YPDat 37uC for 8 h. 5 nM rapamycin was added after 1 h to half of the samples. (D) Nrg1 protein levels and Nrg1-myc or Hda1-myc promoter associationduring hyphal growth in YPD at 37uC with and without 10 nM rapamycin. ChIP DNA were quantitated by qPCR with primers at the UAS region ofHWP1. The ChIP data show the average of three independent qPCR experiments with error bars representing the SEM.doi:10.1371/journal.pbio.1001105.g007
Plasticity of Dimorphism
PLoS Biology | www.plosbiology.org 10 July 2011 | Volume 9 | Issue 7 | e1001105

hypha-specific genes for promoter remodeling that prevents Nrg1
from binding.
Temporal Connection between Hyphal Initiation andMaintenance
To uncover potential molecular connections between the
initiation phase and elongation phase of hyphal development, we
asked whether serum could induce the promoter recruitment of
Hda1 without the initiation phase. We found that Hda1 did not
bind the promoters of hypha-specific genes when cells were grown
at 30uC in the presence of serum (Figure 8A). Therefore, serum is
not sufficient to induce the promoter recruitment of Hda1 without
the initiation step. We further showed that raising the temperature
to 37uC during inoculation was a prerequisite for subsequent
recruitment of Hda1 to the promoters (Figure 8A). Since Nrg1 was
temporally cleared when cells were inoculated into fresh medium
at 37uC, a requirement for hyphal initiation, we then examined
whether serum could induce promoter recruitment of Hda1
independent of temperature in the nrg1 deletion mutant by a time
course ChIP experiment. Hda1 was found to bind the HWP1
promoter in the presence of serum in nrg1 mutants in either 25uCor 37uC (Figure 8B). This result suggests that Nrg1 removal is
required and sufficient for serum-induced Hda1 promoter
recruitment for the sustained hyphal transcriptional program.
This also predicts that the sustained hyphal transcriptional
program could only be established within the time window of
reduced Nrg1 when cells had just been inoculated into fresh media
at 37uC. Indeed, addition of serum after 2 h of hyphal induction at
37uC in YPD could not sustain hyphal development (Figure 8C).
Similar to serum, rapamycin could sustain hyphal growth only
when added during hyphal initiation. Adding rapamycin after 2 h
of hyphal induction showed no effect on hyphal maintenance
Figure 8. The sustained hyphal growth program could only be established during the time window of reduced Nrg1. (A) Serum is notsufficient to induce the promoter recruitment of Hda1 without hyphal initiation at 37uC. ChIP with anti-Myc antibodies in wild-type cells carryingHda1-Myc in YPD with 10% serum under indicated conditions. Primers at the UAS region of HWP1 were used. (B) Nrg1 removal is required andsufficient for serum-induced Hda1 promoter recruitment. The ChIP time course was carried out with anti-Myc antibodies in nrg1/nrg1 cells carryingHda1-Myc under the indicated conditions. Cells were collected at 0 min, 15 min, 30 min, 1 h, 2 h, and 3 h. The ChIP data show the average of threeindependent qPCR experiments with error bars representing the SEM. (C) Serum and rapamycin are unable to sustain hyphal development when theyare added to YPD medium 2 h after inoculation at 37uC. Cells from overnight cultures in YPD at 30uC were diluted at 1:250 into pre-warmed YPDmedium at 37uC; 10% serum or 5 nM rapamycin were added at 1 h, 2 h, or 3 h after inoculation, respectively. The total incubation time is 5 h forserum and 8 h for rapamycin experiments.doi:10.1371/journal.pbio.1001105.g008
Plasticity of Dimorphism
PLoS Biology | www.plosbiology.org 11 July 2011 | Volume 9 | Issue 7 | e1001105

(Figure 8C). Together, our data show a clear temporal connection
between hyphal initiation and maintenance: Hda1 can be
recruited to the promoters of the hypha-specific genes only when
Nrg1 level is low. The time period of reduced Nrg1 during hyphal
initiation can be viewed as a window of opportunity for
establishing the hyphal transcription program. The strength and
duration of the cAMP signal in each cell is expected to affect the
commitment to the hyphal transcriptional program; once
committed, the cAMP-PKA pathway activation is no longer
needed. This is consistent with the observation that farnesol only
inhibits germ-tube formation and has no effect on hyphal
elongation [39]. The duration of hyphal development is
determined by nutrient conditions through the TOR pathway.
Discussion
This study shows that the yeast to hyphae morphogenetic switch
consists of two temporally related phases of regulation of the
promoter chromatin of hypha-specific genes: the first for initiation
and the second for maintenance of hyphal development (Figure 9).
Initiation requires the cAMP-PKA pathway and maintenance
requires reduced Tor1 signaling. A spike of cAMP-PKA activation
[10] and release from the inhibition of quorum sensing molecules,
such as farnesol, initiate hyphal development by transiently
clearing Nrg1 protein. The duration of hyphal development is
under the regulation of Tor1 by controlling promoter access to
Nrg1 through promoter recruitment of Hda1. Therefore, hyphal
maintenance requires active sensing of growth environments.
Hyphal cells convert to yeast when nutrients are replete. This
provides an underlying mechanism for the plasticity of dimor-
phism. Importantly, the sustained transcriptional program, as
measured by promoter recruitment of Hda1, can only be
established during the time period when Nrg1 is absent. This
provides a temporal link at the molecular level between the two
phases of hyphal development. This is the first example of a
temporal integration of two major nutrient-sensing cell growth
pathways in development. Temporal restriction in development of
multicellular organisms is common. For example, nutritional
control of the reproductive status in honeybees is restricted by a
critical ‘‘decision-making’’ period in larval development [61]. Our
study also provides a mechanism for how a transient signal, such as
a cAMP spike, can make a long-lasting impact in transcriptional
reprogramming during development. Such control of cellular
development by a burst of proliferating signal, followed with
sustained action of reduced TOR signaling has been observed for
memory T-cell differentiation [62]. The TOR level is also critical
for maintaining stem cell homeostasis [63–65]. Temporal
regulation of cell fate by different signaling pathways is likely
Figure 9. A schematic diagram depicting initiation and maintenance of hyphal development. (A) Hyphal development in C. albicansrequires two phases of sequential changes in the promoter chromatin of hypha-specific genes. Initiation requires a rapid but temporarydisappearance of Nrg1 in response to the activation of the cAMP-PKA pathway, while the duration of hyphal development is under the regulation ofthe TOR pathway by controlling promoter access to Nrg1 through recruitment of Hda1. Black lines represent active regulatory relationships; grey linesrepresent relationships that are inactive. Dashed circles represent degraded proteins. (B) Hda1 is recruited only during the time period when Nrg1 isabsent. This provides a mechanism for temporal integration of two major nutrient-sensing cell growth pathways in development.doi:10.1371/journal.pbio.1001105.g009
Plasticity of Dimorphism
PLoS Biology | www.plosbiology.org 12 July 2011 | Volume 9 | Issue 7 | e1001105

common in development of multicellular organisms. Coupling two
signaling pathways through the regulation of promoter chromatin
provides a unique mechanism.
Initiation of the hyphal transcriptional program requires
temporary removal of Nrg1 protein. The timing, duration, and
extent of the Nrg1 down-regulation are well correlated with timing
and efficiency of hyphal initiation and are sensitive to multiple
factors, including the state of inoculating cells (e.g., stationary
cells), fold of dilution at inoculation, media, and temperature. Of
those, a rise in temperature to 37uC and release from farnesol
inhibition are essential for the clearance of Nrg1 and hyphal
initiation. The cAMP-PKA pathway is required for the down-
regulation of Nrg1, consistent with the concept that Cyr1 is a hub
that integrates many of the hyphal inducing signals [31–33,40].
Efg1 and Flo8 are both required for the down-regulation of Nrg1,
but a TRA or TRE mutation at the sole conserved PKA site in
Efg1 (T206) does not reduce or enhance hyphal gene expression
(our unpublished data). If Efg1 activity is directly regulated by
PKA phosphorylation, the regulation is not essential or sufficient
for the induction of hypha-specific genes. Efg1 and Flo8 also
function directly on the promoters of hypha-specific genes [16,50],
and the tup1 mutant cannot bypass the requirement of Efg1 for
hyphal transcription [66]. Our finding that Nrg1 is negatively
regulated by the cAMP-PKA pathway and by farnesol is
intriguing. The nrg1 mutant produces 19-fold more farnesol than
the parent wild-type strain [67], and farnesol in turn impacts
activity of the Cyr1 pathway [40]. Together, these findings provide
a loop of three negative feedback regulations from farnesol to
Cyr1, to Nrg1, and then back to farnesol. Removal of farnesol
during inoculation will lead to activation of the cAMP-PKA
pathway and clearance of Nrg1 (Figure 9). Lack of Nrg1 in turn is
expected to lead to increased production of farnesol, and a new
balance among the components in the feedback loop will be
achieved. Another intriguing finding is that the sustained hyphal
transcriptional program can only be established during the time
window of low Nrg1. Commitment to the hyphal program is
therefore determined by the strength and duration of Nrg1 down-
regulation in each cell. If the duration is not long enough to
establish the Hda1-mediated chromatin remodeling, the cell will
be in the yeast growth phase. This explains the cell-to-cell
variation and experiment-to-experiment variation in hyphal
induction in a given culture, and why yeast and hyphal cells can
exist in the same culture. Our model explains a widely accepted
observation that the quality of the initial hyphal induction is
important for the fate of hyphal development. It also provides a
molecular mechanism for why farnesol only inhibits germ-tube
formation and has no effect on hyphal elongation [39].
Our finding of temporally coupled chromatin remodeling under
reduced Tor1 signaling for sustained hyphal transcription provides
a molecular mechanism for why nutrient-poor media are typical
media for sustained hyphal development. It has been puzzling how
C. albicans can respond to both rich and poor media and undergo
hyphal development. Rich medium with serum is the most robust
medium for hyphal induction, and this is consistent with the
requirement of the cAMP-PKA pathway for hyphal growth.
Hyphal induction in nutrient-poor media also requires an
activation of the cAMP-PKA pathway, mostly through the release
from farnesol inhibition. But sustained hyphal growth requires a
nutrient limitation. Considering the conserved functions of Tor1 in
nutrient sensing and growth regulation from yeast to human,
involvement of Tor1 in hyphal development has been studied and
implicated [41,60,68]. However, rapamycin cannot induce hyphal
development, making it hard to reconcile if the cAMP-PKA and
Tor1 pathways function in parallel. We find that rapamycin can
lead to the promoter recruitment of Hda1 only during hyphal
initiation in the absence of Nrg1. The temporal coupling of the
two pathways through the regulation of the promoter chromatin of
hypha-specific genes explains why rapamycin by itself cannot
induce hyphae. Since serum can sustain hyphal development in
YPD, we suspect it may contain certain components that are
inhibitory to the Tor1 pathway. The nuclear localization and the
activity of several nutrient-responsive transcription factors are
regulated by Tor1-mediated phosphorylation in yeast [69]. It is
possible that a Tor1-regulated transcription factor is necessary for
the recruitment of Hda1 to the promoters of hypha-specific genes.
Further studies are needed to address this. In addition to Hda1-
mediated chromatin remodeling for sustained hyphal transcrip-
tion, there is a built-in positive feedback on the hyphal
transcriptional program that is also important for sustained hyphal
development. The transcription factor Ume6, specifically ex-
pressed during hyphal development, controls the level and
duration of hypha-specific genes and is important for hyphal
elongation [53,70,71]. The strength of the cAMP and Tor1
signaling determines the level and duration of UME6 expression,
which in turn dictates the extent of hyphal morphogenesis. Hyphal
morphogenesis and cell chain formation is under the control of
another hypha-specific gene that encodes the G1 cyclin-related
protein Hgc1 [72–76].
Our study should provide insights into understanding C. albicans
pathogenesis. Hyphal initiation is tightly linked to release from
quorum-sensing molecules, a condition parallel to dispersion of C.
albicans cells from biofilms in human hosts, which is a major cause
of disseminated candidiasis [77]. Invasive candidiasis is the most
common invasive fungal infection among organ transplant
recipients [78]. Rapamycin is given to transplant patients,
especially renal transplant patients, to prevent organ rejection.
One speculation from our finding is that rapamycin, known for its
antifungal activity, may also facilitate hyphal development and
invasive infections. The existence of extensive hyphae in various
deep-seeded infection sites also suggests that those host environ-
ments may be perceived as stressful or starvation conditions by
Candida. Our findings provide molecular mechanisms for how C.
albicans adapts to varied host environments and develops from a
benign commensal into a disseminated invasive disease.
We have previously noticed the dynamic increase and decrease in
H4 acetylation concomitant with nucleosome disassembly and
reassembly at the promoters of hypha-specific genes during hyphal
induction [50]. The significance of the temporal dynamic regulation
in promoter chromatin is not clear. Here we find the sequential
dissociation of Rpd3 and association of Hda1 to the promoters is
concomitant with the dynamic changes in H4 acetylation.
Importantly, we show that promoter-associated Hda1 decreases
H4 acetylation level by deacetylating Yng2 and evicting Yng2 and
Esa1 of the NuA4 HAT module from the promoters, leading to
nucleosome reassembly, inhibition of Nrg1 binding, and sustained
hyphal transcription. This provides an example for Hda1 in gene
activation and NuA4 in gene repression. The function of NuA4 in
repression here is revealed by the use of yng2K175Q and yng2K175R
mutations. Temporal analysis of promoter chromatin also delineates
different functions of HDACs and NuA4 in transcription. It shows
that nucleosome reassembly does not necessarily correlate with
transcriptional repression; it can also be used for gene activation by
restricting access of repressors. In S. cerevisiae, Yng2 is deacetylated
by Rpd3 and a similar temporal dynamic H4 acetylation is observed
at DNA double-stranded breaks concomitant with the sequential
recruitment of NuA4 and Rpd3 [56]. Such temporal dynamic
regulation of chromatin is likely also used in transcription in S.
cerevisiae. Indeed Rpd3 is required for the expression of several
Plasticity of Dimorphism
PLoS Biology | www.plosbiology.org 13 July 2011 | Volume 9 | Issue 7 | e1001105

environment-responsive genes [79–82]. Active functions of HDACs
in gene expression are not limited to yeast. A recent genome-wide
mapping of HAT and HDAC binding sites in human T cells shows
that all HDACs analyzed are associated with active genes and
positively correlated with transcription, with some HDACs mainly
in the promoter regions and other HDACs in both the promoter
and gene body regions [83]. Inhibition of HDAC activity with
HDAC inhibitor treatment causes increases in acetylation in the
active genes, suggesting that the majority of HDACs function to
reset chromatin by removing acetylation at active genes [83]. The
temporal dynamic NuA4 regulation by HDACs shown in this study
provides an attractive mechanism likely used for temporal
integration of different signals in transcriptional reprogramming
during development.
Materials and Methods
Media and Growth ConditionsC. albicans strains were routinely grown at 30uC in YPD (2%
Bacto peptone, 2% dextrose, 1% yeast extract). Transformants
were selected on synthetic medium (2% dextrose, 0.17% Difco
yeast nitrogen base w/o ammonium sulfate, 0.5% ammonium
sulfate and auxotrophic supplements). M199 medium (Sigma-
Aldrich) was buffered at pH 8 using 150 mM HEPES. Hyphal
induction was performed as follows. Strains were grown overnight
in liquid YPD at 30uC, pelleted, washed twice in PBS, resuspended
in an equal volume of PBS, and diluted 1:100, unless otherwise
indicated, into YPD+10% serum (Sigma-Aldrich), Lee’s medium
[58] with modifications [5 g (NH4)2SO4, 2.5 g K2HPO4 Anhy-
drous, 5 g NaCl, 0.5 g alanine, 1.3 g leucine, 1 g lysine, 0.1 g
methionine, 0.072 g ornithine, 0.5 g proline, 0.5 g threonine,
10 ml 20 mg/ml MgSO4, 1 ml 2 mg/ml biotin, with pH adjusted
to 7.0, with either 1% glucose or mannitol, filled to 1 L with
water], YEP+2% N-acetyl-glucosamine (Sigma-Aldrich), SCAA
medium [15] with modifications [2% Casein Hydrolysate broth
(Sigma-Aldrich), 0.17% Difco yeast nitrogen base w/o ammonium
sulfate], Spider medium [1% mannitol, 1% nutrient broth, 0.2%
K2HPO4, pH 7.2 before autoclaving] [84], M199 pH 8 or
YPD+5 nM rapamycin. Cultures were grown at 37uC.
Plasmid and Strain ConstructionThe C. albicans strains used in this study are listed in Dataset S1.
Primer sequences are listed in Dataset S2. HDho15 and RPho19
[54] were streaked on 5-fluoro-orotic acid-containing medium to
generate HLY4032 and HLY4039 (Ura2 strains for hda1/hda1
and rpd3/rpd3 mutants). Ura+ strains of TS3.3 [85] and HLY4032
were obtained by transforming AscI digested pBES116 [37], which
were used for hyphal induction. hda1/hda1 yng2/yng2 mutants were
generated by deleting YNG2 in HLY4032 [50]. The disruption was
confirmed by southern blotting (unpublished data).
A 0.9-kb PCR product (primers 1 and 2) containing the C-terminal
NRG1 coding region and a 1.3-kb PCR product (primers 9 and 10)
containing the C-terminal RPD3 coding region were inserted into the
BamHI-MluI sites of pPR673 [50]. The resulting plasmids were
digested with SacI to target integration into their own loci to express
Nrg1-13Myc and Rpd3-13Myc. A 1.3-kb PCR product (primers 5
and 6) containing the C-terminal HDA1 coding region and a 0.8-kb
PCR product (primers 7 and 8) containing the C-terminal YNG2
coding region were digested with BglII and MluI and cloned into the
BamHI-MluI sites of pPR673. The resulting plasmids were digested
with BamHI to target the integration of the plasmid into its own locus
to express Hda1-13Myc and Yng2-13Myc. The pMAL2-NRG1-
13Myc plasmid was constructed by amplifying NRG1-13MYC
(primers 3 and 4) from pACT1-NRG1-13MYC plasmid (our
unpublished data). The PCR product was digested with XbaI and
KpnI to replace WOR1-HA from pMAL2-WOR1-HA plasmid [86].
The resulting plasmids were digested with AscI to target integration
into the ADE2 locus to express Nrg1-13Myc.
The YNG2 coding sequence was amplified using primers 13 and
14. The resulting 0.9-kb PCR product was digested with NotI and
MluI and inserted into the NotI-MluI site of pPR673 to create
pACT1-YNG2-13MYC. A 1.6-kb YNG2 promoter fragment
upstream of the START site of YNG2 (from 21 to 21,600) was
PCR amplified using primers 11 and 12. The resulting purified
PCR product was digested with StuI and BamHI and cloned into
the StuI-BamHI site of pACT1-YNG2-13MYC, displacing the 1-kb
ACT1 promoter region to generate pYNG2-YNG2-13MYC. Two-
step PCR was used to create pYNG2-YNG2K175R-13MYC. Two
pairs of primers (primers 13 and 15, 16 and 14) were used to PCR
amplify overlapping YNG2 fragments with the mutation in the
overlapping region. The resulting PCR products were purified and
mixed as templates for another round of PCR amplification using
the primers 13 and 14, which produced the full-length YNG2K175R
sequence. The resulting mutant, YNG2K175R, was cloned into the
NotI-MluI site of the plasmid pYNG2-YNG2-13MYC, replacing
the wild-type copy, and was confirmed by DNA sequencing. Two-
step PCR was used to create pYNG2-YNG2K175Q-13MYC. Two
pairs of primers (primers 13 and 17, and 18 and 14) were used to
PCR amplify overlapping YNG2 fragments with the mutation in
the overlapping region. Subsequent steps were done by using the
same methods as pYNG2-YNG2K175R-13MYC. These plasmids
were digested with the PmlI within the YNG2 promoter region for
integration into the endogenous YNG2 locus in yng2/yng2 and
hda1/hda1 yng2/yng2 mutant strains.
Northern and Quantitative RT-PCR Expression AnalysisMethods for RNA isolation and Northern blot hybridization
were carried out as previously described [87]. Probes for HWP1,
ALS3, ECE1, and ACT1 were made by PCR amplification of
,500-bp fragments from coding regions of each gene from
SC5314 genomic DNA. The primers used were as follows: HWP1,
primers 19 and 20; ALS3, primers 21 and 22; ECE1, primers 23
and 24; and ACT1, primers 25 and 26. For quantitative real-time
reverse transcription-PCR (qRT -PCR) analysis, 10 mg of total
RNA was DNase-treated at 37uC for 1 h using the DNA-free kit
(Qiagen), cDNA was synthesized using the SuperScript II Reverse
Transcriptase kit (Invitrogen), and qPCR was done using the iQ
SYBR Green Supermix (Bio-Rad) using the primers 27 and 28 for
HWP1, primers 29 and 30 for ALS3, primers 31 and 32 for ECE1,
primers 33 and 34 for ACT1, and primers 35 and 36 for NRG1.
The iCycler iQ detection system (Bio-Rad) was used with the
following program: initial denaturation at 94uC for 5 min,
followed by 40 cycles of 94uC for 20 s, 56uC for 30 s, and 68uCfor 20 s. Amplification specificity was determined by melting curve
analysis.
Chromatin ImmunoprecipitationChromatin immunoprecipitation was performed as described
with modifications [50,57]. Cells were formaldehyde cross-linked
by adding formaldehyde (37%) to a 1% final concentration.
Treated cultures were mixed by shaking and incubated for 15 min
at room temperature. 2.5 M glycine was added to a final
concentration of 125 mM, and treated cultures were mixed and
incubated for 5 min at room temperature. Cells were pelleted at
3,000 g for 5 min at 4uC, washed four times with 20 ml PBS, and
resuspended in 400 ml of 4uC lysis buffer (50 mM HEPES-KOH
[pH 7.5],140 mM NaCl, 1 mM EDTA, 1% Triton X-100, 0.1%
sodium deoxycholate) with protease inhibitors. All subsequent
Plasticity of Dimorphism
PLoS Biology | www.plosbiology.org 14 July 2011 | Volume 9 | Issue 7 | e1001105

ChIP and wash steps were done at 4uC. Cells were lysed using a
Fast-Prep system (FP120; Thermo Electron, Waltham, MA). DNA
was sheared by sonication six times for 20 s at high power on a
Bioruptor (diagenode) with 40 s intervals on ice.
For the IP, 10 ml of anti-Myc (SC-789, Santa Cruz), 4 ml of anti-
H3 (ab1791; Abcam) or anti-acetylated-H4 (06-866; Millipore)
antibodies were used for ,4 mg of chromatin proteins in an
immunoprecipitation volume of 200 ml. The IP was incubated O/
N at 4uC, with agitation. Then 50 ml of a 50% suspension of
protein A-Sepharose beads (GE Healthcare, 17-0974) in lysis
buffer was added to the IP and incubated 2 h at 4uC with
agitation. The beads were pelleted for 1 min at 3,000 g. After
removal of the supernatant, the beads were washed with a series of
buffers for 5 min each wash: twice in lysis buffer, twice in high-salt
lysis buffer (50 mM HEPES-KOH [pH 7.5], 500 mM NaCl,
1 mM EDTA, 1% Triton X-100, 0.1% sodium deoxycholate),
twice in wash buffer (10 mM Tris-HCl [pH 8.0], 250 mM LiCl,
0.5% NP-40, 0.5% sodium deoxycholate, 1 mM EDTA), and
once in TE (10 mM Tris, 1 mM EDTA [pH 8.0]). After the last
wash, 75 ml of elution buffer (50 mM Tris-HCl [pH 8.0], 10 mM
EDTA, 1% SDS) was added to each sample, and the beads were
incubated at 65uC for 10 min. The beads were spun for 1 min at
5000 g, and the supernatant was removed and retained. A second
elution was carried out with 75 ml of elution buffer 2 (TE, 1%
SDS) and eluates from the two elution steps were combined. For
the ChIP input material set aside, 140 ml elution buffer 2 (TE, 1%
SDS) was added to 10 ml WCE. ChIP and input samples were
incubated overnight at 65uC to reverse the formaldehyde cross-
links. 150 ml of proteinase K solution (TE, 60 mg/ml glycogen,
500 mg/ml proteinase K) was added to each sample, and samples
were incubated at 37uC for 2 h. Samples were extracted twice with
400 ml Tris buffer-saturated phenol/chloroform/isoamyl alcohol
solution (25:24:1). 15 ml of 5 M NaCl and 0.8 ml of 100% ethanol
(4uC) were added and the DNA was precipitated for 20 min at
220uC, then pelleted by centrifugation at 14,000 g for 10 min at
4uC, washed once with cold 70% ethanol, and allowed to air dry.
The samples were resuspended in 30 ml of TE containing 100 mg/
ml RNaseA and incubated for 1 h at 37uC. DNA derived from the
whole cell extracts and immunoprecipitation (IP) eluate was
analyzed by quantitative PCR (qPCR). Plotted are the average
and standard error of the mean of three independent qPCR
reactions for each experiment.
ImmunoprecipitationAcetylated Yng2-Myc was immunoprecipitated with 30 ml
protein A-sepharose beads (GE Healthcare, 17-0974) conjugated
with 10 ml of rabbit polyclonal anti-acetylated-lysine (Cell
Signaling, 9441S) at 4uC overnight in 200 ml pre-cleared WCEs,
and detected with a peroxidase-conjugated anti-c-Myc antibody
(Roche).
Supporting Information
Figure S1 Nrg1 is detected in the nucleus of both apical and
subapical cells of hyphae. Wild type cells carrying Nrg1-Myc
(HLY3922) were processed for indirect immunofluorescence, as
described [88]. Cells were grown in YPD+10% Serum medium at
37uC for hyphal induction or in YPD medium for yeast growth.
Cells were fixed at 5 h after induction and stained for Nrg1-Myc
with 9E10 mouse antibodies (Covance) and FITC-conjugated
secondary antibodies (Jackson Laboratory). DNA was stained with
DAPI. An untagged control (SC5314) was included.
(TIF)
Figure S2 Promoter recruitment of Hda1 is required for
hyphal maintenance by inhibiting Nrg1 access to the promoters
of hypha-specific genes. (A) ALS3 and ECE1 mRNA levels were
determined by qRT-PCR as described in Figure 3 (B) Kinetics
of Nrg1-Myc (B) and Hda1-Myc (C) binding at the ALS3 and
ECE1 promoters were determined by ChIP as described in
Figure 3C,D.
(TIF)
Figure S3 Rpd3-Myc disassociates rapidly from the promoters
of hypha-specific genes upon hyphal induction, determined by
ChIP as described in Figure 1B.
(TIF)
Figure S4 The function of Hda1 in sustained hyphal transcrip-
tion is mediated through Yng2 deacetylation (A) ALS3 and ECE1
mRNA levels were determined by qRT-PCR as described in
Figure 4(B) Relative Yng2 enrichment (B), relative H3 occupancy
(C), and H4 acetylation level (D) at ALS3 and ECE1 promoter.
ChIPs were performed as described in Figure 4E,F,G.
(TIF)
Figure S5 Kinetics of Yng2-Myc and Yng2K175R-Myc promoter
binding in hda1 mutants by ChIP with anti-Myc as described in
Figure 5C.
(TIF)
Table S1 Yng2 deacetylation by Hda1 is not required for germ
tube formation. (A) Germ tube formation of wild type and hda1/
hda1. Cells of wild type (TS3.3+pBES116) and hda1/hda1
(HLY4032+pBES116) were diluted 1:100 into indicated medium
at 37uC. The percentage of cells forming germ tubes in YPD+10%
serum medium, Spider medium, and M199 pH 8 medium at
60 min or in Lee’s medium at 180 min was determined by
counting at least 300 cells/sample, in triplicate. The samples from
Spider medium were gently sonicated to disrupt clumping. Mean
(% germ tube formation) 6 SE (standard error) of two
independent experiments. The hda1/hda1 mutant is able to form
germ tube in YPD with serum and Spider media but shows a
dramatically reduced level of germ tube formation in M199 and
Lee’s media. This is likely due to impaired growth of the mutant in
the media. The doubling time of the wild type (TS3.3+pBES116)
and hda1/hda1 (HLY4032+pBES116) in YPD medium at 30uC is
105 min and 135 min, respectively, and in M199 PH 8 medium at
30uC is 150 min and over 18 h, respectively. The defect of hda1/
hda1 cells in germ tube formation in M199 is consistent with the
report by Zacchi et al. [89]. (B) yng2K175Q mutant has no dramatic
defect in germ tube formation. Cells of wild type YNG2
(HLY4035), yng2K175R (HLY4036), and yng2K175Q (HLY4037) were
diluted 1:100 into indicated medium at 37uC. The percentage of
cells forming germ tubes was calculated as described in (A). The
two yng2 mutants show a similar growth rate as the YNG2 strain in
all media examined.
(PDF)
Acknowledgments
We thank Drs. D. Soll, J.Y. Chen, M. Whiteway, A. Mitchell, and J. Ernst
for C. albicans strains, and G.R. Fink for comments on the manuscript.
Author Contributions
Conceived and designed the experiments: YL AW HL. Performed the
experiments: YL CS AW. Analyzed the data: YL CS HL. Wrote the paper:
YL HL.
Plasticity of Dimorphism
PLoS Biology | www.plosbiology.org 15 July 2011 | Volume 9 | Issue 7 | e1001105

References
1. Odds FC (1988) Candida and candidosis. Philadelphia, PA: Bailliere Tindall.
2. Kumamoto CA, Vinces MD (2005) Contributions of hyphae and hypha-co-regulated genes to Candida albicans virulence. Cell Microbiol 7: 1546–1554.
3. Staab JF, Bradway SD, Fidel PL, Sundstrom P (1999) Adhesive and mammaliantransglutaminase substrate properties of Candida albicans Hwp1. Science 283:
1535–1538.
4. Almeida RS, Brunke S, Albrecht A, Thewes S, Laue M, et al. (2008) Thehyphal-associated adhesin and invasin Als3 of Candida albicans mediates iron
acquisition from host ferritin. PLoS Pathog 4: e1000217. doi:10.1371/journal.ppat.1000217.
5. Phan QT, Myers CL, Fu Y, Sheppard DC, Yeaman MR, et al. (2007) Als3 is a
Candida albicans invasin that binds to cadherins and induces endocytosis by hostcells. PLoS Biol 5: e64. doi:10.1371/journal.pbio.0050064.
6. Weissman Z, Kornitzer D (2004) A family of Candida cell surface haem-bindingproteins involved in haemin and haemoglobin-iron utilization. Mol Microbiol
53: 1209–1220.
7. Hogan DA, Sundstrom P (2009) The Ras/cAMP/PKA signaling pathway andvirulence in Candida albicans. Future Microbiol 4: 1263–1270.
8. Rocha CR, Schroppel K, Harcus D, Marcil A, Dignard D, et al. (2001)Signaling through adenylyl cyclase is essential for hyphal growth and virulence in
the pathogenic fungus Candida albicans. Mol Biol Cell 12: 3631–3643.
9. Bahn YS, Sundstrom P (2001) CAP1, an adenylate cyclase-associated proteingene, regulates bud-hypha transitions, filamentous growth, and cyclic AMP
levels and is required for virulence of Candida albicans. J Bacteriol 183:3211–3223.
10. Zou H, Fang HM, Zhu Y, Wang Y (2009) Candida albicans Cyr1, Cap1 and G-actin form a sensor/effector apparatus for activating cAMP synthesis in hyphal
growth. Mol Microbiol.
11. Cassola A, Parrot M, Silberstein S, Magee BB, Passeron S, et al. (2004) Candidaalbicans lacking the gene encoding the regulatory subunit of protein kinase A
displays a defect in hyphal formation and an altered localization of the catalyticsubunit. Eukaryot Cell 3: 190–199.
12. Bockmuhl DP, Krishnamurthy S, Gerads M, Sonneborn A, Ernst JF (2001)
Distinct and redundant roles of the two protein kinase A isoforms Tpk1p andTpk2p in morphogenesis and growth of Candida albicans. Mol Microbiol 42:
1243–1257.
13. Sonneborn A, Bockmuhl DP, Gerads M, Kurpanek K, Sanglard D, et al. (2000)
Protein kinase A encoded by TPK2 regulates dimorphism of Candida albicans.
Mol Microbiol 35: 386–396.
14. Lo HJ, Kohler JR, DiDomenico B, Loebenberg D, Cacciapuoti A, et al. (1997)
Nonfilamentous C. albicans mutants are avirulent. Cell 90: 939–949.
15. Stoldt VR, Sonneborn A, Leuker CE, Ernst JF (1997) Efg1p, an essential
regulator of morphogenesis of the human pathogen Candida albicans, is a
member of a conserved class of bHLH proteins regulating morphogeneticprocesses in fungi. Embo J 16: 1982–1991.
16. Cao F, Lane S, Raniga PP, Lu Y, Zhou Z, et al. (2006) The Flo8 transcriptionfactor is essential for hyphal development and virulence in Candida albicans.
Mol Biol Cell 17: 295–307.
17. Bockmuhl DP, Ernst JF (2001) A potential phosphorylation site for an A-type
kinase in the Efg1 regulator protein contributes to hyphal morphogenesis of
Candida albicans. Genetics 157: 1523–1530.
18. Harcus D, Nantel A, Marcil A, Rigby T, Whiteway M (2004) Transcription
profiling of cyclic AMP signaling in Candida albicans. Mol Biol Cell 15:4490–4499.
19. Braun BR, Johnson AD (1997) Control of filament formation in Candida
albicans by the transcriptional repressor TUP1. Science 277: 105–109.
20. Garcia-Sanchez S, Mavor AL, Russell CL, Argimon S, Dennison P, et al. (2005)
Global roles of Ssn6 in Tup1- and Nrg1-dependent gene regulation in the fungalpathogen, Candida albicans. Mol Biol Cell 16: 2913–2925.
21. Hwang CS, Oh JH, Huh WK, Yim HS, Kang SO (2003) Ssn6, an important
factor of morphological conversion and virulence in Candida albicans. MolMicrobiol 47: 1029–1043.
22. Kadosh D, Johnson AD (2005) Induction of the Candida albicans filamentousgrowth program by relief of transcriptional repression: a genome-wide analysis.
Mol Biol Cell 16: 2903–2912.
23. Khalaf RA, Zitomer RS (2001) The DNA binding protein Rfg1 is a repressor offilamentation in Candida albicans. Genetics 157: 1503–1512.
24. Murad AM, d’Enfert C, Gaillardin C, Tournu H, Tekaia F, et al. (2001)Transcript profiling in Candida albicans reveals new cellular functions for the
transcriptional repressors CaTup1, CaMig1 and CaNrg1. Mol Microbiol 42:981–993.
25. Murad AM, Leng P, Straffon M, Wishart J, Macaskill S, et al. (2001) NRG1
represses yeast-hypha morphogenesis and hypha-specific gene expression inCandida albicans. Embo J 20: 4742–4752.
26. Braun BR, Kadosh D, Johnson AD (2001) NRG1, a repressor of filamentousgrowth in C.albicans, is down-regulated during filament induction. Embo J 20:
4753–4761.
27. Bauer J, Wendland J (2007) Candida albicans Sfl1 suppresses flocculation andfilamentation. Eukaryot Cell 6: 1736–1744.
28. Park YN, Morschhauser J (2005) Tetracycline-inducible gene expression andgene deletion in Candida albicans. Eukaryot Cell 4: 1328–1342.
29. Saville SP, Lazzell AL, Bryant AP, Fretzen A, Monreal A, et al. (2006) Inhibition
of filamentation can be used to treat disseminated candidiasis. AntimicrobAgents Chemother 50: 3312–3316.
30. Enjalbert B, Whiteway M (2005) Release from quorum-sensing molecules
triggers hyphal formation during Candida albicans resumption of growth.
Eukaryot Cell 4: 1203–1210.
31. Klengel T, Liang WJ, Chaloupka J, Ruoff C, Schroppel K, et al. (2005) Fungal
adenylyl cyclase integrates CO2 sensing with cAMP signaling and virulence.Curr Biol 15: 2021–2026.
32. Xu XL, Lee RT, Fang HM, Wang YM, Li R, et al. (2008) Bacterialpeptidoglycan triggers Candida albicans hyphal growth by directly activating the
adenylyl cyclase Cyr1p. Cell Host Microbe 4: 28–39.
33. Shapiro RS, Uppuluri P, Zaas AK, Collins C, Senn H, et al. (2009) Hsp90
orchestrates temperature-dependent Candida albicans morphogenesis via Ras1-PKA signaling. Curr Biol 19: 621–629.
34. Maidan MM, De Rop L, Serneels J, Exler S, Rupp S, et al. (2005) The Gprotein-coupled receptor Gpr1 and the G{alpha} protein Gpa2 act through the
cAMP-protein kinase A pathway to induce morphogenesis in candida albicans.Mol Biol Cell 16: 1971–1986.
35. Miwa T, Takagi Y, Shinozaki M, Yun CW, Schell WA, et al. (2004) Gpr1, aputative G-protein-coupled receptor, regulates morphogenesis and hypha
formation in the pathogenic fungus Candida albicans. Eukaryot Cell 3: 919–931.
36. Sanchez-Martinez C, Perez-Martin J (2002) Gpa2, a G-protein alpha subunit
required for hyphal development in Candida albicans. Eukaryot Cell 1:865–874.
37. Feng Q, Summers E, Guo B, Fink G (1999) Ras signaling is required for serum-induced hyphal differentiation in Candida albicans. J Bacteriol 181: 6339–6346.
38. Zhu Y, Fang HM, Wang YM, Zeng GS, Zheng XD, et al. (2009) Ras1 and Ras2play antagonistic roles in regulating cellular cAMP level, stationary-phase entry
and stress response in Candida albicans. Mol Microbiol 74: 862–875.
39. Hornby JM, Jensen EC, Lisec AD, Tasto JJ, Jahnke B, et al. (2001) Quorum
sensing in the dimorphic fungus Candida albicans is mediated by farnesol. ApplEnviron Microbiol 67: 2982–2992.
40. Davis-Hanna A, Piispanen AE, Stateva LI, Hogan DA (2008) Farnesol anddodecanol effects on the Candida albicans Ras1-cAMP signalling pathway and
the regulation of morphogenesis. Mol Microbiol 67: 47–62.
41. Bastidas RJ, Heitman J, Cardenas ME (2009) The protein kinase Tor1 regulates
adhesin gene expression in Candida albicans. PLoS Pathog 5: e1000294.doi:10.1371/journal.ppat.1000294.
42. Cutler NS, Pan X, Heitman J, Cardenas ME (2001) The TOR signaltransduction cascade controls cellular differentiation in response to nutrients.
Mol Biol Cell 12: 4103–4113.
43. Peterson CL, Laniel MA (2004) Histones and histone modifications. Curr Biol
14: R546–R551.
44. Cairns BR (2005) Chromatin remodeling complexes: strength in diversity,
precision through specialization. Curr Opin Genet Dev 15: 185–190.
45. Sarma K, Reinberg D (2005) Histone variants meet their match. Nat Rev Mol
Cell Biol 6: 139–149.
46. Adkins MW, Howar SR, Tyler JK (2004) Chromatin disassembly mediated by
the histone chaperone Asf1 is essential for transcriptional activation of the yeastPHO5 and PHO8 genes. Mol Cell 14: 657–666.
47. Adkins MW, Tyler JK (2006) Transcriptional activators are dispensable fortranscription in the absence of Spt6-mediated chromatin reassembly of promoter
regions. Mol Cell 21: 405–416.
48. Millar CB, Grunstein M (2006) Genome-wide patterns of histone modifications
in yeast. Nat Rev Mol Cell Biol 7: 657–666.
49. Lee KK, Workman JL (2007) Histone acetyltransferase complexes: one size
doesn’t fit all. Nat Rev Mol Cell Biol 8: 284–295.
50. Lu Y, Su C, Mao X, Raniga PP, Liu H, et al. (2008) Efg1-mediated recruitment
of NuA4 to promoters is required for hypha-specific Swi/Snf binding andactivation in Candida albicans. Mol Biol Cell 19: 4260–4272.
51. Saville SP, Lazzell AL, Monteagudo C, Lopez-Ribot JL (2003) Engineeredcontrol of cell morphology in vivo reveals distinct roles for yeast and filamentous
forms of Candida albicans during infection. Eukaryot Cell 2: 1053–1060.
52. Homann OR, Dea J, Noble SM, Johnson AD (2009) A phenotypic profile of the
Candida albicans regulatory network. PLoS Genet 5: e1000783. doi:10.1371/journal.pgen.1000783.
53. Banerjee M, Thompson DS, Lazzell A, Carlisle PL, Pierce C, et al. (2008)UME6, a novel filament-specific regulator of Candida albicans hyphal extension
and virulence. Mol Biol Cell 19: 1354–1365.
54. Srikantha T, Tsai L, Daniels K, Klar AJ, Soll DR (2001) The histone deacetylase
genes HDA1 and RPD3 play distinct roles in regulation of high-frequencyphenotypic switching in Candida albicans. J Bacteriol 183: 4614–4625.
55. Wu J, Suka N, Carlson M, Grunstein M (2001) TUP1 utilizes histone H3/H2B-specific HDA1 deacetylase to repress gene activity in yeast. Mol Cell 7: 117–126.
56. Lin YY, Qi Y, Lu JY, Pan X, Yuan DS, et al. (2008) A comprehensive syntheticgenetic interaction network governing yeast histone acetylation and deacetyla-
tion. Genes Dev 22: 2062–2074.
57. Pokholok DK, Harbison CT, Levine S, Cole M, Hannett NM, et al. (2005)
Genome-wide map of nucleosome acetylation and methylation in yeast. Cell122: 517–527.
Plasticity of Dimorphism
PLoS Biology | www.plosbiology.org 16 July 2011 | Volume 9 | Issue 7 | e1001105

58. Lee KL, Buckley HR, Campbell CC (1975) An amino acid liquid synthetic
medium for the development of mycelial and yeast forms of Candida Albicans.Sabouraudia 13: 148–153.
59. Wullschleger S, Loewith R, Hall MN (2006) TOR signaling in growth and
metabolism. Cell 124: 471–484.60. Zacchi LF, Gomez-Raja J, Davis DA (2010) Mds3 regulates morphogenesis in
Candida albicans through the TOR pathway. Mol Cell Biol 30: 3695–3710.61. Kucharski R, Maleszka J, Foret S, Maleszka R (2008) Nutritional control of
reproductive status in honeybees via DNA methylation. Science 319:
1827–1830.62. Araki K, Turner AP, Shaffer VO, Gangappa S, Keller SA, et al. (2009) mTOR
regulates memory CD8 T-cell differentiation. Nature 460: 108–112.63. Yilmaz OH, Valdez R, Theisen BK, Guo W, Ferguson DO, et al. (2006) Pten
dependence distinguishes haematopoietic stem cells from leukemia-initiatingcells. Nature 441: 475–482.
64. Hobbs RM, Seandel M, Falciatori I, Rafii S, Pandolfi PP (2010) Plzf regulates
germline progenitor self-renewal by opposing mTORC1. Cell 142: 468–479.65. Castilho RM, Squarize CH, Chodosh LA, Williams BO, Gutkind JS (2009)
mTOR mediates Wnt-induced epidermal stem cell exhaustion and aging. CellStem Cell 5: 279–289.
66. Braun BR, Johnson AD (2000) TUP1, CPH1 and EFG1 make independent
contributions to filamentation in candida albicans. Genetics 155: 57–67.67. Kebaara BW, Langford ML, Navarathna DH, Dumitru R, Nickerson KW, et al.
(2008) Candida albicans Tup1 is involved in farnesol-mediated inhibition offilamentous-growth induction. Eukaryot Cell 7: 980–987.
68. Cruz MC, Goldstein AL, Blankenship J, Del Poeta M, Perfect JR, et al. (2001)Rapamycin and less immunosuppressive analogs are toxic to Candida albicans
and Cryptococcus neoformans via FKBP12-dependent inhibition of TOR.
Antimicrob Agents Chemother 45: 3162–3170.69. Crespo JL, Powers T, Fowler B, Hall MN (2002) The TOR-controlled
transcription activators GLN3, RTG1, and RTG3 are regulated in response tointracellular levels of glutamine. Proc Natl Acad Sci U S A 99: 6784–6789.
70. Carlisle PL, Banerjee M, Lazzell A, Monteagudo C, Lopez-Ribot JL, et al.
(2009) Expression levels of a filament-specific transcriptional regulator aresufficient to determine Candida albicans morphology and virulence. Proc Natl
Acad Sci U S A 106: 599–604.71. Zeidler U, Lettner T, Lassnig C, Muller M, Lajko R, et al. (2009) UME6 is a
crucial downstream target of other transcriptional regulators of true hyphaldevelopment in Candida albicans. FEMS Yeast Res 9: 126–142.
72. Zheng X, Wang Y (2004) Hgc1, a novel hypha-specific G1 cyclin-related protein
regulates Candida albicans hyphal morphogenesis. Embo J 23: 1845–1856.73. Sinha I, Wang YM, Philp R, Li CR, Yap WH, et al. (2007) Cyclin-dependent
kinases control septin phosphorylation in Candida albicans hyphal development.Dev Cell 13: 421–432.
74. Zheng XD, Lee RT, Wang YM, Lin QS, Wang Y (2007) Phosphorylation of
Rga2, a Cdc42 GAP, by CDK/Hgc1 is crucial for Candida albicans hyphalgrowth. EMBO J 26: 3760–3769.
75. Wang A, Raniga PP, Lane S, Lu Y, Liu H (2009) Hyphal chain formation in
Candida albicans: Cdc28-Hgc1 phosphorylation of Efg1 represses cell separation
genes. Mol Cell Biol 29: 4406–4416.
76. Bishop A, Lane R, Beniston R, Chapa-y-Lazo B, Smythe C, et al. (2010) Hyphal
growth in Candida albicans requires the phosphorylation of Sec2 by the Cdc28-
Ccn1/Hgc1 kinase. EMBO J 29: 2930–2942.
77. Uppuluri P, Chaturvedi AK, Srinivasan A, Banerjee M, Ramasubramaniam AK,
et al. (2010) Dispersion as an important step in the Candida albicans biofilm
developmental cycle. PLoS Pathog 6: e1000828. doi:10.1371/journal.ppat.1000828.
78. Pappas PG, Alexander BD, Andes DR, Hadley S, Kauffman CA, et al. (2010)
Invasive fungal infections among organ transplant recipients: results of the
Transplant-Associated Infection Surveillance Network (TRANSNET). Clin
Infect Dis 50: 1101–1111.
79. De Nadal E, Zapater M, Alepuz PM, Sumoy L, Mas G, et al. (2004) The MAPK
Hog1 recruits Rpd3 histone deacetylase to activate osmoresponsive genes.
Nature 427: 370–374.
80. Sertil O, Vemula A, Salmon SL, Morse RH, Lowry CV (2007) Direct role for
the Rpd3 complex in transcriptional induction of the anaerobic DAN/TIR
genes in yeast. Mol Cell Biol 27: 2037–2047.
81. Sharma VM, Tomar RS, Dempsey AE, Reese JC (2007) Histone deacetylases
RPD3 and HOS2 regulate the transcriptional activation of DNA damage-
inducible genes. Mol Cell Biol 27: 3199–3210.
82. Wang A, Kurdistani SK, Grunstein M (2002) Requirement of Hos2 histone
deacetylase for gene activity in yeast. Science 298: 1412–1414.
83. Wang Z, Zang C, Cui K, Schones DE, Barski A, et al. (2009) Genome-wide
mapping of HATs and HDACs reveals distinct functions in active and inactive
genes. Cell 138: 1019–1031.
84. Liu H, Kohler J, Fink GR (1994) Suppression of hyphal formation in Candida
albicans by mutation of a STE12 homolog. Science 266: 1723–1726.
85. Srikantha T, Tsai L, Daniels K, Enger L, Highley K, et al. (1998) The two-
component hybrid kinase regulator CaNIK1 of Candida albicans. Microbiology
144(Pt 10): 2715–2729.
86. Huang G, Wang H, Chou S, Nie X, Chen J, et al. (2006) Bistable expression of
WOR1, a master regulator of white-opaque switching in Candida albicans. Proc
Natl Acad Sci U S A 103: 12813–12818.
87. Lane S, Birse C, Zhou S, Matson R, Liu H (2001) DNA array studies
demonstrate convergent regulation of virulence factors by Cph1, Cph2, and
Efg1 in Candida albicans. J Biol Chem 276: 48988–48996.
88. Wang A, Lane S, Tian Z, Sharon A, Hazan I, et al. (2007) Temporal and spatial
control of HGC1 expression results in Hgc1 localization to the apical cells of
hyphae in Candida albicans. Eukaryot Cell 6: 253–261.
89. Zacchi LF, Schulz WL, Davis DA (2010) HOS2 and HDA1 encode histone
deacetylases with opposing roles in Candida albicans morphogenesis. PLoS One
5: e12171. doi:10.1371/journal.pone.0012171.
Plasticity of Dimorphism
PLoS Biology | www.plosbiology.org 17 July 2011 | Volume 9 | Issue 7 | e1001105
Copyright © 2022 FDOKUMEN




















