Hyperubiquitination of proteins in dilated cardiomyopathy
9
John Weekes 1 * Karen Morrison 1 Anthony Mullen 2 Robin Wait 3 Paul Barton 2 Michael J. Dunn 1 1 Department of Cardiothoracic Surgery, National Heart and Lung Institute, Imperial College School of Medicine, Heart Science Centre, Harefield, UK 2 Department of Cardiothoracic Surgery, National Heart and Lung Institute, Imperial College School of Medicine, London, UK 3 Kennedy Institute of Rheumatology Division, Imperial College School of Medicine, London, UK Hyperubiquitination of proteins in dilated cardiomyopathy Protein degradation by the ubiquitin-proteasome pathway plays an important role in a variety of fundamental cellular processes, including cell cycle regulation, transcription, antigen processing and muscle remodelling. Research into disorders associated with the ubiquitin-proteasome system has been mainly in the field of neurodegenerative diseases. It is however becoming increasingly apparent that defects in the system are responsible for a number of non-neurological pathologies. Based on initial observa- tions made as part of a proteomic analysis of an animal model of dilated cardiomyo- pathy (DCM) which indicated increased activity of the ubiquitin-proteasome system, we sought to determine whether this system was perturbed in hearts of human DCM patients. We studied explanted hearts from 12 DCM, 9 ischaemic (IHD) and 12 unused donor hearts. Protein expression was examined using two-dimensional polyacryl- amide gel electrophoresis, Western blotting and immunohistochemistry. Expression of mRNA was examined using real-time quantitative polymerase chain reaction. Ubiquitinated proteins were affinity purified using a ubiquitin-binding column and iden- tified using peptide mass fingerprinting. All DCM hearts showed significantly higher expression of certain key enzymes of the ubiquitin-proteasome pathway. mRNA expression of ubiquitin carboxyl-terminal hydrolase (UCH) was significantly higher (5.4-fold) in DCM hearts than in control hearts. Myocytes in sections from DCM hearts stained positively for UCH, whereas control hearts were negative. Overall protein ubi- quitination was increased two-fold in DCM relative to IHD hearts and five-fold relative to donor hearts. The ubiquitination of a number of distinct proteins was greatly enhanced in DCM hearts as revealed by anti-ubiquitin Western blots. A number of these proteins were identified using affinity purification and matrix-assisted laser de- sorption/ionisation time-of-flight mass spectrometry. Keywords: Dilated cardiomyopathy / Heart proteins / Protein expression / Ubiquitin / Ubiquitin carboxyl-terminal hydrolase PRO 0347 1 Introduction Dilated cardiomyopathy (DCM) is a severe disease of the heart characterised by impaired systolic function with reduced ejection fraction and increased end-systolic blood volume. Dilation of one or more of the ventricles occurs as the heart adapts to maintain a normal stroke volume [1]. While a large number of factors have been implicated in the disease, including viral infections, toxic agents, genetic factors and chronic alcohol abuse [2], the pathological mechanisms and molecular causes of the disease have yet to be determined. Indeed, the disease is now generally regarded as a syndrome in which a vari- ety of factors give rise to a common cardiac dysfunction. Proteomic analysis of DCM allows us to examine global alterations in protein expression in both human and ani- mal models of the disease [3–6]. Using two-dimensional gel electrophoresis (2-DE) and quantitative computer aided gel analysis we have previously compared the pat- terns of global protein expression in ventricular tissue from patients with DCM to ischaemic heart disease (IHD) and undiseased tissue [4]. This proteomic approach iden- tified a number of quantitative alterations in protein ex- pression specific to the human DCM heart. During the course of this research, we have also used the proteomic approach to investigate global protein expression in bovine hereditary DCM (bDCM), an animal model of human DCM [6]. bDCM is endemic in Switzerland and occurs mainly in the Simmentaler x Red Holstein cross- breed of cattle [7]. The disease occurs in 1- to 4-year- old cattle and progresses rapidly to global heart failure. Correspondence: Dr. Michael J. Dunn, Department of Neu- roscience, Box P045, Institute of Psychiatry, King’s College Lon- don, De Crespigny Park, London SE5 8AF E-mail: [email protected] Fax: +44-207-848-5109 Abbreviations: DCM, dilated cardiomyopathy; IHD, ischaemic heart disease; UCH, ubiquitin carboxyl-terminal hydrolase 208 Proteomics 2003, 3, 208–216 * Present address: Dr. John Weekes, Cytomyx, 6–7 Technopark, Newmarket Road, Cambridge CB5 8PB, UK ª 2003 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim 0173-0835/03/0202–208 $17.501.50/0
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Hyperubiquitination of proteins in dilated cardiomyopathy

John Weekes1*Karen Morrison1
Anthony Mullen2
Robin Wait3
Paul Barton2
Michael J. Dunn1
1Department ofCardiothoracic Surgery,National Heart andLung Institute,Imperial College Schoolof Medicine,Heart Science Centre,Harefield, UK
2Department ofCardiothoracic Surgery,National Heart andLung Institute,Imperial College Schoolof Medicine,London, UK
3Kennedy Institute ofRheumatology Division,Imperial College Schoolof Medicine,London, UK
Hyperubiquitination of proteins in dilatedcardiomyopathy
Protein degradation by the ubiquitin-proteasome pathway plays an important role in avariety of fundamental cellular processes, including cell cycle regulation, transcription,antigen processing and muscle remodelling. Research into disorders associated withthe ubiquitin-proteasome system has been mainly in the field of neurodegenerativediseases. It is however becoming increasingly apparent that defects in the system areresponsible for a number of non-neurological pathologies. Based on initial observa-tions made as part of a proteomic analysis of an animal model of dilated cardiomyo-pathy (DCM) which indicated increased activity of the ubiquitin-proteasome system,we sought to determine whether this system was perturbed in hearts of human DCMpatients. We studied explanted hearts from 12 DCM, 9 ischaemic (IHD) and 12 unuseddonor hearts. Protein expression was examined using two-dimensional polyacryl-amide gel electrophoresis, Western blotting and immunohistochemistry. Expressionof mRNA was examined using real-time quantitative polymerase chain reaction.Ubiquitinated proteins were affinity purified using a ubiquitin-binding column and iden-tified using peptide mass fingerprinting. All DCM hearts showed significantly higherexpression of certain key enzymes of the ubiquitin-proteasome pathway. mRNAexpression of ubiquitin carboxyl-terminal hydrolase (UCH) was significantly higher(5.4-fold) in DCM hearts than in control hearts. Myocytes in sections from DCM heartsstained positively for UCH, whereas control hearts were negative. Overall protein ubi-quitination was increased two-fold in DCM relative to IHD hearts and five-fold relativeto donor hearts. The ubiquitination of a number of distinct proteins was greatlyenhanced in DCM hearts as revealed by anti-ubiquitin Western blots. A number ofthese proteins were identified using affinity purification and matrix-assisted laser de-sorption/ionisation time-of-flight mass spectrometry.
Keywords: Dilated cardiomyopathy / Heart proteins / Protein expression / Ubiquitin / Ubiquitincarboxyl-terminal hydrolase PRO 0347
1 Introduction
Dilated cardiomyopathy (DCM) is a severe disease of theheart characterised by impaired systolic function withreduced ejection fraction and increased end-systolicblood volume. Dilation of one or more of the ventriclesoccurs as the heart adapts to maintain a normal strokevolume [1]. While a large number of factors have beenimplicated in the disease, including viral infections, toxicagents, genetic factors and chronic alcohol abuse [2], thepathological mechanisms and molecular causes of thedisease have yet to be determined. Indeed, the diseaseis now generally regarded as a syndrome in which a vari-
ety of factors give rise to a common cardiac dysfunction.Proteomic analysis of DCM allows us to examine globalalterations in protein expression in both human and ani-mal models of the disease [3–6]. Using two-dimensionalgel electrophoresis (2-DE) and quantitative computeraided gel analysis we have previously compared the pat-terns of global protein expression in ventricular tissuefrom patients with DCM to ischaemic heart disease (IHD)and undiseased tissue [4]. This proteomic approach iden-tified a number of quantitative alterations in protein ex-pression specific to the human DCM heart. During thecourse of this research, we have also used the proteomicapproach to investigate global protein expression inbovine hereditary DCM (bDCM), an animal model ofhuman DCM [6]. bDCM is endemic in Switzerland andoccurs mainly in the Simmentaler x Red Holstein cross-breed of cattle [7]. The disease occurs in 1- to 4-year-old cattle and progresses rapidly to global heart failure.
Correspondence: Dr. Michael J. Dunn, Department of Neu-roscience, Box P045, Institute of Psychiatry, King’s College Lon-don, De Crespigny Park, London SE5 8AFE-mail: [email protected]: +44-207-848-5109
Abbreviations: DCM, dilated cardiomyopathy; IHD, ischaemicheart disease; UCH, ubiquitin carboxyl-terminal hydrolase
208 Proteomics 2003, 3, 208–216
* Present address: Dr. John Weekes, Cytomyx, 6–7 Technopark,Newmarket Road, Cambridge CB5 8PB, UK
ª 2003 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim 0173-0835/03/0202–208 $17.50�.50/0

Proteomics 2003, 3, 208–216 Protein ubiquitination in dilated cardiomyopathy 209
Hearts from diseased animals display important clinicaland biochemical similarities to human DCM [8]. Proteom-ic analysis of bovine left ventricle proteins indicated that35 proteins were differentially expressed in bDCM relativeto undiseased hearts [6]. The most intriguing observa-tion from this study came from the identification of theprotein most highly overexpressed in the diseased hearts.This protein was 7.2-fold more abundant in bDCM heartsthan controls and was identified by N-terminal sequenc-ing as ubiquitin carboxyl-terminal hydrolase (UCH) [6].Enzymes of the UCH family play a key role in the ubiqui-tin-proteasome system of cellular protein degradationand Western blots using an anti-ubiquitin antibody indi-cated that protein ubiquitination was generally elevatedin bDCM. In order to establish whether similar alterationsto the ubiquitin-proteasome system occur in human DCM,we performed a proteomic analysis of human DCM anddonor hearts. Additionally we have applied Western blotanalysis, immunohistochemistry and mRNA analysis toback up the proteomic study and have demonstratedsignificant alterations to this critical pathway that appearto be specific to DCM.
Ubiquitination of proteins plays a central role in nonlyso-somal turnover of cellular proteins [9]. Since the discoveryof ubiquitin in 1975 [10] the ubiquitin-proteasome path-way has emerged to be a complex, highly specific andtightly regulated system of protein degradation. Proteinsubstrates of this pathway are first tagged for degrada-tion by the covalent attachment of chains of ubiquitinmolecules. Proteolysis is then catalysed by the 26S pro-teasome complex in an ATP-dependent manner [11]. Ubi-quitin attachment requires the concerted action of threekey enzymes, termed E1, E2 and E3. Ubiquitin is acti-vated at its C-terminal glycine residue in a reaction requir-ing ATP by the ubiquitin-activating protein (E1). Once acti-vated the ubiquitin bound to E1 is transferred to the sulf-hydryl group of a ubiquitin-conjugating enzyme (E2) [12].A large family of E2 enzymes exists and helps to generatethe specificity of the ubiquitin-proteasome degradationsystem [13]. Finally the activated ubiquitin is transferredfrom E2 to the substrate protein by a ubiquitin-proteinligase (E3), which possesses specific binding sites forubiquitin, E2 and the substrate protein. Ubiquitin can betransferred either to a specific target lysine residue onthe protein, or to lysine-48 of the preceding ubiquitinmolecule to form a chain of polyubiquitin on the targetprotein. It is presumed that the large variety of E3enzymes that exist in the cell play an important role inthe selectivity and regulation of the ubiquitin pathway.
The signal for ubiquitination is present in the target proteinitself, and a variety of structural determinants are recog-nised by different E3s, including the N-terminal residue
of the protein, specific domains such as the ‘destructionbox’, PEST sequences and specific phosphorylated resi-dues [14, 15]. It is apparent therefore that the specificity ofthe E2 and E3 enzymes together with specific ubiquitina-tion signals on the target proteins results in a highly dis-criminating system of protein breakdown. Upon additionof polyubiquitin the target protein is degraded by the 26Sproteasome; a highly complex multisubunit, multifunc-tional protease complex [16, 17]. Destruction of proteinsby the 26S proteasome results in the generation of smallpolypeptides, polypeptides attached to polyubiquitin andpolyubiquitin. UCH hydrolyses the bond between theC-terminus of one ubiquitin and lysine 48 of the next toliberate free ubiquitin moieties which can re-enter thepathway. This enzyme also has a critical role in the supplyof free ubiquitin to the cell, as ubiquitin is only expressedas polyubiquitin or as a ubiquitin fusion protein andrequires processing to free ubiquitin by UCH. It is evidenttherefore that any change in the expression, degradationor activity of UCH would have a dramatic effect on thebalance of protein ubiquitination and proteolysis via theproteasome in the cell.
The importance of the ubiquitin-proteasome pathway tothe cell cannot be overstated. Disruption of any of thefourteen genes encoding subunits of the 20S proteasomecore in yeast produces a lethal phenotype [18]. The path-way has been shown to be necessary for cell growth anddifferentiation by regulating the entry and exit to and fromthe mitotic cycle through the degradation of cyclins,cyclin-dependent kinases and cyclin-dependent kinaseinhibitors [19, 20]. Studies have also shown the systemto be associated with many other important functionsincluding antigen processing [21], degradation of tran-scription factors c-jun, c-fos, MyoD, p53, NF-�B, cell sur-face receptors and short lived regulatory proteins [22, 23].
2 Methods
2.1 Protein preparation
Tissue specimens (typically 0.1–0.2 g) were ground to afine powder under liquid N2 using a pestle and mortar.The resulting powder was collected into 1.5 mL micro-centrifuge tubes and homogenised using a handheldhomogeniser for 1 min in 1 mL lysis buffer, containing9.5 M urea, 1% DTT, 2% CHAPS, and 0.8% PharmalytepH 3–10 (Amersham Biosciences, Amersham, UK). Sam-ples were centrifuged at 15 000 rpm for 1 h and the result-ing supernatants collected. Total homogenate proteinconcentration was measured in duplicate using a modifi-cation of the method described by Bradford [24]. The BSAused for preparation of standard curves and the protein

210 J. Weekes et al. Proteomics 2003, 3, 208–216
samples to be measured were made up to 10 �L with lysisbuffer prior to the addition of 10 �L 0.1 M HCl and 80 �LH20.
2.2 2-DE
First-dimension isoelectric focusing (IEF) was performedusing immobilized pH gradient strips (Amersham Bio-sciences), with pH ranges 4–7 L (linear) and 3–10 NL(nonlinear), using an in-gel rehydration method [25].Samples were diluted with rehydration solution contain-ing 8 M urea, 0.5% CHAPS, 0.2% DTT and 0.2% Phar-malyte pH 3–10 prior to rehydration overnight in a re-swelling tray (Amersham Biosciences). For analytical2-D gels the total protein loaded was 100 �g in a totalvolume of 450 �L. The strips were focused at 0.05A/IPGstrip for 60 kVh at 20�C. After IEF the strips were equili-brated in 1.5 M Tris pH 8.8 buffer containing 6 M urea,30% glycerol, 2% SDS and 0.01% Bromophenol blue,with the addition of 1% DTT for 15 min, followed by thesame buffer with the addition of 4.8% iodoacetamidefor 15 min. Second-dimension SDS-PAGE was per-formed using 22 cm 12%T, 2.6%C separating polyacryl-amide gels without a stacking gel using a Hoefer DALTsystem (Amersham Biosciences). The second-dimensionseparation was carried out overnight at 20 mA/gel at 8�Cand was stopped as the Bromophenol blue dyefront justleft the bottom of the gels.
2.3 Protein visualisation, densitometry andcomputer analysis
Analytical 2-D gels were fixed in 50% methanol, 10%acetic acid and silver stained using the Owl 2-D SilverStaining Kit (Insight Biotechnology, Wembley, UK). Thestained 2-D gels were scanned at 100 �m resolutionusing a Molecular Dynamics Personal SI laser densi-tometer (Amersham Biosciences). Western blots werescanned using a GS-710 optically enhanced imagingdensitometer (Bio-Rad, Hemel Hempstead, UK). Gelswere analysed using Melanie II image analysis software[26] running on a Sun Sparc Ultra1 workstation. Afterdetection of spots the gels were aligned, landmarkedand matched. Gels were then placed into the appropriateexperimental classes and differential analysis performed.All spots that differed between the classes by 50% ormore and passed a Student’s t-test (p � 0.05) wereaccepted as significant differences. All gel spots de-tected as significantly different between the groupswere then highlighted and checked manually to eliminateany artefactual differences due to gel pattern distortions,abnormal silver staining and inappropriately matched orbadly detected spots.
2.4 Real-time quantitative RT-PCR
Human UCH mRNA was detected by PCR amplifica-tion and quantified by 5’ nuclease assay [27] using fluo-rescent labelled TaqMan probes (Applied Biosystems,Warrington, UK) analysed using real time quantitativePCR [28]. Total RNA was extracted from tissue usingthe Qiagen (Crawley, UK) RNeasy minicolumn procedureaccording to the manufacturer’s instructions and RNAquality and quantity was assessed by ethidium bro-mide-agarose gel electrophoresis and by relative ab-sorbance at 260 nm vs. 280 nm. Hexamer-primed cDNAsynthesis was performed on 150 ng of total RNA ina volume of 10 �L using a reverse transcriptase kit(Applied Biosystems, cat # N8080234) and then dilutedto 100 �L with sterile distilled H2O and stored at �20�C.Amplification primers and TaqMan probe for human UCHwere designed using the Primer Express Software(Applied Biosystems) from the published mRNA se-quence (EMBL/GenBank accession number, X04741)with flanking primers located on separate exons andwith the TaqMan probe straddling the exon-exon junc-tion. The primer and TaqMan probe sequences were:forward primer (Tm = 59�C) 5’-GCCACCTCTATGAA-CTTGATGGA-3’, reverse primer (Tm = 58�C) 5’-CTC-ACGCTCGGTGAATTCTCT-3’, TaqMan probe (Tm =70�C) 5’-CAGCGTCCTTCAGCAGGGTGTCC TC-3’ andthe predicted amplicon size was 116 bp. Internal con-trol 18S rRNA primers and TaqMan probe were pro-vided as a pre-optimised kit (Applied Biosystems, cat# 4310893E). PCR reactions were performed using anABI-prism 7700 sequence detector (Applied Biosys-tems). PCR amplifications were performed in a 25 �Lvolume containing 2.5 �L cDNA template in 2x PCRMaster Mix (Applied Biosystems) at 50�C for 2 min,95�C for 10 min followed by 40 cycles of 95�C for15 s and 60�C for 1 min. Each sample was analysedin duplicate. Results were analysed using SequenceDetection Software (Applied Biosystems) and the levelof expression of UCH mRNA normalised to 18S rRNAas outlined in User Bulletin #2 provided by Applied Bio-systems.
2.5 Immunohistochemistry and Westernblotting
Heart tissue was fixed in 10% formalin and processed toparaffin wax. Sections (7 �m) were cut and subsequentlystained for UCH using the rabbit anti-PGP-9.5 antibody(reactive with different isoforms of UCH) at a dilution of1:500 with a biotin-avidin horseradish peroxidase detec-tion system. Controls were performed by omission ofthe PGP-9.5 antibody. Standard Western blotting proto-
Proteomics 2003, 3, 208–216 Protein ubiquitination in dilated cardiomyopathy 211
cols were used except for anti-ubiquitin blots, which wereautoclaved for 30 min at 115�C before blocking with 5%BSA.
2.6 S5a-GST fusion protein purification andidentification of ubiquitinated proteins
Purified S5a-GST attached to glutathione-agarose waspurchased from Affiniti Research Products (Mamhead,UK) and processed according to the protocol supplied.Samples previously extracted from DCM heart using lysisbuffer were loaded onto the column in modified lysisbuffer (4.5 M urea, 0.5% DTT, 1% CHAPS, 5 mM MnCl2,1 �M ubiquitin aldehyde, 50 �M N-acetyl-L-leucyl-L-leu-cyl-L-norleucinal, Complete protease inhibitor cocktail(Roche Diagnostics, Lewes, UK). Proteins were allowed tobind for 30 min at room temperature. The column (0.5 mL)was spun at 2000 rpm, the flow-through collected andresin washed with 6 column volumes of modified lysisbuffer. Bound proteins were eluted with 4 column volumesof modified lysis buffer containing 0.4 M NaCl. Fractionswere concentrated and desalted using Eppendorf centri-fugal filter tubes (4 kDa cutoff). Total eluted proteins (10%of load) plus 100 �g of flowthrough were separated by2-DE. The resulting 2-D gels were stained using a modifiedversion of the PlusOne silver staining kit (Amersham Bio-sciences) that is compatible with MS [29]. Proteins wereexcised from the gels, digested with trypsin and identifiedby peptide mass profiling using MALDI-TOF mass spec-trometry essentially as described in [29].
3 Results
3.1 Global protein expression and UCHabundance in the human DCM heart
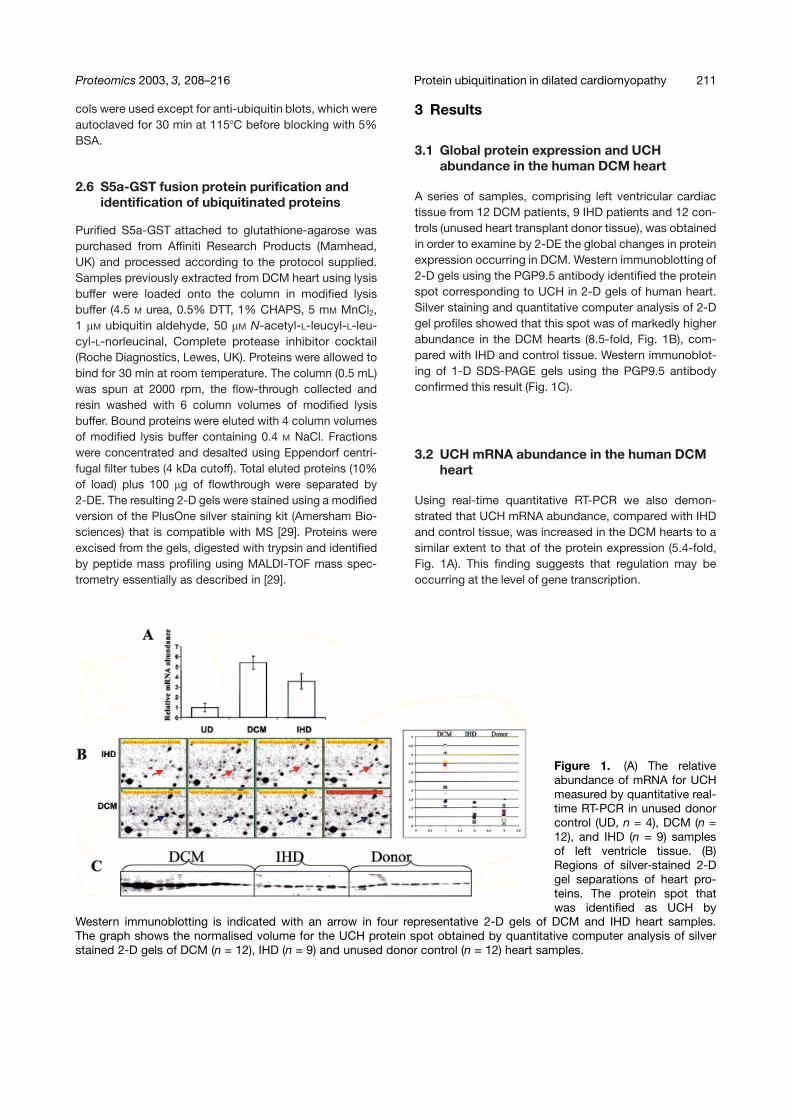
A series of samples, comprising left ventricular cardiactissue from 12 DCM patients, 9 IHD patients and 12 con-trols (unused heart transplant donor tissue), was obtainedin order to examine by 2-DE the global changes in proteinexpression occurring in DCM. Western immunoblotting of2-D gels using the PGP9.5 antibody identified the proteinspot corresponding to UCH in 2-D gels of human heart.Silver staining and quantitative computer analysis of 2-Dgel profiles showed that this spot was of markedly higherabundance in the DCM hearts (8.5-fold, Fig. 1B), com-pared with IHD and control tissue. Western immunoblot-ing of 1-D SDS-PAGE gels using the PGP9.5 antibodyconfirmed this result (Fig. 1C).
3.2 UCH mRNA abundance in the human DCMheart
Using real-time quantitative RT-PCR we also demon-strated that UCH mRNA abundance, compared with IHDand control tissue, was increased in the DCM hearts to asimilar extent to that of the protein expression (5.4-fold,Fig. 1A). This finding suggests that regulation may beoccurring at the level of gene transcription.
Figure 1. (A) The relativeabundance of mRNA for UCHmeasured by quantitative real-time RT-PCR in unused donorcontrol (UD, n = 4), DCM (n =12), and IHD (n = 9) samplesof left ventricle tissue. (B)Regions of silver-stained 2-Dgel separations of heart pro-teins. The protein spot thatwas identified as UCH by
Western immunoblotting is indicated with an arrow in four representative 2-D gels of DCM and IHD heart samples.The graph shows the normalised volume for the UCH protein spot obtained by quantitative computer analysis of silverstained 2-D gels of DCM (n = 12), IHD (n = 9) and unused donor control (n = 12) heart samples.

212 J. Weekes et al. Proteomics 2003, 3, 208–216
3.3 Protein ubiquitination in the human DCMheart
In order to determine whether the up-regulation of UCH inthe human DCM heart leads to an increase in protein-ubi-quitin conjugation, Western immunoblotting using an anti-body reactive with human ubiquitin were performed. Theresults of these experiments (Fig. 2A) indicated that totalprotein-ubiquitin conjugation in DCM was two-fold higherthan that seen in IHD heart samples and five-fold greaterthan in control donor heart tissue (Fig. 2C). An unex-pected feature of these blots is the presence of specificproteins that are ubiquitinated only in the DCM hearts.The ubiquitination of a protein with a molecular mass of16 kDa (Fig. 2A) is increased by over 15-fold in the DCMgroup relative to IHD and controls (Fig. 2B). A protein ofmolecular mass 24.5 kDa (Fig. 2A) is also hyperubiquiti-nated in the DCM group by almost ten-fold comparedwith the IHD and controls. Closer examination of thehigher molecular mass proteins reveals a number of addi-tional proteins, the ubiquitination of which is specificallyincreased in the DCM hearts. The expression of the E1
and E2 enzyme components of the ubiquitin-proteasomesystem is also increased in DCM hearts as measuredusing Western immunoblotting with specific polyclonalantibodies (data not shown).
3.4 Immunohistochemistry
Serial sections of DCM and unused donor hearts wereproduced from formalin-fixed, paraffin embedded tissueand probed with the UCH antibody together with the rele-vant controls. Positive reactions to small nerves were visi-ble in most sections (Fig. 3B) but showed no obviousincrease in abundance in the DCM samples (Fig. 3A). Incontrast cardiac myocytes staining strongly for UCHwere sporadically present throughout most of the DCMsections (Fig. 3A), whilst the donor heart myocytes werealmost completely negative (Fig. 3B). A small number ofpositive cardiac myocytes were seen in the IHD sections(data not shown) but were generally fewer in number thanin the DCM sections.
Figure 2. (A) Western immunoblots of 1-D SDS-PAGE separations of human heart proteins probedwith an antibody reactive with human ubiquitin. The protein bands at 24.5 kDa (**) and 16 kDa (*) thatare hyperubiquitinated in the DCM heart are indicated. (B) Bar graphs of the normalised quantity ofthe 16 kDa band showing that it is increased by more than 15-fold in the DCM group relative to IHDand control samples. (C) Bar graphs of the total ubiquitinated cardiac proteins showing an increase inDCM of five-fold compared with controls and of two-fold compared with IHD.

Proteomics 2003, 3, 208–216 Protein ubiquitination in dilated cardiomyopathy 213
Figure 3. Immunohistochemis-try of sections from humanDCM (A) and unused donorcontrol (B) heart probed withthe PGP9.5 antibody reactivewith multiple isoforms of UCH.In the control heart, cardiacmyocytes are unstained (B), buta proportion of these are stainedstrongly for UCH in the DCMheart (A). Positively stainednerves are indicated with arrowsin the control heart (B).
3.5 Purification and identification ofubiquitinated proteins
Affinity purification of ubiquitinated proteins using an S5a-agarose column and separation of these proteins by 2-DE(Fig. 4) indicated that over 200 distinct proteins werebound to the column, either directly through attached ubi-quitin chains or by association with other bound ubiquiti-
nated proteins. Less than half of these proteins weredeemed of sufficient abundance on the 2-D gels to pro-cess for protein identification by peptide mass profilingusing MALDI-TOF MS. A total of 27 proteins were identi-fied by this approach and their identities are listed inTable 1. The identified proteins, together with a numberof smaller, unidentified protein spots are shown in the2-DE gel in Fig. 4.
Figure 4. IPG 3–10 NL 2-D gelseparation of cardiac proteinspurified by S5a-agarose affinitychromatography and visualisedby silver staining. The identitiesof the proteins established byMALDI-TOF MS (Table 1) areindicated.

214 J. Weekes et al. Proteomics 2003, 3, 208–216
Table 1. Proteins purified by S5a-agarose affinity chromatography and identified by peptide mass profiling using MALDI-TOF MS. These proteins are shown on the 2-D gel in Fig. 4. The arrow in the column “DCM” indicates that theseproteins have previously been shown to be present at reduced abundance [4].
SSP Identification SWISS-PROTprotein name
SWISS-PROTaccession number
DCM
21 Aconitate hydratase (mitochondrial) ACON_HUMAN Q9979822 Aconitate hydratase (mitochondrial) ACON_HUMAN Q9979872 Human serum albumin ALBU_HUMAN P0276873 Human serum albumin ALBU_HUMAN P02768
128 Dihydrolipoamide dehydrogenase precursor DLDH_HUMAN P09622140 ATP synthase beta chain precursor (mitochondrial) ATPB_HUMAN P06576142 ATP synthase alpha chain (mitochondrial) ATPA_HUMAN P25705148 ATP synthase alpha chain (mitochondrial) ATPA_HUMAN P25705165 Myosin heavy-chain (fragment) MYH7_HUMAN P12883177 Long-chain enoyl CoA hydratase ECHA_HUMAN P40939180 C-terminal half of gelsolin (G4-G6) bound to actin GELS_HUMAN P06396182 C-terminal half of gelsolin (G4-G6) bound to actin GELS_HUMAN P06396196 C-terminal half of gelsolin (G4-G6) bound to actin GELS_HUMAN P06396206 Creatine kinase precursor (mitochondrial) KCRS_HUMAN P17540209 Isocitrate dehydrogenase precursor (mitochondrial) IDHA_HUMAN P50213210 Creatine kinase M-chain KCRM_HUMAN P06732212 Creatine kinase M-chain KCRM_HUMAN P06732229 Aspartate aminotransferase precursor (mitochondrial) AATM_HUMAN P00505234 Aldolase A ALFA_HUMAN P04075247 Aminopeptidase N ALFA_HUMAN P04075248 Glyceraldehyde-3-phosphate dehydrogenase G3P1_HUMAN P00354249 Glyceraldehyde-3-phosphate dehydrogenase G3P1_HUMAN P00354250 Malate dehydrogenase precursor (mitochondrial) MDHM_HUMAN P40926294 Alpha crystallin B chain CRAB_HUMAN P02511298 Myoglobin MYG_HUMAN P02144299 Myoglobin MYG_HUMAN P02144300 Haemoglobin beta chain HBB_HUMAN P02023
4 Discussion
UCH has classically been regarded as a marker of nervecells. However it is becoming increasingly apparent thatenzymes belonging to the UCH family are present in avariety of cells and tissues [30, 31]. Although we consid-ered it unlikely that the magnitude of increase in UCHseen in the DCM heart would be due to increased innerva-tion of the diseased heart, we thought it prudent to ruleout such a phenomenon before proceeding with furtherinvestigations. As can be seen from Fig. 3, discretecardiac myocytes staining positive for the presence ofUCH are present in sections from DCM hearts (Fig. 3A),whilst in donor hearts only nerve tissue appeared positive(Fig. 3B).
The results of the studies detailed above clearly demon-strate that protein ubiquitination is greatly increased inDCM cardiac tissue relative to undiseased control hearts.It is also clear that protein ubiquitination is increased inDCM relative to IHD hearts, with proteins that are ubiqui-
tinated in DCM not present in either IHD or undiseasedcontrol hearts (Fig. 2). The increase in ubiquitination inIHD relative to control hearts may be due to an accumula-tion of ubiquitinated proteins, caused by inhibition of theproteasome observed during oxidative stress [32] andcoronary occlusion/reperfusion [33].
It is evident from these studies that the increase in expres-sion of key components of the ubiquitin-proteasome sys-tem in the DCM heart leads to a marked increase in pro-tein ubiquitination in general, as well as dramatic hyper-ubiquitination of certain individual protein species. Proteindegradation by the ubiquitin-proteasome system is ahighly regulated process in the cell. The specificity of thesystem for targeting proteins for ubiquitination and sub-sequent degradation by the proteasome approachesthat seen in phosphorylation and is thought to be broughtabout by the specificity of the ubiquitin ligases. Thiswould appear logical, given the number of essential pro-cesses upon which the ubiquitin system has an effect. It isextremely likely therefore that any disruption to the ubi-

Proteomics 2003, 3, 208–216 Protein ubiquitination in dilated cardiomyopathy 215
quitin-proteasome system would have profound implica-tions to the cell, tissue or organ in which this occurs.
Several areas of human pathology have been linked withthe ubiquitin system, including chronic degenerative dis-eases of the nervous system and muscle breakdown,tumour biology, cell atrophy, programmed cell death, can-cer and antigen presentation to the immune system [34].It is also becoming increasingly evident that ubiquitin-proteasome degradation is involved in skeletal musclebreakdown caused by cancer cachexia, muscle denerva-tion, starvation/malnutrition and other pathological states[35]. Ubiquitin-mediated proteolysis may play a centralrole in the control of protein turnover during skeletal mus-cle breakdown and remodelling and this process is likelyto be conserved in cardiac muscle. Recently Liu et al. [36]have demonstrated that ubiquitin mRNA is increased by50% in streptozotocin-induced diabetic mouse hearts, al-though the amount of free ubiquitin was unchanged. Ubi-quitin is thought to play a major role in protein wasting inskeletal muscle [37]. It is tempting, therefore, to speculatethat an up-regulation of ubiquitination in the DCM heartmay be responsible for the changes in cardiac muscleprotein expression seen during adaptation to maintainnormal stroke volume. The mechanisms leading to anincreased expression of UCH in DCM are unknown.Many genes are up-regulated in heart failure and hyper-trophy, and this often corresponds to re-expression ofgenes associated with foetal cardiac development. Wehave evidence that UCH is readily detected in humanfoetal heart, with a clear decline in expression with devel-opmental age (Weekes et al., unpublished). As such, theexpression profile of UCH fits into the paradigm of rever-sion to a foetal pattern of gene expression in diseasedadult heart.
Activation of the ubiquitin-proteasome system has beenshown to occur after ischaemic damage to neuronal tis-sue [38]. This may also be the case for cardiac ischaemiaand may be part of a cytoprotective mechanism thatinvolves the major stress proteins. If the proteolytic sys-tem remains activated after the initial ischaemic episodethis may be responsible for hyperubiquitination of pro-teins and their subsequent proteolysis. Proteolysis ofmyosin light chain 2 and troponin T and I has recentlybeen implicated in the structural changes in myofilamentproteins found in DCM hearts [39]. Proteolysis of cardiactroponin I has been correlated with stunned myocardium[40] and expression of the major degradation product oftroponin I in transgenic mice leads to diminished contrac-tility and dilation of the ventricles [41]. Although unrelatedto ubiquitination, it is evident that proteolysis underliespathogenesis in these model systems of heart failure.
Perhaps the most intriguing aspect of this work lies in theidentification of the hyper-ubiquitinated proteins them-selves. Figure 2 shows the complex pattern of anti-ubiquitin reactivity seen on Western immunoblots ofSDS-PAGE separations of DCM heart protein extracts.By employing an affinity purification approach in order tofractionate ubiquitinated proteins followed by theirseparation by 2-DE, we have identified a number of theubiquitinated proteins from the DCM heart (Fig. 4). Com-paring the identities of these ubiquitinated proteins to ourpreviously established database of proteins that are dif-ferentially expressed in the DCM heart [4], it is evidentthat a large proportion of these proteins appear in bothlists (Table 1). More striking is the fact that all of the pro-teins found to be ubiquitinated in the present study arepresent at decreased abundance in the DCM heart. Thisevidence strengthens our hypothesis that inappropriateubiquitin conjugation leads to the proteolysis and deple-tion of certain proteins in the DCM heart. Indeed, a num-ber of these proteins have previously been shown to be ofthe utmost importance in determining the phenotype ofcertain hereditary forms of both bovine [42] and human[43] DCM. It is possible, therefore, that the depletion ofthese proteins due to inappropriate ubiquitin conjuga-tion and mutation of these proteins leads to the commonphenotype of DCM. The finding that UCH is present inmyocytes from DCM hearts, together with the markedincrease in protein ubiquitination seen in the same tissueis a novel and interesting observation. This expands ourknowledge of the processes leading to loss of normalcellular activity in the diseased heart and ultimately toheart failure. We believe that the ubiquitin-proteasomesystem may represent a new target for therapeutic inter-vention in heart disease.
This work was supported by a grant from the British HeartFoundation. We are grateful to Dr. Robert Layfield, Univer-sity of Nottingham for useful discussions concerning theisolation of hyperubiquitinated proteins.
Received August 30, 2002
5 References
[1] Richardson, P., McKenna, W., Bristow, M., Maisch, B. et al.,Circulation 1996, 93, 841–842.
[2] Abelmann, W. H., in: Schultheiss, H. P. (Ed.), New Concepts inViral Heart Disease, Springer, Berlin 1998, pp. 2–21.
[3] Pleissner, K.-P., Sander, S., Oswald, H., Regitz-Zagrosek, V.,Fleck, E., Electrophoresis 1996, 17, 1386–1392.
[4] Corbett, J. M., Why, H. J., Wheeler, C. H., Richardson, P. J.et al., Electrophoresis 1998, 19, 2031–2042.
[5] Heinke, M. Y., Wheeler, C. H., Chang, D., Einstein, R. et al.,Electrophoresis 1998, 19, 2021–2030.

216 J. Weekes et al. Proteomics 2003, 3, 208–216
[6] Weekes, J., Wheeler, C. H., Yan, J. X., Weil, J. et al., Electro-phoresis 1999, 20, 898–906.
[7] Tontis, A., Zwahlen, R., Lobsiger, C., Luginbuhl, H., Schweiz.Arch. Tierheilkd. 1990, 132, 105–116.
[8] Eschenhagen, T., Deiderich, M., Kluge, S., Mugnussen, O.et al., J. Mol. Cell. Cardiol. 1995, 27, 357–370.
[9] Hershko, A., Curr. Opin. Cell Biol. 1997, 9, 788–799.[10] Goldstein, G., Steed, M., Hammerling, U., Boyse, E. A. et al.,
Proc. Natl. Acad. Sci. USA 1975, 72, 11–15.[11] Ciechanover, A., Heller, H., Elias, S., Haas, A. L., Hershko,
A., Proc. Natl. Acad. Sci. USA 1980, 77, 1365–1368.[12] Jentsch, S., McGrath, J. B., Varshavsky A., Nature 1987,
329, 131–134.[13] Jentsch, S., Ann. Rev. Genet. 1992, 26, 179–207.[14] Bachmair, A., Finley, D., Varshavsky, A., Science 1986, 234,
179–186.[15] Hoyt, M. A., Cell 1997, 91, 149–151.[16] Peters, J.-M., Harris, J. R., Kleinschmidt, J. A., Eur. J. Cell
Biol. 1991, 56, 422–432.[17] Hilt, W., Wolf, D. H., Trends Biochem. Sci. 1996, 21, 96–102.[18] Heinemeyer, W., Kleinschmidt, J. A., Saidowsky, J., Escher,
C. et al., EMBO J. 1991, 10, 555–562.[19] Hershko, A., Ciechanover, A., Heller, H., Haas, A. L., Rose,
I. A., Proc. Natl. Acad. Sci. USA 1980, 77, 1783–1786.[20] King, R. W., Deshaies, R. J., Peters, J. M., Kirschner, M. W.,
Science 1996, 274, 1652–1659.[21] Goldberg, A. L., Rock, K. L., Nature 1992, 357, 375–379.[22] Ciechanover, A., Cell 1994, 79, 13–22.[23] Maki, C. G., Huibregtse, J. M., Howley, P. M., Cancer Res.
1996, 56, 2649–2654.[24] Bradford, M. M., Anal. Biochem. 1976, 72, 248–253.[25] Rabilloud, T., Valette, C., Lawrence, J. J., Electrophoresis
1994, 15, 1552–1558.[26] Appel, R. D., Palagi, P. M., Walther, D., Vargas, R. J. et al.,
Electrophoresis 1997, 18, 2724–2734.
[27] Holland, P. M., Abramson, R. D., Watson, R., Gelfand, D. H.,Proc. Natl. Acad. Sci. USA 1991, 88, 7276–7280.
[28] Gibson, U. E., Heid, C. A., Williams, P. M., Genome Res.1996, 6, 995–1001.
[29] Yan, J. X., Wait, R., Berkelman, T., Harry, R. A. et al., Electro-phoresis 2000, 21, 3666–3672.
[30] Soldatenkov, V., Dritschilo, A., Cancer Res. 1997, 57, 3881–3885.
[31] Soldatenkov, V., Prasad, S., Voloshin, Y., Dritschilo, A., CellDeath Differ. 1998, 5, 307–312.
[32] Okada, K., Wangpoengtrakul, C., Osawa, T., Toyokuni, S.et al., J. Biol. Chem. 1999, 274, 23787–23793
[33] Bulteau, A. L., Lundberg, K. C., Humphries, K. M., Sadek,H. A. et al., J. Biol. Chem. 2001, 276, 30057–30063
[34] Peters, J.-M., Harris, J. R., Finlay, D., Ubiquitin and the Bio-logy of the Cell, Kluwer, London 1998, pp. 191–452.
[35] Pallares-Trujillo, J., Agell, N., Garcia-Martinez, C., Lopez-Soriano, F. J., Argiles, J. M., Med. Res. Rev. 1997, 17,139–161.
[36] Liu, Z., Miers, W. R., Wei, L., Barrett, E. J., Biochem. Bio-phys. Res. Com. 2000, 276, 1255–1260.
[37] Solomon, V., Goldberg, A., J. Biol. Chem. 1996, 271, 26690–26697.
[38] Jellinger, K. A., J. Neural. Transm. 1996, 47 (Suppl.), 1–29.
[39] Margossian, S. S., Anderson, P. A., Chantler, P. D., Deziel, M.et al., Mol. Cell. Biochem. 1999, 192, 301–313.
[40] Gao, W. D., Atar, D., Liu, Y., Perez, N. G. et al., Circ. Res.1997, 80, 393–399.
[41] Murphy, A. M., Kogler, H., Georgakopoulos, D., McDonough,J. L. et al., Science 2000, 287, 488–491.
[42] Weil, J., Eschenhagen, T., Magnussen, O., Mittmann, C. et al.,J. Mol. Cell. Cardiol. 1997, 29, 743–751.
[43] Marian, A. J., Circ. Res. 2001, 89, 3–5.