
Metformin Inhibits Human Androgen Production by Regulating ...
Upload
independentCategory
view
2download
0
Hepatic Androgen Receptor Suppresses HepatocellularCarcinoma Metastasis Through Modulation of Cell
Migration and AnoikisWen-Lung Ma,1,2* Cheng-Lung Hsu,3* Chun-Chieh Yeh,1 Ming-Heng Wu,2 Chiung-Kuei Huang,2 Long-Bin Jeng,1
Yao-Ching Hung,1 Tze-Yi Lin,1 Shuyuan Yeh,2 and Chawnshang Chang1,2
Early reports suggested androgen/androgen receptor (AR) signals promote hepatocarcinogenesis.However, all antiandrogen clinical trials failed in advanced hepatocellular carcinoma (HCC)without reasonable explanations. We examined AR functions in HCC cancer metastasis in thisstudy. We examined hepatic AR roles in HCC metastasis by comparing liver hepatocyte ARknockout and wildtype in a carcinogen-induced HCCmouse model. We examined tumor histol-ogy, cancer metastatic risks, and cancer survival in vivo, as well as cell anoikis and migrationusing primary hepatic tumor culture in vitro. We also examined therapeutic potentials of ARexpression combined with the molecular targeting agent sorafenib in an HCC metastasis mousemodel. We found a novel cancer phenotype in which mice lacking hepatic AR developed moreundifferentiated tumors and larger tumor size at the metastatic stage. These mice also died earlierwith increased lung metastasis, suggesting that hepatic AR may play dual yet opposite roles topromote HCC initiation but suppress HCC metastasis. Mechanistic dissection found that he-patic AR could enhance anoikis and suppress migration of HCC cells by way of suppression ofp38 phosphorylation/activation and the nuclear factor kappa B (NF-jB)/matrix metallopepti-dase 9 (MMP9) pathway, respectively. In addition, the in vivo preclinical trials concluded that acombination therapy of increased AR expression and reduced multiple-kinase inhibitor (sorafe-nib) exhibited better therapeutic efficacy. Conclusion: Our study demonstrates that AR couldorchestrate intrahepatic signaling hierarchies and cellular behaviors, consequently affect HCCprogression. Results from combination therapy shed light on developing new therapeutic para-digms for battling HCC at later metastatic stages. (HEPATOLOGY 2012;56:176-185)
Hepatocellular carcinoma (HCC) is ranked theseventh cause of cancer death in the UnitedStates and fifth worldwide.1 Androgen and
androgen receptor (AR) signals have been suspected toregulate malignant transformation and progression ofHCC.2,3 However, the amount of AR expression dur-ing HCC remains inconclusive, with reports showingAR is either up- or down-regulated.4-10 Furthermore,clinical studies using antiandrogens had disappointingresults, with few beneficial effects on patients,11,12 oreven worse survival.11
The tumor cell capacity to survive in a detachedenvironment (circulation) or the ability to invade outof primary liver tumor, either homing to distantorgans or micrometastasis to neighboring tissue, canbe critical to cancer metastasis. The recurrence ofHCC, even after hepatic transplantation surgery, couldbe due to re-homing of circulating HCC cells13 resid-ing in the vascular system.14 Because the AR roles in
Abbreviations: AR, androgen receptor; DEN, diethylnitrosamine; HCC,hepatocellular carcinoma; MMP9, matrix metallopeptidase 9; NF-jB: nuclearfactor kappa B.From the 1Sex Hormone Research Centre and Graduate Institute of Clinical
Medical Science, China Medical University/Hospital, Taichung, Taiwan; 2GeorgeWhipple Lab forCancerResearch,Departments of Pathology andUrology and theWilmotCancer Center, University of Rochester Medical Center, Rochester, New York, USA; and3Division of Hematology-Oncology, Department of Internal Medicine, ChangGung Memorial Hospital, Chang Gung University, Taoyuan, Taiwan, ROC.Received August 24, 2011; accepted January 24, 2012.*These authors contributed equally to this study.Supported by George Whipple Professorship Endowment, NIH grant
(CA122295), National Science Council of Taiwan (NSC100-2314-B039001-MY2), Taiwan Department of Health Clinical Trial and Research Center ofExcellence to China Medical University (DOH100-TD-B-111-004), andCMU/H (CMU99-N2-06; CMR-100-159).Address reprint requests to: Chawnshang Chang, George Whipple Lab for
Cancer Research, Departments of Pathology and Urology and the WilmotCancer Center, University of Rochester Medical Center, Rochester, NY 14642.E-mail: [email protected] 2012 by the American Association for the Study of Liver Diseases.View this article online at wileyonlinelibrary.com.DOI 10.1002/hep.25644Potential conflict of interest: Nothing to report.Additional Supporting Information may be found in the online version of
this article.
176

HCC at later metastatic stages remain unclear, using aconditional knockout AR strategy we examined hepaticAR functions in HCC metastasis.
Materials and Methods
Patient Enrollment. From 2005 to 2010, primaryHCC tumors of diameter less than 3 cm and meta-static tumors were collected. Detailed patient informa-tion is described in the Supporting data. Writteninformed consent was obtained from these patients.The studies were approved by the Institutional ReviewBoard of Chang Gung Memorial Hospital, and ChinaMedical University Hospital in Taiwan.Maintenance of Animals, Generation of L-AR2/y
Mice, and HCC Metastasis. All of the animal experi-ments followed the Guide for the Care and Use of Lab-oratory Animals of the U.S. National Institutes ofHealth and with approval from the Department ofLaboratory Animal Medicine at the University ofRochester Medical Center. The strategy to generateflox-AR gene-targeting mice has been described.7
Briefly, we mated male Alb-Cre15 (Cre recombinaseunder control of albumin promoter; Jackson Laborato-ries, B6.Cg-Tg(Alb-cre)21Mgn/J) mice with flox-AR/AR heterozygous (ARflox/X; B6) female mice to pro-duce L-AR�/y males. Each type of transgenic miceexpresses flox-AR and Cre alleles in tail genomicDNA. We genotyped 21-day-old pups from tail snipsby polymerase chain reaction (PCR), as described.16
To induce HCC in the mice liver, we injected 12-day-old pups with HCC initiator, N0-N0-diethylnitros-amine (DEN; 20 mg/kg/mice; Sigma-Aldrich).17 Themale DEN-injected mice were sacrificed at 30, 40, 50,and 60 weeks of age. The nude mice used for tail veininjection experiments were 6-week-old 20-25 g malenude mice (Charles River; Crl: CD1-Foxn1nu Origin).Spontaneous HCC Development and Tail Vein
Injection of HCC Cells for In Vivo Metastasis Assayand Sorafenib Treatments. The carcinogen-inducedmice HCC procedure is further described in the Sup-porting Information and in Ma et al.7 SKAR� andSKARþ cells, parental and AR stable clone of SKhep1cells, respectively, were cultured in a 150-mm flask,maintained in Dulbecco’s modified Eagle’s medium(DMEM) with 10% fetal calf serum (FCS), 1% P/Sand 1% NEAA. When the cells reached �70%-80%confluence they were detached with detaching buffer(0.1 mg/mL trypsin, and 5 mM ethylenediamine-tetra-acetic acid [EDTA]), and 2 � 106 cells/100 lL wereinjected into the tail veins of 8-week-old athymic nude
mice. One month after injection the mice were treatedwith/without sorafenib (Bayer; 30 mg/kg/mice; daily)for another month. The sorafenib stock solutions wereprepared weekly at 4� by dissolving 0.1 g in 4 mLsolvent (Cremophor CL:ethanol ¼ 1:1) and stored at�20�C. For injection, we diluted the 4� sorafenibwith distilled H2O.The experiments consisted of 24 nude mice, ran-
domly assigned to four experimental groups, includingplacebo and sorafenib treatments in SKAR� cells xen-ografted mice; placebo and sorafenib treatments inSKARþ cells xenografted mice. The dosage ofsorafenib was based on the minimal dosages used inmurine models of allograft transplantation. The micerandomized to the sorafenib treatment group weregiven 30 mg/kg/mice in 100 lL by gavage (orally,daily) starting on day 30 after tumor cells injection.All control mice received an equal volume of carriersolution by gavage. The mice were sacrificed 5 weeksafter treatment. At necropsy we observed the visceralorgans and calculated the tumor foci. Both primarytumors and metastatic site tumors were stained forAR and p-p38.Other materials and methods (including mainte-
nance of animals, generation of L-AR�/y mice, HCCmetastasis, in vitro cell culture/maintenance, lentiviral-based gene delivery, reagents, histology, trichromestaining, immunohistochemistry, transfection and re-porter gene assays, cell migration, anoikis assays, statis-tical analysis) are described in the online SupportingMaterials.
Results
Mice Lacking Hepatic AR Developed an EarlierOnset of Metastatic HCC Tumors and Low CancerSurvival Rates. An early study suggested that hepaticAR promotes hepatocarcinogenesis during normal he-patocytes transformation and in mice treated with car-cinogen-DEN.7 This conflicted with the concepts ofclinical trials using antiandrogens to treat HCCpatients.11,18-21 We therefore decided to further dissectthe hepatic AR roles beyond the HCC initiation stage,especially at the HCC later metastatic stage, usingmouse models similar to those we established earlier.7
As expected, we found that male mice lacking liverhepatocyte AR (L-AR�/y, LARKO) developed HCClater as compared with wildtype littermates (ARþ/y,WT), which was consistent with previous studies.7 Yetsurprisingly, we found those L-AR�/y mice died earliercompared with ARþ/y mice (Fig. 1A). Similar resultswith lower survival rates also occurred in female
HEPATOLOGY, Vol. 56, No. 1, 2012 MA ET AL. 177
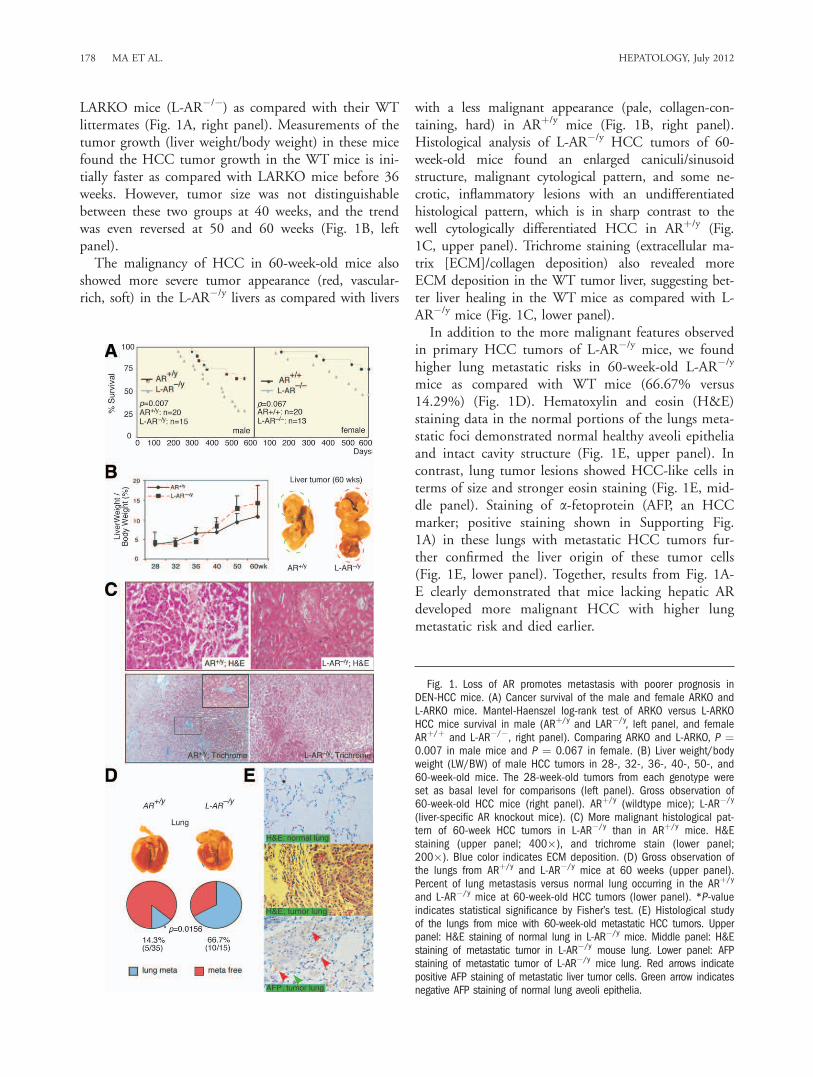
LARKO mice (L-AR�/�) as compared with their WTlittermates (Fig. 1A, right panel). Measurements of thetumor growth (liver weight/body weight) in these micefound the HCC tumor growth in the WT mice is ini-tially faster as compared with LARKO mice before 36weeks. However, tumor size was not distinguishablebetween these two groups at 40 weeks, and the trendwas even reversed at 50 and 60 weeks (Fig. 1B, leftpanel).The malignancy of HCC in 60-week-old mice also
showed more severe tumor appearance (red, vascular-rich, soft) in the L-AR�/y livers as compared with livers
with a less malignant appearance (pale, collagen-con-taining, hard) in ARþ/y mice (Fig. 1B, right panel).Histological analysis of L-AR�/y HCC tumors of 60-week-old mice found an enlarged caniculi/sinusoidstructure, malignant cytological pattern, and some ne-crotic, inflammatory lesions with an undifferentiatedhistological pattern, which is in sharp contrast to thewell cytologically differentiated HCC in ARþ/y (Fig.1C, upper panel). Trichrome staining (extracellular ma-trix [ECM]/collagen deposition) also revealed moreECM deposition in the WT tumor liver, suggesting bet-ter liver healing in the WT mice as compared with L-AR�/y mice (Fig. 1C, lower panel).In addition to the more malignant features observed
in primary HCC tumors of L-AR�/y mice, we foundhigher lung metastatic risks in 60-week-old L-AR�/y
mice as compared with WT mice (66.67% versus14.29%) (Fig. 1D). Hematoxylin and eosin (H&E)staining data in the normal portions of the lungs meta-static foci demonstrated normal healthy aveoli epitheliaand intact cavity structure (Fig. 1E, upper panel). Incontrast, lung tumor lesions showed HCC-like cells interms of size and stronger eosin staining (Fig. 1E, mid-dle panel). Staining of a-fetoprotein (AFP, an HCCmarker; positive staining shown in Supporting Fig.1A) in these lungs with metastatic HCC tumors fur-ther confirmed the liver origin of these tumor cells(Fig. 1E, lower panel). Together, results from Fig. 1A-E clearly demonstrated that mice lacking hepatic ARdeveloped more malignant HCC with higher lungmetastatic risk and died earlier.
Fig. 1. Loss of AR promotes metastasis with poorer prognosis inDEN-HCC mice. (A) Cancer survival of the male and female ARKO andL-ARKO mice. Mantel-Haenszel log-rank test of ARKO versus L-ARKOHCC mice survival in male (ARþ/y and LAR�/y, left panel, and femaleARþ/þ and L-AR�/�, right panel). Comparing ARKO and L-ARKO, P ¼0.007 in male mice and P ¼ 0.067 in female. (B) Liver weight/bodyweight (LW/BW) of male HCC tumors in 28-, 32-, 36-, 40-, 50-, and60-week-old mice. The 28-week-old tumors from each genotype wereset as basal level for comparisons (left panel). Gross observation of60-week-old HCC mice (right panel). ARþ/y (wildtype mice); L-AR�/y
(liver-specific AR knockout mice). (C) More malignant histological pat-tern of 60-week HCC tumors in L-AR�/y than in ARþ/y mice. H&Estaining (upper panel; 400�), and trichrome stain (lower panel;200�). Blue color indicates ECM deposition. (D) Gross observation ofthe lungs from ARþ/y and L-AR�/y mice at 60 weeks (upper panel).Percent of lung metastasis versus normal lung occurring in the ARþ/y
and L-AR�/y mice at 60-week-old HCC tumors (lower panel). *P-valueindicates statistical significance by Fisher’s test. (E) Histological studyof the lungs from mice with 60-week-old metastatic HCC tumors. Upperpanel: H&E staining of normal lung in L-AR�/y mice. Middle panel: H&Estaining of metastatic tumor in L-AR�/y mouse lung. Lower panel: AFPstaining of metastatic tumor of L-AR�/y mice lung. Red arrows indicatepositive AFP staining of metastatic liver tumor cells. Green arrow indicatesnegative AFP staining of normal lung aveoli epithelia.
178 MA ET AL. HEPATOLOGY, July 2012
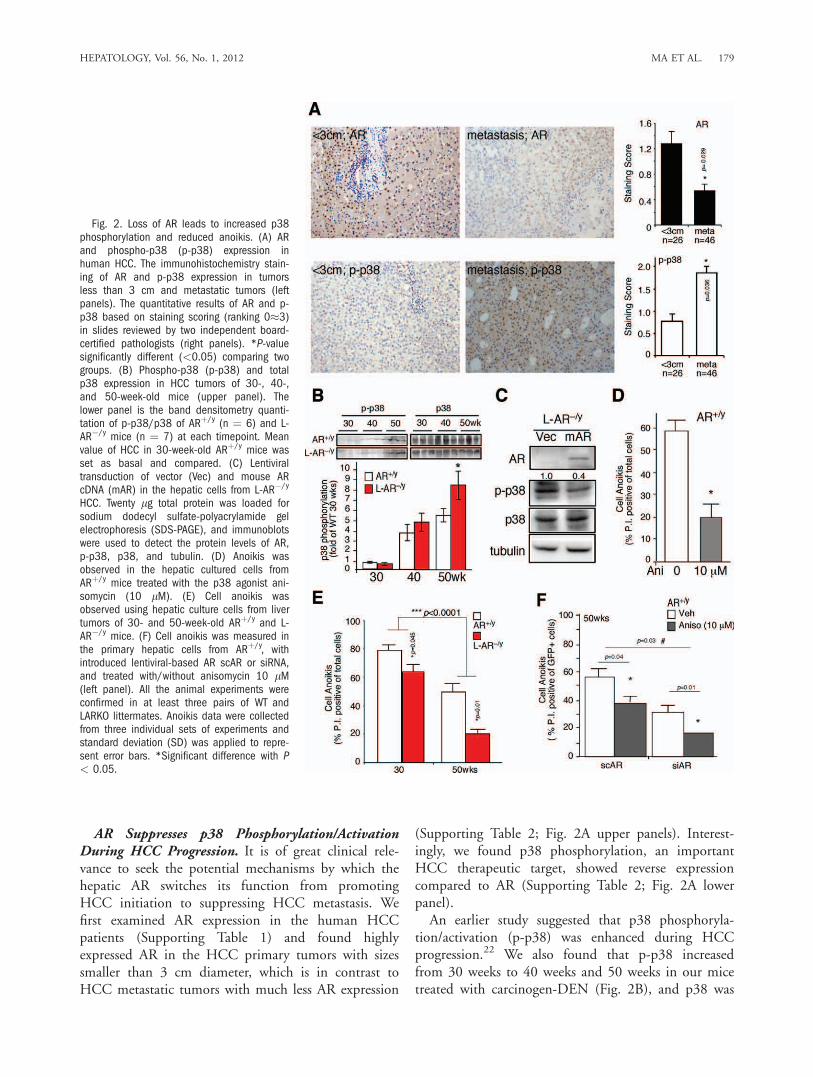
AR Suppresses p38 Phosphorylation/ActivationDuring HCC Progression. It is of great clinical rele-vance to seek the potential mechanisms by which thehepatic AR switches its function from promotingHCC initiation to suppressing HCC metastasis. Wefirst examined AR expression in the human HCCpatients (Supporting Table 1) and found highlyexpressed AR in the HCC primary tumors with sizessmaller than 3 cm diameter, which is in contrast toHCC metastatic tumors with much less AR expression
(Supporting Table 2; Fig. 2A upper panels). Interest-ingly, we found p38 phosphorylation, an importantHCC therapeutic target, showed reverse expressioncompared to AR (Supporting Table 2; Fig. 2A lowerpanel).An earlier study suggested that p38 phosphoryla-
tion/activation (p-p38) was enhanced during HCCprogression.22 We also found that p-p38 increasedfrom 30 weeks to 40 weeks and 50 weeks in our micetreated with carcinogen-DEN (Fig. 2B), and p38 was
Fig. 2. Loss of AR leads to increased p38phosphorylation and reduced anoikis. (A) ARand phospho-p38 (p-p38) expression inhuman HCC. The immunohistochemistry stain-ing of AR and p-p38 expression in tumorsless than 3 cm and metastatic tumors (leftpanels). The quantitative results of AR and p-p38 based on staining scoring (ranking 0�3)in slides reviewed by two independent board-certified pathologists (right panels). *P-valuesignificantly different (<0.05) comparing twogroups. (B) Phospho-p38 (p-p38) and totalp38 expression in HCC tumors of 30-, 40-,and 50-week-old mice (upper panel). Thelower panel is the band densitometry quanti-tation of p-p38/p38 of ARþ/y (n ¼ 6) and L-AR�/y mice (n ¼ 7) at each timepoint. Meanvalue of HCC in 30-week-old ARþ/y mice wasset as basal and compared. (C) Lentiviraltransduction of vector (Vec) and mouse ARcDNA (mAR) in the hepatic cells from L-AR�/y
HCC. Twenty lg total protein was loaded forsodium dodecyl sulfate-polyacrylamide gelelectrophoresis (SDS-PAGE), and immunoblotswere used to detect the protein levels of AR,p-p38, p38, and tubulin. (D) Anoikis wasobserved in the hepatic cultured cells fromARþ/y mice treated with the p38 agonist ani-somycin (10 lM). (E) Cell anoikis wasobserved using hepatic culture cells from livertumors of 30- and 50-week-old ARþ/y and L-AR�/y mice. (F) Cell anoikis was measured inthe primary hepatic cells from ARþ/y, withintroduced lentiviral-based AR scAR or siRNA,and treated with/without anisomycin 10 lM(left panel). All the animal experiments wereconfirmed in at least three pairs of WT andLARKO littermates. Anoikis data were collectedfrom three individual sets of experiments andstandard deviation (SD) was applied to repre-sent error bars. *Significant difference with P< 0.05.
HEPATOLOGY, Vol. 56, No. 1, 2012 MA ET AL. 179

highly phosphorylated/activated in more malignantHCC (Supporting Fig. 1B). More important, wefound loss of hepatic AR in L-AR�/y mice resulted infurther increased p-p38 as compared with that inARþ/y mice at the age of 50 weeks (Fig. 2B). To con-firm that loss of hepatic AR results in enhanced p-p38, primary HCC cells isolated from L-AR�/y micewere cultured and infected with lentiviral-AR cDNA.Figure 2C shows that the addition of AR cDNAresulted in the suppression of p-p38. Together, theresults from Fig. 2A-C suggest that p38 is more activein the advanced state of HCC progression, and loss ofhepatic AR could lead to enhanced p-p38.AR Reverses p38-Mediated Cell Anoikis Resistance.
The pathophysiological consequences of AR-suppressedp-p38 in HCC were then examined by determiningcell anoikis.23 We first demonstrated that p38 couldmodulate cell anoikis in primary cultured HCC cellsisolated from ARþ/y mice. As shown in Fig. 2D, addi-tion of anisomycin, a p38 agonist,24 could reduce cellanoikis significantly. We then examined AR effects oncell anoikis using primary HCC cells from 30-week(early stage) and 50-week (premetastasis stage) -oldmice. We found loss of hepatic AR resulted in differ-ential suppression effects on cell anoikis in 50-week-old tumor cells (WT: L-AR�/y ¼ 55% 6 6%: 20% 64%; P ¼ 0.01) as compared with those at 30 weeks(WT: L-AR�/y ¼ 78% 6 4%: 66% 6 6%; P ¼0.045) (Fig. 2E). The AR differential suppression ontwo stages of cell anoikis reached statistical significance(P < 0.0001). Next, using small interfering RNA(siRNA) knockdown of AR (Supporting Fig. 1C) in50-week-old primary WT tumor cells treated with ani-somycin, we tested AR and p38 effects on cell anoikis.As shown in Fig. 2F, we found that anisomycinreduces anoikis in the 50-week-old WT mice scramble(sc)-treated hepatic cells (57% 6 8% to 39% 6 4%;P ¼ 0.04). However, when comparing cells in the ARsiRNA-treated groups we found that anisomycin has amore dramatic impact on reducing cell anoikis (36%6 4% to 18% 6 0.4%; P ¼ 0.01) (Fig. 2F). TheAR-related anisomycin suppression on cell anoikisreached statistical significance (P ¼ 0.03).Our data consistently show that anisomycin reduces
anoikis and AR enhances anoikis; furthermore, aniso-mycin treatment of sc/siAR-infected WT primary cells(Fig. 2F) showed the anoikis between sc versus siARRNA P ¼ 0.003, which is consistent with our hypothe-sis. When comparing anisomycin effect on scAR (lane1 versus 2; P ¼ 0.04) and siAR (lane 3 versus 4; P ¼0.01) cells, the anisomycin was differentially impactedin sc versus siAR cells. Together, the results from Fig.
2E,F suggest that the hepatic AR enhanced cell anoikis,at least in part, by modulating p38 phosphorylation.AR Enhances Cell Anoikis Through Suppression of
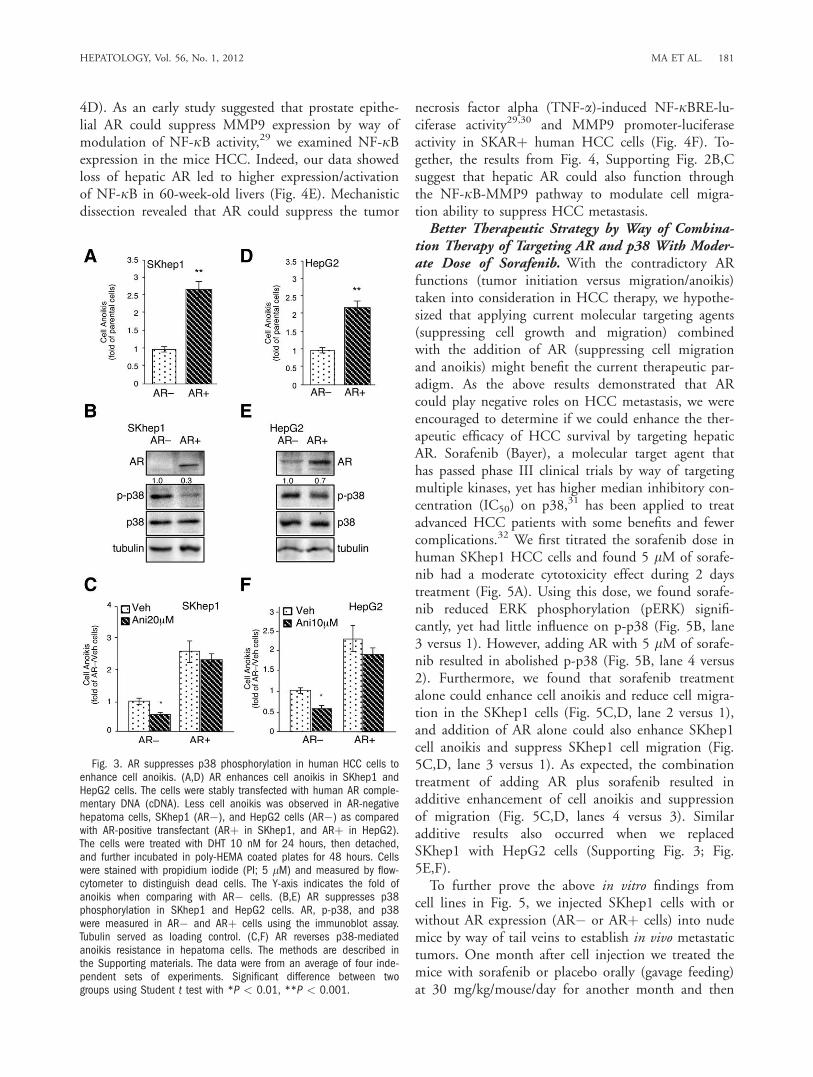
p-p38 in Human HCC Cells. To further confirm thatAR could enhance cell anoikis in the HCC cells, werepeated those experiments using mouse HCC cellswith human HCC cells using previously establishedSKARþ cells7 (SKhep1 cell with AR stable expression)and HepG2-AR cells.25 We demonstrated that addi-tion of AR in the human HCC SKARþ and HepG2-AR cells resulted in increased cell anoikis (Fig. 3A,D).We also demonstrated that addition of AR led to sup-pression of p-p38 (Fig. 3B,E), and addition of the p38agonist anisomycin reduced cell anoikis, whereasexpression of AR reversed that effect (Fig. 3C,F). Aprevious report indicated FasL expression was associ-ated with cell anoikis,26 which was also observed inour system (Supporting Fig. 2A). Furthermore, aniso-mycin could reduce, although addition of AR couldenhance, FasL expression while the cells were detached(Supporting Fig. 2A). Together, the results from Figs.2 and 3 strongly suggested that AR might increase cellanoikis by way of suppression of p-p38. As early stud-ies suggested that cells with anoikis resistance ability ispositively correlated with increased tumor metasta-sis,23,27 it is possible that higher AR expression couldnegatively modulate p38-mediated cell anoikis resist-ance in HCC progression, which might be one reasonwhy hepatic AR could switch from promotion ofHCC initiation to suppression of HCC metastasis atthe metastatic stage.AR Suppresses Cell Migration Through Suppres-
sion of Nuclear Factor kappa B (NF-jB)-MMP9Pathway. In addition to cell anoikis, cell invasiveness(from one foci to multiple foci within the liver) isanother important step contributing to the liver tumormetastasis.27 We noticed that the expression ofMMP9, an important liver cancer migrationmarker,13,28 was higher in the HCC tumors of L-AR�/y
mice as compared with those in ARþ/y mice (Fig. 4A).We then examined the AR effect on cell migration inthe ARþ/y primary cultured HCC cells. As shown inFig. 4B, knocking down AR with siRNA in the ARþ/y
primary cultured HCC cells enhanced cell migration,and addition of AR in L-AR�/y primary cultured HCCcells reduced cell migration.As knockdown of AR also increased migration, but
not cell mobility, in SKARþ human HCC cells (Fig.4C; Supporting Fig. 2C), we further examined MMP9messenger RNA (mRNA)/protein expression in theSKARþ cells and found that the addition of DHTreduced MMP9 mRNA and protein expression (Fig.
180 MA ET AL. HEPATOLOGY, July 2012
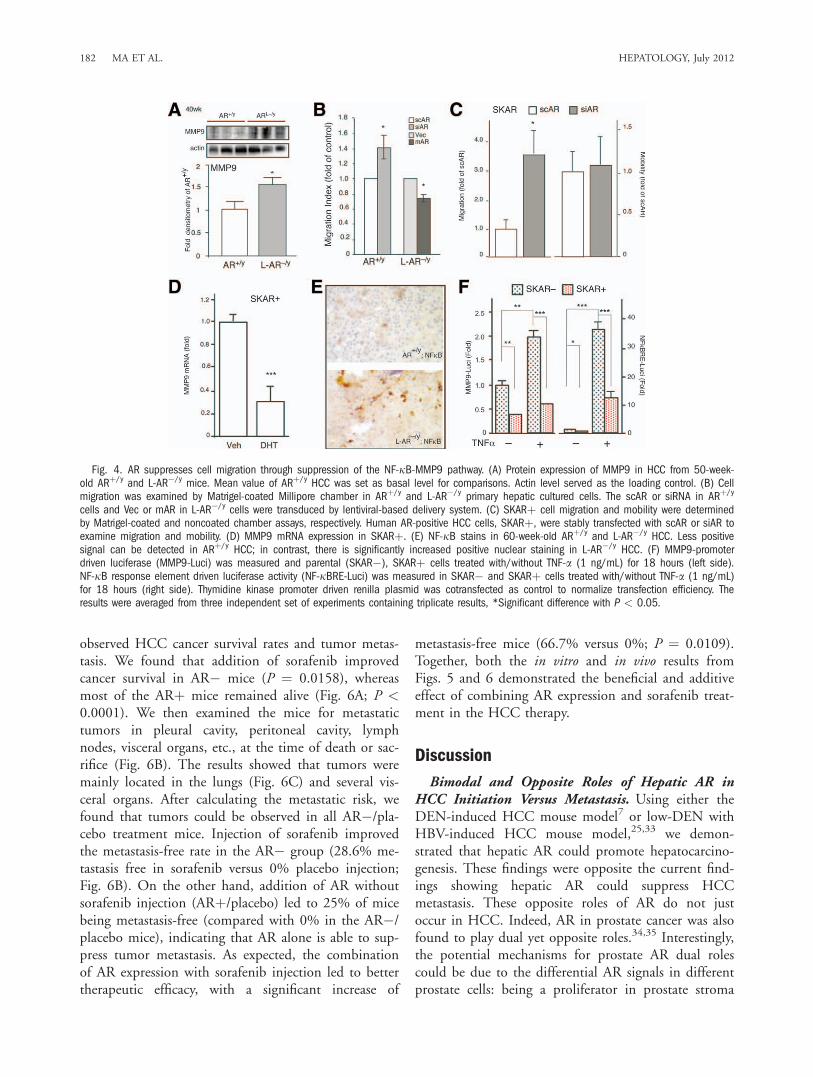
4D). As an early study suggested that prostate epithe-lial AR could suppress MMP9 expression by way ofmodulation of NF-jB activity,29 we examined NF-jBexpression in the mice HCC. Indeed, our data showedloss of hepatic AR led to higher expression/activationof NF-jB in 60-week-old livers (Fig. 4E). Mechanisticdissection revealed that AR could suppress the tumor
necrosis factor alpha (TNF-a)-induced NF-jBRE-lu-ciferase activity29,30 and MMP9 promoter-luciferaseactivity in SKARþ human HCC cells (Fig. 4F). To-gether, the results from Fig. 4, Supporting Fig. 2B,Csuggest that hepatic AR could also function throughthe NF-jB-MMP9 pathway to modulate cell migra-tion ability to suppress HCC metastasis.Better Therapeutic Strategy by Way of Combina-
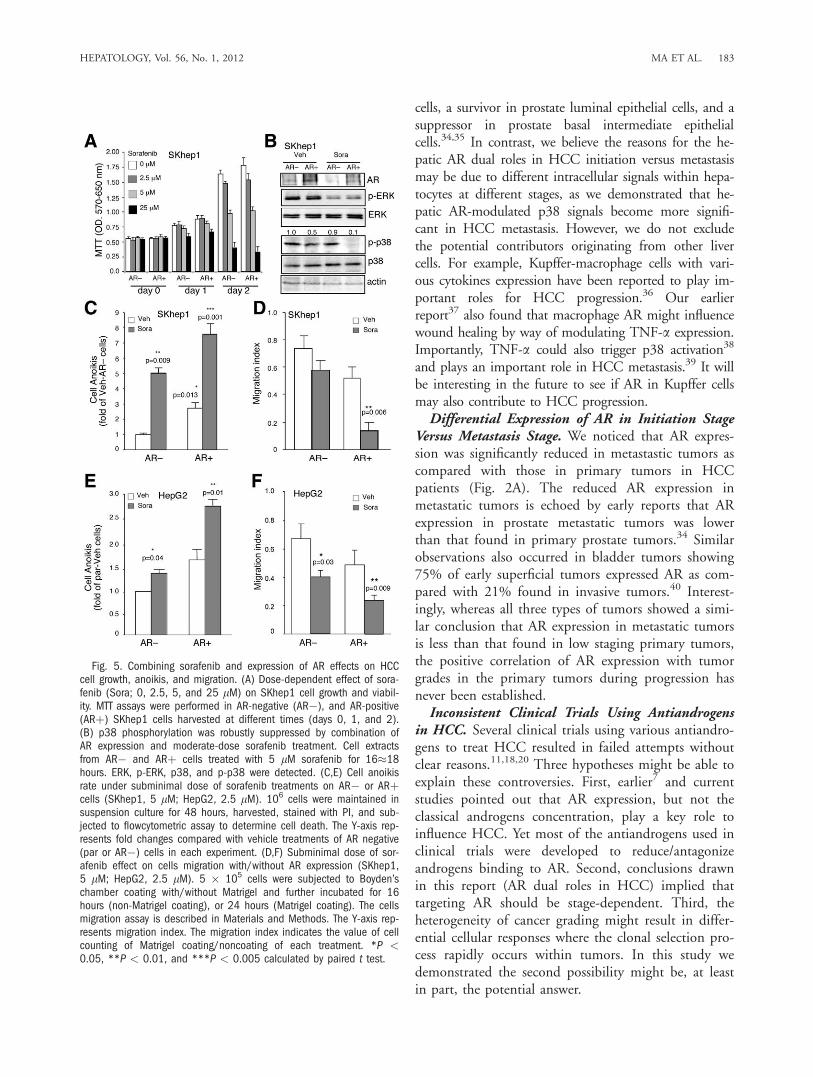
tion Therapy of Targeting AR and p38 With Moder-ate Dose of Sorafenib. With the contradictory ARfunctions (tumor initiation versus migration/anoikis)taken into consideration in HCC therapy, we hypothe-sized that applying current molecular targeting agents(suppressing cell growth and migration) combinedwith the addition of AR (suppressing cell migrationand anoikis) might benefit the current therapeutic par-adigm. As the above results demonstrated that ARcould play negative roles on HCC metastasis, we wereencouraged to determine if we could enhance the ther-apeutic efficacy of HCC survival by targeting hepaticAR. Sorafenib (Bayer), a molecular target agent thathas passed phase III clinical trials by way of targetingmultiple kinases, yet has higher median inhibitory con-centration (IC50) on p38,31 has been applied to treatadvanced HCC patients with some benefits and fewercomplications.32 We first titrated the sorafenib dose inhuman SKhep1 HCC cells and found 5 lM of sorafe-nib had a moderate cytotoxicity effect during 2 daystreatment (Fig. 5A). Using this dose, we found sorafe-nib reduced ERK phosphorylation (pERK) signifi-cantly, yet had little influence on p-p38 (Fig. 5B, lane3 versus 1). However, adding AR with 5 lM of sorafe-nib resulted in abolished p-p38 (Fig. 5B, lane 4 versus2). Furthermore, we found that sorafenib treatmentalone could enhance cell anoikis and reduce cell migra-tion in the SKhep1 cells (Fig. 5C,D, lane 2 versus 1),and addition of AR alone could also enhance SKhep1cell anoikis and suppress SKhep1 cell migration (Fig.5C,D, lane 3 versus 1). As expected, the combinationtreatment of adding AR plus sorafenib resulted inadditive enhancement of cell anoikis and suppressionof migration (Fig. 5C,D, lanes 4 versus 3). Similaradditive results also occurred when we replacedSKhep1 with HepG2 cells (Supporting Fig. 3; Fig.5E,F).To further prove the above in vitro findings from
cell lines in Fig. 5, we injected SKhep1 cells with orwithout AR expression (AR� or ARþ cells) into nudemice by way of tail veins to establish in vivo metastatictumors. One month after cell injection we treated themice with sorafenib or placebo orally (gavage feeding)at 30 mg/kg/mouse/day for another month and then
Fig. 3. AR suppresses p38 phosphorylation in human HCC cells toenhance cell anoikis. (A,D) AR enhances cell anoikis in SKhep1 andHepG2 cells. The cells were stably transfected with human AR comple-mentary DNA (cDNA). Less cell anoikis was observed in AR-negativehepatoma cells, SKhep1 (AR�), and HepG2 cells (AR�) as comparedwith AR-positive transfectant (ARþ in SKhep1, and ARþ in HepG2).The cells were treated with DHT 10 nM for 24 hours, then detached,and further incubated in poly-HEMA coated plates for 48 hours. Cellswere stained with propidium iodide (PI; 5 lM) and measured by flow-cytometer to distinguish dead cells. The Y-axis indicates the fold ofanoikis when comparing with AR� cells. (B,E) AR suppresses p38phosphorylation in SKhep1 and HepG2 cells. AR, p-p38, and p38were measured in AR� and ARþ cells using the immunoblot assay.Tubulin served as loading control. (C,F) AR reverses p38-mediatedanoikis resistance in hepatoma cells. The methods are described inthe Supporting materials. The data were from an average of four inde-pendent sets of experiments. Significant difference between twogroups using Student t test with *P < 0.01, **P < 0.001.
HEPATOLOGY, Vol. 56, No. 1, 2012 MA ET AL. 181
observed HCC cancer survival rates and tumor metas-tasis. We found that addition of sorafenib improvedcancer survival in AR� mice (P ¼ 0.0158), whereasmost of the ARþ mice remained alive (Fig. 6A; P <0.0001). We then examined the mice for metastatictumors in pleural cavity, peritoneal cavity, lymphnodes, visceral organs, etc., at the time of death or sac-rifice (Fig. 6B). The results showed that tumors weremainly located in the lungs (Fig. 6C) and several vis-ceral organs. After calculating the metastatic risk, wefound that tumors could be observed in all AR�/pla-cebo treatment mice. Injection of sorafenib improvedthe metastasis-free rate in the AR� group (28.6% me-tastasis free in sorafenib versus 0% placebo injection;Fig. 6B). On the other hand, addition of AR withoutsorafenib injection (ARþ/placebo) led to 25% of micebeing metastasis-free (compared with 0% in the AR�/placebo mice), indicating that AR alone is able to sup-press tumor metastasis. As expected, the combinationof AR expression with sorafenib injection led to bettertherapeutic efficacy, with a significant increase of
metastasis-free mice (66.7% versus 0%; P ¼ 0.0109).Together, both the in vitro and in vivo results fromFigs. 5 and 6 demonstrated the beneficial and additiveeffect of combining AR expression and sorafenib treat-ment in the HCC therapy.
Discussion
Bimodal and Opposite Roles of Hepatic AR inHCC Initiation Versus Metastasis. Using either theDEN-induced HCC mouse model7 or low-DEN withHBV-induced HCC mouse model,25,33 we demon-strated that hepatic AR could promote hepatocarcino-genesis. These findings were opposite the current find-ings showing hepatic AR could suppress HCCmetastasis. These opposite roles of AR do not justoccur in HCC. Indeed, AR in prostate cancer was alsofound to play dual yet opposite roles.34,35 Interestingly,the potential mechanisms for prostate AR dual rolescould be due to the differential AR signals in differentprostate cells: being a proliferator in prostate stroma
Fig. 4. AR suppresses cell migration through suppression of the NF-jB-MMP9 pathway. (A) Protein expression of MMP9 in HCC from 50-week-old ARþ/y and L-AR�/y mice. Mean value of ARþ/y HCC was set as basal level for comparisons. Actin level served as the loading control. (B) Cellmigration was examined by Matrigel-coated Millipore chamber in ARþ/y and L-AR�/y primary hepatic cultured cells. The scAR or siRNA in ARþ/y
cells and Vec or mAR in L-AR�/y cells were transduced by lentiviral-based delivery system. (C) SKARþ cell migration and mobility were determinedby Matrigel-coated and noncoated chamber assays, respectively. Human AR-positive HCC cells, SKARþ, were stably transfected with scAR or siAR toexamine migration and mobility. (D) MMP9 mRNA expression in SKARþ. (E) NF-jB stains in 60-week-old ARþ/y and L-AR�/y HCC. Less positivesignal can be detected in ARþ/y HCC; in contrast, there is significantly increased positive nuclear staining in L-AR�/y HCC. (F) MMP9-promoterdriven luciferase (MMP9-Luci) was measured and parental (SKAR�), SKARþ cells treated with/without TNF-a (1 ng/mL) for 18 hours (left side).NF-jB response element driven luciferase activity (NF-jBRE-Luci) was measured in SKAR� and SKARþ cells treated with/without TNF-a (1 ng/mL)for 18 hours (right side). Thymidine kinase promoter driven renilla plasmid was cotransfected as control to normalize transfection efficiency. Theresults were averaged from three independent set of experiments containing triplicate results, *Significant difference with P < 0.05.
182 MA ET AL. HEPATOLOGY, July 2012
cells, a survivor in prostate luminal epithelial cells, and asuppressor in prostate basal intermediate epithelialcells.34,35 In contrast, we believe the reasons for the he-patic AR dual roles in HCC initiation versus metastasismay be due to different intracellular signals within hepa-tocytes at different stages, as we demonstrated that he-patic AR-modulated p38 signals become more signifi-cant in HCC metastasis. However, we do not excludethe potential contributors originating from other livercells. For example, Kupffer-macrophage cells with vari-ous cytokines expression have been reported to play im-portant roles for HCC progression.36 Our earlierreport37 also found that macrophage AR might influencewound healing by way of modulating TNF-a expression.Importantly, TNF-a could also trigger p38 activation38
and plays an important role in HCC metastasis.39 It willbe interesting in the future to see if AR in Kupffer cellsmay also contribute to HCC progression.Differential Expression of AR in Initiation Stage
Versus Metastasis Stage. We noticed that AR expres-sion was significantly reduced in metastastic tumors ascompared with those in primary tumors in HCCpatients (Fig. 2A). The reduced AR expression inmetastatic tumors is echoed by early reports that ARexpression in prostate metastatic tumors was lowerthan that found in primary prostate tumors.34 Similarobservations also occurred in bladder tumors showing75% of early superficial tumors expressed AR as com-pared with 21% found in invasive tumors.40 Interest-ingly, whereas all three types of tumors showed a simi-lar conclusion that AR expression in metastatic tumorsis less than that found in low staging primary tumors,the positive correlation of AR expression with tumorgrades in the primary tumors during progression hasnever been established.Inconsistent Clinical Trials Using Antiandrogens
in HCC. Several clinical trials using various antiandro-gens to treat HCC resulted in failed attempts withoutclear reasons.11,18,20 Three hypotheses might be able toexplain these controversies. First, earlier7 and currentstudies pointed out that AR expression, but not theclassical androgens concentration, play a key role toinfluence HCC. Yet most of the antiandrogens used inclinical trials were developed to reduce/antagonizeandrogens binding to AR. Second, conclusions drawnin this report (AR dual roles in HCC) implied thattargeting AR should be stage-dependent. Third, theheterogeneity of cancer grading might result in differ-ential cellular responses where the clonal selection pro-cess rapidly occurs within tumors. In this study wedemonstrated the second possibility might be, at leastin part, the potential answer.
Fig. 5. Combining sorafenib and expression of AR effects on HCCcell growth, anoikis, and migration. (A) Dose-dependent effect of sora-fenib (Sora; 0, 2.5, 5, and 25 lM) on SKhep1 cell growth and viabil-ity. MTT assays were performed in AR-negative (AR�), and AR-positive(ARþ) SKhep1 cells harvested at different times (days 0, 1, and 2).(B) p38 phosphorylation was robustly suppressed by combination ofAR expression and moderate-dose sorafenib treatment. Cell extractsfrom AR� and ARþ cells treated with 5 lM sorafenib for 16�18hours. ERK, p-ERK, p38, and p-p38 were detected. (C,E) Cell anoikisrate under subminimal dose of sorafenib treatments on AR� or ARþcells (SKhep1, 5 lM; HepG2, 2.5 lM). 106 cells were maintained insuspension culture for 48 hours, harvested, stained with PI, and sub-jected to flowcytometric assay to determine cell death. The Y-axis rep-resents fold changes compared with vehicle treatments of AR negative(par or AR�) cells in each experiment. (D,F) Subminimal dose of sor-afenib effect on cells migration with/without AR expression (SKhep1,5 lM; HepG2, 2.5 lM). 5 � 105 cells were subjected to Boyden’schamber coating with/without Matrigel and further incubated for 16hours (non-Matrigel coating), or 24 hours (Matrigel coating). The cellsmigration assay is described in Materials and Methods. The Y-axis rep-resents migration index. The migration index indicates the value of cellcounting of Matrigel coating/noncoating of each treatment. *P <0.05, **P < 0.01, and ***P < 0.005 calculated by paired t test.
HEPATOLOGY, Vol. 56, No. 1, 2012 MA ET AL. 183

Potential New Therapy for HCC. Therapy withsorafenib to treat HCC showed better efficacy withless systemic toxicity.32 However, complications withbleeding or even life-threatening consequences41
remain concerns. Here we found that a combinationof increased AR expression with a moderate dose (5lM) of sorafenib resulted in better efficacy to treatHCC. Early sorafenib phase I clinical trials indicatedthat 6.0�7.7 lM (equal to serum concentration of3.75�4.91 mg/L) of sorafenib resulted in effectivetreatment with tolerable complications.42 Our findingthat AR can be sensitized with a lower dose of sorafe-nib (5 lM) that results in robust therapeutic effectsmay provide an individual approach to treat HCCpatients. Any potential compound(s) to increase/stabi-lize AR expression or technology to increase AR genedelivery into liver might provide a potential method toachieve this purpose.To sum up, there are two major concepts offered in
this study and worthy of future investigation. The firstis to evaluate therapeutic efficiency by distinguishingARþ patients before sorafenib intervention. We expectthat patients with more ARþ in HCC metastatictumors may have a better response and their dose ofsorafenib can be lower to obtain the maximal thera-peutic effect with fewer side effects. The second con-cept is to develop technologies that include increasing/stabilization of AR expression or AR gene delivery in
advanced HCC patients, combined with other molecu-lar targeting agents to evaluate therapeutic effects.
Acknowledgment: We thank Dean Dr. Fu-Jen Tsaifor technical and resources support in MedicalResearch Core Facility, Office of Research & Develop-ment, China Medical University, Taiwan, and KarenWolf for article proofreading. W.L.M. and C.L.H.designed and executed the experiments; C.C.Y.,M.H.W., C.K.H. assisted in some experimental techni-ques; Y.C.H., L.B.J., T.Y.L., and S.Y. helped withpatient tissue histological diagnosis, scientific discus-sion, and editing; C.C. coordinated and supported.
References1. Jemal A, Siegel R, Xu J, Ward E. Cancer statistics. CA Cancer J Clin
2010;60:277-300.
2. Ostrowski JL, Ingleton PM, Underwood JC, Parsons MA. Increased he-patic androgen receptor expression in female rats during diethylnitros-amine liver carcinogenesis. A possible correlation with liver tumordevelopment. Gastroenterology 1988;94:1193-1200.
3. Yoon G, Kim JY, Choi YK, Won YS, Lim IK. Direct activation ofTGF-beta1 transcription by androgen and androgen receptor complexin Huh7 human hepatoma cells and its tumor in nude mice. J CellBiochem 2006;97:393-411.
4. Nagasue N, Yu L, Yukaya H, Kohno H, Nakamura T. Androgen andoestrogen receptors in hepatocellular carcinoma and surrounding liverparenchyma: impact on intrahepatic recurrence after hepatic resection.Br J Surg 1995;82:542-547.
5. Liang L, Lu M, Huang J. [Antiandrogen treatment for nude micemodel with ectopic transplanted human HCC.] Zhonghua Yi Xue ZaZhi 1998;78:299-300.
Fig. 6. Therapeutic evaluation of combining sorafenib and AR expression in HCC metastasis mouse model. (A) Cancer survival of HCC metas-tasis model to observe combining low-dose sorafenib (30 lg/mL/kg) and AR expression. Red line (SKAR� cells injected) and black line(SKARþ cells injected) indicate the cancer survival of mice. Solid line (placebo) and dashed line (sorafenib) indicate differential therapeuticeffects of sorafenib. The cancer survival and P-value were calculated and drawn using the Kaplan-Meier estimator. *P < 0.05, ***P < 0.001.(B) Metastatic tumor in HCC metastasis mouse model. The mice from (A) were sacrificed or autopsied after 1-month treatment and observed formetastasis tumors. The P-values were calculated by two-tailed chi-square test where *P ¼ 0.0109 while comparing metastasis-free lungs in theSKARþ/sorafenib group to the other three treatments. (C) Metastatic lung tumor from tail vein-injected HCC metastasis mouse model. Threegroups of mice lung shown from top to bottom are: AR�/sorafenib, ARþ/placebo, and ARþ/sorafenib. The AR�/placebo group was absentdue to early death during treatment.
184 MA ET AL. HEPATOLOGY, July 2012

6. Tavian D, De Petro G, Pitozzi A, Portolani N, Giulini SM, Barlati S.Androgen receptor mRNA under-expression in poorly differentiatedhuman hepatocellular carcinoma. Histol Histopathol 2002;17:1113-1119.
7. Ma WL, Hsu CL, Wu MH, Wu CT, Wu CC, Lai JJ, et al. Androgenreceptor is a new potential therapeutic target for the treatment ofhepatocellular carcinoma. Gastroenterology 2008;135:947-955, 955e941-945.
8. Barone M, Margiotta M, Scavo MP, Gentile A, Francioso D, PapagniS, et al. Possible involvement of androgen receptor alterations in hepato-carcinogenesis. Dig Liver Dis 2009;41:665-670.
9. Wang AG, Lee KY, Kim SY, Choi JY, Lee KH, Kim WH, et al. Theexpression of estrogen receptors in hepatocellular carcinoma in Koreanpatients. Yonsei Med J 2006;47:811-816.
10. Cho H, Lim IK. Expression of androgen receptor and its implicationin hepatoma cells. Cancer Lett 1997;115:135-140.
11. H�epatocellulaire GdEedTdC. Randomized trial of leuprorelin and flu-tamide in male patients with hepatocellular carcinoma treated with ta-moxifen. HEPATOLOGY 2004;40:1361-1369.
12. Di Maio M, Daniele B, Pignata S, Gallo C, De Maio E, Morabito A,et al. Is human hepatocellular carcinoma a hormone-responsive tumor?World J Gastroenterol 2008;14:1682-1689.
13. Qin LX, Tang ZY. Recent progress in predictive biomarkers for meta-static recurrence of human hepatocellular carcinoma: a review of the lit-erature. J Cancer Res Clin Oncol 2004;130:497-513.
14. Jeng KS, Sheen IS, Tsai YC. Circulating messenger RNA of alpha-feto-protein: a possible risk factor of recurrence after resection of hepatocel-lular carcinoma. Arch Surg 2004;139:1055-1060.
15. Postic C, Shiota M, Niswender KD, Jetton TL, Chen Y, Moates JM,et al. Dual roles for glucokinase in glucose homeostasis as determinedby liver and pancreatic beta cell-specific gene knock-outs using Crerecombinase. J Biol Chem 1999;274:305-315.
16. Yeh S, Tsai MY, Xu Q, Mu XM, Lardy H, Huang KE, et al. Genera-tion and characterization of androgen receptor knockout (ARKO)mice: an in vivo model for the study of androgen functions in selectivetissues. Proc Natl Acad Sci U S A 2002;99:13498-13503.
17. Tejura S, Rodgers GR, Dunion MH, Parsons MA, Underwood JC,Ingleton PM. Sex-steroid receptors in the diethylnitrosamine model ofhepatocarcinogenesis: modifications by gonadal ablation and steroidreplacement therapy. J Mol Endocrinol 1989;3:229-237.
18. Forbes A, Wilkinson ML, Iqbal MJ, Johnson PJ, Williams R. Responseto cyproterone acetate treatment in primary hepatocellular carcinoma isrelated to fall in free 5 alpha-dihydrotestosterone. Eur J Cancer ClinOncol 1987;23:1659-1664.
19. Gupta S, Korula J. Failure of ketoconazole as anti-androgen therapy innonresectable primary hepatocellular carcinoma. J Clin Gastroenterol1988;10:651-654.
20. Matsuura B, Taniguchi Y, Ohta Y. Effect of antiandrogen treatment onchemical hepatocarcinogenesis in rats. J Hepatol 1994;21:187-193.
21. Chao Y, Chan Week, Huang YS, Teng HC, Wang SS, Lui WY, et al.Phase II study of flutamide in the treatment of hepatocellular carci-noma. Cancer 1996;77:635-639.
22. Guo K, Liu Y, Zhou H, Dai Z, Zhang J, Sun R, et al. Involvement ofprotein kinase C beta-extracellular signal-regulating kinase 1/2/p38 mito-gen-activated protein kinase-heat shock protein 27 activation in hepatocel-lular carcinoma cell motility and invasion. Cancer Sci 2008;99:486-496.
23. Frisch SM, Francis H. Disruption of epithelial cell-matrix interactionsinduces apoptosis. J Cell Biol 1994;124:619-626.
24. Cano E, Doza YN, Ben-Levy R, Cohen P, Mahadevan LC. Identifica-tion of anisomycin-activated kinases p45 and p55 in murine cells asMAPKAP kinase-2. Oncogene 1996;12:805-812.
25. Wu MH, Ma WL, Hsu CL, Chen YL, Ou JH, Ryan CK, et al. Andro-gen receptor promotes hepatitis B virus-induced hepatocarcinogenesisthrough modulation of hepatitis B virus RNA transcription. Sci TranslMed;2:32ra35.
26. Laguinge LM, Samara RN, Wang W, El-Deiry WS, Corner G, Augen-licht L, et al. DR5 receptor mediates anoikis in human colorectal carci-noma cell lines. Cancer Res 2008;68:909-917.
27. Sahai E. Illuminating the metastatic process. Nat Rev Cancer 2007;7:737-749.
28. Yuan RH, Jeng YM, Chen HL, Lai PL, Pan HW, Hsieh FJ, et al.Stathmin overexpression cooperates with p53 mutation and osteopontinoverexpression, and is associated with tumour progression, early recur-rence, and poor prognosis in hepatocellular carcinoma. J Pathol 2006;209:549-558.
29. Altuwaijri S, Lin HK, Chuang KH, Lin WJ, Yeh S, Hanchett LA,et al. Interruption of nuclear factor kappaB signaling by the androgenreceptor facilitates 12-O-tetradecanoylphorbolacetate-induced apoptosisin androgen-sensitive prostate cancer LNCaP cells. Cancer Res 2003;63:7106-7112.
30. Yoshizaki T, Sato H, Furukawa M, Pagano JS. The expression of matrixmetalloproteinase 9 is enhanced by Epstein-Barr virus latent membraneprotein 1. Proc Natl Acad Sci U S A 1998;95:3621-3626.
31. Wilhelm SM, Carter C, Tang L, Wilkie D, McNabola A, Rong H,et al. BAY 43-9006 exhibits broad spectrum oral antitumor activity andtargets the RAF/MEK/ERK pathway and receptor tyrosine kinasesinvolved in tumor progression and angiogenesis. Cancer Res 2004;64:7099-7109.
32. Llovet JM, Ricci S, Mazzaferro V, Hilgard P, Gane E, Blanc JF, et al.Sorafenib in advanced hepatocellular carcinoma. N Engl J Med 2008;359:378-390.
33. DeLoia JA, Burk RD, Gearhart JD. Developmental regulation of hepa-titis B surface antigen expression in two lines of hepatitis B virus trans-genic mice. J Virol 1989;63:4069-4073.
34. Niu Y, Altuwaijri S, Lai KP, Wu CT, Ricke WA, Messing EM, et al.Androgen receptor is a tumor suppressor and proliferator in prostatecancer. Proc Natl Acad Sci U S A 2008;105:12182-12187.
35. Niu Y, Chang TM, Yeh S, Ma WL, Wang YZ, Chang C. Differentialandrogen receptor signals in different cells explain why androgen-depri-vation therapy of prostate cancer fails. Oncogene 2010;29:3593-3604.
36. Sander LE, Trautwein C, Liedtke C. Is interleukin-6 a gender-specificrisk factor for liver cancer? HEPATOLOGY 2007;46:1304-1305.
37. Lai JJ, Lai KP, Chuang KH, Chang P, Yu IC, Lin WJ, et al. Monocyte/macrophage androgen receptor suppresses cutaneous wound healing inmice by enhancing local TNF-alpha expression. J Clin Invest 2009.
38. Schindler JF, Monahan JB, Smith WG. p38 pathway kinases as anti-inflammatory drug targets. J Dent Res 2007;86:800-811.
39. Luedde T, Trautwein C. Intracellular survival pathways in the liver.Liver Int 2006;26:1163-1174.
40. Boorjian S, Ugras S, Mongan NP, Gudas LJ, You X, Tickoo SK, et al.Androgen receptor expression is inversely correlated with pathologic tu-mor stage in bladder cancer. Urology 2004;64:383-388.
41. Je Y, Schutz FA, Choueiri TK. Risk of bleeding with vascular endothe-lial growth factor receptor tyrosine-kinase inhibitors sunitinib and sora-fenib: a systematic review and meta-analysis of clinical trials. LancetOncol 2009;10:967-974.
42. Chen HX, Cleck JN. Adverse effects of anticancer agents that targetthe VEGF pathway. Nat Rev Clin Oncol 2009;6:465-477.
HEPATOLOGY, Vol. 56, No. 1, 2012 MA ET AL. 185
Copyright © 2022 FDOKUMEN




















