
Sertraline Treatment of Major Depression in Patients With Acute MI or Unstable Angina
Upload
independentCategory
view
2download
0
Pharmaceuticals and Personal Care Products in the Environment
GROWTH AND DEVELOPMENT OF TADPOLES (XENOPUS LAEVIS) EXPOSED TOSELECTIVE SEROTONIN REUPTAKE INHIBITORS, FLUOXETINE AND SERTRALINE,
THROUGHOUT METAMORPHOSIS
DEANNA E. CONNERS,{ EMILY D. ROGERS,{ KEVIN L. ARMBRUST,{ JEONG-WOOK KWON,{ and
MARSHA C. BLACK*{{Department of Environmental Health Science, University of Georgia, Athens, Georgia 30602, USA
{Mississippi State Chemical Laboratory, Mississippi State University, Mississippi State, Mississippi 39762, USA
(Received 30 September 2008; Accepted 17 February 2009)
Abstract—Selective serotonin reuptake inhibitors (SSRIs) are widely prescribed drugs that are present in sewage effluents and surfacewaters. The objective of the present study was to determine whether low environmentally relevant concentrations of the SSRIsfluoxetine and sertraline could impair growth and development in tadpoles of the African clawed frog (Xenopus laevis) and to evaluateif such effects may be caused by a disruption of the neuroendocrine system. Tadpoles were exposed to SSRIs at concentrations of 0.1,1, and 10 mg/L for 70 d throughout metamorphosis. No effects on deformities were observed. Tadpoles exposed to fluoxetine (10 mg/L)and sertraline (0.1, 1, and 10 mg/L) exhibited reduced growth at metamorphosis. Tadpoles exposed to sertraline (0.1 and 1 mg/L)exhibited an acceleration of development as indicated by an increase in the time to tail resorption. The effects of SSRIs on growth anddevelopment in tadpoles were likely driven by reduced food intake. Reduced feeding rates were observed in SSRI-exposed tadpoles,and nutritional status can influence growth and development in amphibians via effects on the neuroendocrine system. Only sertralinewas capable of causing developmental toxicity in tadpoles at environmentally relevant concentrations. These data warrant additionalresearch to characterize the risks to human health and wildlife from pharmaceutical exposures.
Keywords—Anurans Endocrine disruption Fluoxetine Pharmaceuticals Sertraline
INTRODUCTION
The selective serotonin reuptake inhibitors (SSRIs) fluox-
etine and sertraline are widely prescribed drugs for the
treatment of depression, obsessive compulsive behaviors,
anxiety, and premenstrual dysphoric disorder in humans.
Selective serotonin reuptake inhibitors are an important class
of emerging environmental contaminants along with many
other pharmaceuticals and personal care products [1]. Pres-
ently, the risks associated with environmental exposures to
pharmaceuticals have not been well-characterized.
Low concentrations of SSRIs (#0.1 mg/L) have been
detected in sewage effluents and surface waters [2,3]. Johnson
et al. [4] estimated that predicted environmental concentra-
tions for surface waters may be as high as 0.2 mg/L for
fluoxetine and 0.6 to 1.2 mg/L for sertraline. Fluoxetine and
sertraline were also detected as contaminants in fish tissue [5].
In pregnant women taking prescriptions, these chemicals are
known to contaminate breast milk (fluoxetine ,2–384 mg/L;
sertraline 7–207 mg/L) and infant plasma (fluoxetine ,1–
265 mg/L; sertraline 0–1.5 mg/L) [6,7].
Developmental abnormalities were observed in fish [8]
exposed to low environmentally relevant concentrations of
fluoxetine. Adverse effects on development (e.g., reduced
growth) have been observed in neonatal rats exposed to
fluoxetine [9] and sertraline [10]. In humans, research indicates
that exposure to SSRIs during gestation or via breast milk can
cause developmental toxicity in infants (e.g., hypoglycemia,
respiratory distress, low birth weights, and reduced postnatal
growth) [6,11].
The mechanisms by which SSRIs may cause adverse
developmental effects are currently unknown. The objective
of the present study was to determine whether low environ-
mentally relevant concentrations of fluoxetine and sertraline
can impair growth and development in tadpoles of the African
clawed frog (Xenopus laevis) and to evaluate if such effects may
be caused by a disruption of the neuroendocrine system.
African clawed frogs are ideal organisms to investigate the
ecological and human health implications of exposures to low
concentrations of SSRIs in the environment. Xenopus laevis are
aquatic frogs that are easy to culture and breed in the laboratory,
and standardized protocols exist to explore the effects of
xenobiotics on growth and development of embryos [12] and
tadpoles [13]. Ecologically, frogs are an important test species to
use in toxicity tests because of environmental concern over
chemically induced deformities and declining amphibian popu-
lations [14,15]. In regards to human health, frogs are an
important model for studies of postembryonic organ develop-
ment [16] and chemically mediated thyroid dysfunction [17,18].
Current research indicates that the SSRI fluoxetine may
affect the thyroid system of mammals either by decreasing the
amount of thyrotropin-releasing hormone (TRH) produced by
the hypothalamus or by elevating the concentration of
triiodothyronine (T3), the more potent of two active thyroid
hormones (T3 and T4), in extrathyroidal tissues via effects on
the activities of iodothyronine deiodinases type II and type III
[19,20]. Fluoxetine is also known to affect behaviors controlled
by the production of corticotropin-releasing factor (CRF) in
the hypothalamus of fish and newts [21]. Interestingly, in frogs
CRF is the thyrotrophic hormone responsible for pituitary
production of thyroid-stimulating hormone (TSH), whereas in
mammals TSH release is controlled via TRH [22]. Together* To whom correspondence may be addressed ([email protected]).Published on the Web 7/2/2009.
Environmental Toxicology and Chemistry, Vol. 28, No. 12, pp. 2671–2676, 2009’ 2009 SETAC
Printed in the USA0730-7268/09 $12.00 + .00
2671

these data suggest that there may be a variety of mechanisms
by which SSRIs can interact with the hypothalamus–pituitary–
thyroid axis or the hypothalamus–pituitary–interrenal axis in
frogs and interfere with growth and development (Fig. 1).
Nonendocrine effects (i.e., neurological) may also be an
important contributor to the developmental toxicity of SSRIs
(e.g., serotonin controls left–right patterning in frog and chick
embryos [23] and serotonin plays a trophic role in digestive
processes such as food intake [24] and peristalsis [25]).
In the present study, a randomized block design was used to
expose tadpoles to fluoxetine and sertraline (0.1, 1, and 10 mg/L)
for 70 d throughout development. Biological end points that
were analyzed included mortality, deformities, growth, and
development. Both fluoxetine and sertraline caused develop-
mental toxicity in frogs, but only sertraline was capable of
producing effects at low environmentally relevant concentra-
tions. The type and timing of adverse effects suggest that SSRIs
may disrupt the neuroendocrine system of developing tadpoles.
MATERIALS AND METHODS
Frog breeding
All frog care complied with the University of Georgia’s
Animal Use Guidelines. Two certified breeding pairs of adult
Fig. 1. Hormonal control over morphological traits in developing Xenopus laevis tadpoles and locations where selective serotonin reuptake inhibitors(SSRIs) may interfere with metamorphosis based on amphibian endocrinology information [15,22,39,40] and on information on SSRI and endocrineinteractions [19–21]. CRF 5 corticotropin-releasing factor; TRH 5 thyrotropin-releasing hormone; TSH 5 thyroid-stimulating hormone; ACTH 5adrenocorticotropic hormone; PRL 5 prolactin; TH 5 thyroid hormones T3 and T4.
2672 Environ. Toxicol. Chem. 28, 2009 D.E. Conners et al.

frogs (Xenopus laevis) were obtained from Carolina Biological
Supply Company. Frogs were not fed prior to breeding and
were held for 9 d in a 40-L flow-through aquarium containing
dechlorinated tap water (23uC) (density of four frogs per
1,800 cm3 of water) (12:12 h light/dark cycle). Breeding was
conducted by methods similar to those outlined by the
American Society for Testing and Materials [13]. Briefly,
female frogs (9 and 10 cm in length) were injected with 1,000
International Units (IU) of sterile chorionic gonadotropin
(1 ml total volume) (ChorulonH Intervet, Schering-Plough
Animal Health) via the dorsal lymph sac, and frogs were
placed into individual breeding chambers containing 10 L
FETAX solution (21uC) (water depth of 6 cm above plastic
mesh bottom). Then, male frogs (7 and 8 cm in length) were
injected with 500 IU of chorionic gonadotropin (0.5 ml total
volume) and placed in breeding chambers along with females.
Breeding was allowed to proceed overnight for 14 h. Fertilized
embryos were gently flushed with a plastic transfer pipette into
a glass aquarium containing 40 L of aerated FETAX solution
(23uC). Embryos were allowed to hatch and develop for 3 d
prior to toxicity testing. By 3 d, tadpoles that had reached
Nieuwkoop and Faber [26] stages 42 and 43 of development
and were 7.0 to 8.0 mm in length were used to start the toxicity
tests.
Tadpole exposures
Exposure of tadpoles to SSRIs was conducted similar to the
protocol outlined by Opitz et al. [13], who validated a sensitive
method to detect endocrine-disrupting effects of xenobiotics
on the thyroid system. Fluoxetine was purchased as fluoxetine
hydrochloride (98.7% purity, Spectrum Chemicals and Labo-
ratory Products), and sertraline was purchased as sertraline
hydrochloride (98% purity, Toronto Research Chemicals).
Tadpoles were exposed to fluoxetine and sertraline at
concentrations of 0.1, 1, and 10 mg/L in FETAX solution
(pH 8.0 6 0.2; alkalinity 57 6 7 mg/L; hardness 104 6 7 mg/
L; data are means 6 standard deviation). A negative control
(FETAX solution) and a positive control (10 mg/L ammonium
perchlorate, which is a thyroid hormone inhibitor) were also
used in the experiment. Three replicate glass aquaria were used
for each treatment (n 5 3), except for the positive control (n 5
1). Glass aquaria contained 10 L of aerated test solution, and
28 tadpoles were allocated randomly to each aquarium.
Tadpoles were exposed to SSRIs for 70 days (12:12 h light/
dark cycle). Water in aquaria was renewed by 75% three times
per week. One time each week, prior to a water change, water
was analyzed for temperature (21.4 6 0.6uC), dissolved oxygen
(7.65 6 0.24), conductivity (1,421 6 44 ms), and total ammonia
nitrogen (0.7 6 0.2 mg/L) (data in parentheses are means 6
standard deviation).
Tadpoles were fed daily during exposures. Dry Nasco
Xenopus Frog BrittleTM for tadpoles was mixed with FETAX
solution in a blender (10 g of food to 100 ml of FETAX
solution) and dispensed into aquaria at a rate of 0.5 ml for
exposure days 1 to 7, 1.0 ml for exposure days 8 to 16, 2.0 ml
for exposure days 17 to 24, and 1.0 ml for exposure days 25 to
70. Feeding rates were adjusted during the experiment in an
attempt to have little to no food left remaining on the bottom
of aquaria the morning after feeding. However, this adjust-
ment was only successful for control tadpoles. Tadpoles
exposed to SSRIs ate less food than control tadpoles; hence
more uneaten food was observed on the bottom of these
aquaria.
Concentrations of SSRIs in water and tadpole tissue were
determined periodically throughout the experiment. On expo-
sure days 21 and 39, water samples (50 ml) were collected from
each replicate aquarium and composited (150 ml total volume)
both before and after water changes. Tadpole tissues were
sampled on exposure day 44 (composite of six tadpoles per
treatment, two tadpoles from each replicate aquarium). All
samples were stored frozen (220uC) until analyzed by liquid
chromatography–mass spectrometry (LC/MS). Water samples
from 1 and 10 mg/L treatments were analyzed by direct
injection onto the LC/MS apparatus, whereas water samples
from 0.1 mg/L treatments were analyzed by LC/MS after solid-
phase extraction (SPE, Bakerbond OctadecylH C18, 1,000 mg/
6 ml, Malinckrodt Baker). The recovery range for SPE water
samples was 83.5 to 87.2%. Tadpole tissues were extracted in
acetonitrile/methanol (1:1, v/v). Tissues (0.3 g) were homoge-
nized in 11 ml of acetonitrile/methanol and centrifuged. Then,
supernatants were decanted, evaporated to 1 ml, and com-
bined with 1 ml of the mobile phase solution (0.1% formic acid
in water/0.1% formic acid in acetonitrile, 1:1, v/v). Samples
were sonicated and filtered (0.45-mm Whatman polytetrafluo-
roethylene syringe filters) prior to injection onto the LC/MS
apparatus. The recovery range for tissue samples was 65 to
71%. Composited tissue samples were run in triplicate.
Analyses of adverse effects
Tadpoles were observed daily for mortalities and weekly for
deformities and stage of development. On exposure day 30,
eight tadpoles were sampled destructively from each aquarium
to determine hind limb growth and body mass of prometa-
morphs. Tadpoles were captured with a mesh net and placed in
a lethal solution of 0.2% FinquelH MS-222 (tricaine methane-
sulfonate, Argent Laboratories) for approximately 2 min.
Hind limb length was quantified with a dissecting microscope
by use of an ocular micrometer. Then, four tadpoles were
processed individually to determine body mass, and four
tadpoles were archived (220uC) for later analyses outside the
scope of the present study. On exposure day 44, six tadpoles
were sampled destructively from each aquarium (described
above) to quantify forelimb emergence and tissue concentra-
tions of SSRIs (described previously). When tadpoles com-
pleted metamorphosis, the time to tail resorption was
recorded, and they were processed individually to determine
body mass. Body mass was quantified by drying tadpoles at
100uC for 60 h in a drying oven, then samples were dessicated
for 24 h prior to determining dry weights on an analytical
balance.
Statistics
Data were analyzed at a significance level of p , 0.05.
Fisher’s exact test was used to evaluate the effects of SSRI
exposures on deformities in tadpoles. The effects of SSRI
exposures on growth (body mass) and development (hind limb
length, forelimb emergence, and time to tail resorption) were
evaluated with analysis of variance (ANOVA) by use of
Systat’ version 11 software (Systat Software). All data were
checked for normality (Shapiro–Wilk’s test) and homogeneity
of variance (Bartlett’s test) prior to use in ANOVAs, and data
were transformed if necessary to meet the assumptions of the
statistical tests. When an ANOVA indicated that there were
significant differences among treatments, Dunnett’s multiple
comparison procedure was used to determine which SSRI
treatments differed significantly from the control treatment.
Effects of pharmaceuticals on tadpole growth and development Environ. Toxicol. Chem. 28, 2009 2673

RESULTS
Tadpole exposures
Concentrations of SSRIs were measured in water and
tadpole tissues (Table 1). Concentrations of fluoxetine and
sertraline in water varied during the course of the static-
renewal experiment. Four hours after a water change,
fluoxetine concentrations were 0 to 19% lower than nominal
concentrations. By 48 h after a water change, fluoxetine
concentrations were 70 to 80% lower than nominal concen-
trations. Four hours after a water change, sertraline concen-
trations were 29 to 40% lower than nominal concentrations.
By 48 h after a water change, sertraline concentrations were 15
to 87% lower than nominal concentrations.
On exposure day 44, tadpoles accumulated higher concen-
trations of sertraline than fluoxetine. Pharmaceutical concen-
trations in tadpoles exposed to 10 mg/L sertraline were sixfold
higher than concentrations in tadpoles exposed to 10 mg/L
fluoxetine.
Analyses of adverse effects
Mortality. No significant effects on mortality were observed
in tadpoles exposed to fluoxetine and sertraline for 70 d. The
experiment mortality rate was 18 6 2%. The majority of
mortalities occurred during the first 7 d of exposures (8 6 1%).
During the remaining time of the experiment (i.e., exposure
days 8 to 70), the weekly mortality rate ranged from 0 to 3%.
Deformities. No significant increases in deformities were
observed in tadpoles exposed to fluoxetine and sertraline. A
low number of deformities were observed in all exposure
treatments, including the controls (Table 2). Deformities
observed included bent tails, edemas, missing or partial eyes,
and missing limbs. The overall prevalence for the total number
of deformities was 3.6%. Slight but insignificant increases in
bent tails were observed in tadpoles exposed to 10 mg/L
sertraline (p 5 0.12).
Growth. No significant effects on growth were observed in
prometamorphic tadpoles exposed to fluoxetine and sertraline
for 30 d (data not shown). Body mass among prometamorphic
tadpoles from different treatments was remarkably similar and
averaged 0.024 6 0.001 g dry weight (n 5 21 aquaria).
Significant effects on growth were observed in fluoxetine (F
5 4.702; df 5 3; p 5 0.006) and sertraline (F 5 5.299; df 5 3; p
5 0.003) exposed tadpoles that had completed metamorphosis
(Fig. 2). Reduced growth was observed in tadpoles exposed to
10 mg/L fluoxetine and 0.1, 1, and 10 mg/L sertraline. Reduced
growth was also observed in tadpoles exposed to 1 mg/L
fluoxetine, but data were not statistically significant. Growth
was reduced by approximately 35% in fluoxetine and sertraline
exposed tadpoles.
Development. No significant effects on development were
observed in tadpoles exposed to fluoxetine and sertraline
during early stages of metamorphosis. On exposure day 30,
hind limb length averaged 5.0 6 0.22 mm in tadpoles from all
treatments. On exposure day 44, the percentage of tadpoles
completing forelimb emergence averaged 54.0 6 3.4% for all
treatments.
Significant effects on development were observed in
tadpoles exposed to sertraline (F 5 5.025; df 5 3; p 5 0.030)
during later stages of metamorphosis (Fig. 3). Development
was accelerated in tadpoles exposed to 0.1 and 1 mg/L
sertraline. These tadpoles completed tail resorption 10 d
earlier than control tadpoles. No effects on tail resorption
were observed in tadpoles exposed to 10 mg/L sertraline.
Table 1. Concentrations of fluoxetine and sertraline measured inwater and tadpole tissue. Water samples were collected at 4 and 48 hafter a water change, and data are medians of samples collected onexposure days 22 and 40. Tissue samples are from tadpoles that were
collected on day 44 of exposures
Treatment
Nominalconcentration
in water (mg/L)
Measured concentrations
Water(mg/L) (4 h)
Water(mg/L) (48 h)
Tissue(ng/g wet wt)
Fluoxetine 0.1 0.15 0.02 ,LODa
1.0 0.89 0.24 ,LODa
10.0 8.07 3.00 0.36 6 0.04Sertraline 0.1 0.06 0.09 ,LODa
1.0 0.67 0.16 0.18 6 0.0010.0 7.08 1.26 2.1 6 0.12
a Less than the limit of detection (LOD) of 0.05 ng/g wet weight.
Table 2. Incidence of cumulative deformities (%) in tadpoles exposed to fluoxetine and sertraline for 70 d. Data are percentages calculated fromprobabilities for the number of deformities observed among the 84 tadpoles that were exposed in each treatment
Treatment Concentration (mg/L) Bent tails Edema Missing or partial eyes Missing or deformed limbs Total deformities
Control 0 0.0 1.2 1.2 0.0 2.4Fluoxetine 0.1 1.2 1.2 0.0 1.2 3.6
1.0 2.4 3.5 0.0 0.0 6.010.0 2.4 0.0 0.0 0.0 2.4
Sertraline 0.1 0.0 1.2 0.0 0.0 1.21.0 1.2 0.0 0.0 0.0 1.2
10.0 3.6 1.2 0.0 1.2 6.0
Fig. 2. Effects of fluoxetine and sertraline on growth (body mass) intadpoles exposed to selective serotonin reuptake inhibitors throughoutmetamorphosis. Data are means 6 standard error, n 5 3 aquaria pertreatment. Asterisks indicate significant differences from controls (p, 0.05).
2674 Environ. Toxicol. Chem. 28, 2009 D.E. Conners et al.
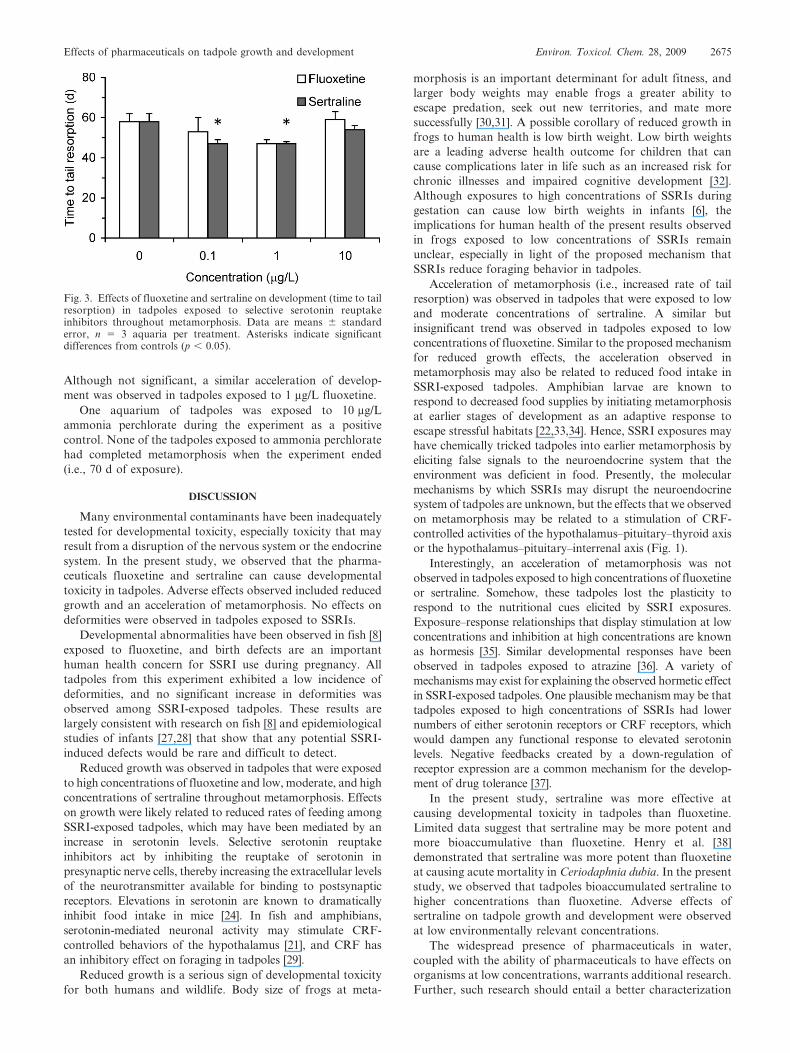
Although not significant, a similar acceleration of develop-
ment was observed in tadpoles exposed to 1 mg/L fluoxetine.
One aquarium of tadpoles was exposed to 10 mg/L
ammonia perchlorate during the experiment as a positive
control. None of the tadpoles exposed to ammonia perchlorate
had completed metamorphosis when the experiment ended
(i.e., 70 d of exposure).
DISCUSSION
Many environmental contaminants have been inadequately
tested for developmental toxicity, especially toxicity that may
result from a disruption of the nervous system or the endocrine
system. In the present study, we observed that the pharma-
ceuticals fluoxetine and sertraline can cause developmental
toxicity in tadpoles. Adverse effects observed included reduced
growth and an acceleration of metamorphosis. No effects on
deformities were observed in tadpoles exposed to SSRIs.
Developmental abnormalities have been observed in fish [8]
exposed to fluoxetine, and birth defects are an important
human health concern for SSRI use during pregnancy. All
tadpoles from this experiment exhibited a low incidence of
deformities, and no significant increase in deformities was
observed among SSRI-exposed tadpoles. These results are
largely consistent with research on fish [8] and epidemiological
studies of infants [27,28] that show that any potential SSRI-
induced defects would be rare and difficult to detect.
Reduced growth was observed in tadpoles that were exposed
to high concentrations of fluoxetine and low, moderate, and high
concentrations of sertraline throughout metamorphosis. Effects
on growth were likely related to reduced rates of feeding among
SSRI-exposed tadpoles, which may have been mediated by an
increase in serotonin levels. Selective serotonin reuptake
inhibitors act by inhibiting the reuptake of serotonin in
presynaptic nerve cells, thereby increasing the extracellular levels
of the neurotransmitter available for binding to postsynaptic
receptors. Elevations in serotonin are known to dramatically
inhibit food intake in mice [24]. In fish and amphibians,
serotonin-mediated neuronal activity may stimulate CRF-
controlled behaviors of the hypothalamus [21], and CRF has
an inhibitory effect on foraging in tadpoles [29].
Reduced growth is a serious sign of developmental toxicity
for both humans and wildlife. Body size of frogs at meta-
morphosis is an important determinant for adult fitness, and
larger body weights may enable frogs a greater ability to
escape predation, seek out new territories, and mate more
successfully [30,31]. A possible corollary of reduced growth in
frogs to human health is low birth weight. Low birth weights
are a leading adverse health outcome for children that can
cause complications later in life such as an increased risk for
chronic illnesses and impaired cognitive development [32].
Although exposures to high concentrations of SSRIs during
gestation can cause low birth weights in infants [6], the
implications for human health of the present results observed
in frogs exposed to low concentrations of SSRIs remain
unclear, especially in light of the proposed mechanism that
SSRIs reduce foraging behavior in tadpoles.
Acceleration of metamorphosis (i.e., increased rate of tail
resorption) was observed in tadpoles that were exposed to low
and moderate concentrations of sertraline. A similar but
insignificant trend was observed in tadpoles exposed to low
concentrations of fluoxetine. Similar to the proposed mechanism
for reduced growth effects, the acceleration observed in
metamorphosis may also be related to reduced food intake in
SSRI-exposed tadpoles. Amphibian larvae are known to
respond to decreased food supplies by initiating metamorphosis
at earlier stages of development as an adaptive response to
escape stressful habitats [22,33,34]. Hence, SSRI exposures may
have chemically tricked tadpoles into earlier metamorphosis by
eliciting false signals to the neuroendocrine system that the
environment was deficient in food. Presently, the molecular
mechanisms by which SSRIs may disrupt the neuroendocrine
system of tadpoles are unknown, but the effects that we observed
on metamorphosis may be related to a stimulation of CRF-
controlled activities of the hypothalamus–pituitary–thyroid axis
or the hypothalamus–pituitary–interrenal axis (Fig. 1).
Interestingly, an acceleration of metamorphosis was not
observed in tadpoles exposed to high concentrations of fluoxetine
or sertraline. Somehow, these tadpoles lost the plasticity to
respond to the nutritional cues elicited by SSRI exposures.
Exposure–response relationships that display stimulation at low
concentrations and inhibition at high concentrations are known
as hormesis [35]. Similar developmental responses have been
observed in tadpoles exposed to atrazine [36]. A variety of
mechanisms may exist for explaining the observed hormetic effect
in SSRI-exposed tadpoles. One plausible mechanism may be that
tadpoles exposed to high concentrations of SSRIs had lower
numbers of either serotonin receptors or CRF receptors, which
would dampen any functional response to elevated serotonin
levels. Negative feedbacks created by a down-regulation of
receptor expression are a common mechanism for the develop-
ment of drug tolerance [37].
In the present study, sertraline was more effective at
causing developmental toxicity in tadpoles than fluoxetine.
Limited data suggest that sertraline may be more potent and
more bioaccumulative than fluoxetine. Henry et al. [38]
demonstrated that sertraline was more potent than fluoxetine
at causing acute mortality in Ceriodaphnia dubia. In the present
study, we observed that tadpoles bioaccumulated sertraline to
higher concentrations than fluoxetine. Adverse effects of
sertraline on tadpole growth and development were observed
at low environmentally relevant concentrations.
The widespread presence of pharmaceuticals in water,
coupled with the ability of pharmaceuticals to have effects on
organisms at low concentrations, warrants additional research.
Further, such research should entail a better characterization
Fig. 3. Effects of fluoxetine and sertraline on development (time to tailresorption) in tadpoles exposed to selective serotonin reuptakeinhibitors throughout metamorphosis. Data are means 6 standarderror, n 5 3 aquaria per treatment. Asterisks indicate significantdifferences from controls (p , 0.05).
Effects of pharmaceuticals on tadpole growth and development Environ. Toxicol. Chem. 28, 2009 2675

of risks to both people and wildlife and an evaluation of risk
management options that would be most effective at reducing
environmental exposures.
Acknowledgement—We thank Samantha Burton for assistance withwater quality analyses and adult frog care. This research wassupported by a U.S. Environmental Protection Agency (U.S. EPA)Science to Achieve Results grant to M.C. Black and K.L. Armbrust(Grant R829006). This research has not been subject to U.S. EPAreview and therefore does not necessarily reflect the view of theAgency.
REFERENCES
1. Daughton CG, Ternes TA. 1999. Pharmaceuticals and personalcare products in the environment: Agents of subtle change?Environ Health Perspect 107(Suppl. 6):907–938.
2. Kolpin DW, Furlong ET, Meyer MT, Thurman EM, Zaugg SD,Barber LB, Buxton HT. 2002. Pharmaceuticals, hormones, andother organic wastewater contaminants in U.S. streams, 1999–2000:A national reconnaissance. Environ Sci Technol 36:1202–1211.
3. Metcalfe CD, Miao XS, Koenig BG, Struger J. 2003. Distributionof acidic and neutral drugs in surface waters near sewage treatmentplants in the lower Great Lakes, Canada. Environ Toxicol Chem22:2881–2889.
4. Johnson DJ, Sanderson H, Brain RA, Wilson CJ, Bestari KT,Solomon KR. 2005. Exposure assessment and microcosm fate ofselected selective serotonin reuptake inhibitors. Regul ToxicolPharmacol 42:313–323.
5. Brooks BW, Chambliss CK, Stanley JK, Ramirez A, Banks KE,Johnson RD, Lewis RJ. 2005. Determination of select antidepres-sants in fish from an effluent-dominated stream. Environ ToxicolChem 24:464–469.
6. Hines RN, Adams J, Buck GM, Faber W, Holson JF, JacobsonSE, Keszler M, McMartin K, Segraves RT, Singer LT, Sipes IG,Williams PL. 2004. NTP-CERHR expert panel report on thereproductive and developmental toxicity of fluoxetine. BirthDefects Res B Dev Reprod Toxicol 71:193–280.
7. Weissman AM, Levy BT, Hartz AJ, Bentler S, Donohue M,Ellingrod VL, Wisner KL. 2004. Pooled analysis of antidepressantlevels in lactating mothers, breast milk, and nursing infants. Am JPsychiatry 161:1066–1078.
8. Foran CM, Weston J, Slattery M, Brooks BW, Huggett DB. 2004.Reproductive assessment of Japanese medaka (Oryzias latipes)following a four-week fluoxetine (SSRI) exposure. Arch EnvironContam Toxicol 46:511–517.
9. Vorhees C, Acuff-Smith K, Schilling M, Fisher J, Moran M,Buelle-Sam J. 1994. A developmental neurotoxicity evaluation ofthe effects of prenatal exposure to fluoxetine in rats. Fundam ApplToxicol 23:194–205.
10. Deiro TCBJ, Manhaes-De-Castro R, Cabral JE, Barreto-MedeirosJM, Souza SL, Maririo SMOC, Castro FMM, Toscano AE, Jesus-Deiro RA, Banos KMFT. 2006. Sertraline delays the somatic growthand reflex ontogeny in neonate rats. Physiol Behav 87:338–344.
11. Oberlander TF, Warburton W, Misri S, Aghajanian J, HertzmanC. 2006. Neonatal outcomes after prenatal exposure to selectiveserotonin reuptake inhibitor antidepressants and maternal depres-sion using population-based linked health data. Arch GenPsychiatry 63:898–906.
12. American Society for Testing and Materials. 2004. Standard guidefor conducting the frog embryo teratogenesis assay–Xenopus(FETAX). E 1439-98. InAnnual Book of ASTM Standards, Vol11.05. West Conshohocken, PA, pp 1–16.
13. Opitz R, Braunbeck T, Bogi C, Pickford DB, Nentwig G,Oehlmann J, Tooi O, Lutz I, Kloas W. 2005. Description andinitial evaluation of a Xenopus metamorphosis assay for detectionof thyroid system—Disrupting activities of environmental com-pounds. Environ Toxicol Chem 24:653–664.
14. Burkhart JG, Ankley G, Bell H, Carpenter H, Fort D, GardinerD, Gardner H, Hale R, Helgen JC, Jepson P, Johnson D, LannooM, Lee D, Lary J, Levey R, Magner J, Meteyer C, Shelby MD,Lucier G. 2000. Strategies for assessing the implications ofmalformed frogs for environmental health. Environ HealthPerspect 108:83–90.
15. Hayes TB, Collins A, Lee M, Mendoza M, Noriega N, Stuart AA,Vonk A. 2002. Hermaphroditic, demasculinized frogs after
exposure to the herbicide atrazine at low ecologically relevantdoses. Proc Natl Acad Sci USA 99:5476–5480.
16. Shi YB. 2000. Amphibian Metamorphosis: From Morphology toMolecular Biology. Wiley-Liss, New York, NY, USA.
17. Brown V. 2003. Disrupting a delicate balance: Environmentaleffects on the thyroid. Environ Health Perspect 111:A642–A649.
18. DeVito M, Biegel L, Brouwer A, Brown S, Brucker-Davis F,Oliver Cheek A, Christensen R, Colborn T, Cooke P, Crissman J,Crofton K, Doerge D, Gray E, Hauser P, Hurley P, Kohn M,Lazar J, McMaster S, McClain M, McConnell E, Meler C, MillerR, Tietge J, Tyl R. 1999. Screening methods for thyroid hormonedisruptors. Environ Health Perspect 107:407–415.
19. Kirkegaard C, Faber J. 1998. The role of thyroid hormones indepression. Eur J Endocrinol 138:1–9.
20. Eravci M, Pinna G, Meinhold H, Baumgartner A. 2000. Effects ofpharmacological and nonpharmacological treatments on thyroidhormone metabolism and concentrations in rat brain. Endocrinol-ogy 141:1027–1040.
21. Lowry CA, Moore FL. 2006. Regulation of behavioral responsesby corticotropin-releasing factor. Gen Comp Endocrinol 146:19–27.
22. Denver RJ. 1997. Proximate mechanisms of phenotypic plasticityin amphibian metamorphosis. Am Zool 37:172–184.
23. Fukumoto T, Blakely R, Levin M. 2005. Serotonin transporterfunction is an early step in left-right patterning in chick and frogembryos. Dev Neurosci 27:349–363.
24. Meguid MM, Fetissov SO, Varma M, Sato T, Zhang L, LavianoA, Rossi-Fanelli F. 2000. Hypothalamic dopamine and serotoninin the regulation of food intake. Nutrition 16:843–857.
25. Olsson C, Homgren S. 2001. The control of gut motility. CompBiochem Physiol A Comp Physiol 128:481–503.
26. Nieuwkoop PD, Faber J. 1994. Normal Table of Xenopus laevis(Daudin): A Systematical and Chronological Survey of Develop-ment from the Fertilized Egg till the End of Metamorphosis, 3rd ed.Garland, New York, NY, USA.
27. Alwan S, Reefhuis J, Rasmussen SA, Olney RS, Friedman JM.2007. Use of selective serotonin-reuptake inhibitors in pregnancyand the risk of birth defects. N Engl J Med 356:2684–2692.
28. Louik C, Lin AE, Werler MM, Hernandez-Dıaz S, Mitchell AA.2007. First-trimester use of selective serotonin-reuptake inhibitorsand the risk of birth defects. N Engl J Med 356:2675–2683.
29. Crespi EJ, Denver RJ. 2004. Ontogeny of corticotropin-releasingfactor effects on locomotion and foraging in the Westernspadefoot toad (Spea hammondii). Horm Behav 46:399–410.
30. Chelgren ND, Rosenberg DK, Heppell SS, Gitelman AI. 2006.Carryover aquatic effects on survival of metamorphic frogs duringpond emigration. Ecol Appl 16:250–261.
31. Relyea RA. 2007. Getting out alive: How predators affect thedecision to metamorphose. Oecologia 152:389–400.
32. Hack M, Flannery DJ, Schluchter M, Cartar L, Borawski E, KleinN. 2002. Outcomes in young adulthood for very-low-birth-weightinfants. N Engl J Med 346:149–157.
33. Semlitsch RD. 1988. Time and size at metamorphosis related toadult fitness in Ambystoma talpoideum. Ecology 69:184–192.
34. Nicieza AG. 2000. Interacting effects of predation risk and foodavailability on larval anuran behavior and development. Oecologia123:497–505.
35. Calabrese EJ. 2008. Hormesis: Why it is important to toxicologyand toxicologists. Environ Toxicol Chem 27:1451–1474.
36. Brodeur JC, Svartz G, Perez-Coll CS, Marino DJG, Herkovits J.2009. Comparative susceptibility to atrazine of three developmen-tal stages of Rhinella arenarum and influence on metamorphosis:Non-monotonous acceleration of the time to climax and delayedtail resorption. Aquat Toxicol 91:161–170.
37. Littleton J. 2001. Receptor regulation as a unitary mechanism fordrug tolerance and physical dependence—Not quite as simple as itseemed! Addiction 96:87–101.
38. Henry TB, Kwon JW, Armbrust KL, Black MC. 2004. Acute andchronic toxicity of five selective serotonin reuptake inhibitors inCeriodaphnia dubia. Environ Toxicol Chem 23:2229–2233.
39. Edwards CJ, Yamamoto K, Kikuyama S, Kelley DB. 1999.Prolactin opens the sensitive period for androgen regulation of alarynx-specific myosin heavy chain gene. J Neurobiol 41:443–451.
40. Huang H, Brown DD. 2000. Prolactin is not juvenile hormone inXenopus laevis metamorphosis. Proc Natl Acad Sci USA 97:195–199.
2676 Environ. Toxicol. Chem. 28, 2009 D.E. Conners et al.
Copyright © 2022 FDOKUMEN




















