
Altered CB 1 receptor-signaling in prefrontal cortex from an animal model of depression is reversed...
11
Departamento de Fisiologı ´a y Farmacologı ´a, Universidad de Cantabria. Instituto de Biomedicina y Biotecnologı ´a de Cantabria (CSIC-UC-IDICAN), Santander, Centro de Investigacio ´n Biome ´dica en Red, Salud Mental (CIBER-SAM), Instituto de Salud Carlos III, Spain The endocannabinoid system (ECs) and the cannabinoid receptors (CB 1 and CB 2 ) have attracted much attention regarding their role in the neurobiology and treatment of mood disorders (Onaivi et al. 2006; Witkin et al. 2005; Vinod and Hungund 2006). CB 1 receptors are highly expressed in the cerebral cortex, striatum, substantia nigra, hippocampus, and other limbic structures (Herkenham et al. 1991; Moldrich and Wenger 2000; Howlett et al. 2002), where they mediate the central effects of exogenous and endogenous CBs, including effects on mood (Chaperon and Thiebot 1999; Howlett et al. 2002). Pharmacological and transgenic studies in animals demonstrate the participa- tion of CB 1 receptors in the control of emotional processes (Martin et al. 2002; Witkin et al. 2005). Moreover, aug- mented levels of ECs, increased CB 1 receptor density, and agonist-stimulated CB receptor activity in the prefrontal cortex (PFC) of depressed suicide victims have been reported (Hungund et al. 2004; Vinod et al. 2005; Pazos et al. 2006). Bilateral olfactory bulbectomy (OBX) in the rat is widely used as an animal model of depression with comorbid anxiety and exhibiting a number of behavioral, neurochem- ical, and structural changes that are reversed by chronic, but not acute, administration of clinically effective antidepressant drugs (Kelly et al. 1997; Song and Leonard 2005). More- over, in the context of the ‘neurogenesis hypothesis of depression’ (Banasr and Duman 2007), it is worth mention- ing that the impaired cell proliferation and/or neuronal degeneration observed following OBX are reduced by some antidepressants (Jaako-Movits et al. 2006; Keilhoff et al. 2006; Jarosik et al. 2007). Received April 29, 2008; revised manuscript received December 13, 2008; accepted December 16, 2008. Address correspondence and reprint requests to Alvaro Dı ´az, MD PhD, Departamento de Fisiologı ´a y Farmacologı ´a, Cardenal Herrera Oria s/n, University of Cantabria, Santander 39011, Spain. E-mail: [email protected] Abbreviations used: 5-HT, 5-hydroxytryptamine or serotonin; CB, cannabinoid; DRN, dorsal raphe nucleus; EC, endocannabinoid; OBX, olfactory bulbectomy; PFC, prefrontal cortex; SSRI, selective serotonin reuptake inhibitor. Abstract Bilateral olfactory bulbectomy in the rat (OBX) induces behavioral, neurochemical, and structural abnormalities simi- lar to those observed in human depression that are normal- ized after chronic, but not acute, treatment with antidepressants. In our study, OBX animals exhibited signifi- cant increases in both CB 1 receptor density ([ 3 H]CP55490 binding) and functionality (stimulation of [ 35 S]GTPcS binding by the cannabinoid (CB) agonist WIN 55212-2) at the pre- frontal cortex (PFC). After chronic treatment with fluoxetine (10 mg/kg/day, 14 days, s.c.), OBX-induced hyperactivity in the open-field test was fully abolished. Interestingly, chronic fluoxetine fully reversed the enhanced CB 1 -receptor signaling in PFC observed following OBX. The CB agonist D 9 -tetra- hydrocannabinol (5 mg/kg, i.p., 1 day) did not produce any behavioral effect in sham-operated animals but returned locomotor activity to control values in OBX rats. As both acute administration of D 9 -tetrahydrocannabinol and chronic fluo- xetine elicited a similar behavioral effect in the OBX rat, it is not unlikely that the regionally selective enhancement of CB 1 receptor-signaling in the PFC could be related with the altered OBX behavior. Our findings reinforce the utility of this animal model to further investigating the implication of the endoc- annabinoid system in the modulation of emotional processes and its potential role in the adaptive responses to chronic antidepressants. Keywords: [ 35 S]GTPcS binding, 9 D-tetrahydrocannabinol, cannabinoid receptor 1, fluoxetine, olfactory bulbectomy, prefrontal cortex. J. Neurochem. (2009) 108, 1423–1433. JOURNAL OF NEUROCHEMISTRY | 2009 | 108 | 1423–1433 doi: 10.1111/j.1471-4159.2009.05898.x Ó 2009 The Authors Journal Compilation Ó 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1423–1433 1423
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Altered CB 1 receptor-signaling in prefrontal cortex from an animal model of depression is reversed...

Departamento de Fisiologıa y Farmacologıa, Universidad de Cantabria. Instituto de Biomedicina y Biotecnologıa de Cantabria
(CSIC-UC-IDICAN), Santander, Centro de Investigacion Biomedica en Red, Salud Mental (CIBER-SAM), Instituto de Salud Carlos
III, Spain
The endocannabinoid system (ECs) and the cannabinoidreceptors (CB1 and CB2) have attracted much attentionregarding their role in the neurobiology and treatment ofmood disorders (Onaivi et al. 2006; Witkin et al. 2005;Vinod and Hungund 2006). CB1 receptors are highlyexpressed in the cerebral cortex, striatum, substantia nigra,hippocampus, and other limbic structures (Herkenham et al.1991; Moldrich and Wenger 2000; Howlett et al. 2002),where they mediate the central effects of exogenous andendogenous CBs, including effects on mood (Chaperonand Thiebot 1999; Howlett et al. 2002). Pharmacologicaland transgenic studies in animals demonstrate the participa-tion of CB1 receptors in the control of emotional processes(Martin et al. 2002; Witkin et al. 2005). Moreover, aug-mented levels of ECs, increased CB1 receptor density, andagonist-stimulated CB receptor activity in the prefrontalcortex (PFC) of depressed suicide victims have been reported(Hungund et al. 2004; Vinod et al. 2005; Pazos et al. 2006).
Bilateral olfactory bulbectomy (OBX) in the rat is widelyused as an animal model of depression with comorbid
anxiety and exhibiting a number of behavioral, neurochem-ical, and structural changes that are reversed by chronic, butnot acute, administration of clinically effective antidepressantdrugs (Kelly et al. 1997; Song and Leonard 2005). More-over, in the context of the ‘neurogenesis hypothesis ofdepression’ (Banasr and Duman 2007), it is worth mention-ing that the impaired cell proliferation and/or neuronaldegeneration observed following OBX are reduced by someantidepressants (Jaako-Movits et al. 2006; Keilhoff et al.2006; Jarosik et al. 2007).
Received April 29, 2008; revised manuscript received December 13,2008; accepted December 16, 2008.Address correspondence and reprint requests to Alvaro Dıaz, MD
PhD, Departamento de Fisiologıa y Farmacologıa, Cardenal Herrera Orias/n, University of Cantabria, Santander 39011, Spain.E-mail: [email protected] used: 5-HT, 5-hydroxytryptamine or serotonin; CB,
cannabinoid; DRN, dorsal raphe nucleus; EC, endocannabinoid; OBX,olfactory bulbectomy; PFC, prefrontal cortex; SSRI, selective serotoninreuptake inhibitor.
Abstract
Bilateral olfactory bulbectomy in the rat (OBX) induces
behavioral, neurochemical, and structural abnormalities simi-
lar to those observed in human depression that are normal-
ized after chronic, but not acute, treatment with
antidepressants. In our study, OBX animals exhibited signifi-
cant increases in both CB1 receptor density ([3H]CP55490
binding) and functionality (stimulation of [35S]GTPcS binding
by the cannabinoid (CB) agonist WIN 55212-2) at the pre-
frontal cortex (PFC). After chronic treatment with fluoxetine
(10 mg/kg/day, 14 days, s.c.), OBX-induced hyperactivity in
the open-field test was fully abolished. Interestingly, chronic
fluoxetine fully reversed the enhanced CB1-receptor signaling
in PFC observed following OBX. The CB agonist D9-tetra-
hydrocannabinol (5 mg/kg, i.p., 1 day) did not produce any
behavioral effect in sham-operated animals but returned
locomotor activity to control values in OBX rats. As both acute
administration of D9-tetrahydrocannabinol and chronic fluo-
xetine elicited a similar behavioral effect in the OBX rat, it is
not unlikely that the regionally selective enhancement of CB1
receptor-signaling in the PFC could be related with the altered
OBX behavior. Our findings reinforce the utility of this animal
model to further investigating the implication of the endoc-
annabinoid system in the modulation of emotional processes
and its potential role in the adaptive responses to chronic
antidepressants.
Keywords: [35S]GTPcS binding, 9D-tetrahydrocannabinol,
cannabinoid receptor 1, fluoxetine, olfactory bulbectomy,
prefrontal cortex.
J. Neurochem. (2009) 108, 1423–1433.
JOURNAL OF NEUROCHEMISTRY | 2009 | 108 | 1423–1433 doi: 10.1111/j.1471-4159.2009.05898.x
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1423–1433 1423

Adaptive changes in the central serotonergic systemappear to play an important role in the OBX-inducedsyndrome. The neurodegeneration of serotoninergic neuronsin dorsal raphe nucleus (DRN) following OBX results in thepermanent deficits in the secretion of 5-hydroxytryptamine orserotonin (5-HT) in the dorsal hippocampus and amygdala(van der Stelt et al. 2005). Abnormalities in 5-HT concen-tration, 5-HT transporter, and receptor expression (5-HT2 and5-HT1A receptors) following OBX in areas such as theamygdaloid cortex, frontal cortex, and midbrain have beenreported (Lumia et al. 1992; Gurevich et al. 1993; Greckschet al. 1997; Zhou et al. 1998; Slotkin et al. 2005). Altera-tions in other neurotransmitter systems (noradrenaline,dopamine, glutamate, and GABA) have been also shown(Jancsar and Leonard 1984; Song and Leonard 2005) but, todate and to our knowledge, the study of the ECs in thisanimal model of depression has not been addressed. Chronicmild/unpredictable stress exposure to rats (Bortolato et al.2007) and to mice (Hillard et al. 2006) results in an increaseCB1 receptor mRNA in PFC. In a very recent paper, Hillet al. (2008) demonstrated that chronic unpredictable stressin rats results in a significant increase in CB1 receptorbinding site density in PFC and a decrease in CB1 receptorbinding site density in hippocampus, hypothalamus, andventral striatum, together with a significant reduction in thecontent of the EC ligand N-arachidonylethanolamide(anandamide) in all the brain area examined.
There are some pre-clinical evidences about the modula-tion of 5-HT neurotransmission by the ECs: CB1 receptorsand the enzyme regulating anandamide brain levels (fattyacid amide hydrolase) are present in the DRN of the mousebrain (Egertova et al. 2003); CBs elicit antidepressant-likebehavior by activating DRN serotonergic neurons (Bambicoet al. 2007); CB1 agonists increase the firing activity of DRNserotonergic neurons (Gobbi et al. 2005), suppress 5-HTrelease in the mouse brain cortex (Nakazi et al. 2000) andblock 5-HT reuptake in the rat brain cortex (Banerjee et al.1975) whereas CB1 antagonists stimulate 5-HT release intothe rat medial PFC (Tzavara et al. 2003); chronic CBtreatment appears to up-regulate 5-HT2A receptor activitywhile concurrently down-regulating 5-HT1A receptor activityin the rat brain, a finding similar to that sometimes observedin depression (Hill et al. 2006b); and impaired functionalityof 5-HT1A and 5-HT2A/C receptors has been found in CB1
knockout mice that exhibit a depressive-like phenotype(Mato et al. 2007).
There are still few evidences about the modulation of theECs by pharmacological manipulation of brain 5-HT neuro-transmission. Acute fluoxetine modulates 9D-tetrahydrocan-nabinol-induced hypothermia in the rat (Malone and Taylor1998), an effect in which somatodendritic 5-HT1A autore-ceptors appear to be involved (Malone and Taylor 2001).Chronic fluoxetine decreases CB1 receptor gene expressionin the caudate–putamen (Oliva et al. 2005) of the rat brain
and chronic citalopram decreases CB receptor-mediated Gprotein coupling in rat hypothalamus and hippocampus, twoareas important in the control of neuroendocrine function andmood (Hesketh et al. 2007); however, chronic administrationof desipramine, a non-selective serotonin reuptake inhibitor(SSRI) drug, results in an increased density of CB1 receptorin the hippocampus and hypothalamus of the rat brain (Hillet al. 2006a).
This study was aimed to investigate CB1 receptor-medi-ated signaling in the brain of olfactory bulbectomized rats, itsimplication in the OBX-induced behavior and its modulationby the chronic administration of the SSRI fluoxetine. Firstly,we have analyzed the changes in the density and function-ality of CB1 receptors in the brain OBX rats. Secondly, wehave evaluated the antidepressant-like activity of chronicfluoxetine in the OBX rat as well as the effect of theantidepressant upon the density and functionality of CB1
receptors in PFC membranes. Finally, we have assessed theeffect of the administration of the CB agonist 9D-tetra-hydrocannabinol in the OBX-induced behavior before andafter chronic treatment with fluoxetine.
Materials and methods
AnimalsExperiments were conducted on male Sprague–Dawley rats (Harlan,
Barcelona, Spain) weighing at the moment of surgery 250–270 g.
Animals were housed in a climate controlled room with a 12-h light/
dark cycle and had ad libitum access to food and water. All surgical
procedures and experiments were carried out with the approval of
the Animal Care Committee of the Universidad de Cantabria and
were performed according to the Spanish legislation and the
European Communities Council Directive on ‘Protection of Animals
Used in Experimental and Other Scientific Purposes’ (86/609/EEC).
MaterialsFluoxetine HCl was kindly supplied by FAES FARMA S.A. Leioa,
Bilbao, Spain. Osmotic minipumps (Alzet osmotic pump model
2ML2; 0.5 lL/h) were supplied by Alzet Corporation (Palo Alto,
CA, USA). 9D-Tetrahydrocannabinol was purchased from THC
Pharm GMBH The Health Concept (Frankfurt, Germany); SR
141716A was kindly supplied by Sanofi Reserche (Montpellier,
France). [3H]CP55940 (160 Ci/mmol) and [35S]GTPcS (1250 Ci/
mmol) were purchased from PerkinElmer Life Sciences (Madrid,
Spain). All other materials, unless otherwise stated, were purchased
from Sigma-Aldrich (St Louis, MO, USA).
Bilateral olfactory bulbectomy and open-field testThe OBX procedure was performed as previously described
(Cairncross et al. 1977). Animals were anesthetized and the bulbs
were excised and removed under stereotactic surgery. Sham-
operated animals underwent the same procedure except for excision
and aspiration of the olfactory bulbs. Behavioral studies started after
a 2-week recovery period. At the end of the experiments, animals
were killed and the success of the operation was confirmed
anatomically. The open field was conducted as previously described
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1423–1433� 2009 The Authors
1424 | A. Rodrıguez-Gaztelumendi et al.

(Gray and Lalljee 1974), recording for a 5 min period the number of
ambulations, rearings, grooming episodes, and defecations (see
Supporting information for details).
Experimental groups and pharmacological treatmentsThree sets of experiments were carried out under this study (see
Fig. S1). In the first set, animals underwent surgery (OBX = 5;
sham = 5) and the behavioral testing in the open field arena was
carried on day 15 post-surgery. Then, animals were killed and
autoradiographic studies in brain sections were performed. In the
second set of experiments, another group of animals underwent
surgery (sham = 14, OBX = 16), an open field test was performed
on day 15 post-surgery and, the following day, they were divided
into four subgroups to receive fluoxetine (10 mg/kg/day, s.c.) or
vehicle for another 2 weeks: vehicle-treated sham-operated (n = 6),
fluoxetine-treated sham-operated (n = 8), vehicle-treated OBX
(n = 8), and fluoxetine-treated OBX (n = 8). Fluoxetine HCl or
vehicle (50% propylene glycol, 10% ethanol, and 40% distilled
water) were administered by osmotic minipumps (Castro et al.2003). An open field session was performed at the end of the
treatment (day 29 following OBX surgery) without removing the
minipumps. Animals were killed after a 24 h washout period and
their brains removed (day 31 post-surgery). The PFC was dissected
and processed for the binding assays in membrane homogenates. In
the third set of experiments, animals underwent surgery (OBX = 10;
sham = 6). The behavioral effects of a challenge dose of 9D-tetrahydrocannabinol (5 mg/kg, i.p.) and its vehicle (5% ethanol,
5% emulphor, and 90% saline) were evaluated, 30 min after
administration, in different open-field sessions performed at days 15
and 16. The following day, sham and OBX animals were implanted
minipumps to receive chronic fluoxetine and the behavioral effect of9D-tetrahydrocannabinol was again evaluated at the end of the
treatment (day 29 post-surgery). The dose of 9D-tetrahydrocannab-inol was selected according to the results obtained from preliminary
studies demonstrating no behavioral effects in naıve animals under
the environmental conditions in which the open field test is
performed in this study.
Autoradiography of [3H]CP55940 binding and [35S]GTPcS bindingCoronal brain sections (20 lm) were obtained from sham and OBX
animals (20 lm). [3H]CP55940 binding autoradiography was
performed according to Glass et al. (1997), with minor modifica-
tions (Mato and Pazos 2004), by incubating brain sections with
3 nM [3H]CP55940 and defining the non-specific binding with
10 lM WIN 55212-2. [35S]GTPcS autoradiography was performed
as previously described (Sim et al. 1996) assessing the stimulation
of [35S]GTPcS binding by incubating brain sections with 0.04 nM
[35S]GTPcS under the following conditions: in the absence (basal
binding) or the presence of the CB agonist WIN 55212-2 (10 lM)
(stimulated binding), in the presence of WIN 55212-2
(10 lM) + SR 141716A (10 lM) and in the presence of 10 lMGTPcS (non-specific binding) (see Supporting information for
details).
[3H]CP55940 Binding and [35S]GTPcS binding in prefrontal cortexmembranesOlfactory bulbectomy and sham animals treated with fluoxetine or
vehicle were killed following the open field test, their brains were
removed, and membrane homogenates were obtained from
prefrontal cortical (PFC) samples. [3H]CP55940 binding assay in
PFC cortical membranes was performed as previously described
(Hungund et al. 2004) by incubating aliquots of PFC membranes
with [3H]CP55940 (0.0125–3.2.0 nM) and defining the non-specific
binding with WIN 55212-2 (10 lM). The CB1 receptor-mediated
[35S]GTPcS binding assay was performed as described previously
(Gonzalez-Maeso et al. 2000) with minor modifications, incubating
aliquots of PFC membranes with 0.1 nM [35S]GTPcS under the
following conditions: in the absence (basal binding) or the presence
of the CB agonist WIN 55212-2 (10)9 to 10)3 M) (stimulated
binding), in the presence of WIN 55212-2 (10 lM) + SR 141716A
(10 lM) and in the presence of 10 lM GTPcS (non-specific
binding) (see Supporting information for details).
Data analysisAutoradiographic densities were determined by densitometry using
the Scion Image software (Scion Corporation, Frederick, MD,
USA). Relative optical density values of brain regions were
averaged over two consecutive sections per rat (bilateral readings)
and converted to nCi/mg of tissue ([3H]CP55940 receptor binding)
or nCi/g of tissue ([35S]GTPcS binding). CB1 receptor autoradio-
graphic densities are given in fmol/mg tissue equivalent (fmol/mg
tissue) and CB1 agonist-stimulation of [35S]GTPcS binding data are
presented as percentage of basal binding (100%). [3H]CP55940 and
[35S]GTPcS binding assays in PFC membranes were carried out
using triplicates and the results represent the mean of the n values
obtained from each experimental group. The Bmax (maximal bound,
fmol/mg protein) and Kd (receptor affinity, nM) for [3H]CP55940
binding studies, and Emax (maximal stimulatory effect expressed as
percentage of basal binding) and pEC50 (potency expressed as
)log EC50) for [35S]GTPcS binding assays in PFC membranes were
determined by non-linear regression analysis. Analysis of the
[3H]CP55940 and [35S]GTPcS binding data were performed using
the computer program PRISM, version 4.03 (GraphPad Software, San
Diego, CA, USA).
Statistical analysisAll data are expressed as mean ± SEM. A two-tailed unpaired
Student’s t-test was used for statistical comparison between two
experimental groups. Differences in drug treatment between sham
and OBX rats were compared using a two-way ANOVA (with surgery
and drug treatment as factors) followed by Bonferroni post hoc testwhen appropriate (PRISM, version 4.03). A probability of p £ 05 was
taken as the level of statistical significance.
Results
Autoradiographic CB1 receptor density and CB1
receptor-mediated stimulation of [35S]GTPcS bindingin the brain of the OBX ratIn the group of animals used for the autoradiographic studies,a characteristic hyperactivity in the open field test wasconfirmed 15 days post-surgery in the OBX group (n = 5)when compared with the sham-operated counterparts (n = 5)as indicated by the increase in the number of ambulations
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1423–1433
Prefrontal CB1 receptors and chronic fluoxetine | 1425

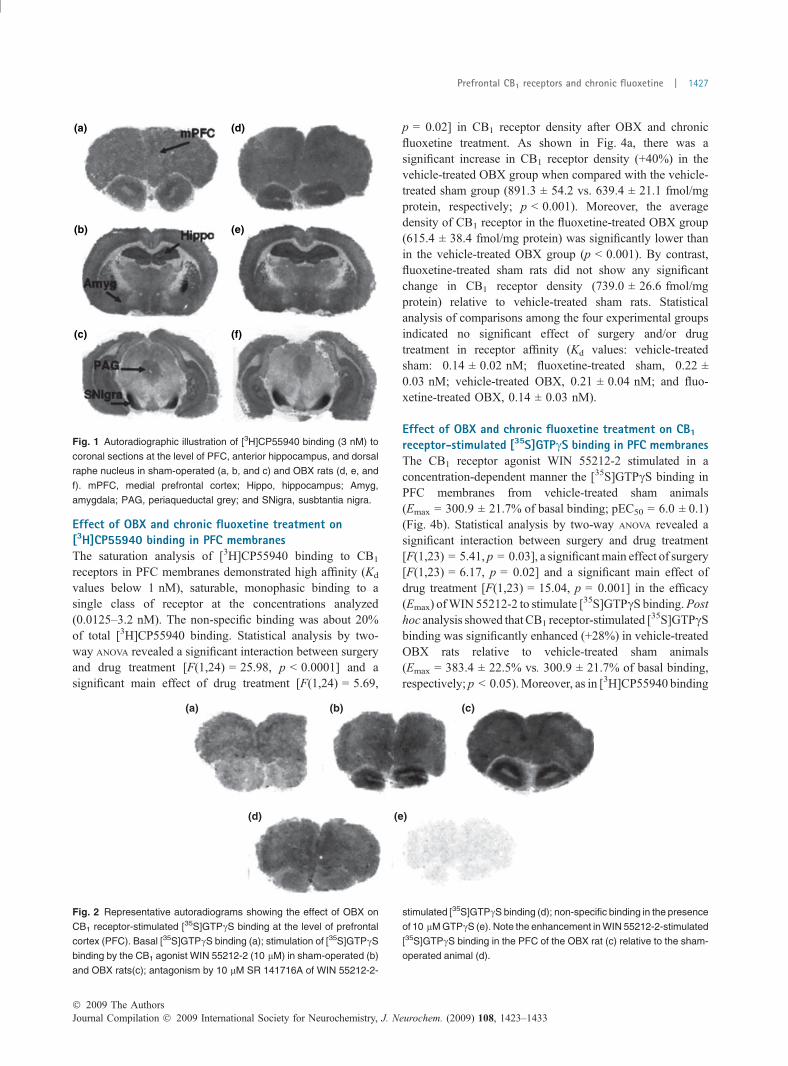
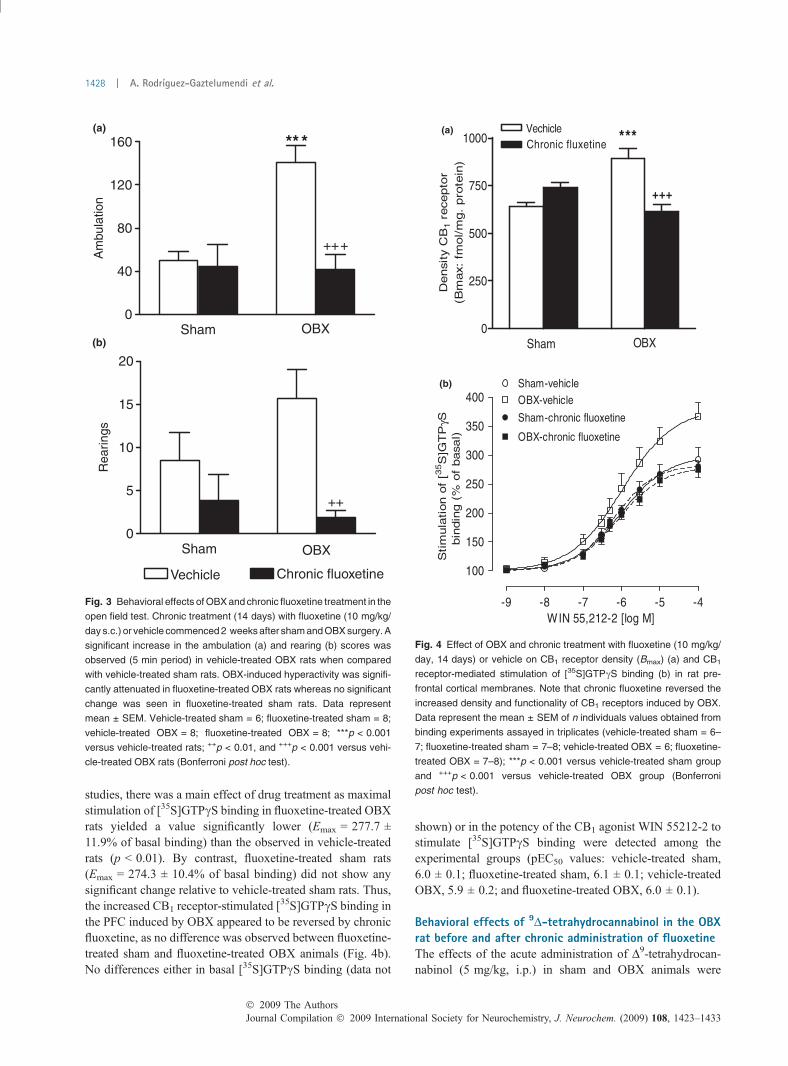
[152.1 ± 13.2 vs. 85.4 ± 9.7; t (8) = 4.07, p = 0.004] andrearings [26.0 ± 4.9 vs. 11.1 ± 2.3; t (8) = 4.66, p = 0.03].No significant differences were observed in the number ofgrooming episodes, the number of defecations or the bodyweights (data not shown). Densitometric analysis of[3H]CP55940 and [35S]GTPcS binding to brain sectionsrevealed that CB1 receptor density ([3H]CP55940 binding)was significantly increased in OBX rats in areas such as themPFC [+29%; t (7) = 2.68, p = 0.03] and the amygdala[+36%; t (8) = 3.00, p = 0.02] whereas no changes weredetected in the caudate–putamen, hippocampus (CA1–CA2fields), and DRN (Table 1 and Fig. 1). CB1 agonist-stimu-lated [35S]GTPcS binding by 10 lM WIN 55212-2 resultedto be significantly enhanced in the mPFC of OBX animals[+40%; t (7) = 2.88, p = 0.02] in comparison to the shamgroup. A slight increase in CB1 agonist-stimulated[35S]GTPcS binding was also observed in the amygdala[+8%; t (8) = 1.39, p = 0.2] although not reaching statisticalsignificance (Table 1 and Fig. 2). No differences weremeasured among the two experimental groups in the levelof basal [35S]GTPcS binding in all the regions examined(data not shown). As significant changes were observed inboth the density and the functionality of CB1 receptors in thePFC from OBX rats, this region was selected to furtherevaluate the effect of OBX and chronic fluoxetine on[3H]CP55940 binding and CB1 agonist-stimulated[35S]GTPcS binding using membrane homogenates.
Effect of chronic fluoxetine on the olfactorybulbectomy-induced behavior in the open fieldTwo weeks after sham (n = 14) or OBX (n = 16) surgery andbefore the initiation of the chronic treatment with fluoxetineor vehicle, the OBX-induced hyperactivity was confirmed onthe open-field test (data not shown). The day after, sham andOBX animals were treated with fluoxetine (10 mg/kg/day,s.c.) or vehicle for another 14 days and the behavioral effectsof the antidepressant were evaluated in the open field test. No
significant differences were detected in body weights amongthe experimental groups either at the start or the end of drugtreatment (data not shown).
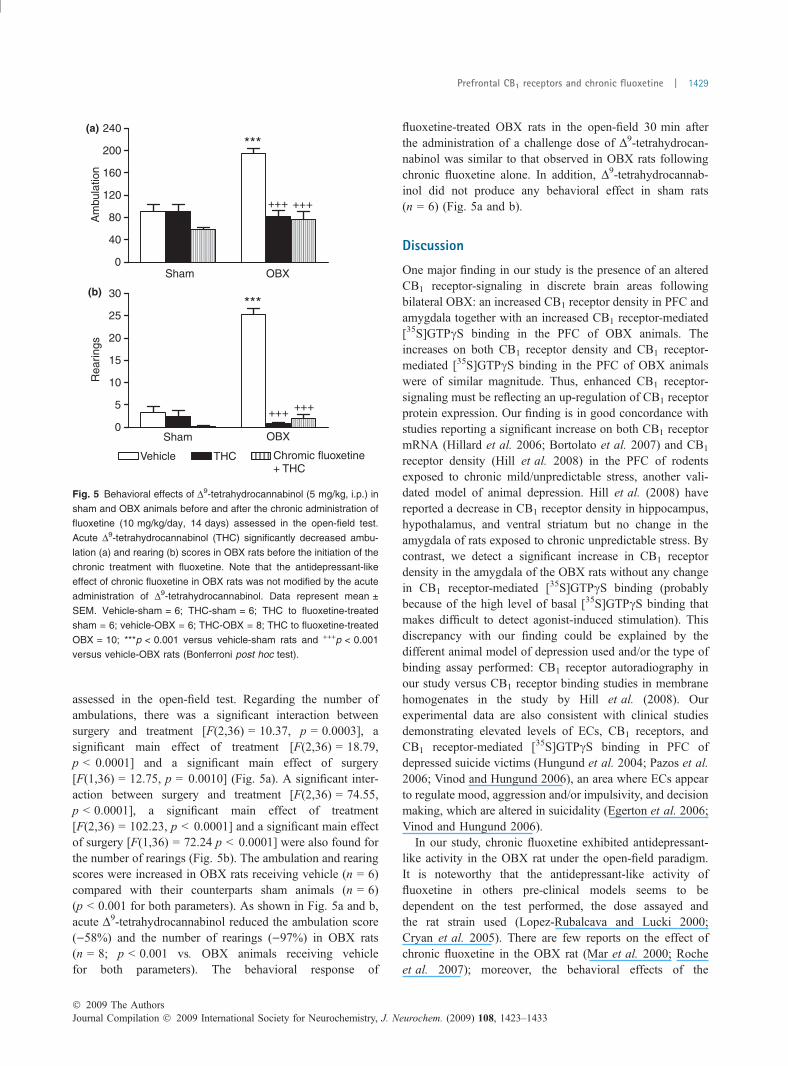
The total ambulation scores of vehicle-treated and fluoxe-tine-treated sham and OBX rats exposed to a 5 min openfield is shown in Fig. 3a. A two-way ANOVA revealed asignificant interaction between surgery and drug treatment[F(1,26) = 8.62, p = 0.007], a significant main effect ofsurgery [F(1,26) = 7.44, p = 0.01] and a significant maineffect of drug treatment [F(1,26) = 10.86, p = 0.003].Post hoc analysis showed that the total ambulation scorewas significantly increased in vehicle-treated OBX ratscompared with vehicle-treated sham-operated controls(p < 0.01). Interestingly, there was a significant and totalattenuation of the number of ambulations in the fluoxetine-treated OBX group relative to the vehicle-treated OBX group(p < 0.001). Thus, chronic fluoxetine appeared to rectify theOBX-induced locomotor hyperactivity, as no difference wasobserved between fluoxetine-treated sham and fluoxetine-treated OBX animals.
The number of rearings following chronic treatment withfluoxetine or vehicle in sham and OBX rats is shown inFig. 3b. There was a significant main effect of drug treatment[F(1,26) = 10.88, p = 0.003], in which fluoxetine-treatedOBX rats showed reduced rearing activity when comparedwith vehicle-treated OBX rats (p < 0.01). Fluoxetine-treatedsham animals did not show any significant change in rearingactivity when compared with vehicle-treated sham rats.
There was a significant main effect of drug treatment onthe number of defecations [F(1,26) = 9.82 p = 0.004].Post hoc analysis showed that defecation was significantlyincreased in fluoxetine-treated OBX rats relative to vehicle-treated OBX rats (p < 0.05). Fluoxetine-treated sham ratsalso exhibited an increased number of defecations compara-ble to fluoxetine-treated OBX rats, though it did not resultsignificant when compared with vehicle-treated sham ani-mals (p = 0.06).
Table 1 Effect of bilateral OBX upon the density and functionality of CB1 receptors in the rat brain
Brain region
[3H]CP55490 binding to CB1 receptors
(fmol/mg tissue)
CB1 receptor-stimulated [35S]GTPcS binding
by WIN 55212-2 (% of basal)
Sham OBX Sham OBX
Medial prefrontal cortex 28.95 ± 2.61 37.24 ± 1.83* 140.25 ± 10.15 196.07 ± 17.78*
Caudate–putamen 52.08 ± 2.91 50.08 ± 1.36 173.71 ± 12.63 173.99 ± 20.24
Hippocampus (CA1–CA2 regions) 66.02 ± 2.58 61.07 ± 2.21 171.04 ± 26.71 168.26 ± 24.93
Amygdala (central-medial nuclei) 20.96 ± 1.60 28.48 ± 1.93* 110.07 ± 2.98 118.85 ± 5.59
Dorsal raphe nucleus 22.86 ± 2.34 20.63 ± 1.49 105.55 ± 5.26 105.84 ± 6.02
OBX, olfactory bulbectomy; CB, cannabinoid. CB1 receptor’s density and functionality were evaluated in consecutive coronal sections of sham-
operated (Sham, n = 4–5) and olfactory bulbectomized rats (OBX, n = 4–5). The density of CB1 receptors was assessed with [3H]CP55490 (3 nM)
and CB1 receptor by measuring the stimulation of [35S]GTPcS binding in the presence of WIN 55212-2 (10 lM). Specific CB1 receptor-stimulated
binding is expressed as the percent of basal binding (100%). See Materials and methods for further details. Results are expressed as the
mean ± SEM; *p < 0.05 versus sham-operated group (two-tailed unpaired Student’s t-test).
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1423–1433� 2009 The Authors
1426 | A. Rodrıguez-Gaztelumendi et al.
Effect of OBX and chronic fluoxetine treatment on[3H]CP55940 binding in PFC membranesThe saturation analysis of [3H]CP55940 binding to CB1
receptors in PFC membranes demonstrated high affinity (Kd
values below 1 nM), saturable, monophasic binding to asingle class of receptor at the concentrations analyzed(0.0125–3.2 nM). The non-specific binding was about 20%of total [3H]CP55940 binding. Statistical analysis by two-way ANOVA revealed a significant interaction between surgeryand drug treatment [F(1,24) = 25.98, p < 0.0001] and asignificant main effect of drug treatment [F(1,24) = 5.69,
p = 0.02] in CB1 receptor density after OBX and chronicfluoxetine treatment. As shown in Fig. 4a, there was asignificant increase in CB1 receptor density (+40%) in thevehicle-treated OBX group when compared with the vehicle-treated sham group (891.3 ± 54.2 vs. 639.4 ± 21.1 fmol/mgprotein, respectively; p < 0.001). Moreover, the averagedensity of CB1 receptor in the fluoxetine-treated OBX group(615.4 ± 38.4 fmol/mg protein) was significantly lower thanin the vehicle-treated OBX group (p < 0.001). By contrast,fluoxetine-treated sham rats did not show any significantchange in CB1 receptor density (739.0 ± 26.6 fmol/mgprotein) relative to vehicle-treated sham rats. Statisticalanalysis of comparisons among the four experimental groupsindicated no significant effect of surgery and/or drugtreatment in receptor affinity (Kd values: vehicle-treatedsham: 0.14 ± 0.02 nM; fluoxetine-treated sham, 0.22 ±0.03 nM; vehicle-treated OBX, 0.21 ± 0.04 nM; and fluo-xetine-treated OBX, 0.14 ± 0.03 nM).
Effect of OBX and chronic fluoxetine treatment on CB1
receptor-stimulated [35S]GTPcS binding in PFC membranesThe CB1 receptor agonist WIN 55212-2 stimulated in aconcentration-dependent manner the [35S]GTPcS binding inPFC membranes from vehicle-treated sham animals(Emax = 300.9 ± 21.7% of basal binding; pEC50 = 6.0 ± 0.1)(Fig. 4b). Statistical analysis by two-way ANOVA revealed asignificant interaction between surgery and drug treatment[F(1,23) = 5.41, p = 0.03], a significantmain effect of surgery[F(1,23) = 6.17, p = 0.02] and a significant main effect ofdrug treatment [F(1,23) = 15.04, p = 0.001] in the efficacy(Emax) ofWIN 55212-2 to stimulate [35S]GTPcS binding.Posthoc analysis showed that CB1 receptor-stimulated [35S]GTPcSbinding was significantly enhanced (+28%) in vehicle-treatedOBX rats relative to vehicle-treated sham animals(Emax = 383.4 ± 22.5% vs. 300.9 ± 21.7% of basal binding,respectively; p < 0.05).Moreover, as in [3H]CP55940 binding
(a)
(d) (e)
(b) (c)
Fig. 2 Representative autoradiograms showing the effect of OBX on
CB1 receptor-stimulated [35S]GTPcS binding at the level of prefrontal
cortex (PFC). Basal [35S]GTPcS binding (a); stimulation of [35S]GTPcS
binding by the CB1 agonist WIN 55212-2 (10 lM) in sham-operated (b)
and OBX rats(c); antagonism by 10 lM SR 141716A of WIN 55212-2-
stimulated [35S]GTPcS binding (d); non-specific binding in the presence
of 10 lM GTPcS (e). Note the enhancement in WIN 55212-2-stimulated
[35S]GTPcS binding in the PFC of the OBX rat (c) relative to the sham-
operated animal (d).
(a) (d)
(b) (e)
(c) (f)
Fig. 1 Autoradiographic illustration of [3H]CP55940 binding (3 nM) to
coronal sections at the level of PFC, anterior hippocampus, and dorsal
raphe nucleus in sham-operated (a, b, and c) and OBX rats (d, e, and
f). mPFC, medial prefrontal cortex; Hippo, hippocampus; Amyg,
amygdala; PAG, periaqueductal grey; and SNigra, susbtantia nigra.
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1423–1433
Prefrontal CB1 receptors and chronic fluoxetine | 1427
studies, there was a main effect of drug treatment as maximalstimulation of [35S]GTPcS binding in fluoxetine-treated OBXrats yielded a value significantly lower (Emax = 277.7 ±11.9% of basal binding) than the observed in vehicle-treatedrats (p < 0.01). By contrast, fluoxetine-treated sham rats(Emax = 274.3 ± 10.4% of basal binding) did not show anysignificant change relative to vehicle-treated sham rats. Thus,the increased CB1 receptor-stimulated [35S]GTPcS binding inthe PFC induced by OBX appeared to be reversed by chronicfluoxetine, as no difference was observed between fluoxetine-treated sham and fluoxetine-treated OBX animals (Fig. 4b).No differences either in basal [35S]GTPcS binding (data not
shown) or in the potency of the CB1 agonist WIN 55212-2 tostimulate [35S]GTPcS binding were detected among theexperimental groups (pEC50 values: vehicle-treated sham,6.0 ± 0.1; fluoxetine-treated sham, 6.1 ± 0.1; vehicle-treatedOBX, 5.9 ± 0.2; and fluoxetine-treated OBX, 6.0 ± 0.1).
Behavioral effects of 9D-tetrahydrocannabinol in the OBXrat before and after chronic administration of fluoxetineThe effects of the acute administration of D9-tetrahydrocan-nabinol (5 mg/kg, i.p.) in sham and OBX animals were
0
40
80
120
160
Sham OBX
** *
+++
Am
bula
tion
0
5
10
15
20
++
Sham OBX
Vechicle Chronic fluoxetine
Rea
rings
(a)
(b)
Fig. 3 Behavioral effects of OBX and chronic fluoxetine treatment in the
open field test. Chronic treatment (14 days) with fluoxetine (10 mg/kg/
day s.c.) or vehicle commenced 2 weeks after sham and OBX surgery. A
significant increase in the ambulation (a) and rearing (b) scores was
observed (5 min period) in vehicle-treated OBX rats when compared
with vehicle-treated sham rats. OBX-induced hyperactivity was signifi-
cantly attenuated in fluoxetine-treated OBX rats whereas no significant
change was seen in fluoxetine-treated sham rats. Data represent
mean ± SEM. Vehicle-treated sham = 6; fluoxetine-treated sham = 8;
vehicle-treated OBX = 8; fluoxetine-treated OBX = 8; ***p < 0.001
versus vehicle-treated rats; ++p < 0.01, and +++p < 0.001 versus vehi-
cle-treated OBX rats (Bonferroni post hoc test).
0
250
500
750
1000Vechicle
Sham OBX
***
+++
Chronic fluxetine
Densi
ty C
B1
rece
pto
r(B
max:
fm
ol/m
g.
pro
tein
)
-9 -8 -7 -6 -5 -4
100
150
200
250
300
350
400Sham-vehicle
Sham-chronic fluoxetine
OBX-chronic fluoxetine
OBX-vehicle
Stim
ula
tion o
f [3
5S
]GT
PγS
bin
din
g (
% o
f basa
l)
W IN 55,212-2 [log M]
(a)
(b)
Fig. 4 Effect of OBX and chronic treatment with fluoxetine (10 mg/kg/
day, 14 days) or vehicle on CB1 receptor density (Bmax) (a) and CB1
receptor-mediated stimulation of [35S]GTPcS binding (b) in rat pre-
frontal cortical membranes. Note that chronic fluoxetine reversed the
increased density and functionality of CB1 receptors induced by OBX.
Data represent the mean ± SEM of n individuals values obtained from
binding experiments assayed in triplicates (vehicle-treated sham = 6–
7; fluoxetine-treated sham = 7–8; vehicle-treated OBX = 6; fluoxetine-
treated OBX = 7–8); ***p < 0.001 versus vehicle-treated sham group
and +++p < 0.001 versus vehicle-treated OBX group (Bonferroni
post hoc test).
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1423–1433� 2009 The Authors
1428 | A. Rodrıguez-Gaztelumendi et al.
assessed in the open-field test. Regarding the number ofambulations, there was a significant interaction betweensurgery and treatment [F(2,36) = 10.37, p = 0.0003], asignificant main effect of treatment [F(2,36) = 18.79,p < 0.0001] and a significant main effect of surgery[F(1,36) = 12.75, p = 0.0010] (Fig. 5a). A significant inter-action between surgery and treatment [F(2,36) = 74.55,p < 0.0001], a significant main effect of treatment[F(2,36) = 102.23, p < 0.0001] and a significant main effectof surgery [F(1,36) = 72.24 p < 0.0001] were also found forthe number of rearings (Fig. 5b). The ambulation and rearingscores were increased in OBX rats receiving vehicle (n = 6)compared with their counterparts sham animals (n = 6)(p < 0.001 for both parameters). As shown in Fig. 5a and b,acute D9-tetrahydrocannabinol reduced the ambulation score()58%) and the number of rearings ()97%) in OBX rats(n = 8; p < 0.001 vs. OBX animals receiving vehiclefor both parameters). The behavioral response of
fluoxetine-treated OBX rats in the open-field 30 min afterthe administration of a challenge dose of D9-tetrahydrocan-nabinol was similar to that observed in OBX rats followingchronic fluoxetine alone. In addition, D9-tetrahydrocannab-inol did not produce any behavioral effect in sham rats(n = 6) (Fig. 5a and b).
Discussion
One major finding in our study is the presence of an alteredCB1 receptor-signaling in discrete brain areas followingbilateral OBX: an increased CB1 receptor density in PFC andamygdala together with an increased CB1 receptor-mediated[35S]GTPcS binding in the PFC of OBX animals. Theincreases on both CB1 receptor density and CB1 receptor-mediated [35S]GTPcS binding in the PFC of OBX animalswere of similar magnitude. Thus, enhanced CB1 receptor-signaling must be reflecting an up-regulation of CB1 receptorprotein expression. Our finding is in good concordance withstudies reporting a significant increase on both CB1 receptormRNA (Hillard et al. 2006; Bortolato et al. 2007) and CB1
receptor density (Hill et al. 2008) in the PFC of rodentsexposed to chronic mild/unpredictable stress, another vali-dated model of animal depression. Hill et al. (2008) havereported a decrease in CB1 receptor density in hippocampus,hypothalamus, and ventral striatum but no change in theamygdala of rats exposed to chronic unpredictable stress. Bycontrast, we detect a significant increase in CB1 receptordensity in the amygdala of the OBX rats without any changein CB1 receptor-mediated [35S]GTPcS binding (probablybecause of the high level of basal [35S]GTPcS binding thatmakes difficult to detect agonist-induced stimulation). Thisdiscrepancy with our finding could be explained by thedifferent animal model of depression used and/or the type ofbinding assay performed: CB1 receptor autoradiography inour study versus CB1 receptor binding studies in membranehomogenates in the study by Hill et al. (2008). Ourexperimental data are also consistent with clinical studiesdemonstrating elevated levels of ECs, CB1 receptors, andCB1 receptor-mediated [35S]GTPcS binding in PFC ofdepressed suicide victims (Hungund et al. 2004; Pazos et al.2006; Vinod and Hungund 2006), an area where ECs appearto regulate mood, aggression and/or impulsivity, and decisionmaking, which are altered in suicidality (Egerton et al. 2006;Vinod and Hungund 2006).
In our study, chronic fluoxetine exhibited antidepressant-like activity in the OBX rat under the open-field paradigm.It is noteworthy that the antidepressant-like activity offluoxetine in others pre-clinical models seems to bedependent on the test performed, the dose assayed andthe rat strain used (Lopez-Rubalcava and Lucki 2000;Cryan et al. 2005). There are few reports on the effect ofchronic fluoxetine in the OBX rat (Mar et al. 2000; Rocheet al. 2007); moreover, the behavioral effects of the
0
40
80
120
160
200
240
Sham OBX
***
+++
Am
bula
tion
+++
0
5
10
15
20
25
30
Sham OBX
Vehicle THC Chromic fluoxetine+ THC
***
+++
Rea
rings
+++
(a)
(b)
Fig. 5 Behavioral effects of D9-tetrahydrocannabinol (5 mg/kg, i.p.) in
sham and OBX animals before and after the chronic administration of
fluoxetine (10 mg/kg/day, 14 days) assessed in the open-field test.
Acute D9-tetrahydrocannabinol (THC) significantly decreased ambu-
lation (a) and rearing (b) scores in OBX rats before the initiation of the
chronic treatment with fluoxetine. Note that the antidepressant-like
effect of chronic fluoxetine in OBX rats was not modified by the acute
administration of D9-tetrahydrocannabinol. Data represent mean ±
SEM. Vehicle-sham = 6; THC-sham = 6; THC to fluoxetine-treated
sham = 6; vehicle-OBX = 6; THC-OBX = 8; THC to fluoxetine-treated
OBX = 10; ***p < 0.001 versus vehicle-sham rats and +++p < 0.001
versus vehicle-OBX rats (Bonferroni post hoc test).
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1423–1433
Prefrontal CB1 receptors and chronic fluoxetine | 1429

antidepressant appear to be influenced by the environmentalconditions. Our results demonstrate the effectiveness ofchronic fluoxetine (10 mg/kg, 14 days) upon the reversal ofthe behavioral deficits exhibited by OBX-rats when exposedto a high luminance open field exposure, thus under morestressful or aversive conditions (Mar et al. 2002). Ourresults are also in good agreement with those reported byRoche et al. (2007) showing that chronic fluoxetine(10 mg/kg, 5 weeks) reverses all the OBX-associatedchanges in behavior, body temperature, heart rate, andneuronal activation in response to open field exposure. Inour study, we did not detect changes in the number ofdefecations between OBX and sham animals, as alreadyreported (Stock et al. 2000), but chronic fluoxetine pro-voked increased defecation in both groups, althoughreaching statistical significance only in the OBX group.By contrast, it has been shown that chronic fluoxetineattenuates the increased defecations observed in rats afterexposure to chronic stress (Zhang et al. 2000; Huang et al.2007) but it induces diarrhea in non-stressed rats (Zhanget al. 2000). We have not a plausible explanation for theincreased defecations induced by chronic fluoxetine. Itcould because of its action on the gastrointestinal 5-HTsystem (James et al. 2005) and, thus, reflecting a side-effectof the antidepressant at this level. Also, we cannot excludethat chronic fluoxetine could increase feeding (food intakewas not measured) but no differences were observed in thebodyweights of the rats at the end of the treatment.
Another major finding is that chronic fluoxetine exhibitsantidepressant-like activity associated to a normalization ofCB1 receptor-signaling in the PFC. Interestingly, the CBagonist D9-tetrahydrocannabinol (5 mg/kg, i.p.) did alsoattenuate OBX-induced hyperactivity when administeredprior to chronic treatment with fluoxetine. As both acute D9-tetrahydrocannabinol and chronic fluoxetine elicited asimilar behavioral effect in the OBX rat, it is not unlikelythat the regionally selective enhancement of CB1 receptor-signaling in the PFC could be related with the altered OBXbehavior. In our study, acute D9-tetrahydrocannabinol didnot affect locomotor activity of control animals in the open-field as previously described in mice (Egashira et al. 2008).However, a decrease in the number of ambulations on theopen field test in control rats after the acute administrationof D9-tetrahydrocannabinol (Jarbe et al. 2002) and WIN55212-2 (Jarbe et al. 2006; Suplita et al. 2008) has beenreported. This discrepancy could be because of the differentenvironmental conditions under which the test is performedas a more stressful or aversive (high luminance) open fieldtest was used in our study (Mar et al. 2002). Themethodological approaches of our study do not allow tofully ascertaining the functional significance of the changeswe have observed in CB1 receptor-signaling with respect tothe OBX-hyperactivity and to the behavioral effects ofchronic fluoxetine It is noteworthy that D9-tetrahydrocan-
nabinol only produced a behavioral change in OBX ratsbefore the initiation of the chronic treatment with fluoxetineand it did not induce any further change in fluoxetine-treated OBX rats, in which ambulatory behavior was fullynormalized. It is important to mention that adaptive changesin the central serotonergic system appear to play animportant role in the OBX-induced syndrome. Thus, itcould be speculated that the enhanced CB1 receptor-signaling in the PFC of the OBX rat could represent acompensatory mechanism to counteract the behavioral andneurochemical consequences of those adaptive changes;conversely, this compensatory mechanism might not benecessary once the antidepressant has exerted the neuro-chemical and cellular actions that normalize brain 5-HTneurotransmission.
There are several pieces of evidence supporting theinteraction between 5-HT neurotransmission and EC/CB1
receptors that could be relevant to the findings of this study.First, OBX-induced neurodegeneration of serotonin neuronsin DRN is counteracted by antidepressant treatment (Nest-erova et al. 1997); interestingly, CB1 receptors and fatty acidamide hydrolase are present in DRN (Egertova et al. 2003)and CBs elicit antidepressant-like behavior by enhancingDRN 5-HT neuronal activity (Gobbi et al. 2005) through thePFC (Bambico et al. 2007). Second, several changes in 5-HTneurotransmission in cortical areas are encountered followingOBX: altered serotonin metabolism (5-hydroxyindoleaceticacid (5-HIAA)/5-HT ratio) in frontal cortex (Lumia et al.1992), increased 5-HT transporter expression in frontalcortex that is reversed by chronic antidepressants (Greckschet al. 1997; Zhou et al. 1998), decreased extracellular basal5-HT levels in amygdaloid cortex and midbrain (Jancsar andLeonard 1984), and lower tissue 5-HT concentration infrontal cortex (Redmond et al. 1997); on the other hand, CB1
receptor agonists suppress 5-HT release in mouse braincortex (Nakazi et al. 2000) and 5-HT reuptake in rat braincortex (Banerjee et al. 1975) while CB1 receptor antagonistsstimulate 5-HT release into the medial PFC in the shrewbrain (Darmani et al. 2003) and in the rat brain (Tzavaraet al. 2003). Third, changes in 5-HT1A and 5-HT2 receptorexpression have been described in brain cortex of bulbec-tomized mice and rats (Gurevich et al. 1993; Slotkin et al.2005); and, chronic CB treatment appears to up-regulate5-HT2A receptor activity while concurrently down-regulating5-HT1A receptor activity in the rat brain, a finding similar tothat sometimes observed in depression (Hill et al. 2006b).
The molecular mechanism by which the elevated CB1
receptor expression and G protein-coupling efficacy in theOBX rat are reverted by chronic fluoxetine remains to beelucidated. Devlin and Christopoulos (2002) have shown that5-HT reduces the proportion of high affinity binding of theCB agonists HU210 and WIN 55212-2 in rat cerebellarmembranes; these data, suggesting crosstalk between CB1
and 5-HT receptors, may help explaining the effect of
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1423–1433� 2009 The Authors
1430 | A. Rodrıguez-Gaztelumendi et al.

chronic fluoxetine in CB1 receptor density in the PFC. Ourpresent results on the effect of chronic fluoxetine in the OBXrat are in good agreement with the findings reported by Hillet al. (2008) in another animal model of depression:increased CB1 receptor density in PFC of rats exposed tochronic unpredictable stress that is attenuated by chronicimipramine (Hill et al. 2008). Also, it has been reported thatchronic fluoxetine decreases CB1 receptor gene expression incaudate–putamen (Oliva et al. 2005) and that chroniccitalopram induces a decrease in CB receptor-mediated Gprotein coupling in rat hypothalamus and hippocampus, twoareas important in the control of neuroendocrine function andmood (Hesketh et al. 2007); however, chronic administrationof desipramine, a non-SSRI drug, results in an increaseddensity of CB1 receptor in hippocampus and hypothalamus(Hill et al. 2006a). In addition, some interventions known toproduce antidepressant effects in humans have been shown tomodulate the ECs: electroconvulsive shock treatment down-regulates EC/CB1 receptor-signaling in rat PFC and hippo-campus (Hill et al. 2007); however, electroconvulsive shocktreatment up-regulates CB1 receptor signaling in rat amyg-dala (Hill et al. 2007) and sleep deprivation increases thecontent of the EC 2-arachidonylglycerol in rat hippocampus(Chen and Bazan 2005).
Regarding the pharmacological relevance of our findings,it is noteworthy that it is still unclear which would be theadequate pharmacological manipulation of the ECs system toimprove mood or anxiety disorders. CB1 receptor knockoutmice exhibited enhanced anxiety- and depressive-like behav-iors (Haller et al. 2002; Martin et al. 2002; Haller et al.2004; Uriguen et al. 2004). In line with our results using D9-tetrahydrocannabinol, CB stimulation has been recentlyshown to induce antidepressant- and anxiolytic-like effectsin rats (Berrendero and Maldonado 2002; Marco et al. 2004;Gobbi et al. 2005; Bambico et al. 2007) likely via promotinghippocampal neurogenesis (Jiang et al. 2005). However,high doses of CB agonists evoke depressive-like behaviors inmice (Shearman et al. 2003; Tzavara et al. 2003). Thisapparent discrepancy has been attributed to several factors,including doses, animal model, and strain used, as well ascontextual conditions (Viveros et al. 2005). CB1 receptorantagonists/inverse agonists have been recently demonstratedto have antidepressant-like activity in laboratory animals(Shearman et al. 2003; Tzavara et al. 2003; Griebel et al.2005), though the OBX model has not yet been investigated.However, relevant incidence of depression has been reportedfor the antagonist rimonabant in some patients (Griebel et al.2005; Van Gaal et al. 2005).
In conclusion, the data obtained from our study demon-strates that the OBX model of depression is associated withup-regulation and hyperfunctionality of CB1 receptors in thePFC. Furthermore, we demonstrate that this enhanced CB1
receptor-signaling is normalized by chronic administration ofthe SSRI fluoxetine.
Acknowledgements
This work was supported by grants from Ministerio de Educacion y
Ciencia (SAF-2004/00941 and SAF-2007/61862), Fundacion Mutua
Madrilena, Fundacion Koplowitz, and Plan Nacional de Drogas
(2006). AR-G was granted a pre-doctoral fellowship from Ministerio
de Educacion y Ciencia and MLR was a pre-doctoral fellow from
Fundacion UPSA.
Supporting information
Additional Supporting information may be found in the online
version of this article:
Detailed description of methods.Figure S1 Diagram depicting the three set of experiments
performed in this study.
Please note: Wiley-Blackwell are not responsible for the content
or functionality of any supporting materials supplied by the authors.
Any queries (other than missing material) should be directed to the
corresponding author for the article.
References
Bambico F. R., Katz N., Debonnel G. and Gobbi G. (2007) Cannabi-noids elicit antidepressant-like behavior and activate serotonergicneurons through the medial prefrontal cortex. J. Neurosci. 27,11700–11711.
Banasr M. and Duman R. S. (2007) Regulation of neurogenesis andgliogenesis by stress and antidepressant treatment. CNS Neurol.Disord. Drug Targets 6, 311–320.
Banerjee S. P., Snyder S. H. and Mechoulam R. (1975) Cannabinoids:influence on neurotransmitter uptake in rat brain synaptosomes.J. Pharmacol. Exp. Ther. 194, 74–81.
Berrendero F. and Maldonado R. (2002) Involvement of the opioidsystem in the anxiolytic-like effects induced by delta(9)-tetra-hydrocannabinol. Psychopharmacology (Berl.) 163, 111–117.
Bortolato M., Mangieri R. A., Fu J. et al. (2007) Antidepressant-likeactivity of the fatty acid amide hydrolase inhibitor URB597 in a ratmodel of chronic mild stress. Biol. Psychiatry 62, 1103–1110.
Cairncross K. D., Cox B., Forster C. and Wren A. F. (1977) Theolfactory bulbectomized rat: a simple model for detecting drugswith antidepressant potential. Br. J. Pharmacol. 61, 497P.
Castro M. E., Diaz A., del Olmo E. and Pazos A. (2003) Chronic flu-oxetine induces opposite changes in G protein coupling at pre andpostsynaptic 5-HT1A receptors in rat brain. Neuropharmacology44, 93–101.
Chaperon F. and Thiebot M. H. (1999) Behavioral effects of cannabinoidagents in animals. Crit. Rev. Neurobiol. 13, 243–281.
Chen C. and Bazan N. G. (2005) Lipid signaling: sleep, synaptic plas-ticity, and neuroprotection. Prostaglandins Other Lipid Mediat. 77,65–76.
Cryan J. F., Page M. E. and Lucki I. (2005) Differential behavioraleffects of the antidepressants reboxetine, fluoxetine, and moc-lobemide in a modified forced swim test following chronic treat-ment. Psychopharmacology (Berl.) 182, 335–344.
Darmani N. A., Janoyan J. J., Kumar N. and Crim J. L. (2003)Behaviorally active doses of the CB1 receptor antagonist SR141716A increase brain serotonin and dopamine levels andturnover. Pharmacol. Biochem. Behav. 75, 777–787.
Devlin M. G. and Christopoulos A. (2002) Modulation of cannabinoidagonist binding by 5-HT in the rat cerebellum. J. Neurochem. 80,1095–1102.
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1423–1433
Prefrontal CB1 receptors and chronic fluoxetine | 1431

Egashira N., Matsuda T., Koushi E. et al. (2008) Delta(9)-tetrahydro-cannabinol prolongs the immobility time in the mouse forced swimtest: involvement of cannabinoid CB(1) receptor and serotonergicsystem. Eur. J. Pharmacol. 28, 117–121.
Egerton A., Allison C., Brett R. R. and Pratt J. A. (2006) Cannabinoidsand prefrontal cortical function: insights from preclinical studies.Neurosci. Biobehav. Rev. 30, 680–695.
Egertova M., Cravatt B. F. and Elphick M. R. (2003) Comparativeanalysis of fatty acid amide hydrolase and CB1 cannabinoidreceptor expression in the mouse brain: evidence of a widespreadrole for fatty acid amide hydrolase in regulation of endocannabi-noid signaling. Neuroscience 119, 481–496.
Glass M., Dragunow M. and Faull R. L. (1997) Cannabinoid receptors inthe human brain: a detailed anatomical and quantitative auto-radiographic study in the fetal, neonatal and adult human brain.Neuroscience 77, 299–318.
Gobbi G., Bambico F. R., Mangieri R. et al. (2005) Antidepressant-likeactivity and modulation of brain monoaminergic transmission byblockade of anandamide hydrolysis. Proc. Natl Acad. Sci. USA102, 18620–18625.
Gonzalez-Maeso J., Rodriguez-Puertas R., Gabilondo A. M. and MeanaJ. J. (2000) Characterization of receptor-mediated [35S]GTPcSbinding to cortical membranes from postmortem human brain. Eur.J. Pharmacol. 390, 25–36.
Gray J. A. and Lalljee B. (1974) Sex differences in emotional behaviourin the rat: correlation between open-field defecation and activeavoidance. Anim. Behav. 22, 856–861.
Grecksch G., Zhou D., Franke C., Schroder U., Sabel B., Becker A. andHuether G. (1997) Influence of olfactory bulbectomy and sub-sequent imipramine treatment on 5-hydroxytryptaminergic pre-synapses in the rat frontal cortex: behavioural correlates. Br. J.Pharmacol. 122, 1725–1731.
Griebel G., Stemmelin J. and Scatton B. (2005) Effects of the cannabi-noid CB1 receptor antagonist rimonabant in models of emotionalreactivity in rodents. Biol. Psychiatry 57, 261–267.
Gurevich E. V., Aleksandrova I. A., Otmakhova N. A., Katkov Y. A.,Nesterova I. V. and Bobkova N. V. (1993) Effects of bulbectomyand subsequent antidepressant treatment on brain 5-HT2 and 5-HT1A receptors in mice. Pharmacol. Biochem. Behav. 45, 65–70.
Haller J., Bakos N., Szirmay M., Ledent C. and Freund T. F. (2002) Theeffects of genetic and pharmacological blockade of the CB1
cannabinoid receptor on anxiety. Eur. J. Neurosci. 16, 1395–1398.Haller J., Varga B., Ledent C. and Freund T. F. (2004) CB1 cannabinoid
receptors mediate anxiolytic effects: convergent genetic and phar-macological evidence with CB1-specific agents. Behav. Pharmacol.15, 299–304.
Herkenham M., Lynn A. B., Johnson M. R., Melvin L. S., de Costa B. R.and Rice K. C. (1991) Characterization and localization of can-nabinoid receptors in rat brain: a quantitative in vitro autoradio-graphic study. J. Neurosci. 11, 563–583.
Hesketh S. A., Brennan A. K., Jessop D. S. and Finn D. P. (2008) Effectsof chronic treatment with citalopram on cannabinoid and opioidreceptor-mediated G-protein coupling in discrete rat brain regions.Psychopharmacology (Berl.) 198, 26–39.
Hill M. N., Ho W. S., Sinopoli K. J., Viau V., Hillard C. J. and GorzalkaB. B. (2006a) Involvement of the endocannabinoid system in theability of long-term tricyclic antidepressant treatment to suppressstress-induced activation of the hypothalamic-pituitary-adrenalaxis. Neuropsychopharmacology 31, 2591–2599.
Hill M. N., Sun J. C., Tse M. T. and Gorzalka B. B. (2006b) Alteredresponsiveness of serotonin receptor subtypes following long-termcannabinoid treatment. Int. J. Neuropsychopharmacol. 9, 277–286.
Hill M. N., Barr A. M., Ho W. S., Carrier E. J., Gorzalka B. B. andHillard C. J. (2007) Electroconvulsive shock treatment differen-
tially modulates cortical and subcortical endocannabinoid activity.J. Neurochem. 103, 47–56.
Hill M. N., Carrier E. J., McLaughlin R. J., Morrish A. C., Meier S. E.,Hillard C. J. and Gorzalka B. B. (2008) Regional alterations in theendocannabinoid system in an animalmodel of depression: effects ofconcurrent antidepressant treatment. J.Neurochem.106, 2322–2336.
Hillard C. J., Hill M. N., Carrier E. J., Shi L., Cullinan W. E. andGorzalka B. B. (2006) Regulation of cannabinoid receptorexpression by chronic unpredictable stress in rats and mice. Soc.Neurosci. Abstr. 746, 19.
Howlett A. C., Barth F., Bonner T. I. et al. (2002) International union ofpharmacology. XXVII. Classification of cannabinoid receptors.Pharmacol. Rev. 54, 161–202.
HuangY. L., Yu J. P.,WangG.H., Chen Z.H.,WangQ. andXiao L. (2007)Effect of fluoxetine on depression-induced changes in the expressionof vasoactive intestinal polypeptide and corticotrophin releasingfactor in rat duodenum.World J. Gastroenterol. 13, 6060–6065.
Hungund B. L., Vinod K. Y., Kassir S. A., Basavarajappa B. S., Yala-manchili R., Cooper T. B., Mann J. J. and Arango V. (2004)Upregulation of CB1 receptors and agonist-stimulated [35S]GTPcSbinding in the prefrontal cortex of depressed suicide victims. Mol.Psychiatry 9, 184–190.
Jaako-Movits K., Zharkovsky T., Pedersen M. and Zharkovsky A.(2006) Decreased hippocampal neurogenesis following olfactorybulbectomy is reversed by repeated citalopram administration.Cell. Mol. Neurobiol. 26, 1559–1570.
James A. N., Ryan J. P. and Parkman H. P. (2005) Effects of the selectiveserotonin reuptake inhibitor, fluoxetine, on regional gastric con-tractility. Neurogastroenterol. Motil. 17, 76–82.
Jancsar S. M. and Leonard B. E. (1984) Changes in neurotransmittermetabolism following olfactory bulbectomy in the rat. Prog.Neuropsychopharmacol. Biol. Psychiatry 8, 263–269.
Jarbe T. U., Andrzejewski M. E. and DiPatrizio N. V. (2002) Interactionsbetween the CB1 receptor agonist Delta 9-THC and the CB1
receptor antagonist SR-141716 in rats: open-field revisited.Pharmacol. Biochem. Behav. 73, 911–919.
Jarbe T. U., Ross T., DiPatrizio N. V., Pandarinathan L. and MakriyannisA. (2006) Effects of the CB1R agonist WIN-55,212-2 and theCB1R antagonists SR-141716 and AM-1387: open-field exami-nation in rats. Pharmacol. Biochem. Behav. 85, 43–52.
Jarosik J., Legutko B., Unsicker K. and von Bohlen Und Halbach O.(2007) Antidepressant-mediated reversal of abnormal behavior andneurodegeneration in mice following olfactory bulbectomy. Exp.Neurol. 204, 20–28.
Jiang W., Zhang Y., Xiao L., Van Cleemput J., Ji S. P., Bai G. and ZhangX. (2005) Cannabinoids promote embryonic and adult hippocam-pus neurogenesis and produce anxiolytic- and antidepressant-likeeffects. J. Clin. Invest. 115, 3104–3116.
Keilhoff G., Becker A., Grecksch G., Bernstein H. G. and Wolf G.(2006) Cell proliferation is influenced by bulbectomy and nor-malized by imipramine treatment in a region-specific manner.Neuropsychopharmacology 31, 1165–1176.
Kelly J. P., Wrynn A. S. and Leonard B. E. (1997) The olfactory bul-bectomized rat as a model of depression: an update. Pharmacol.Ther. 74, 299–316.
Lopez-Rubalcava C. and Lucki I. (2000) Strain differences in thebehavioral effects of antidepressant drugs in the rat forced swim-ming test. Neuropsychopharmacology 22, 191–199.
Lumia A. R., Teicher M. H., Salchli F., Ayers E. and Possidente B.(1992) Olfactory bulbectomy as a model for agitated hyposero-tonergic depression. Brain Res. 587, 181–185.
Malone D. T. and Taylor D. A. (1998) Modulation of delta9-tetrahy-drocannabinol-induced hypothermia by fluoxetine in the rat. Br. J.Pharmacol. 124, 1419–1424.
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1423–1433� 2009 The Authors
1432 | A. Rodrıguez-Gaztelumendi et al.

Malone D. T. and Taylor D. A. (2001) Involvement of somatodendritic5-HT1A receptors in 9D-tetrahydrocannabinol-induced hypothermiain the rat. Pharmacol. Biochem. Behav. 69, 595–601.
Mar A., Spreekmeester E. and Rochford J. (2000) Antidepressantspreferentially enhance habituation to novelty in the olfactory bul-bectomized rat. Psychopharmacology (Berl.) 150, 52–60.
Mar A., Spreekmeester E. and Rochford J. (2002) Fluoxetine-inducedincreases in open-field habituation in the olfactory bulbectomizedrat depend on test aversiveness but not on anxiety. Pharmacol.Biochem. Behav. 73, 703–712.
Marco E. M., Perez-Alvarez L., Borcel E., Rubio M., Guaza C.,Ambrosio E., File S. E. and Viveros M. P. (2004) Involvement of5-HT1A receptors in behavioural effects of the cannabinoidreceptor agonist CP 55,940 in male rats. Behav. Pharmacol. 15,21–27.
Martin M., Ledent C., Parmentier M., Maldonado R. and Valverde O.(2002) Involvement of CB1 cannabinoid receptors in emotionalbehaviour. Psychopharmacology (Berl.) 159, 379–387.
Mato S. and Pazos A. (2004) Influence of age, postmortem delay andfreezing storage period on cannabinoid receptor density andfunctionality in human brain. Neuropharmacology 46, 716–726.
Mato S., Aso E., Castro E., Martin M., Valverde O., Maldonado R. andPazos A. (2007) CB1 knockout mice display impaired functionalityof 5-HT1A and 5-HT2A/C receptors. J. Neurochem. 103, 2111–2120.
Moldrich G. and Wenger T. (2000) Localization of the CB1 cannabinoidreceptor in the rat brain. an immunohistochemical study. Peptides21, 1735–1742.
Nakazi M., Bauer U., Nickel T., Kathmann M. and Schlicker E. (2000)Inhibition of serotonin release in the mouse brain via presynapticcannabinoid CB1 receptors. Naunyn Schmiedebergs Arch. Phar-macol. 361, 19–24.
Nesterova I. V., Gurevich E. V., Nesterov M., Otmakhova N. A. andBobkova N. V. (1997) Bulbectomy-induced loss of raphe neuronsis counteracted by antidepressant treatment. Prog. Neuropsycho-pharmacol. Biol. Psychiatry 21, 127–140.
Oliva J. M., Uriguen L., Perez-Rial S. and Manzanares J. (2005) Timecourse of opioid and cannabinoid gene transcription alterationsinduced by repeated administration with fluoxetine in the rat brain.Neuropharmacology 49, 618–626.
Onaivi E. S., Ishiguro H., Gong J. P. et al. (2006) Discovery of thepresence and functional expression of cannabinoid CB2 receptorsin brain. Ann. NY Acad. Sci. 1074, 514–536.
Pazos A., Mostany R., Mato S. et al. (2006) Constitutive activity ofcannabinoid CB1 receptors in major depression. Soc. Neuro-sci.Abstr. 191, 18.
Redmond A. M., Kelly J. P. and Leonard B. E. (1997) Behavioural andneurochemical effects of dizocilpine in the olfactory bulbectomizedrat model of depression. Pharmacol. Biochem. Behav. 58, 355–359.
Roche M., Harkin A. and Kelly J. P. (2007) Chronic fluoxetine treatmentattenuates stressor-induced changes in temperature, heart rate, andneuronal activation in the olfactory bulbectomized rat. Neuropsy-chopharmacology 32, 1312–1320.
Shearman L. P., Rosko K. M., Fleischer R., Wang J., Xu S., Tong X. S.and Rocha B. A. (2003) Antidepressant-like and anorectic effectsof the cannabinoid CB1 receptor inverse agonist AM251 in mice.Behav. Pharmacol. 14, 573–582.
Sim L. J., Hampson R. E., Deadwyler S. A. and Childers S. R. (1996)Effects of chronic treatment with delta9-tetrahydrocannabinol oncannabinoid-stimulated [35S]GTPcS autoradiography in rat brain.J. Neurosci. 16, 8057–8066.
Slotkin T. A., Cousins M. M., Tate C. A. and Seidler F. J. (2005)Serotonergic cell signaling in an animal model of aging anddepression: olfactory bulbectomy elicits different adaptations inbrain regions of young adult vs aging rats. Neuropsychopharma-cology 30, 52–57.
Song C. and Leonard B. E. (2005) The olfactory bulbectomised rat as amodel of depression. Neurosci. Biobehav. Rev. 29, 627–647.
van der Stelt H. M., Breuer M. E., Olivier B. and Westenberg H. G.(2005) Permanent deficits in serotonergic functioning of olfactorybulbectomized rats: an in vivo microdialysis study. Biol. Psychiatry57, 1061–1067.
Stock H. S., Ford K. and Wilson M. A. (2000) Gender and gonadalhormone effects in the olfactory bulbectomy animal model ofdepression. Pharmacol. Biochem. Behav. 67, 183–191.
Suplita R. L. II, Eisenstein S. A., Neely M. H., Moise A. M. andHohmann A. G. (2008) Cross-sensitization and cross-tolerancebetween exogenous cannabinoid antinociception and endocanna-binoid-mediated stress-induced analgesia. Neuropharmacology 54,161–171.
Tzavara E. T., Davis R. J., Perry K. W., Li X., Salhoff C., Bymaster F. P.,Witkin J. M. and Nomikos G. G. (2003) The CB1 receptorantagonist SR141716A selectively increases monoaminergic neu-rotransmission in the medial prefrontal cortex: implications fortherapeutic actions. Br. J. Pharmacol. 138, 544–553.
Uriguen L., Perez-Rial S., Ledent C., Palomo T. and Manzanares J.(2004) Impaired action of anxiolytic drugs in mice deficient incannabinoid CB1 receptors. Neuropharmacology 46, 966–973.
Van Gaal L. F., Rissanen A. M., Scheen A. J., Ziegler O., Rossner S. andRIO-Europe Study Group (2005) Effects of the cannabinoid-1receptor blocker rimonabant on weight reduction and cardiovas-cular risk factors in overweight patients: 1-year experience fromthe RIO-europe study. Lancet 365, 1389–1397.
Vinod Y. and Hungund B. L. (2006) Role of the endocannabinoid systemin depression and suicide. Trends Pharmacol. Sci. 27, 539–545.
Vinod K. Y., Arango V., Xie S., Kassir S. A., Mann J. J., Cooper T.B. and Hungund B. L. (2005) Elevated levels of endocanna-binoids and CB1 receptor-mediated G-protein signaling in theprefrontal cortex of alcoholic suicide victims. Biol. Psychiatry57, 480–486.
Viveros M. P., Marco E. M. and File S. E. (2005) Endocannabinoidsystem and stress and anxiety responses. Pharmacol. Biochem.Behav. 81, 331–342.
Witkin J. M., Tzavara E. T., Davis R. J., Li X. and Nomikos G. G. (2005)A therapeutic role for cannabinoid CB1 receptor antagonistsin major depressive disorders. Trends Pharmacol. Sci. 26, 609–617.
Zhang Y., Raap D. K., Garcia F., Serres F., Ma Q., Battaglia G. and Vande Kar L. D. (2000) Long-term fluoxetine produces behavioralanxiolytic effects without inhibiting neuroendocrine responses toconditioned stress in rats. Brain Res. 855, 58–66.
Zhou D., Grecksch G., Becker A., Frank C., Pilz J. and Huether G.(1998) Serotonergic hyperinnervation of the frontal cortex in ananimal model of depression, the bulbectomized rat. J. Neurosci.Res. 54, 109–116.
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1423–1433
Prefrontal CB1 receptors and chronic fluoxetine | 1433




















