
Reduced 5-HT1A- and GABAB Receptor Function in Dorsal Raphe Neurons Upon Chronic Fluoxetine...
39
1 Reduced 5-HT 1A - and GABA B receptor function in dorsal raphé neurons upon chronic fluoxetine treatment of socially stressed rats Running head: Chronic fluoxetine treatment reduces GIRK activation Cornelisse LN *†, Van der Harst JE ‡, Lodder JC *, Baarendse, PJJ †, Timmerman AJ *, Mansvelder HD *, Spruijt BM ‡ and Brussaard AB *§ *Department of Experimental Neurophysiology & †Department of Functional Genomics, Center for Neurogenomics and Cognitive Research (CNCR), Vrije Universiteit (VU) and VU Medical Center (VUmc), De Boelelaan 1087, 1081 HV Amsterdam. ‡ Department of Animals, Science and Society, Ethology and Animal Welfare Group, Faculty of Veterinary Medicine, Universiteit Utrecht, Utrecht. § Corresponding author, Prof. Dr. A.B. Brussaard, Department of Experimental Neurophysiology, CNCR, VU Amsterdam, De Boelelaan 1085, 1081 HV Amsterdam, the Netherlands, tel +31 20 5987098, e-mail: [email protected], fax +31 20 5987112 Number of figures: 5, Number of tables: 1, Number of pages: 28 Keywords: Whole-cell patch-clamp, social defeat, fluoxetine, GABA B receptor, 5-HT 1A receptor, GIRK current Acknowledgements: The authors wish to acknowledge the excellent technical assistance of Tessa Lodder, and comments on earlier drafts of the manuscript by Dr. Matthijs Verhage, Dr. Witte Hoogendijk and Dr. Nail Burnashev. Financial support was provided by the CNCR, NeuroBsik and Centre for Medical Systems Biology (CMSB). Page 1 of 39 Articles in PresS. J Neurophysiol (April 25, 2007). doi:10.1152/jn.00109.2007 Copyright © 2007 by the American Physiological Society.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Reduced 5-HT1A- and GABAB Receptor Function in Dorsal Raphe Neurons Upon Chronic Fluoxetine...

1
Reduced 5-HT1A- and GABAB receptor function in dorsal raphé neurons upon chronic fluoxetine treatment of socially stressed rats
Running head: Chronic fluoxetine treatment reduces GIRK activation
Cornelisse LN *†, Van der Harst JE ‡, Lodder JC *, Baarendse, PJJ †, Timmerman
AJ *, Mansvelder HD *, Spruijt BM ‡ and Brussaard AB *§
*Department of Experimental Neurophysiology & †Department of Functional
Genomics, Center for Neurogenomics and Cognitive Research (CNCR), Vrije
Universiteit (VU) and VU Medical Center (VUmc), De Boelelaan 1087, 1081 HV
Amsterdam. ‡ Department of Animals, Science and Society, Ethology and Animal
Welfare Group, Faculty of Veterinary Medicine, Universiteit Utrecht, Utrecht.
§ Corresponding author, Prof. Dr. A.B. Brussaard, Department of Experimental
Neurophysiology, CNCR, VU Amsterdam, De Boelelaan 1085, 1081 HV Amsterdam,
the Netherlands, tel +31 20 5987098, e-mail: [email protected], fax +31 20 5987112
Number of figures: 5, Number of tables: 1, Number of pages: 28
Keywords: Whole-cell patch-clamp, social defeat, fluoxetine, GABAB receptor, 5-HT1A
receptor, GIRK current
Acknowledgements: The authors wish to acknowledge the excellent technical
assistance of Tessa Lodder, and comments on earlier drafts of the manuscript by Dr.
Matthijs Verhage, Dr. Witte Hoogendijk and Dr. Nail Burnashev. Financial support
was provided by the CNCR, NeuroBsik and Centre for Medical Systems Biology
(CMSB).
Page 1 of 39 Articles in PresS. J Neurophysiol (April 25, 2007). doi:10.1152/jn.00109.2007
Copyright © 2007 by the American Physiological Society.

2
Abstract
Autoinhibitory serotonin 1A receptors (5-HT1A) in dorsal raphé nucleus (DRN) have
been implicated in chronic depression and in actions of selective serotonin reuptake
inhibitors (SSRI). Due to experimental limitations, it was never studied at single cell
level whether changes in 5-HT1A receptor functionality occur in depression and during
SSRI treatment. Here we address this question in a social stress paradigm in rats
that mimics anhedonia, a core symptom of depression. We used whole-cell patch-
clamp recordings of 5-HT- and baclophen-induced G-protein-coupled inwardly
rectifying potassium (GIRK) currents as a measure of 5-HT1A- and GABAB receptor
functionality. 5-HT1A- and GABAB receptor-mediated GIRK-currents were not affected
in socially stressed rats, suggesting that there was no abnormal (auto)inhibition in the
DRN upon social stress. However, chronic fluoxetine treatment of socially stressed
rats restored anticipatory behavior and reduced the responsiveness of 5-HT1A
receptor-mediated GIRK currents. Since GABAB receptor-induced GIRK responses
were also suppressed, fluoxetine does not appear to desensitize 5-HT1A receptors
but rather one of the downstream components shared with GABAB receptors. This
fluoxetine effect on GIRK currents was also present in healthy animals and was
independent of the animal’s “depressed” state. Thus, our data show that symptoms
of depression after social stress are not paralleled by changes in 5-HT1A receptor
signaling in DRN neurons, but SSRI treatment can alleviate these behavioral
symptoms while acting strongly on the 5-HT1A receptor signaling pathway.
Page 2 of 39

3
Introduction
5-HT1A autoreceptors in the DRN have been implicated in mood disorders and
suicide although their direct role remains unclear. In the DRN of patients suffering
from major depression, both increased (Stockmeier et al. 1998) and reduced levels
(Arango et al. 2001) of 5-HT1A autoreceptors have been observed. Changes in 5-
HT1A autoreceptors may result from alterations in 5-HT1A receptor gene transcription
(Parsey et al. 2006), due to polymorphisms in the 5-HT1A receptor promoter region
(Albert and Lemonde 2004; Lemonde et al. 2003; Ou et al. 2003).
Moreover, SSRI treatment in patients may have long term effects on 5-HT1A
receptor levels in the DRN, since depressed patients treated with antidepressants
appear to have lower 5-HT1A receptors levels than antidepressant-naïve patients
(Parsey et al. 2006). Also in rodents desensitization of 5-HT1A autoreceptors as a
result of chronic SSRI administration has been observed (Blier et al. 1998; Czachura
and Rasmussen 2000; Elena Castro et al. 2003; Hensler 2002; Le Poul et al. 1995;
Pejchal et al. 2002; Shen et al. 2002; Subhash et al. 2000). Although it remains
unclear by which mechanism SSRI’s exert there antidepressant action, there is a
clear relation between reduced 5-HT1A receptor function and increased 5-HT levels
in DRN innervated forebrain areas after SSRI treatment in rodents. If SSRI’s are
acutely applied, locally enhanced 5-HT levels in the DRN inhibit serotonernergic
neurons through activation of the 5-HT1A autoreceptor. This results in decreased 5-
HT release in the projection areas (Artigas et al. 1996b). However, after chronic SSRI
treatment 5-HT1A receptors are desensitized and firing of DRN cells is restored
resulting in increased 5-HT levels in the frontal cortex (Artigas et al. 1996b; Bel and
Artigas 1993). Co-application of 5-HT1A receptor antagonists appears to prevent the
initial decrease of 5-HT release in the forebrain during SSRI treatment and may
accelerate antidepressant responses in patients (Artigas et al. 1996b; Blier et al.
1998).
Page 3 of 39

4
These studies suggest a role for DRN 5-HT1A autoreceptors in etology of
depression as well as in the antidepressant action of SSRI’s. However, in these
studies radioligand binding essays and in vivo recordings were used that do not allow
detection of changes in 5-HT1A receptor function at the cellular level. Moreover, the
experiments were performed in normal animals only and not in animal models for
depression.
Here we set out to quantify the 5-HT1A receptor function at the cellular level in
the DRN, using 1) control rats chronically treated with the SSRI fluoxetine, 2) a social
stress animal model for depression and 3) socially stressed rats chronically treated
with fluoxetine. Whole-cell patch-clamp recordings of 5-HT and baclophen (BAC)
induced GIRK currents were used as a measure of 5HT1A and GABAB receptor
functionality (Bayliss et al. 1997; Williams et al. 1988). With this approach we directly
tested if 5-HT1A-mediated inhibition is changed and whether this change is exclusive
for the 5-HT1A pathway or that also the GABAB pathway is affected. Thereby, we
discriminate between changes in receptor levels and changes downstream of the
receptor, since 5-HT1A- and GABAB receptors share the same intracellular pathway
through coupling to a pertussis toxin sensitive G-protein (Innis and Aghajanian 1987;
Innis et al. 1988; Williams et al. 1988).
A social stress paradigm in Wistar rats was used, consisting of one week of
daily social defeat encounters with a dominant intruder followed by approximately 3
months of social isolation. This paradigm induces symptoms resembling anhedonia
in depressed patients (Auriacombe et al. 1997), measured as the absence of
anticipatory behavior in response to sucrose rewards in these animals (Von Frijtag et
al. 2000; Von Frijtag et al. 2002). The anhedonia in rodents is a long-term effect that
correlates with impaired social memory and interactions (Rygula et al. 2005; Von
Frijtag et al. 2000), and is counteracted by imipramine (Von Frijtag et al. 2002) or
fluoxetine (Rygula et al. 2006) as also shown in this study. Using this animal model
Page 4 of 39

5
for depression we investigated the putative relationship between social stress, the
effects of chronic fluoxetine and 5-HT1A and GABAB receptor function in the DRN.
Page 5 of 39

6
Methods
Animals, housing and conditioning
Male Wistar rats (app. 8 wks old; HsdCpb:WU, Harlan, the Netherlands) were used in
experiments all approved by the Animal Ethical Committee of the Vrije Universiteit in
Amsterdam, which act in accordance to European law. The social stress procedure
has been described previously (Von Frijtag et al. 2001; Von Frijtag et al. 2000; Von
Frijtag et al. 2002). In short, the social defeat procedure consisted of daily resident-
intruder sessions on five consecutive days. Each defeat session consisted of a
forced introduction of an adult male rat (intruder) in the territory of a dominant male
rat (resident) and was divided in a pre- (5 min), fight- (5-10 min), and a post (5 min)-
phase. To minimize the risk of severe injuries residential male Long-Evans rats
(LE/CpbHsd, Harlan, UK) were selected for their fighting tactics. Only those rats that
terminated their attacks when the intruder showed submissive behaviors were
allowed to the defeat procedure. As part of the social stress paradigm, defeated
animals were housed individually in macrolon type III cages (30 cm long, 25 cm wide
and 15 cm high) following the first defeat session and control rats were housed
socially (two per cage). Three months later the chronic fluoxetine treatment started,
consisting of daily oral gavage with fluoxetine (10 mg/kg in tap water) or vehicle (tap
water) only, during a subsequent period of 21 days before the behavioral testing or
slice recordings started (fluoxetine HCl was a generous gift from Trifarma SpA
(Milan, Italy). Different animals were used for the behavioral assay and slice
recordings to avoid interference of experimental procedures. The behavioral assay
consisted of classical Pavlovian conditioning as previously described (Von Frijtag et
al. 2000); Van der Harst, 2005). A brief conditioning stimulus (CS, 2 x 60 W light-
input with 1 s interval followed by an auditory signal) was repeatedly paired with an
unconditioned stimulus (US, free access to 5% sucrose solution for 5 min, and of
which the consumption was measured). During a training period of 10 days, the
Page 6 of 39

7
offset of the CS and the onset of the US were progressively increased from 30 s to
10 min over 32 trials. Behavior was observed and analysed from videotape using the
computer program ‘The Observer’ (Noldus Information Technology, Wageningen, the
Netherlands). Behavioral activity displayed in the CS-US interval, reflected by the
frequency of behavioural transitions displayed in the time-interval between
announcement and presentation of the reward, was used as a parameter for
anticipation and reflects the level of reward-sensitivity (Von Frijtag et al. 2000; Von
Frijtag et al. 2002). Significant differences within groups between baseline and
posttraining testing was analyzed by paired t-test.
Slice recordings
Rats were anesthetized in a closed compartment with 4 ml Isoflurane (Abbot
Laboratories Ltd, Queenborough, Kent, U.K.) sprinkled on a tissue and decapitated
using a guillotine. Coronal midbrain slices (400 µm) were prepared in ice-cold slice
solution containing (in mM): 3.5 KCl, 2.4 CaCl2, 1.3 MgSO4, 1.2 KH2PO4, 212.5
Sucrose, 26 NaHCO3, 10 D-glucose, carboxygenated in 5% CO2 / 95% O2. Slices
were transferred to ACSF containing (in mM): 125 NaCl, 3 KCl, 1.2 NaH2PO4, 2.4
CaCl2, 1.3 MgSO4, 10 D-glucose, 25 NaHCO3, supplemented with 20 µM DNQX to
prevent overexcitation by glutamatergic inputs. Slices were kept at 37º C for 1 hour to
recover and were subsequently stored at room temperature. The recording chamber
was continuously perfused with ACSF, 20 µM DNQX and 20 µM bicuculline to block
AMPA and GABAA receptors. Cells were recorded in the whole-cell configuration in
voltage-clamp and current-clamp mode. To measure the I-V relation of GIRK currents
slow descending and ascending voltage ramps were applied between -60mV and -
120mV in figure 1-4, and between -60 and -100mV in figure 5. Patch pipettes had a
resistance of 2-2.5 MΩ. The internal solution contained (in mM) 5 NaCl, 1 MgCl2, 130
C6H11O7K (K-Gluconate), 10 HEPES, 2 Na2-ATP, 0.5 Tris-GTP, 0.02 EGTA. Series
Page 7 of 39

8
resistance was not compensated. Average access resistance at the end of the
recording was 9.6 MΩ (±0.3 SEM).
Large DRN neurons were filled through the recording pipette with 120 µM
Alexa 594 (Molecular Probes, Eugene, OR) dissolved in intracellular current clamp
medium. After fully loading with dye (15-30 min after break-in), these cells were
visualized with a Leica MP-RS 2 photon laser microscope (Leica, Mannheim,
Germany) and image projections (at 1 µm intervals) were obtained with Leica LCS
software and ImageJ (NIH), as described previously (de Rover et al. 2004).
The agents used in this study (5-HT, baclophen, DNQX, bicuculline) were
obtained from RBI (Natick, USA). Order of application in figure 2-5 was 5 µM 5-HT,
wash, 100 µM 5-HT or 1mM BAC. For this study we performed our recordings in the
absence of tryptophan loading (Liu et al. 2005). These conditions, which are
unfavorable for sustaining endogenous 5-HT synthesis, make it unlikely that
endogenous 5-HT secretion through spontaneous firing of 5-HT have contributed to
basal current responses in the absence of exogenous 5-HT application.
Data analysis
Current-voltage (I-V) curves were obtained by averaging the I-V responses during the
negative and positive slope of the voltage-clamp protocol in figure 1 C.
Conductances were calculated from the slope of the straight line fitted to the linear
part of the IV curves. 5-HT and BAC induced conductances shown in figure 2-5 were
corrected for basal conductance by subtraction of the basal conductance level at the
start of each experiment (Reanalysing the data by normalizing the induced GIRK
conductances to the basal conductance in each recording did not change the results;
not shown). Basal conductances were not different between the experimental groups
shown in figure 2-5 (figure 2: control + water 1.06±0.07 nS, control + fluoxetine
1.20±0.14 nS; figure 3: control 1.43±0.18 nS, social stress 1.65±0.16 nS; figure 4:
social stress + water 1.25±0.09 nS, social stress + fluoxetine 1.26±0.07 nS; figure 5:
Page 8 of 39

9
control + fluoxetine 1.73±0.54 nS, social stress + fluoxetine 1.65±0.61 nS) For
statistical analysis of the effects of different pharmaca within a given experimental
group (ie. comparison of the effect of 5 versus 100 µM 5-HT within the control for
instance) we used a two sample paired t-test assuming unequal variance (two tailed).
For comparison between experimental groups of the induced effect of 5-HT or BAC
at any given concentration and the conductance levels after wash we used a regular
ANOVA followed by a post hoc test (Boniferoni mode) for multiple comparisons. In
some instances, to increase statistical power for the data presented, we pooled the
data if experiments were conducted under identical experimental conditions; this was
done in particular for data shown in Fig. 2, where the control + fluoxetine data were
pooled with those also shown in Fig. 5. Data of Fig. 3 and 4 could not be pooled or
compared since the animal handling conditions were not identical, i.e. in Fig. 3
experimental animals were not treated orally either with water or fluoxetine, whereas
for data obtained for Fig. 4 they were. The significance found is indicated in the
figures with NS, *, ** or *** for not significant, p < 0.05, 0.01 and 0.005 respectively.
Error-bars show SEM. Not shown is the fact that all “wash conductances” were not
significantly different between experimental groups and that all 5-HT or BAC induced
conductances in figures 2-5 were significantly different from conductances before
application (at p < 0.01) unless mentioned otherwise explicitly (see figure 4 and figure
5, responses to 1 mM BAC). In addition, we used a multivariate ANOVA to test the
putative interaction between social defeat and SSRI treatment on GIRK current
activation by either 5-HT or BAC. Reanalysing the data by normalizing the induced
GIRK conductances to the basal conductance in each recording did not change the
results (not shown).
Page 9 of 39

10
Results
Assessing 5-HT1A receptor function in DRN neurons: GIRK currents
5-HT mediated auto-inhibition of serotonergic cells is extensively studied in rodents.
5-HT induces a hyperpolarizing current by activating the 5-HT1A receptor that is
coupled to a G-protein coupled potassium (GIRK) channel (Bayliss et al. 1997;
Katayama et al. 1997; Penington et al. 1993a; 1993b; Williams et al. 1988). Besides
5-HT1A receptors also GABAB receptors are coupled to GIRK channels in raphe
nucleus 5-HT cells (Bayliss et al. 1997; Innis and Aghajanian 1987; Innis et al. 1988;
Williams et al. 1988). This allows an accurate quantification on the cellular level of
the 5-HT1A receptor-mediated inhibition relative to the GABAB receptor-mediated
inhibition.
In whole-cell recordings from neurons in the ventral portion of the DRN we
distinguished between 5-HT-containing neurons and non-5-HT neurons based on
morphology and electrophysiological characteristics. The putative 5-HT neurons had
a morphology of magnocellular neurons (figure 1 A) and showed membrane potential
profiles in response to a series of current steps that were typical of 5-HT neurons
(figure 1 B, see (Li et al. 2001). In our experiments 89 % (n = 196) of the cells
displayed the characteristic electrophysiological profile of 5-HT neurons. The vast
majority of these cells fired spontaneously at the onset of recordings, unless slices
were exposed to tryptophan prior to recording (Liu et al. 2005) (n = 15, not shown). In
that case, DRN neurons secrete 5-HT within the nucleus, which hyperpolarizes DRN
neurons and reduce spontaneous activity (Johnson et al. 2002; Liu et al. 2005). In
the absence of the 5-HT precursor tryptophan, endogenous paracrine 5-HT release
is low or absent in DRN slices (Johnson et al. 2002), providing a good starting point
for quantitative measures of the exogenous 5-HT-induced GIRK responses. Current
responses were quantified by calculating the conductances from the measured IV
curves.
Page 10 of 39

11
Application of 5-HT induced a robust hyperpolarization of putative 5-HT
neurons (figure 1 C) but not of non-5-HT neurons (not shown). Subsequently, in the
same recording we switched to voltage-clamp and applied descending and
ascending voltage ramps, both in the absence and presence of various
concentrations of 5-HT (figure 1 C bottom traces). In the presence of 5-HT, the slope
of the current response during voltage ramps was much steeper, indicating that 5-HT
activated a GIRK conductance as reported previously (Bayliss et al. 1997; Katayama
et al. 1997; Penington et al. 1993a; 1993b; Williams et al. 1988) Subtraction of the
control current from the current during 5-HT application yielded the 5-HT activated
current with a reversal potential of -86 ± 7 mV (SD, figure 1 D, n = 38). This is in line
with the Nernst equilibrium potential for GIRK potassium currents calculated from the
intracellular and extracellular solutions used and corrected for the liquid junction
potential (-81 mV). Comparing conductances at different 5-HT concentrations
resulted in a dose-response relation with an EC50 of 2.7 µM 5-HT and 100 µM 5-HT
induced maximal GIRK conductance (figure 1 E).
Chronic fluoxetine reduces 5-HT1A and GABAB induced GIRK-currents
It has been suggested that during treatment with SSRIs, 5-HT1A receptors on DRN
neurons desensitize (Elena Castro et al. 2003; Hensler 2002; Le Poul et al. 1995;
Pejchal et al. 2002; Shen et al. 2002; Subhash et al. 2000). To test whether
fluoxetine treatment would reduce 5-HT1A receptor-mediated inhibition, healthy rats
were treated for 21 days with daily oral administrations of fluoxetine. The 5-HT-
induced GIRK responses were probed with two 5-HT concentrations: 5 µM, close to
the EC50, and 100 µM. In line with data shown in figure 1 E, in DRN slices from
water-treated animals 100 µM 5-HT induced a significantly larger GIRK conductance
than 5 µM 5-HT (figure 2 A-B, E). In contrast, in fluoxetine-pretreated animals, paired
comparison of the GIRK responses induced by these two 5-HT concentrations
Page 11 of 39

12
showed that they were not significantly different (figure 2 A-B, E). Moreover, while
the response to 5µM 5-HT upon chronic fluoxetine was similar as in the water
treated group, the 100µM 5-HT response was significantly reduced, This implies that
5-HT1A receptor functionality is reduced after chronic fluoxetine treatment.
To examine whether the effect of fluoxetine treatment was unique to the 5-
HT1A receptor pathway, we also tested whether the GABAB receptor-induced GIRK
currents were affected. GIRK responses induced by a maximal dose of the GABAB
receptor agonist BAC (1 mM) were significantly reduced in 5-HT neurons after
fluoxetine treatment (figure 2 C-D, and F). This suggests that fluoxetine treatment
affects 5-HT1A receptor functionality downstream of the 5-HT1A receptor and at a level
of the signaling pathway that is shared with the GABAB receptor to the GIRK-
pathway.
Recently, it was claimed (Takahashi et al. 2006) that fluoxetine may suppress
basal GIRK conductance acutely and directly. To test whether the suppression of the
maximal GIRK current activation in our experiments was due to a chronic, rather than
to an acute or direct effect of fluoxetine, we measured basal conductance levels
before and after the acute application of 33 µM fluoxetine for 4 min. This
concentration corresponds to the theoretical maximal concentration of fluoxetine
when a dose of 10 mg/kg is used in vivo and is about 100 times higher than plasma
levels of fluoxetine measured after 10mg/kg injected i.p. or with osmotic minipumps
(Czachura and Rasmussen 2000). We found that acute exposure to fluoxetine
increased, rather than decreased, the normalized basal conductance (from 1 ± 0.15
nA to 1.33 ± 0.09 nA, n = 10, paired t-test, p < 0.002). We cannot exclude that this is
due to a fluoxetine-mediated increase in residual endogenously released,
extracellular 5-HT levels as a consequence of the acute block of 5-HT reuptake in the
brain slices. Furthermore, 5 µM 5-HT elicited normal GIRK responses in the
presence of fluoxetine (2.73 ± 0.38 fold increase, n = 6). Thus, in our recordings we
Page 12 of 39

13
find no evidence for a direct inhibition by fluoxetine of GIRK conductance level or of
5-HT receptor mediated GIRK currents.
Social defeat does not affect 5-HT1A receptor responses
To test whether 5-HT1A receptor function is affected in DRN neurons of animals that
exhibit symptoms of depression, we recorded 5-HT-induced GIRK currents in DRN 5-
HT neurons from rats that were subjected to a social stress paradigm (Von Frijtag et
al. 2000; Von Frijtag et al. 2002). These rats were compared to control rats that had
not received social defeat encounters nor isolation. It has previously been reported
that rats subjected to this social stress paradigm show anhedonia, a symptom of
depression, reflected by a suppression of in anticipatory behavior in expectation of a
reward. In the present experiments, socially stressed rats were also observed to
show a supressed anticipatory behavior as an indication for anhedonia (Table I).
When 5-HT-induced GIRK currents were recorded in DRN 5-HT neurons, no
differences were found between control and socially stressed animals. In both groups
GIRK currents were activated by 5-HT in a dose-dependent manner and to a similar
extent (figure 3 A-B and E). This suggests that the social stress paradigm does not
change 5-HT1A receptor function in DRN 5-HT neurons. Similarly, the maximal GIRK
conductance induced by the GABAB receptor agonist BAC was also not affected
upon social defeat stress (figure 3 C-D, and F). Thus, the induction of depression-like
symptoms in rats by social defeat stress does not affect 5-HT1A receptors or one of
the downstream components.
Rescue of anticipatory behavior after social stress by fluoxetine correlates with
reduced 5-HT1A- and GABAB receptor functionality
This insensitivity for reward (anhedonia), reflected by the impaired anticipatory
behavior in socially stressed rats was previously shown to be reversed by chronic
treatment of the antidepressant imipramine (Von Frijtag et al. 2002). We also
Page 13 of 39

14
observed relief from anhedonia-like symptoms in socially stressed rats after
treatment with fluoxetine for 21 days. In fluoxetine treated animals, anticipatory
behavior toward sucrose rewards was restored after three weeks of treatment
whereas in vehicle treated animals anticipatory behavior remained impaired (Table I).
No effect of social stress and/or fluoxetine treatment on the intake of 5% sucrose
consumption was observed (data not shown). To test whether the behavioral effect of
chronic fluoxetine treatment correlated with a cellular effect of fluoxetine in the DRN,
we examined GIRK currents induced by 5-HT and BAC in DRN 5-HT neurons of
animals subjected to the same social stress paradigm with or without subsequent
chronic fluoxetine treatment (figure 4 A-B, and E). In 5-HT neurons of socially-
stressed fluoxetine-treated animals, 100 µM 5-HT did not produce larger responses
than 5µM 5-HT (figure 4 B and E), similar to the effect seen of chronic fluoxetine
treatment in control animals (figure 2 B and E). Moreover the responsiveness to
either 5µM or 100µM 5-HT was significantly reduced when compared to socially
stressed animals not treated with fluoxetine (figure 4 E). Thus the 5-HT
responsiveness of DNR neurons in terms of 5-HT induced GIRK activation appeared
to be suppressed by fluoxetine-treatment in socially stressed animals, similar to the
apparent effect of chronic fluoxetine in control animals. However, the apparent
relative effect of fluoxetine upon social stress appeared to be larger than in control
animals (compare figure 4 E and figure 2 E respectively; see also below).
To test whether fluoxetine treatment reduced 5-HT1A receptor functionality
downstream of the 5-HT1A receptor, we also examined BAC-induced GIRK currents.
Similar to the effect of fluoxetine-treatment in healthy rats, upon social stress
conditioning, the BAC-induced GIRK currents were significantly reduced in 5-HT
neurons after fluoxetine treatment (figure 4 C-D, and F). This suggests that in social
defeated animals a reduced functionality of the 5-HT1A receptor coupling to GIRK
induced by fluoxetine treatment is expressed downstream of the 5-HT1A receptor and
Page 14 of 39

15
most likely at a level of the signaling pathway that is shared with the GABAB receptor
to the GIRK-pathway. Again the relative effect of fluoxetine upon social defeat
appeared larger than in control animals (compare figure 4 F and figure 2 F
respectively).
Fluoxetine acts on 5-HT1A receptor functionality independent of social
conditions
Fluoxetine appears to have mood-elevating effects predominantly in depressive
patients. In healthy human subjects, no significant effects of fluoxetine on mood and
other psychological variables were found (Gelfin et al. 1998). As indicated above, in
our experiments, fluoxetine treatment appeared to affect 5-HT- and GABAB GIRK
activation in DRN 5-HT neurons of both control and socially stressed rats, but
possibly to a different extent. We directly addressed the hypothesis that fluoxetine
treatment has a stronger impact on 5-HT1A receptor responses in DRN 5-HT neurons
of animals upon social stress compared to control conditioning. To this end, we
tested 5-HT-induced GIRK activation in fluoxetine-treated, socially stressed rats and
in fluoxetine-treated, control rats. Recordings of DRN 5-HT neurons of these animals
showed that, while 5 µM 5-HT induced GIRK responses were significantly different
from basal levels, the 5-HT-induced GIRK conductance at 5 µM and 100 µM were not
different in either group (figure 5 A, B). Thus, no significant differences were found in
the relative effect of fluoxetine between the control and socially stressed groups
(figure 5 E). Similarly, BAC-induced GIRK currents were similarly suppressed in both
groups (figure 5 F), indicating that also in these experiments fluoxetine acted
downstream of the 5-HT1A receptor. The BAC-induced GIRK conductance change
was also not different from 100 µM 5-HT responses, either in control and in socially
stressed animals (figure 5 E and F). These data show that fluoxetine acts on DRN 5-
HT neurons independent of whether animals show symptoms of depression.
Page 15 of 39

16
Subsequent multivariate ANOVA revealed no interaction between social
defeat and fluoxetine treatment. Simple contrast (K matrix) testing, for all data pooled
in this study, showed no significant effect of social stress (compared to controls, p =
0.563 and p = 0.154). Basal conductances were not different between social stress
and control animals. In contrast, chronic treatment with fluoxetine produced robust
overall effects (p = 0.006 and p = 0.001 respectively for 5 and 100 µM 5-HT, with no
effects on basal responses). Additional multivariate analysis of possible interaction
between social stress and chronic SSRI treatment revealed no significance. This
confirms that DRN 5-HT neurons in socially stressed animals are not more sensitive
to fluoxetine treatment than 5-HT neurons of control animals.
Page 16 of 39

17
Discussion
In this study, we addressed the question of whether 5-HT1A receptor function in
dorsal raphé nucleus 5-HT neurons is affected in animals that show symptoms of
depression and upon fluoxetine treatment. Rats exposed to social stress exhibited
behavioral symptoms of depression that could be relieved by fluoxetine treatment. In
socially stressed animals, neither 5-HT1A - nor GABAB receptor-induced GIRK
currents in DRN 5-HT neurons were different in amplitude from GIRK currents in
healthy animals. However, when these animals were treated with fluoxetine for 21
days, both 5-HT1A- and GABAB receptor mediated GIRK responses were markedly
reduced. This implies that fluoxetine acts on downstream components shared by
both receptor-types. We found that fluoxetine treatment also affects both 5-HT1A- and
GABAB receptor functionality in 5-HT neurons of healthy animals. When directly
tested, the effects of fluoxetine treatment on 5-HT1A- and GABAB induced GIRK
currents were not different in healthy and socially stressed animals, indicating that
the action of fluoxetine is independent of the animal’s “depressed” state.
5-HT1A receptor function in the DRN and depression
Previous studies on depressed patients suggest either a positive or an inverse
correlation between chronic depression and 5-HT1A receptor levels in the brain. In
postmortem brains of patients suffering from major depression, increased
(Stockmeier et al. 1998) and decreased (Arango et al. 2001) levels of 5-HT1A
receptors were reported. In patients with bipolar or late-life depression, reduced
levels of 5-HT1A receptors were found (Drevets et al. 2000; Drevets et al. 1999;
Meltzer et al. 2004). In socially stressed rats that show behavioral symptoms of
depression, we found no changes in levels or functionality of 5-HT1A-receptors at the
level of DRN neurons. Our results suggest that in DRN neither receptor levels, nor
downstream components, such as G-proteins or GIRK channel activity, are affected
Page 17 of 39

18
after severe social stress. This suggests that the behavioral symptoms of depression
observed in these animals, such as a loss of reward-sensitivity, an impaired social
memory and social interaction behavior that resemble anhedonia and motivational
deficits in humans (Von Frijtag et al. 2001; Von Frijtag et al. 2000; Von Frijtag et al.
2002), are brought about by other mechanisms than an alteration in 5-HT1A-receptor
signaling in DRN 5-HT neurons. These results appear in line with findings in
monkeys that show symptoms of behavioral depression. In these animals, 5-HT1A
receptor binding at the level of the raphé nucleus (Shively et al. 2006) were not
affected.
Chronic treatment with the SSRI fluoxetine did affect 5-HT1A receptor
functionality in DRN 5-HT neurons, both in healthy and socially stressed animals.
Fluoxetine treatment also restored anticipatory responses in the latter group (Table
I), just as the antidepressant imipramine reverses the symptoms of anhedonia after
social stress (Von Frijtag et al. 2001). Previous studies have shown that fluoxetine
treatment affects 5-HT1A receptor-stimulated G-protein-binding in the DRN in healthy
rats (Elena Castro et al. 2003; Hensler 2002; Pejchal et al. 2002; Shen et al. 2002).
In transgenic mice lacking 5-HT transporter protein, similar adaptive changes in 5-
HT1A receptor signaling were found in DRN (Fabre et al. 2000; Gobbi et al. 2001;
Holmes et al. 2003; Li et al. 2000; Mannoury la Cour et al. 2001; Mannoury la Cour et
al. 2004). Recently, it was confirmed that fluoxetine treatment can alleviate
behavioral deficits evoked by chronic social stress in rats (Rygula et al. 2006). How
reduced 5-HT1A receptor functionality in the DRN contributes to the suppression of
depressive symptoms in socially stressed animals is not clear. However, previous
studies show a clear relation between 5-HT1A receptor mediated autoinhibition at the
DRN level and 5-HT levels in the projection areas. 5-HT levels in DRN innervated
regions and the activity of serotonergic neurons are both reduced after acute
application of SSRI’s (Artigas et al. 1996a; Artigas et al. 1996b; Czachura and
Rasmussen 2000). This is attributed to the inhibiting action of 5-HT1A receptors that
Page 18 of 39

19
are activated by the increased concentration of 5-HT in the DRN after blocking 5-HT
reuptake. Long-term treatment with SSRI’s produces a robust increase in
extracellular 5-HT levels in the projection area’s and restores firing activity of
serotonergic cells, which is consistent with the desensitization of 5-HT1A receptors in
the DRN (Bel and Artigas 1993; Ferrer and Artigas 1994; Czachura and Rasmussen
2000). The similar time course of 5-HT levels in frontal cortex and the antidepressant
response by SSRI’s suggests that these are related. This idea is supported by the
fact that coaplication of SSRI’s with a 5-HT1AR antagonist like pindolol prevents the
initial reduction of 5-HT in forebrain projection areas and accelerates antidepressant
action (Artigas et al. 1996b). Our findings therefore suggests that the restoration of
hedonic behavior in socially stressed animals after treatment with fluoxetine might
possibly be due to augmented serotonergic output of DRN as a result of reduced 5-
HT1A receptor functionality in DRN.
Action of fluoxetine at the cellular level
The observed reduction of 5-HT-induced GIRK currents after chronic SSRI treatment
may result from desensitization of 5-HT1A receptors or may be due to desensitization
of a component downstream of the receptor. The fluoxetine-induced reduction in 5-
HT1A receptor functionality is paralleled by a reduction in GABAB receptor-mediated
GIRK activation after treatment with fluoxetine, both in healthy as well as in socially
stressed animals. Although a parallel effect of fluoxetine on both receptors can not be
excluded it is more likely that fluoxetine treatment affects 5-HT1A receptor signaling in
DRN 5-HT neurons downstream of the receptors, since both 5-HT1A and GABAB
receptors couple to the same type of GIRK channel through Gi-proteins in these
neurons (Innis and Aghajanian 1987; Innis et al. 1988; Williams et al. 1988).
Radioligand binding assays suggest that chronic fluoxetine treatment reduces the
coupling of 5-HT1A-receptors and G-proteins without changing 5-HT1A-receptor levels
Page 19 of 39

20
(Elena Castro et al. 2003; Hensler 2002; Pejchal et al. 2002; Shen et al. 2002). In
transgenic mice in which 5-HT reuptake is prevented by knocking out the 5-HT
transporter (5-HTT null mice), functionality of 5-HT1A- as well as GABAB receptors is
strongly reduced in DRN 5-HT cells (Mannoury la Cour et al. 2004), supporting the
idea that both receptor types partly share a common signal transduction pathway in
5-HT neurons.
Thus far, the best supported candidate mechanism by which fluoxetine
treatment leads to inhibition of GIRK channel activity is through uncoupling of Gi-
proteins from their receptors (Elena Castro et al. 2003; Hensler 2002; Pejchal et al.
2002; Shen et al. 2002). Alternative mechanisms could be through PKC-dependent
phosphorylation of GIRK channels (Mao et al. 2004), depletion of PIP2, an
endogenous co-factor necessary for GIRK channel activity (Cho et al. 2005; Lei et al.
2003) or through a glucocorticoid-mediated modulation of GIRK responsiveness
(Fairchild et al. 2003). Recently, the background potassium channel TREK-1 was
proposed as a target for antidepressant action in DRN neurons ((Heurteaux et al.
2006)). In this model, fluoxetine acts as antidepressant through direct inhibition of
TREK-1 channels. However, in our experiments, we could not detect a direct
inhibition of background whole-cell potassium currents by fluoxetine in DRN 5-HT
neurons that would support this model.
GABAB receptors and the action of SSRI’s
There is an emerging body of data purporting a role for GABAB receptors as potential
targets for antidepressant actions (Mombereau et al. 2004a; Mombereau et al. 2005;
Petty 1995; Slattery et al. 2005). Although previous studies reported no change in
GABAB receptor levels (Cross et al. 1988) or functionality (Monteleone et al. 1990) in
depressed patients, more recent studies showed antidepressant effects of GABAB
antagonists. Transgenic mice lacking either GABAB1 or GABAB2 receptor subunits
show enhanced resistance against symptoms of depression in the forced swim test
Page 20 of 39

21
(Mombereau et al. 2005; Mombereau et al. 2004b; Slattery et al. 2005). We show
that chronic SSRI treatment suppresses GABAB receptor functionality of DRN
neurons, confirming a possible role for GABAB receptors as target for
antidepressants. Since local GABAergic interneurons in the DRN are activated by 5-
HT through 5-HT2A/C receptors (Liu et al. 2000), GABA signaling through GABAB
receptors is likely to provide an additional negative feedback loop of 5-HT neuron
activity. Therefore, reduction of GABAB receptor functionality may have a similar
disinhibiting effect of 5-HT neuron activity as decreased 5-HT1A receptor functionality.
This model may further suggest that co-application of GABAB antagonists might
speed up the antidepressant action of SSRI’s by blocking GABAB receptor activity.
Significance from a clinical perspective
The prevalent view on the cellular mechanisms underlying the origin of depression in
humans and the cellular mechanisms of SSRI treatment is that they both reside in
the serotonergic system. Our results suggest that these mechanisms could diverge.
On the one hand, severe social stress can induce behavioral symptoms of
depression, such as anhedonia, without affecting 5-HT1A receptor functionality in 5-
HT neurons. On the other hand, fluoxetine treatment can alleviate these symptoms of
depression by reducing 5-HT1A receptor functionality in DRN 5-HT neurons. Thus,
although the possibility remains that stress hormones affects components in the
serotonergic cell system other than GIRK currents (Abrams et al. 2005; Lowry et al.
2000), it becomes more and more likely that non-aminergic pathways located in
target areas of the DRN may also be involved (Berton and Nestler 2006). Also in the
latter model, antidepressants are thought to act primarily on the serotonergic system
(Duman 2002; Duman et al. 1999). Cause and therapy of depression may therefore
diverge. Using animal models, such as social stress in rodents, to identify the
neuronal mechanisms underlying both cause and therapy of mood disorders is
Page 21 of 39

22
indispensable to eventually get from palliative to curative treatments of these
disorders in the future.
Page 22 of 39

23
References
Abrams JK, Johnson PL, Hay-Schmidt A, Mikkelsen JD, Shekhar A, and Lowry CA. Serotonergic systems associated with arousal and vigilance behaviors following administration of anxiogenic drugs. Neuroscience 133: 983-997, 2005. Albert PR, and Lemonde S. 5-HT1A receptors, gene repression, and depression: guilt by association. Neuroscientist 10: 575-593, 2004. Arango V, Underwood MD, Boldrini M, Tamir H, Kassir SA, Hsiung S, Chen JJ, and Mann JJ. Serotonin 1A receptors, serotonin transporter binding and serotonin transporter mRNA expression in the brainstem of depressed suicide victims. Neuropsychopharmacology 25: 892-903, 2001. Artigas F, Bel N, Casanovas JM, and Romero L. Adaptative changes of the serotonergic system after antidepressant treatments. Adv Exp Med Biol 398: 51-59, 1996a. Artigas F, Romero L, de Montigny C, and Blier P. Acceleration of the effect of selected antidepressant drugs in major depression by 5-HT1A antagonists. Trends Neurosci 19: 378-383, 1996b. Auriacombe M, Reneric JP, and Le Moal M. Animal models of anhedonia. Psychopharmacology (Berl) 134: 337-338; discussion 371-337, 1997. Bayliss DA, Li YW, and Talley EM. Effects of serotonin on caudal raphe neurons: activation of an inwardly rectifying potassium conductance. J Neurophysiol 77: 1349-1361, 1997. Bel N, and Artigas F. Chronic treatment with fluvoxamine increases extracellular serotonin in frontal cortex but not in raphe nuclei. Synapse 15: 243-245, 1993. Berton O, and Nestler EJ. New approaches to antidepressant drug discovery: beyond monoamines. Nat Rev Neurosci 7: 137-151, 2006. Blier P, Pineyro G, el Mansari M, Bergeron R, and de Montigny C. Role of somatodendritic 5-HT autoreceptors in modulating 5-HT neurotransmission. Ann N Y Acad Sci 861: 204-216, 1998. Cho H, Lee D, Lee SH, and Ho WK. Receptor-induced depletion of phosphatidylinositol 4,5-bisphosphate inhibits inwardly rectifying K+ channels in a receptor-specific manner. Proc Natl Acad Sci U S A 102: 4643-4648, 2005. Cross JA, Cheetham SC, Crompton MR, Katona CL, and Horton RW. Brain GABAB binding sites in depressed suicide victims. Psychiatry Res 26: 119-129, 1988. Czachura JF, and Rasmussen K. Effects of acute and chronic administration of fluoxetine on the activity of serotonergic neurons in the dorsal raphe nucleus of the rat. Naunyn Schmiedebergs Arch Pharmacol 362: 266-275, 2000. de Rover M, Mansvelder HD, Lodder JC, Wardeh G, Schoffelmeer AN, and Brussaard AB. Long-lasting nicotinic modulation of GABAergic synaptic transmission in the rat nucleus accumbens associated with behavioural sensitization to amphetamine. Eur J Neurosci 19: 2859-2870, 2004. Drevets WC, Frank E, Price JC, Kupfer DJ, Greer PJ, and Mathis C. Serotonin type-1A receptor imaging in depression. Nucl Med Biol 27: 499-507, 2000. Drevets WC, Frank E, Price JC, Kupfer DJ, Holt D, Greer PJ, Huang Y, Gautier C, and Mathis C. PET imaging of serotonin 1A receptor binding in depression. Biol Psychiatry 46: 1375-1387, 1999. Duman RS. Structural alterations in depression: cellular mechanisms underlying pathology and treatment of mood disorders. CNS Spectr 7: 140-142, 144-147, 2002.
Page 23 of 39

24
Duman RS, Malberg J, and Thome J. Neural plasticity to stress and antidepressant treatment. Biol Psychiatry 46: 1181-1191, 1999. Elena Castro M, Diaz A, del Olmo E, and Pazos A. Chronic fluoxetine induces opposite changes in G protein coupling at pre and postsynaptic 5-HT1A receptors in rat brain. Neuropharmacology 44: 93-101, 2003. Fabre V, Beaufour C, Evrard A, Rioux A, Hanoun N, Lesch KP, Murphy DL, Lanfumey L, Hamon M, and Martres MP. Altered expression and functions of serotonin 5-HT1A and 5-HT1B receptors in knock-out mice lacking the 5-HT transporter. Eur J Neurosci 12: 2299-2310, 2000. Fairchild G, Leitch MM, and Ingram CD. Acute and chronic effects of corticosterone on 5-HT1A receptor-mediated autoinhibition in the rat dorsal raphe nucleus. Neuropharmacology 45: 925-934, 2003. Ferrer A, and Artigas F. Effects of single and chronic treatment with tranylcypromine on extracellular serotonin in rat brain. Eur J Pharmacol 263: 227-234, 1994. Gelfin Y, Gorfine M, and Lerer B. Effect of clinical doses of fluoxetine on psychological variables in healthy volunteers. Am J Psychiatry 155: 290-292, 1998. Gobbi G, Murphy DL, Lesch K, and Blier P. Modifications of the serotonergic system in mice lacking serotonin transporters: an in vivo electrophysiological study. JPharmacol Exp Ther 296: 987-995, 2001. Hensler JG. Differential regulation of 5-HT1A receptor-G protein interactions in brain following chronic antidepressant administration. Neuropsychopharmacology 26:565-573, 2002. Heurteaux C, Lucas G, Guy N, El Yacoubi M, Thummler S, Peng XD, Noble F, Blondeau N, Widmann C, Borsotto M, Gobbi G, Vaugeois JM, Debonnel G, and Lazdunski M. Deletion of the background potassium channel TREK-1 results in a depression-resistant phenotype. Nat Neurosci 9: 1134-1141, 2006. Holmes A, Lit Q, Murphy DL, Gold E, and Crawley JN. Abnormal anxiety-related behavior in serotonin transporter null mutant mice: the influence of genetic background. Genes Brain Behav 2: 365-380, 2003. Innis RB, and Aghajanian GK. Pertussis toxin blocks 5-HT1A and GABAB receptor-mediated inhibition of serotonergic neurons. Eur J Pharmacol 143: 195-204, 1987. Innis RB, Nestler EJ, and Aghajanian GK. Evidence for G protein mediation of serotonin- and GABAB-induced hyperpolarization of rat dorsal raphe neurons. Brain Res 459: 27-36, 1988. Johnson DA, Gartside SE, and Ingram CD. 5-HT1A receptor-mediated autoinhibition does not function at physiological firing rates: evidence from in vitro electrophysiological studies in the rat dorsal raphe nucleus. Neuropharmacology 43:959-965, 2002. Katayama J, Yakushiji T, and Akaike N. Characterization of the K+ current mediated by 5-HT1A receptor in the acutely dissociated rat dorsal raphe neurons. Brain Res 745: 283-292, 1997. Le Poul E, Laaris N, Doucet E, Laporte AM, Hamon M, and Lanfumey L. Early desensitization of somato-dendritic 5-HT1A autoreceptors in rats treated with fluoxetine or paroxetine. Naunyn Schmiedebergs Arch Pharmacol 352: 141-148, 1995. Lei Q, Jones MB, Talley EM, Garrison JC, and Bayliss DA. Molecular mechanisms mediating inhibition of G protein-coupled inwardly-rectifying K+ channels. Mol Cells 15: 1-9, 2003.
Page 24 of 39

25
Lemonde S, Turecki G, Bakish D, Du L, Hrdina PD, Bown CD, Sequeira A, Kushwaha N, Morris SJ, Basak A, Ou XM, and Albert PR. Impaired repression at a 5-hydroxytryptamine 1A receptor gene polymorphism associated with major depression and suicide. J Neurosci 23: 8788-8799, 2003. Li Q, Wichems C, Heils A, Lesch KP, and Murphy DL. Reduction in the density and expression, but not G-protein coupling, of serotonin receptors (5-HT1A) in 5-HT transporter knock-out mice: gender and brain region differences. J Neurosci 20: 7888-7895, 2000. Li YQ, Li H, Kaneko T, and Mizuno N. Morphological features and electrophysiological properties of serotonergic and non-serotonergic projection neurons in the dorsal raphe nucleus. An intracellular recording and labeling study in rat brain slices. Brain Res 900: 110-118, 2001. Liu R, Jolas T, and Aghajanian G. Serotonin 5-HT(2) receptors activate local GABA inhibitory inputs to serotonergic neurons of the dorsal raphe nucleus. Brain Res 873: 34-45, 2000. Liu RJ, Lambe EK, and Aghajanian GK. Somatodendritic autoreceptor regulation of serotonergic neurons: dependence on L-tryptophan and tryptophan hydroxylase-activating kinases. Eur J Neurosci 21: 945-958, 2005. Lowry CA, Rodda JE, Lightman SL, and Ingram CD. Corticotropin-releasing factor increases in vitro firing rates of serotonergic neurons in the rat dorsal raphe nucleus: evidence for activation of a topographically organized mesolimbocortical serotonergic system. J Neurosci 20: 7728-7736, 2000. Mannoury la Cour C, Boni C, Hanoun N, Lesch KP, Hamon M, and Lanfumey L. Functional consequences of 5-HT transporter gene disruption on 5-HT(1a) receptor-mediated regulation of dorsal raphe and hippocampal cell activity. JNeurosci 21: 2178-2185, 2001. Mannoury la Cour C, Hanoun N, Melfort M, Hen R, Lesch KP, Hamon M, and Lanfumey L. GABA(B) receptors in 5-HT transporter- and 5-HT1A receptor-knock-out mice: further evidence of a transduction pathway shared with 5-HT1A receptors. JNeurochem 89: 886-896, 2004. Mao J, Wang X, Chen F, Wang R, Rojas A, Shi Y, Piao H, and Jiang C.Molecular basis for the inhibition of G protein-coupled inward rectifier K(+) channels by protein kinase C. Proc Natl Acad Sci U S A 101: 1087-1092, 2004. Meltzer CC, Price JC, Mathis CA, Butters MA, Ziolko SK, Moses-Kolko E, Mazumdar S, Mulsant BH, Houck PR, Lopresti BJ, Weissfeld LA, and Reynolds CF. Serotonin 1A receptor binding and treatment response in late-life depression. Neuropsychopharmacology 29: 2258-2265, 2004. Mombereau C, Kaupmann K, Froestl W, Sansig G, van der Putten H, and Cryan JF. Genetic and pharmacological evidence of a role for GABA(B) receptors in the modulation of anxiety- and antidepressant-like behavior. Neuropsychopharmacology 29: 1050-1062, 2004a. Mombereau C, Kaupmann K, Gassmann M, Bettler B, van der Putten H, and Cryan JF. Altered anxiety and depression-related behaviour in mice lacking GABAB(2) receptor subunits. Neuroreport 16: 307-310, 2005. Mombereau C, Kaupmann K, van der Putten H, and Cryan JF. Altered response to benzodiazepine anxiolytics in mice lacking GABA B(1) receptors. Eur J Pharmacol 497: 119-120, 2004b. Monteleone P, Maj M, Iovino M, and Steardo L. GABA, depression and the mechanism of action of antidepressant drugs: a neuroendocrine approach. J Affect Disord 20: 1-5, 1990.
Page 25 of 39

26
Ou XM, Lemonde S, Jafar-Nejad H, Bown CD, Goto A, Rogaeva A, and Albert PR. Freud-1: A neuronal calcium-regulated repressor of the 5-HT1A receptor gene. JNeurosci 23: 7415-7425, 2003. Parsey RV, Oquendo MA, Ogden RT, Olvet DM, Simpson N, Huang YY, Van Heertum RL, Arango V, and Mann JJ. Altered serotonin 1A binding in major depression: a [carbonyl-C-11]WAY100635 positron emission tomography study. Biol Psychiatry 59: 106-113, 2006. Pejchal T, Foley MA, Kosofsky BE, and Waeber C. Chronic fluoxetine treatment selectively uncouples raphe 5-HT(1A) receptors as measured by [(35)S]-GTP gamma S autoradiography. Br J Pharmacol 135: 1115-1122, 2002. Penington NJ, Kelly JS, and Fox AP. Unitary properties of potassium channels activated by 5-HT in acutely isolated rat dorsal raphe neurones. J Physiol 469: 407-426, 1993a. Penington NJ, Kelly JS, and Fox AP. Whole-cell recordings of inwardly rectifying K+ currents activated by 5-HT1A receptors on dorsal raphe neurones of the adult rat. J Physiol 469: 387-405, 1993b. Petty F. GABA and mood disorders: a brief review and hypothesis. J Affect Disord 34: 275-281, 1995. Rygula R, Abumaria N, Domenici E, Hiemke C, and Fuchs E. Effects of fluoxetine on behavioral deficits evoked by chronic social stress in rats. Behav Brain Res 2006.Rygula R, Abumaria N, Flugge G, Fuchs E, Ruther E, and Havemann-Reinecke U. Anhedonia and motivational deficits in rats: impact of chronic social stress. Behav Brain Res 162: 127-134, 2005. Shen C, Li H, and Meller E. Repeated treatment with antidepressants differentially alters 5-HT1A agonist-stimulated [35S]GTP gamma S binding in rat brain regions. Neuropharmacology 42: 1031-1038, 2002. Shively CA, Friedman DP, Gage HD, Bounds MC, Brown-Proctor C, Blair JB, Henderson JA, Smith MA, and Buchheimer N. Behavioral depression and positron emission tomography-determined serotonin 1A receptor binding potential in cynomolgus monkeys. Arch Gen Psychiatry 63: 396-403, 2006. Slattery DA, Desrayaud S, and Cryan JF. GABAB receptor antagonist-mediated antidepressant-like behavior is serotonin-dependent. J Pharmacol Exp Ther 312: 290-296, 2005. Stockmeier CA, Shapiro LA, Dilley GE, Kolli TN, Friedman L, and Rajkowska G. Increase in serotonin-1A autoreceptors in the midbrain of suicide victims with major depression-postmortem evidence for decreased serotonin activity. J Neurosci 18: 7394-7401, 1998. Subhash MN, Srinivas BN, Vinod KY, and Jagadeesh S. Modulation of 5-HT1A receptor mediated response by fluoxetine in rat brain. J Neural Transm 107: 377-387, 2000. Takahashi T, Kobayashi T, Ozaki M, Takamatsu Y, Ogai Y, Ohta M, Yamamoto H, and Ikeda K. G protein-activated inwardly rectifying K+ channel inhibition and rescue of weaver mouse motor functions by antidepressants. Neurosci Res 54: 104-111, 2006. Von Frijtag JC, Kamal A, Reijmers LG, Schrama LH, van den Bos R, and Spruijt BM. Chronic imipramine treatment partially reverses the long-term changes of hippocampal synaptic plasticity in socially stressed rats. Neurosci Lett 309: 153-156, 2001.
Page 26 of 39

27
Von Frijtag JC, Reijmers LG, Van der Harst JE, Leus IE, Van den Bos R, and Spruijt BM. Defeat followed by individual housing results in long-term impaired reward- and cognition-related behaviours in rats. Behav Brain Res 117: 137-146, 2000. Von Frijtag JC, Van den Bos R, and Spruijt BM. Imipramine restores the long-term impairment of appetitive behavior in socially stressed rats. Psychopharmacology (Berl) 162: 232-238, 2002. Williams JT, Colmers WF, and Pan ZZ. Voltage- and ligand-activated inwardly rectifying currents in dorsal raphe neurons in vitro. J Neurosci 8: 3499-3506, 1988.
Page 27 of 39

28
Figure 1 . Morphological and biophysical identification of 5-HT neurons. A: 2-photon
laser scanning microscopy of a serotonergic cell. Serotonergic cells displayed a
current clamp profile shown in B. C: Currents in response to a voltage clamp protocol
(-60 mV to -120 mV) before, during, and after application of 5-HT (5 µM). Membrane
potential changes after wash-in and wash-out of 5-HT were monitored in current
clamp. D: I-V plots of the currents as induced in C. The conductance was calculated
from the slope of the linear part of the I-V. E: Dose response curve of GIRK
conductance induction by 5-HT (EC 50 = 2.66 µM).
Figure 2 . Chronic application of fluoxetine affects both 5-HT and GABAB mediated
GIRK responses. A-D: Example traces of 5-HT- and BAC induced currents in control
animals not treated (water) (A, C) and treated with fluoxetine (B, D). E: 5-HT-
activated GIRK conductances in control animals not treated (5µM 5-HT 2.3±0.3,
n=19; wash 0.2±0.1, n=19; 100µM 5-HT 4.9±0.5, n=15; wash 0.8±0.4, n=11) and
treated with fluoxetine (5µM 5-HT 2.6±0.2, n=42; wash 0.4±0.1, n=35; 100µM 5-HT
3.3±0.4, n=30; wash 1.3±0.2, n=24). F: BAC-affected GIRK conductances in control
animals not treated (wash 0.6±0.3, n=7; 1mM BAC 5.1±0.7, n=7) and treated with
fluoxetine (wash 0.8±0.2, n=16; 1mM BAC 2.2±0.3, n=16). All induced effects, with
the exception of the BAC effect in unconditioned animals treated with fluoxetine,
were significantly different from their controls (p<0.05). The 100 µM 5-HT effect was
significantly different from the 5 µM 5-HT (p<0.01) in the control animals-not
fluoxetine treated group but not in the fluxetine-treated group. The BAC effect in the
fluoxetine treated group was significantly different from 100 µM 5-HT effect (p<0.05).
Figure 3 . Social stress does not affect 5-HT induced GIRK responses. A-D: GIRK
currents in response to 5-HT and BAC in control (A, C) and socially stressed animals
(B, D). E: 5-HT-activated GIRK conductances in control (5µM 5-HT 3.6±0.5, n=13;
Page 28 of 39

29
wash 0,3±0.1, n=12; 100µM 5-HT 6.3±0.8, n=10; wash 0.6±0.2, n=9) and socially
stressed animals (5µM 5-HT 3.3±0.3, n=19; wash 1,1±0.4, n=19; 100µM 5-HT
6.0±0.8, n=11; wash 2.0±0.5, n=10). F: BAC-activated GIRK conductances in control
(wash 0.3±0.1, n=9; 1mM BAC 2.9±0.6, n=9) and socially stressed animals (wash
1.5±0.6, n=10; 1mM BAC 4.6±1.2, n=10). All induced effects were significantly
different from their control (p<0.05) but not from the induced effects in the other
experimental group. The 100 µM 5-HT effect was significantly different from the 5 µM
5-HT effect (*=p<0.05, **=p<0.01) but not from the 1 mM BAC effect, within each
experimental group.
Figure 4. Chronic application of fluoxetine to socially stressed animals leads to
reduction of both 5-HT and GABAB mediated GIRK responses. A-D: Example traces
of 5-HT- and BAC induced currents in socially stressed animals not treated (A, C) or
treated with fluoxetine (B, D). E: 5-HT-activated GIRK conductances in socially
stressed animals not treated (5µM 5-HT 4.7±0.6, n=19; wash 0,3±0.2, n=19; 100µM
5-HT 6.3±0.7, n=11; wash 0.9±0.3, n=10) and treated with fluoxetine (5µM 5-HT
2.7±0.2, n=31; wash 0.3±0.1, n=31; 100µM 5-HT 2.7±0.5, n=15; wash 1.4±0.5, n=8).
F: BAC-activated GIRK conductances in socially stressed animals treated not treated
(wash 0.1±0.1, n=4; 1mM BAC 4.1±0.3, n=4) and treated with fluoxetine (wash
0.2±0.1, n=14; 1mM BAC 0.6±0.2, n=13). All induced effects, with the exception of
the BAC effect in socially stressed animals treated with fluoxetine, were significantly
different from their control (p<0.05) and from the induced effects in the other
experimental group (**<0.01, ***<0.005). The 100 µM 5-HT effect was significantly
different from the 5 µM 5-HT effect (p<0.05) in the social stress-not treated with
fluoxetine- group, but not in the social stress -treated with fluoxetine- group. The BAC
effect is significantly different from 100 µM 5-HT effect (p<0.05) in both groups.
Page 29 of 39

30
Figure 5 . No interaction between effect of chronic fluoxetine treatment and social
defeat conditioning. A-D: Example traces of 5-HT- and BAC induced currents in
control animals (A, C) and socially stressed animals (B, D), both treated with
fluoxetine (10mg/kg) for 21 days. E: 5-HT-activated GIRK conductances in control
animals treated with fluoxetine (5µM 5-HT 2.4±0.3, n=22; wash 0.5±0.1, n=15;
100µM 5-HT 2.5±0.4, n=13; wash 1.1±0.4, n=9) and socially stressed animals treated
with fluoxetine (5µM 5-HT 2.1±0.2, n=19; wash 0,5±0.1, n=18; 100µM 5-HT 2.1±0.2,
n=12; wash 1.2±0.2, n=7) . F: BAC-activated GIRK conductances in control animals
treated with fluoxetine (wash 0.3±0.4, n=5; 1mM BAC 1.4±0.4, n=5) and socially
stressed animals treated with fluoxetine (wash 0.4±0.2, n=5; 1mM BAC 1.5±0.4,
n=5). All 5-HT induced effects were significantly different from their control (p<0.05)
but not from the induced effects in the other experimental group. The BAC effect was
not significantly different from its control or from the induced effect in the other group.
Tabel I. Relief from anhedonia-symptoms upon chronic fluoxetine treatment in
socially stressed animals. Mean number of transition between behavioural elements
(± SEM), displayed in the interval (10 min) between the announcement (light+sound)
and the arrival of the reward (sucrose 5%, 5 min), reflecting baseline activity at trial 0,
and anticipatory activity (post-training) in expectation of the reward at trial 32.
Page 30 of 39
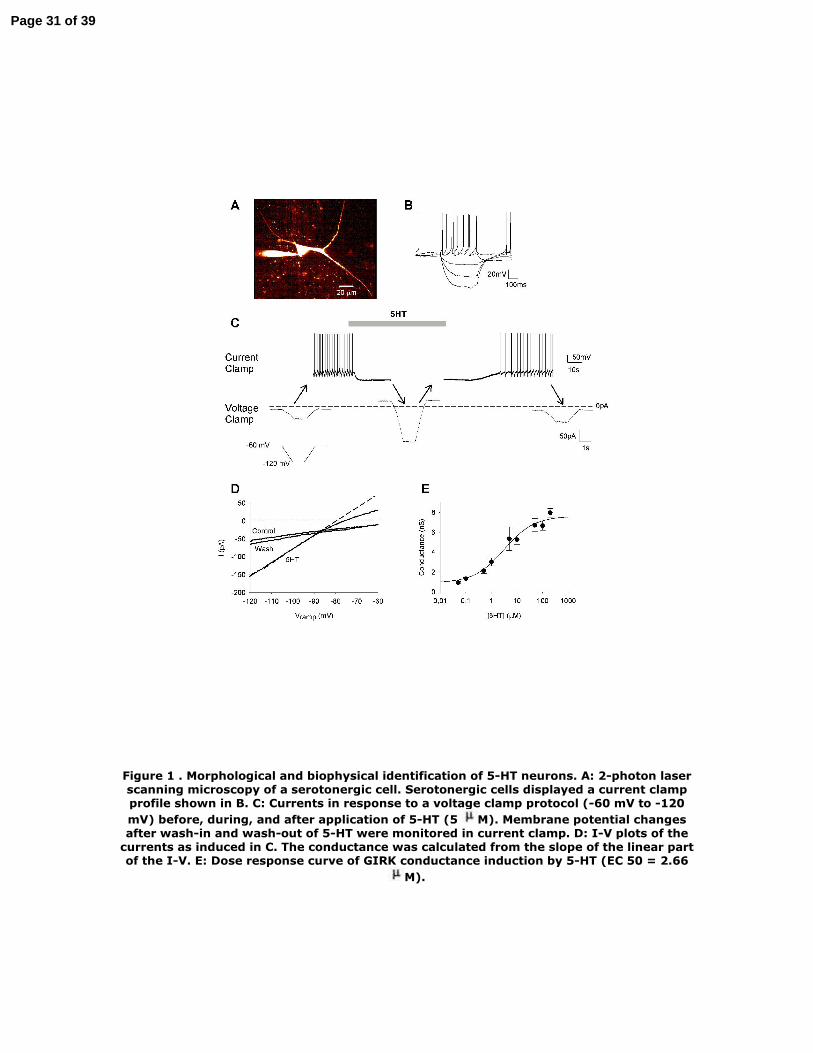
Figure 1 . Morphological and biophysical identification of 5-HT neurons. A: 2-photon laser scanning microscopy of a serotonergic cell. Serotonergic cells displayed a current clamp profile shown in B. C: Currents in response to a voltage clamp protocol (-60 mV to -120 mV) before, during, and after application of 5-HT (5 M). Membrane potential changes after wash-in and wash-out of 5-HT were monitored in current clamp. D: I-V plots of the
currents as induced in C. The conductance was calculated from the slope of the linear part of the I-V. E: Dose response curve of GIRK conductance induction by 5-HT (EC 50 = 2.66
M).
Page 31 of 39
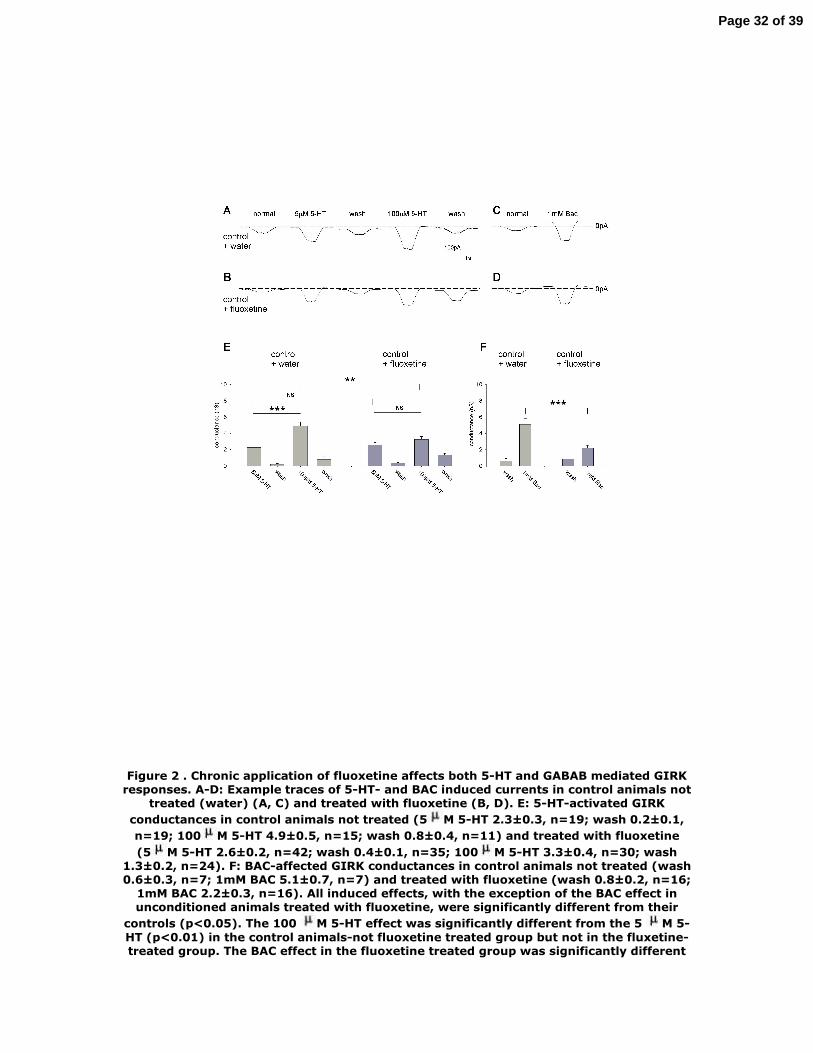
Figure 2 . Chronic application of fluoxetine affects both 5-HT and GABAB mediated GIRK responses. A-D: Example traces of 5-HT- and BAC induced currents in control animals not
treated (water) (A, C) and treated with fluoxetine (B, D). E: 5-HT-activated GIRK conductances in control animals not treated (5 M 5-HT 2.3±0.3, n=19; wash 0.2±0.1, n=19; 100 M 5-HT 4.9±0.5, n=15; wash 0.8±0.4, n=11) and treated with fluoxetine (5 M 5-HT 2.6±0.2, n=42; wash 0.4±0.1, n=35; 100 M 5-HT 3.3±0.4, n=30; wash
1.3±0.2, n=24). F: BAC-affected GIRK conductances in control animals not treated (wash 0.6±0.3, n=7; 1mM BAC 5.1±0.7, n=7) and treated with fluoxetine (wash 0.8±0.2, n=16;
1mM BAC 2.2±0.3, n=16). All induced effects, with the exception of the BAC effect in unconditioned animals treated with fluoxetine, were significantly different from their
controls (p<0.05). The 100 M 5-HT effect was significantly different from the 5 M 5-HT (p<0.01) in the control animals-not fluoxetine treated group but not in the fluxetine-treated group. The BAC effect in the fluoxetine treated group was significantly different
Page 32 of 39

from 100 M 5-HT effect (p<0.05).
Page 33 of 39

Figure 3 . Social stress does not affect 5-HT induced GIRK responses. A-D: GIRK currents in response to 5-HT and BAC in control (A, C) and socially stressed animals (B, D). E: 5-HT-activated GIRK conductances in control (5 M 5-HT 3.6±0.5, n=13; wash 0,3±0.1, n=12; 100 M 5-HT 6.3±0.8, n=10; wash 0.6±0.2, n=9) and socially stressed animals (5 M 5-HT 3.3±0.3, n=19; wash 1,1±0.4, n=19; 100 M 5-HT 6.0±0.8, n=11; wash
2.0±0.5, n=10). F: BAC-activated GIRK conductances in control (wash 0.3±0.1, n=9; 1mM BAC 2.9±0.6, n=9) and socially stressed animals (wash 1.5±0.6, n=10; 1mM BAC 4.6±1.2, n=10). All induced effects were significantly different from their control
(p<0.05) but not from the induced effects in the other experimental group. The 100 M5-HT effect was significantly different from the 5 M 5-HT effect (*=p<0.05,
**=p<0.01) but not from the 1 mM BAC effect, within each experimental group.
Page 34 of 39
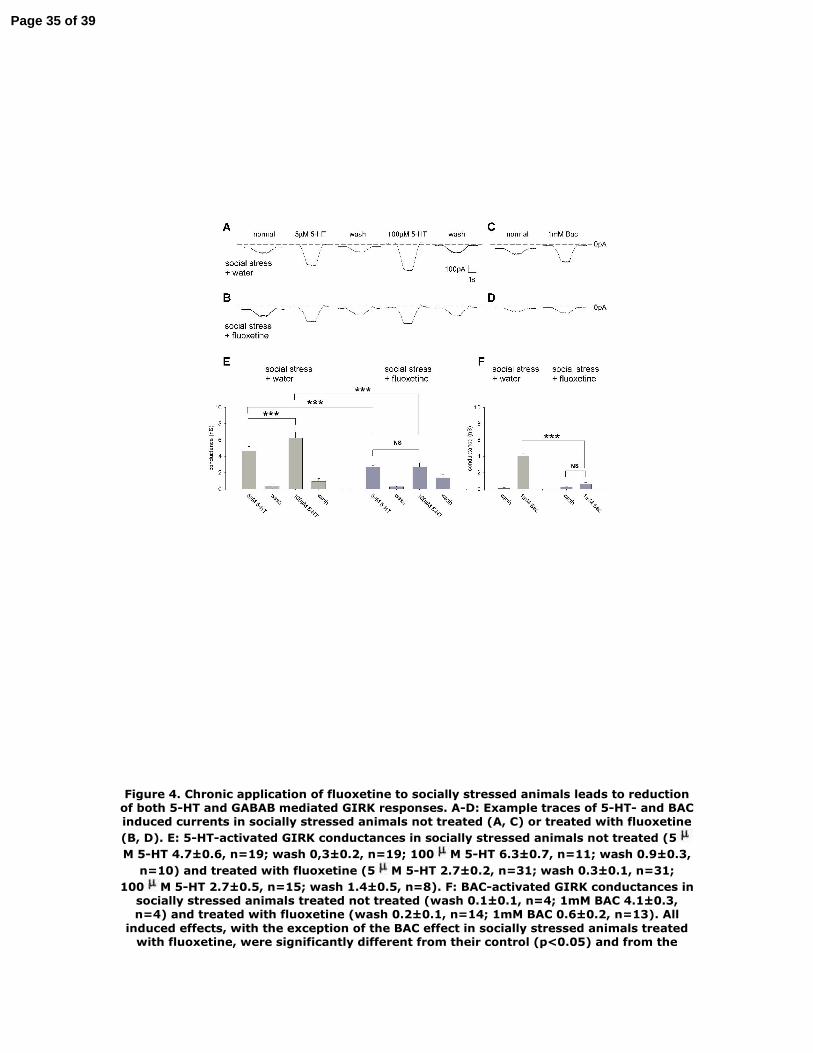
Figure 4. Chronic application of fluoxetine to socially stressed animals leads to reduction of both 5-HT and GABAB mediated GIRK responses. A-D: Example traces of 5-HT- and BAC induced currents in socially stressed animals not treated (A, C) or treated with fluoxetine (B, D). E: 5-HT-activated GIRK conductances in socially stressed animals not treated (5M 5-HT 4.7±0.6, n=19; wash 0,3±0.2, n=19; 100 M 5-HT 6.3±0.7, n=11; wash 0.9±0.3,
n=10) and treated with fluoxetine (5 M 5-HT 2.7±0.2, n=31; wash 0.3±0.1, n=31; 100 M 5-HT 2.7±0.5, n=15; wash 1.4±0.5, n=8). F: BAC-activated GIRK conductances in
socially stressed animals treated not treated (wash 0.1±0.1, n=4; 1mM BAC 4.1±0.3, n=4) and treated with fluoxetine (wash 0.2±0.1, n=14; 1mM BAC 0.6±0.2, n=13). All
induced effects, with the exception of the BAC effect in socially stressed animals treated with fluoxetine, were significantly different from their control (p<0.05) and from the
Page 35 of 39

induced effects in the other experimental group (**<0.01, ***<0.005). The 100 M 5-HT effect was significantly different from the 5 M 5-HT effect (p<0.05) in the social
stress-not treated with fluoxetine- group, but not in the social stress -treated with fluoxetine- group. The BAC effect is significantly different from 100 M 5-HT effect
(p<0.05) in both groups
Page 36 of 39

Figure 5 . No interaction between effect of chronic fluoxetine treatment and social defeat conditioning. A-D: Example traces of 5-HT- and BAC induced currents in control animals
(A, C) and socially stressed animals (B, D), both treated with fluoxetine (10mg/kg) for 21 days. E: 5-HT-activated GIRK conductances in control animals treated with fluoxetine (5 M 5-HT 2.4±0.3, n=22; wash 0.5±0.1, n=15; 100 M 5-HT 2.5±0.4, n=13; wash
1.1±0.4, n=9) and socially stressed animals treated with fluoxetine (5 M 5-HT 2.1±0.2, n=19; wash 0,5±0.1, n=18; 100 M 5-HT 2.1±0.2, n=12; wash 1.2±0.2, n=7) . F: BAC-activated GIRK conductances in control animals treated with fluoxetine (wash 0.3±0.4,
n=5; 1mM BAC 1.4±0.4, n=5) and socially stressed animals treated with fluoxetine (wash 0.4±0.2, n=5; 1mM BAC 1.5±0.4, n=5). All 5-HT induced effects were significantly different from their control (p<0.05) but not from the induced effects in the other
Page 37 of 39

experimental group. The BAC effect was not significantly different from its control or from the induced effect in the other group
Page 38 of 39

Baseline Post-training p-value
social defeat 16.9 ±1.09 16.0 ±0.82 0.4 (n = 9)
social defeat+ fluoxetine
17.0 ±1.01 21.1 ±0.92 0.02 (n = 10)
Table I
Page 39 of 39




















