
Geography and ontogeny influence the stable oxygen and carbon isotopes of otoliths of Pacific...
10
Fisheries Research 154 (2014) 1–10 Contents lists available at ScienceDirect Fisheries Research j ourna l ho me page: www.elsevier.com/locate/fishres Geography and ontogeny influence the stable oxygen and carbon isotopes of otoliths of Pacific sardine in the California Current Barbara Javor a,∗ , Emmanis Dorval b a Ocean Associates, Inc., 4007 Abingdon Street, Arlington, VA 22207, USA 1 b Southwest Fisheries Science Center, National Marine Fisheries Service, 8901 La Jolla Shores Drive, La Jolla, CA 92037, USA a r t i c l e i n f o Article history: Received 5 November 2013 Received in revised form 21 January 2014 Accepted 26 January 2014 Keywords: Otolith Stable oxygen isotope Sardine Sardinops sagax a b s t r a c t Pacific sardine (Sardinops sagax), a commercially valuable species, have a broad distribution along the North American coast that spans Canada, the United States, and Mexico. The goal of this research was to evaluate water temperature history of the fish inferred from stable oxygen isotopes in otoliths in order to differentiate and connect stocks across regions, and between juveniles and adults. Local seawater composition in the Pacific Northwest affected major north–south trends in 18 O composition of juvenile otoliths. Inferred temperature correlated inversely with the age (otolith weight) of juveniles within a region, possibly resulting from changes in depth preferences as the fish grew. The correlations between 13 C and 18 O in juvenile otoliths were relatively weak in the northernmost and southernmost samples, but comparisons of the sample means indicated significant differences between some regions. Otoliths from adult sardine captured between California and Canada recorded 18 O values reflecting cooler tem- peratures than otoliths from juveniles, and without regional differentiation. These results are consistent with a northern stock that mixes during annual migrations. © 2014 Elsevier B.V. All rights reserved. 1. Introduction The distribution of Pacific sardine (Sardinops sagax) in North America, spanning approximately 5000 km of coastline that includes Canada, the United States (US), and Mexico, poses a chal- lenge for fishery management (Fig. 1; southeast Alaska not shown). After the commercial sardine fishery collapsed during the 1940s and 1950s due to overfishing and/or decadal climatic shifts in the California Current large marine ecosystem (Norton and Mason, 2005), the population rebounded sufficiently by the 1980s to sup- port managed commercial fishing again in the three countries. In spite of the broad geographic range and the commercial impor- tance of Pacific sardine, knowledge of their life history, including depth distribution, spawning activity, juvenile recruitment, and migratory habits, is incomplete. Three North American stocks are generally assumed: (1) a cold, northern stock along the California coast that migrates as far as southern Alaska in the summer as adults; (2) a temperate, southern stock along most of the Pacific coast of Baja California, Mexico, that migrates seasonally into south- ern California; and (3) a warm, Gulf of California stock that migrates out of the Gulf as far as Bahía Magdalena on the Pacific coast of Baja ∗ Corresponding author. Tel.: +1 858 546 5679. E-mail addresses: [email protected], [email protected] (B. Javor), [email protected] (E. Dorval). 1 Present address. California (Wing et al., 2000; Félix-Uraga et al., 2004, 2005; Smith, 2005). The basis for dividing Pacific sardine into stocks is not well defined. Little genetic structure has been found in their present dis- tribution (Hedgecock et al., 1989; Grant and Bowen, 1998; Pereyra et al., 2004; García-Rodríguez et al., 2011). Other methods have had some success in identifying regional stocks and movements of sardine. They include egg, larval, and adult surveys; tempera- ture at catch; vertebral counts; tags; fish and otolith morphometric analysis; and otolith chemistry, including trace elements and stable isotopes (Félix-Uraga et al., 2004, 2005; Smith, 2005; Lo et al., 2005, 2010, 2011; Valle and Herzka, 2008; Dorval et al., 2011; Javor et al., 2011; Javor, 2013). The extent of drift and migration among young juveniles that are not large enough to be commercially captured remains unknown. Temperature history of sardine provides a rationale to divide them into stocks. It is reflected in region and season of spawn- ing, and is recorded in stable isotopes of oxygen in otoliths. Pacific sardine spawn primarily in the spring in central and southern California waters (Lo et al., 2005), although spawning adults are caught in the Pacific Northwest (off Oregon and Washington) in summer months where juveniles are also periodically captured (Emmett et al., 2005; Lo et al., 2010). They spawn in the summer in Bahía Magdalena, and in the late fall and winter in the Gulf of California (Smith, 2005). The temperature range of Pacific sardine is broad, from less than 10 ◦ C off Oregon and Washington (Emmett et al., 2005) to over 25 ◦ C in their southern distribution in Mexico 0165-7836/$ – see front matter © 2014 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.fishres.2014.01.016
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Geography and ontogeny influence the stable oxygen and carbon isotopes of otoliths of Pacific...

Gi
Ba
b
a
ARRA
KOSSS
1
AilAaC2pstdmgcaceo
e
0h
Fisheries Research 154 (2014) 1–10
Contents lists available at ScienceDirect
Fisheries Research
j ourna l ho me page: www.elsev ier .com/ locate / f i shres
eography and ontogeny influence the stable oxygen and carbonsotopes of otoliths of Pacific sardine in the California Current
arbara Javora,∗, Emmanis Dorvalb
Ocean Associates, Inc., 4007 Abingdon Street, Arlington, VA 22207, USA1
Southwest Fisheries Science Center, National Marine Fisheries Service, 8901 La Jolla Shores Drive, La Jolla, CA 92037, USA
r t i c l e i n f o
rticle history:eceived 5 November 2013eceived in revised form 21 January 2014ccepted 26 January 2014
eywords:tolith
a b s t r a c t
Pacific sardine (Sardinops sagax), a commercially valuable species, have a broad distribution along theNorth American coast that spans Canada, the United States, and Mexico. The goal of this research was toevaluate water temperature history of the fish inferred from stable oxygen isotopes in otoliths in orderto differentiate and connect stocks across regions, and between juveniles and adults. Local seawatercomposition in the Pacific Northwest affected major north–south trends in �18O composition of juvenileotoliths. Inferred temperature correlated inversely with the age (otolith weight) of juveniles within a
table oxygen isotopeardineardinops sagax
region, possibly resulting from changes in depth preferences as the fish grew. The correlations between�13C and �18O in juvenile otoliths were relatively weak in the northernmost and southernmost samples,but comparisons of the sample means indicated significant differences between some regions. Otolithsfrom adult sardine captured between California and Canada recorded �18O values reflecting cooler tem-peratures than otoliths from juveniles, and without regional differentiation. These results are consistent
t mix
with a northern stock tha. Introduction
The distribution of Pacific sardine (Sardinops sagax) in Northmerica, spanning approximately 5000 km of coastline that
ncludes Canada, the United States (US), and Mexico, poses a chal-enge for fishery management (Fig. 1; southeast Alaska not shown).fter the commercial sardine fishery collapsed during the 1940snd 1950s due to overfishing and/or decadal climatic shifts in thealifornia Current large marine ecosystem (Norton and Mason,005), the population rebounded sufficiently by the 1980s to sup-ort managed commercial fishing again in the three countries. Inpite of the broad geographic range and the commercial impor-ance of Pacific sardine, knowledge of their life history, includingepth distribution, spawning activity, juvenile recruitment, andigratory habits, is incomplete. Three North American stocks are
enerally assumed: (1) a cold, northern stock along the Californiaoast that migrates as far as southern Alaska in the summer asdults; (2) a temperate, southern stock along most of the Pacific
oast of Baja California, Mexico, that migrates seasonally into south-rn California; and (3) a warm, Gulf of California stock that migratesut of the Gulf as far as Bahía Magdalena on the Pacific coast of Baja∗ Corresponding author. Tel.: +1 858 546 5679.E-mail addresses: [email protected], [email protected] (B. Javor),
[email protected] (E. Dorval).1 Present address.
165-7836/$ – see front matter © 2014 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.fishres.2014.01.016
es during annual migrations.© 2014 Elsevier B.V. All rights reserved.
California (Wing et al., 2000; Félix-Uraga et al., 2004, 2005; Smith,2005).
The basis for dividing Pacific sardine into stocks is not welldefined. Little genetic structure has been found in their present dis-tribution (Hedgecock et al., 1989; Grant and Bowen, 1998; Pereyraet al., 2004; García-Rodríguez et al., 2011). Other methods havehad some success in identifying regional stocks and movementsof sardine. They include egg, larval, and adult surveys; tempera-ture at catch; vertebral counts; tags; fish and otolith morphometricanalysis; and otolith chemistry, including trace elements and stableisotopes (Félix-Uraga et al., 2004, 2005; Smith, 2005; Lo et al., 2005,2010, 2011; Valle and Herzka, 2008; Dorval et al., 2011; Javor et al.,2011; Javor, 2013). The extent of drift and migration among youngjuveniles that are not large enough to be commercially capturedremains unknown.
Temperature history of sardine provides a rationale to dividethem into stocks. It is reflected in region and season of spawn-ing, and is recorded in stable isotopes of oxygen in otoliths. Pacificsardine spawn primarily in the spring in central and southernCalifornia waters (Lo et al., 2005), although spawning adults arecaught in the Pacific Northwest (off Oregon and Washington) insummer months where juveniles are also periodically captured(Emmett et al., 2005; Lo et al., 2010). They spawn in the summer
in Bahía Magdalena, and in the late fall and winter in the Gulf ofCalifornia (Smith, 2005). The temperature range of Pacific sardineis broad, from less than 10 ◦C off Oregon and Washington (Emmettet al., 2005) to over 25 ◦C in their southern distribution in Mexico
2 B. Javor, E. Dorval / Fisheries Re
(jt(ts
n(1crtvtcmDu12dSB2
t�a2pcHg2
Fig. 1. Map of the North American collection sites of S. sagax samples.
Félix-Uraga et al., 2004, 2005). Small pelagic fish such as young,uvenile sardine lack the energetic requirements to migrate far fromheir natal region in the absence of currents that allow them to driftNøttestad et al., 1999). Therefore tracking the habitat tempera-ures of young sardine may be a useful key for delineating spawningtocks.
Fish deposit 18O in their calcium carbonate otoliths in orear thermodynamic isotopic equilibrium with their environmentKalish, 1991a; Iacumin et al., 1992; Thorrold et al., 1997; Campana,999). 13C deposition is partially under metabolic control, witharbon derived both from bicarbonate in seawater and CO2 fromespiration. Physicochemical factors such as bicarbonate concen-ration and temperature control the offsets from seawater �18Oalues recorded in otoliths in a predictable way. Under experimen-al conditions, the linear response of 18O deposition to temperaturean be calibrated for a fish species and used to calculate environ-ental temperatures in otoliths of wild-caught fish (Kalish, 1991b;orval et al., 2011). Stable oxygen isotope profiles have also beensed to distinguish Australian S. sagax (Edmonds and Fletcher,997) and other fish stocks (Ashford and Jones, 2007; Rooker et al.,008), and to identify changes in habitat temperature of fish atifferent life stages (Gao and Beamish, 2003; Gao et al., 2004;hephard et al., 2007) or seasons (Weidman and Millner, 2000;egg and Weidman, 2001; Gao et al., 2001a; Høie and Folkvord,006).
The global water cycle modulates the distribution of H218O in
he ocean because evaporation favors the removal of H216O. The
18O signature of seawater generally correlates to salinity (Craignd Gordon, 1965; Bigg and Rohling, 2000; LeGrande and Schmidt,006), but climate and geography affect the regional distributionatterns of stable isotopes in freshwater sources that mix with
oastal seawater. While some coastal regions drain freshwater with218O content close to that of seawater, others discharge waterreatly depleted in H2
18O (Kipphut, 1990; Kendall and Coplen,001).
search 154 (2014) 1–10
Freshwater influence is important in assessing stable isotopesin otoliths of a coastal species such as the Pacific sardine. Tempera-tures will be recorded in a predictable way, i.e., cold water promoteshigher �18O values in otoliths (Kalish, 1991a, 1991b; Thorrold et al.,1997; Høie et al., 2003, 2004; Dorval et al., 2011), but local �18O val-ues of seawater due to freshwater dilution might counteract suchpredicted thermal effects (Elsdon and Gillanders, 2002). Interpreta-tions of fish habitat may be even more complex if sardine undergodiurnal changes in depths (Dorval et al., 2011) or if they migratenear freshwater sources (Gao et al., 2001b).
The overall aims of this investigation were to use otolith sta-ble isotopes to differentiate or connect regional stocks of youngPacific sardine throughout their range in North America, and tocompare stable isotope signatures of whole otoliths from juvenilesand adults. The study evaluated stable isotopes in several ways: (1)variations in �18O values within individual fish; (2) regional sig-natures; (3) age effects on �18O composition in local populationswithin the first year of life; (4) growth rate effects on �18O values;and (5) correlations between �18O and �13C values.
This study was part of a broader research effort to evaluate sta-ble isotopes (Dorval et al., 2011), morphology (Javor et al., 2011;Javor, 2013), and trace element chemistry of otoliths (to be reportedelsewhere) as methods to understand stock characteristics and lifehistory of Pacific sardine, especially pre-adult fish.
2. Materials and methods
2.1. Field collections
The study analyzed sardine otoliths collected between Vancou-ver Island (British Columbia, Canada) and the Gulf of California(Mexico) at various dates between 1986 and 2008 (Fig. 1 andTable 1). The older samples collected by the California Depart-ment of Fish and Game were archived by collection date, vessel,and catch site, and stored dry in gelatin capsules. Otoliths wereaged by microscopic examination of daily or annual growth bands(Yaremko, 1996) in conjunction with otolith weight. Otolithscollected in monthly samples of sardine caught near San Diego typ-ically weighed 0.8–1.0 mg when they reached age-1 and 1.2–1.5 mgat the time they turned age-2. The sardine collected near San Diegoranged in standard length (SL) from 67 to 190 mm, and their otolithsranged in weight from 0.3 to 1.8 mg.
2.2. Growth experiment
The effect of growth rate on otolith stable isotope composi-tion was tested in the laboratory at Southwest Fisheries ScienceCenter (SWFSC). Young sardine were captured in a beach seineat the Scripps Institution of Oceanography beach in San Diego inDecember, 2004, and raised in a flow-through seawater systemat constant salinity (33 psu) and at ambient seawater temper-ature from December until the following July (14–17 ◦C). Theseawater was collected and filtered at the Scripps Institution ofOceanography pier. The initial fish were nearly uniform in size,averaging 67.4 ± 3.9 mm SL (±standard deviation [S.D.]), 2.6 ± 0.4 gfish weight, and 0.339 ± 0.027 mg otolith weight. Counting theincrements in the otoliths indicated they were born the previousMay, making them about 7 months of age at the start of the exper-iment. They were maintained on a 12-h L/12-h D light cycle in a
single, circular tank (1.83-m diameter, 1600 L) with approximatelyten volumes turnover per day. They were fed Bio-diet brood formu-lation pellets (Bio-Oregon), ad libitum. Fish were sampled at Time-0and after 8 months.
B. Javor, E. Dorval / Fisheries Research 154 (2014) 1–10 3
Table 1Pacific sardine (S. sagax) otolith collection used for stable isotope studies.
Area Year Collection Collectora
Canada 2003 Vancouver Island (adults) FAOCanada 2005 Vancouver Island (age 1) FAOOregon–Washington 2003 Cruise FR0307 (3/03), 4 trawls SWFSCOregon–Washington 2003 Cruise MF0313 (11/03), 6 trawls NWFSCOregon–Washington 2004 Cruise FR0403 (3/04), 3 trawls SWFSCHumboldt Bay 1996 Port samples, 3 dates CDFGMonterey 1996–7 Port samples, 8 dates CDFGSo. California Bight 1986–88 Port samples, juveniles CDFGSo. California Bight 1995–2003 Port samples, March-April, age 1 CDFGSan Diego 2003–09 Bait receiver, monthly samples SWFSCSan Diego 2004 Beach seine, juveniles SWFSCEnsenada 1991–92 Port samples, spring and fall CDFGBahía Magdalena 2004 Spring and fall, 4 dates CICM (R. Félix-Uraga)Gulf of California 2006 February and December CICESE (Y. Ríos)
a FAO (Fisheries and Oceans, Canada); SWFSC (Southwest Fisheries Science Center); NWFSC (Northwest Fisheries Science Center); CDFG (California Department of Fish andG Sur);B
2
mswOooogst(wt(
gabLcssBcasodm
oi(0mtdcUsvtn
ame); CICM (Centro Interdisciplinario de Ciencias Marinas, La Paz, Baja Californiaaja California).
.3. Stable isotope analysis
The survey utilized primarily right sagittal otoliths after deter-ining there was no significant difference between left and right
ides (discussed below). Many of the corresponding left otolithsere used for trace element analysis (to be reported elsewhere).toliths with vaterite were excluded. Because sardine capturedff southern California were represented in multiple collectionsver time (years), and by different ages and sizes (weights) oftoliths, samples were selected to test variations with time orrowth for that region. Over 500 otoliths were analyzed for thetudy. After initially cleaning in purified water (Milli-QTM water),he otoliths were dried, weighed on a Cahn C-33 microbalance0.005 mg accuracy), and stored in plastic Eppendorf tubes. Otolithsere further cleaned in a Class 100 clean room using acid-washed
ubes, probes, pipet tips, slides, etc., by the method of Dorval et al.2011).
Powdered samples (60–100 �g) were analyzed for stable oxy-en and carbon isotopic composition with a Micromass IsoPrimeutomated carbonate analyzer (GV Instruments, Manchester, UK)ased on the dual inlet measurement at the Isotope Geochemistryaboratory of the University of Maryland (UMd), or with a tripleollector gas ratio mass spectrometer (VG Prism) at the Univer-ity of Southern California (USC). Samples of otolith powder werepiked with concentrated phosphoric acid at 90 ◦C. Vienna Pee Deeelemnite (VPDB) reference standards (NBS-19 or Ultiss, which wasalibrated to NBS-19) were run before, between sample blocks,nd after each set analyzed on any one day. The average preci-ion was ±0.07‰ (S.D.) determined from replicate measurementsf standards. The raw data were corrected and reported as the ‰ifference from the standard Pee Dee Belemnite (PDB) referenceaterial using the customary � notation.To cross-check the results from the two laboratories, 11 pairs
f otoliths from two different collections were split to run onen each instrument using similar reference standards. The mean±standard error, S.E.) �18O values for the USC instrument were.297‰ ± 0.051‰ greater than those analyzed by the UMd instru-ent, and the mean �13C values were 0.223‰ ± 0.109‰ less than
he results from the UMd instrument. The discrepancies were likelyue to differences in reference gases (Miguel Rincon, USC, personalommunication). The corrections were applied to conform to theMd instrument results where data from both instruments are pre-
ented in the figures. Comparable differences in oxygen isotopealues of otolith material determined on the UMd instrument andwo other instruments (+0.25‰ and −0.24‰) have previously beenoted (Rooker et al., 2008).
CICESE (Centro de Investigación Científica y de Educación Superior de Ensenada,
The �18O values of coastal seawater in the eastern Pacific(2–57◦ N), estimated from the graphic data of Craig and Gordon(1965), were calculated to compare �18O signatures of surface sea-water and otoliths according to latitude. The study by Craig andGordon is the most comprehensive database to date for oxygenisotopes and salinities of the region that overlaps with the distri-bution of Pacific sardine. The relationship between seawater �18Oand latitude was:
�18Owater = (−0.024 × latitude) + 0.443 (R2 = 0.915, n = 37) (1)
Salinities for the latitudinal range were 32.55–34.88 psu. Salinitiesfor seawater collected at the Scripps Institution of Oceanographypier ranged from 33.48 to 33.83 psu, n = 16.
2.4. Data analysis
Temperature calculations from �18O values in otoliths werebased on the formula determined by Dorval et al. (2011) for sardinereared in San Diego seawater:
�18Ootolith(‰) − �18Owater(‰)
= (−0.132(±0.003S.E.) × Temperature ◦C)
+ 2.4455(±0.043S.E.), (2)
where S.E. is standard error equal to 1.Those authors used data from the UMd mass spectrome-
ter. Temperature and salinity experienced during the lifespanof immature fish in California waters were estimated usingCalifornia Cooperative Oceanic Fisheries Investigations (CalCOFI)data (www.calcofi.org, accessed 18 Jan, 2013) and compared totemperature values of otoliths calculated by the method of Dorvalet al. (2011). CalCOFI temperature and salinity values are recordedquarterly each year for a large grid of nearshore and offshorestations including stations near San Diego; the database also pro-vides 50-year averages for all the stations. Because little freshwaterenters the ocean off California, salinity fluctuations were assumedto be minimal in the Southern California Bight (Dorval et al., 2011).
Paired t-tests were used to determine whether left and rightotoliths had similar isotopic composition. Fourteen pairs of otolithswere randomly selected from several monthly collections dur-ing 2008. Normality assumptions were tested based on theShapiro–Wilk test. Homogeneity of variances was determined
based on the F-ratio test, by taking the ratio of the greater varianceover the lesser one (Sokal and Rohlf, 1992).If immature sardine spend more time at depth as they grow,their oxygen isotopic signatures would change to reflect the
4 B. Javor, E. Dorval / Fisheries Research 154 (2014) 1–10
-2.0
-1.5
-1.0
-0.5
0.0
0.5
1.0
50454035302520
δ18 O
, ‰
Latitude, ºN
Gulf
Bahía Magdalen a
Ensenada
San Diego
SoCal
Age-2 Montere y
Humboldt
PNW
Canada
Age 3-5
Age 1-2
Canada
A
Sea wate r δδ18O
Age- 0
Age- 1
-2.5
-2.0
-1.5
-1.0
-0.5
0.0
1.21.00.80.60.40.20.0
δ18 O
, ‰
Weight, mg
Canada
PNW '03
Humbold tMon terey
PNW '04
B
BMagGulf
Ensenada
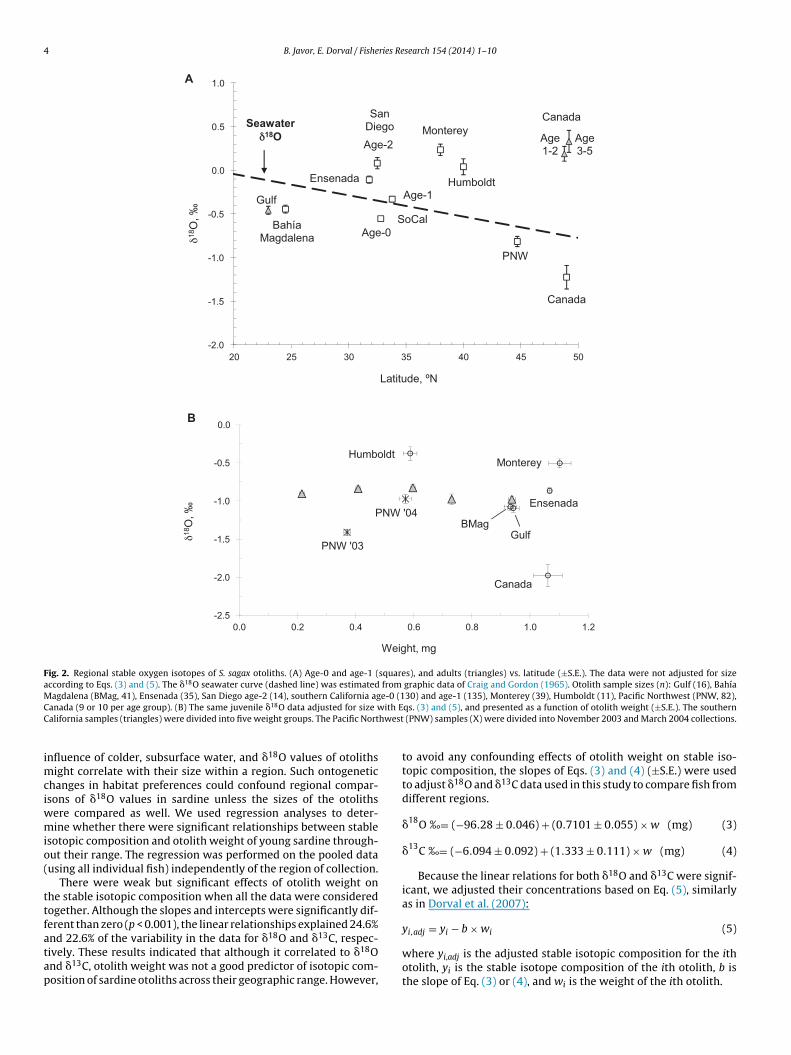
Fig. 2. Regional stable oxygen isotopes of S. sagax otoliths. (A) Age-0 and age-1 (squares), and adults (triangles) vs. latitude (±S.E.). The data were not adjusted for sizeaccording to Eqs. (3) and (5). The �18O seawater curve (dashed line) was estimated from graphic data of Craig and Gordon (1965). Otolith sample sizes (n): Gulf (16), BahíaMagdalena (BMag, 41), Ensenada (35), San Diego age-2 (14), southern California age-0 (130) and age-1 (135), Monterey (39), Humboldt (11), Pacific Northwest (PNW, 82),C ith EC hwest
imciwmio(
ttfatap
anada (9 or 10 per age group). (B) The same juvenile �18O data adjusted for size walifornia samples (triangles) were divided into five weight groups. The Pacific Nort
nfluence of colder, subsurface water, and �18O values of otolithsight correlate with their size within a region. Such ontogenetic
hanges in habitat preferences could confound regional compar-sons of �18O values in sardine unless the sizes of the otoliths
ere compared as well. We used regression analyses to deter-ine whether there were significant relationships between stable
sotopic composition and otolith weight of young sardine through-ut their range. The regression was performed on the pooled datausing all individual fish) independently of the region of collection.
There were weak but significant effects of otolith weight onhe stable isotopic composition when all the data were consideredogether. Although the slopes and intercepts were significantly dif-erent than zero (p < 0.001), the linear relationships explained 24.6%
nd 22.6% of the variability in the data for �18O and �13C, respec-ively. These results indicated that although it correlated to �18Ond �13C, otolith weight was not a good predictor of isotopic com-osition of sardine otoliths across their geographic range. However,qs. (3) and (5), and presented as a function of otolith weight (±S.E.). The southern (PNW) samples (X) were divided into November 2003 and March 2004 collections.
to avoid any confounding effects of otolith weight on stable iso-topic composition, the slopes of Eqs. (3) and (4) (±S.E.) were usedto adjust �18O and �13C data used in this study to compare fish fromdifferent regions.
�18O ‰= (−96.28 ± 0.046) + (0.7101 ± 0.055) × w (mg) (3)
�13C ‰= (−6.094 ± 0.092) + (1.333 ± 0.111) × w (mg) (4)
Because the linear relations for both �18O and �13C were signif-icant, we adjusted their concentrations based on Eq. (5), similarlyas in Dorval et al. (2007):
y = y − b × w (5)
i,adj i iwhere yi,adj is the adjusted stable isotopic composition for the ithotolith, yi is the stable isotope composition of the ith otolith, b isthe slope of Eq. (3) or (4), and wi is the weight of the ith otolith.
B. Javor, E. Dorval / Fisheries Research 154 (2014) 1–10 5
-1.2
-0.8
-0.4
0.0
0.4
0.0 0.4 0.8 1.2 1.6
δ18 O
, ‰
Weight, mg
Age-1 1995-2003
Age-1 2006 & 2008
Age 2 2007
A
10
14
18
22
0.0 0.4 0. 8 1.2 1. 6
Tem
pera
ture
, ºC
Weight, mg
B
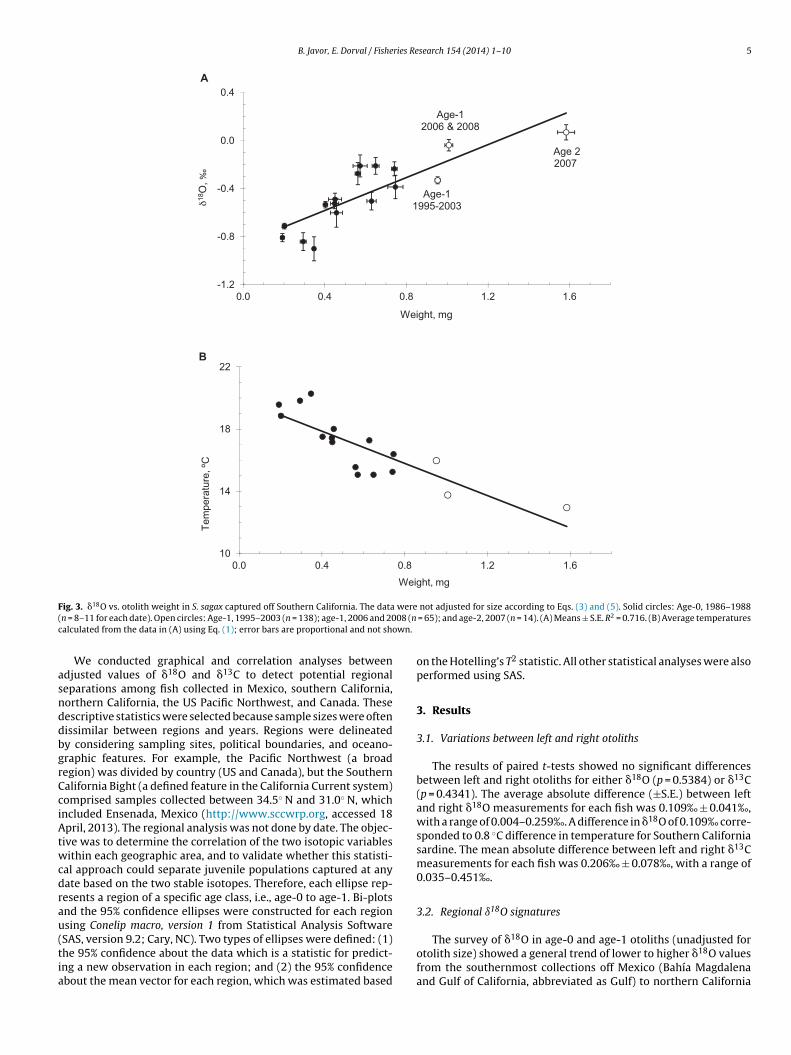
F were( 08 (nc wn.
asnddbgrCciAtwcdrau(tia
ig. 3. �18O vs. otolith weight in S. sagax captured off Southern California. The datan = 8–11 for each date). Open circles: Age-1, 1995–2003 (n = 138); age-1, 2006 and 20alculated from the data in (A) using Eq. (1); error bars are proportional and not sho
We conducted graphical and correlation analyses betweendjusted values of �18O and �13C to detect potential regionaleparations among fish collected in Mexico, southern California,orthern California, the US Pacific Northwest, and Canada. Theseescriptive statistics were selected because sample sizes were oftenissimilar between regions and years. Regions were delineatedy considering sampling sites, political boundaries, and oceano-raphic features. For example, the Pacific Northwest (a broadegion) was divided by country (US and Canada), but the Southernalifornia Bight (a defined feature in the California Current system)omprised samples collected between 34.5◦ N and 31.0◦ N, whichncluded Ensenada, Mexico (http://www.sccwrp.org, accessed 18pril, 2013). The regional analysis was not done by date. The objec-
ive was to determine the correlation of the two isotopic variablesithin each geographic area, and to validate whether this statisti-
al approach could separate juvenile populations captured at anyate based on the two stable isotopes. Therefore, each ellipse rep-esents a region of a specific age class, i.e., age-0 to age-1. Bi-plotsnd the 95% confidence ellipses were constructed for each regionsing Conelip macro, version 1 from Statistical Analysis Software
SAS, version 9.2; Cary, NC). Two types of ellipses were defined: (1)he 95% confidence about the data which is a statistic for predict-ng a new observation in each region; and (2) the 95% confidencebout the mean vector for each region, which was estimated basednot adjusted for size according to Eqs. (3) and (5). Solid circles: Age-0, 1986–1988 = 65); and age-2, 2007 (n = 14). (A) Means ± S.E. R2 = 0.716. (B) Average temperatures
on the Hotelling’s T2 statistic. All other statistical analyses were alsoperformed using SAS.
3. Results
3.1. Variations between left and right otoliths
The results of paired t-tests showed no significant differencesbetween left and right otoliths for either �18O (p = 0.5384) or �13C(p = 0.4341). The average absolute difference (±S.E.) between leftand right �18O measurements for each fish was 0.109‰ ± 0.041‰,with a range of 0.004–0.259‰. A difference in �18O of 0.109‰ corre-sponded to 0.8 ◦C difference in temperature for Southern Californiasardine. The mean absolute difference between left and right �13Cmeasurements for each fish was 0.206‰ ± 0.078‰, with a range of0.035–0.451‰.
3.2. Regional ı18O signatures
The survey of �18O in age-0 and age-1 otoliths (unadjusted forotolith size) showed a general trend of lower to higher �18O valuesfrom the southernmost collections off Mexico (Bahía Magdalenaand Gulf of California, abbreviated as Gulf) to northern California
6 B. Javor, E. Dorval / Fisheries Research 154 (2014) 1–10
15
17
19
21
0.3 0.4 0.5 0.6 0.7 0.8
Tem
pera
ture
, ºC
Otolith, mg
11/04
2/03
2/0410/05
9/05
A
10
14
18
22
2002 200 3 200 4 200 5 200 6
Tem
pera
ture
, ºC
Year
Otol ithsSW, 0 mSW, 30 m
0.741
0.747
0.338
0.450
0.565
B
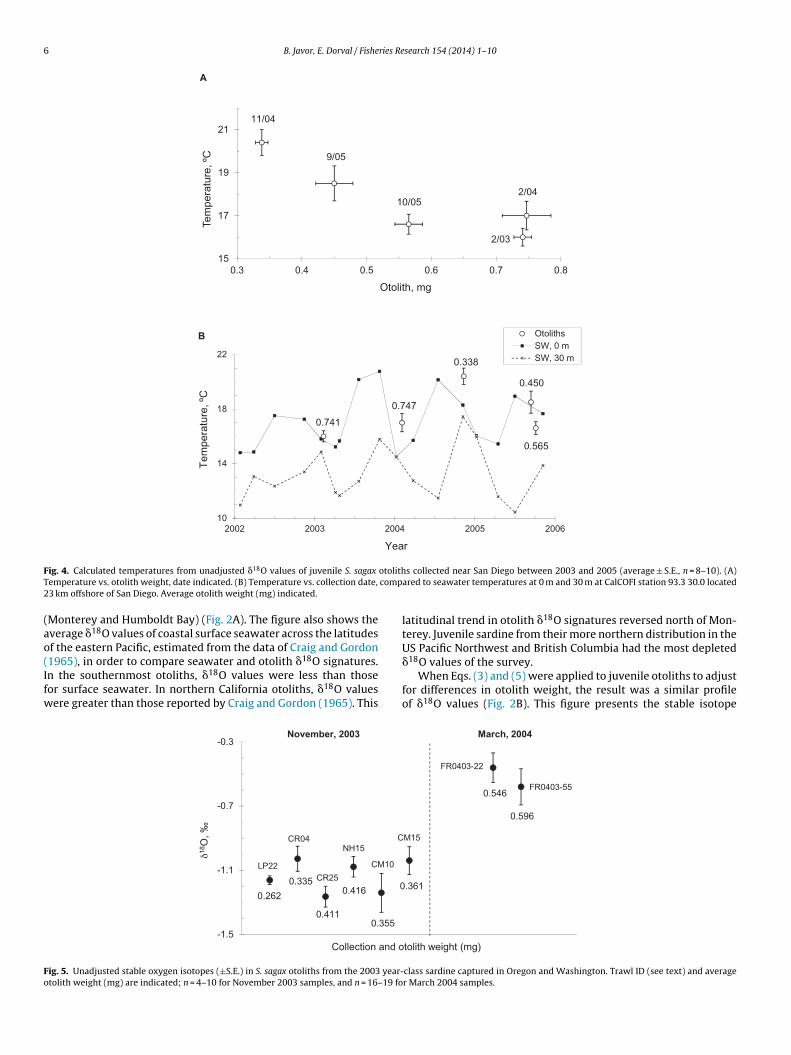
F otolithT comp2
(ao(Ifw
Fo
ig. 4. Calculated temperatures from unadjusted �18O values of juvenile S. sagax
emperature vs. otolith weight, date indicated. (B) Temperature vs. collection date,3 km offshore of San Diego. Average otolith weight (mg) indicated.
Monterey and Humboldt Bay) (Fig. 2A). The figure also shows theverage �18O values of coastal surface seawater across the latitudesf the eastern Pacific, estimated from the data of Craig and Gordon
1965), in order to compare seawater and otolith �18O signatures.n the southernmost otoliths, �18O values were less than thoseor surface seawater. In northern California otoliths, �18O valuesere greater than those reported by Craig and Gordon (1965). ThisLP22
CR04
CR25
NH15
CM10
C
-1.5
-1.1
-0.7
-0.3
0 2 4
δ18 O
, ‰
Collect ion and o
0.2620.335
0.411
0.416
0.355
0
3002 ,rebmevoN
ig. 5. Unadjusted stable oxygen isotopes (±S.E.) in S. sagax otoliths from the 2003 year-tolith weight (mg) are indicated; n = 4–10 for November 2003 samples, and n = 16–19 fo
s collected near San Diego between 2003 and 2005 (average ± S.E., n = 8–10). (A)ared to seawater temperatures at 0 m and 30 m at CalCOFI station 93.3 30.0 located
latitudinal trend in otolith �18O signatures reversed north of Mon-terey. Juvenile sardine from their more northern distribution in theUS Pacific Northwest and British Columbia had the most depleted
�18O values of the survey.When Eqs. (3) and (5) were applied to juvenile otoliths to adjustfor differences in otolith weight, the result was a similar profileof �18O values (Fig. 2B). This figure presents the stable isotope
M15
FR0403 -22
FR0403 -55
6 8 10 12tolith weight (mg)
.361
0.546
0.596
4002 ,hcraM
class sardine captured in Oregon and Washington. Trawl ID (see text) and averager March 2004 samples.

B. Javor, E. Dorval / Fisheries Research 154 (2014) 1–10 7
Table 2Growth, otolith stable isotopes, and calculated temperatures of Pacific sardine (S. sagax) in the tank experiment. The �18O values were not adjusted with Eqs. (3) and (5).Measurements are reported as ranges or ±S.E. The correlation coefficients were determined from the otolith weight–�18O relationships in each the Time-0 and Time-8months sample sets.
Sample set n SL (mm) Otolith (mg) �18O (‰) Calculated (◦C) Correl. coeff.
vCs(usni
3
eit(sfoceat((a
dctar(aala
tic(csscwts
3
twso
Time-0 8 69± 1 0.338 ± 0.010
Time-8 mos 27 71–141 0.494–1.177
alues versus otolith weight rather than latitude. Southernalifornia otoliths were divided into five size groups to demon-trate the validity of the adjustments after applying Eqs. (3) and5). Northern California juveniles still had the greatest �18O val-es, and Canadian juveniles had the most depleted values. Whentable oxygen isotope values were adjusted for otolith weight, juve-ile samples from southern California and Mexico were essentially
ndistinguishable.
.3. Otolith weight or age effects
As sardine grew during their first year of life in the South-rn California Bight, their otolith �18O signatures also generallyncreased (Fig. 3A). To demonstrate the effects of otolith weight,he data in the figure were not adjusted by applying Eqs. (3) and5). The data represent three decades of samples, including age-0ardine collected on 16 dates during 1986–1988, pre-adult samplesrom 2003 to 2005, and averaged yearly or multi-yearly samplesf age-1 and age-2 sardine. The averaged data for age-1 otolithsollected as spring samples during 1995–2003 are from Dorvalt al. (2011). The samples included collections near Los Angelesnd San Diego, up to about 200 km apart in the Bight. A linearrend described the relationship between the mean �18O valuesy) and otolith weights (x) in the Southern California datasetsy = 0.6853x − 0.8557, R2 = 0.7134). The average calculated temper-tures correlated inversely to weight (Fig. 3B).
Five sample sets of juvenile sardine collected off San Diegouring 2003–2005 also showed a similar weight-temperatureorrelation in unadjusted data (Fig. 4A). The calculated otolithemperatures were compared with local seawater temperaturest 0 m and 30 m (CalCOFI station 93.3 30.0, 23 km from shore)ecorded over the course of the study (Fig. 4B). The smallest otolithsNovember, 2004, and September, 2005) appeared to track theverage surface temperatures between their springtime spawn andutumn. The larger juvenile otoliths appeared to record accruedifetime temperatures that corresponded to averages between 0 mnd 30 m depth.
A juvenile sardine cohort that presumably overwintered alonghe Oregon and Washington coasts during 2003–2004 also exhib-ted greater �18O values (i.e., colder temperatures) in the Marchollection compared to samples collected the previous NovemberFig. 5). Two of the November sample sets, CR04 and CR25, wereaptured in the Columbia River plume. Their �18O signatures wereimilar to those of sardine caught away from the direct freshwaterource along the central Washington coast (LP22), central Oregonoast (NH15), and southern Oregon coast (CM10 and CM15). Like-ise, the �18O signatures of the March 2004 otoliths from sardine
aken in the Columbia River plume (FR0403-55) resembled those ofample FR0403-22 captured further south during the same cruise.
.4. Effect of growth rate
Otoliths from juveniles maintained for 8 months in a single
ank at ambient seawater temperature were analyzed to evaluatehether the trend of increasing stable isotope values with otolithize in wild sardine might be a result of vital effects or growth raten aragonite deposition rather than as a response to temperature
−0.881 ± 0.088 20.1 ± 0.7 −0.0417−0.615 ± 0.027 18.1 ± 0.2 −0.1641
shifts in the environment. In this experiment, some fish hardly grewwhile others doubled in size (Table 2). The isotopic values corre-sponded to average local surface seawater temperatures (Fig. 4B).There was negligible correlation between otolith weight and �18Ovalues in either the Time-0 juveniles or in the sardine that weremaintained for 8 months, demonstrating that vital effects wereundetectable.
3.5. Graphical and correlation analyses of ı13C and ı18O
Oxygen and carbon isotopic values correlated positively in juve-nile otoliths, but the strength of their relationships differed byregion. Both original data and adjusted compositions of �18O and�13C (from Eqs. (3) and (5)) correlated well for fish collected insouthern and northern California (correlation factors >0.7). Sar-dine collected in Mexico and Canada showed modest correlationsbetween these two variables (correlation factors = 0.3).
The shapes of the 95% confidence ellipses about the data andabout the mean vectors for each region are graphical represen-tations of the strength of the correlation between �18O and �13C(Fig. 6). The correlation results for the samples from Canada werepoor, likely due to the small sample size (n = 10). The 95% confidenceellipses about the mean vectors showed clear separations betweensome of the regions (Fig. 6B). For example, otoliths from the USPacific Northwest and Mexico had similar 95% confidence intervals,but they were different from those estimated for juveniles collectedin Canada and both northern and southern California. The resultsfor the juvenile otoliths from Canada did not follow the generalpattern of the other otoliths because the �18O values were moredepleted (Figs. 2 and 6A).
4. Discussion
This investigation evaluated several factors that might influencethe use of stable oxygen isotopes in otoliths of pre-adults for distin-guishing and connecting Pacific sardine stocks in North America:variability within an individual fish; size (weight) effects withinthe first year of life; and growth rate effects in juveniles. The sur-vey profiled �18O values in otoliths of juvenile and adult sardinethroughout most of their range, and determined the validity ofregional correlations between �18O and �13C values in juveniles.
The mean difference in �18O values between left and rightotoliths was low relative to the typical standard errors of the aver-aged measurements for each region sampled in this study as wellas those reported in the Pacific sardine studies of Valle and Herzka(2008) and Dorval et al. (2011). Our measurements were similar to<0.5 ◦C differences recorded in stable oxygen isotopes between leftand right otoliths in juvenile cod, Gadus morhua (Høie et al., 2003,2004).
The survey of juvenile Pacific sardine otoliths revealed two con-trasting latitudinal trends between southern and northern regions.One trend was that otoliths tended to have lighter �18O signaturesin their southern distribution in Mexico and southern California,
and heavier �18O values in northern California fish, the predictedresponse to sea surface temperature in these latitudes (23◦ N to40◦ N). This stretch of coastline has few freshwater effluents andgenerally low rainfall: ≤25 cm/year between the southern tip of
8 B. Javor, E. Dorval / Fisheries Re
-9
-7
-5
-3
0-1-2-3
δ13 C
δ18O
A
Can
SCA
NCA
MexPNW
-9
-7
-5
-3
0-1-2-3
δ13 C
δ18O
B
Can NCA
SCAMex
PNW
Fig. 6. Relationships between stable oxygen and carbon isotopes in juvenile S. sagaxotoliths. Ellipses represent 95% confidence limits of the dataset (A) and of the means(B). Samples are grouped from Canada (Can); US Pacific Northwest (PNW, Oregonand Washington); Northern California (NCA); Southern California and Ensenada(r
BAtsa
oajTs(tsaWCT(u1lo
SCA); and Mexico (Mex, Bahía Magdalena and Gulf of California). Sample sizes areeported in the legend of Fig. 2.
aja California and San Diego to nearly 50 cm/year in Monterey. general south-to-north trend of decreasing salinity and surface
emperature along this section of coastline corresponds to surfaceeawater �18O values also decreasing from south to north (Craignd Gordon, 1965), a trend opposite that of the otoliths.
North of California, the latitudinal trend reversed, and juveniletoliths were more depleted in �18O than those from both Mexicond northern California sardine. The most isotopically depleteduvenile otoliths came from the northernmost collection in Canada.he isotopic signatures of the northern juveniles likely reflectedeawater temperatures as well as coastal dilution from riversColumbia and Fraser), streams, and abundant precipitation alonghe coastlines of Oregon and Washington (about 100 cm/year) tooutheast Alaska (nearly 400 cm/year at Ketchikan, 54◦ N). Theverage �18O composition of freshwater entering Puget Sound fromashington is about −10‰ to −16‰, and at the mouth of the
olumbia River is about −8‰ to −10‰ (Kendall and Coplen, 2001).he �18O of seawater near Ketchikan is influenced by snowmelt−11.7‰) and streams (−13.9‰), resulting in seawater �18O val-es of −1.08‰ to −1.87‰, and salinities of 31.32–32.42‰ (Kipphut,
990). The average �18O value measured in the juvenile otoliths col-ected off Canada, −1.225‰, likely reflected the isotopic signaturef seawater in that coastal region.
search 154 (2014) 1–10
�18O differences were not detected between otoliths fromjuvenile sardine captured near the mouth of the Columbia Riverand those caught farther north or south in the Pacific North-west during the same surveys, suggesting the young fish migratedin and out of �18O-depleted plume water, or else the plumewas broad and diffuse (Kaltenberg et al., 2010). The relativelywide range of �18O values in the March 2004 juveniles (±1.113‰to ±1.464‰) corresponded to calculated differences in otolithtemperatures of up to 8–11 ◦C within a trawl if the �H2
18O his-tory was presumed to be uniform for all the fish. Because thesedifferences exceeded the variations in Oregon coastal tempera-tures during this period of growth, the most likely explanationis the isotopic signatures reflected local variations in �18O ofseawater.
Other published surveys of stable isotopes of oxygen in otolithsof S. sagax found small to large differences within collections.The average variation of 0.5‰ determined in Australian S. sagax(Edmonds and Fletcher, 1997) was similar to the typical range ofvalues found in single trawls in the present study. Pacific sardinefrom Mexico showed variations up to 2.0‰ in single collections(Valle and Herzka, 2008). In related species, the European sardine(Sardina pilchardis) varied 1.4‰ within a collection (Iacumin et al.,1992) while the Australian herring Arripis georgiana varied 1.6‰(Ayvazian et al., 2004). Large differences were found in otoliths ofPacific herring (Clupea pallasi) that spawned in Puget Sound, Wash-ington, a basin highly influenced by freshwater (Gao et al., 2001b).Otolith nuclei (natal signature) yielded �18O values of −1‰ to −8‰whereas otolith material extracted from the second summer ringyielded values of −1‰ to about −4‰.
Unlike otoliths from juveniles captured in Canada, otoliths fromadults caught off Vancouver Island and the US Pacific Northwestwere enriched in �18O and had similar isotopic signatures as juve-niles fished near Monterey and adults captured near San Diego.Catch records in Canada and the Pacific Northwest indicate largeradult sardine undergo annual northward migrations as far as BritishColumbia in the summer and return south to California spawninggrounds by mid-October. Juveniles and adults occasionally over-winter in the north (McFarlane et al., 2005; Emmett et al., 2005).
Because of migration, the calculated temperatures recorded inthe otoliths of older adults cannot reflect the �18O composition ofseawater with any certainty. Using the temperature–�18O equationof Dorval et al. (2011) based on the isotopic signature of San Diegoseawater as an estimate, the average temperatures recorded in theotoliths of Canadian adults shown in Fig. 2 would be 11–13 ◦C. Thesevalues concur with average summer sea surface temperatures (typ-ically 11–14 ◦C) recorded near Vancouver Island where sardine arefished (McFarlane et al., 2005). In Oregon and Washington, smallsardine (<130 mm fork length) were captured in 12–16 ◦C water,whereas larger sardine were caught in water as cold as 10–12 ◦C(Emmett et al., 2005).
If otoliths of adult sardine reflected the �18O composition ofsurface seawater near Monterey (Fig. 2, calculated from Craig andGordon (1965)), the estimated otolith temperature for those fishwould be 12–14 ◦C, which is the range of sea surface temperaturesdetermined for stock that spawns offshore central and southernCalifornia (Reiss et al., 2008). Dorval et al. (2011) demonstratedage-1 sardine otoliths in the Southern California Bight had calcu-lated temperatures that were less than surface temperatures andlikely reflected a habitat ranging from 0 m to 30 m depth, or about14–17 ◦C.
These conclusions have several ramifications for interpretingthe stock structure of Pacific sardine. Because �18O values changedwith juvenile growth in nature, it is critical to know their size
and age to validate comparisons between regions and times ofcollection. The wide variation in �18O values measured in juve-niles captured in March 2004 in Oregon and Washington, and in
ries Re
Msted
ScwntTn�ztaWan1ba2
s(lCctutcs
w22edcsdHohOaa
fia1(actttaotdis
B. Javor, E. Dorval / Fishe
exican sardine (Valle and Herzka, 2008), confounds the preci-ion of stable oxygen isotopes as signatures of regional stocks inhose areas. In addition, variations in values determined by differ-nt mass spectrometers may make comparisons between studiesifficult without cross-validation.
Otoliths from wild-caught juvenile sardine captured in theouthern California Bight integrated higher �18O values, henceolder temperatures, as they grew during their first year, no matterhich year they were collected. In the Pacific Northwest, this phe-omenon may have also reflected seasonal changes in sea surfaceemperatures in juveniles that presumably overwintered there.he surveys of southern California sardine suggest younger juve-iles tracked sea surface temperatures while older juveniles had18O signatures that integrated temperatures within the mixingone of the top 30 m. Dorval et al. (2011) used this explana-ion to describe otolith temperature history of age-1 sardine in
9-year survey in the Southern California Bight. Trawl data (Billatson, SWFSC, personal communication), acoustic data (Cutter
nd Demer, 2008; Kaltenberg et al., 2010), and comparisons toorthern anchovy (Engraulis mordax) behavior in this region (Mais,974) support the conclusion that Pacific sardine generally rangeetween 0 and 30 m or deeper with diurnal migrations to for-ge for zooplankton and phytoplankton (van der Lingen et al.,006).
In contrast, the correlation between size of the juveniles andtable isotope values was negligible in the study Valle and Herzka2008) conducted on the population of Mexican sardine. They ana-yzed samples from Ensenada, Bahía Magdalena, and the Gulf ofalifornia. In their sample sets that included two different sizelasses of juveniles collected from the same sites, the otoliths fromhe larger sardine did not record colder temperatures. The �18O val-es of the otoliths from their study and ours overlapped, indicatinghe differences were not likely due to instrumentation or analyti-al procedures. It is unclear why their samples did not demonstrateize effects.
Eggs, larvae, and juveniles of sardine (S. sagax and S. pilchardis)orldwide are found in the surface mixed layer (Uehara et al.,
005; Tsagarakis et al., 2008; Osman et al., 2010; Inda-Díaz et al.,010) where they are distributed according to temperature prefer-nces. In anchovy (Engraulis encrasicolus), differences in planktonicistributions by size suggested that larger larvae exhibited verti-al diving behavior in South Africa (Osman et al., 2010). A similarhift in behavior with age may have occurred in young Pacific sar-ine if they spent less and less time at the surface as they grew.abitat temperature change during juvenile growth, recorded intolith oxygen isotopes, has been noted in other fish as well, e.g.,addock (Melanogrammus aeglefinus) from the northwest Atlanticcean (Begg and Weidman, 2001) and orange roughy (Hoplostethustlanticus) that are pelagic during their juvenile stage and benthics adults (Shephard et al., 2007).
It is generally accepted that oxygen isotope discrimination insh otoliths is not influenced by vital effects or growth rates,lthough such effects have been described in corals (Emiliani et al.,978; Spiro et al., 2000; López Correa et al., 2010) and foraminiferaSpero and Lea, 1996). Vital effects, i.e., isotope fractionation as
factor of size or age rather than temperature, could hypotheti-ally explain the trend in �18O values in age-0 sardine. Culturinghe sardine in a single tank eliminated environmental variations inemperature between fish, and behavioral preferences for depth oremperature. Our experiment at ambient surface seawater temper-tures that resulted in differential growth did not produce a slopef �18O discrimination responses found in wild juvenile sardine of
he same sizes. The most likely explanation is that in nature, sar-ine controlled their habitat temperature as they grew, i.e., theyncreased their time in colder water by remaining longer at sub-urface depths.
search 154 (2014) 1–10 9
Although �13C and �18O values in juvenile otoliths generallycorrelated positively, the correlations were not very strong in thenorthernmost and southernmost collections. The apparent uncou-pling of the fractionation response of the two elements in theMexican sardine was most likely due to the variable influence ofmetabolic 13C in the bicarbonate pool of the endolymph where theotoliths grew. This phenomenon is well documented in fish (Kalish,1991a,b; Iacumin et al., 1992; Thorrold et al., 1997; Ashford andJones, 2007). Our results suggest �13C–�18O correlations in sardineotoliths may not be a robust statistic for discriminating betweenstocks.
Recognizing the qualitative and descriptive nature of this study,future investigations of otolith stable isotopes in Pacific sardine canbe designed to expand the tests for regional differences in pre-adults to identify stock structure. This study also highlights theimportance of evaluating age-0 juveniles that are usually too smallto be captured by commercial seiners, and hence are lacking in mostport samples of commercial landings.
In summary, across the range of Pacific sardine on theNorth American coast, local seawater composition in the PacificNorthwest apparently affected the north-south trends in �18O com-position of juvenile otoliths that temperature alone did not predict.Within the Southern California Bight, temperature histories of juve-niles followed a pattern that correlated with the size (weight) ofthe otoliths which likely resulted from changes in depth prefer-ences as the fish grew. These results demonstrated the importanceof comparing otoliths of similar weight when evaluating stable iso-tope signatures in sardine. Otoliths of adult sardine in the northernstock, collected between California and Canada, recorded coolertemperatures than most otoliths from juveniles, and no regionaldifferentiation. These results are consistent with an adult stock thatmixes during annual migrations.
Acknowledgments
We thank the agencies and individuals cited in the text for col-lecting fish and otoliths. We thank L. Robertson for culturing theexperimental sardine. We thank Jeff Seminoff and Nancy Lo forcritical discussions of the manuscript.
References
Ashford, J., Jones, C., 2007. Oxygen and carbon stable isotopes in otoliths recordspatial isolation of Patagonian toothfish (Dissostichus eleginoides). Geochim. Cos-mochim. Acta 71 (1), 87–94.
Ayvazian, S.G., Bastow, T.P., Edmonds, J.S., How, J., Nowara, G.B., 2004. Stock structureof Australian herring (Arripis georgiana) in southwestern Australia. Fish. Res. 67,39–53.
Begg, G.A., Weidman, C.R., 2001. Stable �13C and �18O isotopes in otoliths of haddockMelanogrammus aeglefinus from the northwest Atlantic Ocean. Mar. Ecol. Prog.Ser. 216, 223–233.
Bigg, G.R., Rohling, E.J., 2000. An oxygen isotope data set for marine waters. J. Geo-phys. Res. 105 (C4), 8517–8535.
Campana, S.E., 1999. Chemistry and composition of fish otoliths. Mar. Ecol. Prog. Ser.188, 263–297.
Craig, H., Gordon, L.I., 1965. Deuterium and oxygen 18 variations in the ocean andmarine atmosphere. In: Tongiorgi, E. (Ed.), Stable isotopes in oceanographicstudies and paleotemperatures. Cons. Naz. Di Rech., Spoleto, Italy, pp. 9–130.
Cutter Jr., G., Demer, D.A. (Eds.), 2008. California Current Ecosystem Survey 26 Acous-tic Cruise Reports for NOAA FRV Oscar Dyson and NOAA FRV David Starr Jordan.NOAA-TM-NMFS-SWFSC-415.
Dorval, E., Jones, C.M., Hannigan, R., van, Montfrans, 2007. Relating otolith chemistryto surface water chemistry in a coastal plain estuary. Can. J. Fish Aquat. Sci. 64,411–424.
Dorval, E., Piner, K., Robertson, L., Reiss, C.S., Javor, B., Vetter, R., 2011. Temperaturerecord in the oxygen stable isotopes of Pacific sardine otoliths: experimental vs.
wild stocks from the Southern California Bight. J. Exp. Mar. Biol. Ecol. 397 (2),136–143.Edmonds, J.S., Fletcher, W.J., 1997. Stock discrimination of pilchards Sardinops sagaxby stable isotope ratio analysis of otolith carbonate. Mar. Ecol. Prog. Ser. 152,241–247.

1 ries Re
E
E
E
F
F
G
G
G
G
G
G
H
H
H
H
I
I
J
J
K
K
K
K
K
L
Atlantic cod. Fish. Res. 46 (1–3), 327–342.
0 B. Javor, E. Dorval / Fishe
lsdon, T.S., Gillanders, B.M., 2002. Interactive effects of temperature and salinity onotolith chemistry: challenges for determining environmental histories of fish.Can. J. Fish. Aquat. Sci. 59 (11), 1796–1808.
miliani, C., Hudson, J.H., Shinn, E.A., George, R.Y., 1978. Oxygen and carbon isotopicgrowth record in a reef coral from the Florida Keys and a deep-sea coral fromBlake Plateau. Science 202 (4368), 627–629.
mmett, R.L., Brodeur, R.D., Miller, T.W., Pool, S.S., Krutzikowsky, G.K., Bentley, P.J.,McCrae, J., 2005. Pacific sardine (Sardinops sagax) abundance, distribution, andecological relationships in the Pacific Northwest. Calif. Coop. Oceanic Fish. Invest.Rep. 46, 122–143.
élix-Uraga, R., Gómez-Munoz, V.M., Quinonez-Velázquez, C., Melo-Barrera, F.N.,García-Franco, W., 2004. On the existence of Pacific sardine groups off thewest coast of the Baja California Peninsula and southern California. Calif. Coop.Oceanic Fish. Invest. Rep. 45, 146–151.
élix-Uraga, R., Quinónez-Velázquez, C., Hill, K.T., Gómez-Munoz, V.M., Melo-Barrera, F.N., García-Franco, W., 2005. Pacific sardine (Sardinops sagax) stockdiscrimination off the west coast of Baja California and southern California usingotolith morphometry. Calif. Coop. Oceanic Fish. Invest. Rep. 46, 113–121.
ao, Y.W., Beamish, R.J., 2003. Stable isotope variations in otoliths of Pacific halibut(Hippoglossus stenolepis) and indications of the possible 1990 regime shift. Fish.Res. 60 (2–3), 393–404.
ao, Y., Schwarcz, H.P., Brannd, U., Moksness, E., 2001a. Seasonal stable isotoperecords of otoliths from ocean-pen reared and wild cod, Gadus morhua. Environ.Biol. Fishes 61 (4), 445–453.
ao, Y.W., Joner, S.H., Bargmann, G.G., 2001b. Stable isotopic composition in iden-tification of spawning stocks of Pacific herring (Clupea pallasi) in Puget Sound.Can. J. Fish. Aquat. Sci. 58 (11), 2113–2120.
ao, Y., Joner, S.H., Svec, R.A., Weinberg, K.L., 2004. Stable isotopic comparison inotoliths of juvenile sablefish (Anoplopoma fimbria) from waters off the Wash-ington and Oregon coast. Fish. Res. 68 (1–3), 351–360.
arcía-Rodríguez, F.J., García-Gasca, S.A., De La Cruz-Agüero., J., Cota-Gómez, V.M.,2011. A study of the population structure of the Pacific sardine Sardinops sagax(Jenyns, 1942) in Mexico based on morphometric and genetic analyses. Fish. Res.107 (1–3), 169–176.
rant, W.S., Bowen, B.W., 1998. Shallow population histories in deep evolutionarylineages of marine fishes: insights from sardines and anchovies and lessons forconservation. J. Heredity 89 (5), 415–426.
edgecock, D., Hutchinson, E.S., Li, G., Sly, F.L., Nelson, K., 1989. Genetic and morpho-metric variations in the Pacific sardine Sardinops sagax caerulea: comparisonsand contrasts with historical data and with variability in northern anchovyEngraulis mordax. Fish. Bull. U.S. 87 (3), 653–671.
øie, H., Folkvord, A., 2006. Estimating the timing of growth rings in Atlantic codotoliths using stable oxygen isotopes. J. Fish Biol. 68 (3), 826–837.
øie, H., Folkvord, A., Otterlei, E., 2003. Effect of somatic and otolith growth rate onstable isotopic composition of early juvenile cod (Gadus morhua L.) otoliths. J.Exp. Mar. Biol. Ecol. 289, 41–58.
øie, H., Otterlei, E., Folkvord, A., 2004. Temperature-dependent fractionation ofstable oxygen isotopes in otoliths of juvenile cod (Gadus morhua L.). ICES J. Mar.Sci. 61 (90), 243–251.
acumin, P., Bianucci, G., Longinelli, A., 1992. Oxygen and carbon isotopic composi-tion of fish otoliths. Mar. Biol. 113 (4), 537–542.
nda-Díaz, E.A., Sánchez-Velasco, L., Lavín, M.E., 2010. Three-dimensional distribu-tion of small pelagic fish larvae (Sardinops sagax and Engraulis mordax) in atidal-mixing front and surrounding waters (Gulf of California). J. Plankton Res.32 (9), 1241–1254.
avor, B.J., 2013. Do shifts in otolith morphology of young Pacific sardine (Sardinopssagax) reflect changing recruitment contributions from northern and southernstocks? Calif. Coop. Oceanic Fish. Invest. Rep. 54, 85–96.
avor, B., Lo, N., Vetter, R., 2011. Otolith morphometrics and population structureof Pacific sardine (Sardinops sagax) along the west coast of North America. Fish.Bull. 109 (4), 402–415.
alish, J.M., 1991a. 13C and 18O isotopic composition disequilibria in fish otoliths:metabolic and kinetic effects. Mar. Ecol. Prog. Ser. 75 (2–3), 191–203.
alish, J.M., 1991b. Oxygen and carbon stable isotopes in the otoliths of wild and lab-oratory maintained Australian salmon (Arripis trutta). Mar. Biol. 110 (1), 37–47.
altenberg, A.M., Emmett, R.L., Benoit-Bird, K.J., 2010. Timing of forage fish seasonalappearance in the Columbia River plume and link to ocean conditions. Mar. Ecol.Prog. Ser. 419, 171–184.
endall, C., Coplen, T.B., 2001. Distribution of oxygen-18 and deuterium in river
waters across the United States. Hydrol. Processes 15 (7), 1363–1393.ipphut, G.W., 1990. Glacial meltwater input to the Alaska coastal current: evidencefrom oxygen isotope measurements. J. Geophys. Res. 95 (C4), 5177–5181.
eGrande, A.N., Schmidt, G.A., 2006. Global gridded data set of the oxygen isotopiccomposition in seawater. Geophys. Res. Lett. 33 (L12604), 1–5.
search 154 (2014) 1–10
Lo, N.C.H., Macewicz, B.J., Griffith, D.A., 2005. Spawning biomass of Pacific sar-dine (Sardinops sagax), from 1994–2004 off California. Calif. Coop. Oceanic Fish.Invest. Rep. 46, 93–112.
Lo, N.C.H., Macewicz, B.J., Griffith, D.A., 2010. Biomass and reproduction of Pacificsardine (Sardinops sagax) off the Pacific northwestern United States, 2003–2005.Fish. Bull. 108 (2), 174–192.
Lo, N.C.H., Macewicz, B.J., Griffith, D.A., 2011. Migration of Pacific sardine (Sardinopssagax) off the west coast of the United States. Bull. Mar. Sci. 87 (3),395–412.
López Correa, M., Montagna, P., Vendrell-Simón, B., McCulloch, M., Taviani, M., 2010.Stable isotopes (�18O and �13C), trace and minor element compositions of Recentscleractinians and Last Glacial bivalves at the Santa Maria di Leuca deep-watercoral province, Ionian Sea. Deep-Sea Res. II: Top. Stud. Oceanogr. 57 (5–6),471–486.
Mais, K.F., 1974. Pelagic fish surveys in the California Current. Calif. Dep. Fish GameFish. Bull. 162, 1–79.
McFarlane, G.A., Schweigert, J., MacDougall, L., Hrabok, C., 2005. Distribution andbiology of Pacific sardine (Sardinops sagax) off British Columbia, Canada. Calif.Coop. Oceanic Fish. Invest. Rep. 46, 144–160.
Norton, J.G., Mason, J.E., 2005. Relationship of California sardine (Sardinops sagax)abundance to climate-scale ecological changes in the California Current System.Calif. Coop. Oceanic Fish. Invest. Rep. 46, 83–92.
Nøttestad, L., Giske, J., Holst, J.C., Huse, G., 1999. A length-based hypothesis forfeeding migrations in pelagic fish. Can. J. Fish Aquat. Sci. 56 (Suppl. 1), 24–34.
Osman, W., Moloney, C.L., van der Lingen, C.D., 2010. Vertical distribution of smallpelagic fish eggs and larvae on the eastern Agulhas Bank, South Africa. Afr. J.Mar. Sci. 32 (1), 115–125.
Pereyra, R.T., Saillant, E., Pruett, C.L., Rexroad, C.E., Rocha-Olivares, A., Gold, A.R.,2004. Characterization of polymorphic microsatellites in the Pacific sardineSardinops sagax (Clupeidae). Mol. Ecol. Notes 4 (4), 739–741.
Reiss, C.S., Checkley Jr., D.M., Bograd, S.J., 2008. Remotely sensed spawning habitatof Pacific sardine (Sardinops sagax) and Northern anchovy (Engraulis mordax)within the California Current. Fish. Oceanogr. 17 (2), 126–136.
Rooker, J.R., Secor, D.H., DeMetrio, G., Kaufman, A.J., Belmonte Ríos, A., Ticina, V.,2008. Evidence of trans-Atlantic movement and natal homing of bluefin tunafrom stable isotopes in otoliths. Mar. Ecol. Prog. Ser. 368, 231–239.
Shephard, S., Trueman, C., Rickaby, R., Rogan, E., 2007. Juvenile life history of NEAtlantic orange roughy from otolith stable isotopes. Deep-Sea Res. I 54 (8),1221–1230.
Smith, P.E., 2005. A history of proposals for subpopulation structure in the Pacificsardine (Sardinops sagax) population off western North America. Calif. Coop.Oceanic Fish. Invest. Rep. 46, 75–82.
Sokal, R.R., Rohlf, F.J., 1992. Biometry, the Principles and Practice of Statistic inBiological Research, third ed. WH Freeman & Company, New York.
Spero, H.J., Lea, D.W., 1996. Experimental determination of stable isotope variabil-ity in Globigerina bulloides: implications for paleoceanographic reconstructions.Mar. Micropaleontol. 28 (3–4), 231–246.
Spiro, B., Roberts, M., Gage, J., Chenery, S., 2000. 18O/16O and 13C/12C in an aher-matypic deep-water coral Lophelia pertusa from the North Atlantic: a case ofdisequilibrium isotope fractionation. Rapid Commun. Mass Spectrom. 14 (15),1332–1336.
Thorrold, S.R., Campana, S.E., Jones, C.M., Swart, P.K., 1997. Factors determining�13C and �18O fractionation in aragonitic otoliths of marine fish. Geochim. Cos-mochim. Acta 61 (14), 2909–2919.
Tsagarakis, K., Machias, A., Somarakis, S., Giannoulaki, M., Palialexis, A., Valavanis,V.D., 2008. Habitat discrimination of juvenile sardines in the Aegean Sea usingremotely sensed environmental data. Hydrobiologia 612, 215–223.
Uehara, S., Syahailatua, A., Suthers, I.M., 2005. Recent growth rate of pilchardsSardinops sagax in relation to their stable isotope composition, in an upwellingzone of the East Australian Current. Mar. Freshwater Res. 56 (5), 549–560.
Valle, S.R., Herzka, S.Z., 2008. Natural variability in �18O values of otoliths of youngPacific sardine captured in Mexican waters indicates subpopulation mixingwithin the first year of life. ICES J. Mar. Sci. 65 (2), 174–190.
van der Lingen, C.D., Hutchings, L., Field, J.G., 2006. Comparative trophodynamicsof anchovy Engraulis encrasicolus and sardine Sardinops sagax in the southernBenguela: are species alternations between small pelagic fish trophodynami-cally mediated? Afr. J. Mar. Sci. 28 (3–4), 465–477.
Weidman, C.R., Millner, R., 2000. High-resolution stable isotope records from North
Wing, B.L., Murphy, J.M., Rutecki, T.L., 2000. Occurrence of Pacific sardine, Sardinopssagax, off southeastern Alaska. Fish. Bull. 98 (4), 881–883.
Yaremko, M.L., 1996. Age determination in Pacific sardine, Sardinops sagax. NOAATech Memo NMFS-SWFSC-223, 32 pp.




















