
Ageing seabreams: A comparative study between scales and otoliths
12
Available online at www.sciencedirect.com Fisheries Research 89 (2008) 37–48 Ageing seabreams: A comparative study between scales and otoliths D. Abecasis ∗ , L. Bentes, R. Coelho, C. Correia, P.G. Lino, P. Monteiro, J.M.S. Gonc ¸alves, J. Ribeiro, K. Erzini Centro de Ciˆ encias do Mar (CCMAR), Universidade do Algarve, Faculdade de Ciˆ encias do Mar e Ambiente, Campus de Gambelas, 8005-139 Faro, Portugal Received 23 January 2007; received in revised form 21 August 2007; accepted 27 August 2007 Abstract Otoliths and scales were used to determine age and growth of: Boops boops (Linnaeus, 1758), Diplodus vulgaris (Geoffroy Saint-Hilaire, 1817), Diplodus sargus (Linnaeus, 1758), Lithognathus mormyrus (Linnaeus, 1758), Pagellus acarne (Risso, 1827), Pagellus erythrinus (Linnaeus, 1758) and Spondyliosoma cantharus (Linnaeus, 1758). These structures came from previous studies conducted in southern Portugal, and complemented by market sampling and beach seining. Von Bertalanffy growth functions were estimated with otolith and scale readings. Results indicate that otoliths are better structures for ageing these species but scales can also be used as a non-destructive technique and with satisfactory results. The exceptions were P. erythrinus and S. cantharus for which scales provided better results. © 2007 Elsevier B.V. All rights reserved. Keywords: Age estimation techniques; Growth; Otolith; Scale; Sparidae 1. Introduction Seabreams (Sparidae) are found in coastal waters world-wide and sustain important recreational and commercial fisheries (Fischer et al., 1987). This family can be found in a wide vari- ety of marine habitats, from rocky to sand bottoms, at depths between 0 to 500 m, although they are usually more common at less than 150 m deep. In Southern Portugal, seabreams are an abundant group found on the continental shelf, lagoons and estu- aries, with more than 20 species regularly caught (Gonc ¸alves, 2000; Gomes et al., 2001; Sousa et al., 2005; Ribeiro et al., 2006). Age information is important as it forms the basis for the calculations of growth and mortality rates and productivity estimates (Campana, 2001), making it essential for fisheries management (Casselman, 1987; Cailliet et al., 2001). One of the main problems facing age and growth estimates is the selection of the most suitable structure to age the fish. Scales have been used widely for age estimation however, the use of scales has been criticized mainly because the ages of older fish are frequently underestimated (Beamish and McFarlane, 1983; Carlander, 1987). Otolith age determination is thought ∗ Corresponding author. E-mail address: [email protected] (D. Abecasis). to be more accurate because otoliths have a higher priority in utilization of calcium (Carlander, 1987). Furthermore, unlike scales otoliths continue to grow as the fish ages (Beamish and McFarlane, 1983; Casselman, 1987). The objective of this study is to compare the use of two different structures, otoliths and scales, for the study of age and growth, for seven sparid fishes, bogue, Boops boops (Linnaeus, 1758), common two-banded seabream, Diplodus vul- garis (Geoffroy Saint-Hilaire, 1817), white seabream, Diplodus sargus (Linnaeus, 1758), the striped seabream, Lithognathus mormyrus (Linnaeus, 1758), axillary seabream, Pagellus acarne (Risso, 1827), common pandora, Pagellus erythrinus (Linnaeus, 1758), and black seabream, Spondyliosoma cantharus (Lin- naeus, 1758). 2. Materials and methods 2.1. Sampling Samples were obtained from gear selectivity studies con- ducted in south Portugal, complemented by market sampling and beach seining for juveniles inside a temperate coastal lagoon (Ria Formosa, south Portugal). Procedures and sampling details are described in Erzini et al. (1996, 1998, 2003) and Gonc ¸alves (2000). Data collection during gear selectivity studies was made 0165-7836/$ – see front matter © 2007 Elsevier B.V. All rights reserved. doi:10.1016/j.fishres.2007.08.013
Transcript of Ageing seabreams: A comparative study between scales and otoliths

A
Daboe©
K
1
a(ebaaa22
cem
iSufi1
0d
Available online at www.sciencedirect.com
Fisheries Research 89 (2008) 37–48
Ageing seabreams: A comparative study between scales and otoliths
D. Abecasis ∗, L. Bentes, R. Coelho, C. Correia, P.G. Lino,P. Monteiro, J.M.S. Goncalves, J. Ribeiro, K. Erzini
Centro de Ciencias do Mar (CCMAR), Universidade do Algarve, Faculdade de Ciencias do Mar e Ambiente,Campus de Gambelas, 8005-139 Faro, Portugal
Received 23 January 2007; received in revised form 21 August 2007; accepted 27 August 2007
bstract
Otoliths and scales were used to determine age and growth of: Boops boops (Linnaeus, 1758), Diplodus vulgaris (Geoffroy Saint-Hilaire, 1817),iplodus sargus (Linnaeus, 1758), Lithognathus mormyrus (Linnaeus, 1758), Pagellus acarne (Risso, 1827), Pagellus erythrinus (Linnaeus, 1758)
nd Spondyliosoma cantharus (Linnaeus, 1758). These structures came from previous studies conducted in southern Portugal, and complemented
y market sampling and beach seining. Von Bertalanffy growth functions were estimated with otolith and scale readings. Results indicate thattoliths are better structures for ageing these species but scales can also be used as a non-destructive technique and with satisfactory results. Thexceptions were P. erythrinus and S. cantharus for which scales provided better results.2007 Elsevier B.V. All rights reserved.
tusM
da(gsm(1n
2
2
eywords: Age estimation techniques; Growth; Otolith; Scale; Sparidae
. Introduction
Seabreams (Sparidae) are found in coastal waters world-widend sustain important recreational and commercial fisheriesFischer et al., 1987). This family can be found in a wide vari-ty of marine habitats, from rocky to sand bottoms, at depthsetween 0 to 500 m, although they are usually more commont less than 150 m deep. In Southern Portugal, seabreams are anbundant group found on the continental shelf, lagoons and estu-ries, with more than 20 species regularly caught (Goncalves,000; Gomes et al., 2001; Sousa et al., 2005; Ribeiro et al.,006).
Age information is important as it forms the basis for thealculations of growth and mortality rates and productivitystimates (Campana, 2001), making it essential for fisheriesanagement (Casselman, 1987; Cailliet et al., 2001).One of the main problems facing age and growth estimates
s the selection of the most suitable structure to age the fish.cales have been used widely for age estimation however, the
se of scales has been criticized mainly because the ages of oldersh are frequently underestimated (Beamish and McFarlane,983; Carlander, 1987). Otolith age determination is thought∗ Corresponding author.E-mail address: [email protected] (D. Abecasis).
da(a(
165-7836/$ – see front matter © 2007 Elsevier B.V. All rights reserved.oi:10.1016/j.fishres.2007.08.013
o be more accurate because otoliths have a higher priority intilization of calcium (Carlander, 1987). Furthermore, unlikecales otoliths continue to grow as the fish ages (Beamish and
cFarlane, 1983; Casselman, 1987).The objective of this study is to compare the use of two
ifferent structures, otoliths and scales, for the study of agend growth, for seven sparid fishes, bogue, Boops boopsLinnaeus, 1758), common two-banded seabream, Diplodus vul-aris (Geoffroy Saint-Hilaire, 1817), white seabream, Diplodusargus (Linnaeus, 1758), the striped seabream, Lithognathusormyrus (Linnaeus, 1758), axillary seabream, Pagellus acarne
Risso, 1827), common pandora, Pagellus erythrinus (Linnaeus,758), and black seabream, Spondyliosoma cantharus (Lin-aeus, 1758).
. Materials and methods
.1. Sampling
Samples were obtained from gear selectivity studies con-ucted in south Portugal, complemented by market sampling
nd beach seining for juveniles inside a temperate coastal lagoonRia Formosa, south Portugal). Procedures and sampling detailsre described in Erzini et al. (1996, 1998, 2003) and Goncalves2000). Data collection during gear selectivity studies was made
3 ies R
fw
cmwtibtl
2
ebbOuwta
aaoeo
uOeotso
2
ifl(atmw
osowfvg
(mM
t(pte
3
rmiftvpvhbw
3
bmrtmsrtifpVtamtuvmlw
3
8 D. Abecasis et al. / Fisher
rom 1992 to 1999 using longline and gillnet, and beach seiningas used for data collection from 1999 through 2001.All fish were measured in total length (TL, to the nearest
m), weighed, and had their sex and maturity determined byacroscopic observation of the gonads. Several scales (5–6)ere removed from the area below the pectoral fin, making sure
hat they were not regeneration scales, washed and stored dry inndividually labelled envelopes. Sagittal otoliths were extractedy a transverse section on the posterior dorsal part of the head ofhe fish, cleaned in distilled water, and stored dry in individuallyabelled plastic vials.
.2. Reading procedures
The otoliths collected were read whole by immersion in glyc-rol and observed under reflected white light against a darkackground, with a compound stereoscope with amplificationsetween 10 and 40×, as described by Bagenal and Tesch (1978).tolith growth increments were counted with the anti-sulcal facep and from the nucleus towards the post-rostrum end. Readingsere carried out by three readers, independently, and only when
he three were in agreement the otolith was used on subsequentnalyses.
Regarding scales, these were placed between two glass slidesnd a slide projector was used to project the mounted scales ontoscreen. Three-independent readings of each scale were carriedut by one reader. All scales showing signs of regeneration wereliminated. After calibration, measurements and quantificationf growth increments were carried out.
The counts were converted into ages assuming the 1st of Jan-ary as the designated birthday (Williams and Bedford, 1974).toliths and scales were never read simultaneously and the read-
rs never had prior access to information on size, sex or datef capture while they were counting growth increments. Forhe marginal increment analysis 10 scales and 10 otoliths wereelected for each month and each species covering the full rangef size classes, whenever possible.
.3. Statistical analyses
Age was determined from otoliths and scales and marginalncrement analysis was used to validate annual growth incrementormation (Beamish and McFarlane, 1983). The von Berta-anffy growth function (VBGF) Lt = L∞(1 − exp[−k(t − t0)]L∞: asymptotic mean length; k: growth rate; and t0: hypotheticge at zero length), was fitted to length-at-age data obtained fromhe otolith and scale readings with least squares procedures by
eans of a non-linear regression (StatSoft Inc., 1996). VBGFsere fitted to pooled data from both scale and otolith.For the marginal increment analysis, the growth increments
f the otoliths were measured under the compound stereo-cope with an ocular micrometer and the growth incrementsf the scales measured directly on the screen where they
ere projected. The radius of the structure (R: the distancerom the focus to the margin) was measured along the dorso-entral axis, as was the distance from the focus to the lastrowth increment (Rn) and the penultimate growth increment
sTiA
esearch 89 (2008) 37–48
Rn − 1) which included both translucent and opaque zones. Theonthly mean marginal increment (MI) was then calculated byI = (R − Rn)/(Rn − Rn − 1).Statistical comparisons of growth equations between struc-
ures (otoliths and scales), were conducted using likelihood ratioLR) tests (Kimura, 1980; Hilborn and Mangel, 1997). Growtherformance indexes (Φ = log10 k + 2 log10 L∞) were calculatedo compare results obtained in this study with results publishedlsewhere (Munro and Pauly, 1983).
. Results
Overall, otoliths and scales of all seven species showed aegular growth pattern, alternating opaque and translucent incre-ents. Marginal increment analysis suggesting an annual growth
ncrement formation, validating the use of otoliths and scalesor age determination (Fig. 1). The peak of the increment variedhrough the year for each different species. For the same indi-iduals scales provided, on average, more growth incrementser years than otoliths in the following species: D. sargus, D.ulgaris, L. mormyrus and S. cantharus (Fig. 2). On the otherand, scales provided, in average, less years than otoliths in B.oops. Regarding P. erythrinus and P. acarne age estimationsere very similar for otolith and scales.
.1. B. boops
A total of 608 otoliths and 454 scales were used to investigateogue (B. boops) age and growth. Age was successfully deter-ined in 585 of the otoliths. The remaining 23 otoliths were
ejected due to the disagreement between readers or becausehey were considered unreadable. Ages were successfully deter-
ined from 425 scales, 29 were rejected either because thecales could not be read or because of disagreement betweeneadings. The range size, number of females, males and unde-ermined sex for otoliths and scales used in the analyses is shownn Table 1. The maximum observed age was 11 years (28 cm TL)or otoliths and 10 years (25 cm TL) for scales. LR tests com-aring otoliths with scales revealed significant differences in theBGF (P < 0.05). The von Bertalanffy growth curve parame-
ers estimated for bogue are presented in Fig. 3. The theoreticalsymptotic mean lengths (L∞) obtained based on ages deter-ined from otoliths (28.14 cm) and scales (26.76 cm) were less
han the maximum observed length (30.5 cm), suggesting annderestimation of L∞. Growth performance indexes (Φ) andon Bertalanffy parameters obtained by other authors are sum-arised in Table 2. The variance explained by the regression
ines (R2) was slightly higher when fitted to scales (0.91) thanhen fitted to otolith readings (0.83).
.2. D. sargus
A total of 737 white seabream (D. sargus) otoliths and 347
cales were used to determine age and growth (Fig. 4; Table 1).he otoliths proved easy to read, with clearly identifiable growthncrements and age was successfully determined in 715 otoliths.lthough double growth increments were quite common when
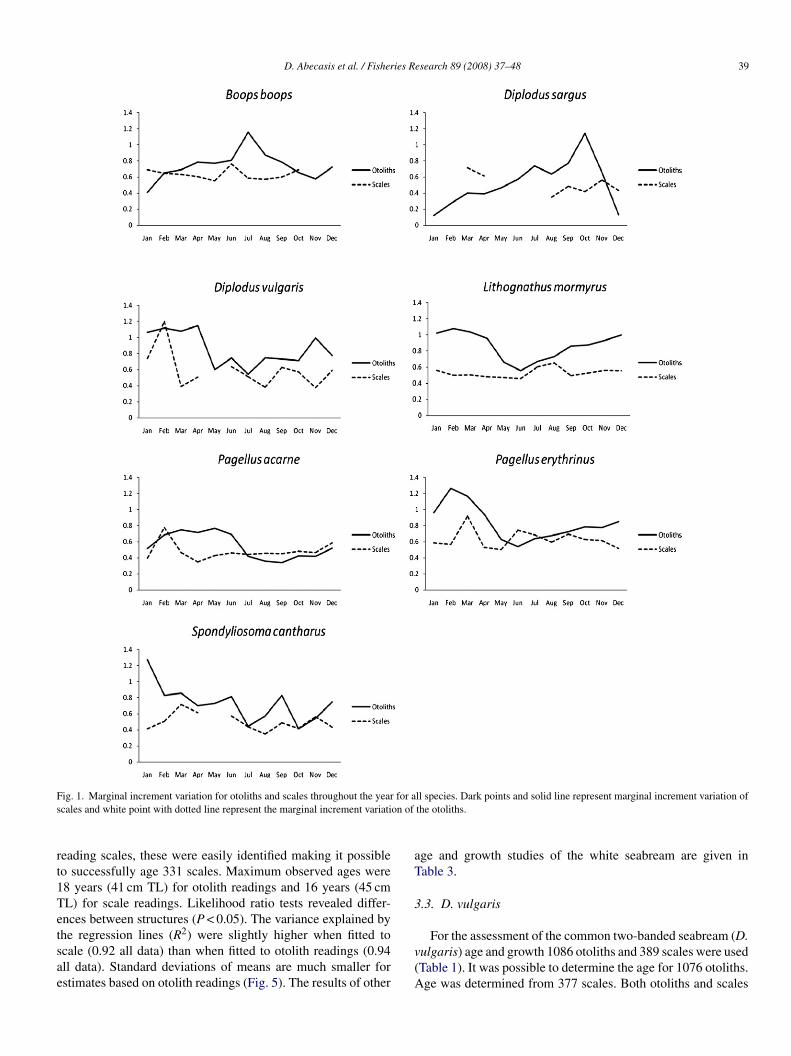
D. Abecasis et al. / Fisheries Research 89 (2008) 37–48 39
F for as on of
rt1Tetsae
aT
3
ig. 1. Marginal increment variation for otoliths and scales throughout the yearcales and white point with dotted line represent the marginal increment variati
eading scales, these were easily identified making it possibleo successfully age 331 scales. Maximum observed ages were8 years (41 cm TL) for otolith readings and 16 years (45 cmL) for scale readings. Likelihood ratio tests revealed differ-nces between structures (P < 0.05). The variance explained by
he regression lines (R2) were slightly higher when fitted tocale (0.92 all data) than when fitted to otolith readings (0.94ll data). Standard deviations of means are much smaller forstimates based on otolith readings (Fig. 5). The results of otherv(A
ll species. Dark points and solid line represent marginal increment variation ofthe otoliths.
ge and growth studies of the white seabream are given inable 3.
.3. D. vulgaris
For the assessment of the common two-banded seabream (D.ulgaris) age and growth 1086 otoliths and 389 scales were usedTable 1). It was possible to determine the age for 1076 otoliths.ge was determined from 377 scales. Both otoliths and scales
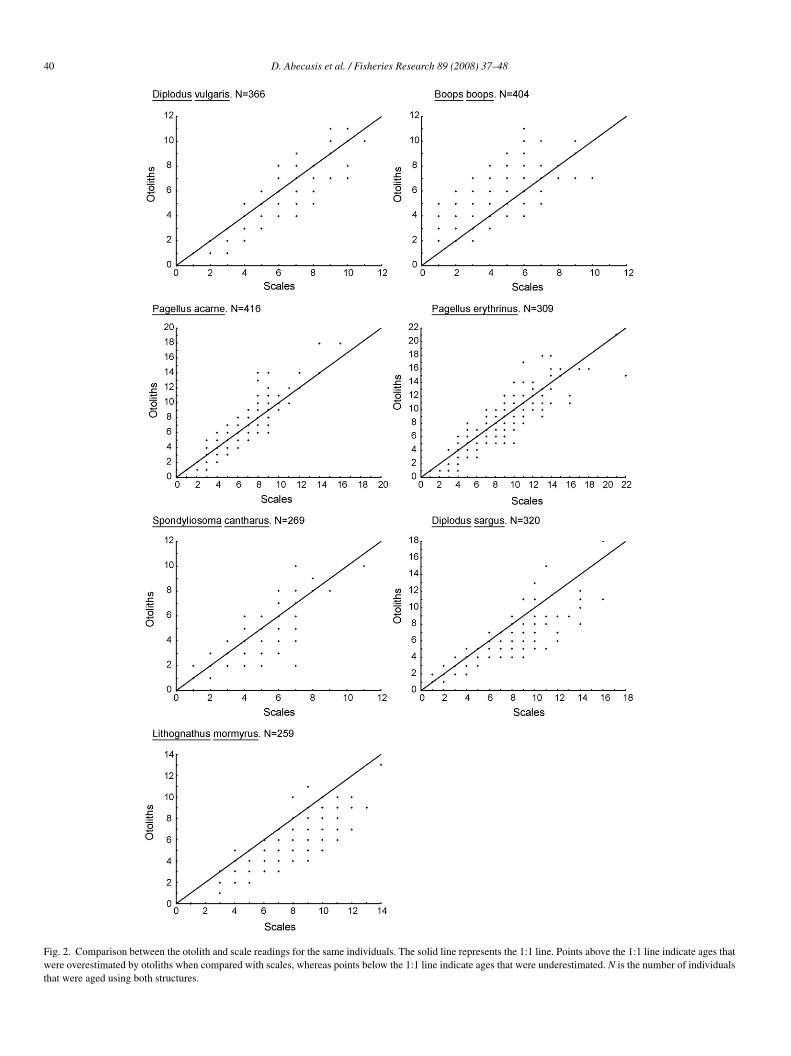
40 D. Abecasis et al. / Fisheries Research 89 (2008) 37–48
Fig. 2. Comparison between the otolith and scale readings for the same individuals. The solid line represents the 1:1 line. Points above the 1:1 line indicate ages thatwere overestimated by otoliths when compared with scales, whereas points below the 1:1 line indicate ages that were underestimated. N is the number of individualsthat were aged using both structures.
D. Abecasis et al. / Fisheries Research 89 (2008) 37–48 41
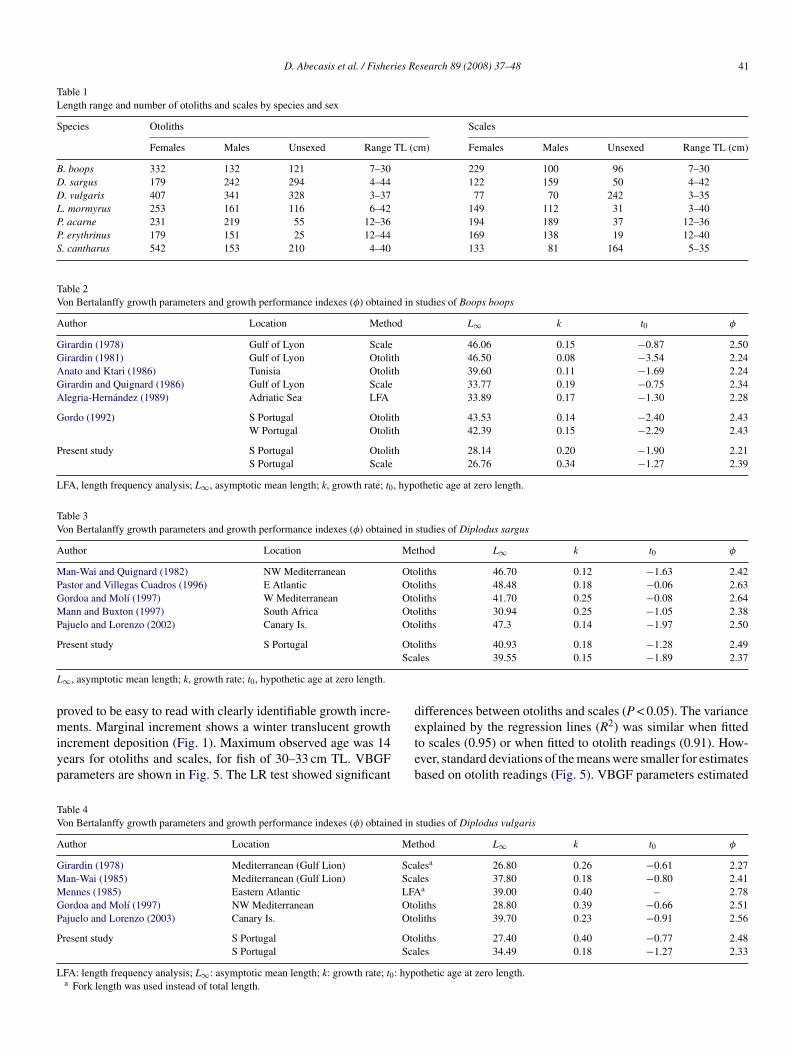
Table 1Length range and number of otoliths and scales by species and sex
Species Otoliths Scales
Females Males Unsexed Range TL (cm) Females Males Unsexed Range TL (cm)
B. boops 332 132 121 7–30 229 100 96 7–30D. sargus 179 242 294 4–44 122 159 50 4–42D. vulgaris 407 341 328 3–37 77 70 242 3–35L. mormyrus 253 161 116 6–42 149 112 31 3–40P. acarne 231 219 55 12–36 194 189 37 12–36P. erythrinus 179 151 25 12–44 169 138 19 12–40S. cantharus 542 153 210 4–40 133 81 164 5–35
Table 2Von Bertalanffy growth parameters and growth performance indexes (φ) obtained in studies of Boops boops
Author Location Method L∞ k t0 φ
Girardin (1978) Gulf of Lyon Scale 46.06 0.15 −0.87 2.50Girardin (1981) Gulf of Lyon Otolith 46.50 0.08 −3.54 2.24Anato and Ktari (1986) Tunisia Otolith 39.60 0.11 −1.69 2.24Girardin and Quignard (1986) Gulf of Lyon Scale 33.77 0.19 −0.75 2.34Alegria-Hernandez (1989) Adriatic Sea LFA 33.89 0.17 −1.30 2.28
Gordo (1992) S Portugal Otolith 43.53 0.14 −2.40 2.43W Portugal Otolith 42.39 0.15 −2.29 2.43
Present study S Portugal Otolith 28.14 0.20 −1.90 2.21S Portugal Scale 26.76 0.34 −1.27 2.39
LFA, length frequency analysis; L∞, asymptotic mean length; k, growth rate; t0, hypothetic age at zero length.
Table 3Von Bertalanffy growth parameters and growth performance indexes (φ) obtained in studies of Diplodus sargus
Author Location Method L∞ k t0 φ
Man-Wai and Quignard (1982) NW Mediterranean Otoliths 46.70 0.12 −1.63 2.42Pastor and Villegas Cuadros (1996) E Atlantic Otoliths 48.48 0.18 −0.06 2.63Gordoa and Molı (1997) W Mediterranean Otoliths 41.70 0.25 −0.08 2.64Mann and Buxton (1997) South Africa Otoliths 30.94 0.25 −1.05 2.38Pajuelo and Lorenzo (2002) Canary Is. Otoliths 47.3 0.14 −1.97 2.50
P OtoSca
L
pmiyp
d
TV
A
GMMGP
P
L
resent study S Portugal
∞, asymptotic mean length; k, growth rate; t0, hypothetic age at zero length.
roved to be easy to read with clearly identifiable growth incre-
ents. Marginal increment shows a winter translucent growthncrement deposition (Fig. 1). Maximum observed age was 14ears for otoliths and scales, for fish of 30–33 cm TL. VBGFarameters are shown in Fig. 5. The LR test showed significant
eteb
able 4on Bertalanffy growth parameters and growth performance indexes (φ) obtained in
uthor Location Me
irardin (1978) Mediterranean (Gulf Lion) Scaan-Wai (1985) Mediterranean (Gulf Lion) Scaennes (1985) Eastern Atlantic LFAordoa and Molı (1997) NW Mediterranean Otoajuelo and Lorenzo (2003) Canary Is. Oto
resent study S Portugal OtoS Portugal Sca
FA: length frequency analysis; L∞: asymptotic mean length; k: growth rate; t0: hypa Fork length was used instead of total length.
liths 40.93 0.18 −1.28 2.49les 39.55 0.15 −1.89 2.37
ifferences between otoliths and scales (P < 0.05). The variance
xplained by the regression lines (R2) was similar when fittedo scales (0.95) or when fitted to otolith readings (0.91). How-ver, standard deviations of the means were smaller for estimatesased on otolith readings (Fig. 5). VBGF parameters estimatedstudies of Diplodus vulgaris
thod L∞ k t0 φ
lesa 26.80 0.26 −0.61 2.27les 37.80 0.18 −0.80 2.41a 39.00 0.40 – 2.78
liths 28.80 0.39 −0.66 2.51liths 39.70 0.23 −0.91 2.56
liths 27.40 0.40 −0.77 2.48les 34.49 0.18 −1.27 2.33
othetic age at zero length.
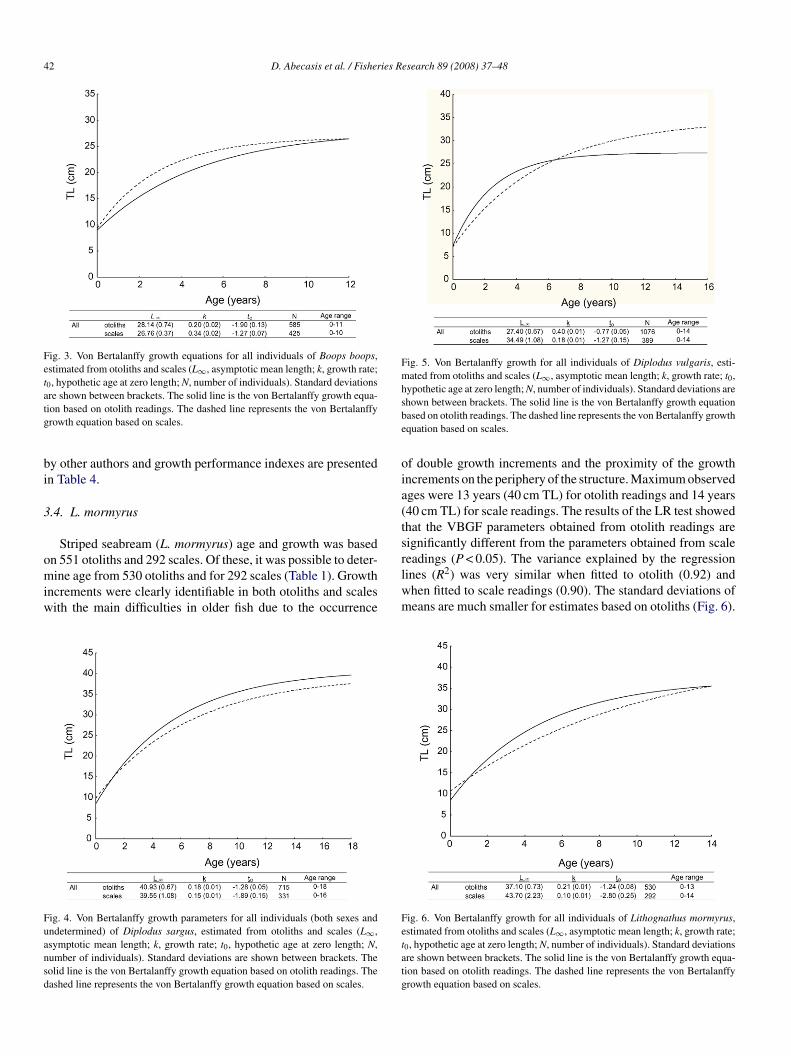
42 D. Abecasis et al. / Fisheries Research 89 (2008) 37–48
Fig. 3. Von Bertalanffy growth equations for all individuals of Boops boops,estimated from otoliths and scales (L∞, asymptotic mean length; k, growth rate;t0, hypothetic age at zero length; N, number of individuals). Standard deviationsare shown between brackets. The solid line is the von Bertalanffy growth equa-tg
bi
3
omiw
Fuansd
Fig. 5. Von Bertalanffy growth for all individuals of Diplodus vulgaris, esti-mated from otoliths and scales (L∞, asymptotic mean length; k, growth rate; t0,hypothetic age at zero length; N, number of individuals). Standard deviations aresbe
oia(ts
ion based on otolith readings. The dashed line represents the von Bertalanffyrowth equation based on scales.
y other authors and growth performance indexes are presentedn Table 4.
.4. L. mormyrus
Striped seabream (L. mormyrus) age and growth was based
n 551 otoliths and 292 scales. Of these, it was possible to deter-ine age from 530 otoliths and for 292 scales (Table 1). Growthncrements were clearly identifiable in both otoliths and scalesith the main difficulties in older fish due to the occurrence
ig. 4. Von Bertalanffy growth parameters for all individuals (both sexes andndetermined) of Diplodus sargus, estimated from otoliths and scales (L∞,symptotic mean length; k, growth rate; t0, hypothetic age at zero length; N,umber of individuals). Standard deviations are shown between brackets. Theolid line is the von Bertalanffy growth equation based on otolith readings. Theashed line represents the von Bertalanffy growth equation based on scales.
rlwm
Fetatg
hown between brackets. The solid line is the von Bertalanffy growth equationased on otolith readings. The dashed line represents the von Bertalanffy growthquation based on scales.
f double growth increments and the proximity of the growthncrements on the periphery of the structure. Maximum observedges were 13 years (40 cm TL) for otolith readings and 14 years40 cm TL) for scale readings. The results of the LR test showedhat the VBGF parameters obtained from otolith readings areignificantly different from the parameters obtained from scaleeadings (P < 0.05). The variance explained by the regression
ines (R2) was very similar when fitted to otolith (0.92) andhen fitted to scale readings (0.90). The standard deviations ofeans are much smaller for estimates based on otoliths (Fig. 6).ig. 6. Von Bertalanffy growth for all individuals of Lithognathus mormyrus,stimated from otoliths and scales (L∞, asymptotic mean length; k, growth rate;
0, hypothetic age at zero length; N, number of individuals). Standard deviationsre shown between brackets. The solid line is the von Bertalanffy growth equa-ion based on otolith readings. The dashed line represents the von Bertalanffyrowth equation based on scales.
D. Abecasis et al. / Fisheries Research 89 (2008) 37–48 43
Table 5Von Bertalanffy growth parameters and growth performance indexes (φ) obtained in studies of Lithognathus mormyrus
Author Location Method L∞ k t0 φ
Suau (1970) NW Mediterranean No data 33.30 0.28 0.06 2.48
Kraljevic et al. (1995) Northern Adriatic Battacharya 37.30 0.26 −0.39 2.56Middle Adriatic Battacharya 36.20 0.30 −0.08 2.59
Kraljevic et al. (1996) Adriatic Scales 40.10 0.20 −0.95 2.50Lorenzo et al. (2002) Canary Is. Otoliths 42.90 0.19 −1.37 2.54Kallianiotis et al. (2005) Thracean Sea Greece Otoliths 30.94 0.21 −1.00 2.30
P ths 37.10 0.21 −1.24 2.46s 43.70 0.10 −2.80 2.29
L
Ti
3
soasTirTffis
3
iAscwle
Fig. 7. Von Bertalanffy growth for all individuals of Pagellus acarne, estimatedfrom otoliths and scales (L∞, asymptotic mean length; k, growth rate; t0, hypo-thetic age at zero length; N, number of individuals). Standard deviations areshown between brackets. The solid line is the von Bertalanffy growth equationbe
iw
TV
A
A
MPP
P
L
resent study S Portugal OtoliScale
∞: asymptotic mean length; k: growth rate; t0: hypothetic age at zero length.
able 5 presents VBGF parameters and growth performancendexes estimated by other authors.
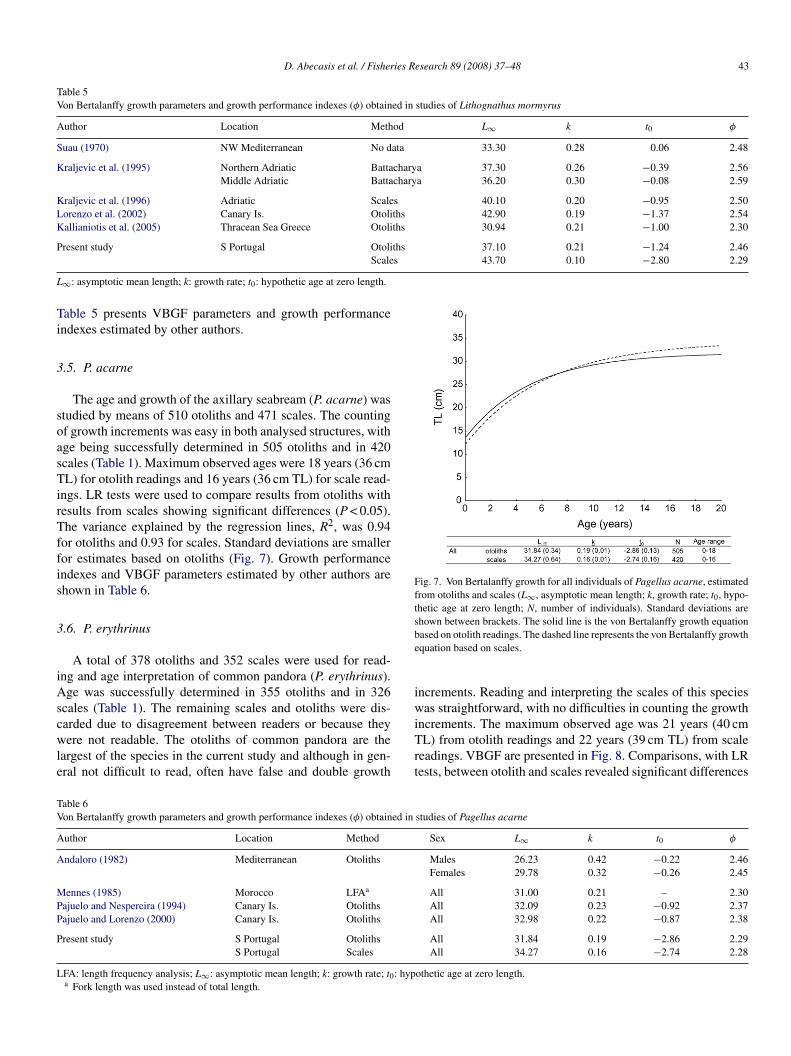
.5. P. acarne
The age and growth of the axillary seabream (P. acarne) wastudied by means of 510 otoliths and 471 scales. The countingf growth increments was easy in both analysed structures, withge being successfully determined in 505 otoliths and in 420cales (Table 1). Maximum observed ages were 18 years (36 cmL) for otolith readings and 16 years (36 cm TL) for scale read-
ngs. LR tests were used to compare results from otoliths withesults from scales showing significant differences (P < 0.05).he variance explained by the regression lines, R2, was 0.94
or otoliths and 0.93 for scales. Standard deviations are smalleror estimates based on otoliths (Fig. 7). Growth performancendexes and VBGF parameters estimated by other authors arehown in Table 6.
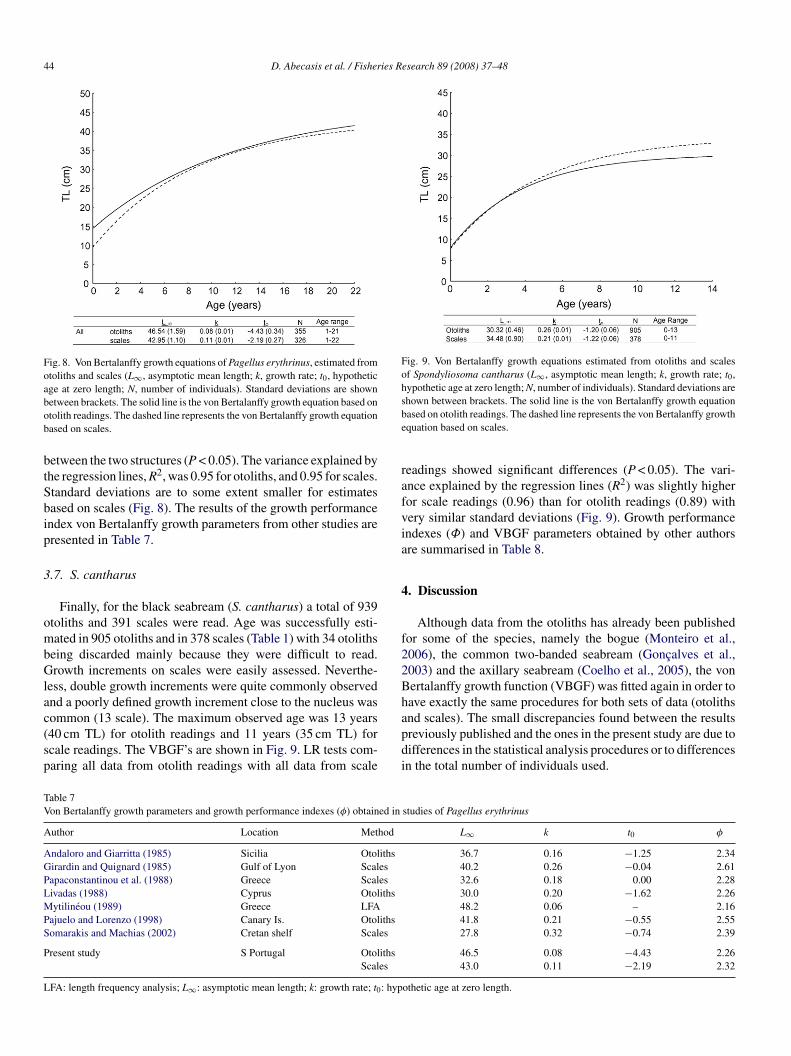
.6. P. erythrinus
A total of 378 otoliths and 352 scales were used for read-ng and age interpretation of common pandora (P. erythrinus).ge was successfully determined in 355 otoliths and in 326
cales (Table 1). The remaining scales and otoliths were dis-
arded due to disagreement between readers or because theyere not readable. The otoliths of common pandora are theargest of the species in the current study and although in gen-ral not difficult to read, often have false and double growth
iTrt
able 6on Bertalanffy growth parameters and growth performance indexes (φ) obtained in
uthor Location Method
ndaloro (1982) Mediterranean Otoliths
ennes (1985) Morocco LFAa
ajuelo and Nespereira (1994) Canary Is. Otolithsajuelo and Lorenzo (2000) Canary Is. Otoliths
resent study S Portugal OtolithsS Portugal Scales
FA: length frequency analysis; L∞: asymptotic mean length; k: growth rate; t0: hypa Fork length was used instead of total length.
ased on otolith readings. The dashed line represents the von Bertalanffy growthquation based on scales.
ncrements. Reading and interpreting the scales of this speciesas straightforward, with no difficulties in counting the growth
ncrements. The maximum observed age was 21 years (40 cm
L) from otolith readings and 22 years (39 cm TL) from scaleeadings. VBGF are presented in Fig. 8. Comparisons, with LRests, between otolith and scales revealed significant differences
studies of Pagellus acarne
Sex L∞ k t0 φ
Males 26.23 0.42 −0.22 2.46Females 29.78 0.32 −0.26 2.45
All 31.00 0.21 – 2.30All 32.09 0.23 −0.92 2.37All 32.98 0.22 −0.87 2.38
All 31.84 0.19 −2.86 2.29All 34.27 0.16 −2.74 2.28
othetic age at zero length.
44 D. Abecasis et al. / Fisheries Research 89 (2008) 37–48
Fig. 8. Von Bertalanffy growth equations of Pagellus erythrinus, estimated fromotoliths and scales (L∞, asymptotic mean length; k, growth rate; t0, hypotheticage at zero length; N, number of individuals). Standard deviations are shownbob
btSbip
3
ombGlac(sp
Fig. 9. Von Bertalanffy growth equations estimated from otoliths and scalesof Spondyliosoma cantharus (L∞, asymptotic mean length; k, growth rate; t0,hypothetic age at zero length; N, number of individuals). Standard deviations areshown between brackets. The solid line is the von Bertalanffy growth equationbe
rafvia
4
f22Bh
TV
A
AGPLMPS
P
L
etween brackets. The solid line is the von Bertalanffy growth equation based ontolith readings. The dashed line represents the von Bertalanffy growth equationased on scales.
etween the two structures (P < 0.05). The variance explained byhe regression lines, R2, was 0.95 for otoliths, and 0.95 for scales.tandard deviations are to some extent smaller for estimatesased on scales (Fig. 8). The results of the growth performancendex von Bertalanffy growth parameters from other studies areresented in Table 7.
.7. S. cantharus
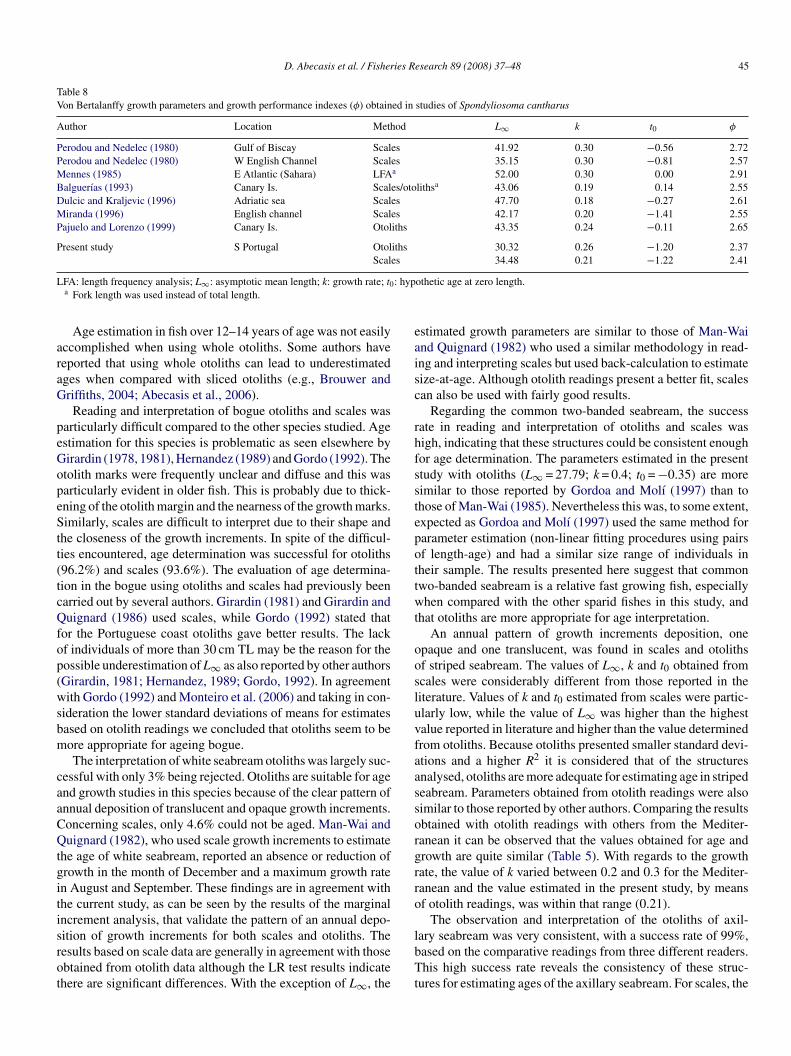
Finally, for the black seabream (S. cantharus) a total of 939toliths and 391 scales were read. Age was successfully esti-ated in 905 otoliths and in 378 scales (Table 1) with 34 otoliths
eing discarded mainly because they were difficult to read.rowth increments on scales were easily assessed. Neverthe-
ess, double growth increments were quite commonly observednd a poorly defined growth increment close to the nucleus was
ommon (13 scale). The maximum observed age was 13 years40 cm TL) for otolith readings and 11 years (35 cm TL) forcale readings. The VBGF’s are shown in Fig. 9. LR tests com-aring all data from otolith readings with all data from scaleapdi
able 7on Bertalanffy growth parameters and growth performance indexes (φ) obtained in
uthor Location Method
ndaloro and Giarritta (1985) Sicilia Otolithsirardin and Quignard (1985) Gulf of Lyon Scalesapaconstantinou et al. (1988) Greece Scalesivadas (1988) Cyprus Otolithsytilineou (1989) Greece LFA
ajuelo and Lorenzo (1998) Canary Is. Otolithsomarakis and Machias (2002) Cretan shelf Scales
resent study S Portugal OtolithsScales
FA: length frequency analysis; L∞: asymptotic mean length; k: growth rate; t0: hyp
ased on otolith readings. The dashed line represents the von Bertalanffy growthquation based on scales.
eadings showed significant differences (P < 0.05). The vari-nce explained by the regression lines (R2) was slightly higheror scale readings (0.96) than for otolith readings (0.89) withery similar standard deviations (Fig. 9). Growth performancendexes (Φ) and VBGF parameters obtained by other authorsre summarised in Table 8.
. Discussion
Although data from the otoliths has already been publishedor some of the species, namely the bogue (Monteiro et al.,006), the common two-banded seabream (Goncalves et al.,003) and the axillary seabream (Coelho et al., 2005), the vonertalanffy growth function (VBGF) was fitted again in order toave exactly the same procedures for both sets of data (otolithsnd scales). The small discrepancies found between the results
reviously published and the ones in the present study are due toifferences in the statistical analysis procedures or to differencesn the total number of individuals used.studies of Pagellus erythrinus
L∞ k t0 φ
36.7 0.16 −1.25 2.3440.2 0.26 −0.04 2.6132.6 0.18 0.00 2.2830.0 0.20 −1.62 2.2648.2 0.06 – 2.1641.8 0.21 −0.55 2.5527.8 0.32 −0.74 2.39
46.5 0.08 −4.43 2.2643.0 0.11 −2.19 2.32
othetic age at zero length.
D. Abecasis et al. / Fisheries Research 89 (2008) 37–48 45
Table 8Von Bertalanffy growth parameters and growth performance indexes (φ) obtained in studies of Spondyliosoma cantharus
Author Location Method L∞ k t0 φ
Perodou and Nedelec (1980) Gulf of Biscay Scales 41.92 0.30 −0.56 2.72Perodou and Nedelec (1980) W English Channel Scales 35.15 0.30 −0.81 2.57Mennes (1985) E Atlantic (Sahara) LFAa 52.00 0.30 0.00 2.91Balguerıas (1993) Canary Is. Scales/otolithsa 43.06 0.19 0.14 2.55Dulcic and Kraljevic (1996) Adriatic sea Scales 47.70 0.18 −0.27 2.61Miranda (1996) English channel Scales 42.17 0.20 −1.41 2.55Pajuelo and Lorenzo (1999) Canary Is. Otoliths 43.35 0.24 −0.11 2.65
Present study S Portugal Otoliths 30.32 0.26 −1.20 2.37Scales 34.48 0.21 −1.22 2.41
L : hyp
araG
peGopeStt(tcQfop(wsbm
caaCQtgitisrot
eaisc
rhfsstepottwt
oosluvfaassorgrro
FA: length frequency analysis; L∞: asymptotic mean length; k: growth rate; t0a Fork length was used instead of total length.
Age estimation in fish over 12–14 years of age was not easilyccomplished when using whole otoliths. Some authors haveeported that using whole otoliths can lead to underestimatedges when compared with sliced otoliths (e.g., Brouwer andriffiths, 2004; Abecasis et al., 2006).Reading and interpretation of bogue otoliths and scales was
articularly difficult compared to the other species studied. Agestimation for this species is problematic as seen elsewhere byirardin (1978, 1981), Hernandez (1989) and Gordo (1992). Thetolith marks were frequently unclear and diffuse and this wasarticularly evident in older fish. This is probably due to thick-ning of the otolith margin and the nearness of the growth marks.imilarly, scales are difficult to interpret due to their shape and
he closeness of the growth increments. In spite of the difficul-ies encountered, age determination was successful for otoliths96.2%) and scales (93.6%). The evaluation of age determina-ion in the bogue using otoliths and scales had previously beenarried out by several authors. Girardin (1981) and Girardin anduignard (1986) used scales, while Gordo (1992) stated that
or the Portuguese coast otoliths gave better results. The lackf individuals of more than 30 cm TL may be the reason for theossible underestimation of L∞ as also reported by other authorsGirardin, 1981; Hernandez, 1989; Gordo, 1992). In agreementith Gordo (1992) and Monteiro et al. (2006) and taking in con-
ideration the lower standard deviations of means for estimatesased on otolith readings we concluded that otoliths seem to beore appropriate for ageing bogue.The interpretation of white seabream otoliths was largely suc-
essful with only 3% being rejected. Otoliths are suitable for agend growth studies in this species because of the clear pattern ofnnual deposition of translucent and opaque growth increments.oncerning scales, only 4.6% could not be aged. Man-Wai anduignard (1982), who used scale growth increments to estimate
he age of white seabream, reported an absence or reduction ofrowth in the month of December and a maximum growth raten August and September. These findings are in agreement withhe current study, as can be seen by the results of the marginalncrement analysis, that validate the pattern of an annual depo-
ition of growth increments for both scales and otoliths. Theesults based on scale data are generally in agreement with thosebtained from otolith data although the LR test results indicatehere are significant differences. With the exception of L∞, thelbTt
othetic age at zero length.
stimated growth parameters are similar to those of Man-Waind Quignard (1982) who used a similar methodology in read-ng and interpreting scales but used back-calculation to estimateize-at-age. Although otolith readings present a better fit, scalesan also be used with fairly good results.
Regarding the common two-banded seabream, the successate in reading and interpretation of otoliths and scales wasigh, indicating that these structures could be consistent enoughor age determination. The parameters estimated in the presenttudy with otoliths (L∞ = 27.79; k = 0.4; t0 = −0.35) are moreimilar to those reported by Gordoa and Molı (1997) than tohose of Man-Wai (1985). Nevertheless this was, to some extent,xpected as Gordoa and Molı (1997) used the same method forarameter estimation (non-linear fitting procedures using pairsf length-age) and had a similar size range of individuals inheir sample. The results presented here suggest that commonwo-banded seabream is a relative fast growing fish, especiallyhen compared with the other sparid fishes in this study, and
hat otoliths are more appropriate for age interpretation.An annual pattern of growth increments deposition, one
paque and one translucent, was found in scales and otolithsf striped seabream. The values of L∞, k and t0 obtained fromcales were considerably different from those reported in theiterature. Values of k and t0 estimated from scales were partic-larly low, while the value of L∞ was higher than the highestalue reported in literature and higher than the value determinedrom otoliths. Because otoliths presented smaller standard devi-tions and a higher R2 it is considered that of the structuresnalysed, otoliths are more adequate for estimating age in stripedeabream. Parameters obtained from otolith readings were alsoimilar to those reported by other authors. Comparing the resultsbtained with otolith readings with others from the Mediter-anean it can be observed that the values obtained for age androwth are quite similar (Table 5). With regards to the growthate, the value of k varied between 0.2 and 0.3 for the Mediter-anean and the value estimated in the present study, by meansf otolith readings, was within that range (0.21).
The observation and interpretation of the otoliths of axil-
ary seabream was very consistent, with a success rate of 99%,ased on the comparative readings from three different readers.his high success rate reveals the consistency of these struc-ures for estimating ages of the axillary seabream. For scales, the

4 ies R
lhtmsTaT(oms
hFpseoQprcwtteiuo1oLLfsa1atgoctwQM
saotocaad
wsaa(satow(
5
satressaemst
A
pFdta
R
A
A
A
A
A
B
6 D. Abecasis et al. / Fisher
ower success rate of 89.2% was, however, also considered to beigh. Even though significant differences were found betweenhe growth curves estimated by otoliths and by scales, the esti-
ated parameters are very similar, which suggests that bothtructures can be used to estimate ages for the axillary seabream.he estimated k values of 0.19 for otoliths and 0.16 for scalesre lower than the ones reported by other authors (Table 6).he estimated values of L∞ of 31.84 cm (otoliths) and 34.27 cm
scales) are reasonable, given the maximum observed lengthf 36.5 cm. The smaller standard deviations and the higher R2
ay indicate that otoliths are more suitable for ageing axillaryeabream.
Regarding common pandora, both scales and otoliths hadigh readability rates (93.9% for otoliths and 92.6% for scales).ew studies on common pandora had specimens as large as theresent study (44.8 cm 18 years old). The oldest fish in thisample are apparently much older than those reported in the lit-rature. The oldest fish reported for this species was a 15-yearld female measuring 43.7 cm FL (∼50.7 cm TL) (Girardin anduignard, 1985). This could be related to difficulties in inter-reting growth marks. However, since otoliths and scales wereead independently by different observers and the results wereonsistent between the two structures, we are confident that theyere not over aged. Parameters estimated by both methods lead
o higher L∞ and lower k values than for other studies. The0 value had a relatively large absolute value and is probablyxplained by the absence of 0 age specimens. On the other hand,t was not expected that the value of t0 should be close to 0 whensing the VBGF since other authors report that the first partf the curve corresponding to juveniles is linear (Macpherson,996, in Gordoa and Molı, 1997; Monteiro, 1989). The absencef this class also explains the low k and the higher than usual∞. Pajuelo and Lorenzo (1998) who also reported a value of∞ that is higher than usual (41.78 cm) stated that these dif-erences with other authors are due to the largest individualsampled in each area. In all the literature consulted, maximumges and L∞ values are much smaller (Andaloro and Giarritta,985; Papaconstantinou et al., 1988; Livadas, 1988; Somarakisnd Machias, 2002). This may indicate that this species can growo large sizes and reach a relatively old age in southern Portu-al when compared with other regions. The comparison of agesbtained by otoliths and scales, although statistically different,ould not lead to an objective exclusion of either of these struc-ures as valid for age reading and interpretation. However, scalesere easier to read as has been noted elsewhere (Girardin anduignard, 1985; Papaconstantinou et al., 1988; Somarakis andachias, 2002).The success rates in reading otoliths and scales of black
eabream were very high. The marginal increment analysis,lthough not providing absolute validation of an annual patternf growth increment formation, suggests that the pair of opaque-ranslucent bands are deposited annually. The deposition of thepaque growth increment in otoliths in the summer months was
onfirmed by Pajuelo and Lorenzo (1999). The existence of anlternating pattern of translucent and opaque bands laid downnnually shows that both otoliths and scales can be used for ageetermination in this species.B
esearch 89 (2008) 37–48
The largest fish sampled measured approximately 40 cmhich is smaller than the maximum reported sizes from other
tudies. The maximum age in the current study was 13 years forfish of 40.8 cm while Miranda (1996), using scales, recordedmaximum age of 20 years for fish between 39 and 44 cm
mean of 41.8 cm) in length. Although the growth increments incales were not easy to count, mainly because of poorly definednd sometimes double growth increments, scales showed bet-er fittings (higher R2) and similar S.D. when compared withtoliths which made us conclude that scales are better structureshen ageing the black seabream, as also suggested by Goncalves
2000).
. General conclusions
Growth parameters were successfully estimated for all sevenpecies. In general the parameters estimated based on data forll individuals combined were reasonable. The consequence ofhe lack of juveniles in the samples of some of the species iseflected in the relatively large absolute values of the param-ter t0. Overall, one can say that otoliths are generally bettertructures to age these sparid fishes because they presentedmaller standard deviations and higher R2. Scales may be useds an alternative non-destructive technique with good results,specially for white seabream and striped seabream. Com-on pandora and black seabream were the only species where
cales revealed better results than otoliths as they are easiero read.
cknowledgments
This work was funded in part by the Commission of the Euro-ean Communities DG XIV through the project ref. 98/082:isheries biology and assessment of demersal species (Spari-ae) from the South of Portugal. The authors would also like tohank two anonymous reviewers that made valuable commentsnd suggestions.
eferences
becasis, D., Costa, A.R., Pereira, J.G., Pinho, M.R., 2006. Age and growth ofbluemouth, Helicolenus dactylopterus (Delaroche, 1809) from the Azores.Fish. Res. 79, 148–154.
legria-Hernandez, V., 1989. Study on the age and growth of Bogue (Boopsboops (L.)) from Central Adriatic Sea. Cybium 13, 281–288.
nato, C.B., Ktari, M.H., 1986. Reproduction de Boops boops (Linne, 1758) etde Sarpa salpa (Linne, 1758), Poissons, Teleosteens, Sparides du Golfe deTunis. Bull. Inst. natn. scient. tech. Oceanog. Peche Salammbo 10, 49–53.
ndaloro, F., 1982. Resume des parametres biologiques sur Pagellus acarne dela mer Tyrrhenienne meridionale et de la mer Ionienne septentrionale. FAO,Fish. Rapport. 266 (Annexe M), 89–92.
ndaloro, F., Giarritta, S.P., 1985. Contribution to the knowledge of the age,growth and feeding of pandora, Pagellus erythrinus (L. 1758) in the SicilianChannel. FAO Fish. Rep. 336 (Annex L), 85–92.
agenal, T.B., Tesch, F.W., 1978. Age and growth. In: Bagenal, T.B. (Ed.),
Methods for Assessment of Fish Production in Fresh Waters. BlackwellScientific Publications, Oxford, pp. 101–136.alguerıas, E., 1993. La pesca de los costeros canarios en el banco sahariano:antecedentes historicos y situacion actual. La chopa (Spondyliosoma can-tharus Linnaeus, 1758) como ejemplo de aplicacion de metodos simples

ies R
B
B
C
C
C
C
C
D
E
E
E
F
G
G
G
G
G
G
G
G
G
H
H
K
K
K
K
L
L
M
M
M
M
M
M
M
M
M
P
D. Abecasis et al. / Fisher
para la evaluacion de stocks en la pesquera artesanal canaria. Universidadde La Laguna, Departamento de Biologia Animal, U.D.I. Ciencias Marinas,La Laguna, PhD Thesis, 382 p.
eamish, R.J., McFarlane, G.A., 1983. Validation of age determination esti-mates: the forgotten requirement. In: Prince, E.D., Pulos, L.M. (Eds.),Proceedings of the International Workshop on Age Determination of OceanicPelagic Fishes: Tunas, Billfishes and Sharks. NOAA Technical ReportNMFS 8, U.S. Department of Commerce, Miami, pp. 29–33.
rouwer, S.L., Griffiths, M.H., 2004. Age and growth of Argyrozona argyro-zona (Pisces: Sparidae) in a marine protected area: an evaluation of methodsbased on whole otoliths, sectioned otoliths and mark-recapture. Fish. Res. 67,1–12.
ailliet, G.M., Andrews, A.H., Burton, E.J., Watters, D.L., Kline, D.E., Ferry-Graham, L.A., 2001. Age determination and validation studies of marinefishes: do deep-dwellers live longer? Exp. Gerontol. 36, 739–764.
ampana, S.E., 2001. Accuracy, precision and quality control in age determi-nation, including a review of the use and abuse of age validation methods.J. Fish Biol. 59, 197–242.
arlander, J.M., 1987. A history of scale age and growth studies of North Ameri-canfresh water fish. In: Summerfelt, R.C., Hall, G.E. (Eds.), Age and Growthof Fish. Iowa State University Press, Ames, pp. 3–14.
asselman, J.M., 1987. Determination of age and growth. In: Weatherley, A.H.,Gill, H.S. (Eds.), The Biology of Fish Growth. Academic Press, London, pp.209–242.
oelho, R., Bentes, L., Correia, C., Goncalves, J.M.S., Lino, P.G., Monteiro, P.,Ribeiro, J., Erzini, K., 2005. Age, growth and reproduction of the axillaryseabream, Pagellus acarne (Risso, 1827), from the South coast of Portugal.Thalassas 21 (1), 79–84.
ulcic, J., Kraljevic, M., 1996. Growth of the black sea bream Spondyliosomacantharus (L.) in the eastern middle Adriatic. Arch. Fish. Mar. Res. 44 (3),279–293.
rzini, K., Goncalves, J.M.S., Bentes, L., Lino, P.G., Cruz, J., 1996. Species andsize selectivity in a Portuguese multispecies artisanal longline fishery. ICESJ. Mar. Sci. 53, 811–819.
rzini, K., Goncalves, J.M.S., Bentes, L., Lino, P.G., Ribeiro, J., 1998. Speciesand size selectivity in a ‘red’ seam bream longline ‘metier’ in the Algarve(southern Portugal). Aquat. Living Res. 11, 1–11.
rzini, K., Goncalves, J.M.S., Bentes, L., Lino, P.G., Ribeiro, J., Stergiou, K.I.,2003. Quantifying the roles of competing static gears: comparative selectiv-ity of longlines and monofilament gill nets in a multi-species fishery of theAlgarve (southern Portugal). Sci. Mar. 67 (3), 341–352.
ischer, W., Schneider, M., Bauchot, M.L., 1987. Fiches FAO d’identificationdes especes pour les besoins de la peche. Mediterranee et Mer Noire (zonede peche 37) FAO, Rome.
irardin, M., 1978. Les Sparidae (Pisces, Teleostei) du Golfe du Lion—Ecologieet Biogeographie. Universite des Sciences et Techniques du Languedoc, Lab-oratoire D’Ichthyologie et de Parasitologie Generale, Montpellier, DiplomeD’Estudes Approfundies D’Ecologie Generale et Apliquee—Option Ecolo-gie Aquatique, 146 pp.
irardin, M., 1981. Pagellus erythrinus (Linnaeus, 1758) et Boops boops (Lin-naeus, 1758) Sparidae (Pisces, Sparidae) du Golfe du Lion. Ecobiologie.Prises commerciales et Modeles de gestion. Universite des Sciences etTechniques du Languedoc, Laboratoire D’Ichthyologie et de Parasitolo-gie Generale, Montpellier, These de Docteur de 3eme Cycle d’EcologieGenerale et Apliquee—Option Ecologie Aquatique, 295 pp.
irardin, M., Quignard, G.-P., 1985. Croissance de Pagellus erythrinus (Pices:Teleosteen, Sparidae) dans le Golfe du Lion. Cybium 9 (4), 359–374.
irardin, M., Quignard, G.-P., 1986. Croissance de Boops boops Linne. 1758(poissons, Sparides) dans le Golfe du Lion. J. Appl. Ichthyol. 2, 22–32.
omes, M.C., Serrao, E., Borges, M.d.F., 2001. Spatial patterns of groundfishassemblages on the continental shelf of Portugal. ICES J. Mar. Sci. 58 (3),633–647.
oncalves, J.M.S., 2000. Biologia Pesqueira e Dinamica Populacional de Diplo-
dus vulgaris (Geoffr.) e Spondyliosoma cantharus (L.) (Pisces, Sparidae) naCosta Sudoeste de Portugal. Universidade do Algarve, UCTRA, Faro, Ph.D.Thesis, 369 pp.oncalves, J.M.S., Bentes, L., Coelho, R., Correia, C., Lino, P.G., Monteiro, P.,Ribeiro, J., Erzini, K., 2003. Age and growth, maturity, mortality and yield
P
esearch 89 (2008) 37–48 47
per recruit for two banded bream (Diplodus vulgaris Geoffr.) from the southcoast of Portugal. Fish. Res. 62 (3), 349–359.
ordo, L.S., 1992. Contribuicao para o conhecimento da biologia e do estado deexploracao do stock de boga (Boops boops Linne. 1758) da costa portuguesa.Faculdade de Ciencias, Ph.D. Thesis, Lisboa, 361 pp.
ordoa, A., Molı, B., 1997. Age and growth of the sparids Diplodus vulgaris,D. sargus and D. annularis in adult populations and the differences in theirjuvenile growth patterns in the north-western Mediterranean Sea. Fish. Res.33, 123–129.
ernandez, V.A., 1989. Study on the age and growth of Bogue (Boops boops(L.)) from Central Adriatic Sea. Cybium 13 (3), 281–288.
ilborn, R., Mangel, M., 1997. The Ecological Detective Confronting Modelswith Data. Princeton University Press, New Jersey.
allianiotis, A., Torre, M., Argyri, A., 2005. Age, growth, mortality, reproduc-tion, and feeding habits of the striped seabream, Lithognathus mormyrus(Pisces: Sparidae), in the coastal waters of the Thracian Sea, Greece. Sci.Mar. 69 (3), 391–404.
imura, D.K., 1980. Likelihood methods for the von Bertalanffy growth curve.Fish. Bull. 77, 765–776.
raljevic, M., Dulcic, J., Cetinic, P., Pallaoro, A., 1996. Age, growth and mor-tality of the striped sea bream, Lithognathus mormyrus L. in the NorthernAdriatic. Fish. Res. 28, 361–370.
raljevic, M., Dulcic, J., Pallaoro, A., Cetinic, P., Jug-Dujakovic, J., 1995.Sexual maturation, age and growth of the striped sea bream, Lithognathusmormyrus L. on the eastern coast of the Adriatic sea. J. Appl. Ichthyol. 11,1–8.
ivadas, R.J., 1988. A study of the biology and population dynamics of pandora(Pagellus erythrinus L., 1758), Family Sparidae, in the sea of Cyprus. FAOFish. Rep. 412, 58–76.
orenzo, J.G., Pajuelo, J.M., Mendez-Villamil, M., Coca, J., Ramos, A.G., 2002.Age, growth, reproduction and mortality of the striped seabream, Lithog-nathus mormyrus (Pisces, Sparidae), off the Canary Islands (Central-eastAtlantic). J. Appl. Ichthyol. 18, 204–209.
ann, B.Q., Buxton, C.D., 1997. Age and growth of Diplodus sargus capensisand D. cervinus hottentotus (Sparidae) on the Tsitsikamma coast, SouthAfrica. Cybium 21 (2), 135–147.
an-Wai, R., 1985. Les sars du Golfe du Lion. Diplodus sargus, D. vulgaris, D.annularis (Pisces, Sparidae). Ecobiologie-Peche. Universite des Sciences etTechniques du Languedoc, These Docteur., 361 pp.
an-Wai, R., Quignard, J.-P., 1982. The seabream Diplodus sargus (Linne,1758) in Gulf of Lions: growth of the seabream and characteristics of land-ings from the commercial fishing grounds of Sete and Grau-du-Roi. Rev.Trav. Inst. Peches Marit. Nantes 46 (3), 173–194.
ennes, F., 1985. Multispecies assessment of fish stocks off the Western Sahararegion with emphasis on the family Sparidae. NAGA 3 (3), 5–10.
iranda, J.R., 1996. La Dorade grise en Manche: production, croissance,evaluation et analyse des parametres d’exploitation. Univesite de BretagneOccıdentale-IFREMER Diplome d’Etudes Approfondies, 30 pp.
onteiro, C.C., 1989. La faune ichthyologique de la lagune Ria Formosa (SudPortugal). Repartition et organization spatio-temporelle des communautes:aplication a l’amenagement des ressources. Univ. Sci. Technol. Languedoc,Montpellier, These Doctorat, 219 pp.
onteiro, P., Bentes, L., Coelho, R., Correia, C., Goncalves, J.M.S., Lino, P.,Ribeiro, J., Erzini, K., 2006. Age and growth, mortality, reproduction andrelative yield per recruit of the bogue, Boops boops Linne, 1758 (Sparidae),from the Algarve (south of Portugal) longline fishery. J. Appl. Ichthyol. 22,345–352.
ytilineou, C., 1989. Donnees biologiques sur le Pageot, Pagellus erythrinus,des cotes orientales de la Grece centrale. FAO Fish Rep. 412 (Annexe H),77–82.
unro, J.L., Pauly, D., 1983. A simple method for comparing growth of fishesand invertebrates. ICLARM Fishbyte 1 (1), 5–6.
ajuelo, J.G., Nespereira, J.M.L., 1994. Parametros biologicos del besugo Pag-
ellus acarne (Pisces: Sparidae) en Gran Canaria (islas Canarias). BoletinInstituto Espanol Oceanografia 10 (2), 155–164.ajuelo, J.G., Lorenzo, J.M., 1998. Population biology of the common pandoraPagellus erythrinus (Pisces: Sparidae) off the Canary Islands. Fish. Res. 36(2–3), 75–86.

4 ies R
P
P
P
P
P
P
P
R
S
S
S
S
8 D. Abecasis et al. / Fisher
ajuelo, J.G., Lorenzo, J.M., 1999. Life history of black seabream, Spondylio-soma cantharus off the Canary Islands, Central-east Atlantic. Environ. Biol.Fish. 54, 325–336.
ajuelo, J.G., Lorenzo, J.M., 2000. Reproduction, age, growth and mortal-ity of axillary seabream, Pagellus acarne (Sparidae), from the Canarianarchipelago. J. Appl. Ichthyol. 16, 41–47.
ajuelo, J.G., Lorenzo, J.M., 2002. Growth and age estimation of Diplodussargus cadenati (Sparidae) off the Canary Islands. Fish. Res. 59 (1–2),93–100.
ajuelo, J.G., Lorenzo, J.M., 2003. The growth of the common two-bandedseabream, Diplodus vulgaris (Teleostei, Sparidae), in Canarian waters, esti-mated by reading otoliths and by back-calculation. J. Appl. Ichthyol. 19,79–83.
apaconstantinou, C., Mytilineou, C., Panos, T., 1988. Aspects of the life his-
tory and fishery of Red Pandora, Pagellus erythrinus (Sparidae) off WesternGreece. Cybium 12 (4), 267–280.astor, C.M., Villegas Cuadros, M.L., 1996. Edad, crecimiento y reproduccionde Diplodus sargus Linnaeus, 1758 (Sparidae) en aguas asturianas (norte deEspana). Boletin Instituto Espanol Oceanografia 12 (1), 65–76.
W
esearch 89 (2008) 37–48
erodou, J.-B., Nedelec, D., 1980. Bilan d’explotation du stock de dorade grise.Science et Peche, Bull. Inst. Peches marit. 308, 1–7.
ibeiro, J., Bentes, L., Coelho, R., Goncalves, J.M.S., Lino, P.G., Monteiro, P.,Erzini, K., 2006. Seasonal, tidal and diurnal changes in fish assemblagesin the Ria Formosa lagoon (Portugal). Estuar. Coast. Shelf Sci. 67 (3),461–474.
omarakis, S., Machias, A., 2002. Age, growth and bathymetric distributionof red pandora (Pagellus erythrinus) on the Cretan shelf (eastern Mediter-ranean). J. Mar. Biol. Assoc. U.K. 82 (1), 149–160.
ousa, P., Azevedo, M., Gomes, M.C., 2005. Demersal assemblages off Portugal:mapping, seasonal, and temporal patterns. Fish. Res. 75, 120–137.
tatSoft, Inc., 1996. STATISTICA for Windows [Computer program manual].StatSoft, Inc., Tulsa, OK.
uau, P., 1970. Contribuition al estudio de la biologia Lithognathus
(=Pagellus) mormyrus L. (Pesces esparideos). Invest. Pesq. 34, 237–265.illiams, T., Bedford, B.C., 1974. The use of otoliths for age determination.In: Bagenal, T.B. (Ed.), The Ageing of Fish, Proceedings of an InternationalSymposium. Unwin Brothers, Surrey, pp. 114–123.




















