
Genetic strategies to determine the mode of 2n egg formation in diploid potatoes
14
Euphytica 38: 247-260 (1988) © Kluwer Academic Publishers, Dordrecht - Printed in the Netherlands Genetic strategies to determine the mode of 2n egg formation in diploid potatoes D.S. Douches and C.F. Quiros Department of Vegetable Crops" University of California, Davis, CA 95616, USA Received 28 April 1987; accepted in revised form 15 July 1987 Key words: Solanum, potato, half-tetrad analysis, isozymes, 2x-4x, megasporogenesis. Summary In this study, nineteen diploid potato clones (Solanum spp. 2n = 2x = 24) were identified as 2n egg producers on the basis of fruit set in 2x-4x crosses. The segregation of three genes mapped close to the centromere, Got-l (1.1 cM), Pgm-2 (2.0 cM), and Sdh-1 (8.3 cM), were analyzed in the tetraploid offspring in these 2x-4x crosses to discriminate between First Division Restitution (FDR) and Second Division Restitution (SDR) modes of 2n egg formation. The co-dominant nature of these markers lead to more precise estimates of the recombinational frequencies as a result of completely classifying the segregating progenies. 2x-4x data revealed a predominance of SDR mechanisms occurring in 20 of the 21 families analyzed. With a SDR mode established, half-tetrad analysis (HTA) of four distal loci, 6-Pgdh-3, Mdh-1, Pgi-l, and Aps-l, revealed two SDR segregation patterns in some of the families. One pattern fit the expectations for the distal arm position. The gene-centromere map distances based upon SDR modes in the families following this pattern, were generally close to 4x-2x (FDR) estimates suggesting similar recombina- tion rates between micro- and mega-sporogenesis. Heterozygosity transmission, on average, was 39.1 0/0. In the other segregation pattern, in which the diploid parents were derived from S. chacoense PI 230580, higher than expected homozygosity levels were found in the female 2n gametophyte populations. A post-meiotic doubling of the reduced megaspore, which generates homozygous 2n eggs, is suggested to operate in three families. The common genetic background of the diploid clones suggested a heritable nature of this mechanism. Pooled data from these three deviant families calculated that 1.80/0 of the heterozygosity was transmitted to the tetraploid progeny. It is concluded that utilization of seven enzyme-coding loci, with previously established gene-centromere map distances, in 2x-4x crosses improved half-tetrad analysis (HTA) as a means to determine the mode of2n gamete formation in megasporogenesis and megagametogenesis of diploid Solanum species. Introduction synthesize highly heterozygous clones and to pro- duce uniform true seed potato cultivars (Mendibu- Nunlerically unreduced or 2n gametes offer a ru & Peloquin, 1977; Peloquin, 1982). It is impor- unique approach to maximizing heterozygosity in tant to identify the mechanisms of 2n gamete for- polyploid species. Their discovery in diploid potato mation occurring in the diploid parent since the species (Group Tuberosum 2n = 2x = 24) has led level of heterozygosity transmitted can be extreme- to the proposal of various breeding schemes to ly different. In general, 2n gametes formed by first
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Genetic strategies to determine the mode of 2n egg formation in diploid potatoes

Euphytica 38: 247-260 (1988) © Kluwer Academic Publishers, Dordrecht - Printed in the Netherlands
Genetic strategies to determine the mode of 2n egg formation in diploid potatoes
D.S. Douches and C.F. Quiros Department of Vegetable Crops" University of California, Davis, CA 95616, USA
Received 28 April 1987; accepted in revised form 15 July 1987
Key words: Solanum, potato, half-tetrad analysis, isozymes, 2x-4x, megasporogenesis.
Summary
In this study, nineteen diploid potato clones (Solanum spp. 2n = 2x = 24) were identified as 2n egg producers on the basis of fruit set in 2x-4x crosses. The segregation of three genes mapped close to the centromere, Got-l (1.1 cM), Pgm-2 (2.0 cM), and Sdh-1 (8.3 cM), were analyzed in the tetraploid offspring in these 2x-4x crosses to discriminate between First Division Restitution (FDR) and Second Division Restitution (SDR) modes of 2n egg formation. The co-dominant nature of these markers lead to more precise estimates of the recombinational frequencies as a result of completely classifying the segregating progenies. 2x-4x data revealed a predominance of SDR mechanisms occurring in 20 of the 21 families analyzed. With a SDR mode established, half-tetrad analysis (HTA) of four distal loci, 6-Pgdh-3, Mdh-1, Pgi-l, and Aps-l, revealed two SDR segregation patterns in some of the families. One pattern fit the expectations for the distal arm position. The gene-centromere map distances based upon SDR modes in the families following this pattern, were generally close to 4x-2x (FDR) estimates suggesting similar recombination rates between micro- and mega-sporogenesis. Heterozygosity transmission, on average, was 39.10/0. In the other segregation pattern, in which the diploid parents were derived from S. chacoense PI 230580, higher than expected homozygosity levels were found in the female 2n gametophyte populations. A post-meiotic doubling of the reduced megaspore, which generates homozygous 2n eggs, is suggested to operate in three families. The common genetic background of the diploid clones suggested a heritable nature of this mechanism. Pooled data from these three deviant families calculated that 1.80/0 of the heterozygosity was transmitted to the tetraploid progeny.
It is concluded that utilization of seven enzyme-coding loci, with previously established gene-centromere map distances, in 2x-4x crosses improved half-tetrad analysis (HTA) as a means to determine the mode of2n gamete formation in megasporogenesis and megagametogenesis of diploid Solanum species.
Introduction synthesize highly heterozygous clones and to produce uniform true seed potato cultivars (Mendibu
Nunlerically unreduced or 2n gametes offer a ru & Peloquin, 1977; Peloquin, 1982). It is imporunique approach to maximizing heterozygosity in tant to identify the mechanisms of 2n gamete forpolyploid species. Their discovery in diploid potato mation occurring in the diploid parent since the species (Group Tuberosum 2n = 2x = 24) has led level of heterozygosity transmitted can be extremeto the proposal of various breeding schemes to ly different. In general, 2n gametes formed by first

248
division restitution (FDR) are considered superior to those formed by second division restitution (SDR) because, in the former, most of the heterozygosity that is present in the diploid parent is transmitted to the tetraploid progeny (Mok & Peloquin, 1975). This superiority of FDR 2n pollen has been well established in 4x-2x crosses (Quinn & Peloquin, 1973; Mok & Peloquin, 1975; Mendiburu & Peloquin, 1977; Tai & Dejong, 1980; McHale & Lauer, 1981).
Cytological identification of the meiotic mechanisms which generate diplandroids, is routine and diagnostic, hence, most of the information concerning 2n gamete mechanisms has been derived from studies of microsporogenesis (Mok & Peloquin, 1975; Ramanna, 1979; Veilleux, et al., 1982). Direct information concerning the mechanisms of 2n egg formation has been limited because of the difficulties of cytologically studying megasporogenesis. However, recently improved cytological techniques have begun to examine megasporogenesis and megagametogenesis, and mechanisms involved in diplogynoid formation (Stelly & Peloquin, 1984; Jongedijk, 1985).
As an alternative, Mendiburu and Peloquin (1979) proposed that if a chromosome marker could be identified close to its centromere, the mode of 2n egg formation could be determined through half-tetrad analysis (HTA) of the segregating marker in the progeny. This approach resulted in varying degrees of success in the potato (Ross & Langton, 1974; Mok & Peloquin, 1975; Iwanaga & Peloquin, 1979; Mok, 1981; Stelly & Peloquin, 1986). In these cases, the limitations of HTA were strongly influenced by the chromosome arm position of the segregating marker in the 2x-4x crosses. To improve HTA as a means to study diplogynoid formation, new markers closer to the centromere need to be mapped. Douches & Quiros (1986b) recently determined gene-centromere map distances for ten co-dominant electrophoretic markers in the potato. These enzyme-coding loci offer a more precise testing of the theoretical expectations for FDR and SDR segregations in 2x-4x crosses.
In this paper we report on the identification of a number of a 2n egg producing diploid clones that
were heterozygous for these markers. The segregation of three proximally positioned enzyme-coding loci were used to discriminate between FDR and SDR modes of2n egg formation through 2x-4x crosses, while the segregation of the more distal loci were also exanlined to gain insight into these mechanisms. In addition, heterozygosity levels transmitted through SDR 2n eggs were compared to theoretical expectations.
Material and methods
2x-4x crosses: The diploid clones P129-4 (S. tuberosum L. x S. berthaltii Hawkes) and P178-1 (S. tuberosum x S. tarijense Hawkes) kindly supplied by Dr. S. Jansky (North Dakota State University) and DM56-4 supplied by Dr. F.L. Haynes Jr. (North Carolina State University), were crossed to the tetraploid clones NDD277-2, Nooksack, and maintained by Dr. R.E. Voss (University of California, Davis). Additional diploid parents were species selections chosen for their ability to set seed in 2x-4x crosses. These included S. chacoense Bitt., S. kurtzianum Bitt. et Witt., S. phureja Juz. et Buk., and S. tarijense which were obtained from the Potato Introduction Station, Sturgeon Bay, Wisconsin. In 1985, ten diploid selections were identified as 2n egg producers. In 1986, three additional 2n egg producing clones were selected fronl the cross 106-2-1 x DM56-4, while four others were selected from an open pollinated berry of 111-2-1. Table 1 lists these selections.
Seed from 2x-4x crosses were treated with 1000 ppm gibberellic acid to break dormancy. Germinated seedlings were transplanted into trays (50 plants per tray). Chromosomes of off-types were counted to rule out diploids and triploids (Ramanna, 1979). In four to six weeks, leaf samples from healthy and vigorously growing seedlings were sampled for starch gel electrophoretic analysis. To assay for Aps-l, a tuber specific locus, the seedlings were allowed to tuberize in the trays. Small mature tubers were harvested in three to four months for analysis.
We assayed the diploid and tetraploid parental clones in 11 enzyme systems which revealed 14
249
enzyme-coding loci: Glutamate oxaloacetate transaminase (GOT), Phosphoglucomutase (PGM) , Peroxidase (PRX) , Triose phosphate isomerase (TPI), Diaphorase (DIA), Shikimic acid dehydrogenase (SDH) , 6-Phosphogluconic acid (6PDGH) , Malate dehydrogenase (MDH) , Phosphoglucose isomerase (PGl), Isocitrate dehydrogenase «(IDH) , and Acid phosphatase (APS) as described by Quiros & McHale (1985) and Douches & Quiros (1986a; 1986b). Gene-centromere map distances were previously estimated by Douches & Quiros (1986b), except for Tpi-l. Specific electrophoretic and enzyme staining procedures are described by Quiros (1981) and Vallejos (1983) respectively. Table 1 lists the diploid clones and the heterozygous loci identified for each individual.
Half-tetrad analysis: HTA is ideally ascertained by a locus tightly linked to its centromere (00/0 recombination frequency). The expectations ofFDR and SDR segregations in diploids are the inverse of
each other, under this model. If FDR occurs, all tetraploid progeny would be simplex heterozygotes. When SDR occurs, the progeny would be equally divided between the nulliplex and duplex classes. A mixture of the three genotypes for this locus would be found if both FDRand SDR 2n eggs were operating.
Three loci, Got-l, Pgm-2, and Sdh-l, were identified by 4x-2x (FDR) crosses to be proximal to their centromeres (Douches & Quiros, 1986b) (Table 2) hence, approaching the ideal HTA case. Observed multinomial data for these three loci resulting from 4x-2x (FDR) crosses, were used to establish probabilities for the genotypic classes, shown in Table 2. The validity of these probabilities rests upon the assumption that the gene-centromere map distances were well-estimated and that reconlbination levels are similar in both sexes. From pooled family sizes of 624, 1122, and 396 we estimated the centromeric linkages for Got-l, Pgm-2, and Sdh-l, respectively, and considered
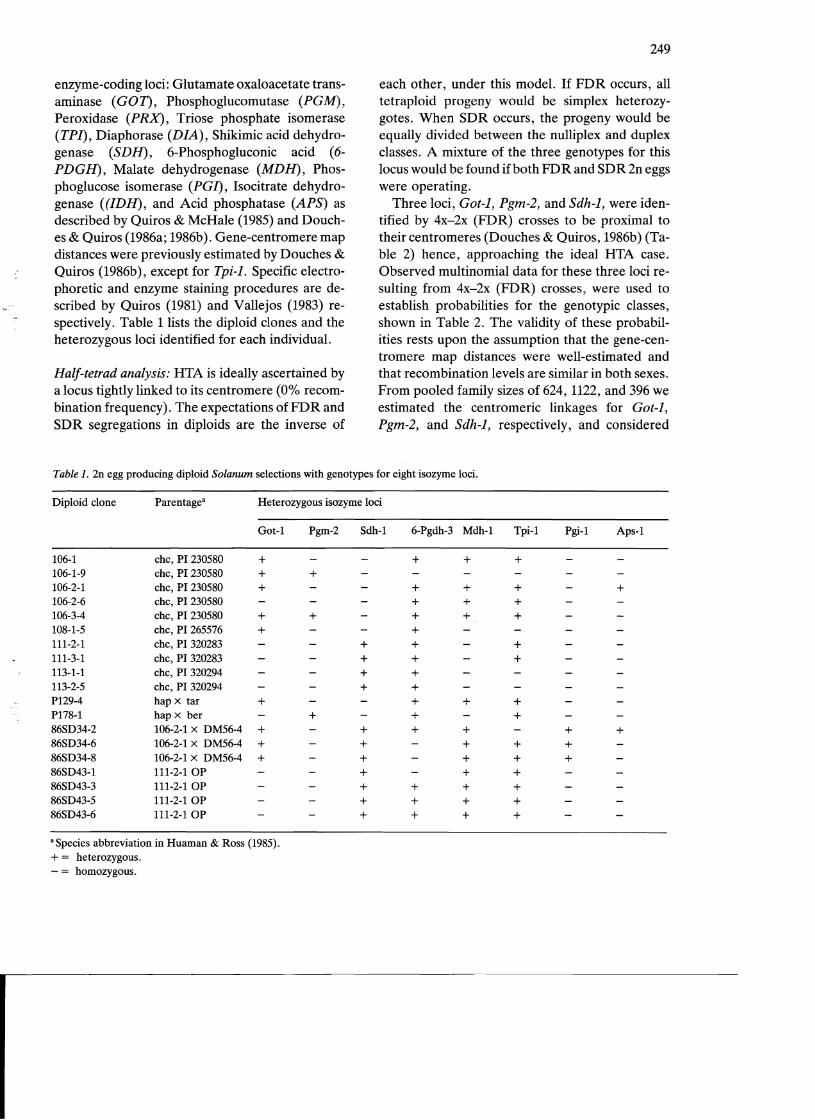
Table 1. 2n egg producing diploid Solanum selections with genotypes for eight isozyme loci.
Diploid clone Parentagea Heterozygous isozyme loci
Got-1 Pgm-2 Sdh-1 6-Pgdh-3 Mdh-l Tpi-1 Pgi-1 Aps-1
106-1 chc, PI 230580 + + + + 106-1-9 chc, PI 230580 + 106-2-1 chc, PI 230580 + + + + + 106-2-6 chc, PI 230580 + + + 106-3-4 chc, PI 230580 + + + + 108-1-5 chc, PI 265576 + + 111-2-1 chc, PI 320283 + + 111-3-1 chc, PI 320283 + + 113-1-1 chc, PI 320294 + 113-2-5 chc, PI 320294 + P129-4 hap x tar + + + + P178-1 hap x ber + + 86SD34-2 106-2-1 x DM56-4 + + + + + 86SD34-6 106-2-1 x DM56-4 + + + + 86SD34-8 106-2-1 x DM56-4 + + + + 86SD43-1 111-2-1 OP + + 86SD43-3 111-2-1 OP + + + 86SD43-5 111-2-10P + + + 86SD43-6 111-2-1 OP + + +
a Species abbreviation in Huaman & Ross (1985). + = heterozygous. - = honlozygoUs.
250
these to be unbiased estimates (Douches & Quiros, 1986b, 1988). Chi-square tests were performed to test the segregation ratios observed at these loci in 2x-4x crosses under FDR and SDR modes. For small family sizes(n~8), Baye's Rule was used as an alternative to calculate posterior probabilities for the SDR and FDR segregations from the observed data (Mendenhall, 1986).
To calculate gene-centromere map distances with an SDR mode operating, the following formula was used: Gene-centromere map distance =
.50-freq(nulliplex + duplex)/2 x 100 eM (adapted from Mendiburu and Peloquin, 1979.)
Transmission of heterozygosity: Estimates were based upon the segregation patterns of the larger families, which were divided into two subpopulations: 1) expected SDR segregation, and 2) deviant SDR segregation. The percent heterozygosity was calculated for each locus within these subpopulations. To obtain a value of average amount of heterozygosity transmitted through SDR 2n eggs, we combined the data over all loci as follows: average heterozygosity = Simplex genotype/(Nulliplex + Duplex + 8in1plex).
Results
Selection of 2n egg producing diploid clones: Ap
proximately 100 seedlings from 20 diploid potato species accessions (S. chacoense, S. kurtzianum, S. phureja, S. tarijense) and other selected clones described in materials and methods were sampled for 2n egg production using fruit set in 2x-4x crosses as a criteria for selection. Fruit set ranged from 65 to zero berries; however, fruit set was not a good indicator of seed yield (r = 0.264). In general, seed yield on a seed per fruit basis was low with a range of 5.1 to 0.1 seeds per fruit and a mean of 1.6 (Table 3).
Assays for 14 enzyme-coding loci revealed nine polymorphic loci in the selected diploid parents: Aps-l, Got-I, Idh-I, Mdh-l, 6-Pgdh-3, Pgi-I, Pgm-2, Sdh-I, and Tpi-l. When heterozygosity was ascertained for these loci, most of the respective clones were used directly for 2x-4x crosses. Of the diploid clones tested, 95% were heterozygous for at least one of the three proximally positioned loci. Got-I, Pgm-2, and Sdh-l were heterozygous, on an individual basis, in 43% , 140/0 and 520/0 of the clones, respectively. The 6-Pgdh-3 and Tpi-l loci were the most polymorphic with 67% of the diploid clones being heterozygous, while 53% were heterozygous for Mdh-l. The Pgi-I locus was heterozygous in only 140/0 of the plants, while the tuber specific locus, Aps-l, was heterozygous in two of 13 (15%) parents assayed. The Idh-llocus was heterozygous in 140/0 of the plants, but all the tetraploid parents were also heterozygotes, thus, the segregation of this locus was not amenable to HTA in the 2x-4x crosses.
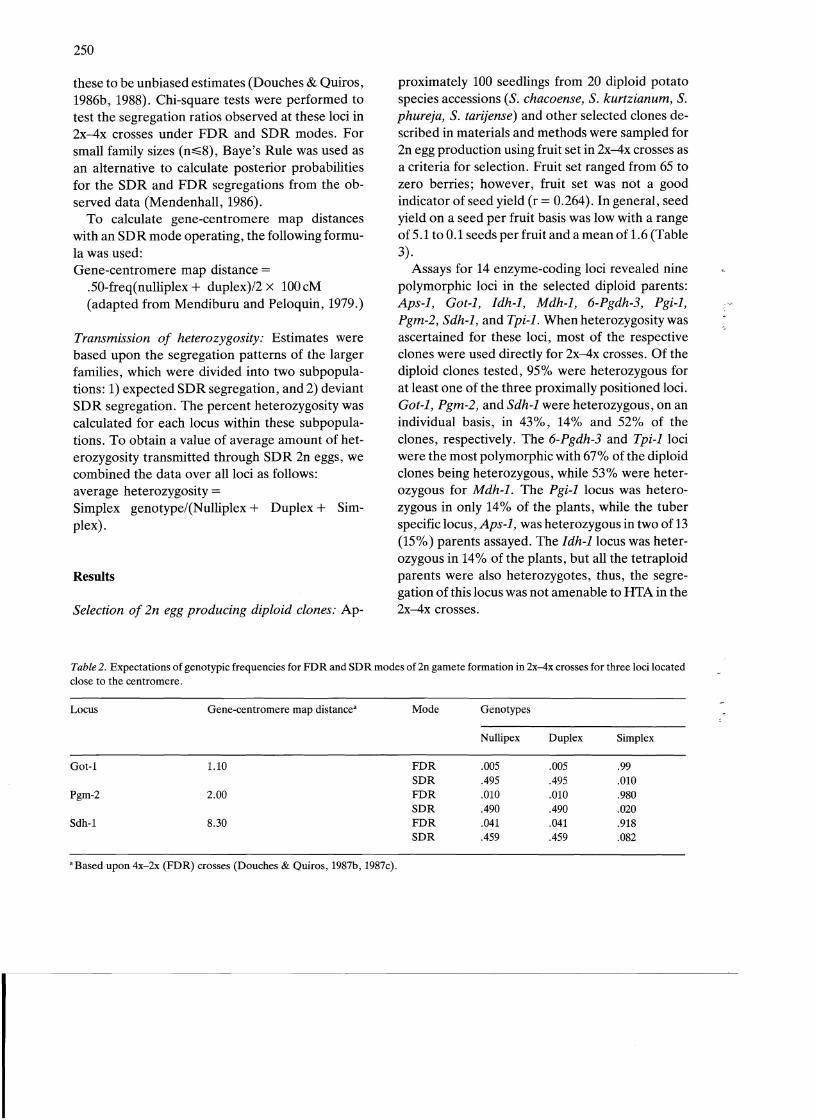
Table 2. Expectations of genotypic frequencies for FDR and SDR modes of2n ganlete formation in 2x-4x crosses for three loci located close to the centromere.
Locus Gene-centromere map distancea Mode Genotypes
Nullipex Duplex Simplex
Got-1 1.10 FDR .005 .005 .99 SDR .495 .495 .010
Pgm-2 2.00 FDR .010 .010 .980 SDR .490 .490 .020
Sdh-1 8.30 FDR .041 .041 .918 SDR .459 .459 .082
a Based upon 4x-2x (FDR) crosses (Douches & Quiros, 1987b, 1987c).
251
Segregation ofproximal loci in large families: Eight 2x-4x families were analyzed (Table 4). The Got-I, Pgm-2, and Sdh-Iloci were segregating in 4,1, and 5 of the families respectively. In addition, two families (86SD35 and 87SD4) segregated simultaneously for two of these loci. Specific triallelic conditions were found for each of these three segregating loci and are described below.
For the Pgm-2 locus (Fig. 1), a monomeric enzyme, the triallelic condition resulting after the 2x-4x cross was more ideal than the typical diallelic model. that is referenced for HTA (Mendiburu & Peloquin, 1979; Mok, 1981). Unlike HTA utilizing a dominantly controlled marker, the use of the Pgm-2 locus insured complete classification of the segregating progeny. In family 86SD35, the diploid parent was heterozygous for alleles Pgm-21 and Pgm-23, while NDD277-2 was homozygous for the Pgm-22 allele. Considering the proximity of this locus to the centromere (2.0 cM), a preponderance of three-banded phenotypes would indicate FDR, while a 1: 1 ratio of the duplex genotypes
(Pgm_21212222 and Pgm-22222323) recognized as twobanded phenotypes would manifest SDR (Fig. 1B). A comparison of chi-square probabilities for the FDR and SDR expectations demonstrated by the high frequency of homozygous gametes that 2n eggs generated by the diploid clone 106-3-4 originate via SDR (Table 4).
The Got-I locus (Fig. 2), which has the tightest linkage to a centromere (1.1 cM) of all the proximal loci tested, segregated in four 2x-4x families: 86SD35, 86SD40, 86SD41, and 87SD4 (Table 4). A triallelic segregation was found in the resulting 4x progenies. The tetraploid parent of all these crosses, NDD277-2, was heterozygous for this locus (Got-1313J414), while all four diploid parents had a heterozygous genotype Got-J4J5. The three expected genotype classes for the gametes of the diploid parents were Got-14J4, Got-J5J5, and Got-J4J5. With a common allele between the two parents, discrimination between the progeny classes was based upon the presence, absence, or dosage of the Got-15 allele. The genotype 00t-1415 would be expected if an
Table 3. Fruit set and seed yield of 2n egg producing diploids in 2x-4x crosses.
Family Cross
86SD74 P178-1 x NDD 86SD73 P129-4 x NDD 86SD77 P129-4 x Nooksack 86SD70 106-1-3 x NDD 86SD36 106-1-9 x NDD 86SD40 106-2-1 x NDD 86SD41 108-1-1 x NDD 86SD71 106-2-6 x NDD 86SD35 106-3-4 x NDD 86SD75 111-2-1 x NDD 86SD72 111-3-1 x NDD 86SD22 113-2-5 x NDD 86SD42 113-1-1 x NDD 87SDI 106-1 x NDD 87SD2 86SD34-2 x NDD 87SD3 86SD34-6 x NDD 87SD4 86SD34-8 x NDD 87SD5 86SD43-1 x NDD 87SD6 86SD43-3 x NDD 87SD8 86SD43-5 x NDD 87SD9 86SD43-6 x NDD
Fruit Seed SeedlFruit
20 9 0.45 20 8 0.40 3 2 0.67
33 3 0.09 12 4 0.33 55 195 3.55 4 12 3.00 4 14 3.50
18 46 2.56 65 14 0.22 15 7 0.47 NA 13+ NA NA 76+ NA 6 7 1.17 5 6 1.20
17 17 1.00 23 60 2.61 5 6 1.20
16 13 0.81 9 18 2.00
16 81 5.06
NDD = NDD277-2; NA = Not available; mean = 1.6 SeedslFruit; Standard error = 0.32; r = 0.514a•
a Correlation between fruit set and seed set.
252
FDR mode of 2n egg formation was occurring, while a SDR segregation would give in an equal proportion of Got-J4J4- and Got-J5J5_genotypes. Chi-square tests for fit to FDR expectations have highly significant deviations (P~.OOOl). Meanwhile, chi-square tests for fit to SDR expectations did not deviate significantly in three of the four families. The lack of fit for the family 86SD40 was due to an excess of the Got-J5J5_genotype in comparison to the Got-J4J4_genotype (Fig. 2B). Hence, SDR was concluded to occur in these clones. Heterogeneity between these four families precluded pooling segregation data for the Got-Jlocus, how
ever, the lack of the Got-J4J5_genotype (none in 254 offspring) also reaffirmed a tight centromeric linkage for the Got-Jlocus.
Five 2x-4x families (86SD22, 86SD42, 86SD42, 87SD4, 87SD8, and 87SD9) segregated for the Sdh-J locus (8.3cM) (Fig. 3). In all crosses, NDD277-2 was used as the pollen parent. The heterozygous nature of the Sdh-Jlocus (Sdh-J2J2]5J5) in the tester parent did not hinder out ability to discriminate between the homozygous and heterozygous gametes produced by the diploid parents. Each family ratio fit SDR expectations while at the same time deviating significantly from FDR expec-
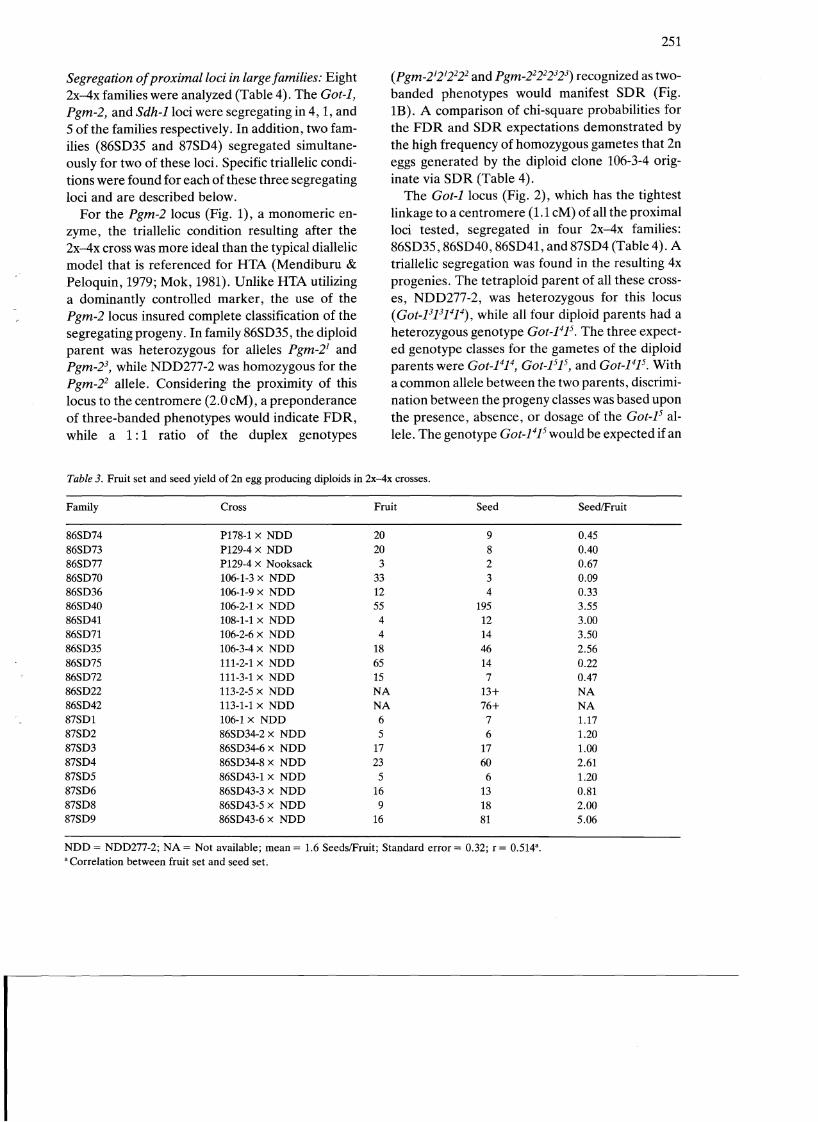
Table 4. Segregation of various isozyme markers in 2x-4x crosses in large families and its applications to determine the mode of 2n egg formation and gene-centromere map distances.
Family Diploid Genotype 4x-2x Genotypesb Prob Prob Mode of Map parent (FDR) Map _ SORc FOR 2n egg distance
distancea Nulliplex Duplex Simplex formation
86SD22 113-2-5 Sdh-1113 8.3 19 23 o 0.91 0.00 SDR 0.00 6-pgdh-3132 30.1 16 13 49 30.9
86SD41 108-1-5 Got-1315 1.1 3 6 o 0.82 0.00 SOR 0.00 6-pgdh-3132 30.1 2 o 7 38.9
86SD42 113-1-1 Sdh-1113 8.3 7 6 o 1.00 0.00 SDR 0.00 6-pgdh-3132 30.1 2 1 13 40.6
87SD8 86SD43-5 Sdh-1113 8.3 9 3 o 0.40 0.00 SDR 0.00 6-pgdh-3132 30.1 7 8 22 29.7 Mdh-1316 33.5 5 8 23 31.9
87SD9 86SD43-6 Sdh-1131 8.3 21 15 o 0.73 0.00 SDR 0.00 6-pgdh-3132 30.1 7 8 22 29.7 Mdh-1316 33.5 5 8 23 31.9
86SD40 106-2-1 Got-1315 1.1 65 118 o 0.002 0.00 SDR 0.00 6-pgdh-3132 30.1 109 58 4 1.2 Mdh-1315 33.5 86 85 3 0.9 Tpi-1113 98 81 4 1.1 Aps-1112 13.4 65 51 2 1.1
86SD35 106-3-4 Pgm-2123 2.0 19 12 o 0.60 0.00 SDR 0.00 Got-1315 1.1 12 14 o 0.99 0.00 SDR 0.00 6-pgdh-3132 30.1 17 11 2 3.3 Mdh-1315 33.5 9 6 1 1.9 Tpi-1112 7 17 2 3.8
87SD4 86SD34-8 Got_1315 1.1 12 24 o 0.20 0.00 SDR 0.00 Sdh-1113 8.3 15 21 o 0.73 0.00 SDR 0.00 Pgi-1213 26.0 13 23 o 0.00 Mdh-1315 33.5 23 13d 0.00 Tpi-1112 19 14 3 4.17
a Obtained from Douches & Quiros (1986b; 1988c); b Nulliplex and duplex refer to genotypes fornled from homozygous 2n eggs; simplex refer to genotypes formed from heterozygous 2n eggs; c Probabilities based upon chi-square tests; d Simplex and duplex classes were pooled.
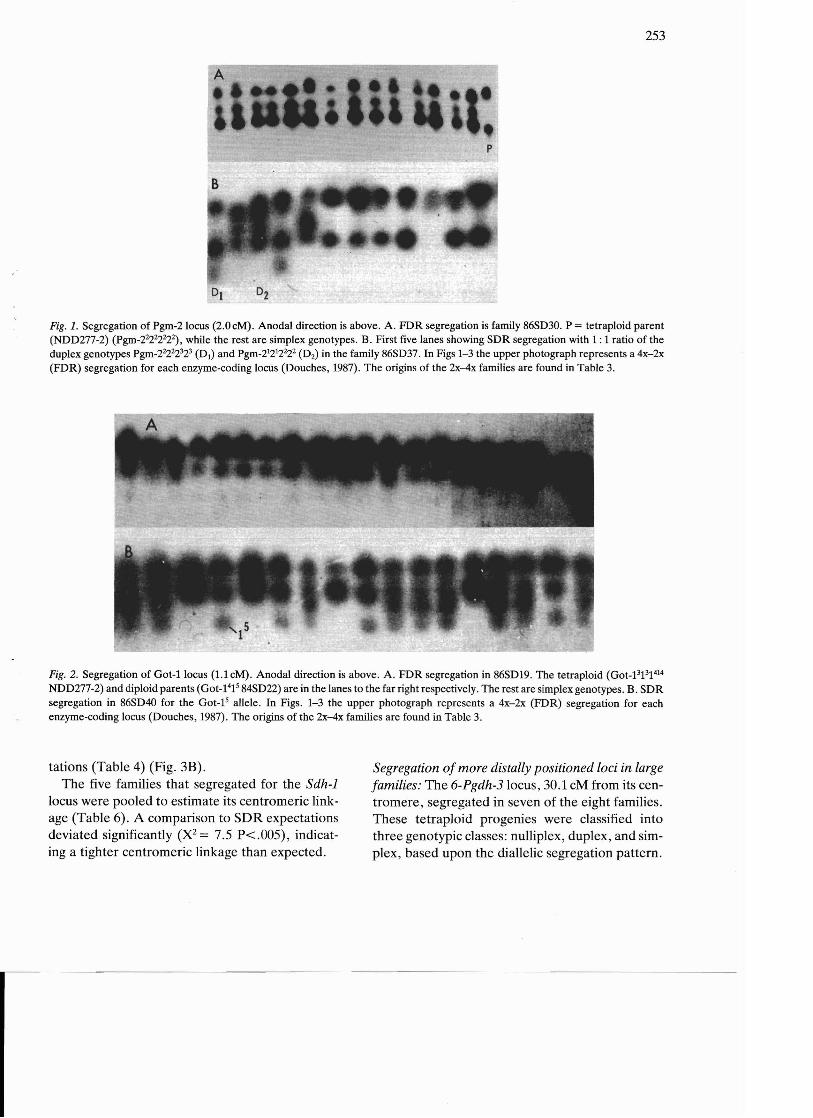
253
p
Fig. 1. Segregation of Pgm-2locus (2.0 cM). Anodal direction is above. A. FDR segregation is family 86SD30. P = tetraploid parent (NDD277-2) (Pgm_2~22222), while the rest are simplex genotypes. B. First five lanes showing SDR segregation with 1: 1 ratio of the duplex genotypes Pgm_2~22323 (D1) and Pgm_21212222(D2) in the family 86SD37. In Figs 1-3 the upper photograph represents a 4x-2x (FDR) segregation for each enzyme-coding locus (Douches, 1987). The origins of the 2x-4x families are found in Table 3.
Fig. 2. Segregation of Got-1 locus (1.1 cM). Anodal direction is above. A. FDR segregation in 86SD19. The tetraploid (Got_13131414 NDD277-2) and diploid parents (Got-1415 84SD22) are in the lanes to the far right respectively. The rest are simplex genotypes. B. SDR segregation in 86SD40 for the Got-15 allele. In Figs. 1-3 the upper photograph represents a 4x-2x (FDR) segregation for each enzyme-coding locus (Douches, 1987). The origins of the 2x-4x families are found in Table 3.
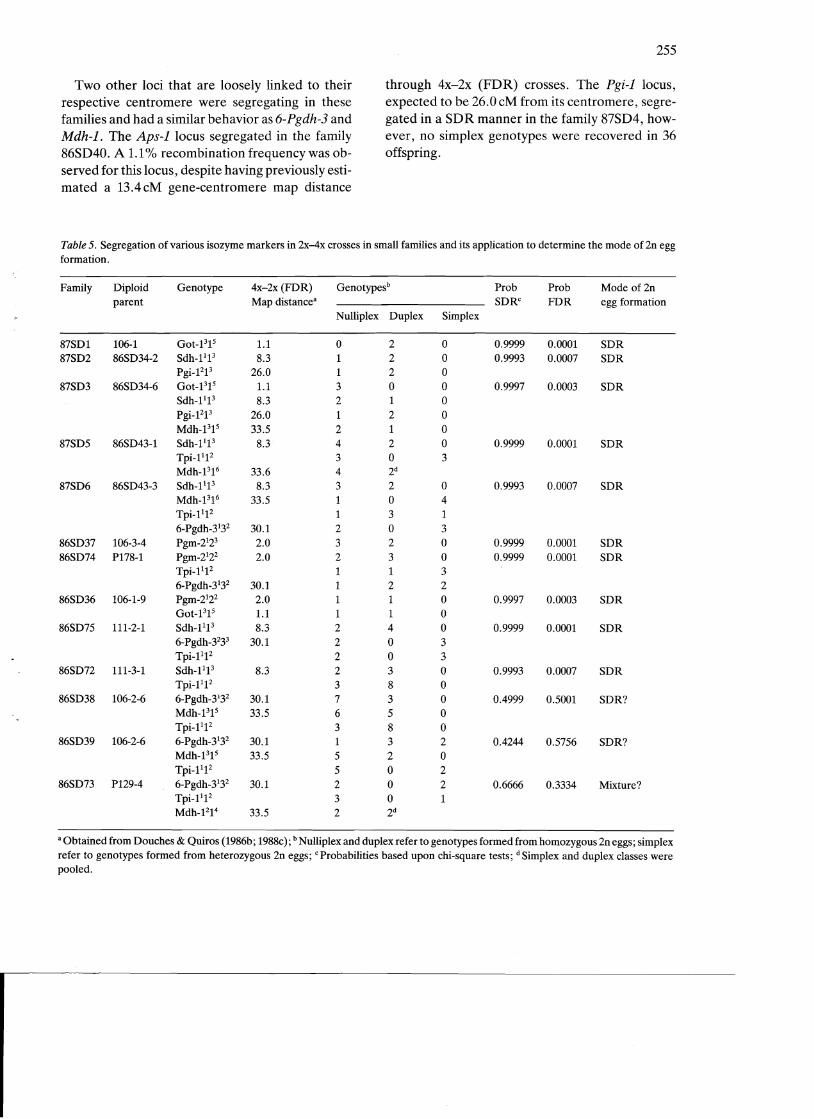
tations (Table 4) (Fig. 3B). Segregation of more distally positioned loci in large The five families that segregated for the Sdh-l families: The 6-Pgdh-310cus, 30.1 cM from its cen
locus were pooled to estimate its centromeric link tromere, segregated in seven of the eight families. age (Table 6). A comparison to SDR expectations These tetraploid progenies were classified into deviated significantly (X2 = 7.5 P<.005), indicat three genotypic classes: nulliplex, duplex, and siming a tighter centromeric linkage than expected. plex, based upon the diallelic segregation pattern.

254
.: ."~F:. l'
. '~ '. .. .••.
~
-..'
" ..
Fig. 3. Segregation of Sdh-l locus (8.3 cM). Anodal direction is above. A. FDR segregation in 85SD41. R denotes recombinant offspring. B. SDR segregation in 87SD8. Note the 1: 1segregationofSdh-13 allozyme band (slowest migrating). Tdenotes a Sdh-13131215
genotype, while D is an example of Sdh-1215-phenotype. In Figs. 1-3 the upper photograph represents a 4x-2x (FDR) segregation for each enzyme-coding locus (Douches, 1987). The origins of the 2x-4x families are found in Table 3.
Two types of segregation patterns were observed for this locus: 1) a SDR segregation typical for its distal arm position, and 2) SDR segregation as if it was proximally positioned. The first segregation pattern was observed in families 86SD22, 86SD41, 86SD42, 87SD8, and 87SD9, which are known to produce 2n eggs by SDR as suggested by the segregation of the proximal loci (Table 4). A pooled estimate of 152 progeny allowed us to detect a 30.9 cM gene-centromere distance for 6-Pgdh-3, based on SDR expectations (Table 6). This value was not significant from the distance determined by 4x-2x (FDR) crosses (X2 = 0.177 P = .90). When using the 6-Pgdh-3 locus to attempt to predict the mode of 2n egg formation, the segregation data also marginally fit FDR (X2 = 5.08 P = .08). In the families 86SD35 and 86SD40, this locus segregated in a SDR pattern typical of proximal position. Hence, the data fit poorly FDR and SDR expectations assuming normal recombination levels. A gene-centromere recombination frequency, based upon SDR, was calculated to be 1.5% in the two deviant families (Table 6).
The Mdh-llocus, which is 33.5 cM from its centromere, segregated in five 2x-4x families (Table 4). Two new Mdh-l alleles were identified in the diploid parents of these crosses. The clones 106-2-1 and 106-3-4 carried the Mdh-P allele, while 86SD43-5 and 86SD43-6 carried the Mdh-1 6 allele
(Fig. 4). The upper band of the two-banded Mdh-P allele migrated more slowly than the upper band of Mdh-J3, while the upper band of the two-banded Mdh-1 6 allele migrated further than the upper band of the Mdh-13 allele. NDD277-2 was homozygous for the Mdh-F allele, which overlaps with the slower-migrating band of Mdh-13 , thus a triallelic segregation pattern was observed in the tetraploid progenies. Analysis of the progenies for this locus was similar to the reasoning used for the Pgm-2 locus. The gametic types were completely classified on the presence and absence of Mdh-l alleles in the four families generated from these parents.
The segregation patterns of Mdh-l for these families were separated into two groups in the same manner as that for 6-Pgdh-3. The Mdh-l locus in the pooled data from the two families, 87SD8 and 87SD9, segregated according to SDR expectations. A 29.2 cM centromeric distance was estimated from a total progeny size of 48 (Table 6). This estimate was not significantly different from those obtained by 4x-2x (FDR) crosses. The segregation of the Mdh-l locus (33.5 cM) could not discriminate between the two modes since expectations are practically the same for both FDR and SDR (X2 =
2.5 P = .33). In contrast, from the pooled segregation data of progenies 86SD40 and 86SD35, a 1.1% recombination rate was observed for the Mdh-l-centromere chromosome segment on the basis of SDR (Table 6).
255
Two other loci that are loosely linked to their through 4x-2x (FDR) crosses. The Pgi-l locus, respective centromere were segregating in these expected to be 26.0 cM from its centromere, segrefamilies and had a similar behavior as 6-Pgdh-3 and gated in a SDR manner in the family 87SD4, howMdh-l. The Aps-l locus segregated in the family ever, no simplex genotypes were recovered in 36 86SD40. A 1.1010 recombination frequency was ob offspring. served for this locus, despite having previously estimated a 13.4cM gene-centromere map distance
Table 5. Segregation of various isozyme markers in 2x-4x crosses in small families and its application to determine the mode of 2n egg formation.
Family Diploid Genotype 4x-2x (FDR) Genotypesb Prob Prob Mode of2n parent Map distancea SDRc FDR egg formation
Nulliplex Duplex Simplex
87SD1 106-1 Got-1315 1.1 0 2 0 0.9999 0.0001 SDR 87SD2 86SD34-2 Sdh-1113 8.3 1 2 0 0.9993 0.0007 SDR
Pgi-1213 26.0 1 2 0 87SD3 86SD34-6 Got-1315 1.1 3 0 0 0.9997 0.0003 SDR
Sdh-1113 8.3 2 1 0 Pgi-1213 26.0 1 2 0 Mdh-1315 33.5 2 1 0
87SD5 86SD43-1 Sdh-1113 8.3 4 2 0 0.9999 0.0001 SDR Tpi-1112 3 0 3 Mdh-1316 33.6 4 2d
87SD6 86SD43-3 Sdh-1113 8.3 3 2 0 0.9993 0.0007 SDR Mdh-1316 33.5 1 0 4 Tpi-1112 1 3 1 6-Pgdh-3132 30.1 2 0 3
86SD37 106-3-4 Pgm-2123 2.0 3 2 0 0.9999 0.0001 SDR 86SD74 P178-1 Pgm-2122 2.0 2 3 0 0.9999 0.0001 SDR
Tpi-1112 1 1 3 6-Pgdh-3132 30.1 1 2 2
86SD36 106-1-9 Pgm-2122 2.0 1 1 0 0.9997 0.0003 SDR Got-1315 1.1 1 1 0
86SD75 111-2-1 Sdh-1113 8.3 2 4 0 0.9999 0.0001 SDR 6-Pgdh-3233 30.1 2 0 3 Tpi-1112 2 0 3
86SD72 111-3-1 Sdh-1113 8.3 2 3 0 0.9993 0.0007 SDR Tpi-1112 3 8 0
86SD38 106-2-6 6-Pgdh-JI.32 30.1 7 3 0 0.4999 0.5001 SDR? Mdh-1315 33.5 6 5 0 Tpi-1112 3 8 0
86SD39 106-2-6 6-Pgdh-3132 30.1 1 3 2 0.4244 0.5756 SDR? Mdh-1315 33.5 5 2 0 Tpi-1112 5 0 2
86SD73 P129-4 6-Pgdh-3132 30.1 2 0 2 0.6666 0.3334 Mixture? Tpi-1112 3 0 1 Mdh-1214 33.5 2 2d
a Obtained from Douches & Quiros (1986b; 1988c); b Nulliplex and duplex refer to genotypes formed from homozygous 2n eggs; simplex refer to genotypes formed from heterozygous 2n eggs; C Probabilities based upon chi-square tests; d Simplex and duplex classes were pooled.
256
HTA ofsparse family data: Thirteen 2x-4x families gations based upon these two loci could not disof small size were analyzed using Bayes' Rule (Ta criminate between a SDR and FDR TIl0de in the ble 5). Three families each segregated for the families 86SD38 and 86SD39 (P = .4999 and .4244 Pgm-2 and Got-l loci respectively, one of these respectively) . families segregating for both (86SD36). The Sdh-l Pooled family data for the 6-Pgdh-3, Pgi-l, and locus segregated in six of the families while one Mdh-lloci revealed a progeny distribution through segregating for both Got-l and Sdh-l (87SD3). the three genotypic classes (Table 5). The progeny
In ten of the 13 families, the discrimination be sizes were too small to calculate accurate genetween SDR and FDR modes was straightforward centromere values, howeve~, two families, 87SD2 and sinlple. All these probability values were based and 87SD3, were unique in that they lacked simupon the segregation of at least one of the three plex genotypes for the 2x-4x segregations of the proximally positioned loci. A SDR mode was con Pgi-l and Mdh-lloci. cluded for these ten families with the data summarized in Table 5. Gene-centromere relationship ofTpi-llocus: Segre
The diploid parent, 106-2-6, involved in families gation of the Tpi-llocus in six 2x-4x families gave 86SD38 and 86SD39, was homozygous for the some insight into the centromeric relationship of three proximal loci, but was heterozygous for the this previously unreported enzyme-coding locus distal loci 6-Pgdh-3 and Mdh-l. The 2x-4x segre- (Fig. 5). The pooled data from 86SD72, 86SD75,
Table 6. Heterozygosity transmission through 2x-4x crosses and comparison of gene-centromere distances estimated by 4x-2x and 2x-4x crosses.
a) Normal2x-4x families
Locus Number of Nulliplex Duplex Sinlplex Percent (SDR) 2x-4x 4x-2x (FDR) families heterozygosity Map distance Map distancea
Got-1 1 3 6 0 0.0 0.0 1.1 Sdh-1 4 56 47 0 0.0 0.0 8.3 6-Pgdh-3 5 30 28 92 61.3 30.9 30.1 Mdh-1 2 8 12 28 58.3 29.2 33.5 Tpi-1 1 4 2 6 50.0 25.0 NA
Total 101 95 126 39.1
b) Deviant 2x-4x families (86SD40, 86SD35, 87SD4)
Locus Number of Nulliplex Duplex Simplex Percent (SDR) 2x-4x 4x-2x (FDR) families heterozygosity Map distance Map distancea
Got-1 3 89 156 0 0.0 0.0 1.1 ,,Pgm-2 1 19 12 0 0.0 0.0 2.0 Sdh-1 1 15 21 0 0.0 0.0 8.3 Aps-1 1 65 51 2 1.1 0.6 13.4 Pgi-1 1 13 23 0 0.0 0.0 26.0 6-Pgdh-3 2 126 69 6 3.0 1.5 30.1 Mdh-1 2 118 114 4 1.7 0.9 33.5 Tpi-1 3 124 112 9 3.7 1.9 NA
Total 569 558 21 1.83
a Obtained from Douches & Quiros (1986b, 1988c). NA = not available.

257
Fig. 4. SDR segregation for Mdh-l locus (33.5 cM). Anodal direction is above. A. Progeny from 86SD40 segregating 1: 1 for Mdh_12121515 (M) and Mdh_12121313 (M2 genotypes). B. Progeny from 87SD9 segregating in a SDR manner. S, T, and F denote Mdh_12121313, Mdh_12121316, and Mdh_12121616 genotypes respectively.
N 0"
Fig. 5. SDR segregation for Tpi-llocus in 87SD4. D and N denote the Tpi_11111212 and Tpi_12121212 genotypes respectively.
86SD74, 87SD5, 87SD6, and 87SD8 suggests a more distal chromosome arm position based upon a SDR mode of 2n egg formation, however, the small total progeny size of 33 precluded any precise chromosome arm location. Tpi-l also segregated in the three deviant families 86SD35, 86SD40, and 87SD4. Pooled data for the Tpi-llocus had a segregation pattern that was similar to the other distal loci in these deviant families. Few simplex genotypes were observed. Pooled data of 255 offspring estimated a 1.8% recombination frequency on the basis of SDR.
Heterozygosity transmission through 2x-4x crosses: Estimates of gene-centromere map distances are simply another way of expressing the recombination frequency for that chromosome segment. The 2x-4x segregation data from this study offered an opportunity to examine the ability of the female 2n gametophyte to transmit its heterozygosity through an SDR mechanism. This analysis was limited to
the large 2x-4x families. The isozyme data were subdivided into two gametic populations for this analysis: normal recombination vs. deviant SDR segregations (Table 6).
The gametic population in which the large families showed normal recombination levels was believed to be the least biased population since most centromeric linkages were within expectations. The SDR gametes transmitted on average 39.1% of their heterozygosity (Table 6). The other large families (86SD35, 86SD40, and 87SD4) which showed abnormal SDR segregations had a much different gametic population structure. For the eight chromosome segments sampled, only 1.83% of the heterozygosity was transmitted on average, indicating an almost completely inbred population of 2n gametes. Almost all the heterozygosity was found between rather than within the gametes.

258
Discussion
We conclude from the segregation of these isozyme loci that SDR is more prevalent than FDR in 2n egg formation. Similar conclusions were drawn by Stelly & Peloquin (1986) when a different population of diplogynoid producing parents was analyzed in 2x-4x crosses. Prevalence of SDR mechanisms is predicted as a result of sequential type of embryo sac development that the female gametophyte follows in Solanum (Jongedijk, 1985). In alfalfa, which has a sinlilar embryo sac development, SDR 2n eggs also have been identified cytologically (Pfeiffer & Bingham, 1983). FDR 2n eggs have been previously identified through HTA in potato (Ross & Langton, 1974; Mok & Peloquin, 1976; Iwanaga & Peloquin, 1979; Mok, 1981), however in these instances, the frequency was very low or in combination with a SDR nlechanism. A FDR mechanism comparable to the parallel/fused spindles during nletaphase II in meiosis during microsporogenesis is not feasible in megasporogenesis because of successive divisions associated with its development. Hence, a reduction in chromosome pairing is a prerequisite for FDR 2n egg formation through abnormal megasporogenesis (Jongedijk, 1985). Under normal synaptic conditons as seen in this study, SDR 2n eggs should prevail.
Five of the eight large 2x-4x families segregated in an expected manner. A SDR mechanism such as second division omission or premature cytokinesis could account for the observed results. In these families, recombination levels were generally close to expectations extrapolated from 4x-2x (FDR) data, suggesting similar recombination levels in micro- and megasporogenesis. Gene-centromere map distance estimates were within expectations for Pgm-2, Got-l, 6-Pgdh-3, and Mdh-l, while the Sdh-llocus in megasporogenesis showed a slightly tighter linkage than expected (0.0 vs. 8.3 eM).
HTA of the more distal loci estimated severely reduced recombination rates in the families 86SD35, 86SD40, and 87SD4. These discordant results indicate that a different SDR mechanism other than second division omission was occurring. It is unlikely that a low frequency of simplex genotypes would result from a reduction in recombina
tion. For instance, a strong expression of a desynaptic gene would lead to univalents at metaphase I and an unbalanced first division (Jongedijk, 1985; Hermsen, 1984). Sterility would be extremely high if a SDR mechanism was exclusively operating in this synaptic genotype, while, only FDR-type gametes would be expected to be fertile under these desynaptic conditions. The segregation data for the proximal loci in this study did not support an FDR mode of 2n egg formation. Under less extreme desynaptic conditions, a mixture of SDR and FDR gametes could be possible, however" the data did not support this contention either.
Jongedijk (1985) has suggested that a postmeiotic doubling of reduced megaspores taking place during gametogenesis could lead to the formation of homozygous 2n eggs. Stelly & Peloquin (1986) also noted this mechanism as a potential source of excess homozygous genotypes in certain 2x-4x crosses. To account for this heterogeneous population of almost completely homozygous 2n eggs in our study, it is likely that two separate mechanisms of 2n egg formation occur in these diploid parents: 1) a high frequency of post meiotic doubling of the reduced megaspore generating completely homozygous 2n eggs and, 2) an omission of the second division generating SDR-type gametes with normal meiosis occurring at a low frequency in the gametophytic population.
Transmission ofheterozygosity through SDR mechanisms: The SDR gametes, produced in non-deviant families, transmitted on the average 39.1% of their heterozygosity (Table 6). In comparison, theoretical expectations, based upon a number of cytological assumptions, estimated SDR gametes can transmit 35-40% of their heterozygosity (Peloquin, 1981; Hermsen, 1984). Our observed value, based upon six chromosome segments, supports theoretical expectations that SDR mechanisms are unable to transmit heterozygosity at high levels.
A SDR mechanism which promotes homozygous 2n gametes is less desirable for transmitting heterozygosity in 2x-4x crosses than SDR gametes formed through second division ommission of the megaspore. A mechanism such as post-meiotic doubling of the reduced megaspore, prevents any

259
heterozygosity from being transferred through the female 2n gametophyte (Jongedijk, 1985). SDR gametes of this type appear undesirable, considering the importance of heterozygosity when breeding a tetrasomic autoploid crop like the potato. FDR gametes transmit a large percentage of the parental genome intact (81.5-98.00/0), which tends toward homogeneous populations of highly heterozygous 2n gametes (Douches & Quiros, 1988). SDR eggs generated through a post-meiotic doubling mechanism produce a population inbred 2n ganletophytes.
The feasibility of producing inbred lines through SDR mechanisms has been studied in several organisms. Matromorphoric embryos generated through SDR mechanisms have been identified in Brassica spp. (Eenick, 1974a, 1974b) and in S. andigena (Taylor, 1978), however, heterozygosity has been found in them. In rainbow trout, diploid gynogenesis, a potential method to produce inbred lines, has been suggested through retention of the second polar body (Allendorf et aI., 1986), or by suppression of the first mitotic cell division (Purdom, 1983). A system to generate completely inbred potato clones would be established if these 2n gametes produced through post-meiotic doubling, cou~d be induced to develop parthenogenetically. Parthenogenetic seed development has been routine in generating potato haploids (2n = 2x = 24) from tetraploid clones (Hougas & Peloquin, 1957; Montelongo-Escobedo & Rowe, 1969) and has also been extended to generating monoploids (n =
x = 12) (Van Breukelen et aI., 1975). The generation of inbred diploid clones could have important genetic and breeding implications; i.e. inheritance studies and the use of F1 hybrid and double-cross breeding methodologies. The testing of the efficacy of generating inbred diploid clones through haploid pollinator and tissue culture techniques would be of interest.
Value ofHTA: The limitations of HTA, in the past, were mainly influenced by the markers available. For example, the dominant nature of chromosome position of the yellow tuber flesh locus (y) limited its value to determine the composition of the 2n gametophyte populations (Stelly & Peloquin,
1986). The enzyme-coding loci used in this study expand HTA as a means to study megasporogenesis and megagametogenesis. Their co-dominant nature allowed the segregating progeny to be completely classified to obtain more precise estimates of segregation patterns through 2n gamete mechanisms. In addition, the identification of markers with tighter centromeric linkage has improved our ability to discriminate between SDR and FDR modes even in small progenies. The analytic powers of HTA were greatly improved by following the segregation of both proximal and distal loci in a 2x-4x family. Initially, the segregation of proximal markers was used to determine the mode of 2n eggformation. Secondly, the segregation of a distal locus provided information concerning recombination levels within the 2n gametic pool, which has been extremely difficult to assess cytologically. In this study, a post-meiotic doubling of the reduced megaspore was suggested to be operating in at least three of the 19 clones, leading to homozygous SDR gametes. Hence, caution should be exercised when SDR mechanisms are utilized in 2x-4x and 2x-2x breeding schemes, since heterozygosity transmission can vary greatly (39.1% vs. 1.8°/0).
An analytic system as described above would be useful in: 1) discriminating between FDR/SDR and exclusive FDR populations of female 2n gametophytes, and 2) quantifying the asynapsis associated with FDR 2n gametophytes, if sufficient progeny sizes can be generated.
References
Alleendorf, F.W., J.E. Seeb, K.L. Knudsen, G.H. Thorgaard & R.F. Leary, 1986. Gene-centromere mapping in rainbow trout. J Heredity 77: 307-312.
Douches, D.S., 1987. Development of genetic strategies to determine the mode of 2n egg formation in diploid potatoes. Ph.D. Thesis, University of California, Davis.
Douches, D.S. & C.F. Quiros, 1986a. Additional isozyme loci in tuber-bearing Solanums: Inheritance and linkage relationships. J Heredity (in press).
Douches, D.S. & C.F. Quiros, 1986b. Use of 4x-2x crosses to determine gene-centromere map distances of isozyme loci in Solanum species. Can J Genet Cytol (in press).
Douches, D.S. & C.F. Quiros, 1988. Genetic recombination in a diploid synaptic mutant and a Solanum tuberosum x S.

260
I
chacoense diploid hybrid. Heredity 60: 183-191. Eenink, A.H., 1974. Matromorphy in Brassica oleracea L. IV.
Formation of homozygous and heterozygous diploid products of gametogenesis and qualitative genetical research on matromorphic plants. Euphytica 23: 719-724.
Eenink, A.H. 1975. Matromorphy in Brassica oleracea L. VII. Research on products of microsporogenesis and gametogenesis from prickle pollinated plants. Euphytica 24: 45-52.
Hermsen, J. G.Th., 1984. Mechanisms and Genetic Implications of 2n-Gamete Formation. Iowa State Journal of Research 58(4): 421-434.
Hougas, R.W. & S.J. Peloquin, 1957. A haploid plant of the potato variety Katahdin. Nature 180: 1209-1210.
Huaman, Z. & R.W. Ross, 1985. Updated listing of potato species names, abbreviations and taxonomic status. Am Potato J 62: 629-641.
Iwanaga, M. & S.F. Peloquin, 1979. Synaptic mutant affecting only megasporogenesis in potatoes. J Heridity 70: 385-389.
Jongedijk, E., 1985. The pattern of megasporogensis and megagametogenesis in diploid Solanum species hybrids; its relevance to the origin of 2n eggs and the induction of apomixis. Euphytica 34: 599-611.
McHale, N.A. & F.I. Lauer, 1981. Breeding value of2n pollen diploid from hybrids and S. phureja in 4x-2x crosses in potatoes. Am Potato J 58: 365-374.
Mendenhall, W., R.L. Schaeffer & D.D. Wackerly, 1986. Mathematical Applications. Third Edition. Duxberry Press, Boston, 62 p.
Mendiburu, A.O. & S.J. Peloquin, 1977a. The significance of 2n gametes in potato breeding. Theor Appli Genet 49: 53-61.
Mendiburu, A.O. & S.J. Peloquin, 1977b. Bilateral sexual polyploidization in potatoes. Euphytica 26: 573-583.
Mendiburu, A.O. & S.J. Peloquin, 1979. Gene-centromere mapping by 4x-2x matings in potatoes. Theor Appl Genet 54: 177-180.
Mok, D.W.S. & S.J. Peloquin, 1975a. Breeding value of 211 pollen (diplandroids) in tetraploid x diploid crosses in potatoes. Theor Appl Genet 46: 307-314.
Mok, D.W.S. & S.J. Peloquin, 1975b. Three mechanisms of2n pollen formation in diploid potatoes. Can J Genet Cytol17: 217-225.
Mok, D.W.S., S.J. Peloquin & A.O. Mendiburu, 1976. Genetic evidence for mode of 2n pollen formation and S-locus mapping in potatoes. Potato Res 19: 157-164.
Mok, I., 1981. Sexual polyploidization and protein diversity in potatoes. Ph.D. Thesis, University of Wisconsin, Madison.
Montelongo-Escobedo, H. & P.R. Rowe, 1969. Haploid induction in potato: Cytological basis for the pollinator effect. Euphytica 18: 116-123.
Peloquin, S.J., 1981. Breeding methods for achieving phenotypic uniformity. In: The International Potato Center. Report of
the Planning Conference on the Production of Potato from True Seed. Manila Philippines, pp. 151-155.
Peloquin, S.J., 1983. New approaches to breeding for the potato in the year 2000. In: Proceedings of the International Congress on 'Research for the Potato in the year 2000'. International Potato Center, Lima, Peru, pp. 32-34.
Pfeiffer, T.W. & E.T. Bingham, 1983. Abnormal meiosis in alfalfa, Medicago sativa: cytology of 2n egg and 4n pollen formation. Can J Genet Cytol25: 107-112.
Purdom, C.E., 1983. Genetic engineering by the manipulation of chromosomes. Aquaculture 33: 287-300.
Quinn, A.A. & S.J. Peloquin, 1973. Use of experimental tetraploids in potato breeding. Am Potato J 50: 415-420.
Quiros, C.F., 1981. Starch gel electrophoresis techniques used with alfalfa and other Medicago species. Can J Plant Sci 61: 745-749.
Quiros, C.F. & N. McHale, 1985. Genetic analysis of isozyme variants in diploid and tetraploid potatoes. Genetics 111: 131-145.
Ramanna, M.S., 1979. A re-examination of the mechanisms of 2n gamete formation in potato and its implications for breeding. Euphytica 28: 537-561.
Ross, H. & F.A. Langton, 1974. Origin of unreduced embryo sacs in diploid potatoes. Nature 247: 378-379.
Stelly, D.M., S.J. Peloquin, R.G. Palmer & C.F. Crane, 1984. Mayer's Hemalum-Methylsalicylate: A stain-clearing technique for observations within while ovules. Stain Technology 59(3): 155-161.
Stelly, D .M. & S.J. Peloquin, 1986. Diploid female gametophyte formation in 24-chromosome potatoes: Genetic evidence for the prevalence of the second meiotic division restitution mode. Can J Genet Cytol28: 101-108..
Tai, G.C.C. & H. Dejong, 1980. Multivariate analysis of potato hybrids. I. Discrimination between tetraploid-diploid hybrid families and their relationship to cultivars. Can J Genet Cytol 22: 227-235.
Taylor, L.M., 1978. Variation patterns of parthenogenetic plants derived from unreduced embryo sacs of Solanum tuberosum subspecies andigena (Juz. et Buk.). Theor Appl Genet 52: 241-249.
Vallejos, C.E., 1983. Enzyme activity staining. In: Isozymes in plant genetics and breeding (eds. Tanksley, S.D. and T.J. Orton) Vol A, Elsevier, Amsterdam, pp. 469-516.
Van Breukelen, E.W.M., M.S. Ramanna & J.G.Th. Hermsen, 1975. Monoploids (n = x = 12) from autotetraploid Solanum tuberosum (2n = 4x = 48) through two successive cycles of female parthenogenesis. Euphytica 24: 567-574.
Veilleux, R.E., N.A. McHale & F.I. Lauer, 1982. 2n ganletes in diploid Solanum: frequency and types of spindle abnormalities. Can J Genet Cytol 24: 301-314.




















