
Cryopreservation of Shoot Tips of Tetraploid Potato ( Solanum tuberosumL.) Clones by Vitrification
Upload
independentCategory
view
1download
0
Phenotypic characterization and nuclear microsatelliteanalysis reveal genomic changes and rearrangementsunderlying androgenesis in tetraploid potatoes(Solanum tuberosum L.)
Sushruti Sharma Æ Debabrata Sarkar ÆSuman Kumar Pandey
Received: 10 October 2008 / Accepted: 17 June 2009 / Published online: 1 July 2009
� Springer Science+Business Media B.V. 2009
Abstract Two (di)haploids (2n = 2x = 24) and
nine tetraploids (2n = 4x = 48) obtained from Sola-
num tuberosum through anther culture were character-
ized for nDNA variation, phenotypic variation and
nuclear microsatellite polymorphism. Androgenic
(di)haploids were also characterized for late blight
resistance. The (di)haploid C-13 was derived from
Indian tetraploid potato cv. Kufri Chipsona-2, while
D4 from TPS (true potato seed) parental line JTH/C-
107, which is an interspecific hybrid between Indian
tetraploid cv. Kufri Jyoti and diploid (2n = 2x = 24)
cultivated species S. phureja Juz. & Buk. IVP-35. C-13
and D4 (both male-fertile) could be distinguished from
their corresponding tetraploid anther donors based on
plant height, shoot number, terminal leaflet length and
width, leaf ratio, anther length, pollen diameter and
corolla width and radius. A complete reversal of flower
color occurred in D4, and C-13 was highly resistant to
late blight. Most interestingly, about 3–7% increase in
nDNA content occurred in most of the anther-derived
tetraploids. Both the androgenic (di)haploids and their
anther donors had unique genotypes at the microsat-
ellite loci POTM1-2, STM0015 and STM0019b. How-
ever, the nine anther-derived tetraploids shared the
same allelic profiles with their anther donor JTH/C-107
at all the microsatellite loci, except at STM0019a
where they were characterized by the absence of a
standard donor allele (186-bp). A typical (di)haploid-
specific allele was detected for the locus STWAX-2
where the standard donor alleles were replaced by a
230-bp allele in both C-13 and D4. The over-expres-
sion of microsatellite variation in D4 that also shows
triallelic profiles at the microsatellite loci POTM1-2
and STM0015 can perhaps be attributed to its chimeric
structure, which might have been formed through
incomplete fusion of two different pro-embryos during
the first steps of microspore division.
Keywords Androgenesis � Anther culture �(Di)haploid � Microsatellite � Nuclear DNA content �Phenotyping � Potato � Simple sequence repeats �Solanum tuberosum L.
Abbreviations
AFLP Amplified fragment length polymorphism
AUDPC Area under the disease progress curve
CRBC Chicken red blood cell
CTAB Cetyltrimethylammonium bromide
Sushruti Sharma passed away on November 10, 2008.
S. Sharma � D. Sarkar � S. K. Pandey
Cell and Molecular Biology Laboratory, Division of Crop
Improvement, Central Potato Research Institute (CPRI),
Shimla, Himachal Pradesh 171 001, India
D. Sarkar (&)
Biotechnology Unit, Division of Crop Improvement,
Central Research Institute for Jute & Allied Fibres
(CRIJAF), Barrackpore, Kolkata, West Bengal 700 120,
India
e-mail: [email protected]; [email protected];
123
Euphytica (2010) 171:313–326
DOI 10.1007/s10681-009-9983-7

DAPI 40,6-Diamidino-2-phenylindole
EPU Eyepiece unit
nDNA Nuclear DNA
KI Potassium iodide
RAPD Random amplified polymorphic DNA
RCBD Randomized complete block design
RFLP Restriction fragment length
polymorphism
SSR Simple sequence repeat
TPS True potato seed
Introduction
Tetraploid potatoes (Solanum tuberosum L.; 2n =
4x = 48) are highly heterozygous, with complicated
genetic segregation due to tetrasomic mode of inher-
itance. Since (di)haploids (2n = 2x = 24) are thought
to be genetically pure S. tuberosum, they can be
regarded as gametic samples of their tetraploid parents
(Wilkinson et al. 1995). (Di)haploid potato can be
obtained either paternally through androgenesis or
maternally through gynogenesis. The gynogenic
method relies on the development of (di)haploids from
unfertilized ovules by parthenogenesis (Samitsu and
Hosaka 2002), or from fertilized triploid zygotes by
chromosome elimination (Clulow et al. 1991), or
through somatic translocations (Wilkinson et al. 1995).
However, gynogenic (di)haploids are not always
completely parthenogenetic in origin (Clulow et al.
1991; Waugh et al. 1992; Wilkinson et al. 1995). The
haploid-inducing S. phureja pollinator, which is
essentially used for the production of gynogenic
(di)haploids in potato, frequently produces not only
aneusomatic (di)haploids with variable chromosome
numbers (Clulow et al. 1991; Samitsu and Hosaka
2002), but also (di)haploids with substantial amount of
inducer’s nuclear genome (Waugh et al. 1992;
Wilkinson et al. 1995).
Androgenesis through anther culture is, therefore,
the most reliable method of (di)haploid-derivation in
tetraploid potatoes (Wenzel 1994). It is also consid-
ered more efficient than gynogenesis because there
are greater numbers of microspores among which
superior recombinants can occur than ovules in a
flower (Jacobsen and Ramanna 1994). However,
androgenic (di)haploidization via anther culture has
been proved to be frustratingly difficult in potato
(Wenzel 1994; Cappadocia and Ramulu 1998). There
are only few reports on the induction of androgenesis
in tetraploid S. tuberosum (Rokka et al. 1996 and
references therein) and in disomic tetraploid potato
species S. acaule ssp. acaule (Rokka et al. 1998),
leading to successful regeneration of (di)haploid
plants. Although flowering has been reported in
androgenic (di)haploids of S. tuberosum (Meyer et al.
1993; Calleberg 1996; Song et al. 2005), they
generally show low levels of male fertility similar
to their gynogenic counterparts. In androgenic
(di)haploids of S. acaule ssp. acaule, flowers could
develop, but they were infertile (Rokka et al. 1998).
Phenotypic characterization of anther-derived
plants represents a major means for distinguishing
the haploids from their parental lines according to
ploidy level. This approach was successfully
employed in diploid (2n = 2x = 24) S. phureja not
only to group anther-derived plants (Pehu et al.
1987), but also to compare the performance of
monoploid potatoes developed through androgenesis
versus gynogenesis (Lough et al. 2001). Potato
(di)haploids derived from specific cultivars of tetra-
ploid S. tuberosum exhibited varying levels of
phenotypic and genetic variation in comparison to
their donor cultivars (Matsubayashi 1979; De Maine
1984; Kotch et al. 1992). However, in S. acaule ssp.
acaule, the anther-derived plants were normal look-
ing and as a group morphologically similar to each
other and to the tetraploid anther donor (Rokka et al.
1998). There were no differences in growth, color,
number and size of leaflets and color and size of floral
corollas and anthers between anther-derived (di)hap-
loids or tetraploids and their tetraploid anther donor
S. acaule. This may be because S. acaule is largely
self-pollinating (Watanabe et al. 1994), which results
in a high degree of homozygosity and thus a low level
of variation in its (di)haploid population. In the
present study, we obtained androgenic (di)haploids of
tetraploid S. tuberosum, which not only flower but
also are male fertile. These androgenic (di)haploids
were characterized in relation to their corresponding
tetraploid anther donors for a large number of
phenotypic traits and late blight resistance. In this
study, the androgenic competence of tetraploid
potatoes was also assessed, and the anther-derived
plants obtained were characterized for ploidy level,
chromosome number and nuclear DNA variation.
314 Euphytica (2010) 171:313–326
123

DNA markers have been extensively used for
assessing the genetic variation and for determining
the genetic composition of anther-derived plants in
potato (Birhman et al. 1994; Rivard et al. 1994;
Veilleux et al. 1995; Chani et al. 2000, 2002). All
these studies were, however, confined to the genetic
analyses of monoploid and/or diploid anther-derived
plants, which originated from diploid S. chacoense or
S. phureja anther donors, or their interspecific
hybrids. Rokka et al. (1995) analyzed the androgenic
triploids of hexaploid somatic hybrids (S. brevi-
dens 9 S. tuberosum) by RAPD markers. The utility
of androgenic (di)haploids for genetic marker devel-
opment in potato was demonstrated using AFLP and
SSR markers (Song et al. 2005). However, DNA
markers were never used to characterize androgenic
(di)haploids in relation to their tetraploid anther
donors for assessing the genomic changes and
rearrangements underlying androgenesis in tetraploid
potatoes.
Microsatellites, also termed as simple sequence
repeats (SSRs), have been used to distinguish homo-
zygous doubled monoploids from heterozygous dip-
loid plants in a monoploid anther-derived population
originated from an interspecific diploid hybrid
between S. chacoense and S. phureja (Veilleux
et al. 1995; Chani et al. 2000, 2002). They offer the
same discriminatory potential as RFLPs because both
alleles of a heterozygote can be detected (Veilleux
et al. 1995). Nuclear microsatellite analysis has
proved to be very useful for genotyping tetraploid
potato cultivars and for assessing the comparative
diversity among the potato Groups (McGregor et al.
2000; Ashkenazi et al. 2001; Ghislain et al. 2006).
However, the effectiveness of microsatellites for
characterizing the genetic variation and for determin-
ing the genetic composition of androgenic (di)hap-
loids has not been assessed so far. Since meiotic
processes and the modes of gamete formation in
tetraploid potatoes differ greatly from those in diploid
potatoes, there is no one-to-one relationship between
the phenotype and the genotype of DNA markers
scored in tetraploid individuals (Luo et al. 2004).
Hence, genotyping and characterization of the segre-
gation pattern in androgenic (di)haploids using more
informative genetic markers, such as DNA microsat-
ellites, are not as simple and straightforward as in an
androgenic monoploid population that originated
from a diploid potato species or hybrid.
It is thus of potential interest to resolve the
discriminatory ability of SSRs for assessing the
androgenic origin of anther-derived (di)haploids and
for determining the genetic composition of sporo-
phytic gamete genotypes of tetraploid potatoes. The
present study assessed the potential of nuclear
microsatellite markers for characterizing the andro-
genic (di)haploids in relation to their tetraploid
anther-derived sibs and anther donors. This study
also goes in some way towards determining whether
differential sporophytic pattern of microspore
embryogenesis is reflected in nSSR polymorphism
scored in the regenerated sporophytes of the resulting
gamete genotypes.
Materials and methods
Plant material
For the induction of androgenesis, 14 agronomically
superior tetraploid (2n = 4x = 48) potato (Solanum
tuberosum L. ssp. tuberosum) cultivars were used:
FL-1533, Kufri Badshah, Kufri Chandramukhi, Kufri
Chipsona-1, Kufri Chipsona-2, Kufri Giriraj, Kufri
Jyoti, Kufri Kanchan, Kufri Lalima, Kufri Megha,
Kufri Sindhuri, Kufri Sutlej, MF-II and TPS-13.
Besides, a tetraploid interspecific hybrid JTH/C-107
(S. tuberosum ssp. tuberosum cv. Kufri Jyoti 9 S.
phureja IVP-35; TPS parental line) was also used.
They were raised from tubers in the experimental
field of the Central Potato Research Institute (CPRI),
Shimla (31.06�N, 77.10�E; 2202 m above msl)
during the summer (May–August) following the
recommended cultural practices. The flower buds
were harvested and used for initiating the anther
culture during June–July (mean day/night tempera-
ture: 24.4/16.1�C; RH: 80–90%).
Anther culture
The harvested flower buds (4–6 mm in length) were
processed as described earlier (Sarkar et al. 2004a).
Anthers were isolated aseptically and cultured on MS
(Murashige and Skoog 1962) medium supplemented
with 60 g l-1 sucrose, 1.0 mM ascorbic acid, 2.0 mg l-1
D-calcium pantothenate, 0.5 mM L-cysteine, 60 g l-1
potato starch, 5.0 g l-1 activated charcoal, 17.74 lM
BA and 2.0 g l-1 gelrite (modified from Rokka et al.
Euphytica (2010) 171:313–326 315
123

1996). The pH was adjusted to 5.8 before autoclaving at
121�C for 20 min. The cultures were incubated under a
16 h photoperiod (20 lmol m-2 s-1 PFD) at 28�C.
After 8 weeks’ culture, the androgenic response was
assessed by scoring the frequency of androgenesis. The
embryoids that did not develop into shoots were cultured
on � MS medium supplemented with 2.89 lM GA3,
5.69 lM zeatin riboside, 20 g l-1 sucrose and 2.0 g l-1
gelrite. The regeneration cultures were incubated under
a 16 h photoperiod (60 lmol m-2 s-1 PFD) at 20�C.
Anther-derived regenerants were maintained and mul-
tiplied through single-node cuttings (SNCs) as
described earlier (Sarkar et al. 2004b).
Ploidy diagnosis
For L1-ploidy estimation, the chloroplast density per
stomatal guard cell pair was examined in subterminal
leaflet of the fourth-leaf from the top of 2-month-old
greenhouse-grown plants with a solution of 0.5%
iodine in 5% aqueous KI solution for 2 min. The
length and width of stomatal guard cells were
measured using an eyepiece graticule at 9400 mag-
nification, and all measurements were recorded to the
nearest whole eyepiece unit (1 EPU = 2.0 lm). For
L2- and L3-ploidy estimation, young flower buds and
healthy root tips, respectively were fixed in Carnoy’s
fluid for 24 h. The roots were hydrolyzed in 1 N HCl
for 10 min at 60�C, stained in 2% aceto-orcein, and
squash preparations were made in 45% acetic acid.
The chromosomes in pollen mother cells were stained
with the fluorescent dye DAPI (1.0 ml of 0.35 mg ml-1
DAPI stock was used to prepare a 100 ml working
solution in 0.4 M Na2HPO4). Counts were obtained
from a minimum of 15 cells and from at least five
separate flower buds or roots.
Flow cytometry
Leaf tissues (*50 mg), collected from 4-week-old
microplants, were macerated to prepare nuclei sus-
pensions (Arumuganathan and Earle 1991). Relative
fluorescence of the propidium iodide-stained leaf
nuclei was measured in a FACSCalibur flow cytom-
eter (Becton Dickinson, San Jose, USA) using CRBC
as an internal standard. The objects were analysed
for forward (FSC) versus side (SSC) scatter signals
for at least 5,000 nuclei in each sample. The peak
corresponding to the CRBC nuclei was adjusted to
around channel 200 set on a linear scale of fluores-
cence intensity. The nuclear DNA amount (2C value
in pg) was calculated by direct comparison of the
mean position of plant nuclear peak to the mean
position of CRBC nuclear peak (2.33 pg). For each
sample, there were three independent replicated
measurements.
Field studies
Field studies were conducted on the two androgenic
dihaploids (C-13 and D4) and their corresponding
tetraploid parents (Kufri Chipsona-2 and JTH/C-107).
As potato plants flower only under long-day condi-
tions of Shimla hills (mid Himalayas) in northern
India, this experiment was conducted in the summer
(May–August). The tubers were planted in a ran-
domized complete block design (RCBD) with five
replications. Each clone was represented by three
rows of five plants each at a spacing of 20 cm in rows
60 cm apart per replication. Fertilizer schedules and
cultural practices were as described earlier (Naik
et al. 1998). At 60 days after planting, the following
data were collected: plant height, number of shoots,
terminal leaflet length and width, leaf ratio (average
terminal leaflet length divided by average terminal
leaflet width), leaflet number, leaflet length and
width, internode length, stem width, specific leaf
area, number of inflorescences, number of flower
buds, anther length at dehiscence, corolla radius to
petal tip and corolla width at widest diameter. The
data on internode length, number of tubers, average
tuber weight, harvest index and tuber dry matter were
collected at harvest (120 days after planting). Total
soluble sugars, reducing sugars and starch content of
freshly harvested tubers were estimated as described
earlier (Sharma et al. 2004). Pollen stainability and
quality were assessed by carmino acetic acid and
fluorochromatic tests (2.0 mg ml-1 fluorescein diac-
etate in acetone and few drops of this stock solution
were added to 2.0 ml of a 0.5 M sucrose solution),
respectively.
Foliar assessment of late blight disease
Assessments of the percentage of foliage affected
with late blight were made on the two middle rows of
each plot at an interval of 7 days based on the British
316 Euphytica (2010) 171:313–326
123

Mycological Society scale, starting on 16 July and
continuing for a total of six observations up to 20
August. Late blight susceptible cv. Kufri Jyoti was
used as a spreader. The area under the disease
progress curve (AUDPC) was calculated in percent-
age-day according to the midpoint rule method
(Campbell and Madden 1990). The relative area
under the disease progress curve (RAUDPC = AU-
DPC/(tn-t1) 9 100 where (tn-t1) is the duration of
the epidemic) was calculated to quantify the disease
level that ranged from 0.0 to 1.0 without any unit.
Nuclear microsatellite analysis
Total genomic DNA was extracted from 0.2 to 0.3 g
leaf tissues of 3-week-old microplants, according to a
modified CTAB-dichloromethane protocol (Saghai-
Maroof et al. 1984). Ten nuclear microsatellite loci
were characterized in the study (Table 1). The
reaction mixture (25 ll) was as follows: 19 assay
buffer, 2.5 mM MgCl2, 200 lM of each dNTP, 1.0 U
Taq polymerase (Qiagen GmbH, Hilden, Germany),
0.2 lM of each primer and 50 ng of template DNA.
A PCR protocol consisting of 30 cycles at 93�C for
30 s, Ta�C for 1 min and 72�C for 1 min was used
(Mastercycler� gradient; Eppendorf AG, Hamburg,
Germany). Negative controls were run with each
amplification. Samples were separated on a 18.0 cm
long 9 16.0 cm wide, 1.5 mm thick 8% non-dena-
turing polyacrylamide (29:1) gel, which were pre-run
at 180 V constant voltage for 10 min in 0.59 TBE
buffer using a Hoefer SE 600 (Amersham Biosci-
ences AB, Uppsala, Sweden) vertical electrophoresis
system; samples were loaded and separated under the
same conditions. The amplified products were
exposed with a modified silver staining protocol
(Sanguinetti et al. 1994). For each clone, the SSR
assay was performed at least five times using
five independent DNA extracts. For nSSR allele
detection and scoring, Gene Profiler Version 4.05
(Scanalytics, Fairfax, Virginia, USA), which allowed
single base differences to be detected, was used. Only
Table 1 Oligonucleotide sequences and other associated information for primer pairs used for the amplification from nuclear microsatellite
sequences for the characterization of an anther-derived population in potato
Locus
name
Accession
number
nSSR motif Primer sequence 50–30 Gene function
(LG)
Expected
allele size (bp)
Ta (�C)
POTM 1-2 U237558 (AT)20 AATAATACTGTGATGCCACAATGG
GTGGCATGTCTTCGAAGGTAC
POTM 1-2 (ND)a 221 55
STM0015 NAa (AC)13 (AT)10 GATTGTGAGAAGGCACTGA
CACTTGATATACTAGTGTGTTTGG
PAC26 (ND) 193 53
STM0019ab NA (AT)7 (GT)10 (AT)4
(GT)5 (GC)4 (GT)4
AATAGGTGTACTGACTCTCAATG
TTGAAGTAAAAGTCCTAGTATGTG
PAC33 (VI) 204 (161–239) 47
STM0019bb NA (AT)7 (GT)10 (AT)4
(GT)5 (GC)4 (GT)4
AATAGGTGTACTGACTCTCAATG
TTGAAGTAAAAGTCCTAGTATGTG
ND (83–119) 47
STM1043 U24088 (T)14.....(TAA)5 ATTTGAATTGAAGAACTTAATAGAA
CACAAACAAAATACTGTTAACTCA
STSUSY2 (X) 226 47
STM1069 X78271 (TCC)4 ATGCTAACTTGGACACTTA
AGTCTCTCAGGAGGATTAC
LEPDSGEN (III) 393 td 64–57
STM3003 NA (CT)4, (CA) CATAAATCACTAAAACTAGATAACC
TATTTCATATCAATGTCGAATCCC
ND 317 55
STWAX-1 X58453 (CTT)4 TGATTCTCTTGCCTACTGTAATCG
AGTCAGAGTATGGTTCCTGAGTCC
Waxy gene;
STGBSS (ND)
246 55
STWAX-2 X52417 (ACTC)5 CCCATAATACTGTCGATGAGCA
GAATGTAGGGAAACATGCATGA
G28WXST waxy
gene (VIII)
223 (224–243) 55
STWIN12G X13497 (AAT)6 AAATCGACACAGACGGAAATG
CGAGGGACTTTAATTTGTTGGA
STWIN12G (ND) 231–249 55
a NA not available; ND not determinedb Because using the same primer pairs STM0019 shows two distinct loci, they have been labeled as STM0019a and STM0019b
Euphytica (2010) 171:313–326 317
123

reproducible fragments were selected for data anal-
ysis. The alleles were generated using the Gene
Profiler linkage analysis database module.
Statistical analysis
Prior to univariate analysis, normality and equal
variance assumptions were tested, and accordingly,
the data on pollen stainability/quality (%) and tuber
dry matter (%) were transformed into arc sine square
roots and square roots, respectively. The data were
analyzed by ANOVA followed by all pairwise
multiple comparisons of mean values using the
post-hoc Tukey’s honestly significant difference test
(MSTAT-C; Michigan State University, Michigan,
USA). The term significant has been used to indicate
differences for which P B 0.05.
Results
Androgenic competence
A total of 27,715 anthers of the 15 genotypes gave
rise to three types of androgenic responses, viz.,
embryogenesis, callus induction, or a combination of
the both. Except for Kufri Chandramukhi, all
genotypes were responsive to anther culture; Kufri
Badshah, Kufri Chipsona-1, Kufri Megha and
TPS-13 were characterized by only callus induction,
whereas embryogenesis occurred in the other geno-
types (data not shown). However, complete plants
were obtained only in Kufri Chipsona-2 (henceforth
KCS-2) and JTH/C-107 (henceforth JTH). The
regenerated androgenic plantlets of K. Lalima did
not survive in cultures because they gradually turned
brown and degenerated. Distinct embryogenesis
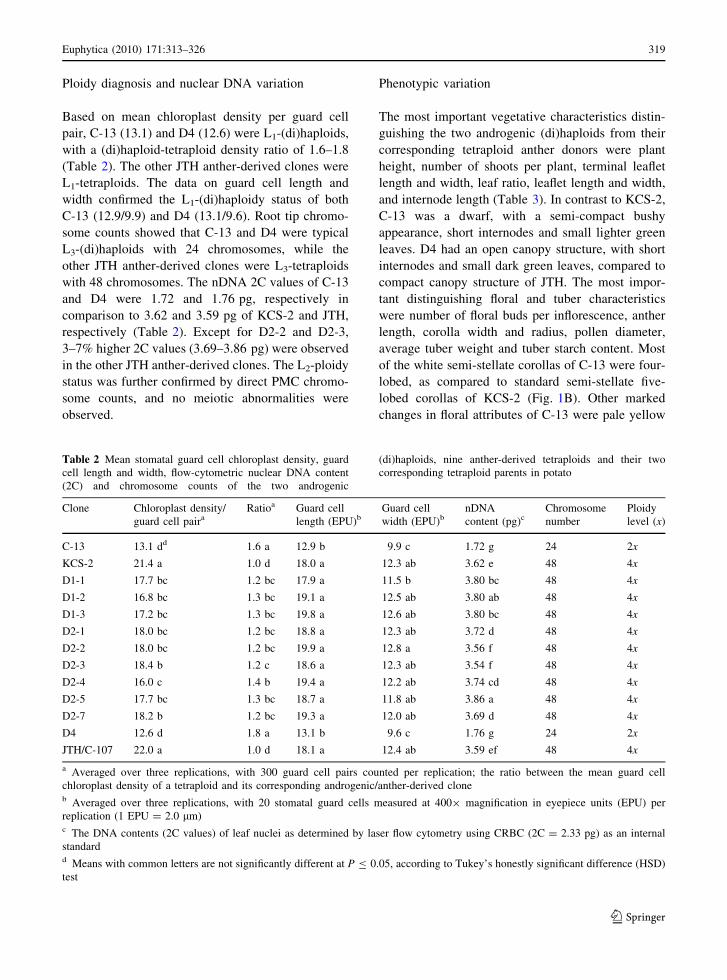
patterns were observed in KCS-2 and JTH (Fig. 1A,
C). The embryogenesis was exclusively direct
(primary microspore embryogenesis without any
intermediary callus phase) in KCS-2, while both
direct and indirect (somatic embryogenesis from
anther walls via intermediary callus dedifferentia-
tion) embryogenesis occurred in JTH. The progres-
sion of callus dedifferentiation from anther walls
was visualized by sterio-microscopic observations;
however, the somatic origin of the calluses could
only be ascertained by ploidy diagnosis, nDNA
content and nuclear microsatellite analysis of the
regenerants (see below). Most microspore-embryoid-
derived regenerants degenerated during various
stages of cultures in vitro. In contrast, the regener-
ation potential of somatic embryoids was higher in
JTH. Altogether 11 plants were obtained: one in
KCS-2 (C-13 from a microspore embryoid) and 10
in JTH (D4 from a microspore embryoid; D1-1 to
D1-3, D2-1 to D2-5 and D2-7 from somatic
embryoids).
Fig. 1 Androgenesis in
tetraploid Solanumtuberosum L. A The
androgenic (di)haploid C-
13 of tetraploid cv. Kufri
Chipsona-2 (KCS-2)
developing through primary
microspore embryogenesis.
B Five-lobed corollas of
KCS-2 (left) in comparison
to four-lobed corollas of C-
13 (right). C Primary
microspore embryogenesis
in tetraploid interspecific
hybrid JTH/C-107. D A
complete reversal of flower
color in androgenic
(di)haploid D4 (right) in
comparison to its anther
donor JTH/C-107 (left)
318 Euphytica (2010) 171:313–326
123
Ploidy diagnosis and nuclear DNA variation
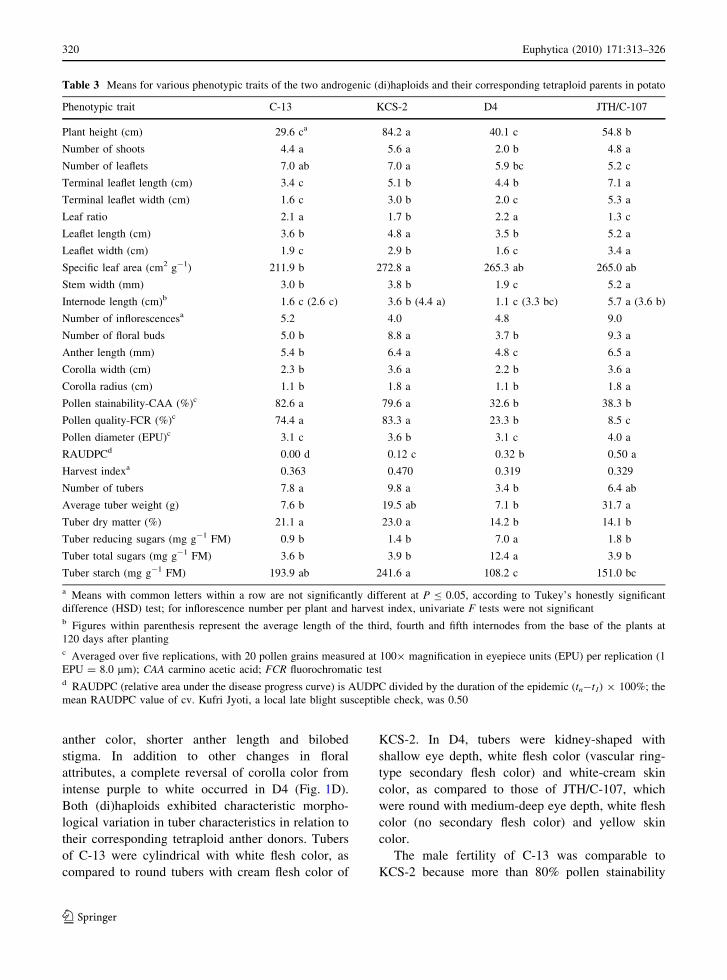
Based on mean chloroplast density per guard cell
pair, C-13 (13.1) and D4 (12.6) were L1-(di)haploids,
with a (di)haploid-tetraploid density ratio of 1.6–1.8
(Table 2). The other JTH anther-derived clones were
L1-tetraploids. The data on guard cell length and
width confirmed the L1-(di)haploidy status of both
C-13 (12.9/9.9) and D4 (13.1/9.6). Root tip chromo-
some counts showed that C-13 and D4 were typical
L3-(di)haploids with 24 chromosomes, while the
other JTH anther-derived clones were L3-tetraploids
with 48 chromosomes. The nDNA 2C values of C-13
and D4 were 1.72 and 1.76 pg, respectively in
comparison to 3.62 and 3.59 pg of KCS-2 and JTH,
respectively (Table 2). Except for D2-2 and D2-3,
3–7% higher 2C values (3.69–3.86 pg) were observed
in the other JTH anther-derived clones. The L2-ploidy
status was further confirmed by direct PMC chromo-
some counts, and no meiotic abnormalities were
observed.
Phenotypic variation
The most important vegetative characteristics distin-
guishing the two androgenic (di)haploids from their
corresponding tetraploid anther donors were plant
height, number of shoots per plant, terminal leaflet
length and width, leaf ratio, leaflet length and width,
and internode length (Table 3). In contrast to KCS-2,
C-13 was a dwarf, with a semi-compact bushy
appearance, short internodes and small lighter green
leaves. D4 had an open canopy structure, with short
internodes and small dark green leaves, compared to
compact canopy structure of JTH. The most impor-
tant distinguishing floral and tuber characteristics
were number of floral buds per inflorescence, anther
length, corolla width and radius, pollen diameter,
average tuber weight and tuber starch content. Most
of the white semi-stellate corollas of C-13 were four-
lobed, as compared to standard semi-stellate five-
lobed corollas of KCS-2 (Fig. 1B). Other marked
changes in floral attributes of C-13 were pale yellow
Table 2 Mean stomatal guard cell chloroplast density, guard
cell length and width, flow-cytometric nuclear DNA content
(2C) and chromosome counts of the two androgenic
(di)haploids, nine anther-derived tetraploids and their two
corresponding tetraploid parents in potato
Clone Chloroplast density/
guard cell pairaRatioa Guard cell
length (EPU)bGuard cell
width (EPU)bnDNA
content (pg)cChromosome
number
Ploidy
level (x)
C-13 13.1 dd 1.6 a 12.9 b 9.9 c 1.72 g 24 2x
KCS-2 21.4 a 1.0 d 18.0 a 12.3 ab 3.62 e 48 4x
D1-1 17.7 bc 1.2 bc 17.9 a 11.5 b 3.80 bc 48 4x
D1-2 16.8 bc 1.3 bc 19.1 a 12.5 ab 3.80 ab 48 4x
D1-3 17.2 bc 1.3 bc 19.8 a 12.6 ab 3.80 bc 48 4x
D2-1 18.0 bc 1.2 bc 18.8 a 12.3 ab 3.72 d 48 4x
D2-2 18.0 bc 1.2 bc 19.9 a 12.8 a 3.56 f 48 4x
D2-3 18.4 b 1.2 c 18.6 a 12.3 ab 3.54 f 48 4x
D2-4 16.0 c 1.4 b 19.4 a 12.2 ab 3.74 cd 48 4x
D2-5 17.7 bc 1.3 bc 18.7 a 11.8 ab 3.86 a 48 4x
D2-7 18.2 b 1.2 bc 19.3 a 12.0 ab 3.69 d 48 4x
D4 12.6 d 1.8 a 13.1 b 9.6 c 1.76 g 24 2x
JTH/C-107 22.0 a 1.0 d 18.1 a 12.4 ab 3.59 ef 48 4x
a Averaged over three replications, with 300 guard cell pairs counted per replication; the ratio between the mean guard cell
chloroplast density of a tetraploid and its corresponding androgenic/anther-derived cloneb Averaged over three replications, with 20 stomatal guard cells measured at 4009 magnification in eyepiece units (EPU) per
replication (1 EPU = 2.0 lm)c The DNA contents (2C values) of leaf nuclei as determined by laser flow cytometry using CRBC (2C = 2.33 pg) as an internal
standardd Means with common letters are not significantly different at P B 0.05, according to Tukey’s honestly significant difference (HSD)
test
Euphytica (2010) 171:313–326 319
123
anther color, shorter anther length and bilobed
stigma. In addition to other changes in floral
attributes, a complete reversal of corolla color from
intense purple to white occurred in D4 (Fig. 1D).
Both (di)haploids exhibited characteristic morpho-
logical variation in tuber characteristics in relation to
their corresponding tetraploid anther donors. Tubers
of C-13 were cylindrical with white flesh color, as
compared to round tubers with cream flesh color of
KCS-2. In D4, tubers were kidney-shaped with
shallow eye depth, white flesh color (vascular ring-
type secondary flesh color) and white-cream skin
color, as compared to those of JTH/C-107, which
were round with medium-deep eye depth, white flesh
color (no secondary flesh color) and yellow skin
color.
The male fertility of C-13 was comparable to
KCS-2 because more than 80% pollen stainability
Table 3 Means for various phenotypic traits of the two androgenic (di)haploids and their corresponding tetraploid parents in potato
Phenotypic trait C-13 KCS-2 D4 JTH/C-107
Plant height (cm) 29.6 ca 84.2 a 40.1 c 54.8 b
Number of shoots 4.4 a 5.6 a 2.0 b 4.8 a
Number of leaflets 7.0 ab 7.0 a 5.9 bc 5.2 c
Terminal leaflet length (cm) 3.4 c 5.1 b 4.4 b 7.1 a
Terminal leaflet width (cm) 1.6 c 3.0 b 2.0 c 5.3 a
Leaf ratio 2.1 a 1.7 b 2.2 a 1.3 c
Leaflet length (cm) 3.6 b 4.8 a 3.5 b 5.2 a
Leaflet width (cm) 1.9 c 2.9 b 1.6 c 3.4 a
Specific leaf area (cm2 g-1) 211.9 b 272.8 a 265.3 ab 265.0 ab
Stem width (mm) 3.0 b 3.8 b 1.9 c 5.2 a
Internode length (cm)b 1.6 c (2.6 c) 3.6 b (4.4 a) 1.1 c (3.3 bc) 5.7 a (3.6 b)
Number of inflorescencesa 5.2 4.0 4.8 9.0
Number of floral buds 5.0 b 8.8 a 3.7 b 9.3 a
Anther length (mm) 5.4 b 6.4 a 4.8 c 6.5 a
Corolla width (cm) 2.3 b 3.6 a 2.2 b 3.6 a
Corolla radius (cm) 1.1 b 1.8 a 1.1 b 1.8 a
Pollen stainability-CAA (%)c 82.6 a 79.6 a 32.6 b 38.3 b
Pollen quality-FCR (%)c 74.4 a 83.3 a 23.3 b 8.5 c
Pollen diameter (EPU)c 3.1 c 3.6 b 3.1 c 4.0 a
RAUDPCd 0.00 d 0.12 c 0.32 b 0.50 a
Harvest indexa 0.363 0.470 0.319 0.329
Number of tubers 7.8 a 9.8 a 3.4 b 6.4 ab
Average tuber weight (g) 7.6 b 19.5 ab 7.1 b 31.7 a
Tuber dry matter (%) 21.1 a 23.0 a 14.2 b 14.1 b
Tuber reducing sugars (mg g-1 FM) 0.9 b 1.4 b 7.0 a 1.8 b
Tuber total sugars (mg g-1 FM) 3.6 b 3.9 b 12.4 a 3.9 b
Tuber starch (mg g-1 FM) 193.9 ab 241.6 a 108.2 c 151.0 bc
a Means with common letters within a row are not significantly different at P B 0.05, according to Tukey’s honestly significant
difference (HSD) test; for inflorescence number per plant and harvest index, univariate F tests were not significantb Figures within parenthesis represent the average length of the third, fourth and fifth internodes from the base of the plants at
120 days after plantingc Averaged over five replications, with 20 pollen grains measured at 1009 magnification in eyepiece units (EPU) per replication (1
EPU = 8.0 lm); CAA carmino acetic acid; FCR fluorochromatic testd RAUDPC (relative area under the disease progress curve) is AUDPC divided by the duration of the epidemic (tn-t1) 9 100%; the
mean RAUDPC value of cv. Kufri Jyoti, a local late blight susceptible check, was 0.50
320 Euphytica (2010) 171:313–326
123
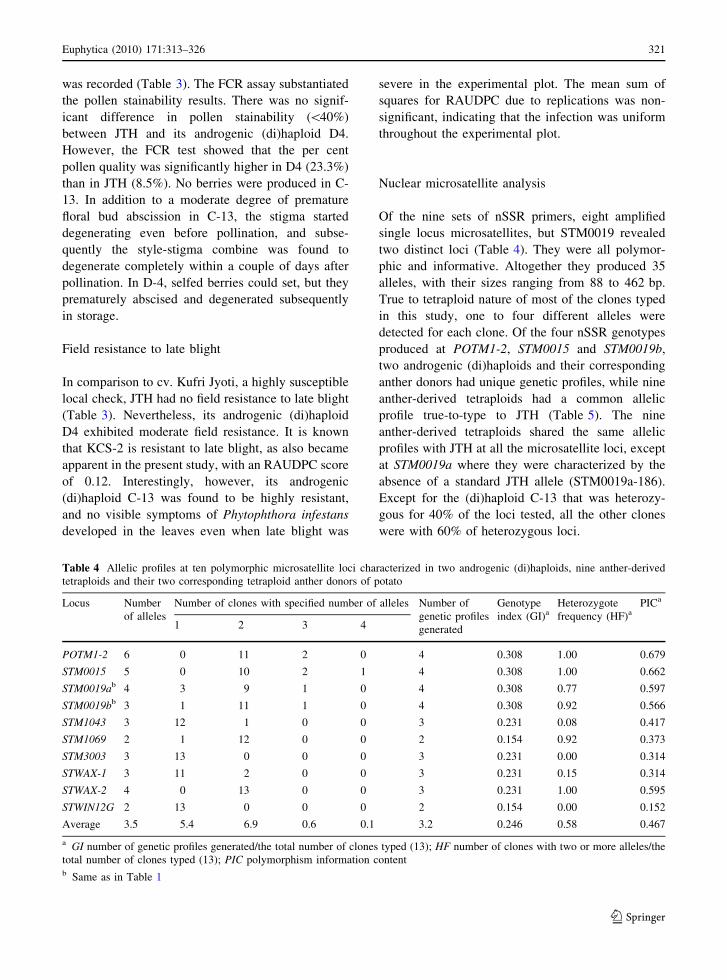
was recorded (Table 3). The FCR assay substantiated
the pollen stainability results. There was no signif-
icant difference in pollen stainability (\40%)
between JTH and its androgenic (di)haploid D4.
However, the FCR test showed that the per cent
pollen quality was significantly higher in D4 (23.3%)
than in JTH (8.5%). No berries were produced in C-
13. In addition to a moderate degree of premature
floral bud abscission in C-13, the stigma started
degenerating even before pollination, and subse-
quently the style-stigma combine was found to
degenerate completely within a couple of days after
pollination. In D-4, selfed berries could set, but they
prematurely abscised and degenerated subsequently
in storage.
Field resistance to late blight
In comparison to cv. Kufri Jyoti, a highly susceptible
local check, JTH had no field resistance to late blight
(Table 3). Nevertheless, its androgenic (di)haploid
D4 exhibited moderate field resistance. It is known
that KCS-2 is resistant to late blight, as also became
apparent in the present study, with an RAUDPC score
of 0.12. Interestingly, however, its androgenic
(di)haploid C-13 was found to be highly resistant,
and no visible symptoms of Phytophthora infestans
developed in the leaves even when late blight was
severe in the experimental plot. The mean sum of
squares for RAUDPC due to replications was non-
significant, indicating that the infection was uniform
throughout the experimental plot.
Nuclear microsatellite analysis
Of the nine sets of nSSR primers, eight amplified
single locus microsatellites, but STM0019 revealed
two distinct loci (Table 4). They were all polymor-
phic and informative. Altogether they produced 35
alleles, with their sizes ranging from 88 to 462 bp.
True to tetraploid nature of most of the clones typed
in this study, one to four different alleles were
detected for each clone. Of the four nSSR genotypes
produced at POTM1-2, STM0015 and STM0019b,
two androgenic (di)haploids and their corresponding
anther donors had unique genetic profiles, while nine
anther-derived tetraploids had a common allelic
profile true-to-type to JTH (Table 5). The nine
anther-derived tetraploids shared the same allelic
profiles with JTH at all the microsatellite loci, except
at STM0019a where they were characterized by the
absence of a standard JTH allele (STM0019a-186).
Except for the (di)haploid C-13 that was heterozy-
gous for 40% of the loci tested, all the other clones
were with 60% of heterozygous loci.
Table 4 Allelic profiles at ten polymorphic microsatellite loci characterized in two androgenic (di)haploids, nine anther-derived
tetraploids and their two corresponding tetraploid anther donors of potato
Locus Number
of alleles
Number of clones with specified number of alleles Number of
genetic profiles
generated
Genotype
index (GI)aHeterozygote
frequency (HF)aPICa
1 2 3 4
POTM1-2 6 0 11 2 0 4 0.308 1.00 0.679
STM0015 5 0 10 2 1 4 0.308 1.00 0.662
STM0019ab 4 3 9 1 0 4 0.308 0.77 0.597
STM0019bb 3 1 11 1 0 4 0.308 0.92 0.566
STM1043 3 12 1 0 0 3 0.231 0.08 0.417
STM1069 2 1 12 0 0 2 0.154 0.92 0.373
STM3003 3 13 0 0 0 3 0.231 0.00 0.314
STWAX-1 3 11 2 0 0 3 0.231 0.15 0.314
STWAX-2 4 0 13 0 0 3 0.231 1.00 0.595
STWIN12G 2 13 0 0 0 2 0.154 0.00 0.152
Average 3.5 5.4 6.9 0.6 0.1 3.2 0.246 0.58 0.467
a GI number of genetic profiles generated/the total number of clones typed (13); HF number of clones with two or more alleles/the
total number of clones typed (13); PIC polymorphism information contentb Same as in Table 1
Euphytica (2010) 171:313–326 321
123
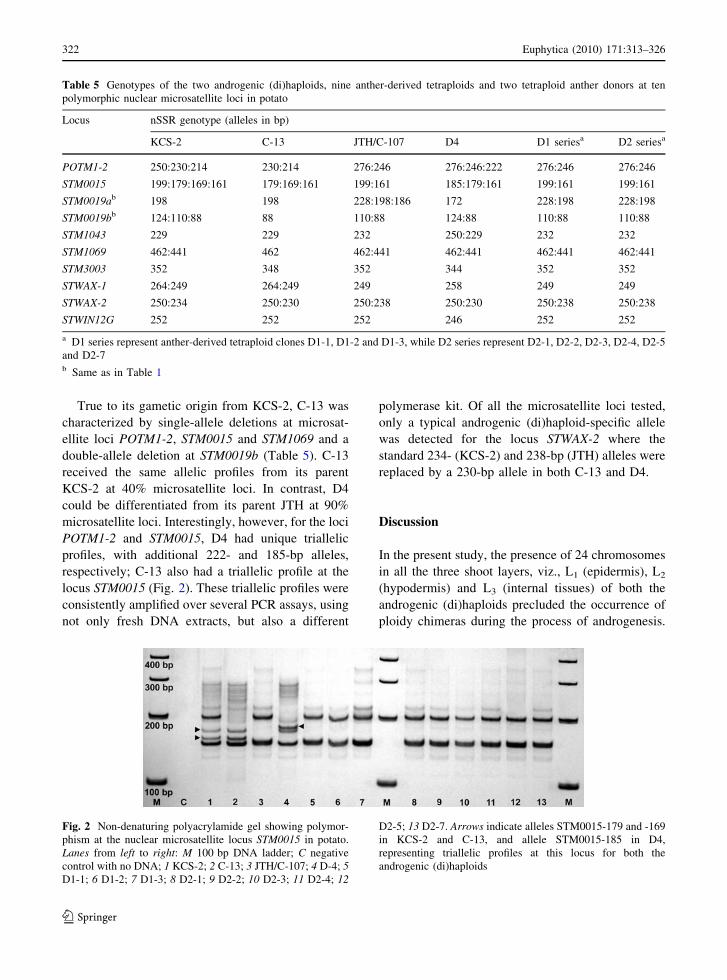
True to its gametic origin from KCS-2, C-13 was
characterized by single-allele deletions at microsat-
ellite loci POTM1-2, STM0015 and STM1069 and a
double-allele deletion at STM0019b (Table 5). C-13
received the same allelic profiles from its parent
KCS-2 at 40% microsatellite loci. In contrast, D4
could be differentiated from its parent JTH at 90%
microsatellite loci. Interestingly, however, for the loci
POTM1-2 and STM0015, D4 had unique triallelic
profiles, with additional 222- and 185-bp alleles,
respectively; C-13 also had a triallelic profile at the
locus STM0015 (Fig. 2). These triallelic profiles were
consistently amplified over several PCR assays, using
not only fresh DNA extracts, but also a different
polymerase kit. Of all the microsatellite loci tested,
only a typical androgenic (di)haploid-specific allele
was detected for the locus STWAX-2 where the
standard 234- (KCS-2) and 238-bp (JTH) alleles were
replaced by a 230-bp allele in both C-13 and D4.
Discussion
In the present study, the presence of 24 chromosomes
in all the three shoot layers, viz., L1 (epidermis), L2
(hypodermis) and L3 (internal tissues) of both the
androgenic (di)haploids precluded the occurrence of
ploidy chimeras during the process of androgenesis.
Table 5 Genotypes of the two androgenic (di)haploids, nine anther-derived tetraploids and two tetraploid anther donors at ten
polymorphic nuclear microsatellite loci in potato
Locus nSSR genotype (alleles in bp)
KCS-2 C-13 JTH/C-107 D4 D1 seriesa D2 seriesa
POTM1-2 250:230:214 230:214 276:246 276:246:222 276:246 276:246
STM0015 199:179:169:161 179:169:161 199:161 185:179:161 199:161 199:161
STM0019ab 198 198 228:198:186 172 228:198 228:198
STM0019bb 124:110:88 88 110:88 124:88 110:88 110:88
STM1043 229 229 232 250:229 232 232
STM1069 462:441 462 462:441 462:441 462:441 462:441
STM3003 352 348 352 344 352 352
STWAX-1 264:249 264:249 249 258 249 249
STWAX-2 250:234 250:230 250:238 250:230 250:238 250:238
STWIN12G 252 252 252 246 252 252
a D1 series represent anther-derived tetraploid clones D1-1, D1-2 and D1-3, while D2 series represent D2-1, D2-2, D2-3, D2-4, D2-5
and D2-7b Same as in Table 1
Fig. 2 Non-denaturing polyacrylamide gel showing polymor-
phism at the nuclear microsatellite locus STM0015 in potato.
Lanes from left to right: M 100 bp DNA ladder; C negative
control with no DNA; 1 KCS-2; 2 C-13; 3 JTH/C-107; 4 D-4; 5D1-1; 6 D1-2; 7 D1-3; 8 D2-1; 9 D2-2; 10 D2-3; 11 D2-4; 12
D2-5; 13 D2-7. Arrows indicate alleles STM0015-179 and -169
in KCS-2 and C-13, and allele STM0015-185 in D4,
representing triallelic profiles at this locus for both the
androgenic (di)haploids
322 Euphytica (2010) 171:313–326
123

The involvement of primary microspore embryogen-
esis in (di)haploid-derivation mechanism may be
responsible for this ploidy stability, which has been
otherwise shown to progress through varying degrees
of instabilities in potato anther cultures (Birhman
et al. 1994; Wenzel 1994; Calleberg 1996). The 2C
nDNA contents of both the androgenic (di)haploids
were about half the amounts of their corresponding
tetraploid anther donors. These values not only are in
good agreement with the estimates obtained in
tetraploid potatoes (Valkonen 1994), but also exactly
reflect the same variation between (di)haploids and
their corresponding tetraploids (Valkonen 1994;
Rokka et al. 1995, 1998). As we could not find any
cytological basis for a significant increase in nDNA
content in most of the anther-derived tetraploid
clones of JTH, these changes are perhaps associated
with somaclonal variation induced by somatic
embryogenesis via an intermediary callus phase.
Studies on various plant species have shown that
changes in nDNA content are related to growth
environmental stress and even somaclonal variation
(Cullis 1990).
In a cross-pollinated crop such as potato, genetic
variation among (di)haploids is expected because of
the high level of heterozygosity in the parental
cultivars (Ross 1986). The variation between the
(di)haploids and their corresponding tetraploid par-
ents is also common in potato. In the present study, in
the comparison between androgenic (di)haploids and
their tetraploid anther donors, the phenotypic varia-
tion was expressed at all the stages of plant growth.
This might have accrued from genomic changes
inherent in (di)haploid-derivation process, although
differential selection pressures during anther culture
(Rivard et al. 1994) cannot be ignored. A greater
selection pressure during monoploid-derivation pro-
cess through androgenesis than through gynogenesis
has been shown to occur in diploid potatoes (Lough
et al. 2001). The extent of phenotypic variation was
greater in D4 than in C-13. As JTH is an interspecific
hybrid, it is quite expected that the androgenic
process would result in yielding more variability in
its androgenic (di)haploid D4. Though both the
androgenic (di)haploids exhibited improved field
resistance to late blight, of particular interest is the
high level of field resistance expressed by C-13. This
points to extensive genomic rearrangements resulting
in the accumulation of favourable alleles in a gametic
phase under the sporophytic influence. The transmis-
sion and maintenance of virus resistance in potato
during haploidization via anther culture have been
reported (Rokka et al. 1995; Grammatikaki et al.
2007). The restoration of male fertility in androgenic
(di)haploids can be attributed to the mode of
(di)haploid-derivation mechanism through primary
microspore embryogenesis, perhaps coincident with
sporophytic determinants carried over and expressed
in the form of a high degree of meiotic stability
(Heberle-Bors 1985). Indeed, we could not detect any
apparent meiotic abnormalities in both the (di)hap-
loids. However, the inability to set berries in C-13
indicates that some sorts of (self) incompatibilities
might be operating at some levels. It is well known
that potato (di)haploids have an extremely low
capacity to produce berries (Uijtewaal et al. 1987;
Singh et al. 1988; Paz and Veilleux 1999; Phumichai
et al. 2005).
The microsatellite results are indicative of exten-
sive changes and rearrangements underlying andro-
genic (di)haploid-derivation process. The high
polymorphism averaged over ten microsatellite loci
is comparable with the results obtained from nSSR-
based cultivar discrimination for tetraploid potatoes
(Ashkenazi et al. 2001 and references therein). An
average genotype index of 0.246 is also comparable
with that reported for the four single-locus SSRs
(0.360) over 39 potato cultivars (McGregor et al.
2000). Since both the androgenic (di)haploids repre-
sent the gametes directly and SSR is a codominant
marker, it is possible to explain the reasons for such
high polymorphism based on recombination vis-a-vis
crossover events. The microsatellite locus POTM1-2
representing a dinucleotide repeat (AT) has been
mapped at the farthest (42.65 cM) from the centro-
mere (Chani et al. 2002) amongst the other loci
characterized in this study, for which gene-centro-
mere map positions are available. This provides a
strong basis for high polymorphism detected at this
locus because the probability of a crossover in
meiosis increases with distance from the centromere
(Chani et al. 2000), accepting the hypothesis of a
single crossover per chromosome arm in potato
microspores (Bastiaanssen et al. 1996). This also
explains the lowest level of polymorphism at the
locus STWIN12G representing a trinucleotide (AAT)
repeat, which has been mapped \33 cM from the
centromere (at 24.24 cM; Chani et al. 2002). The
Euphytica (2010) 171:313–326 323
123

similar frequency of heterozygous loci in D4 and its
parent JTH may be due to the fact that the later
represents an interspecific hybrid between two repro-
ductively isolated parental species (Veilleux et al.
1995). Incidentally, the frequency of heterozygous
loci was lower in C-13 than in KCS-2, an intraspe-
cific hybrid.
For the locus STWAX-2 representing a tetranucle-
otide (ACTC) repeat that has been mapped at
40.58 cM from the centromere (Chani et al. 2002)
on chromosome VIII, a 230-bp allele was favored in
both the androgenic (di)haploids. Since tetranucleo-
tide repeats are in general least susceptible to
polymerase template slippage (Valdes et al. 1993),
this mutant allele might have been generated due to
unequal crossing-over between sister chromatids. Its
presence in both the (di)haploids can be attributed to
the selection of alleles that influence regeneration
during androgenesis (Rivard et al. 1996). The fact
that all the JTH anther-derived tetraploids have the
common microsatellite allelic profiles that are true-
to-type to their tetraploid anther donor unambigu-
ously indicates their origin from somatic anther
tissues. If these anther-derived tetraploid clones
would have originated from reduced microspores
followed by spontaneous chromosome doubling
either during culture or subsequently, they would
have been expected to display microsatellite variation
not only between themselves, but also in relation to
their tetraploid anther donor JTH due to meiotic
recombination and segregation. In diploid potato
species, anther-derived diploids that originated from
somatic anther tissues were found to be genetically
identical to the diploid anther donor, except for
possible somaclonal variation (Veilleux et al. 1995;
Chani et al. 2000). Indeed, in the present study, the
deletion of a standard 186-bp allele at the locus
STM0019a in all the JTH anther-derived tetraploid
clones suggests the occurrence of somaclonal varia-
tion during the process of somatic embryogenesis
from anther walls. Somatic embryogenesis from
grapevine anthers was also associated with somacl-
onal variation as revealed by microsatellite analysis
(Bertsch et al. 2005).
In the present study, inordinately high allele
numbers for the loci POTM1-2 and STM0015 were
observed in androgenic (di)haploids. The presence of
more alleles than expected for a diploid species has
been related to higher ploidy levels (Ghislain et al.
2006). However, both C-13 and D4 are (di)haploids
in all the three shoot layers. A more probable
hypothesis explaining this phenomenon in D4 that
shows triallelic profiles at both the loci can be based
upon its chimeric structure, which might have been
formed through incomplete fusion of two different
pro-embryos during the first steps of microspore
division (Murigneux et al. 1993). It may perhaps
explains the basis for enormous phenotypic as well as
genetic variation observed in D4 as compared to its
tetraploid anther donor JTH/C-107. However, the
triallelic profile in C-13 for the locus STM0015 is
difficult to explain, even if one considers that its
tetraploid parent KCS-2 is a periclinal chimera
consisting of two different allelic profiles in L1 and
L2 cell layers, and this chimeric genotype has been
dissociated during floral bud formation (from the L2
cell layer). The separation of a periclinal chimera by
passage through anther-derived somatic embryogenic
calli has been detected in grapevine using microsat-
ellite markers (Bertsch et al. 2005). A detailed
microsatellite analysis of both C-13 and D4 in
relation to their parents using DNA from different
plant parts that represent distinct cell layers will be
carried out in our laboratory in the future.
Acknowledgments We thank Poonam Chandel for her
assistance in stomatal guard cell characterization, and Sheesh
Ram Thakur and Ram Dayal for greenhouse and field
management. We also thank Dr. P.H. Singh for his guidance
on the evaluation of field resistance to late blight disease and
Dr. P.S. Ahuja, Director, IHBT, Palampur, Himachal Pradesh
for allowing us to use the flow cytometry facility. The financial
assistance received from the Indian Council of Agricultural
Research (ICAR), New Delhi in the form of an ad-hoc research
project (F. No. 8-45/2004-Hort. II) is also acknowledged.
Comments and suggestions on the manuscript from an
anonymous reviewer are gratefully acknowledged.
References
Arumuganathan K, Earle ED (1991) Estimation of nuclear
DNA content of plants by flow cytometry. Plant Mol Biol
Rep 9:229–241
Ashkenazi V, Chani E, Lavi U, Levy D, Hillel J, Veilleux RE
(2001) Development of microsatellite markers in potato
and their use in phylogenetic and fingerprinting analyses.
Genome 44:50–62
Bastiaanssen HJM, Ramanna MS, Sawor Z, Mincione A, Steen
AVD, Jacobsen E (1996) Pollen markers for gene cen-
tromere mapping in diploid potato. Theor Appl Genet
93:1040–1047
324 Euphytica (2010) 171:313–326
123

Bertsch C, Kieffer F, Maillot P, Farine S, Butterlin G, Merdinoglu
D, Walter B (2005) Genetic chimerism of Vitis vinifera cv.
Chardonnay 96 is maintained through organogenesis but not
somatic embryogenesis. BMC Plant Biol 5:20
Birhman RK, Rivard SR, Cappadocia M (1994) Restriction
fragment length polymorphism analysis of anther-culture-
derived Solanum chacoense. HortScience 29:206–208
Calleberg E (1996) Anther culture of potato. Dissertation,
Swedish University of Agricultural Sciences
Campbell CL, Madden LV (1990) Introduction to plant disease
epidemiology. Wiley, New York
Cappadocia M, Ramulu KS (1998) Anther culture in potato
improvement. In: Paul Khurana SM, Chandra R, Upadhya
MD (eds) Comprehensive potato biotechnology. Malhotra
Publishing House, New Delhi, pp 1–30
Chani E, Veilleux RE, Boluarte-Medina T (2000) Improved
androgenesis of interspecific potato and efficiency of SSR
markers to identify homozygous regenerants. Plant Cell
Tissue Organ Cult 60:101–112
Chani E, Ashkenazi V, Hillel J, Veilleux RE (2002) Micro-
satellite marker analysis of an anther-derived potato
family: skewed segregation and gene-centromere map-
ping. Genome 45:236–242
Clulow SA, Wilkinson MJ, Waugh R, Baird E, DeMaine MJ,
Powell W (1991) Cytological and molecular observations
on Solanum phureja-induced dihaploid potatoes. Theor
Appl Genet 82:545–551
Cullis CA (1990) DNA rearrangements in response to envi-
ronmental stress. Adv Genet 28:73–97
De Maine MJ (1984) Patterns of variation in potato dihaploid
families. Potato Res 27:1–11
Ghislain M, Andrade D, RodrAguez F, Hijmans RJ, Spooner
DM (2006) Genetic analysis of the cultivated potato
Solanum tuberosum L. Phureja group using RAPDs and
nuclear SSRs. Theor Appl Genet 113:1515–1527
Grammatikaki G, Avgelis A, Sonnino A (2007) Behavior of
potato gametoclonal plants against the necrotic strain of
potato Y potyvirus. Russian J Plant Physiol 54:507–512
Heberle-Bors E (1985) In vitro haploid formation from pollen:
a critical review. Theor Appl Genet 71:361–374
Jacobsen E, Ramanna MS (1994) Production of monohaploids
of Solanum tuberosum L. and their use in genetics,
molecular biology and breeding. In: Bradshaw JE, Mac-
kay GR (eds) Potato genetics. CAB International, Wal-
lingford, pp 155–170
Kotch GP, Ortiz R, Peloquin SJ (1992) Genetic analysis by use
of potato haploid populations. Genome 35:103–108
Lough RC, Varrieur JM, Veilleux RE (2001) Selection inherent
in monoploid derivation mechanisms for potato. Theor
Appl Genet 103:178–184
Luo ZW, Zhang RM, Kearsey MJ (2004) Theoretical basis for
genetic linkage analysis in autotetraploid species. Proc
Natl Acad Sci USA 101:7040–7045
Matsubayashi M (1979) Genetic variation in dihaploid potato
clones, with special reference to phenotypic segregations
in some characters (preliminary report). Sci Rep Fac Agr
Kobe Univ 13:185–192
McGregor CE, Lambert CA, Greyling MM, Louw JH, Warnich
L (2000) A comparative assessment of DNA finger-
printing techniques (RAPD, ISSR, AFLP and SSR) in
tetraploid potato (Solanum tuberosum L.) germplasm.
Euphytica 113:135–144
Meyer R, Salamini F, Uhring H (1993) Isolation and character-
ization of potato diploid clones generating a high frequency
of monohaploid or homozygous diploid androgenic plants.
Theor Appl Genet 85:905–912
Murashige T, Skoog F (1962) A revised medium for rapid
growth and bioassays with tobacco tissue cultures. Physiol
Plant 15:473–497
Murigneux A, Barloy D, Leroy P, Beckert M (1993) Molecular
and morphological evaluation of doubled haploid lines in
maize. 1. Homogeneity within DH lines. Theor Appl
Genet 86:837–842
Naik PS, Sarkar D, Gaur PC (1998) Yield components of
potato microtubers: in vitro production and field perfor-
mance. Ann Appl Biol 133:91–99
Paz MM, Veilleux RE (1999) Influence of culture medium and
in vitro conditions on shoot regeneration in Solanumphureja monoploids and fertility of regenerated doubled
monoploids. Plant Breed 118:53–57
Pehu E, Veilleux RE, Hilu KW (1987) Cluster analysis of anther-
derived plants of Solanum phureja (Solanaceae) based on
morphological characteristics. Am J Bot 74:47–52
Phumichai C, Mori M, Kobayashi A, Kamijima O, Hosaka K
(2005) Toward the development of highly homozygous
diploid potato lines using the self-compatibility control-
ling Sli gene. Genome 48:977–984
Rivard SR, Saba-El-Leil MK, Landry BS, Cappadocia M
(1994) RFLP analyses and segregation of molecular
markers in plants produced by in vitro anther culture,
selfing, and reciprocal crosses of two lines of self-
incompatible Solanum chacoense. Genome 37:775–783
Rivard SR, Cappadocia M, Landry BS (1996) A comparison of
RFLP maps based on anther culture derived, selfed, and hybrid
progenies of Solanum chacoense. Genome 39:611–621
Rokka V-M, Valkonen JPT, Pehu E (1995) Production and
characterization of haploids derived from somatic hybrids
between Solanum brevidens and S. tuberosum through
anther culture. Plant Sci 112:85–95
Rokka V-M, Pietila L, Pehu E (1996) Enhanced production of
dihaploid lines via anther culture of tetraploid potato (Solanumtuberosum L. ssp. tuberosum) clones. Am Potato J 73:1–12
Rokka V-M, Ishimaru CA, Lapitan NLV, Pehu E (1998) Pro-
duction of androgenic dihaploid lines of the disomic tet-
raploid potato species Solanum acaule ssp. acaule. Plant
Cell Rep 18:89–93
Ross H (1986) Potato breeding-problems and perspectives.
Verlag Paul Parey, Berlin
Saghai-Maroof MA, Soliman KM, Jorgensen RA, Allard RW
(1984) Ribosomal DNA spacer-length polymorphisms in
barley: mendelian inheritance, chromosomal locations,
and population dynamics. Proc Natl Acad Sci USA 81:
8014–8018
Samitsu Y, Hosaka K (2002) Molecular marker analysis of 24-
and 25-chromosome plants obtained from Solanum tu-berosum L. ssp. andigena (2n = 4x = 48) pollinated with
a Solanum phureja haploid inducer. Genome 45:577–583
Sanguinetti CJ, Dias Neto E, Simpson AJG (1994) Rapid silver
staining and recovery of PCR products separated on
polyacrylamide gels. Biotechniques 17:915–919
Euphytica (2010) 171:313–326 325
123

Sarkar D, Maity S, Das PK (2004a) Induction of androgenic
embryoids from anther culture of Madhuca indica. J Trop
For Sci 16:429–436
Sarkar D, Pandey SK, Sud KC, Chanemougasoundharam A
(2004b) In vitro characterization of manganese toxicity in
relation to phosphorus nutrition in potato (Solanum tuberosumL.). Plant Sci 167:977–986
Sharma S, Chanemougasoundharam A, Sarkar D, Pandey SK
(2004) Carboxylic acids affect induction, development
and quality of potato (Solanum tuberosum L.) microtubers
grown in vitro from single-node explants. Plant Growth
Regul 44:219–229
Singh AK, Uhrig H, Salamini F (1988) Implications of chro-
mosome pairing in a monoploid and its colchidiploid of
Solanum tuberosum (x = 12). Genome 30:347–351
Song Y-S, Hepting L, Schweizer G, Hartl L, Wenzel G,
Schwarzfischer A (2005) Mapping of extreme resistance
to PVY (Rysto) on chromosome XII using anther-culture-
derived primary dihaploid potato lines. Theor Appl Genet
111:879–887
Uijtewaal BA, Jacobsen E, Hermsen JGTh (1987) Morphology
and vigour of monohaploids potato clones, their corre-
sponding homozygous diploids and tetraploids and their
heterozygous diploid parent. Euphytica 36:745–753
Valdes M, Slatkin M, Freimer NB (1993) Allele frequencies at
microsatellite loci: the stepwise mutation model revisited.
Genetics 133:737–749
Valkonen JPT (1994) Nuclear DNA content of the Solanumspp. in the series etuberosa as determined by laser flow
cytometry. Ann Appl Biol 125:589–600
Veilleux RE, Shen LY, Paz MM (1995) Analysis of the genetic
composition of anther-derived potato by randomly
amplified polymorphic DNA and simple sequence repeats.
Genome 38:1153–1162
Watanabe K, Orrillo M, Vega S, Masuelli R, Ishiki K (1994)
Potato germplasm enhancement with disomic tetraploid
Solanum acaule. II. Assessment to breeding value of tet-
raploid F1 hybrids between S. acaule and tetrasomic tet-
raploid potatoes. Theor Appl Genet 88:135–140
Waugh R, Baird E, Powell W (1992) The use of RAPD
markers for the detection of gene introgression in potato.
Plant Cell Rep 11:466–469
Wenzel G (1994) Tissue culture. In: Bradshaw E, Mackay GR
(eds) Potato genetics. CAB International, Wallingford, pp
173–195
Wilkinson MJ, Bennett ST, Clulow SA, Allainguillaume J,
Harding K, Bennett MD (1995) Evidence for somatic
translocation during potato dihaploid induction. Heredity
74:146–151
326 Euphytica (2010) 171:313–326
123
Copyright © 2022 FDOKUMEN




















