
COLORED POTATOES (SOLANUM TUBEROSUM L.) DRIED FOR ANTIOXIDANT-RICH VALUE-ADDED FOODS
Upload
independentCategory
view
2download
0
Abstract The effects of jasmonic acid (JA) and
methyl jasmonate (MeJA) on potato (Solanum
tuberosum L.) tuberization were studied in
relation to cytokinins using single-node cuttings
(SNCs) in vitro. In three potato cultivars differing
in maturity levels, JA or MeJA (0.0, 2.5, 5.0, 7.5
and 10.0 lM) action was examined with N6-
benzyladenine (BA: 0.0, 22.0 and 44.0 lM) under
optimum tuber-inducing treatment with 80 g L–1
sucrose. Although jasmonates had a stimulatory
role in root growth from SNCs, BA inhibited the
root growth and antagonized the jasmonates
action on root growth promotion. There was a
strong inhibitory effect of BA on stoloniferous
shoot growth, and in combination with jasmo-
nates it could exert a much pronounced inhibitory
effect. Jasmonates did not have any role in tuber
induction in terms of tuber number and source-
sink coordination (harvest index), rather their
effects were counteracted in the presence of
22.0 lM BA in early cultivar. However, they
had a promoting effect on tuber growth after
induction in early cultivar, possibly due to
its lower levels of endogenous gibberellins.
Cytokinins detrimentally affected the tuber
growth, and antagonized the jasmonates action
irrespective of the maturity levels of the culti-
vars. It could, however, increase the tuber dry
matter concomitant with a higher accumulation
of starch. The promoting effect of jasmonates on
tuber dry matter and starch accumulation was
apparent only in early cultivar. There were
significant interactions between cytokinin and
jasmonates for accumulation of reducing and
total sugars in tubers. The results show that
exogenous cytokinins antagonize the jasmonates
action, and the correlated effects of these two
growth hormones interact with the maturing
time of the cultivar during potato tuber forma-
tion in vitro.
Keywords Cytokinins Æ Jasmonic acid Æ Methyl
jasmonate Æ Solanum tuberosum L. Æ Starch ÆTuberization
AbbreviationsBA N6-benzyladenine
JA Jasmonic acid
KA Kufri Ashoka
KBD Kufri Badshah
KS Kufri Sindhuri
MeJA Methyl jasmonate
MS Murashige and Skoog’s medium (1962)
SNC Single-node cutting
D. Sarkar (&) Æ S. K. Pandey Æ S. SharmaDivision of Crop Improvement, Central PotatoResearch Institute, Shimla,171 001 Himachal Pradesh, Indiae-mails: [email protected]; [email protected]
Plant Cell Tiss Organ Cult (2006) 87:285–295
DOI 10.1007/s11240-006-9166-3
123
ORIGINAL PAPER
Cytokinins antagonize the jasmonates actionon the regulation of potato (Solanumtuberosum) tuber formation in vitro
Debabrata Sarkar Æ Suman Kumar Pandey ÆSushruti Sharma
Received: 9 April 2006 / Accepted: 12 September 2006 / Published online: 10 November 2006� Springer Science+Business Media B.V. 2006

Introduction
Besides specific environmental cues, such as
short-photoperiod, high-light intensity and low-
nitrogen level, the growth hormones have been
suggested to play a prominent role in the control
of tuberization in potato (Vreugdenhil and
Struik 1989). However, no unequivocal tuberizing
factor has been identified till date, and, therefore,
the recognition of a tuber-inducing signal still
remains an elusive goal (Jackson 1999). Tuberonic
acid (TA) and tuberonic acid glucoside (TAG)
with strong tuber-inducing activities were isolated
from potato leaves (Koda and Okazawa 1988;
Yoshihara et al. 1989). TA was later found to be
structurally related to jasmonic acid (JA) having
similar levels of tuber-inducing activity in vitro
(Koda et al. 1991). JA and its volatile derivative
methyl jasmonate (MeJA) have been detected in a
large number of plant species (Mithofer et al.
2005). They are involved in various morphogenic
events, such as tuberization, tuberous root forma-
tion, bulb formation (Koda 1997) and its dor-
mancy development (Jasik and de Klerk 2006),
senescence, developmental process, wounding,
plant structural determination, thigmomorpho-
genesis (van den Berg and Ewing 1991; Koda
1997), abiotic stress responses and mechanotrans-
duction (Wasternack and Hause 2002). In potato
stolons, shoot cuttings and plantlets cultured in
vitro, JA stimulated tuberization (Koda et al.
1991; Pelacho and Mingo-Castel 1991; Ravnikar
et al. 1992; Pruski et al. 2001, 2002). Even its
endogenous presence was confirmed in roots,
stolons and periderm of newly formed tubers
(Abdala et al. 1996). However, a number of
studies demonstrated that JA was not directly
involved in potato tuberization (Helder et al.
1993; Jackson and Willmitzer 1994).
Now, it is well known that potato tuberization
is regulated by a balance between the levels of
jasmonates and gibberellin (Koda 1997). JA
exerts its effect principally by antagonizing the
effect of GA on microtubule orientation during
potato tuberization (Jackson 1999). It has also
been shown that JA caused a total reversion of
the delaying effect of GA3 on tuberization of
potato shoot cuttings in vitro, and the stimulatory
effect of JA on potato tuberization was antago-
nized in rooted plantlets having a high level of
endogenous GAs (Castro et al. 2000).
Cytokinins have long been suggested to be
involved in potato tuberization (Palmer and Smith
1969), but in contrast to GA and ABA less
attention has been paid to cytokinins. A direct
role of cytokinins in tuberization vis-a-vis tuber
induction has been confirmed by transgenic
approach (Vreugdenhil 2004). For in vitro
tuberization, exogenous cytokinins, especially
N6-benzyladenine (BA), are extensively used to
induce microtubers on a variety of explants
including stolons, shoot cuttings and intact
microplantlets (see review Donnelly et al. 2003).
However, although the presence of cytokinin is
required for tuberization, it is not the stimulus
provided by photoperiodic induction. This essen-
tially shows that cytokinins cannot be the only
factor regulating tuber induction in potato. The
response of a stolon to high-cytokinin levels
depends on the interaction with other hormones,
particularly the levels of GAs (Vreugdenhil and
Struik 1989). It has also been shown that exogenous
cytokinins do not have any role in stolon elonga-
tion and tuber formation in vitro implying that
cytokinins are not the limiting factor for tuber
formation (Xu et al. 1998).
It is not known how the jasmonates (JA and
MeJA) regulate tuber induction and development
in relation to cytokinins. Pelacho and Mingo-
Castel (1991) showed that JA was the stronger
promoter of stolon tuberization in vitro than
kinetin. But the concentration of kinetin tested by
them was about four-times lesser than that
optimally used for cytokinin-induced tuberization
in vitro. Is there any interaction between jasmo-
nates and cytokinins for tuberization in potato? It
has also been proved that BA is more potent than
kinetin for tuberization in vitro. Moreover, in
comparison with JA, the role of MeJA in potato
tuberization is also least investigated to our
knowledge.
In this study, we employed a well-defined in
vitro tuberization system using single-node
cuttings (SNCs) to explore the interaction
between JA/MeJA and BA for tuber formation
in potato. The objectives were (1) to assess the
286 Plant Cell Tiss Organ Cult (2006) 87:285–295
123

comparative efficacy of JA and MeJA for
tuberization under cytokinin-aided inducing and
non-inducing conditions and (2) to examine the
individual and correlated effects of JA/MeJA
and BA on tuber formation in relation to
sucrose-aided optimum tuber-inducing condition.
The study also goes in some way towards
examining the effects JA and MeJA on tuber
formation from SNCs, as compared to that from
stolons and leafy shoot cuttings employed in
other studies.
Materials and methods
Plant material
In potato (Solanum tuberosum L.), tuberization in
vitro is cultivar-specific, essentially modulated by
maturity levels with distinct differences in
response between early and maincrop cultivars
(Naik et al. 1998). Therefore, three tetraploid
(2n = 4x = 48) Indian potato (S. tuberosum
L. subsp. tuberosum) cultivars belonging to dif-
ferent maturity groups, viz., Kufri Ashoka (KA;
early maturing), Kufri Badshah (KBD; medium
maturing) and Kufri Sindhuri (KS; late maturing)
were used in the present study.
Culture of single-node cuttings in vitro
In vitro culture of these three cultivars were
established from indexed tubers (sprouts), and
they were made virus-free following standard
meristem culture and virus diagnostic procedures
as described earlier (Sarkar et al. 2004). SNCs
from disease-free microplants were cultured in
vitro as described by Sarkar et al. (1997). The
basal medium consisted of MS medium (Murash-
ige and Skoog 1962) supplemented with 20 g L–1
sucrose and solidified with 2 g L–1 gelrite (Sigma,
St. Louis, MI, USA). Five SNCs were cultured
per Magenta� vessel (GA-7; Sigma) containing
50 mL of medium. The cultures were incubated
under a 16 h photoperiod from cool white
fluorescent lights (~50–60 lmol m–2 s–1 PPFD)
at 24 ± 1�C and subcultured on fresh medium at
an interval of 21 days.
In vitro tuberization
For tuber induction in vitro, SNCs with a leaf
subtending the axillary buds from 21-day-old
microplants grown under above conditions were
used as explants. To maintain the explant homo-
geneity in relation to tuber induction and devel-
opment, the SNCs were prepared from middle
nodes of the source microplants. They were
layered horizontally over 30 mL of microtuber
induction medium (Sarkar and Naik 1998) in
90 mm disposable Petri dishes (Greiner Bio-One,
Frickenhausen, Germany). The induction medium
consisted of MS basal medium supplemented with
80 g L–1 sucrose and different concentrations of
BA plus jasmonates (JA and or MeJA) and
solidified with 2 g L–1 gelrite. The presence of
80 g L–1 sucrose in the medium provided an
optimum tuber-inducing conditions in vitro to
counteract the effects of endogenous GAs. Tuber
induction and development were also tested in
hormone-free medium supplemented only with
80 g L–1 sucrose. The pH of the medium was
adjusted to 5.8 before autoclaving at 121�C for
20 min. The Petri dishes were sealed with Parafilm
MTM
(American National Can, Chicago, USA),
and the tuber induction cultures were incubated in
the dark at 20�C (Sarkar and Naik 1998).
Jasmonates and cytokinin bioassays
In the present study, both (±)-JA (Sigma) and
MeJA (Duchefa, Haarlem, The Netherlands)
were supplemented in tuber induction medium
at varying combinations with BA (Sigma). Five
different concentrations of JA or MeJA (0.0, 2.5,
5.0, 7.5 and 10.0 lM) were tested with three
different concentrations of BA (0.0, 22.0 and
44.0 lM). The growth hormones were applied to
the medium by filter sterilization. The standard
concentration of BA for cytokinin-induced in
vitro tuberization is 44.0 lM. Many workers have
confirmed that optimum microtuberization occurs
at this concentration, especially in the absence of
light (Wang and Hu 1982; Wattimena 1983;
Slimmon et al. 1989; Sarkar and Naik 1998).
However, when growth retardants, like CCC is
used, the optimum concentration of BA has been
found to be half of this standard concentration,
Plant Cell Tiss Organ Cult (2006) 87:285–295 287
123

i.e. 22.0 lM (Estrada et al. 1986). Therefore, in
the present study, BA was tested at both 22.0 and
44.0 lM. For each jasmonate treatment, the
experiment was laid out in a factorial (5 · 3 · 3)
completely randomized design (CRD) involving
five concentrations of JA or MeJA, three
concentrations of BA and three cultivars with
five replicate Petri dishes, each containing ten
SNCs.
Data observations
The total duration of in vitro (tuber) induction
cultures was 8 weeks. The in vitro-induced
tubers were harvested, and the observations
were recorded on number of tubers per SNC,
average tuber fresh weight (mg), average stolon
fresh weight (mg), average root fresh weight
(mg) and harvest index (the ratio of tuber yield
to total biomass measured as fresh weight). The
harvested tubers were dried at 70�C for 2 days
for the estimation of percentage tuber dry
matter.
Sugars and starch determination
Sugars and starch in freshly harvested tubers
were estimated as described by Sharma et al.
(2004). Total soluble sugars, reducing sugars
and starch were expressed as mg glucose
equivalents per g microtuber fresh weight. For
each treatment, there were three independent
replications for the estimation of soluble sugars
and starch.
Statistical analyses
Prior to statistical analyses, the data on average
stolon fresh weight (mg), average root fresh
weight and tuber dry matter (%) were trans-
formed into square roots. For each jasmonate
(JA or MeJA) treatment, the three-way analy-
ses of variance (ANOVAs) were calculated
using the standard procedure, and the
means were separated by Least Significant
Difference (LSD) test. The term significant has
been used to indicate differences for which
p £ 0.05.
Results
All tested factors, viz., cultivar, JA/MeJA and BA
significantly affected tuber induction and devel-
opment from SNCs. The main effects of BA were
highly significant for tuber induction, growth and
biochemical parameters. JA had a significant
interaction with BA for number of tubers per
SNC, tuber weight, stolon fresh weight, root fresh
weight and tuber dry matter (%). For MeJA, the
significant interactions with BA were apparent
only for root fresh weight, harvest index and
tuber dry matter (%). Since three-way interac-
tions involving the cultivar, jasmonates and BA
were significant for most of the characters, only
the means and mean comparisons for the three-
way interactions are presented.
Effects of jasmonates on root
and stoloniferous shoot growth
JA significantly stimulated root growth, measured
as fresh weight, from SNCs. Independent of the
cultivars, the stimulatory effect of JA on root
growth was apparent up to 5.0 lM. Averaged
over the cultivars, about three-times increase in
root growth occurred at 5.0 lM JA (75.0 mg FW/
SNC) as compared to control 80 g L–1 sucrose
medium (25.5 mg FW/SNC). When the medium
contained 22.0–44.0 lM BA, rooting was com-
pletely inhibited, and JA even at higher concen-
trations (‡5.0 lM) could not repress this BA-
induced inhibitory effect. MeJA also had a
stimulatory effect on root growth, but its effect
was not so much pronounced (40.0 mg FW/SNC
at 7.5 lM MeJA averaged over the cultivars) and
uniform over the cultivars. Like JA, it also failed
to repress the inhibitory effect of BA on root
growth, especially in medium (KBD) and late
(KS) cultivars.
JA significantly inhibited stoloniferous shoot
(SS) growth, measured as fresh weight, even at
2.5 lM in early (KA) and late (KS) cultivars. The
addition of BA to the induction medium resulted
in a considerable decline in SS growth (2.4–6.8-
fold reduction as compared to the control). When
the BA-medium contained 2.5 lM JA, there was
a further decline in SS growth in all the cultivars.
The inhibitory effect of MeJA on SS growth was
288 Plant Cell Tiss Organ Cult (2006) 87:285–295
123

observed at higher concentrations (7.5–10.0 lM)
only in early cultivar (KA). In general, MeJA did
not have any effect on BA-induced inhibition of
SS growth, except in early cultivar (KA) where it
could significantly promote the SS growth at
5.0 lM.
Effects of jasmonates on tuber induction
and development
In cytokinin-aided in vitro tuberization under
optimum tuber-inducing treatment with 80 g L–1
sucrose, generally a single tuber is induced per
SNC. Therefore, it is not surprising that there was
not much variation in tuber number per SNC in
the present experiments. Both JA and MeJA did
not have any effect on tuber number; however,
there was a significant JA · BA interaction for
tuber induction (Fig. 1). JA at 2.5–5.0 lM was
found to inhibit tuber induction when the medium
contained 22.0 lM BA, but this JA-induced
inhibitory effect on tuber induction did not occur
in the presence of 44.0 lM BA. The inhibitory
effect of MeJA on tuber induction in the presence
of 22.0 lM BA was observed only in early cv. KA
(data not shown).
For tuber growth measured as fresh weight, the
promoting effect of JA was observed only in early
cultivar, but not in medium and late ones (Figs. 1,
2a). However, at higher concentrations, it not
only caused a reduction in tuber growth, but also
induced morphological changes in tuber shape
(Fig. 2b). In general, BA inhibited tuber growth.
Irrespective of the cultivars, an inhibition of tuber
growth occurred when JA was supplemented in
the medium with BA, although its effect was not
uniform over the BA levels (Figs. 1, 2c, d). The
trend of MeJA effect on tuber growth was similar
to that of JA effect; at 2.5 lM it had a promoting
effect on tuber growth only in early cultivar when
the nutrient medium contained only 80 g L–1
sucrose (Fig. 1). There was no significant Me-
JA · BA interaction, and in combination with
BA it depressed the tuber growth further only in
early cultivar (KA).
Harvest index is a function of total biomass
(source) and tuber yield (sink) in vitro. In the
present study, BA was found to significant
increase the harvest indices in all the cultivars
(Fig. 1). Both JA and MeJA did not have any
positive effect on harvest index. In combination
with BA, JA did not show any synergistic effect
on harvest index in relation to what obtained
when BA was supplemented alone in the med-
ium. In medium (KBD) and late (KS) cultivars,
there was a significant decline in harvest index
with increasing concentrations of MeJA (Fig. 1).
Effects of jasmonates on tuber quality
Tuber dry matter was between 15 and 17% when
the tubers were induced on medium containing
only 80 g L–1 sucrose (data not shown). BA
caused a significant increase in tuber dry matter
(19.2–22.9%) in medium (KBD) and late (KS)
cultivars. When JA was supplemented at 2.5 lM,
there was a significant increment in tuber dry
matter (20.5%) only in early cultivar (KA). Like
JA, MeJA at 2.5 lM also had a promoting effect
on tuber dry matter (18.1% as compared to
16.2% in the control) in early cultivar. In general,
MeJA in combination with BA significantly
increased the tuber dry matter in all the cultivars,
although this promoting effect was more conspic-
uous in medium (20.9–25.3%) and late cultivars
(22.8–25.5%). In contrast, the combined effect of
JA and BA on tuber dry matter was not so much
prominent as that of MeJA and BA.
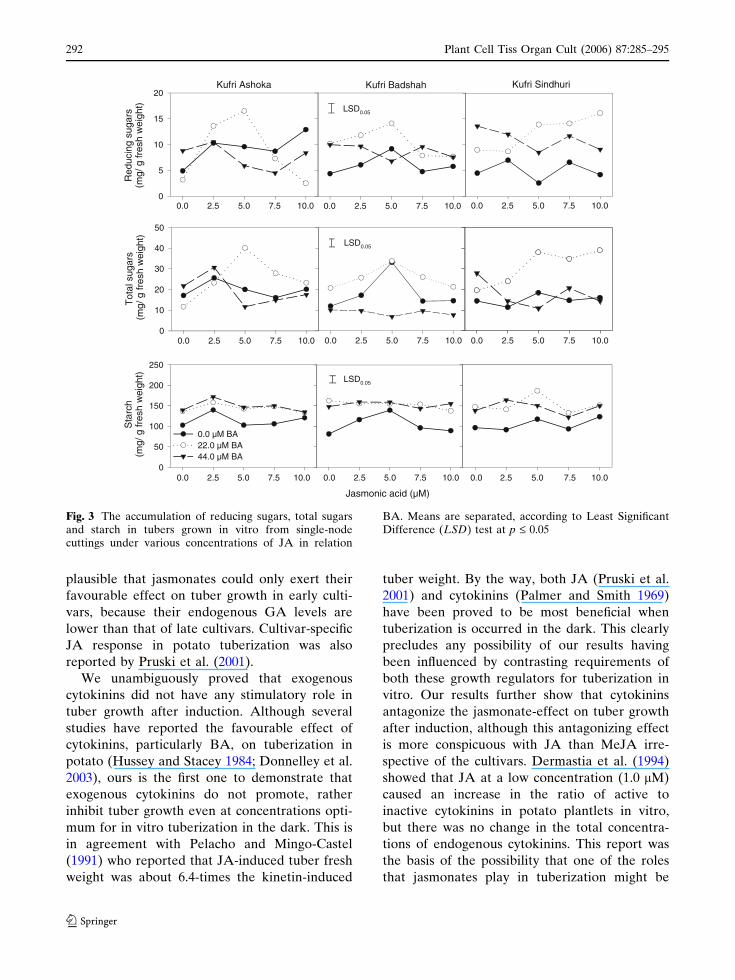
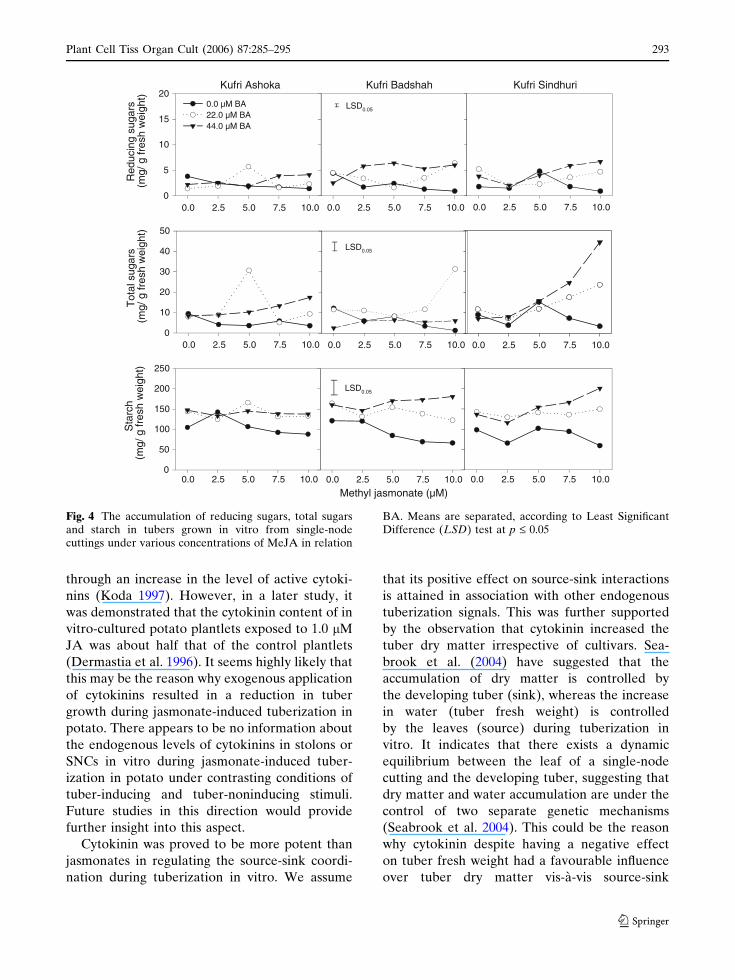
There were significant interactions between
jasmonates and BA for reducing sugars, total
sugars and starch in tubers induced on SNCs in
vitro. In general, BA was found to have a
promoting effect on reducing and total sugar
levels in tubers (Figs. 3, 4). JA significantly
increased the levels of reducing and total sugars
in all the cultivars at 2.5–5.0 lM when the
medium contained only 80 g L–1 sucrose
(Fig. 4). In the presence of BA, JA could exert
a further promoting effect on reducing and total
sugar contents. No significant promoting effect of
MeJA on reducing and total sugar contents could
be detected in the present study (Fig. 4).
When the tubers were induced on medium
containing only 80 g L–1 sucrose, the tuber starch
content was in the range of 80–120 mg g–1 FW
(Figs. 3, 4). There was a significant increase in
starch content when the medium contained BA.
On medium containing 2.5–5.0 lM JA there was
Plant Cell Tiss Organ Cult (2006) 87:285–295 289
123

a significant increase in starch content in all
the cultivars, as compared to that obtained at
80 g L–1 sucrose-medium. In the presence of
22.0 lM BA, JA increased the starch levels
further, and increasing BA level did not have
any further promoting effect on starch levels. In
contrast, MeJA could increase the starch
content only in early cultivar (KA). However,
Jasmonic acid (µM)
0.0 2.5 5.0 7.5 10.0
Har
vest
inde
x
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Tub
er fr
esh
wei
ght (
mg)
0
50
100
150
200
250
300
0.0 2.5 5.0 7.5 10.0
Har
vest
inde
x
0.0
0.2
0.4
0.6
0.8
1.0
1.2
0.0 2.5 5.0 7.5 10.0
Methyl jasmonate (µM)
LSD0.05
LSD0.05
Kufri Ashoka Kufri Badshah Kufri Sindhuri
Tub
ers
per
expl
ant
0.0
0.5
1.0
1.5
LSD0.05
Tub
er fr
esh
wei
ght (
mg)
0
50
100
150
200
250
300
LSD0.05
0.0 2.5 5.0 7.5 10.0 0.0 2.5 5.0 7.5 10.0
LSD0.5
0.0 2.5 5.0 7.5 10.0
0.0 µM BA22.0 µM BA44.0 µM BA
Fig. 1 Effects of jasmonic acid and methyl jasmonate ontuber induction and development from single-node cut-tings in relation to BA in three potato cultivars differing in
maturity levels. Means are separated, according to LeastSignificant Difference (LSD) test at p £ 0.05
290 Plant Cell Tiss Organ Cult (2006) 87:285–295
123

MeJA at 5.0–10.0 lM could significantly in-
crease the starch levels in the presence of
22.0 lM BA, irrespective of the cultivars
(Fig. 4).
Discussion
Although several reports have been published
concerning the effect of JA on potato tuberiza-
tion (Koda et al. 1991; Pelacho and Mingo-Castel
1991; Ravnikar et al. 1992; Helder et al. 1993;
Jackson and Willmitzer 1994; Castro et al. 2000;
Pruski et al. 2001, 2002), our study is the first one
to our knowledge to analyze the efficacy of
jasmonates (JA and MeJA) for potato tuber
induction and development in relation to exoge-
nous cytokinins. Our results clearly showed that
jasmonates did not affect tuber induction. Neither
did they have any favourable effect on in vitro
source-sink coordination. The inhibitory effects
of jasmonates on tuber induction in the presence
of 22.0 lM BA, especially in early cultivar is
difficult to explain, but may perhaps be due to an
altered balance of ethylene/gibberellin ratio
(Vreugdenhil and Struik 1989). However, the
promoting effects of both JA and MeJA on tuber
growth only in early cultivar suggested that their
effects are maturity-specific. A favourable effect
of JA on tuber growth after induction was also
reported by Pelacho and Mingo-Castel (1991).
Koda and Kikuta (2001) observed that JA-
induced in vitro tuberization in potato was
dependent on the maturing time of the cultivar;
the later the maturing time of the cultivar, the
lower the response. The relative higher JA-
responsiveness of early cultivar is assumed to be
due to the lower levels of endogenous GAs (Koda
1997; Koda and Kikuta 2001), which are known to
antagonize the JA action (Jackson 1999; Castro
et al. 2000). Since relatively high GA levels
reduce or stop tuber growth, it seems highly
Fig. 2 Development of tubers from single-node cuttingsof potato cv. Kufri Ashoka in vitro on nutrient mediasupplemented with sucrose and various combinations ofJA plus BA after 8 weeks of culture. (a) Eighty g L–1
sucrose (bottom) and 80 g L–1 sucrose plus 2.5 lM JA(top). (b) Eighty g L–1 sucrose plus 10.0 lM JA. (c) Eighty
g L–1 sucrose plus 22.0 lM BA and a series of JAconcentrations: 2.5, 5.0, 7.5 and 10.0 lM JA from left toright. (d) Eighty g L–1 sucrose plus 44.0 lM BA and aseries of JA concentrations: 2.5, 5.0, 7.5 and 10.0 lM JAfrom left to right
Plant Cell Tiss Organ Cult (2006) 87:285–295 291
123
plausible that jasmonates could only exert their
favourable effect on tuber growth in early culti-
vars, because their endogenous GA levels are
lower than that of late cultivars. Cultivar-specific
JA response in potato tuberization was also
reported by Pruski et al. (2001).
We unambiguously proved that exogenous
cytokinins did not have any stimulatory role in
tuber growth after induction. Although several
studies have reported the favourable effect of
cytokinins, particularly BA, on tuberization in
potato (Hussey and Stacey 1984; Donnelley et al.
2003), ours is the first one to demonstrate that
exogenous cytokinins do not promote, rather
inhibit tuber growth even at concentrations opti-
mum for in vitro tuberization in the dark. This is
in agreement with Pelacho and Mingo-Castel
(1991) who reported that JA-induced tuber fresh
weight was about 6.4-times the kinetin-induced
tuber weight. By the way, both JA (Pruski et al.
2001) and cytokinins (Palmer and Smith 1969)
have been proved to be most beneficial when
tuberization is occurred in the dark. This clearly
precludes any possibility of our results having
been influenced by contrasting requirements of
both these growth regulators for tuberization in
vitro. Our results further show that cytokinins
antagonize the jasmonate-effect on tuber growth
after induction, although this antagonizing effect
is more conspicuous with JA than MeJA irre-
spective of the cultivars. Dermastia et al. (1994)
showed that JA at a low concentration (1.0 lM)
caused an increase in the ratio of active to
inactive cytokinins in potato plantlets in vitro,
but there was no change in the total concentra-
tions of endogenous cytokinins. This report was
the basis of the possibility that one of the roles
that jasmonates play in tuberization might be
0.0 2.5 5.0 7.5 10.0
Red
ucin
g su
gars
(mg/
g fr
esh
wei
ght)
0
5
10
15
20
0.0 2.5 5.0 7.5 10.0 0.0 2.5 5.0 7.5 10.0
0.0 2.5 5.0 7.5 10.0
Tot
al s
ugar
s(m
g/ g
fres
h w
eigh
t)
0
10
20
30
40
50
0.0 2.5 5.0 7.5 10.0 0.0 2.5 5.0 7.5 10.0
0.0 2.5 5.0 7.5 10.0
Sta
rch
(mg/
g fr
esh
wei
ght)
0
50
100
150
200
250
0.0 2.5 5.0 7.5 10.0 0.0 2.5 5.0 7.5 10.0
0.0 µM BA22.0 µM BA44.0 µM BA
Jasmonic acid (µM)
LSD0.05
LSD0.05
LSD0.05
Kufri Ashoka Kufri Badshah Kufri Sindhuri
Fig. 3 The accumulation of reducing sugars, total sugarsand starch in tubers grown in vitro from single-nodecuttings under various concentrations of JA in relation
BA. Means are separated, according to Least SignificantDifference (LSD) test at p £ 0.05
292 Plant Cell Tiss Organ Cult (2006) 87:285–295
123
through an increase in the level of active cytoki-
nins (Koda 1997). However, in a later study, it
was demonstrated that the cytokinin content of in
vitro-cultured potato plantlets exposed to 1.0 lM
JA was about half that of the control plantlets
(Dermastia et al. 1996). It seems highly likely that
this may be the reason why exogenous application
of cytokinins resulted in a reduction in tuber
growth during jasmonate-induced tuberization in
potato. There appears to be no information about
the endogenous levels of cytokinins in stolons or
SNCs in vitro during jasmonate-induced tuber-
ization in potato under contrasting conditions of
tuber-inducing and tuber-noninducing stimuli.
Future studies in this direction would provide
further insight into this aspect.
Cytokinin was proved to be more potent than
jasmonates in regulating the source-sink coordi-
nation during tuberization in vitro. We assume
that its positive effect on source-sink interactions
is attained in association with other endogenous
tuberization signals. This was further supported
by the observation that cytokinin increased the
tuber dry matter irrespective of cultivars. Sea-
brook et al. (2004) have suggested that the
accumulation of dry matter is controlled by
the developing tuber (sink), whereas the increase
in water (tuber fresh weight) is controlled
by the leaves (source) during tuberization in
vitro. It indicates that there exists a dynamic
equilibrium between the leaf of a single-node
cutting and the developing tuber, suggesting that
dry matter and water accumulation are under the
control of two separate genetic mechanisms
(Seabrook et al. 2004). This could be the reason
why cytokinin despite having a negative effect
on tuber fresh weight had a favourable influence
over tuber dry matter vis-a-vis source-sink
0.0 2.5 5.0 7.5 10.0
Red
ucin
g su
gars
(mg/
g fr
esh
wei
ght)
0
5
10
15
20
0.0 2.5 5.0 7.5 10.0 0.0 2.5 5.0 7.5 10.0
0.0 2.5 5.0 7.5 10.0
Tot
al s
ugar
s(m
g/ g
fres
h w
eigh
t)
0
10
20
30
40
50
0.0 2.5 5.0 7.5 10.0 0.0 2.5 5.0 7.5 10.0
0.0 2.5 5.0 7.5 10.0
Sta
rch
(mg/
g fr
esh
wei
ght)
0
50
100
150
200
250
0.0 2.5 5.0 7.5 10.0 0.0 2.5 5.0 7.5 10.0
0.0 µM BA22.0 µM BA44.0 µM BA
Methyl jasmonate (µM)
LSD0.05
LSD0.05
LSD0.05
Kufri Ashoka Kufri Badshah Kufri Sindhuri
Fig. 4 The accumulation of reducing sugars, total sugarsand starch in tubers grown in vitro from single-nodecuttings under various concentrations of MeJA in relation
BA. Means are separated, according to Least SignificantDifference (LSD) test at p £ 0.05
Plant Cell Tiss Organ Cult (2006) 87:285–295 293
123

coordination. Cytokinin was also found to in-
crease the starch accumulation in tubers. As
starch is the major component of the dry matter
content of potato tubers accounting for about
70% of the total solids, this may explain the
occurrence of high-dry matter in cytokinin-
induced tubers in vitro. The beneficial effect of
jasmonates on tuber dry matter was apparent only
in early cultivar in which they had also stimula-
tory influence over tuber growth. Incidentally,
the promoting effects of jasmonates on starch
accumulation in tubers at relatively low concen-
trations were also evident in early cultivar. This
again substantiates the beneficial role of jasmo-
nates in tuber growth after induction depending
on the maturing time of the cultivars through a
regulation of endogenous GA levels. Our results
on starch accumulation in relation to jasmonates
and cytokinins based on in vitro bioassays can
well generalize their effects on potato tuberiza-
tion in vivo since starch synthesis and composi-
tion in tubers grown in vitro have been shown to
be identical to field-grown tubers (Veramendi
et al. 1999).
We also observed that both cytokinin and JA
had a promoting effect on the accumulation of
reducing and total sugars in tubers. The biochem-
ical machinery for starch synthesis and break-
down is present during all stages of tuber
development, and is coordinately up- or down-
regulated (Vreugdenhil 2004). Therefore, it is not
surprising as to why these two growth hormones
resulted in a high levels of sugars concomitant
with increased starch accumulation in tubers
induced in vitro. However, cytokinin, JA and
cultivars interacted among themselves to regulate
the levels of reducing sugars, total sugars and
starch in tubers. This invariably indicates that
these two growth hormones affect endogenous
sugar and starch levels of developed tubers
depending on the maturing time of the cultivars.
However, it must be noted that our study
describes the partitioning of sugars and starch in
tubers after they have developed fully vis-a-vis
matured in vitro, and therefore, might be its
result, not the cause. A time-course estimation of
the partitioning of sugars and starch during
various stages of tuber development from single-
node cuttings under varying treatments of exog-
enous cytokinins and jasmonates will be needed
to examine the specific roles of these two growth
hormones in tuber formation in vitro. Reducing
sugar levels can be an estimate of the maturity of
tubers grown in vitro (Xiong et al. 2002; Sharma
et al. 2004). Since both BA and JA caused an
increase in reducing sugars in tubers, their yet
another role in tuberization can be attributed to
tuber maturity, possibly through an endogenous
regulation of GA: ABA balance. However, fur-
ther investigation is needed to study the effect of
jasmonates in relation to cytokinins on tuber
dormancy release.
Acknowledgements Comments and suggestions on themanuscript from the Editor and two anonymous reviewersare gratefully acknowledged.
References
Abdala G, Castro G, Guinazu R, Tizio R, Miersch O(1996) Occurrence of jasmonic acid in organs ofSolanum tuberosum L. and its effect on tuberization.Plant Growth Regul 19:139–143
Castro G, Abdala G, Aguero C, Tizio R (2000) Interactionbetween jasmonic and gibberellic acids on in vitrotuberization of potato plantlets. Potato Res 43:83–88
Dermastia M, Ravnikar M, Vilhar B, Kovac M (1994)Increased level of cytokinin ribosides in jasmonicacid-treated potato (Solanum tuberosum) stem nodecultures. Physiol Plant 92:241–246
Dermastia M, Ravnikar M, Kovac M (1996) Morphologyof potato (Solanum tuberosum L. cv. Sante) stemnode cultures in relation to the level of endogenouscytokinins. J Plant Growth Regul 15:105–108
Donnelly DJ, Coleman WK, Coleman SE (2003) Potatomicrotuber production and performance: a review.Am J Potato Res 80:103–115
Estrada R, Tovar P, Dodds JH (1986) Induction of in vitrotubers in a broad range of potato genotypes. PlantCell Tiss Org Cult 7:3–10
Helder H, Miersch O, Vreugdenhil D, Sembdner G (1993)Occurrence of hydroxylated jasmonic acids in leafletsof Solanum demissum plants grown under long- andshort-day conditions. Physiol Plant 88:647–653
Hussey G, Stacey NJ (1984) Factors affecting the forma-tion of in vitro tubers of potato (Solanum tuberosumL.). Ann Bot 53:565–578
Jackson SD (1999) Multiple signalling pathways controltuber induction in potato. Plant Physiol 119:1–8
Jackson SD, Willmitzer L (1994) Jasmonic acid sprayingdoes not induce tuberization in short-day requiringpotato species kept in non-inducing conditions. Planta194:155–159
Jasik J, de Klerk G-J (2006) Effect of methyl jasmonate onmorphology and dormancy development in lily bul-
294 Plant Cell Tiss Organ Cult (2006) 87:285–295
123

bets regenerated in vitro. J Plant Growth Regul25:45–51
Koda Y (1997) Possible involvement of jasmonates invarious morphogenic events. Physiol Plant 100:639–646
Koda Y, Kikuta Y (2001) Effects of jasmonates on in vitrotuberization in several potato cultivars that differgreatly in maturity. Plant Prod Sci 4:66–70
Koda Y, Okazawa Y (1988) Detection of potato tuber-inducing activity in potato leaves and old tubers. PlantCell Physiol 29:1047–1051
Koda Y, Kikuta Y, Takazi H, Tsujino Y, Sakamura S,Yoshihara T (1991) Potato tuber-inducing activities ofjasmonic acid and related compounds. Phytochemis-try 30:1435–1438
Mithofer A, Maitrejean M, Boland W (2005) Structural andbiological diversity of cyclic octadecanoids, jasmo-nates and mimetics. J Plant Growth Regul 23:170–178
Murashige T, Skoog F (1962) A revised medium for rapidgrowth and bioassays with tobacco tissue cultures.Physiol Plant 15:473–497
Naik PS, Sarkar D, Gaur PC (1998) Yield components ofpotato microtubers: in vitro production and fieldperformance. Ann Appl Biol 133:91–99
Palmer CE, Smith OE (1969) Cytokinins and tuberinitiation in the potato Solanum tuberosum L. Nature221:279–280
Pelacho AM, Mingo-Castel AM (1991) Jasmonic acidinduces tuberization of potato stolons cultured invitro. Plant Physiol 97:1253–1255
Pruski K, Duplessis P, Lewis T, Astatkie T, Nowak J,Struik PC (2001) Jasmonate effect on in vitro tuber-ization of potato (Solanum tuberosum L.) cultivarsunder light and dark conditions. Potato Res 44:315–325
Pruski K, Astatkie T, Nowak J (2002) Jasmonate effects onin vitro tuberization and tuber bulking in two potatocultivars (Solanum tuberosum L.) under differentmedia and photoperiod conditions. In Vitro CellDevelop Biol Plant 38:203–209
Ravnikar M, Vilhar B, Gogala N (1992) Stimulatoryeffects of jasmonic acid on potato node and protoplastculture. J Plant Growth Regul 11:29–33
Sarkar D, Naik PS (1998) Effect of inorganic nitrogennutrition on cytokinin-induced potato microtuberproduction in vitro. Potato Res 41:211–217
Sarkar D, Chandra R, Naik PS (1997) Effect of inoculationdensity on potato micropropagation. Plant Cell TissOrg Cult 48:63–66
Sarkar D, Pandey SK, Sud KC, ChanemougasoundharamA (2004) In vitro characterization of manganesetoxicity in relation to phosphorous nutrition in potato(Solanum tuberosum L.). Plant Sci 167:977–986
Seabrook JEA, Douglass LK, Arnold DA (2004) Effect ofleaves on microtubers produced from potato single-node cuttings in vitro. Am J Potato Res 81:1–5
Sharma S, Chanemougasoundharam A, Sarkar D, PandeySK (2004) Carboxylic acids affect induction, develop-ment and quality of potato (Solanum tuberosum L.)microtubers grown in vitro from single-node explants.Plant Growth Regul 44:219–229
Slimmon T, Souza Machado V, Coffin R (1989) The effectof light on in vitro microtuberization of potatocultivars. Am Potato J 66:843–848
van den Berg JH, Ewing EE (1991) Jasmonates and theirrole in plant growth and development, with specialreference to the control of potato tuberization: areview. Am Potato J 68:781–794
Veramendi J, Willmitzer L, Trethewey RN (1999) In vitrogrown potato microtubers are a suitable system forthe study of primary carbohydrate metabolism. PlantPhysiol Biochem 37:693–697
Vreugdenhil D (2004) Comparing potato tuberization andsprouting: opposite phenomena. Am J Potato Res81:275–280
Vreugdenhil D, Struik PC (1989) An integrated view ofthe hormonal regulation of tuber formation in potato(Solanum tuberosum). Physiol Plant 75:525–531
Wang PJ, Hu CY (1982) In vitro mass tuberization andvirus-free seed potato production in Taiwan. AmPotato J 59:33–39
Wasternack C, Hause B (2002) Jasmonates and octadeca-noids. Progr Nucleic Acid Res 72:165–221
Wattimena GA (1983) Micropropagation as an alternativetechnology for potato production in Indonesia. PhDdissertation, University of Wisconsin, Madison
Xiong X, Tai GCC, Seabrook JEA (2002) Effectivenessof selection for quality traits during the early stagein potato breeding population. Plant Breed 121:441–444
Xu X, van Lammeren AAM, Vermeer E, Vreugdenhil D(1998) The role of gibberellin, abscisic acid, andsucrose in the regulation of potato tuber formation invitro. Plant Physiol 117:575–584
Yoshihara T, Omer E-LA, Koshino H, Sakamura S,Kikuta Y, Koda Y (1989) Structure of a tuber-inducing stimulus from potato leaves (Solanumtuberosum L.). Agr Biol Chem 53:2835–2837
Plant Cell Tiss Organ Cult (2006) 87:285–295 295
123
Copyright © 2022 FDOKUMEN




















