
Ability of a synthetic coumestan to antagonize Bothrops snake venom activities
9
Ability of a synthetic coumestan to antagonize Bothrops snake venom activities Paulo A. Melo a, * , Diogo A. Pinheiro a , Hilmar Dias Ricardo a , Fabrı ´cio F.A. Fernandes a , Marcelo A. Tomaz a , Camila Z. El-Kik a , Marcelo A. Strauch a , Tatiane F. da Fonseca a , Daniel N. Sifuentes d , Sabrina Calil-Elias c , Camilla D. Buarque b , Fla ´via V. Brito b , Paulo R.R. Costa b , Alcides J.M. Da Silva b a Departamento de Farmacologia Ba ´sica e Clı ´nica – ICB, CCS, Universidade Federal do Rio de Janeiro, Rio de Janeiro, RJ 21941-590, Brazil b Laborato ´rio de Quı ´mica Bioorga ˆnica (LQB), Nu ´cleo de Pesquisas de Produtos Naturais, CCS, Universidade Federal do Rio de Janeiro, Rio de Janeiro, RJ, Brazil c Departamento de Farma ´cia e Administraça ˜o Farmace ˆutica, Faculdade de Farma ´cia, Universidade Federal Fluminense, Nitero ´i, RJ, Brazil d Programa de Po ´s-graduaça ˜o em Biologia Animal, Universidade de Brası ´lia, Brası ´lia, DF, Brazil article info Article history: Received 22 July 2009 Received in revised form 26 August 2009 Accepted 30 September 2009 Available online 31 October 2009 Keywords: Synthetic coumestans Wedelolactone Bothrops snake venoms Myotoxicity and cardiotoxicity abstract We investigated a synthetic coumestan named LQB93 and similar compounds abilities to antagonize activities of Bothrops jararacussu and Bothrops jararaca crude venoms in different protocols. The antimyotoxic activity was evaluated in vitro by the rate of release of creatine kinase (CK) from isolated mouse extensor digitorum longus muscle (EDL) induced by B. jararacussu (25 g/ml). For in vivo studies, B. jararacussu venom (1.0 mg/kg) was preincubated with LQB93 (0.1–30 mg/kg), during 30 min, for later injection in mouse tight and evaluation of the antimyotoxic and anti-edematogenic effects. LQB93 antagonized in vitro, the increase of CK release from the EDL muscle (IC 50 ¼ 0.0291 M). It also showed in vivo, antimyotoxic and anti-edematogenic effects that were dose-dependent with ID50 of 0.17 mg/kg and 0.14 mg/kg, respectively. The hemorrhage induced by B. jararaca (1.0 mg/ kg) venom in the mouse skin, was abolished by LQB93 (10.0 mg/kg) preincubated with venom. Like wedelolactone, LQB93 protected rat isolated heart on a Langendorff prepa- ration, from the cardiotoxicity of B. jararacussu venom. LQB93 inhibit the effects of Bothrops venoms like wedelolactone, a natural compound isolated from the plant Eclipta prostrata. Ó 2009 Elsevier Ltd. All rights reserved. 1. Introduction Snake venoms contain several cytotoxic agents such as peptides, myotoxins, hemorrhagins and active enzymes with proteolytic and phospholipase activities which induce systemic dysfunctions as well as local swelling hemorrhage and myonecrosis in the different mammal tissues (Mebs et al., 1983; Watt, 1989; Sanches et al., 1992; Matsui et al., 2000; Kawano et al., 2002; Gutie ´ rrez and Ownby, 2003; Fuly et al., 2003; Harvey, 2006). Envenomation by snakes of the genus Bothrops (family Viperidae), whose venoms present many of these components, is an important public health issue in the Americas. The only specific therapy known so far is the specific antivenom (Antibothropic), which has low and limited effectiveness against local myonecrosis and cytotoxicity (Rosenfeld, 1971; Watt, 1989; Cardoso et al., 1993; Chippaux, 1998; Chippaux and Goyf- fon, 1998; Calil-Elias et al., 2002; Gutie ´rrez and Ownby, 2003; Quesada et al., 2006; Da Silva et al., 2007). A previous report of our group has shown that an herbaceous plant of pantropical and subtropical * Corresponding author. Tel.: þ55 21 2280 4694; fax: þ55 21 2610 3726. E-mail address: [email protected] (P.A. Melo). Contents lists available at ScienceDirect Toxicon journal homepage: www.elsevier.com/locate/toxicon 0041-0101/$ – see front matter Ó 2009 Elsevier Ltd. All rights reserved. doi:10.1016/j.toxicon.2009.09.021 Toxicon 55 (2010) 488–496
Transcript of Ability of a synthetic coumestan to antagonize Bothrops snake venom activities

ilable at ScienceDirect
Toxicon 55 (2010) 488–496
Contents lists ava
Toxicon
journal homepage: www.elsevier .com/locate/ toxicon
Ability of a synthetic coumestan to antagonize Bothrops snakevenom activities
Paulo A. Melo a,*, Diogo A. Pinheiro a, Hilmar Dias Ricardo a, Fabrıcio F.A. Fernandes a,Marcelo A. Tomaz a, Camila Z. El-Kik a, Marcelo A. Strauch a, Tatiane F. da Fonseca a,Daniel N. Sifuentes d, Sabrina Calil-Elias c, Camilla D. Buarque b, Flavia V. Brito b,Paulo R.R. Costa b, Alcides J.M. Da Silva b
a Departamento de Farmacologia Basica e Clınica – ICB, CCS, Universidade Federal do Rio de Janeiro, Rio de Janeiro, RJ 21941-590, Brazilb Laboratorio de Quımica Bioorganica (LQB), Nucleo de Pesquisas de Produtos Naturais, CCS, Universidade Federal do Rio de Janeiro, Rio de Janeiro, RJ, Brazilc Departamento de Farmacia e Administraçao Farmaceutica, Faculdade de Farmacia, Universidade Federal Fluminense, Niteroi, RJ, Brazild Programa de Pos-graduaçao em Biologia Animal, Universidade de Brasılia, Brasılia, DF, Brazil
a r t i c l e i n f o
Article history:Received 22 July 2009Received in revised form 26 August 2009Accepted 30 September 2009Available online 31 October 2009
Keywords:Synthetic coumestansWedelolactoneBothrops snake venomsMyotoxicity and cardiotoxicity
* Corresponding author. Tel.: þ55 21 2280 4693726.
E-mail address: [email protected] (P.A. Me
0041-0101/$ – see front matter � 2009 Elsevier Ltddoi:10.1016/j.toxicon.2009.09.021
a b s t r a c t
We investigated a synthetic coumestan named LQB93 and similar compounds abilities toantagonize activities of Bothrops jararacussu and Bothrops jararaca crude venoms indifferent protocols. The antimyotoxic activity was evaluated in vitro by the rate of release ofcreatine kinase (CK) from isolated mouse extensor digitorum longus muscle (EDL) inducedby B. jararacussu (25 g/ml). For in vivo studies, B. jararacussu venom (1.0 mg/kg) waspreincubated with LQB93 (0.1–30 mg/kg), during 30 min, for later injection in mouse tightand evaluation of the antimyotoxic and anti-edematogenic effects. LQB93 antagonized invitro, the increase of CK release from the EDL muscle (IC50¼ 0.0291 M). It also showed invivo, antimyotoxic and anti-edematogenic effects that were dose-dependent with ID50 of0.17 mg/kg and 0.14 mg/kg, respectively. The hemorrhage induced by B. jararaca (1.0 mg/kg) venom in the mouse skin, was abolished by LQB93 (10.0 mg/kg) preincubated withvenom. Like wedelolactone, LQB93 protected rat isolated heart on a Langendorff prepa-ration, from the cardiotoxicity of B. jararacussu venom. LQB93 inhibit the effects ofBothrops venoms like wedelolactone, a natural compound isolated from the plant Ecliptaprostrata.
� 2009 Elsevier Ltd. All rights reserved.
1. Introduction
Snake venoms contain several cytotoxic agents such aspeptides, myotoxins, hemorrhagins and active enzymeswith proteolytic and phospholipase activities which inducesystemic dysfunctions as well as local swelling hemorrhageand myonecrosis in the different mammal tissues (Mebset al., 1983; Watt, 1989; Sanches et al., 1992; Matsui et al.,
4; fax: þ55 21 2610
lo).
. All rights reserved.
2000; Kawano et al., 2002; Gutierrez and Ownby, 2003;Fuly et al., 2003; Harvey, 2006). Envenomation by snakes ofthe genus Bothrops (family Viperidae), whose venomspresent many of these components, is an important publichealth issue in the Americas. The only specific therapyknown so far is the specific antivenom (Antibothropic),which has low and limited effectiveness against localmyonecrosis and cytotoxicity (Rosenfeld, 1971; Watt, 1989;Cardoso et al., 1993; Chippaux, 1998; Chippaux and Goyf-fon, 1998; Calil-Elias et al., 2002; Gutierrez and Ownby,2003; Quesada et al., 2006; Da Silva et al., 2007).
A previous report of our group has shown that anherbaceous plant of pantropical and subtropical

P.A. Melo et al. / Toxicon 55 (2010) 488–496 489
distribution, named Eclipta prostrata (EP), known in Brazilas erva-botao, has been widely used in folk medicineagainst snakebite. These data have shown that the crudeextracts of EP prevented the lethality and antagonizedmyotoxic and hemorrhagic activities of some South Amer-ican crotalid venoms (Mors et al., 1989; Melo et al., 1994).Among the active compounds isolated from EP withantivenom activity, we have tested wedelolactone,a natural coumestan which is one of the most activeconstituent of EP. This coumestan not only reproduced theEP crude extract effects, but also antagonized the muscledamage, the hemorrhagic effect, and the proteolytic andphospholipase activities of different Brazilian and NorthAmerican snake venoms and their isolated toxins, in vitroand in vivo (Melo et al., 1994; Melo and Ownby, 1999). Wehave also investigated the antimyotoxic, anti-proteolyticand PLA2 inhibitory properties of the synthetic compoundEdunol, a pterocarpan isolated from Harpalyce brasiliana,a plant used against snakebites in the Northeastern Brazil.These properties could be improved through the synthesisof a bioisoster, where the prenyl group was replaced by thebenzyl group (Da Silva et al., 2004). We worked synthe-sizing new coumestans with different patterns of oxygen-ation and screened for their antimyotoxic activity. Theyreproduced the activity of wedelolactone and one of thecompounds showed antimyotoxic activity with an IC50 inthe range of wedelolactone (Da Silva et al., 2001).
In the present work, in a continuation of our program’sobjective to develop biologically active compounds againstcrotalid snake venoms, we report part of the synthesis ofnatural product named 8-methoxycoumestrol. It presentsa very low yielded product when naturally isolated fromMedicago sativa (Bickoff et al., 1966). We have also studiedthe protective effect of this coumestan in the natural andsynthetic forms against Bothrops jararaca and Bothropsjararacussu venoms using isolated mouse extensor dig-itorum longus skeletal muscle and Langendorff rat heartpreparations, as well as phospholipase and proteolyticassays. We extended our investigation to the in vivoexperiments against venom myotoxic effect, edema andhemorrhage induced by Bothrops venoms in mice.
2. Materials and methods
B. jararaca and B. jararacussu venoms were obtainedfrom Instituto Vital Brazil, Niteroi, RJ, Brazil. The anti-bothropic antivenom (PAV) was provided by the HospitalUniversitario Clementino Fraga Filho-UFRJ, Rio de Janeiro,RJ, Brazil. Each milliliter of the PAV is ascribed to be able toneutralize 2.5–5.0 mg of the Bothrops crude venoms.Triphenyl tetrazolium chloride (TTC) was purchased fromSigma Co. St Louis. Adult Wistar male rats weighing200–250 g and male Swiss mice (25.0� 4.0 g) weremaintained under a 12 h light–dark cycle, in a temperature-controlled environment (22–24 �C), with food and waterfreely available. Animal procedures were in accordancewith the guidelines for animal care prepared by theCommittee on Care and Use of Laboratory AnimalResources, National Research Council, USA, which isadopted by our University Committee (CEUA–UFRJ). The
creatine kinase (CK) activity was determined using a kit (CKNAC or CK MB).
2.1. Synthesis of 8-methoxy-3,9-dihydroxy coumestandissodium salt (LQB93)
In most of our experiment the compound 8-methox-ycoumestrol was dissodium salt derivative (Fig. 1) obtainedby reaction of the 8-methoxycoumestrol with sodiumhydride (NaH) according to previously described method-ology (Poças et al., 2006, 2007). For testing purpose, westabilished the nomenclature LQB08, LQB10, LQB12, LQB93.
2.2. Myotoxicity in vitro
In vitro CK assays were performed as previouslydescribed (Melo and Suarez-Kurtz,1988a; Melo et al.,1993).Briefly, mouse isolated extensor digitorum longus (EDL)muscle was removed, weighed and continuously super-fused with a physiologic saline solution (PSS), whosecomposition in mM was: NaCl, 135; KCl, 5; CaCl2, 2; MgCl2,1; NaHPO4, 1; NaHCO3, 15 and dextrose, 11. The pH of thissolution was equilibrated to 7.3 with 5% CO2/95% O2. Duringsuperfusion, muscles were exposed to B. jararacussu venom(25 mg/mL) or the venom preincubated with the testedcompounds at different concentrations. Perfusion batcheswere collected at 30 min intervals and replaced by freshsolution. The collected samples were stored at 4 �C andtheir CK activity determined according to procedurespreviously described (Melo and Suarez-Kurtz, 1988a,b).Muscle preparations were mounted in the sample collect-ing units for a minimum of 1 h prior to the experiment,when the basal release rate was defined as the enzyme lossfrom the muscles into the perfusion bath prior to theaddition of venom. The CK release was subsequentlydetermined prior to the addition of venom alone or thevenom plus the investigated compound. The loss of creatinekinase from the isolated muscles was expressed as enzymeunits released into the medium per gram of muscle, perhour of collection (U g�1 h�1). Pre-incubation of venomwith the coumestans (LQB93 and others) was performed atroom temperature 30 min prior to addition to the perfusionbath solution. Enzyme activities were expressed as inter-national units, where 1 U is the amount that catalyzes thetransformation of 1.0 mM of substrate at 25 �C/min.
2.3. Myotoxicity in vivo and edema
In vivo myotoxicity was evaluated by intramuscular(i.m.) injection of the venom (1 mg/kg) dissolved in phys-iological saline solution (PSS, 0.1 mL) alone or preincubatedwith 8-methoxycoumestrol (15 min, 37 �C). We performedthe i.m. injection on the middle of the back part of rightthigh with PSS, the venom alone or the venom plus8-methoxycoumestrol. The animals were lightly anes-thetized with diethyl-ether and blood were collected byorbital puncture, immediately before and 2 h after the i.m.injection. Plasma was separated by centrifugation andstored at 4 �C for subsequent determination of CK activityas described previously (Melo and Suarez-Kurtz, 1988b;Melo et al., 1993, 1994). In this same group of mice at 2 h,

O
O
OHO
R2
R3
O
O
OHO
OCH3
OH
8-metoxy-coumestrol-(LQB 93)
NaH
THF
O
O
ONaO
OCH3
ONa
dissodium salt (LQB 93)
LQB 08 = R2
= R3
= OCH2OLQB 10 = R
2= OH; R3 = OCH3
LQB 12 = R2
= R3
= OHLQB 93 = R
2= OCH3; R
3= OH
wedelolactone
Fig. 1. Wedelolactone, the synthetic coumestan derivatives compounds and the compound 8-methoxycoumestrol dissodium salt derivative (LQB93) at finalpreparation according to Poças et al. (2006, 2007).
P.A. Melo et al. / Toxicon 55 (2010) 488–496490
before the blood collect we determined with a micrometercaliper the thickness in middle of the right thigh (thelateral–lateral diameter) and evaluated the edema. Theanimals that received i.m. injection with PSS were used ascontrol.
2.4. Hemorrhagic activity
The hemorrhagic lesions were induced by intradermic(i.d.) injections of Bothrops venoms and quantified aspreviously described (Melo et al., 1994; Da Silva et al.,2007). Briefly, the experiment was performed by a premixof the venom with the compounds at different doses. Thevenom or the preincubated mixtures were kept at roomtemperature for 30 min and then injected (i.d.; 0.1 mL) inthe ventral abdominal area of mice. Two hours after venominjection, animals were euthanized by inhalation ofdiethyl-ether, and then the skin covering the abdomen wasremoved, stretched and dried at room temperature for72 h. The skin was fixed to a lucite base plate, and the wholearea in the site of injection and surrounding area weretransilluminated using an incandescent light. Light trans-mitted over a skin area of 109 mm2 was read, and lighttransmission or the absorbance was normalized by takingthe mean readings of each group, i.e. PSS (calibrated tozero), Bothrops jararacaaraca venom and treatments, withthe values being expressed as arbitrary units of absorbance.
2.5. Proteolytic and phospholipase activities
The proteolytic assay was performed with B. jararacavenom and the phospholipase assay with Bothrops jarar-acussucussu venom. The proteolytic assay was made aspreviously described (Garcia et al., 1978). B. jararaca venomat concentration of 10 mg/mL was preincubated with theinvestigated compound at room temperature during30 min. The solutions containing venom or venom plus
LQB93 were added in a preparation containing azocasein(0.2%), CaCl2 (20 mM), and Tris–HCl (0.2 M, pH 8.8). Thereaction was performed for 90 min at 37 �C and stopped bythe addiction of 0.4 mL of trichloracetic acid (15% vv) andthen centrifuged. We then took 1.0 mL of the supernatantand mixed with 0.5 mL of NaOH (2.0 M). This final solutionwas analyzed by spectrophotometry at an absorbance of420 nm. The readings for B. jararaca venom (10 mg/mL) andfor PSS were taken as 100 and 0% of proteolytic activity,respectively. Phospholipase A2 activity was carried out byadapting the turbidimetric assay previously described(Marinetti, 1965). The substrate was prepared by shakingone chicken egg yolk in a solution of NaCl (150 mM) toa final volume of 100 mL and stored at 4 �C prior to thereaction. In each assay, several tubes were prepared bytaking a final volume (0.33 mL) of a 10% dilution of the eggsuspension and adding it to a solution of NaCl (150 mM),CaCl2 (10 mM), taurocholic acid (0.01%) and Tris–HCl(5.0 mM, pH 7.4), until a spectrophotometrical absorbancebetween 0.62 and 0.65 at 925 nm was achieved. The tubeswere kept at 37 �C, under mild and constant agitationduring the procedure. The reactions begun by the additionof B. jararacussu crude venom at a concentration of or bythe addition of the mixtures of preincubated compound(concentration) plus the venom (10 mg/mL).
2.6. Langendorff isolated perfused rat heart
Langendorff isolated perfused rat heart experimentswere performed as previously described (Sifuentes et al.,2008). Briefly, the rats were anesthetized with diethyl-ether and their hearts were rapidly excised and immersedin the physiological saline solution (NaCl: 135; KCl: 5;CaCl2: 2; MgCl2: 1; NaH2PO4: 1; NaHCO3: 15 and dextrose:11, in mmol/L) at 4 �C. The aorta was then cannulated witha 20-gauge (G) needle, and connected to the Langendorffretrograde perfusion apparatus. The heart was initially

LQB93 (µM)0.01 0.1 1 10 100
LO
RTN
OC
FO
%
0
25
50
75
100B
GROUPS
g.U(
ESAELER
KC
1-h.
1-)
0
5
10
15
20
25
ACONTROLVENOM 25 µg/mLV. + LQB08 30 µMV. + LQB93 30 µMV. + LQB10 30 µMV. + LQB12 30 µM
Fig. 2. Effect of synthetic coumestans on the myotoxicity of Bothrops jararacussu crude venom on mouse EDL isolated muscle. On panel A data show the rate of CKrelease induced by B. jararacussu crude venom (25 mg/mL) on isolated EDL mouse muscles after 60 min of exposure. Effect of the different coumestans at 30 mM(LQB08; LQB10; LQB12; LQB93) on the myotoxic effect of B. jararacussu venom. On panel B data show the percent of control after inhibition by LQB93 on the rateof CK release induced by B. jararacussu venom (25 mg/mL). Data are expressed as the percent of the rate of CK released from EDL mouse muscle after 60 min ofexposure to the venom alone or associated to LQB93 at different concentrations (0.001–100 mM). The curve was determined by non-linear fitting to the Hillequation. Each point represents the mean standard error (n¼ 4–6; IC50¼ 0.03 mM).
P.A. Melo et al. / Toxicon 55 (2010) 488–496 491
perfused at a pressure of 40–70 mmHg (2–5 mL/min), withthe PSS maintained at 37 �C and constantly aerated (95% O2
and 5% CO2). The electrocardiogram (EKG) as well as theheart tension were continuously recorded by a transducer(A strain gauge Grass, FT 03) connected to a 7D GrassPolygraph. In some experiments with wedelolactone, theeffluent from bath solution was collected each 15 min andcreatine kinase (CK) activity was analyzed, and expressedas U g�1 h�1. The heart activities were assessed in differentgroups: B. jararacussu crude venom (2.5–10.0 mg/mL) alone;the venom (2.5–10.0 mg/mL) preincubated (for 15 min)with the compounds at different concentrations(1–100 mM). Before addition of the treatments, a PSSperfused the heart for approximately 15 min to stabilize thebaseline, which was used as control. Every alteration inwave amplitude, frequency and length was evaluated asa percentage of control. The measurements were takeneach 5 min before and after exposure to the venom, untilthe complete heart stoppage or 90 min. Values werestatistically analyzed using analysis of variance (ANOVA) ofrepeated measures, considering the size of the waves asparameters. For the assessment of the damaged myocardialarea, by the end of perfusion the heart ventricles weresliced into uniform sections 2–3 mm thick to investigatethe presence of lactate dehydrogenase (LDH). The slices
were incubated in 1% triphenyl tetrazolium chloride (TTC)at 37 �C (pH 7.4) for 4 min, and then the slices were placedin formaldehyde solution, which not only fixes the tissuebut also enhances the colour contrast developed. Thenormal myocardium gets brick red stained, while theinfarcted part remains unstained. Infarct size wasmeasured by WCIF Image J program and is expressed aspercentage of total area.
2.7. Statistical analysis
The statistical analysis of the data was performed usingthe Student’s t-test; for the repeated measures we usedANOVA and the Tukey test for comparisons among groups.Data are expressed as the mean � standard errors. Differ-ences were considered statistically significant for p< 0.05.
3. Results
3.1. Effect of synthetic coumestans on the in vitro myotoxicityof B. jararacussu crude venom on mouse isolated EDL muscle
Mouse EDL muscle exposed to the venom alone(25 mg/mL) showed an increase in the rate of CK releasefrom the basal level, 0.36� 0.03 (n¼ 75), to

LQB93 (mg/kg)0,1 1 10 100
PLAS
MA
CK
(U/L
)
0
200
400
600
800
1000
1200
1400
PSS
LQB93 (mg/kg)0,1 1 10 100
EDEM
A (m
m2 )
0
1
2
3
4
PSS
A B
Fig. 3. Inhibition of the myotoxicity and edema induced by B. jararacussu venom in mice by LQB93. Data show experiments performed on mice which receivedi.m. injection of Bothrops jararacussu venom alone (1.0 mg/kg) or preincubated with LQB93 (0.1–30.0 mg/kg). On panel A and B the white columns represent thei.m. injection of PSS, while the black columns represent the i.m. injection of the crude venom alone. The dotted circles represent the myotoxic effect, panel A, ofthe edema, panel B, of B. jararacussu venom plus the LQB93 at different doses. Each value represents the mean � standard error (n¼ 6). The curve was determinedby non-linear fitting to the Hill equation. The LQB93 ID50 for the antimyotoxic effect and edema inhibition was 0.17 mg/kg and 0.14 mg/kg, respectively.
P.A. Melo et al. / Toxicon 55 (2010) 488–496492
18.01�2.11 U g�1 h�1 (n¼ 28). When the EDL muscleswere exposed to the venom (25 mg/mL), a myotoxic effectwas produced. When each compound named LQB08,LQB10, LQB12 or LQB93 (0–100 mM), where addedtogether with the venom, at 30 mM they decreased themyotoxic effect in different intensities (Fig. 2A). Particu-larly, the compound LQB93 showed a complete inhibitionof the venom effect at the concentration range of10–100 mM. The IC50 determined by non-linear fitting tothe Hill equation for the compound LQB93 was 0.03 mM(Fig. 2B).
LQB93 (µM)0.1 1 10 100
PHO
SPH
OLI
PASE
AC
TIVI
TY(%
OF
CO
NTR
OL)
0
25
50
75
100
Fig. 4. Effect of LQB93 on the phospholipase activity of Bothrops jararacussu crude vshow the inhibition of phospholipase A2 activity of B. jararacussu crude venom (10between the control, venom alone, and the values for the compound LQB93 plus vencurve was determined by non-linear fitting to the Hill equation. The LQB93 IC50 for tof compound LQB93 on proteolytic activity of B. jararaca venom. The data were exprcrude venom effect alone, as a control, while the white columns represent the effectEach value represents the mean � standard error (n¼ 6). The curve was determiproteolytic activity was 09.51 mM.
3.2. Effect of 8-methoxycumestrol (LQB93) on the in vivomyotoxicity and edema induced by B. jararacussu venom inmice
Intramuscular injections of B. jararacussu crude venom(1 mg/kg of body weight) induced a significant increase onplasma CK activity when compared to control level (PSSinjection), from 52.68� 9.20 (n¼ 5) to 1196.37� 99.76 U/L(n¼ 5), 120 min after injection. The pre-incubation withLQB93 (0.1–30 mg/kg) antagonized the increase of plasmaCK activity in a dose-dependent way. The concentration of
PAV + LQB 1 µM
VENOM
PAV 2,0µL
LQB93 (µM)1 10 100
PRO
TEO
LYTI
C A
CTI
VITY
(% O
F C
ON
TRO
L)
0
25
50
75
100
enom and on proteolytic activity of Bothrops jararaca venom. On panel A data.0 mg/mL) by the compound LQB93 (0.1–300 mM). p< 0.05 for the differenceom (ANOVA). Each value represents the mean � standard error (n¼ 4–6). Thehe anti-phospholipase activity was 50.04 mM. On panel B data show the effectessed in percent of control enzyme activity. The black column represents theof the venom plus the PAV (2.0 mL) or the venom plus PAVþ 1.0 mM of LQB93.ned by non-linear fitting to the Hill equation. The LQB93 IC50 for the anti-

Time (min)0 10 20 30 40 50 60
noisneTcaidra
C)lortno
Cfo%(
0
25
50
75
100
125VENOM + LQB93
VENOM
Fig. 5. Effect of synthetic coumestans on the cardiotoxicity of Bothropsjararacussu crude venom on perfused isolated heart in a Langendorff prep-aration. The data express the percent of control tension on isolated rat heartsunder perfusion with PSS plus B. jararacussu venom 10.0 mg/mL alone or inthe presence of LQB93 (10.0 mM). Data are expressed as percentage of thetension recorded at the time zero when the heart was in control conditions,under a perfusion only with PSS. The values express the mean � standarderror (n¼ 6–10) and they are significant at p< 0.05 (ANOVA).
Fig. 6. Records of the cardiac response to Bothrops jararacussu crude venomalone or associated with 10.0 mM of LQB93 in the Langendorff preparation.Tension and EKG records of the isolated rat heart under perfusion of PSS,with B. jararacussu venom (10.0 mg/mL) alone in A, or in the presence of thevenom plus LQB93 (10.0 mM) in B and C. The heart tension values wererecorded continuously until heart stoppage in the presence of the venomalone or more than 90 min in the presence of the coumestan.
P.A. Melo et al. / Toxicon 55 (2010) 488–496 493
3 mg/kg showed more than 90% of inhibition in the venommyotoxic effect and edema. The LQB93 ID50 for the anti-myotoxic effect and edema inhibition was 0.17 mg/kg and0.14 mg/kg, respectively (Fig. 3, panel A and B).
3.3. Effect of 8-methoxycumestrol (LQB93) on B. jararacussuvenom phospholipase and B. jararaca venom proteolyticactivities
The phospholipase activity of B. jararacussu crude venom(10 mg/mL) was inhibited by the compound LQB93 ina concentration-dependent way (0.1–300 mM). Concentra-tions over 10 mM of LQB93 inhibited significantly the phos-pholipase activity of B. jararacussu crude venom (p< 0.05 forthe difference between the control and the venom alone),and LQB93 IC50 for the anti-phospholipase activity was50.04 mM. The ability of B. jararaca venom to hydrolyzeazocasein was also antagonized by LQB93 (1–100 mM) ina concentration-dependent way reaching a maximum inhi-bition of 70% at the concentration of 30 mg/mL, and LQB93IC50 for the anti-proteolytic activity was 09.51 mM. Thepolyvalent antivenom (PAV 2.0 mL) inhibited circa of 50% ofthe 10 mg/mL of B. jararaca venom proteolytic activity andthe PAVþ 1.0 mM plus LQB93 1 mM showed an additive effect(Fig. 4).
3.4. Effect of 8-methoxycumestrol (LQB93) on Hemorrhageinduced by B. jararaca venom in mice
The intradermic injection of B. jararaca crude venom(1.0 mg/kg) induced a large vascular damage in the skinwith local hemorrhage which can be quantified by changesin the light absorbance by skin (see Methods). Incubation ofLQB93 (10 mg/g) inhibited completely the hemorrhagiceffects of the venom (not shown).
3.5. Effect of coumestans on the cardiotoxic effect ofB. jararacussu venom on the Langendorff isolated perfusedrat heart
Rat isolated heart exposed to the B. jararacussu crudevenom showed a decrease on the tension amplitude,besides a time-related progressive ventricular contracture.These results are in agreement with a previous observation(Sifuentes et al., 2008). In addition to the cardiac depres-sion, on the EKG many extra systoles were observed, supra-and ventricular fibrillation and heart stoppage afterprolonged (60 min) exposure to the venom. These effectswere time and concentration-dependent. The pre-incuba-tion with LQB93 in all concentrations decreased the venomcardiotoxic effect and no stoppage occurred when LQB93concentrations were higher than 3 mM (not shown). Figs. 5and 6 show the inhibition of the venom cardiotoxic effectobserved with 10 mM of LQB93, and Fig. 6 shows records ofthe rat isolated heart tension and the EKG. The venomeffect was almost completely inhibited by LQB93 even after80 min of continuous perfusion. Fig. 7 shows the damagedmyocardial area by the end of perfusion. While theinfarcted are as of the myocardium remained unstained(clear) after incubation with TTC (see Methods), normalportions stained red. The venom decreased about of 50% of
the stained area, showing the damaging effect. WhenLQB93 is present in the perfusion solution, the damagedarea percentage reduces to 10%. This is another way ofseeing the protective effect of LQB93 on rat hearts. Anotherexperiment was performed with 2.5 mg/mL B. jararacussucrude venom plus wedelolactone (1 mM), which antago-nized the cardiotoxic effect of the venom protecting the rat

Fig. 7. Area of tissue damaged produced by Bothrops jararacussu crude venom in the rat isolated heart on the Langendorff preparation and the effect of LQB93,after stained by triphenyl tetrazolium chloride (TTC). The panel on left expresses the damaged area after exposure to B. jararacussu crude venom (10 mg/mL) aloneor in presence of LQB93 (10.0 mM). On the right the images from rat isolated heart slices in control conditions, PSS (A) or in the presence of the B. jararacussuvenom alone in B, in presence of the venom plus LQB93, in C. The normal tissue areas were stained by TTC (brick red), and the damaged areas were not stainedremaining clear. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)
P.A. Melo et al. / Toxicon 55 (2010) 488–496494
hearts from the effect on the tension and the EKG (Fig. 8, Aand B), as well as decreasing the release of CK in theperfusionate induced by the venom (Fig. 8C).
4. Discussion
The present results show that synthetic and naturalcoumestans are able to inhibit the tested Bothrops snakevenoms activities. It confirms previous studies that thesesubstances are antivenom compounds and that B. jarar-acussu venom is a powerful myotoxic and cardiotoxic agentwith many well characterized components (Melo et al.,1994; Arni et al., 1995; Bortoleto et al., 1996; Melo andOwnby, 1999; Da Silva et al., 2001; Sifuentes et al., 2008).All the synthetic coumestans tested in the isolated mousemuscle preparation showed some antimyotoxic effect indifferent grades and the compound LQB93 was the mosteffective even at low concentrations. This coumestanshowed a dose-dependent ability in vivo neutralizing themyotoxic effect of B. jararacussu crude venom as well as thethigh edema. Its ability reproduced the wedelolactoneantimyotoxic effect against many different crotalid venoms(Melo et al., 1994; Melo and Ownby, 1999). Myonecrosisand edema are widely described by many previous obser-vations as central effects induced by crotalid snake venoms(Queiroz et al., 1984; Homsi-Brandeburgo et al., 1988; Mebsand Ownby, 1990; Andriao-Escarso et al., 2000; Gutierrezand Ownby, 2003; Panunto et al., 2006). The compoundLQB93 inhibited these main activities as well as thephospholipase activity of B. jararacussu crude venom andthe B. jararaca venom proteolytic activity. There are manyprevious observations describing that B. jararacussu andother Bothrops venoms present phospholipases and theycould be involved in the thigh edema response (Homsi-Brandeburgo et al., 1988; Gutierrez et al., 1991; Cintra et al.,
1993; Gutierrez and Lomonte, 1995; Montecucco et al.,2008). These enzymes catalyze the hydrolysis of cellularphospholipids at the sn-2 position to liberate arachidonicacid and lysophospholipid to generate a family of pro-inflammatory eicosanoids (Kini and Evans, 1989; NarendraSharath Chandra et al., 2007). Panunto et al. (2006)described a lectin from B. jararacussu venom whichincreased local capillary permeability and this local effectcould be associated with proteolytic or phospholipaseactivity. One way to explain the inhibition of edema couldbe by the association of anti-phospholipase and anti-proteolytic effects of the compound. Although there aremany steps on Bothrops venom local effects that are notwell understood (Gutierrez and Lomonte, 1995; Gutierrezand Rucavado, 2000), we showed that the compoundLQB93 was able to inhibit many tested effects. In addition,LQB93 enhanced the inhibition of the PAV against B. jar-araca venom proteolytic activity. Among these antivenomeffects we can add the data that pre-incubation of B. jar-araca venom with LQB93 prevented completely thehemorrhagic effect observed by i.d. injection as well as theprocoagulant effect of this venom. The hemorrhage isa very important issue in the snakebite accident withB. jararaca and it involves the venom local tissue damageand the systemic changes in the blood coagulationcomponents (Kamiguti et al., 1986, 1991). These resultssupport our previous observations that the coumestan8-metoxycoumestrol, like wedelolactone, can inhibitBothrops snake venoms activities. We believe that theprotective effect of this compound is due to its abilities toinhibit the enzymes activities (phospholipase and proteo-lytic) of these Bothrops venoms. We suggest new studies onwhich of these enzymes are responsible for the activities,and how the interaction with the coumestans occur. Withthese new data and these pre-clinical tests we are moving
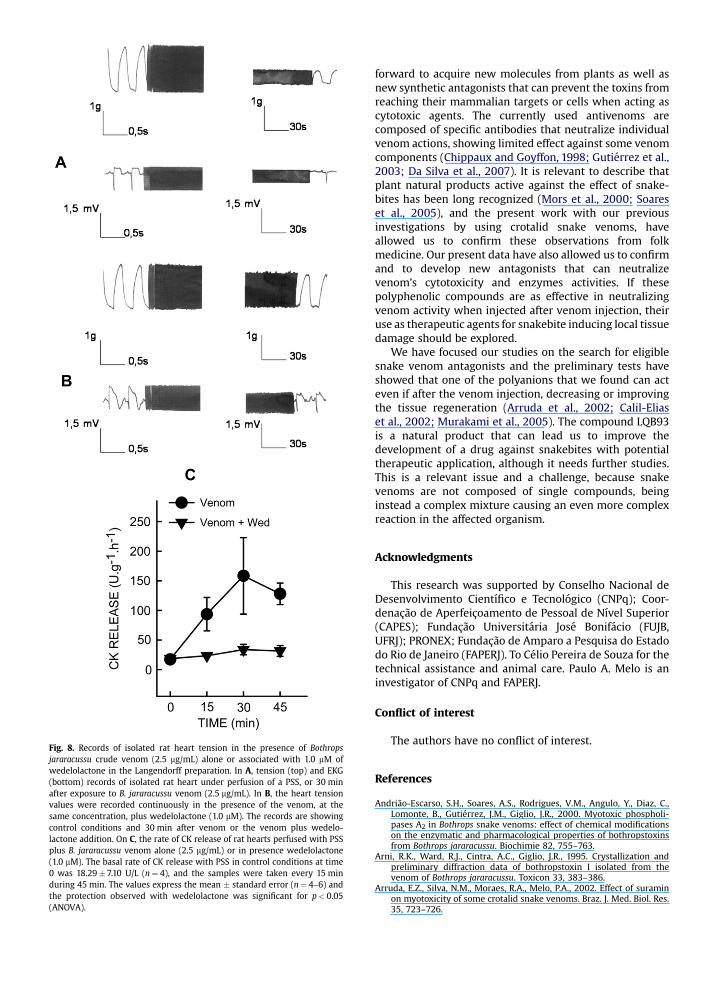
Fig. 8. Records of isolated rat heart tension in the presence of Bothropsjararacussu crude venom (2.5 mg/mL) alone or associated with 1.0 mM ofwedelolactone in the Langendorff preparation. In A, tension (top) and EKG(bottom) records of isolated rat heart under perfusion of a PSS, or 30 minafter exposure to B. jararacussu venom (2.5 mg/mL). In B, the heart tensionvalues were recorded continuously in the presence of the venom, at thesame concentration, plus wedelolactone (1.0 mM). The records are showingcontrol conditions and 30 min after venom or the venom plus wedelo-lactone addition. On C, the rate of CK release of rat hearts perfused with PSSplus B. jararacussu venom alone (2.5 mg/mL) or in presence wedelolactone(1.0 mM). The basal rate of CK release with PSS in control conditions at time0 was 18.29� 7.10 U/L (n¼ 4), and the samples were taken every 15 minduring 45 min. The values express the mean � standard error (n¼ 4–6) andthe protection observed with wedelolactone was significant for p< 0.05(ANOVA).
forward to acquire new molecules from plants as well asnew synthetic antagonists that can prevent the toxins fromreaching their mammalian targets or cells when acting ascytotoxic agents. The currently used antivenoms arecomposed of specific antibodies that neutralize individualvenom actions, showing limited effect against some venomcomponents (Chippaux and Goyffon, 1998; Gutierrez et al.,2003; Da Silva et al., 2007). It is relevant to describe thatplant natural products active against the effect of snake-bites has been long recognized (Mors et al., 2000; Soareset al., 2005), and the present work with our previousinvestigations by using crotalid snake venoms, haveallowed us to confirm these observations from folkmedicine. Our present data have also allowed us to confirmand to develop new antagonists that can neutralizevenom’s cytotoxicity and enzymes activities. If thesepolyphenolic compounds are as effective in neutralizingvenom activity when injected after venom injection, theiruse as therapeutic agents for snakebite inducing local tissuedamage should be explored.
We have focused our studies on the search for eligiblesnake venom antagonists and the preliminary tests haveshowed that one of the polyanions that we found can acteven if after the venom injection, decreasing or improvingthe tissue regeneration (Arruda et al., 2002; Calil-Eliaset al., 2002; Murakami et al., 2005). The compound LQB93is a natural product that can lead us to improve thedevelopment of a drug against snakebites with potentialtherapeutic application, although it needs further studies.This is a relevant issue and a challenge, because snakevenoms are not composed of single compounds, beinginstead a complex mixture causing an even more complexreaction in the affected organism.
Acknowledgments
This research was supported by Conselho Nacional deDesenvolvimento Cientıfico e Tecnologico (CNPq); Coor-denaçao de Aperfeiçoamento de Pessoal de Nıvel Superior(CAPES); Fundaçao Universitaria Jose Bonifacio (FUJB,UFRJ); PRONEX; Fundaçao de Amparo a Pesquisa do Estadodo Rio de Janeiro (FAPERJ). To Celio Pereira de Souza for thetechnical assistance and animal care. Paulo A. Melo is aninvestigator of CNPq and FAPERJ.
Conflict of interest
The authors have no conflict of interest.
References
Andriao-Escarso, S.H., Soares, A.S., Rodrigues, V.M., Angulo, Y., Diaz, C.,Lomonte, B., Gutierrez, J.M., Giglio, J.R., 2000. Myotoxic phospholi-pases A2 in Bothrops snake venoms: effect of chemical modificationson the enzymatic and pharmacological properties of bothropstoxinsfrom Bothrops jararacussu. Biochimie 82, 755–763.
Arni, R.K., Ward, R.J., Cintra, A.C., Giglio, J.R., 1995. Crystallization andpreliminary diffraction data of bothropstoxin I isolated from thevenom of Bothrops jararacussu. Toxicon 33, 383–386.
Arruda, E.Z., Silva, N.M., Moraes, R.A., Melo, P.A., 2002. Effect of suraminon myotoxicity of some crotalid snake venoms. Braz. J. Med. Biol. Res.35, 723–726.

P.A. Melo et al. / Toxicon 55 (2010) 488–496496
Bickoff, E.M., Spencer, R.R., Knuckles, B.E., Lundin, R.E., 1966. 3-Methox-ycoumestrol from alfalfa – isolation and characterization. J. Agric.Food Chem. 14, 444–446.
Bortoleto, R.K., Ward, R.J., Giglio, J.R., Cintra, A.C., Arni, R.K., 1996. Crys-tallization of bothropstoxin II isolated from the venom of Bothropsjararacussu. Toxicon 34, 614–617.
Calil-Elias, S., Martinez, A.M., Melo, P.A., 2002. Effect of heparin andantivenom on skeletal muscle damage produced by Bothrops jarar-acussu venom. Histol. Histopathol. 17, 463–470.
Cardoso, J.L., Fan, H.W., Franca, F.O., Jorge, M.T., Leite, R.P., Nishioka, S.A.,Avila, A., Sano-Martins, I.S., Tomy, S.C., Santoro, M.L., Chudizinski, A.M.,Castro, S.C.B., Kamiguti, A.S., Kelen, E.M.A., Hirata, M.H., Mirandola, R.M.S., Theakston, R.D.A., Warrel, D.A., 1993. Randomized comparative trialof three antivenoms in the treatment of envenoming by lance-headedvipers (Bothrops jararaca) in Sao Paulo, Brazil. Q. J. Med. 86, 315–325.
Chippaux, J.P., 1998. Snake-bites: appraisal of the global situation. Bull.WHO 76, 515–524.
Chippaux, J.P., Goyffon, M., 1998. Venoms, antivenoms and immuno-therapy. Toxicon 36, 823–846.
Cintra, A.C., Marangoni, S., Oliveira, B., Giglio, J.R., 1993. Bothropstoxin-I:amino acid sequence and function. J. Protein Chem. 12, 57–64.
Da Silva, A.J.M., Melo, P.A., Silva, N.M., Brito, F.V., Buarque, C.D., deSouza, D.V., Rodrigues, V.P., Poças, E.S., Noel, F., Albuquerque, E.X.,Costa, P.R., 2001. Synthesis and preliminary pharmacological evalua-tion of coumestans with different patterns of oxygenation. Bioorg.Med. Chem. Lett. 12, 283–286.
Da Silva, A.J., Coelho, A.L., Simas, A.B., Moraes, R.A., Pinheiro, D.A.,Fernandes, F.F., Arruda, E.Z., Costa, P.R., Melo, P.A., 2004. Synthesis andpharmacological evaluation of prenylated and benzylated pter-ocarpans against snake venom. Bioorg. Med. Chem. Lett. 14, 431–435.
Da Silva, N.M., Arruda, E.Z., Murakami, Y.L., Moraes, R.A., El-Kik, C.Z.,Tomaz, M.A., Fernandes, F.F., Oliveira, C.Z., Soares, A.M., Giglio, J.R.,Melo, P.A., 2007. Evaluation of three Brazilian antivenom ability toantagonize myonecrosis and hemorrhage induced by Bothrops snakevenoms in a mouse model. Toxicon 50, 196–205.
Fuly, A.L., Calil-Elias, S., Martinez, A.M., Melo, P.A., Guimaraes, J.A., 2003.Myotoxicity induced by an acidic Asp-49 phospholipase A2 isolatedfrom Lachesis muta snake venom. Comparison with lysophosphati-dylcholine. Int. J. Biochem. Cell Biol. 35, 1470–1481.
Garcia, E.S., Guimaraes, J.A., Prado, J.L., 1978. Purification and character-ization of sulfhydryl-dependent protease from Rhodnius prolixusmidgut. Arch. Biochem. Biophys. 188, 315–322.
Gutierrez, J.M., Nunez, J., Diaz, C., Cintra, A.C., Homsi-Brandeburgo, M.I.,Giglio, J.R., 1991. Skeletal muscle degeneration and regeneration afterinjection of bothropstoxin-II, a phospholipase A2 isolated from thevenom of the snake Bothrops jararacussu. Exp. Mol. Pathol. 55, 217–229.
Gutierrez, J.M., Leon, G., Lomonte, B., 2003. Pharmacokinetic–pharmaco-dynamic relationships of immunoglobulin therapy for envenomation.Clin. Pharmacokinet. 42, 721–741.
Gutierrez, J.M., Lomonte, B., 1995. Phospholipase A2 myotoxins fromBothrops snake venoms. Toxicon 33, 1405–1424.
Gutierrez, J.M., Ownby, C.L., 2003. Skeletal muscle degeneration inducedby venom phospholipases A2: insights into the mechanisms of localand systemic myotoxicity. Toxicon 42, 915–931.
Gutierrez, J.M., Rucavado, A., 2000. Snake venom metalloproteinases: theirrole in the pathogenesis of local tissue damage. Biochimie 82, 841–850.
Harvey, A.L., 2006. Snake venom peptides. In: Kastin, A.J. (Ed.), Handbook ofBiologically Active Peptides. Academic Press-Elsevier, USA, pp. 355–362.
Homsi-Brandeburgo, M.I., Queiroz, L.S., Santo-Neto, H., Rodrigues-Simioni, L., Giglio, J.R., 1988. Fractionation of Bothrops jararacussusnake venom: partial chemical characterization and biologicalactivity of bothropstoxin. Toxicon 26, 615–627.
Kamiguti, A.S., Matsunaga, S., Spir, M., Sano-Martins, I.S., Nahas, L., 1986.Alterations of the blood coagulation system after accidental humaninoculationby Bothrops jararacavenom. Braz. J. Med. Biol. Res.19,199–204.
Kamiguti, A.S., Cardoso, J.L.C., Theakston, R.D., Sano-Martins, I.S.,Hutton, R.A., Rugman, F.P., Warrel, D.A., Hay, C.R.M., 1991. Coagulop-athy and haemorrhage victims of Bothrops jararaca envenoming inBrazil. Toxicon 29, 961–972.
Kawano, J., Anai, K., Sugiki, M., Yoshida, E., Maruyama, M., 2002. Vascularendothelial cell injury induced by Bothrops jararaca venom; non–significance of hemorrhagic metalloproteinase. Toxicon 40, 1553–1562.
Kini, R.M., Evans, H.J., 1989. A model to explain the pharmacologicaleffects of snake venom phospholipases A2. Toxicon 27, 613–635.
Marinetti, G.V., 1965. The action of phospholipase A on lipoproteins.Biochem. Biophys. Acta 98, 554–565.
Matsui, T., Fujimura, Y., Titani, K., 2000. Snake venom proteasesaffecting hemostasis and thrombosis. Biochim. Biophys. Acta1477, 146–156.
Mebs, D., Ehrenfeld, M., Samejima, Y., 1983. Local necrotizing effect ofsnake venoms on skin and muscle: relationship to serum creatinekinase. Toxicon 21, 393–404.
Mebs, D., Ownby, C.L., 1990. Myotoxic components of snake venoms:their biochemical and biological activities. Pharmacol. Ther. 48,223–236.
Melo, P.A., Homsi-Brandeburgo, M.I., Giglio, J.R., Suarez-Kurtz, G., 1993.Antagonism of the myotoxic effects of Bothrops jararacussu venomand bothropstoxin by polyanions. Toxicon 31, 285–291.
Melo, P.A., Nascimento, M.C., Mors, W.B., Suarez-Kurtz, G., 1994. Inhibition ofthe myotoxic and hemorrhagic activities of crotalid venoms by Ecliptaprostrata (Asteraceae) extracts and constituents. Toxicon 32, 595–603.
Melo, P.A., Ownby, C.L., 1999. Ability of wedelolactone, heparin, and para-bromophenacyl bromide to antagonize the myotoxic effects of twocrotaline venoms and their PLA2 myotoxins. Toxicon 37, 199–215.
Melo, P.A., Suarez-Kurtz, G., 1988a. Release of creatine kinase from skel-etal muscle by Bothrops venoms: heparin potentiation of inhibition byantivenin. Braz. J. Med. Biol. Res. 21, 548–558.
Melo, P.A., Suarez-Kurtz, G., 1988b. Release of sarcoplasmic enzymes fromskeletal muscle by Bothrops jararacussu venom: antagonism byheparin and by the serum of South American marsupials. Toxicon 26,87–95.
Montecucco, C., Gutierrez, J.M., Lomonte, B., 2008. Cellular pathologyinduced by snake venom phospholipase A2 myotoxins and neuro-toxins: common aspects of their mechanisms of action. Cell Mol. LifeSci. 65, 2897–2912.
Mors, W.B., do Nascimento, M.C., Parente, J.P., da Silva, M.H., Melo, P.A.,Suarez-Kurtz, G., 1989. Neutralization of lethal and myotoxicactivities of South American rattlesnake venom by extracts andconstituents of the plant Eclipta prostrata (Asteraceae). Toxicon 27,1003–1009.
Mors, W.B., Nascimento, M.C., Pereira, B.M., Pereira, N.A., 2000. Plantnatural products active against snake bite – the molecular approach.Phytochemistry 55, 627–642.
Murakami, M.T., Arruda, E.Z., Melo, P.A., Martinez, A.B., Calil-Elias, S.,Tomaz, M.A., Lomonte, B., Gutierrez, J.M., Arni, R.K., 2005. Inhibitionof myotoxic activity of Bothrops asper myotoxin II by the anti-trypa-nosomal drug suramin. J. Mol. Biol. 350, 416–426.
Narendra Sharath Chandra, J.N., Ponnappa, K.C., Sadashiva, C.T.,Priya, B.S., Nanda, B.L., Veerabasappa Gowda, T., Vishwanath, B.S.,Rangappa, K.S., 2007. Chemistry and structural evaluation ofdifferent phospholipase A2 inhibitors in arachidonic acid pathwaymediated inflammation and snake venom toxicity. Curr. Top. Med.Chem. 7, 787–800.
Panunto, P.C., Da Silva, M.A., Linardi, A., Buzin, M.P., Melo, S.E., Mello, S.M.,Prado-Franceschi, J., Hyslop, S., 2006. Biological activities of a lectinfrom Bothrops jararacussu snake venom. Toxicon 47, 21–31.
Poças, E.S., Lopes, D.V., da Silva, A.J., Pimenta, P.H., Leitao, F.B., Netto, C.D.,Buarque, C.D., Brito, F.V., Costa, P.R., Noel, F., 2006. Structure–activityrelationship of wedelolactone analogues: structural requirements forinhibition of Naþ, Kþ-ATPase and binding to the central benzodiaze-pine receptor. Bioorg. Med. Chem. 14, 7962–7966.
Poças, E.S., Touza, N.A., da Silva, A.J., Costa, P.R., Noel, F., 2007. Synergisticinteraction between ouabain and 8-methoxy-3,9-dihydroxy coume-stan, a non-steroidal synthetic inhibitor of Naþ, Kþ-ATPase. Life Sci.22, 1199–1204.
Queiroz, L.S., Neto, H.S., Rodrigues-Simioni, L., Prado-Franceschi, J., 1984.Muscle necrosis and regeneration after envenomation by Bothropsjararacussu snake venom. Toxicon 22, 339–346.
Quesada, L., Sevcik, C., Lomonte, B., Rojas, E., Gutierrez, J.M., 2006. Phar-macokinetics of whole IgG equine antivenom: comparison betweennormal and envenomed rabbits. Toxicon 48, 255–263.
Rosenfeld, G., 1971. Symptomatology, pathology and treatment of snakebites. In: South America. Venomous Animals and their Venoms.Academic Press, New York, pp. 345–384.
Sanches, E.F., Freitas, T.V., Ferreira-Alves, D.L., Velarde, D.T., Diniz, M.R.,Cordeiro, M.N., Agostini-Cotta, G., Diniz, C.R., 1992. Biological activi-ties of venoms from South American snakes. Toxicon 30, 95–103.
Sifuentes, D.N., El-Kik, C.Z., Ricardo, H.D., Tomaz, M.A., Strauch, M.A., Calil-Elias, S., Arruda, E.Z., Schwartz, E.F., Melo, P.A., 2008. Ability ofsuramin to antagonize the cardiotoxic and some enzymatic activitiesof Bothrops jararacussu venom. Toxicon 51, 28–36.
Soares, A.M., Ticli, F.K., Marcussi, S., Lourenço, M.V., Januario, A.H.,Sampaio, S.V., Giglio, J.R., Lomonte, B., Pereira, P.S., 2005. Medicinalplants with inhibitory properties against snake venoms. Curr. Med.Chem. 12, 2625–2641.
Watt, G., 1989. Snakebite treatment and first aid. In: Campbell, J.A.,Lamar, W.W. (Eds.), The Venomous Reptiles of Latin America. CornellUniversity Press, New York, pp. 6–13.




















