
Características biológicas e inmunológicas del veneno de Bothrops cotiara (Serpentes: Viperidae)
Upload
anhangueraCategory
view
0download
0
EXPERIMENTAL AND MOLECULAR PATHOLOGY 52,25-36 (I!?.@)
Changes in Myofibrillar Components after Skeletal Muscle Necrosis Induced by a Myotoxin Isolated from the Venom of
the Snake Bothrops asper
Jo& MARIA GUTIBRREZ,’ VILMA ARCE,* FERNANDO BFENES,~ AND FERNANDO CHAVES’
‘Institute Clodomiro Picado, Facultad de Microbiologia, and ‘Centro de Znvestigacibn en Biologia Celular y Molecular, Universidad de Costa Rica, San Jose, Costa Rica; and ‘Laboratorio de Patologia Experimental, Hospital Mkico, Caja Costarricense de1 Seguro Social, Costa Rica
Received May 12, 1989, and in revised form August 28, 1989
The effects of a myotoxic phospholipase A, isolated from the venom of the crotaline snake Bothrops asper on skeletal muscle myotibrils were studied by histological, ultrastructural, immunohistochemical, and biochemical parameters. Myotoxin induced a rapid and promi- nent muscle necrosis after intramuscular injection in mice. In this process, myofibrils were affected and three main changes were observed: (A) Initially, they were hypercontracted, eventually forming “clumped,” dense masses which alternated with spaces devoid of my- ofilaments in the cytoplasm. This initial stage is probably due to hypercontraction resulting from a calcium influx after toxin-induced sarcolemmal damage. (B) A second change oc- curred between 3 and 6 hr, when the clumped or hypercontracted pattern changed to a “hyaline” pattern in which myotilaments were relaxed and had a more uniform distribution in the cellular space. Although there was not a widespread degradation of myofibrillar components at this stage, desmin started to be lost in samples obtained as early as 15 min after toxin injection, and a-actinin was almost absent by 7 hr. Thus, it is proposed that this shift may be due to a selective proteolytic degradation of structurally relevant components, particularly a-a&tin. As a consequence, the mechanical integration of myofdaments is impaired, precluding hypercontraction. (C) Finally, at later time periods (24,48, and 72 hr), there was widespread degradation of myotibrillar proteins, probably caused by proteases derived from intlanunatory cells such as neutrophils and macrophages, whose numbers in necrotic muscle increased markedly at these time periods. Q EJW Academic has, hc.
INTRODUCTION
Many snake venoms induce skeletal muscle necrosis clinically as well as in experimental animals (Tu, 1977). Some of them contain myotoxins, i.e., proteins that directly damage muscle cells (Harris et al., 1980; Gutierrez et al., 1984a), whereas in other cases muscle damage is indirect, being due to the ischemia which develops as a consequence of microvessel damage (Gleason et al., 1983; Gutierrez et al., 1984b). In both cases myolibrils are drastically affected as a consequence of venom action. However, little is known about the detailed morphological and biochemical changes occurring in myolibrils after injection of snake venoms and toxins.
A basic myotoxic phospholipase A2 was isolated from the venom of the Central American pit viper Bothrops usper (Gutierrez ef al., 1984a), and has recently been named Bothrops usper myotoxin I (Lomonte and Gutierrez, 1989). This toxin induces muscle damage by affecting initially the plasma membrane (Gutierrez et al., 1984b; Brenes et al., 1987). After this initial alteration, there is a rapid se- quence of degenerative events in myotibrils, mitochondria, sarcoplasmic reticu- lum, and nuclei, a process that rapidly ends in irreversible cell damage. Other myotoxic agents such as tarantula venom (Ownby and Odell, 1983), bupivacaine
0014-4800190 $3.00 Copyright 0 1990 by Academic Press, Inc. AU rights of reproduction in any form reserved.

26 GUTIlhREZ ET AL.
(Nonaka et al., 1983), and plasmocid (Ishiura et al., 1984) also induce irreversible muscle cell damage which has morphological similarities with myonecrosis in- duced by B. asper myotoxin. Thus, the study of myofibrillar alterations that take place after injection of this myotoxin represents a potentially useful model to gain a better understanding of structural protein degradation following acute muscle damage. In this work we have followed these changes by using histological, ul- trastructural, immunohistochemical, and biochemical techniques.
MATERIALS AND METHODS
Venom and toxin. Crude B. asper venom was a pool obtained from adult spec- imens collected in the Atlantic region of Costa Rica. Once collected, venom was freeze-dried and stored at -40°C. The myotoxin was isolated according to Guti- Crrez et al. (19&c). Homogeneity was demonstrated by polyacrylamide disc gel electrophoresis (Reisfeld et al., 1962).
Biochemical changes of myofibrils in vivo. Six groups of four mice each (18 20 g body wt) were injected intramuscularly in the right gastrocnemius with 100 p,g of the toxin (dissolved in 100 p,l of phosphate-buffered saline solution, pH 7.2). Four mice were killed at each of the following time intervals: 1, 3, 6,24, 48, and 72 hr. Then, gastrocnemius muscles were obtained and homogenized in phos- phate-buffered saline, pH 7.2, containing 0.1% Triton X-100. Homogenates were centrifuged at 9OOOg and an aliquot of the pellet was analyzed by slab SDS- polyacrylamide gel electrophoresis (SDS-PAGE) using 12% acrylamide (Laem- mli, 1970). Control mice were injected with 100 ~1 of phosphate-buffered saline solution.
Structural changes in myofibrils. Six groups of four mice each (18-20 g body wt) were injected with 100 p.g of the toxin as described above. Four mice were killed at each of the following time intervals: 30 min and 1,3,6, 12, and 24 hr, and injected gastrocnemius muscles were obtained. The tissue was cut into small pieces and processed for light and electron microscopy as previously described (Arroyo and Gutierrez, 1981). Thick sections (1 p,m) were stained with methylene blue and thin sections (silver to light gold) were stained with methanolic uranyl acetate and with lead citrate, and examined in a Hitachi HU-12A electron micro- scope. Control mice in these experiments were injected with 100 l.~l of phosphate- buffered saline solution, pH 7.2.
Zmmunohistochemistry. Five groups of three mice each (18-20 g body wt) were injected as described previously. Three mice were killed at each of the following time intervals: 15 and 30 min and 1,2, and 7 hr, and tissue samples from injected gastrocnemius muscles were collected. Muscle tissue was fixed in BG-HgCl, (6% mercuric chloride in 1% sodium acetate, containing 0.1% glutaraldehyde) for 6 hr at room temperature, dehydrated in alcohols, embedded in paraffin, and cut at 5 pm. Sections were stained with rabbit anti-desmin and mouse monoclonal anti-ol-actinin using the avidin-biotin peroxidase method. Briefly, some sections were treated with trypsin for 10 min, incubated with anti-desmin 1:80 (Dakopatts, Denmark), and incubated with biotinylated goat anti-rabbit 1500 (Sigma Chemical Co., St. Louis, MO). Other sections were treated with protease XIV for 20 min, incubated with anti-ol-actinin I:200 (Sigma), and incubated with biotinylated goat anti-mouse 1:160 (Sigma). Finally, the sections were incubated with avidin- peroxidase (Sigma), developed with diaminobenzidine (Sigma), and counter- stained with Harris hematoxylin. Negative controls included omission of the pri-

EFFECT OF MYOTOXIN ON MYOFIBRILS 27
mat-y antibody and replacement with rabbit normal serum and mouse normal ascites, followed by the procedure described above.
Znflammatory infiltrate. The technique of Maskrey et al. (1977) was used with some modifications. Five groups of four mice each (N-20 g body wt) were in- jected with 100 p,g of the toxin as described above. Then, four mice were killed at each of the following time intervals: 3,6,24,48, and 72 hr. Immediately, injected gastrocnemius were removed and chopped with scissors in 2.0 ml of phosphate- buffered saline solution, pH 7.2. The suspensions were incubated at 37°C for 30 min and then filtered through bolting silk. Inflammatory cells were counted in a hemocytometer. Suspensions were then centrifuged and smears were prepared and stained in order to identify intlammatory cell types.
RESULTS
Myofibrillar changes. Control samples had the typical normal structure of skel- etal muscle. In toxin-injected samples, necrotic muscle cells were observed at all time periods, and pathological changes were similar to those described in detail in previous publications (Gutierrez et al., 1984b). Regarding myofibrillar changes, samples obtained at 30 min and 1 hr had a similar pattern of alterations, charac- terized by clumping of myofilaments with formation of dense, amorphous masses alternating with intracellular spaces devoid of myotibrils (Fig. IA). Ultrastructur- ally, in some necrotic cells, regions of hypercontracted myofilaments alternated with areas of overstretched myotibrils. In other fibers, clumping was so strong that individual myofilaments could not be detected in these masses, and there was no preservation of striated structure (Fig. 1B). After 3 hr, some myotibrillar masses were shifting from this clumped pattern to a more relaxed and hyaline appearance; this change was well advanced by 6 hr when the majority of necrotic cells had a hyaline and homogeneous myotibrillar material (Fig. 2A). Ultrastruc- turally, myofilaments in these cells had a more uniform distribution in the cellular space, and individual myofilaments could be distinguished (Fig. 2B).
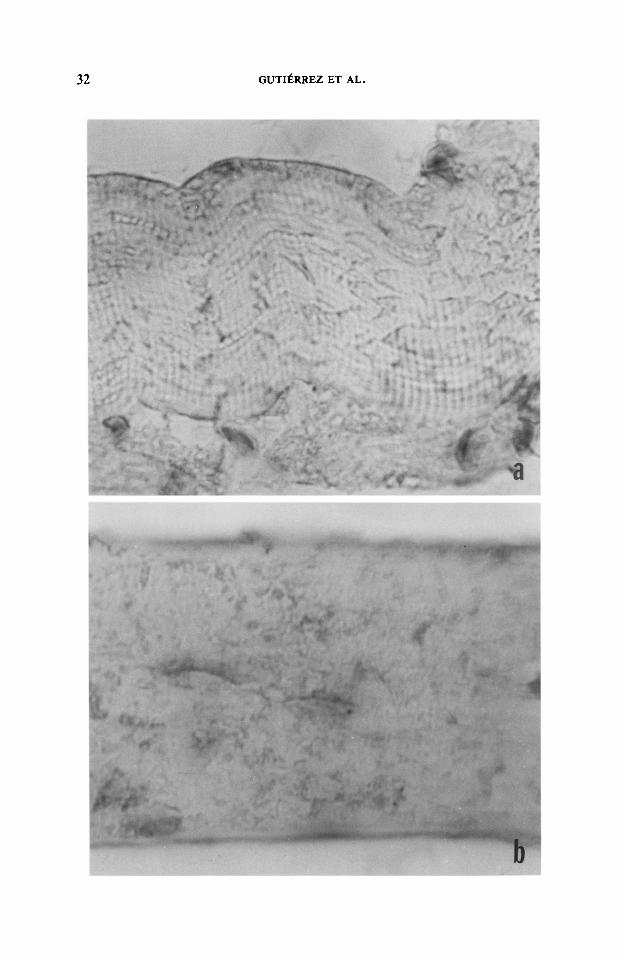
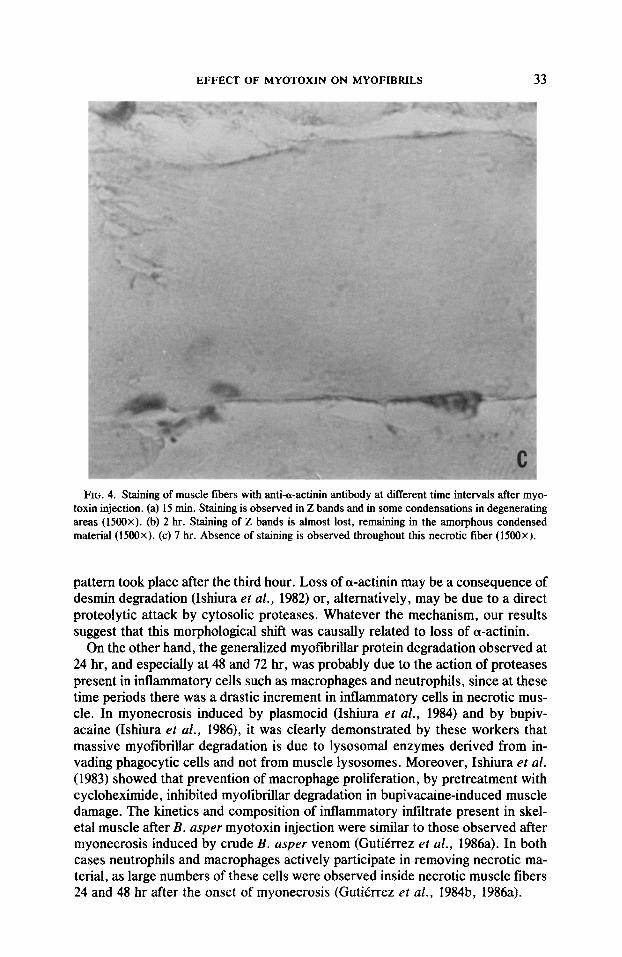
Zmmunohistochemistry. Results obtained with anti-desmin and anti-a-actimim are summarized in Table I. Most of the necrotic fibers were localized at the periphery of the muscle 15 min after myotoxin injection. Desmin staining was lost in necrotic areas as early as I5 min after myotoxin administration (Fig. 3). A similar lack of staining for desmin was observed at all time intervals studied. On the other hand, cw-actinin staining was observed in partially preserved Z bands and in condensations of positively stained material surrounding necrotic areas (Fig. 4). At 1 hr most necrotic myofibers presented this staining pattern for a-actinin, while others had lost Z band staining, but maintaining granular positive stained material. At 2 hr there were more fibers without Z bands, and the condensed material became smaller and less intensely stained. By 7 hr most necrotic fibers had lost positively stained Z bands and condensed material, while very few fibers still retained these positively stained granular deposits (Fig. 4).
Electrophoretic analysis. Electrophoresis of myotibrillar proteins demonstrated few changes in samples obtained 1, 3, and 6 hr after toxin injection. Samples collected at 24 hr showed only a very small decrease in the intensity of some protein bands, whereas samples collected at 48 and 72 hr presented a widespread decrease in myofibrillar components, since the majority of the bands were less stained than those of control muscle (Fig. 5).
Znjlammatory infiltrate. The amount of inflammatory cells present in injected

28 GUTIl%BEZ ET AL.
FIG. 1. Skeletal muscle taken 1 hr after injection of B. asper myotoxin. (A) Light micrograph showing necrotic cells with a “clumped” pattern of myotibrillar disorganization. Notice that the clumps alternate with spaces devoid of myofibrils (300X). (B) Electron micrograph of part of a necrotic muscle cell containing a dense clump of myofilaments (arrows). Individual myotilaments cannot be discerned within the clump. The basal lamina (BL) is the only structure observed at the periphery of the cell. The rest of the cytoplasm is almost totally devoid of myofilaments (7500x).

EFFECT OF MYOTOXIN ON MYOFIBRILS 29
FIG. 2. Skeletal muscle taken 6 hr after injection of B. asper myotoxin. (A) Light micrograph showing necrotic muscle cells with a “hyaline” pattern of myotibrillar disorganization. Myotibrils present a more uniform and hyaline distribution in the cellular space and there are no clumps. One muscle cell was not affected by the toxin (arrow) (350x). (B) Electron micrograph of a portion of a muscle necrotic cell having a hyaline pattern of myotibrillar disorganization. Myotilaments are not forming dense clumps, but are instead distributed more homogeneously in the cellular space, leaving no spaces devoid of myotilaments (6000~).

30 GUTIEBBEZ ET AL.
TABLE I Immunobistochemical Staining of Necrotic Muscle Fibers with Anti-Desmin and Anti-a-Actinin
Antibodies at Different Time Intervals following Myotoxin Injection”
Time
15 mill 30 min 6Omin 2hr 7hr
Desminb a-Actinin’ Percentage of necrotic
fibers
0 0 0 0 0 3 3 3 2 0
l-S% 3m 70% 80% 80-90%
0 Staining intensity is based on a semiquantitative scale ranging from 0 (unstained) to 4 (strongly stained).
b Staining for desmin was localized along the Z bands. c Staining for a-actinin was localized in Z bands and/or in granular deposits.
gastrocnemius at different time periods is given in Fig. 6. A significant increase occurred at 24 hr, but the largest increment was observed at 48 hr. When inflam- matory cells were identified by light microscopy, changes in cellular composition were studied. Neutrophil was the predominant cell type at 24 hr, whereas mac- rophages were predominant by 48 and 72 hr.
DISCUSSION
B. asper myotoxin induces muscle cell damage by first binding and affecting the integrity of skeletal muscle plasma membrane (Gutierrez et al., 1984a, b, 1986~; Brenes et al., 1987). This early membrane alteration induces an efflux of creatine kinase and creatine and a calcium intlux (Gutierrez et al., 1984a, 1986c). The net result is a complex series of degenerative events in mitochondria, sarcoplasmic reticulum, T tubules, and myofilaments that bring the cells beyond the “point of no return,” as has been described previously by Gutierrez et al. (1984b).
Myofibrillar structure changed drastically in myonecrosis induced by B. usper myotoxin. The initial hypercontraction and clumping of myofilaments was obvi- ously not related to a generalized protein degradation, since electrophoretic stud- ies detected very few changes in myofibrillar proteins during the first hour after toxin injection. Thus, the initial changes in myofibrils were structural alterations in the spatial arrangement of myofilaments and may be caused by the hypercon- traction that occurs after the elevation of cytosolic calcium levels as a conse- quence of the initial disruption of sarcolemmal integrity. A rapid increase in cytosolic calcium levels after injection of this myotoxin was demonstrated by Gutierrez ef al. (1984a). Hypercontraction may cause a mechanical disruption of myotibrillar continuity so that hypercontracted myofibrils would be clumped into separate masses, with no integration between them. The observation that, at early time periods, there were areas of overcontracted myofilaments alternating with regions of overstretched myofilaments indicates that this may represent a stage that precedes myofibrilIar clumping.
Two significant changes occurred in myotibrils at later time periods: (a) Myo- flbrillar material changed to a more hyaline and homogeneous appearance 3 to 6 hr after toxin injection, with no evidence of widespread protein degradation, and (b) there was a degradation of all myotibrillar components at 24, 48, and 72 hr, as judged by electrophoretic analysis. The first change also seemed to occur in other
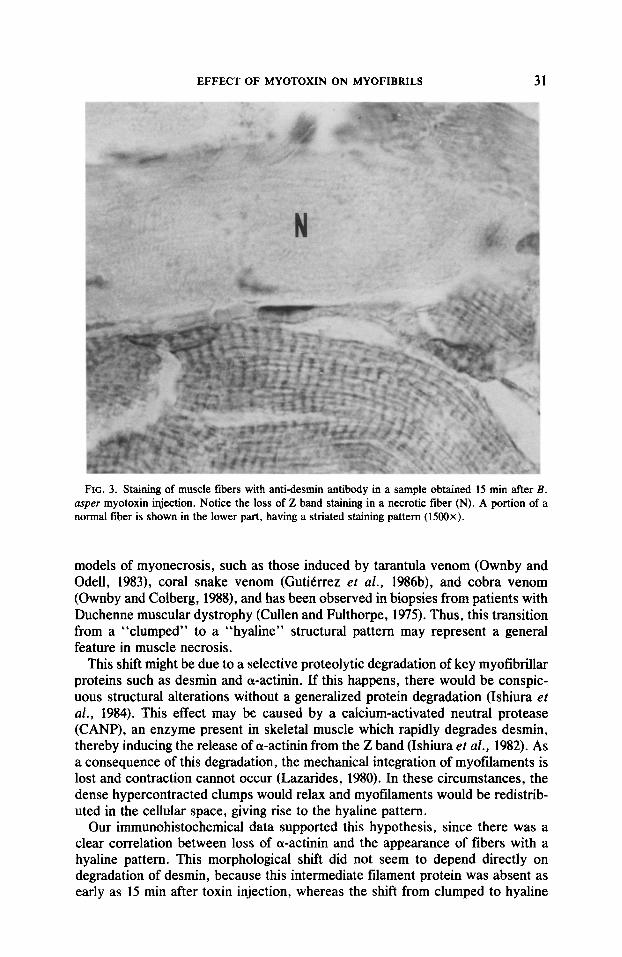
EFFECT OF MYOTOXIN ON MYOFIBIULS 31
FIG. 3. Staining of muscle fibers with anti-desmin antibody in a sample obtained 15 min after B. asper myotoxin injection. Notice the loss of Z band staining in a necrotic fiber (N). A portion of a normal fiber is shown in the lower part, having a striated staining pattern (1500x).
models of myonecrosis, such as those induced by tarantula venom (Ownby and Odell, 1983), coral snake venom (Gutierrez et al., 1986b), and cobra venom (Ownby and Coiberg, 1988), and has been observed in biopsies from patients with Duchenne muscular dystrophy (Cullen and Fulthorpe, 1975). Thus, this transition from a “clumped” to a “hyaline” structural pattern may represent a general feature in muscle necrosis.
This shift might be due to a selective proteolytic degradation of key myofibrillar proteins such as desmin and ol-actinin. If this happens, there would be conspic- uous structural alterations without a generalized protein degradation (Ishiura et al., 1984). This effect may be caused by a calcium-activated neutral protease (CANP), an enzyme present in skeletal muscle which rapidly degrades desmin, thereby inducing the release of ol-actinin from the Z band (Ishiura et al., 1982). As a consequence of this degradation, the mechanical integration of myofilaments is lost and contraction cannot occur (Lazarides, 1980). In these circumstances, the dense hypercontracted clumps would relax and myofilaments would be redistrib- uted in the cellular space, giving rise to the hyaline pattern.
Our immunohistochemical data supported this hypothesis, since there was a clear correlation between loss of a-actinin and the appearance of fibers with a hyaline pattern. This morphological shift did not seem to depend directly on degradation of desmin, because this intermediate filament protein was absent as early as 15 min after toxin injection, whereas the shift from clumped to hyaline
32 GUTIfiRREZ ET AL.
EFFECT OF MYOTOXIN ON MYOFIBRILS 33
FIG. 4. Staining of muscle fibers with anti-a-actinin antibody at different time intervals after myo- toxin injection. (a) 15 min. Staining is observed in Z bands and in some condensations in degenerating areas (1500x). (b) 2 hr. Staining of Z bands is almost lost, remaining in the amorphous condensed material (1500x). (c) 7 hr. Absence of staining is observed throughout this necrotic fiber (1500x).
pattern took place after the third hour. Loss of o-actinin may be a consequence of desmin degradation (Ishiura et al., 1982) or, alternatively, may be due to a direct proteolytic attack by cytosolic proteases. Whatever the mechanism, our results suggest that this morphological shift was causally related to loss of ol-actinin.
On the other hand, the generalized myotibrillar protein degradation observed at 24 hr, and especially at 48 and 72 hr, was probably due to the action of proteases present in inflammatory cells such as macrophages and neutrophils, since at these time periods there was a drastic increment in inllammatory cells in necrotic mus- cle. In myonecrosis induced by plasmocid (Ishiura et al., 1984) and by bupiv- acaine (Ishiura et al., 1986), it was clearly demonstrated by these workers that massive myofibrillar degradation is due to lysosomal enzymes derived from in- vading phagocytic cells and not from muscle lysosomes. Moreover, Ishiura et al. (1983) showed that prevention of macrophage proliferation, by pretreatment with cycloheximide, inhibited myotibrillar degradation in bupivacaine-induced muscle damage. The kinetics and composition of inflammatory infiltrate present in skel- etal muscle after B. asper myotoxin injection were similar to those observed after myonecrosis induced by crude B. asper venom (Gutierrez et al., 1986a). In both cases neutrophils and macrophages actively participate in removing necrotic ma- terial, as large numbers of these cells were observed inside necrotic muscle fibers 24 and 48 hr after the onset of myonecrosis (Gutierrez et at., 1984b, 1986a).

34 GUTIhRJ5Z ET AL.
l-
2- T
3-
4-
5-
a b c FIG. 5. SDS-PAGE of myofibrillar proteins obtained from mouse gastrocnemius muscle at different
time intervals after injection of B. asper myotoxin. (Lane a) Proteins of control muscle. (Lane b) Proteins of muscle obtained 6 hr after toxin injection. (Lane c) Proteins of muscle obtained 24 hr after toxin injection. (Lane d) Proteins of muscle obtained 48 hr after toxin injection. The numbers to the left represent the position of the following molecular weight standards: (1) phosphorylase b (94K); (2) albumin (67K); (3) ovalbumin (43K); (4) carbonic anhydrase (30K); and (5) trypsin inhibitor (20K).
In conclusion, our findings on myofibrillar changes after B. asper myotoxin injection agree with the “two-step” model proposed by Ishiura et al. (1984) to explain myofibrillar degradation after plasmocid-induced muscle damage. In the case of B. asper myotoxin, after the early hypercontraction of myofilaments due
400 -
d
‘0, r< 300 - v)
= 3
6 & 200 -
-is
z
100 -
Time (hr) FIG. 6. Changes in the number of inflammatory cells present in mouse gastrocnemius muscle at
different time periods after injection of E. asper myotoxin. Each value represents the mean 5 SEM (n = 4).

EFFECT OF MYOTOXIN ON MYOFIBRILS 35
to a massive calcium influx, there was a first stage of myofibrillar degradation characterized by a restricted hydrolysis of particular components such as desmin and a-actinin. The loss of o-actinin was probably involved in the morphological shift from a clumped to a hyaline pattern. The second stage was responsible for the widespread myofibrillar protein degradation and was probably caused by pro- teases present in neutrophils and macrophages.
ACKNOWLEDGMENTS
The authors thank Marcia Munoz and Albert0 Delgado for their collaboration in the laboratory work and Rocfo Monge for typing the manuscript. This work was supported by Vicerrectorfa de Investiga- cibn, Universidad de Costa Rica, Projects 741-86-080 and 801-87-108, by CENDEISSS, Costa Rica, and by the Third World Academy of Sciences (Research Grant BC 88-54). J. M. Gutitrrez and F. Chaves were recipients of a research career award from the Costa Rican National Scientific and Technological Research Council (CONICIT).
REFERENCES ARROYO, O., and GtJTn%REZ, J. M. (1981). Estudio ultraestructural de la mionecrosis inducida en
ratbn por el veneno de terciopelo (gothrops asper) de Costa Rica. Toxicon 19, 773-782. BRENES, F., GUTI~~RREZ, J. M., and LOMONTE, B. (1987). Immunohistochemical demonstration of the
binding of Bothrops asper myotoxin to skeletal muscle sarcolemma. Toxicon 25, 574-577. CULLEN, M. J., and FULTHORPE, J. J. (1975). Stages in Bbre breakdown in Duchenne muscular
dystrophy. An electron microscopic study. J. Neural. Sci. 24, 179-200. GLEASON, M. L., ODELL, G. V., and OWNBY, C. L. (1983). Isolation and biological activity of viri-
ditoxin and a viriditoxin variant from Crotalus viridis viridis venoms. J. Toxicol. Toxin Rev. 2, 235-265.
GUTIBRREZ, J. M., OWNBY, C. L., and ODELL, G. V. (1984a). Isolation of a myotoxin fromBothrops asper venom: Partial characterization and action on skeletal muscle. Toxicon 22, 115-128.
GUTI~?RREZ, J. M., OWNBY, C. L., and ODELL, 6. V. (1984b). Pathogenesis of myonecrosis induced by crude venom and a myotoxin of Bothrops asper. Exp. Mol. Parhol. 40, 367-379.
GUTI~RREZ, J. M., CHAVES, F., and CERDAS, L. (1986a). Inflammatory infiltrate in skeletal muscle injected with Bothrops asper venom. Rev. Biol. Trap. 34, 209-219.
GUTIBRREZ, J. M., ARROYO, O., CHAVES, F., LOMONTE, B., and CERDAS, L. (1986b). Pathogenesis of myonecrosis induced by coral snake (Micrurus nigrocinctus) venom in mice. Brit. J. Exp. Pathol. 67, 1-12.
GUTIBRREZ, J. M., LOMONTE, B., CHAVES, F., MORENO, E., and CERDAS, L. (1986c). Pharmacolog- ical activities of a toxic phospholipase A isolated from the venom of the snake Bothrops asper.
Camp. Biochem. Physiol. 84C, 159-164. HARRIS, J. B., JOHNSON, M. A., and MACDONELL, C. A. (1980). Muscle necrosis induced by some
presynaptically active neurotoxins. In “Natural Toxins” (D. Eaker and T. Wadstrom, Eds.), pp. 569-578. Pergamon Press, Oxford.
ISHIURA, S., NONAKA, I., and SUGITA, H. (1982). Calcium-activated neutral protease: Its degradative role in muscle cells. In “Muscular Dystrophy” (S. Ebashi, Ed.), pp. 265-282. University of Tokyo Press, Tokyo.
ISHIURA, S., NONAKA, I., FUJITA, T., and SUGITA, H. (1983). Effect of cycloheximide administration on bupivacaine-induced acute muscle degradation. J. Biochem. 94, 1631-1636.
ISHIURA, S., NONAKA, I., NAKASE, H., TADA, A., and SUGITA, H. (1984). Two-step mechanism of myofibrillar protein degradation in acute plasmocid-induced muscle necrosis. Biochim. Biophys. Acta 198, 333-342.
ISHIURA, S., NONAKA, I., and SUGITA, H. (1986). Biochemical aspects of bupivacaine-induced acute muscle degradation. J. Cell Sci. 83, 197-212.
LAEMMLI, U. K. (1970). Cleavage of structural proteins during the assembly of the head of bacterio- phage T4. Nature (London) 227, 680-685.
LAZARIDES, E. (1980). Intermediate filaments as mechanical integrators of the cellular space. Nature (London) 283, 249-253.
LOMONTE, B., and GUTIBRREZ, J. M. (1989). A new muscle damaging toxin, myotoxin II, from the venom of the snake Bothrops asper (terciopelo). Toxicon, 27, 725-733.

36 GUTIlbREZ ET AL.
MASKREY, P., PLUSKAL, M. G., HARRIS, J. B., and PENNINGTON, R. J. T. (1977). Studies on in- creased acid hydrolase activities in denervated muscle. J. Neurochem. 28, 403-409.
NONAKA, I., TAKAGI, A., ISHIURA, S., NAKASE, H., and SUGITA, H. (1983). Pathophysiology of muscle fiber necrosis induced by bupivacaine-hydrochloride. Acta Neuropathol. 60, 167-174.
OWNBY, C. L., and ODELL, G. V. (1983). Pathogenesis of skeletal muscle necrosis induced by taran- tula venom. Exp. Mol. Puthol. 38, 283-296.
OWNBY, C. L., and COLBERG, T. (1988). Classification of myonecrosis induced by snake venoms: Venoms from the prairie rattlesnake (Crotalus viridis viridis), Western diamondback rattlesnake (Crotalus atrox) and the Indian cobra (Nuja naja naja). Toxicon 26,459-474.
REISFELD, R. A., LEWIS, U. J., and WILLIAMS, D. E. (1%2). Disk electrophoresis of basic proteins and peptides on polyacrylamide gels. Nature (London) 195, 281-283.
Tu, A. T. (1977). “Venoms: Chemistry and Molecular Biology,” 1st ed. Wiley, New York.
Copyright © 2022 FDOKUMEN




















