
Comparative effects of post-mortem storage and low-calcium-requiring neutral proteinase on bovine...
11
J, Sci. Food Agric. 1983, 34,466476 Comparative Effects of Post-mortem Storage and Low-calcium-requiring Neutral Proteinase on Bovine and Rabbit Myofibrillar Proteins Ahmed Oualia, Alain Obleda, Patrick Cottinb, Nadia Merdacib, Andrk Ducastaingb and Christian Valina a Station de Recherches sur la Viande, INRA de Theix, 63110 Beaumont, France; Laboratoire de Biochimie de la Nutrition, Universite'de Bordeaux I, All.4e des Facultis, 33405 Talence, Cedex, France (Manuscript received 11 May 1982) The effects of post-mortem storage and of a low-calcium-requiring neutral proteinase on the myofibrils of beef Longissimus dorsi and Rectus abdominis muscles and rabbit Longissirnus dorsi muscle were studied by measuring the Mg-Ca-enhanced myofibrillar ATPase activity and the changes in banding patterns on electrophoresis in sodium dodecyl sulphate. During ageing, the changes in the ATPase activity and myofibrillar proteins were qualitatively different between rabbit and bovine muscles, whilst differ- ences in intensity were only observed between the two bovine muscles. The most prevalent component appearing upon conditioning had a mol. wt of 27 000 in rabbit muscle and of 30 000 in bovine muscles. In both species, the incubation of myofibrils with the low-calcium-requiring neutral proteinase mimicked the post-mortem changes. This would suggest that this enzyme has a qualitatively different effect on rabbit and bovine myofibrils. 1. Introduction Many studies have been carried out on the changes in myofibrillar proteins during post-mortem storage, their causes and on the relationship between these alterations and meat tenderi~ation.l-~ The intensity and the speed of these modifications are species-dependent4and, within a given species, muscle-dependent.3,57 Two changes have been well documented: firstly, the alteration of the actin-myosin interactions, which can be monitored by the modification of the Mg-Ca-enhanced triphosphatase (ATPase) activity of actomyosin6-10; and, secondly, the hydrolysis of myofibrillar proteins.11-13 From the in-vitro results reported, these two myofibrillar protein changes can be ascribed to the action of calcium-activated neutral proteinase (Ca-ANP) and lysosomal proteinases.1910, 14e19 CaANP, a neutral proteinase activated by millimolar (mmol) concentrations of calcium (high- calcium-requiring neutral proteinase) has been studied extensively,l6* 20-23 but one major problem associated with assessing the physiological significance of this proteinase activity and its role in the post-mortem ageing process of meat is that it requires calcium of mmol order, which is much higher than in-vivo and post-mortem concentrations of intracellular calcium. Another problem is that this activity is more or less abolished by sarcoplasmic inhibitors of both high24 and low25-27 molecular weight. Recently Mellgren2*and Kubota et aLZ9 obtained a low-calcium-requiring form of the protease from canine cardiac and chicken skeletal muscle respectively. This enzyme may be derived from the high-calcium-requiring neutral proteinase by an autolytic p r o c e ~ s , ~ ~ ~ 31 but no investigations con- cerning the identity between the low-calcium-requiring enzyme obtained from the high-calcium- requiring form by autolysis and the in-situ form have been reported. A form of this proteinase with low calcium requirements and its inhibitor were recently purified by Cottin et ~1.25.26 from rabbit skeletal muscle. 466
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Comparative effects of post-mortem storage and low-calcium-requiring neutral proteinase on bovine...

J, Sci. Food Agric. 1983, 34,466476
Comparative Effects of Post-mortem Storage and Low-calcium-requiring Neutral Proteinase on Bovine and Rabbit Myofibrillar Proteins
Ahmed Oualia, Alain Obleda, Patrick Cottinb, Nadia Merdacib, Andrk Ducastaingb and Christian Valina
a Station de Recherches sur la Viande, INRA de Theix, 63110 Beaumont, France; Laboratoire de Biochimie de la Nutrition, Universite' de Bordeaux I , All.4e des Facultis, 33405 Talence, Cedex, France
(Manuscript received 11 May 1982)
The effects of post-mortem storage and of a low-calcium-requiring neutral proteinase on the myofibrils of beef Longissimus dorsi and Rectus abdominis muscles and rabbit Longissirnus dorsi muscle were studied by measuring the Mg-Ca-enhanced myofibrillar ATPase activity and the changes in banding patterns on electrophoresis in sodium dodecyl sulphate. During ageing, the changes in the ATPase activity and myofibrillar proteins were qualitatively different between rabbit and bovine muscles, whilst differ- ences in intensity were only observed between the two bovine muscles. The most prevalent component appearing upon conditioning had a mol. wt of 27 000 in rabbit muscle and of 30 000 in bovine muscles. In both species, the incubation of myofibrils with the low-calcium-requiring neutral proteinase mimicked the post-mortem changes. This would suggest that this enzyme has a qualitatively different effect on rabbit and bovine myofibrils.
1. Introduction
Many studies have been carried out on the changes in myofibrillar proteins during post-mortem storage, their causes and on the relationship between these alterations and meat tenderi~ation.l-~ The intensity and the speed of these modifications are species-dependent4 and, within a given species, muscle-dependent.3,57
Two changes have been well documented: firstly, the alteration of the actin-myosin interactions, which can be monitored by the modification of the Mg-Ca-enhanced triphosphatase (ATPase) activity of actomyosin6-10; and, secondly, the hydrolysis of myofibrillar proteins.11-13 From the in-vitro results reported, these two myofibrillar protein changes can be ascribed to the action of calcium-activated neutral proteinase (Ca-ANP) and lysosomal proteinases.1910, 14e19
CaANP, a neutral proteinase activated by millimolar (mmol) concentrations of calcium (high- calcium-requiring neutral proteinase) has been studied extensively,l6* 20-23 but one major problem associated with assessing the physiological significance of this proteinase activity and its role in the post-mortem ageing process of meat is that it requires calcium of mmol order, which is much higher than in-vivo and post-mortem concentrations of intracellular calcium. Another problem is that this activity is more or less abolished by sarcoplasmic inhibitors of both high24 and low25-27 molecular weight.
Recently Mellgren2* and Kubota et aLZ9 obtained a low-calcium-requiring form of the protease from canine cardiac and chicken skeletal muscle respectively. This enzyme may be derived from the high-calcium-requiring neutral proteinase by an autolytic p r o c e ~ s , ~ ~ ~ 31 but no investigations con- cerning the identity between the low-calcium-requiring enzyme obtained from the high-calcium- requiring form by autolysis and the in-situ form have been reported. A form of this proteinase with low calcium requirements and its inhibitor were recently purified by Cottin et ~1.25.26 from rabbit skeletal muscle.
466

Effects of storage and neutral proteinase on rnyofibriller proteins 467
The aim of this study was to look at the effects of low-calcium-requiring neutral proteinase in the absence or presence of its inhibitor on bovine and rabbit myofibrils and to see whether the post- mortem changes affecting the myofibrillar structure could be reproduced with this enzyme.
2. Experimental 2.1. Muscle conditioning Bovine Rectus abdominis (RA) and Longissimus dorsi (LD) muscles were removed from one side of the carcass 24 h post-slaughter. Three 3 cm thick transverse portions of each muscle were then taken, vacuum-packed and stored at 6°C. Rabbit LD muscles were removed about 6 h post mortem and portions stored as for the bovine muscles.
2.2. Preparation of myofibrils For the incubation with the low-calcium-requiring neutral proteinase, myofibrils were prepared according to the method of Bendall32 from rabbit and bovine muscles removed 6 and 24 h after death respectively. Myofibrils used to study the post-mortem ageing effect were prepared from the same muscles of different animals after varying storage lengths as indicated in the corresponding figures.
2.3. Preparation of low-calcium-requiring neutral proteinase and inhibitor Low-calcium-requiring neutral proteinase was purified from rabbit muscles by Cottin et al. (un- published). The high-calcium-requiring proteinase inhibitor was purified from rabbit skeletal muscle according to Cottin et ~ 1 . ~ ~
2.4. Enzymatic treatment of the myofibrils Myofibrils suspended in 0 . 0 5 ~ KC1 containing 2 mmol mercaptoethanol (MCE) and 1 mmol NaN3 were washed with 0 . 0 5 ~ tris-acetate buffet pH 7.0 containing 0 . 1 ~ KCl, 2 mmol MCE and 1 mmol NaN3 just before use. Myofibrillar proteins (240-250 mg) were incubated in tris buffer with 1.2 ml of low-calcium-requiring neutral proteinase (50 units ml-1) in the presence of CaC12 at a final concen- tration of 50 pmol and with or without 1.2 ml of the inhibitor preparation in a final volume of 26.5 ml. One unit of enzyme is defined as previously reported.16 In controls, myofibrils were incubated in the same conditions but without enzyme. The assays were carried out at 30°C and the reaction stopped at indicated times, by centrifuging at 4000g for 10 min. The myofibrils pellet was washed twice with 0 . 0 5 ~ KCl containing 2 mmol MCE and 1 mmol NaN3. For the ATPase activity measure- ments, the myofibrils were suspended by gentle homogenisation in 10 ml of the washing solution. An aliquot of this suspension was taken for electrophoretic analysis.
2.5. Myofibrillar ATPase measurements The ATPase activity was measured at different ionic strenths with a Radiometer TTA 31 titrator as previously described.10333 The assays were carried out at 30°C and pH 7.4 with a 0 . 0 1 ~ KOH titrant in a total volume of 6.5 ml. The mixtures contained 4 mmol ATP, 4 mmol MgC12,0.2 mmol CaC12, myofibrils 1-2 mg ml-1 and various concentrations of KCl as indicated.
2.6. Polyacrylamide gel electrophoresis Electrophoresis was performed in the presence of sodium dodecyl sulphate on slab gels (80 x 80 x 0.8 mm) at 200 V (LKB 2103 power supply) overnight in a Pharmacia GE 4 gel electrophoresis appara- tus with tris 0 . 0 5 ~ , boric acid 0 . 0 7 ~ , 2-mercaptoethanol5 mmol, SDS 0.8 % as electrophoresis buffer. Enzyme-treated and post-mortem aged muscle myofibrils were dissolved in 0 . 0 5 ~ tris-HC1 buffer, pH 8.8, containing 7.5 % glycerol, 1 % 2-mercaptoethanol and 2% SDS and heated for 5 min at 100°C. Either homogeneous I0 % acrylamide slab gels (10 % acrylamide, 0.1 % biscrylamide) or gradient slab gels from 10% (10% acrylamide, 0.06% bisacrylamide) to 30% (30% acrylamide,
468 A. Ouali et a/.
1 % bisacrylamide) were used. The gels were stained for 10 min in 0.2% Coomassie brilliant blue R 250 and destained for 10 min in 30 % ethanol-5 % acetic acid with a Pharmacia GD 4 gel destainer attached to a Shandon SAE 2760 destainer power supply.
2.7. Protein determinations Protein concentrations were determined by the Lowry method34 as modified by Hartree35 with bovine serum albumin as standard. Myofibrillar proteins were quantified by the biuret rneth~d.~G
2.8. Data analysis When the ATPase activity was plotted versus KCl concentrations, a linear relationship was obtained over most of the range (Figures 1 and 2). The linear regression for this part (4-6 values) was deter- mined and the slope (B) calculated together with the standard deviation (SB).
For the comparison of different samples, Student's t-test was used with N-2 degrees of freedom.
3. Results
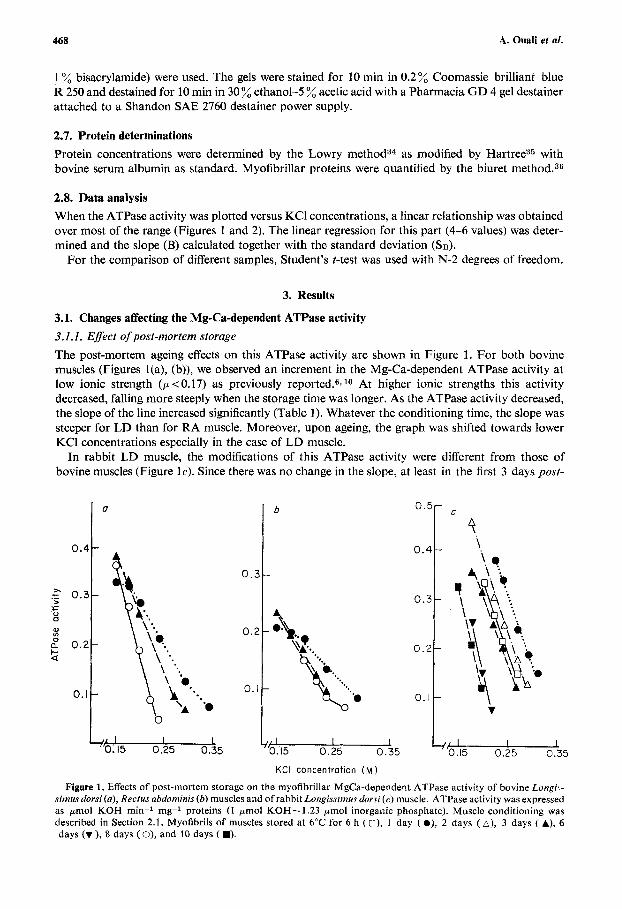
3.1. Changes affecting the Mg-Ca-dependent ATPase activity 3.1.1. Efect of post-mortem storage The post-mortem ageing effects on this ATPase activity are shown in Figure 1. For both bovine muscles (Figures l(a), (b)), we observed an increment in the Mg-Ca-dependent ATPase activity at low ionic strength ( p < 0.17) as previously reported.6, lo At higher ionic strengths this activity decreased, falling more steeply when the storage time was longer. As the ATPase activity decreased, the slope of the line increased significantly (Table 1) . Whatever the conditioning time, the slope was steeper for LD than for RA muscle. Moreover, upon ageing, the graph was shifted towards lower KCl concentrations especially in the case of LD muscle.
In rabbit LD muscle, the modifications of this ATPase activity were different from those of bovine muscles (Figure lc). Since there was no change in the slope, at least in the first 3 days post-
0.4 -
.2 0.3- L;
2 0.2- W In
t- a
0.1 -
O.?
0.2
0. I
b
I I 1 /0.'15 0.25 0.35
KCI concentration ( M )
0.:
0 . L
0.:
0.2
0. I
C
4, \ '. .
Figure 1. Effects of post-mortem storage on the myofibrillar MgCa-dependent ATPase activity of bovine Longi-- simus dorsi (a), Rectus abdoniinis (b) muscles and of rabbit Longissintus dorsi (c) muscle. ATPase activity was expressed as pmol KOH min-1 mg-l proteins (1 pmol KOH= 1.23 pmol inorganic phosphate). Muscle conditioning was described in Section 2.1. Myofibrils of muscles stored at 6°C for 6 h (U), 1 day ( O), 2 days (A), 3 days ( A), 6 days (v ), 8 days ( 0), and 10 days f W).
Effects of storage and neutral proteinase on myofibrillar proteins 469
0.3.'- U 0.3- b 0.8'
0.
> .- z 0.2- 0.2- Q;... 0.4- .- +
Table 1. Changes affecting the slope of the ATPase activity straight-line plots versus KCI concentration (B+SB) upon ageing. The number of experimental values used for the determination of B + SB are in parentheses. Longissimus dorsi (LD) and Rectus a6dominis (RA) muscle conditioning is described in the Section 2.1. Slope values of each muscle were compared and for bovine muscles, comparison between muscles was made. No comparison between
bovine and rabbit muscles could be made.
Time of storage (days)
C
4. a,,
Muscles 1 2 3 6 8 10
0
0 0
I- a
0.1-
Bovine LD 1.522a*
k0.088 (4)
(5)
RA 0.914a k0.050
Rabbit LD 2.4705
k0.178 (5)
0.1- h
I I I I I I I I I I i I I
2.2710 - +0.098
(5)
t0 .084 (4)
I . 584"
+2.808f 2.442s t0 .201 t0 .341
(5) (5)
3 .705~ k 0 . 134
I . 7518 k0.066
(5)
( 5 )
3.783s 3.8098 4.345s - +0.341 k0.208 k0.764
(4) (4) (4)
* Slope values followed by different superior letters were significantly different at the 5 % level according to Student's t-test.
mortem (Table 1). After 6 days of storage, the slope was significantly higher than at 3 days but did not change further since at the eighth and tenth conditioning days, the slopes did not differ signifi- cantly from the sixth day slope. The increased sensitivity of the ATPase activity to ionic strength upon ageing is associated with a shift of the graph towards higher KCl concentrations between 7 and 24 hpost mortem and to lower ones after 1 day. When the LD muscle was excised 6 h after death, the onset of rigor mortis had not been completed (the pH was about 5.9-6.0) and this might explain the graph shift to higher ionic strength between 7 and 24 h post-mortem. Indeed, it was noted that the straight-line plots occurred at the highest KCI concentrations soon after the completion of rigor mortis.
KCI Concentration ( M )
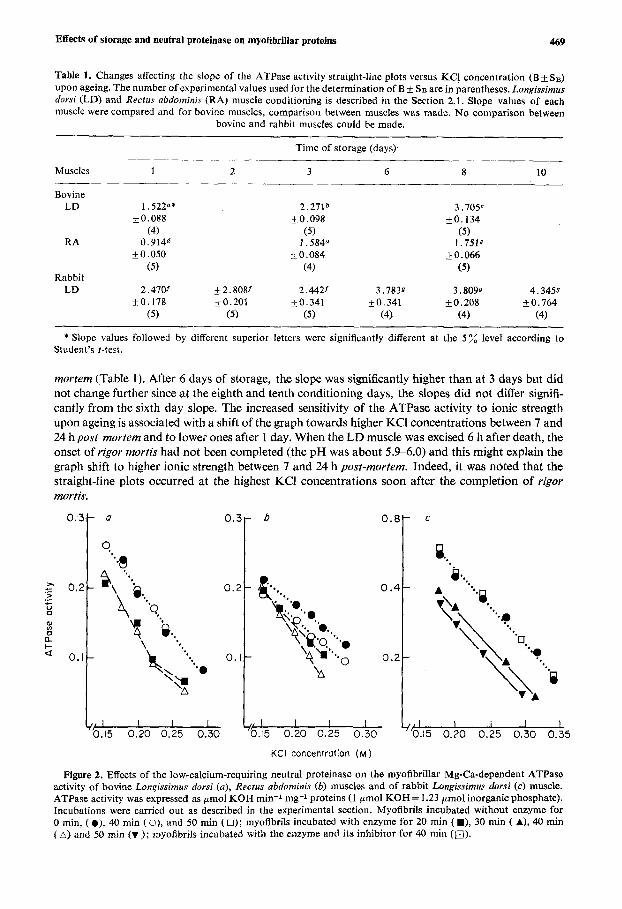
Figure 2. Effects of the low-calcium-requiring neutral proteinase on the myofibrillar Mg-Ca-dependent ATPase activity of bovine Lonpissimus dorsi (a), Rectus abdominis (6) muscles and of rabbit Longissimrts dorsi (c) muscle. ATPase activity was expressed as pmol KOH min-1 mg-1 proteins (1 pmol KOH = 1.23 pmol inorganic phosphate). Incubations were carried out as described in the experimental section. Myofibrils incubated without enzyme for 0 min, ( e), 40 rnin ( o), and 50 rnin ( 0); myofibrils incubated with enzyme for 20 rnin ( m), 30 min ( A), 40 rnin [A) and 50 min (v ); myofibrjls incubated with the enzyme and its inhibitor for 40 min (a).

470 A. Ouali et al.
3.1.2. Effect of low-calcium-requiring neutral proteinase When bovine muscle myofibrils were incubated with the low-calcium-requiring neutral proteinase, the Mg-Ca-dependent ATPase activity underwent changes (Figures 2a and 6 ) similar to those seen upon conditioning of meat (Figures l a and b). Although enhanced ATPase activity at low ionic strength was not observed as upon ageing, an increment in the straight-line slope of LD muscle treated myofibrils and a shift of the graph towards lower KC1 concentrations were noted (Figure 2a and Table 2). For this muscle, the effect of the low-calcium-requiring neutral proteinase reached a maximum after 20 rnin and although significant, the slope increment was much lower than that observed during post-mortem storage (compare Table 1 results with Table 2).
Table 2. Effect of the low-calcium requiring neutral proteinase on the slope of the ATPase activity straight-line plots versus KCl concentration (B f SB). The number of experimental values used for the determination of B k SB are in parentheses. Incubation of Lungissimus dursi (LD) and Rectus abduminis (RA) muscle myofibrils with this proteinase in the absence or presence of its inhibitor were carried out as described in Section 2.2. Slope values of each muscle were compared and for bovine muscles, comparison between muscles was made. No comparison between bovine
and rabbit muscles could be made. ~~~~ ~
Treated myofibrils (min)
Untreated myofibrils (min)
Muscles 0 40 50
With inhibitor Without inhibitor
~ _ _ _ 20 30 40 50 40
Bovine LD 1 .303a*
- + O . 137 ( 5 )
RA 0. 78SC 50.039
( 5 ) Rabbit
LD 2.0651 +o . 121
(6)
2.154f i0 .116
( 5 )
1 .663b i0 .222
(4) 1.051"
50.014 (4)
2.173f i0 .031
(4)
1.597b - +0.076
(4) 1 . 536b 0.74sc
- f0.035 k0.080 (4) (6)
2.152f k0.097
(4)
* Slope values followed by different superior letters were significantly different at the 5 % levels according to Student's t-test.
Upon incubation of RA muscle myofibrils, the control straight-line plot was shifted to lower ionic strengths without any change in slope (Figure 2b and Table 2). This indicates that myofibrils were slightly altered. Nevertheless, when compared to the controls, enzyme treatment increased the slopes significantly (Table 2). Unlike LD muscle, the proteolytic action of the low-calcium-requiring neutral proteinase was not at a maximum after 20 min and significantly went on until at least 40 rnin of treatment. This suggests that the effect of this proteinase was more limited for LD than for RA muscle. When compared to the action of high-calcium-requiring neutral proteinase reported elsewhere,1° it appears that these two muscle proteinases have a quite similar effect on the Mg-Ca- dependent ATPase activity of bovine muscle.
With rabbit LD muscle myofibrils as a substrate, the low-calcium-requiring neutral proteinase faithfully reproduced the effect of post-mortem storage on the Mg-Ca-dependent ATPase activity (Figure 2c). The assay graphs were then shifted towards lower ionic strengths without any change in their slope (Figure 2c and Table 2). However, if the treatment time was longer or if the ratio of enzyme to myofibrillar proteins was higher, an increment in the slope, as was seen upon ageing (Figure l c and Table l), might have occurred.
Whatever the muscle, if the myofibrils were treated by the low-calcium-requiring neutral pro- teinase in the presence of its inhibitor, no change was detectable even after the longer incubation time. This is shown for bovine RA muscle only in Figure 2a and Table 2.
Effects of storage and neutral proteinase on myofibrillar proteins 471
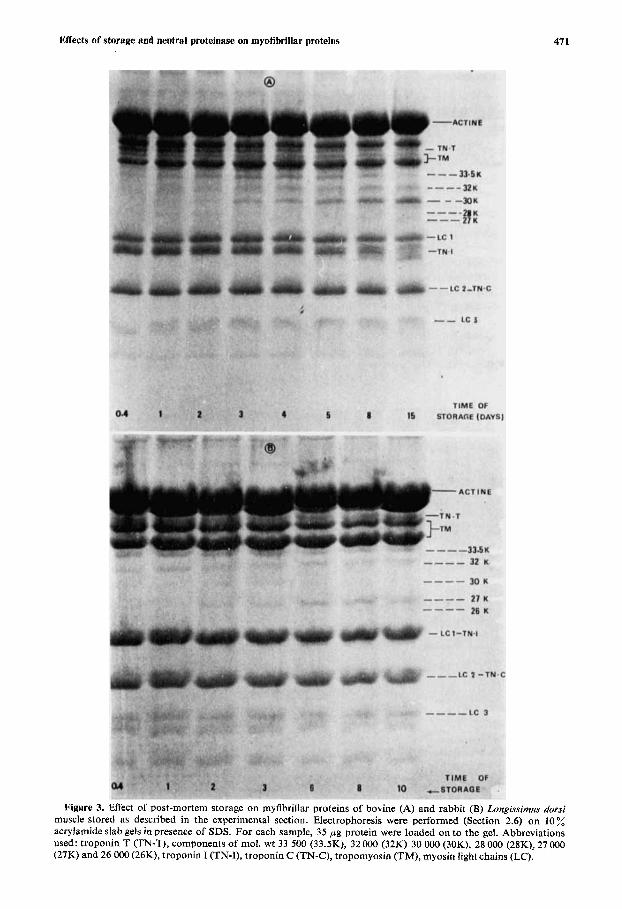
Figure 3. Effect of post-mortem storage on myfibrillar proteins of bovine (A) and rabbit (B) Longissirnus dorsi muscle stored as described in the experimental section. Electrophoresis were performed (Section 2.6) on 10 % acrylamide slab gels in presence of SDS. For each sample, 35 pg protein were loaded on to the gel. Abbreviations used: troponin T (TN-T), components of mol. wt 33 500 (33.5K), 32000 (32K) 30 000 (30K), 28 000 (28K), 27 000 (27K) and 26 000 (26K), troponin I (TN-I), troponin C (TN-C), tropomyosin (TM), myosin light chains (LC).
412 A. Ouali et al.
3.2. Changes affecting the myofibrillar proteins Only the myofibrillar components which run below actin on SDS-polyacrylamide gel electro- phoresis were examined in detail.
3.2.1. Effect of post-mortem storage Upon storage of bovine LD muscle, troponin T disappeared progressively with time. Troponin I was also broken down but much later than troponin T (Figure 3A). Moreover, between tropo- myosin and myosin light chain 1 , five new bands of unknown origin were resolved. This corroborates other resultslS which showed the presence of several bands of similar electrophoretic mobility in bovine Sternomandibuluris muscle stored for 6 days at 15°C. Their mol. wt ranged from 27000 to 33500. The main band corresponded to a component mol. wt of 30 000. The appearance of a similar component in meat has been reported by many workers in different muscles of various species.l,11vs7 This material is thought to arise from proteolysis of troponin T but only indirect probes supported this hypothesis. Therefore, this will be further discussed later.
In addition to this main band, four other components of mol. wt 33 500,32000,28000 and 27000 appeared upon ageing (Figure 3A). The first detected was the mol. wt 33 500 component which in- creased in intensity until the eighth day of storage and then decreased. Thenext band (mol. wt 32000) continually increased in intensity even after 15 days of conditioning. The proteolytic product (mol. wt 28 000) which was just perceptible from days 1-3 post-slaughter had completely disappeared after the fifth day of storage. The last one of mol. wt 27000 which was visible on the eighth day post-
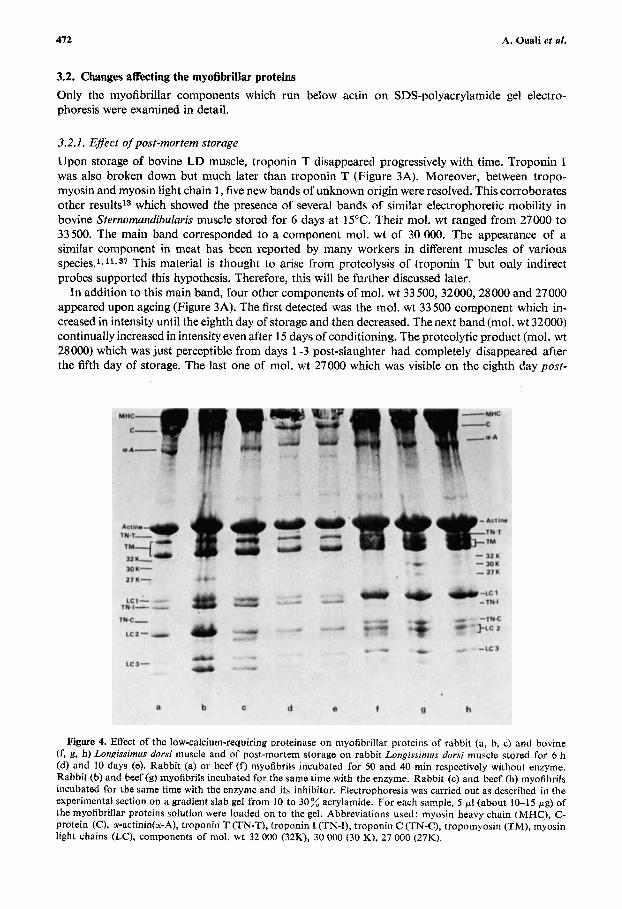
Figure 4. Effect of the low-calcium-requiring proteinase on myofibrillar proteins of rabbit (a, b, c) and bovine (f, g, h) Longissirnus dorsi muscle and of post-mortem storage on rabbit Longissirnus dorsi muscle stored for 6 h (d) and 10 days (e). Rabbit (a) or beef (f) myofibrils incubated for 50 and 40 min respectively without enzyme. Rabbit (b) and beef (g) myofibrils incubated for the same time with the enzyme. Rabbit (c) and beef (h) myofibrils incubated for the same time with the enzyme and its inhibitor. Electrophoresis was carried out as described in the experimental section on a gradient slab gel from 10 to 30% acrylamide. For each sample, 5 pl (about 10-15 pg) of the myofibrillar proteins solution were loaded on to the gel. Abbreviations used: myosin heavy chain (MHC), C- protein (C), or-actinin(a-A), troponin T (TN-T), troponin I (TN-I), troponin C (TN-C), tropomyosin (TM), myosin light chains (LC), components of mol. wt 32 000 (32K), 30 000 (30 K), 27 000 (27K).

Effects of storage and neutral proteinase on myofibrillar proteins 473
mortem increased very markedly in intensity between days 8 and 15. According to their intensity, three of the five components resolved seemed to be more predominant. These have a mol. wt of 30000, 32000 and 33 500.
For RA beef muscle (not shown), the same three main proteolytic components were seen clearly only after 8 days of storage. Therefore, in these two beef muscles, the post-mortem ageing effect was qualitatively similar but the intensity and the speed of the proteolysis was much lower for RA than for LD muscle.
Unlike LD beef muscle, in rabbit LD muscle troponin T did not seem to be altered upon con- ditioning (Figure 3; see also Figures 4(d), and 4(e)). For this muscle, four bands were resolved of mol. wt 26000, 27000, 30000 and 32000 (Figure 3). The predominant ones differed from those of LD bovine muscle since they had mol. wt of 27000 and 32000. Ikeuchi et aZ.8 also reported the appear- ance of a mol. wt 27000 component in rabbit muscles. The 30000 band began to decrease after day 2 of storage and was quite absent after day 10. On the tenth post-portem day, a new band of mol. wt 26000 which was observed had not been seen in beef LD muscle. Therefore, the main difference between rabbit and bovine muscles involved the main proteolytic component that appears upon storage. This has a mol. wt of 27000 in rabbit muscle and of 30000 in beef muscle.
3.2.2. EHect of low-calcium-requiring neutral proteinase When bovine LD muscle myofibrils were treated with the low-calcium-requiring neutral proteinase, two main proteolytic products of mol. wt 27000 and 30000 appeared; the latter was more abundant (Figure 4(g)). This was very similar to the proteolytic components appearing in beef muscle homo- genates incubated with calcium at pH 6.5.38 In the presence of the proteinase inhibitor, three bands were visible (Figure 4(h)). Therefore, with the one-to-one enzyme /inhibitor ratio (v/v) used in this ex- periment, the proteinase was not fully inactivated and the mol. wt 32000 component which was appar- ent in this case was degraded further, since it had disappeared when the inhibitor was not added (Figure 4(g)). Although the low-calcium-requiring neutral proteinase and post-mortem ageing effects were very similar, some differences were found. The 27000 band was much more intense in enzyme-treated myofibrils than in conditioned muscle. Moreover, the 32000 component, the intensity of which increased upon storage, was broken down in myofibrils incubated with this proteinase. Nothing can be concluded about the 33 500 product because, in this electrophoretic system, it migrated with tropomyosin. Nevertheless, the main 30000 component appeared in both post-mortem muscle and enzyme-treated myofibrils.
With rabbit LD muscle myofibrils, the low-calcium-requiring neutral proteinase action led to the appearance of one major band which ran as the 27000 component observed in aged meat (Figures 4(b), 4(e)). Two minor bands of 32000 and 30000 were also seen. The last one was scarcely detectable (Figure 4(b)). As with beef myofibrils, the inhibitor did not completely prevent these proteolytic changes in rabbit myofibrils (Figure 4(c)).
4. Discussion
Many recent resultsl+4-6, 33 prompted us to determine whether the biochemical changes affecting myofibrillar proteins either upon storage or after low-calcium-requiring neutral proteinase treatment were both muscle and species-dependent.
It was found that the sensitivity of Mg-Ca-enhanced ATPase activity to ionic strength increased upon ageing. In beef muscle, this modification is concomitant with an increment in the straight-line slope as previously reported.6.10 It was more noticeable for LD than for RA muscle which confirms the previous results6 and supports the idea that the slope of the ATPase activity graph versus KCl concentration could characterise the meat ageing process. Similar ATPase activity changes were observed on enzyme-treated myofibrils but the action of the low-calcium-requiring neutral pro- teinase seemed to be more restricted for LD than for RA muscle.
The difference in the slopes between these beef muscles was significant until 20 min of treatment by this proteinase and then it levelled off. Upon storage of meat, this difference was always signifi- cant and, moreover, it increased with storage time. This restricted effect on bovine LD muscle might

474 A. Ouali et ol.
mean that, in meat, the action of the low-calcium-requiring neutral proteinase alone did not explain the more marked increment in the slope observed for this muscle during post-mortem storage. These results show that the origin of the myofibrils is very important since the low-calcium-requiring neutral proteinase effect differs in intensity between these two beef muscles despite the similar incubation conditions used.
For rabbit LD muscle, tle alteration in ATPase activity is concomitant with a shift in the straight- line plots towards lower ionic strengths without any change in their slope except for longer storage time. Over the entire range of KCI concentrations examined, the Mg-Ca-dependent ATPase activity increased up to the second daypost mortem and then decreased. This did not agree with the results of Ikeuchi et aZ.7 who showed that, for rabbit LD muscle, this ATPase activity was always higher during the post-mortem storage period than at the time of death. This discrepancy might arise from differences either in the KCI concentrations used for the ATPase activity measurements or in the conditioning temperature. For instance, their muscle samples were stored at 0°C and in this case the ATPase activity modifications were slower than at 6°C. As with beef muscles, the low- calcium-requiring neutral proteinase closely reproduced the post-mortem ageing effects on this ATPase actikity as far as quality and intensity are concerned but the modification of ATPase activity differed qualitatively from that of beef muscle.
Proteolysis is known to occur during conditioning of meat.l> 11-13, 37, 39 In beef LD muscle five new components, ranging from mol. wt 27000 to 33000, were resolved between tropomyosin and myosin light chain 1. The main one was the 30000 component. In rabbit muscle four bands were seen in this region and the main one was the 27000 component. Up to date, the appearance of a 30 000 component either in meat or in CaANP-treated myofibrils has been reported by many workers in chicken,ll, 409 41
pork,37, 42 beefl. 13943 and rabbits, 16 muscle myofibrils. Whatever the species and the muscle, this component was assumed to be the same. Our results showed that post-mortem ageing and low- calcium-requiring neutral proteinase effects were similar but led to a different proteolytic component, depending on the species studied.
This mol. wt 30000 component is thought to arise from the hydrolysis of troponin T first because of the concomitant disappearance of this protein and appearance of the 30000 band and then because the in-vitro proteolysis of purified troponin T by the high-calcium-requiring neutral proteinase led to the formation of several products, one of which had a mol. wt of 30000.1943 However, in contrast to these findings, Azznza et a1.17 did not observe a 30000 component in high-calcium- requiring neutral proteinase-treated rabbit troponin preparations. Moreover, for beef troponin preparations, this proteolytic product was not the final step in troponin T hydrolysis, since it disappeared further and, therefore, did not accumulate in the incubation mixture.1343 Another noticeable feature was that, unlike the appearance of the 30000 component3, 44 the disappearance of troponin T was not related to post-mortem meat tender i~a t ion .~~ Therefore, further investigations are needed to define the origin of this component.
In rabbit LD muscle, the mol. wt 30000 component disappeared soon after death and the main hydrolytic products were those of 27000 and 32000. Unlike beef muscle, the appearance of these proteolytic components was not concomitant with an alteration of troponin T which was not broken down (Figures 3, 4(d) and 4(e)). This agrees with the results of Ikeuchi et aZ.8 and Yang et ~ 1 . ~ ~ who failed to demonstrate any modification in the activity of rabbit muscle regulatory proteins. Whatever the muscle, the low-calcium-requiring neutral proteinase action led to the same main proteolytic components as those observed during storage of the same muscles.
Concerning the efficiency of the high-calcium-requiring neutral proteinase inhibitor previously describedz5, 26 on the low-calcium-requiring form, we have shown that it fully prevented the changes in the ATPase activity whatever the origin of the myofibrils. In contrast, the proteolytic changes detected by SDS-gel electrophoresis are not totally prevented either in bovine or in rabbit myofibrils. The discrepancy in the assessment of the inhibitor efficiency might arise from a difference in sen- sitivity between the ATPase activity method and the SDS-gel electrophoresis one, the latter being the more sensitive. Nevertheless, it appears that this inhibitor is able to act on both forms of the enzyme.
These results show that the changes affecting myofibrillar proteins upon conditioning differed in intensity6 between muscles of the same animal and in quality between the same muscle of the two

Effects of storage and neutral proteinase on myofibriller proteins
species studied. The cause of these variations is unknown and calls for further research. In addition, the low-calcium-requiring neutral proteinase is able to reproduce the post-mortem changes in rabbit and beef muscles qualitatively and its action differs according to the origin of the myofibrils. This has confirmed the major role ascribed to the calcium-activated neutral proteinases in the meat ageing process. Nevertheless, further investigations are needed to improve the knowledge about the enzyme-inhibitor interactions either with low or high-calcium-requiring neutral proteinase and the possible alteration of these interactions during post-mortem storage of meat.
Acknowledgements We thank Dr Jean Chevallier (Institut de Biologie Cellulaire du CNRS., Bordeaux, France) for his very kind assistance and Mrs Nicole Nee1 for her skilful technical contribution.
References
475
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12. 13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
Olson, D. G.; Parrish, F. C.; Dayton, W. R.; Goll, D. G. Effect ofpost mortem storage and calcium activated factor on the myofibrillar proteins of bovine skeletal muscle. J . Food Sci. 1977, 42, 117-124. Penny, I. F.; Dransfield, E. Relationship between toughness and troponin T in conditioned beef. Meat Sci. 1979, 3, 135-141. Parrish, F. C.; Selvig, C. J . ; Culler, R. D.; Zeece, M. G. CAF activity, calcium concentration and the 30 000 D component of tough and tender bovine Longissimus muscle. J . Food Sci. 1981,46, 308-309. Dransfield, E.; Jones, R. C. D.; MacFie, H. J. H. Tenderising in M . Longissirnus dorsi of beef, veal, rabbit, lamb and pork. Meat Sci. 1981, 5, 139-147. Dransfield, E.; Jones, R. C. D. Relationship between tenderness of three beef muscles. J. Sci. Food Agric. 1981,32, 300-304. Ouali, A. Variation between muscles of the effect of ageing on myofibrillar ATPase activity. Science des Aliments 1981, 1, 1-6. Ikeuchi, Y.; Ito, T.; Fukazawa, T. Change in regulation of myofibrillar ATPase activity by calcium during post morteni storage of muscle. J. Food Sci. 1978, 43, 1338-1339. Ikeuchi, Y.; Ito, T.; Fukazawa, T. Change of regulatory activity of tropomyosin and troponin on acto-hedvy- meromyosin ATPase duringpost mortem storage of muscle. J. Food Sci. 1980, 45, 13-16. Chin-Sheng Cheng; Parrish, F. C . Effects ofpost mortem storage conditions on myofibrillar ATPase activity of porcine red and white Semitendinusus muscle. J. Food Sci. 1978, 43, 17-21. Ouali, A. ; Valin, C. Effect of muscle lysosomal enzymes and calcium activated neutral proteinase on myo- fibrillar ATPase activity; relationship with ageing changes. Meat Sci. 1981, 5, 233-245. Hay, J. D.; Currie, R. W.; Wolfe, F. H. Polyacrylamide disc gel electrophoresis of fresh and aged chicken muscle proteins in sodium dodecyl sulphate. J. Food Sci. 1973, 38, 987-990. Takahashi, K.; Saito, H. Post mortem changes in skeletal muscle connectin. J. Biochem. 1979, 85, 1539-1542. Young, 0. A,; Graafhuis, A. E.; Davey, C . L. Post mortem changes in cytoskeleal proteins of muscle. Meat Sci. 1980, 5, 41-55. Valin, C. Post mortem changes in myofibrillar proteins and their relationship to the speed of meat aging. 24th European Meeting of Meat Research Workers Kulmbach, 1978, 11, E3. Okitani, A. ; Matsukura, V. ; Matsumoto, T. ; Fujimaki, M. ; Kato, H. Identification of the proteinases respons- ible for the post nzortem degradation of myofibrillar proteins. 27th European Meeting of Meat Research Workers Wien, 1981, I, 121-124. Azanza, J. L.; Raymond, J.; Robin, J. M.; Cottin, P.; Ducastaing, A. Purification and some physio-chemical and enzymic properties of a calcium ion-activated neutral proteinase from rabbit skeletal muscle. Biochem. J . 1979, 183, 339-347. Azanza, J. L.; Raymond, J.; Robin, J. M . ; Cottin, P.; Ducastaing, A. Proteinase neutre calcium dependante de muscle squelettique de lapin: activiti sur les proteines myofibrillaires. Biochimie 1980, 62, 481-486. Noda, T. ; Isogai, K. ; Hayashi, H. ; Kutunuma, N. Susceptibilities of various myofibrillar proteins to cathepsin B and morphological alteration of isolated myofibrils by this enzyme. J. Biochem. 1981, 90, 371-379. King, N. L.; Kurth, L.; Shortose, W. R. Proteolytic degradation of connectin, a high molecular weight myo- fibrillar protein during heating of meat. Meat Sci. 1981, 5, 389-396. Ouali, A.; Valin, C. Effect of a neutral proteinase on the myofibrillar structure. 22nd European Meeting of Meat Research Workers Malmo, 1976, I, B3. Dayton, W. R.; Reville, W. J.; Goll, D. E.; Stromer, M. H. A calcium-activated protease possibly involved in myofibrillar protein turnover. Partial characterization of the purified enzyme. Biochemistry 1976, 15, 2159- 2167. Ishiura, S.; Murofushi, H.; Ssuzuki, K.; Imahori, K. Studies of a calcium activated neutral proteinase from chicken skeletal muscle. Purification and characterization. J. Biochem. 1978, 84, 225-230.
23. Tsuji, S.; Ishiura, S.; Takahashi-Nakamura, M.; Katamoto, T.; Suzuki, K. ; Iamhori, K. Studies on a Ca-

476 A. Ouali et al.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
activated neutral proteinase of rabbit skeletal muscle. Characterization of sulfhydryl groups and role of calcium ions in this enzyme. J. Biochem. 1981, 90, 1405-1411. Murachi, T. ; Tanaka, K. ; Hatanaka, M. ; Murakami, T. Iiitracellular calcium-dependent protease (calpain) and its high molecular weight endogenous inhibitor (calpastatin). Adv. Enzyme Regulation 1981, 9, 407-427. Cottin, P.; Azanza, J. L.; Vidalenc, P.; Ducastaing, A.; Valin, C.; Ouali, A. Characterizatioii and purification of a calcium-ion activated neutral proteinase inhibitor in rabbit skeletal muscle. Reprod. Nuir. Devel. 1981,
Cottin, P. ; Vidalenc, P. ; Ducastaing, A. Calcium-dependent association between calcium-activated neutral proteinase (CaANP) and its specific inhibitor. FEBS Letters 1981, 136, 221-224. Takahshi-Nakamura, M.; Tsuji, S. ; Suzuki, K. ; Imahori K. Purification and characterization of an inhibitor of calcium activated neutral protease from rabbit skeletal muscle. J. Biochem. 1981, 90, 1583-1589. Mellgren, R. L. Canine cardiac calcium dependent proteases: resolution of two forms with different require- ments for calcium. FEBS Letters 1980, 1009, 129-133. Kutoba, S. I . ; Suzuki, K ; Imahori, K. A new method for the preparation of a calcium-activated neutral pro- tease highly sensitive to calcium ions. Biochem. Biophys. Res. Commun. 1981, 100, 1189-1194. Suzuki, K.; Tsuji, S.; Kubota, S.; Kimura, Y.; Imahori, K. Limited autolysis of calcium-activated neutral protease (CANP) changes its sensitivity to calcium-ions. J. Biothem. 1981, 90, 275-278. Suzuki, K.; Tsuji, S . ; Ishiura, S.; Kimura, Y . ; Kubota, S.; Imahori, K. Autolysis of calcium activated neutral protease of chicken skeletal muscle. J . Biochem. 1981, 90, 1787-1793. Bendall, J. R. A study of the kinetics of the myofibrillar adenosine triphosphatase of rabbit skeletal muscle. Biochem. J. 1961, 81, 520-535. Valin, C.; Touraille, C.; Ouali, A.; Lacourt, A. Effects of electrical stimulation on ageing and eating quality of beef. Sciences des Aliments 1981, 19, 467-476. Lowry, 0. N.; Rosenbrough, N. J.; Farr, A. I . ; Randall, R. J. Protein measurement with the Folin phenol reagent. J . Biol. Chem. 1951, 193, 265-268. Hartree, E. F. Determination of protein: a modification of the Lowry method that gives a linear photometric response. Anal. Biochem. 1972, 48, 422-427. Gornall, A. G.; Bardawill, C. T.; David, M. M. Determination of serum proteins by means of the biuret reagent. J. Biol. Chem. 1949, 117, 751-766. Penny, 1. F. The effect of conditioning on the myofibrillar proteins of pork muscle. J . Sci. Food Agric. 1976, 27, 1147-1155.
21, 309-317.
Penny, I. F.; Fergusson-Pryce, R. Measurement of autolysis in beef muscle homogenates. Meat Sci. 1979, 3, 121-134. Ito, T.; Sung, S . K.; Fukazawa, T. Change of acto-heavy meromyosin ATPase of rabbit skeletal muscle during post mortem storage. J. Agric. Food Chem. 1978, 26, 324-326. Samejima, K.; Wolfe, F. H. Degradation of myofibrillar protein components during post mortem ageing of chicken muscle. J. Food Sci. 1976, 41, 250-254. Yamamoto, K.; Samejima, K.; Yasui, T. A comparative study of the changes in hen pectoral muscle during storage at 4°C and -20°C. J . Food Sci. 1977,42, 1642-1645. Kang, C. G.; Mugurama, M.; Fukazawa, T.; Ito, T. Difference in 2-line removal between normal and PSE porcine myofibrils. J. Food Sci. 1978, 43, 508-514. Penny, I. F. The action of a muscle proteinase on the myofibrillar proteins of bovine muscle. J . Sci. Food Agric. 1974, 25, 1273-1284. MacBride, M. A.; Parrish, F. C. The 30 000 dalton component of tender bovine Longissirnus muscle. J. Food Sci. 1977, 42, 1627-1629. George, A. R.; Bendall, J. R.; Jones, R. C. D. The tenderising effect of electrical stimulation of beef car- casses. Meat Science 1980, 4, 51-68. Yang, R.; Okitani, A.; Fujimaki, M. Post mortem changes in regulatory proteins of rabbit muscle. Agric. Biol. Chem. 1978,42,555-563.




















