
Investigation on CAST, CAPN1 and CAPN3 porcine gene polymorphisms and expression in relation to...
7
Investigation on CAST, CAPN1 and CAPN3 porcine gene polymorphisms and expression in relation to post-mortem calpain activity in muscle and meat quality G. Gandolfi a , L. Pomponio b , P. Ertbjerg b , A.H. Karlsson b , L. Nanni Costa a , R. Lametsch b , V. Russo a , R. Davoli a, ⁎ a Department of Agri-food Protection and Valorization, Faculty of Agriculture, University of Bologna, via F.lli Rosselli 107, 42123 Reggio Emilia, Italy b Department of Food Science, Faculty of Life Sciences, University of Copenhagen, Rolighedsvej 30, DK-1958 Frederiksberg C, Denmark abstract article info Article history: Received 22 July 2010 Received in revised form 22 February 2011 Accepted 28 February 2011 Keywords: Calpain activity CAST Pork quality Polymorphism Proteolysis This study aimed to detect variability in CAST, CAPN1 and CAPN3 porcine genes and to investigate the effect of CAST and CAPN1 polymorphisms on the activity of native and autolyzed μ-calpain and m-calpain, measured from 1 to 72 h post-mortem in Longissimus dorsi (LD) muscle of 30 pigs. Effects of polymorphisms on meat quality parameter such as pH, color and drip loss were also evaluated. Samples carrying CAST EU137105: g.76,872AA genotype showed higher autolyzed μ-calpain activity 24 and 72 h post-mortem, as well as lower drip loss values. Expression of CAST, CAPN1 and CAPN3 was assessed in LD muscles divergent for shear force. Higher CAST and CAPN3 expression was found in LD with high shear force (P b 0.2), confirming a direct role for calpastatin but not for calpain 3 in meat tenderization. In conclusion, CAST gene affected post-mortem activation time of calpain and drip loss. © 2011 Elsevier Ltd. All rights reserved. 1. Introduction Post-mortem muscle proteolysis affects many quality traits of pig meat, such as tenderness, drip loss and color, which in turn have a strong effect on consumer acceptance. In particular, tenderness is considered as the most important parameter regarding eating quality (Moeller et al., 2010). The calpain system is thought to play an important role in post- mortem muscle proteolysis and meat tenderization processes (Huff- Lonergan & Lonergan, 2005; Koohmaraie, 1992). In skeletal muscle, the calpain system consists of the ubiquitously expressed μ-calpain and m-calpain, the skeletal muscle specific calpain 3, and the endogenous inhibitor of μ-calpain and m-calpain, calpastatin (Goll, Thompson, Li, Wei, & Cong, 2003). The ubiquitous m- and μ-calpains are calcium dependent proteases composed of a catalytic subunit of 80 kDa and a regulative subunit of 28 kDa, which both autolyze in presence of calcium (Cong, Goll, Peterson, & Kapprell, 1989). By autolysis, the catalytic subunit is reduced to a 76-kDa form, which has a reduced calcium requirement for activity (Goll et al., 2003). The detection of the autolyzed form of the ubiquitous calpains in muscle is thought to indicate proteolytic activity (Geesink & Koohmaraie, 1999) and an earlier appearance of autolysis was reported to be associated to an earlier degradation of myofibrillar protein (Melody et al., 2004). High calpain activity induces the cleavage of specific myofibrillar protein such as titin, desmin and vinculin, and leads to improvement of tenderness, whereas high levels of calpastatin are associated with decreased proteolysis and increased meat toughness (Kemp, Sensky, Bardsley, Buttery, & Parr, 2010; Kent, Spencer, & Koohmaraie, 2004). Several studies demonstrated that, among the ubiquitous calpains, μ-calpain has the most significant role in post- mortem proteolysis and meat tenderization (Kemp et al., 2010; Geesink, Kuchay, Chishti, & Koohmaraie, 2006), while m-calpain is thought to have a marginal role, at least in bovine and ovine muscle (Camou, Marchello, Thompson, Mares, & Goll, 2007; Veiseth, Shackelford, Wheeler, & Koohmaraie, 2001). However, the detection of autolyzed m-calpain activity, along with the decrease in activity of native form, supported a role of m-calpain in post-mortem proteolysis in pork (Pomponio et al., 2008). Calpain 3 is expressed almost exclusively in skeletal muscle (Sorimachi et al., 1989). The facts that calpain 3 binds the giant myofibrillar protein titin and it autolyses in post-mortem muscle initially suggested that it could have a role in meat tenderization (Kemp et al., 2010). However, discordant results have been obtained in this regard: a positive correlation between calpain 3 expression level and tenderness has been detected in sheep (Ilian, Bekhit, & Bickerstaffe, 2004; Ilian et al., 2001) but not in pig meat (Parr et al., 1999). Further study on calpain 3 knock out mice suggested that calpain 3 is not involved in post-mortem muscle proteolysis (Geesink, Taylor & Koohmaraie, 2005). Meat Science 88 (2011) 694–700 ⁎ Corresponding author at: Department of Agri-food Protection and Valorization, Faculty of Agriculture, Università di Bologna, via F.lli Rosselli 107, 42123 Reggio Emilia, Italy. Tel.: +39 0522290507; fax: +39 0522290523. E-mail addresses: greta.gandolfi[email protected] (G. Gandolfi), [email protected] (L. Pomponio), [email protected] (P. Ertbjerg), [email protected] (A.H. Karlsson), [email protected] (L. Nanni Costa), [email protected] (R. Lametsch), [email protected] (V. Russo), [email protected] (R. Davoli). 0309-1740/$ – see front matter © 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.meatsci.2011.02.031 Contents lists available at ScienceDirect Meat Science journal homepage: www.elsevier.com/locate/meatsci
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Investigation on CAST, CAPN1 and CAPN3 porcine gene polymorphisms and expression in relation to...

Meat Science 88 (2011) 694–700
Contents lists available at ScienceDirect
Meat Science
j ourna l homepage: www.e lsev ie r.com/ locate /meatsc i
Investigation on CAST, CAPN1 and CAPN3 porcine gene polymorphisms andexpression in relation to post-mortem calpain activity in muscle and meat quality
G. Gandolfi a, L. Pomponio b, P. Ertbjerg b, A.H. Karlsson b, L. Nanni Costa a, R. Lametsch b,V. Russo a, R. Davoli a,⁎a Department of Agri-food Protection and Valorization, Faculty of Agriculture, University of Bologna, via F.lli Rosselli 107, 42123 Reggio Emilia, Italyb Department of Food Science, Faculty of Life Sciences, University of Copenhagen, Rolighedsvej 30, DK-1958 Frederiksberg C, Denmark
⁎ Corresponding author at: Department of Agri-fooFaculty of Agriculture, Università di Bologna, via F.lli RosItaly. Tel.: +39 0522290507; fax: +39 0522290523.
E-mail addresses: [email protected] (G. Gand(L. Pomponio), [email protected] (P. Ertbjerg), [email protected]@unibo.it (L. Nanni Costa), [email protected]@unibo.it (V. Russo), roberta.davoli@unib
0309-1740/$ – see front matter © 2011 Elsevier Ltd. Aldoi:10.1016/j.meatsci.2011.02.031
a b s t r a c t
a r t i c l e i n f oArticle history:Received 22 July 2010Received in revised form 22 February 2011Accepted 28 February 2011
Keywords:Calpain activityCASTPork qualityPolymorphismProteolysis
This study aimed to detect variability in CAST, CAPN1 and CAPN3 porcine genes and to investigate the effect ofCAST and CAPN1 polymorphisms on the activity of native and autolyzed μ-calpain and m-calpain, measuredfrom 1 to 72 h post-mortem in Longissimus dorsi (LD) muscle of 30 pigs. Effects of polymorphisms on meatquality parameter such as pH, color and drip loss were also evaluated. Samples carrying CAST EU137105:g.76,872AA genotype showed higher autolyzed μ-calpain activity 24 and 72 h post-mortem, as well as lowerdrip loss values. Expression of CAST, CAPN1 and CAPN3 was assessed in LD muscles divergent for shear force.Higher CAST and CAPN3 expression was found in LDwith high shear force (Pb0.2), confirming a direct role forcalpastatin but not for calpain 3 in meat tenderization. In conclusion, CAST gene affected post-mortemactivation time of calpain and drip loss.
d Protection and Valorization,selli 107, 42123 Reggio Emilia,
olfi), [email protected] (A.H. Karlsson),ku.dk (R. Lametsch),o.it (R. Davoli).
l rights reserved.
© 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Post-mortem muscle proteolysis affects many quality traits of pigmeat, such as tenderness, drip loss and color, which in turn have astrong effect on consumer acceptance. In particular, tenderness isconsidered as the most important parameter regarding eating quality(Moeller et al., 2010).
The calpain system is thought to play an important role in post-mortem muscle proteolysis and meat tenderization processes (Huff-Lonergan & Lonergan, 2005; Koohmaraie, 1992). In skeletal muscle,the calpain system consists of the ubiquitously expressed μ-calpainand m-calpain, the skeletal muscle specific calpain 3, and theendogenous inhibitor of μ-calpain and m-calpain, calpastatin (Goll,Thompson, Li, Wei, & Cong, 2003). The ubiquitous m- and μ-calpainsare calcium dependent proteases composed of a catalytic subunit of80 kDa and a regulative subunit of 28 kDa, which both autolyze inpresence of calcium (Cong, Goll, Peterson, & Kapprell, 1989). Byautolysis, the catalytic subunit is reduced to a 76-kDa form, which hasa reduced calcium requirement for activity (Goll et al., 2003).
The detection of the autolyzed form of the ubiquitous calpains inmuscle is thought to indicate proteolytic activity (Geesink&Koohmaraie,
1999) and an earlier appearance of autolysis was reported to beassociated to an earlier degradation of myofibrillar protein (Melodyet al., 2004). High calpain activity induces the cleavage of specificmyofibrillar protein such as titin, desmin and vinculin, and leads toimprovement of tenderness, whereas high levels of calpastatin areassociated with decreased proteolysis and increased meat toughness(Kemp, Sensky, Bardsley, Buttery, & Parr, 2010; Kent, Spencer, &Koohmaraie, 2004). Several studies demonstrated that, among theubiquitous calpains, μ-calpain has the most significant role in post-mortem proteolysis andmeat tenderization (Kemp et al., 2010; Geesink,Kuchay, Chishti, & Koohmaraie, 2006), while m-calpain is thought tohave a marginal role, at least in bovine and ovine muscle (Camou,Marchello, Thompson, Mares, & Goll, 2007; Veiseth, Shackelford,Wheeler, & Koohmaraie, 2001). However, the detection of autolyzedm-calpain activity, along with the decrease in activity of native form,supported a role of m-calpain in post-mortem proteolysis in pork(Pomponio et al., 2008).
Calpain 3 is expressed almost exclusively in skeletal muscle(Sorimachi et al., 1989). The facts that calpain 3 binds the giantmyofibrillar protein titin and it autolyses in post-mortem muscleinitially suggested that it could have a role in meat tenderization(Kemp et al., 2010). However, discordant results have been obtainedin this regard: a positive correlation between calpain 3 expressionlevel and tenderness has been detected in sheep (Ilian, Bekhit, &Bickerstaffe, 2004; Ilian et al., 2001) but not in pig meat (Parr et al.,1999). Further study on calpain 3 knock out mice suggested thatcalpain 3 is not involved in post-mortemmuscle proteolysis (Geesink,Taylor & Koohmaraie, 2005).

695G. Gandolfi et al. / Meat Science 88 (2011) 694–700
The porcine calpastatin (CAST) gene is located on SSC2 (2q2.1–2q2.4; Ernst, Robic, Yerle, Wang & Rothschild, 1998), where QTL formeat quality were described (Meyers, Rodriguez-Zas & Beever, 2007;Stearns, Beever, Southey, Ellis, McKeith & Rodriguez-Zas, 2005).CAPN1 gene, encoding for μ-calpain large subunit, is located on SSC 2(2p14–p27; Fridolfsson et al., 1997), and calpain 3 (CAPN3) gene onSSC1 (1q15–q17; Briley, Riggs, Womack, Hancock & Bidwell, 1996).
Several CAST polymorphisms have been described and some havebeen associated with carcass quality and production traits (Škrlepet al., 2010; Lindholm-Perry et al., 2009; Meyers & Beever, 2008;Ciobanu et al., 2004). In cattle, association with meat tenderness hasbeen reported for CAPN3 and CAPN1 gene polymorphisms (Barendse,Harrison, Bunch & Thomas, 2008; Page et al., 2002), but noinvestigation have been performed in pigs.
The effect of porcine CAST and CAPN1 single nucleotide polymor-phisms (SNP) on calpain activity in muscle during post-mortemstorage has never been investigated. Moreover, analysis of geneexpression of CAST in relation to pork quality parameter has beenperformed, pointing out a relation between lower expression leveland favorable pH values (Sieczkowska et al., 2010), but relationshipbetween gene expression and tenderness has never been reported.
2. Purpose of the research
The present study aimed to detect SNP in CAST, CAPN1 and CAPN3porcine genes and to investigate the effects of CAST and CAPN1 SNP onthe time course activity of the native and autolyzed μ-calpain and ofm-calpain in pig Longissimus dorsi (LD) muscle, from 1 to 72 h post-mortem. The effect of the SNP on meat quality parameters was alsoevaluated. Moreover, we analyzed the expression of CAST, CAPN1 andCAPN3 genes in LD muscles differing in shear force.
3. Material and methods
3.1. Samples and meat parameter measurements
Forty Italian Duroc×(Landrace×Large White) crossbred pigs, pen-fed and finished in Northern Italy under standard Parma ham practice,were slaughtered on two different days (10 pigs slaughtered on day 1and30pigs slaughteredonday2). Average carcassweightwas127.84±9.86 kg. Post-slaughter storage conditions and measurements were thesame as described in Pomponio, Ertbjerg, Karlsson, Nanni Costa andLametsch (2010). Briefly, after slaughter the carcasseswere divided intocuts and the loinswere stored in a chilling roomat 2 °C. In LDmuscle, pHwasmeasured at 1, 3, 6, 24 and 72 h post-mortem (Eutech InstrumentspH 6), at the 7th rib of the right side of the carcass. Color measurements(lightness L*, redness a* and yellowness b*) were recorded 1 h, 4.5 h,6 h, 24 h, 72 h and 144 h post-mortem (Minolta CRC 300 instrument,Minolta Co., Tokyo, Japan) at the same loin location.
Drip loss was determined as percentage of fluid loss in a 5 cm sliceof LD muscle, collected 24 post-mortem between the 7th and the 9thrib, placed in a commercial tray coveredwith plastic film and stored at4 °C for 2 days (72 h post-mortem).
Cooking loss and shear force (Warner–Bratzler test) weredetermined following Honikel (1998) in LD muscle samples at 24 h,72 h and 144 h post-mortem. For the 30 pigs slaughtered on day 2,samples for calpain activity were taken 1 h and 6 h post-mortem atthe 8th rib and stored at −80 °C. The next days, samples fromdeboned LD stored at 4 °C were collected and frozen at−80 °C, for thedetermination of calpain activity at 24 h and 72 h and myofibrilfragmentation at 24 h, 72 h and 144 h post-mortem.
For DNA and RNA extraction, LD muscle samples were collectedimmediately after slaughter, snap frozen in liquid nitrogen and storedat −20 °C or −80 °C.
3.2. Myofibril fragmentation and calpain activity
Myofibril fragmentation, expressed as particle size, was determinedat 24 h, 72 h and 144 h post-mortem as described in Pomponio et al.(2010). The sizes of myofibrils were determined as surface meandiameter D (3,2). Calpain activity at 1, 6, 24, 72 h post-mortem wasmeasured using a casein zymographymethod, as reported in Pomponioet al. (2010).
3.3. DNA extraction, PCR amplification, sequencing and genotyping
Total DNA was extracted from muscle tissue samples by standardmethod. The primer pairs CAST_SNP, CAPN1_SNP and CAPN3_SNP(Table 1) were designed on the GenBank accession numberAY594692.1, DQ674244.1 and DQ388766.1, respectively (http://www.ncbi.nlm.nih.gov/genbank/), using Primer3 software v.0.4.0(http://frodo.wi.mit.edu/primer3/). PCR amplifications were per-formed in a total volume of 20 μl containing 2 μl of standard buffer,1.5 mM MgCl2, 0.5 μM of each primer, 160 μM dNTP, 1 U EuroTaqpolymerase (Promega, Mannheim, Germany) and 50–100 ng oftemplate DNA. Cycling conditions consisted of an initial denaturationat 95 °C for 5 min, 35 cycles of 95 °C for 30 s, annealing temperature of65 °C or 60 °C (for CAST or CAPN1 and CAPN3, respectively) for 30 sand 72 °C for 30 s, followed by a final step of 72 °C for 5 min. The PCRproducts, purified with ExoSAP-IT® (USB Corporation, Cleveland,Ohio, USA) following the manufacturer's protocol, were sequenced onboth strands using the BigDye v. 3.1 Cycle sequencing kit (AppliedBiosystems, Foster City, CA, USA) and the four-capillary system ABIPRISM 3100-Avant Genetic Analyzer (Applied Biosystems).
For the genotyping of CAST and CAPN1 SNP by RFLP method, 5 μl ofPCR products were digested by 3 U of HinfI or HhaI restrictionenzymes (Fermentas, Hanover, MD 21076, USA), respectively, and theresulting products were resolved on 10% polyacrylamide/bisacryla-mide gels stained with ethidium bromide.
3.4. RNA isolation, cDNA synthesis and quantitative real-time PCR
Total RNA was isolated using TRIzol® Reagent (Invitrogen,Carlsbad, CA, USA), following the manufacturer's protocol. The RNAwas quantified by NanoDrop (ND-1000, NanoDrop Technologies,USA) and the quality was checked by electrophoresis of 500 ng RNAon ethidium bromide stained 1% agarose gels. After DNase treatment(TURBO DNA-free™, Ambion, Applied Biosystem), 1 μg of total RNAwas reverse transcribed at 42 °C using Improm-II™ reverse transcrip-tase and an Oligo(dT) (Promega) according to the manufacturer'sinstructions.
Quantitative real-time PCR reactions were performed on a Light-Cycler 1.0 System (Roche Diagnostics, Mannheim, Germany) using5 μl of the SYBR® Premix Ex Taq™ (Takara Bio Inc., Otsu, Shiga, Japan),0.5 μM of each primer (CAST_Expr, CAPN1_Expr and CAPN3_Expr,Table 1) and about 100 ng of cDNA, in a total volume of 10 μl. Allreactions were performed in triplicate, and consisted of an initialdenaturation step at 95 °C for 30 s and 45 cycles of 95 °C for 5 s and63 °C for 20 s. After the real-time PCR reactions, melting curveanalysis was performed to confirm specificity of the amplification.Threshold cycles were converted to mRNA molecules/μl using astandard curve (Bustin, 2004) generated from serial dilutions (108–50copies/μl) of the PCR amplicon obtained with primers pairs CAST-St,CAPN1-St and CAPN3-St (Table 1), flanking the annealing region ofthe primers used in the real-time PCR. The calculated average mRNAmolecules/μl for each sample was normalized by dividing with thegeometric mean of the expression of two housekeeping genes in thegiven sample, as suggested by Bustin (2004) and by Vandesompeleet al. (2002). The housekeeping genes used in this study were beta-2-microglobulin (B2M) and RNA polymerase II polypeptide A (POLR2A),and the primer pairs used are listed in Table 1.
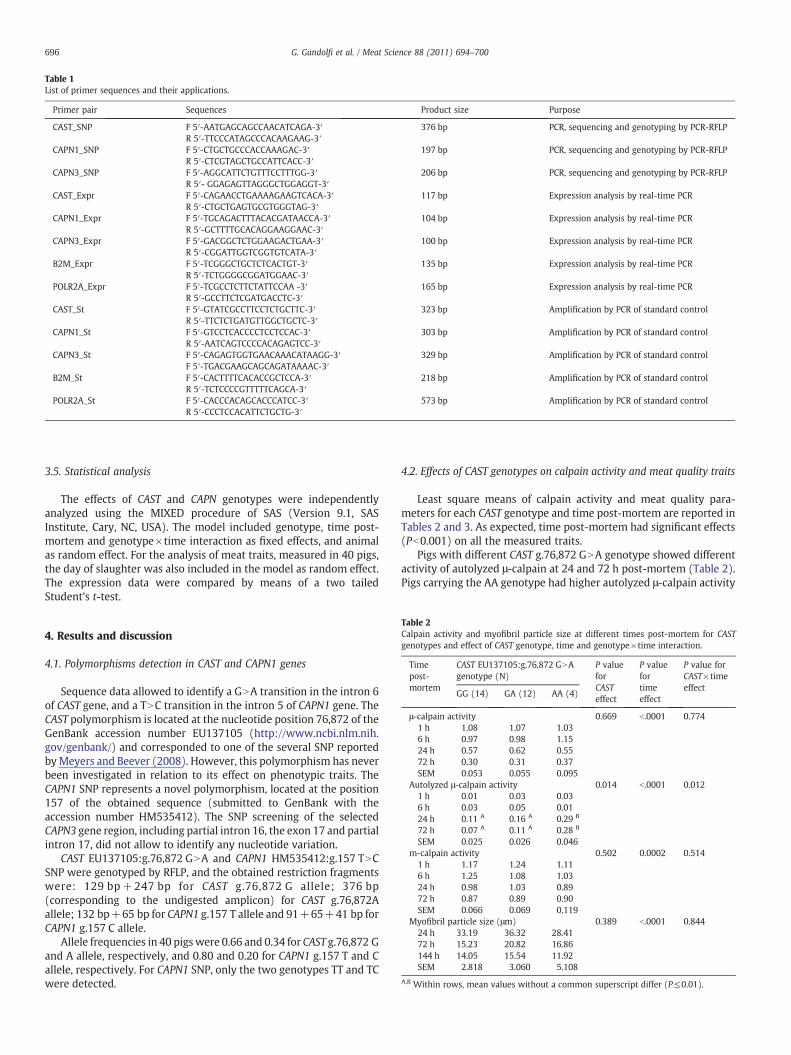
Table 1List of primer sequences and their applications.
Primer pair Sequences Product size Purpose
CAST_SNP F 5′-AATGAGCAGCCAACATCAGA-3′ 376 bp PCR, sequencing and genotyping by PCR-RFLPR 5′-TTCCCATAGCCCACAAGAAG-3′
CAPN1_SNP F 5′-CTGCTGCCCACCAAAGAC-3′ 197 bp PCR, sequencing and genotyping by PCR-RFLPR 5′-CTCGTAGCTGCCATTCACC-3′
CAPN3_SNP F 5′-AGGCATTCTGTTTCCTTTGG-3′ 206 bp PCR, sequencing and genotyping by PCR-RFLPR 5′- GGAGAGTTAGGGCTGGAGGT-3′
CAST_Expr F 5′-CAGAACCTGAAAAGAAGTCACA-3′ 117 bp Expression analysis by real-time PCRR 5′-CTGCTGAGTGCGTGGGTAG-3′
CAPN1_Expr F 5′-TGCAGACTTTACACGATAACCA-3′ 104 bp Expression analysis by real-time PCRR 5′-GCTTTTGCACAGGAAGGAAC-3′
CAPN3_Expr F 5′-GACGGCTCTGGAAGACTGAA-3′ 100 bp Expression analysis by real-time PCRR 5′-CGGATTGGTCGGTGTCATA-3′
B2M_Expr F 5′-TCGGGCTGCTCTCACTGT-3′ 135 bp Expression analysis by real-time PCRR 5′-TCTGGGGCGGATGGAAC-3′
POLR2A_Expr F 5′-TCGCCTCTTCTATTCCAA -3′ 165 bp Expression analysis by real-time PCRR 5′-GCCTTCTCGATGACCTC-3′
CAST_St F 5′-GTATCGCCTTCCTCTGCTTC-3′ 323 bp Amplification by PCR of standard controlR 5′-TTCTCTGATGTTGGCTGCTC-3′
CAPN1_St F 5′-GTCCTCACCCCTCCTCCAC-3′ 303 bp Amplification by PCR of standard controlR 5′-AATCAGTCCCCACAGAGTCC-3′
CAPN3_St F 5′-CAGAGTGGTGAACAAACATAAGG-3′ 329 bp Amplification by PCR of standard controlF 5′-TGACGAAGCAGCAGATAAAAC-3′
B2M_St F 5′-CACTTTTCACACCGCTCCA-3′ 218 bp Amplification by PCR of standard controlR 5′-TCTCCCCGTTTTTCAGCA-3′
POLR2A_St F 5′-CACCCACAGCACCCATCC-3′ 573 bp Amplification by PCR of standard controlR 5′-CCCTCCACATTCTGCTG-3′
696 G. Gandolfi et al. / Meat Science 88 (2011) 694–700
3.5. Statistical analysis
The effects of CAST and CAPN genotypes were independentlyanalyzed using the MIXED procedure of SAS (Version 9.1, SASInstitute, Cary, NC, USA). The model included genotype, time post-mortem and genotype×time interaction as fixed effects, and animalas random effect. For the analysis of meat traits, measured in 40 pigs,the day of slaughter was also included in the model as random effect.The expression data were compared by means of a two tailedStudent's t-test.
Table 2Calpain activity and myofibril particle size at different times post-mortem for CASTgenotypes and effect of CAST genotype, time and genotype×time interaction.
Timepost-mortem
CAST EU137105:g.76,872 GNAgenotype (N)
P valueforCASTeffect
P valuefortimeeffect
P value forCAST×timeeffect
GG (14) GA (12) AA (4)
μ-calpain activity 0.669 b.0001 0.7741 h 1.08 1.07 1.036 h 0.97 0.98 1.1524 h 0.57 0.62 0.5572 h 0.30 0.31 0.37SEM 0.053 0.055 0.095
Autolyzed μ-calpain activity 0.014 b.0001 0.0121 h 0.01 0.03 0.036 h 0.03 0.05 0.0124 h 0.11 A 0.16 A 0.29 B
72 h 0.07 A 0.11 A 0.28 B
SEM 0.025 0.026 0.046m-calpain activity 0.502 0.0002 0.514
1 h 1.17 1.24 1.116 h 1.25 1.08 1.0324 h 0.98 1.03 0.8972 h 0.87 0.89 0.90SEM 0.066 0.069 0.119
Myofibril particle size (μm) 0.389 b.0001 0.84424 h 33.19 36.32 28.4172 h 15.23 20.82 16.86144 h 14.05 15.54 11.92SEM 2.818 3.060 5.108
A,B Within rows, mean values without a common superscript differ (P≤0.01).
4. Results and discussion
4.1. Polymorphisms detection in CAST and CAPN1 genes
Sequence data allowed to identify a GNA transition in the intron 6of CAST gene, and a TNC transition in the intron 5 of CAPN1 gene. TheCAST polymorphism is located at the nucleotide position 76,872 of theGenBank accession number EU137105 (http://www.ncbi.nlm.nih.gov/genbank/) and corresponded to one of the several SNP reportedbyMeyers and Beever (2008). However, this polymorphism has neverbeen investigated in relation to its effect on phenotypic traits. TheCAPN1 SNP represents a novel polymorphism, located at the position157 of the obtained sequence (submitted to GenBank with theaccession number HM535412). The SNP screening of the selectedCAPN3 gene region, including partial intron 16, the exon 17 and partialintron 17, did not allow to identify any nucleotide variation.
CAST EU137105:g.76,872 GNA and CAPN1 HM535412:g.157 TNCSNP were genotyped by RFLP, and the obtained restriction fragmentswere: 129 bp+247 bp for CAST g.76,872 G allele; 376 bp(corresponding to the undigested amplicon) for CAST g.76,872Aallele; 132 bp+65 bp for CAPN1 g.157 T allele and 91+65+41 bp forCAPN1 g.157 C allele.
Allele frequencies in 40 pigswere 0.66 and 0.34 for CAST g.76,872 Gand A allele, respectively, and 0.80 and 0.20 for CAPN1 g.157 T and Callele, respectively. For CAPN1 SNP, only the two genotypes TT and TCwere detected.
4.2. Effects of CAST genotypes on calpain activity and meat quality traits
Least square means of calpain activity and meat quality para-meters for each CAST genotype and time post-mortem are reported inTables 2 and 3. As expected, time post-mortem had significant effects(Pb0.001) on all the measured traits.
Pigs with different CAST g.76,872 GNA genotype showed differentactivity of autolyzed μ-calpain at 24 and 72 h post-mortem (Table 2).Pigs carrying the AA genotype had higher autolyzed μ-calpain activity
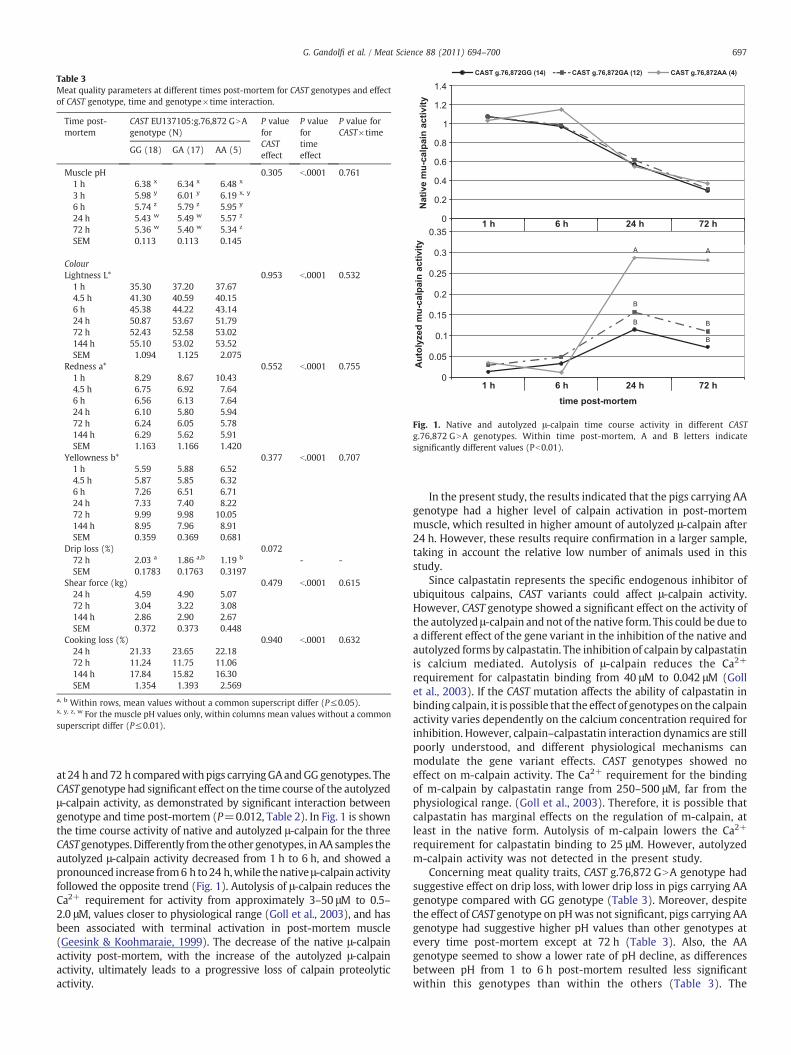
Table 3Meat quality parameters at different times post-mortem for CAST genotypes and effectof CAST genotype, time and genotype×time interaction.
Time post-mortem
CAST EU137105:g.76,872 GNAgenotype (N)
P valueforCASTeffect
P valuefortimeeffect
P value forCAST×time
GG (18) GA (17) AA (5)
Muscle pH 0.305 b.0001 0.7611 h 6.38 x 6.34 x 6.48 x
3 h 5.98 y 6.01 y 6.19 x, y
6 h 5.74 z 5.79 z 5.95 y
24 h 5.43 w 5.49 w 5.57 z
72 h 5.36 w 5.40 w 5.34 z
SEM 0.113 0.113 0.145
ColourLightness L* 0.953 b.0001 0.532
1 h 35.30 37.20 37.674.5 h 41.30 40.59 40.156 h 45.38 44.22 43.1424 h 50.87 53.67 51.7972 h 52.43 52.58 53.02144 h 55.10 53.02 53.52SEM 1.094 1.125 2.075
Redness a* 0.552 b.0001 0.7551 h 8.29 8.67 10.434.5 h 6.75 6.92 7.646 h 6.56 6.13 7.6424 h 6.10 5.80 5.9472 h 6.24 6.05 5.78144 h 6.29 5.62 5.91SEM 1.163 1.166 1.420
Yellowness b* 0.377 b.0001 0.7071 h 5.59 5.88 6.524.5 h 5.87 5.85 6.326 h 7.26 6.51 6.7124 h 7.33 7.40 8.2272 h 9.99 9.98 10.05144 h 8.95 7.96 8.91SEM 0.359 0.369 0.681
Drip loss (%) 0.07272 h 2.03 a 1.86 a,b 1.19 b - -SEM 0.1783 0.1763 0.3197
Shear force (kg) 0.479 b.0001 0.61524 h 4.59 4.90 5.0772 h 3.04 3.22 3.08144 h 2.86 2.90 2.67SEM 0.372 0.373 0.448
Cooking loss (%) 0.940 b.0001 0.63224 h 21.33 23.65 22.1872 h 11.24 11.75 11.06144 h 17.84 15.82 16.30SEM 1.354 1.393 2.569
a, b Within rows, mean values without a common superscript differ (P≤0.05).x, y, z, w For the muscle pH values only, within columns mean values without a commonsuperscript differ (P≤0.01).
Fig. 1. Native and autolyzed μ-calpain time course activity in different CASTg.76,872 GNA genotypes. Within time post-mortem, A and B letters indicatesignificantly different values (Pb0.01).
697G. Gandolfi et al. / Meat Science 88 (2011) 694–700
at 24 h and72 h comparedwithpigs carryingGAandGGgenotypes. TheCAST genotype had significant effect on the time course of the autolyzedμ-calpain activity, as demonstrated by significant interaction betweengenotype and time post-mortem (P=0.012, Table 2). In Fig. 1 is shownthe time course activity of native and autolyzed μ-calpain for the threeCASTgenotypes.Differently from theother genotypes, in AA samples theautolyzed μ-calpain activity decreased from 1 h to 6 h, and showed apronounced increase from6 h to24 h,while thenative μ-calpain activityfollowed the opposite trend (Fig. 1). Autolysis of μ-calpain reduces theCa2+ requirement for activity from approximately 3–50 μM to 0.5–2.0 μM, values closer to physiological range (Goll et al., 2003), and hasbeen associated with terminal activation in post-mortem muscle(Geesink & Koohmaraie, 1999). The decrease of the native μ-calpainactivity post-mortem, with the increase of the autolyzed μ-calpainactivity, ultimately leads to a progressive loss of calpain proteolyticactivity.
In the present study, the results indicated that the pigs carrying AAgenotype had a higher level of calpain activation in post-mortemmuscle, which resulted in higher amount of autolyzed μ-calpain after24 h. However, these results require confirmation in a larger sample,taking in account the relative low number of animals used in thisstudy.
Since calpastatin represents the specific endogenous inhibitor ofubiquitous calpains, CAST variants could affect μ-calpain activity.However, CAST genotype showed a significant effect on the activity ofthe autolyzed μ-calpain and not of the native form. This could be due toa different effect of the gene variant in the inhibition of the native andautolyzed forms by calpastatin. The inhibition of calpain by calpastatinis calcium mediated. Autolysis of μ-calpain reduces the Ca2+
requirement for calpastatin binding from 40 μM to 0.042 μM (Gollet al., 2003). If the CAST mutation affects the ability of calpastatin inbinding calpain, it is possible that the effect of genotypes on the calpainactivity varies dependently on the calcium concentration required forinhibition. However, calpain–calpastatin interaction dynamics are stillpoorly understood, and different physiological mechanisms canmodulate the gene variant effects. CAST genotypes showed noeffect on m-calpain activity. The Ca2+ requirement for the bindingof m-calpain by calpastatin range from 250–500 μM, far from thephysiological range. (Goll et al., 2003). Therefore, it is possible thatcalpastatin has marginal effects on the regulation of m-calpain, atleast in the native form. Autolysis of m-calpain lowers the Ca2+
requirement for calpastatin binding to 25 μM. However, autolyzedm-calpain activity was not detected in the present study.
Concerning meat quality traits, CAST g.76,872 GNA genotype hadsuggestive effect on drip loss, with lower drip loss in pigs carrying AAgenotype compared with GG genotype (Table 3). Moreover, despitethe effect of CAST genotype on pHwas not significant, pigs carrying AAgenotype had suggestive higher pH values than other genotypes atevery time post-mortem except at 72 h (Table 3). Also, the AAgenotype seemed to show a lower rate of pH decline, as differencesbetween pH from 1 to 6 h post-mortem resulted less significantwithin this genotypes than within the others (Table 3). The
Table 4Calpain activity and myofibril particle size at different times post-mortem for CAPN1genotypes and effect of CAPN1 genotype, time and genotype×time interaction.
Timepost-mortem
CAPN1 HM535412:g.157 TNC genotype(N)
P valueforCAPN1effect
P valuefor timeeffect
P value forCAPN1×time
TT (19) TC (11)
μ-calpain activity 0.934 b.0001 0.6411 h 1.07 1.066 h 1.02 0.9624 h 0.58 0.6072 h 0.29 0.35SEM 0.043 0.057
Autolyzed μ-calpain activity 0.971 b.0001 0.2051 h 0.03 0.016 h 0.04 0.0324 h 0.17 0.1472 h 0.09 0.15SEM 0.023 0.030
m-calpain activity 0.448 b.0001 0.3991 h 1.19 1.196 h 1.09 1.2524 h 1.01 0.9772 h 0.88 0.89SEM 0.054 0.071
Myofibril particle size (μm) 0.558 b.0001 0.05324 h 30.90 a 38.94 b
72 h 18.01 17.06144 h 15.09 13.10SEM 2.286 3.005
a,b Within rows, mean values without a common superscript differ (P≤0.05).
698 G. Gandolfi et al. / Meat Science 88 (2011) 694–700
differences among genotypes in pH and drip loss values found in thisstudy were limited, possibly due to the analyzed sample size, andthese results are worthwhile of further validation in a larger numberof animals.
The relation between pH values and drip loss has been wellestablished, with low ultimate pH values, or accelerated pH decline,resulting indecreasedwaterholding capacity of themeat (Huff-Lonergan& Lonergan, 2005). However, the relation between calpain activity, pHdecline anddrip loss are controversial. Calpain activitywas reported tobeinfluencedby early pHdecline,with fast pHdecline associatedwith fasteractivation of μ-calpain (Pomponio et al., 2010; Claeys, De Smet, Demeyer,Geers&Buys, 2001;Hwang&Thompson, 2001). On theother hand, it hasbeen suggested that higher calpain-mediated proteolysis, through thedegradation of specific myofibril structural protein, could lead to animproved water holding capacity (Huff-Lonergan & Lonergan, 2005). Ithas been proposed that the early activation of calpain caused by fast pHdrop can lead to earlier loss of activity, resulting in higher drip loss (Bee,Anderson, Lonergan & Huff-Lonergan, 2007). In accordance with thishypothesis, in thepresent studypigs carryingCASTAAgenotypes showedthe slowest pH decline till 24 h, an increased level of autolyzed μ-calpainactivity at 24 h and the lowest drip loss, and thus resulted in morefavourable meat quality traits.
Different CAST polymorphisms have been described associatedwithmeat quality traits such as shear force, cooking loss and juiciness,and with CAST expression level (Lindholm-Perry et al., 2009; Ciobanuet al., 2004). This is the first analysis on the effect of CAST SNP on timecourse calpain activity. The obtained results suggested that CAST genevariation has an effect on calpain activity; however this should beconfirmed in larger samples. It seems unlikely that the identified SNPwithin CAST intron 6 represents the causative mutation for the effecton calpain activity and meat quality. Previously reported SNP thatassociated with tenderness and CAST expression, have been proposedto be in linkage with a causative mutation (Lindholm-Perry et al.,2009), probably located within the block of heterozygosity includingthe 5′ region of the gene (Meyers & Beever, 2008). It is possible thatthe intronic SNP analyzed in this study is in linkage with a causativemutation that could affect gene expression, alternative splicing ordirectly the activity of calpastatin in calpain inhibition.
The further validation of the effect of the SNP analyzed, or theidentification of the causative mutation affecting calpain activity,could be useful to develop a DNA test for proteolytic activity in post-mortemmuscle. This could also providemarkers for the improvementof pork tenderization by marker-assisted selection (MAS).
4.3. Effects of CAPN1 genotypes on calpain activity and meat qualitytraits
Least square means of calpain activity and meat quality para-meters for each CAPN1 genotype and time post-mortem are reportedin Tables 4 and 5.
CAPN1 g.157TNC genotype showed no effect on calpain activity,but effect on myofibrillar particle size at 24 h post-mortemwas found(Table 4). Pigs carrying CAPN1 g.157TC genotype showed highervalues of surface mean diameter of myofibrils, indicating a lowerfragmentation rate, at least in the first 24 hours. These differencesdisappeared at 3 and 6 days post-mortem.
The SNP CAPN1 g.157TNC showed effect onmeat color, with highervalues of redness a* at 1 h, 3 h and 6 h post-mortem for the genotypeTC compared with the homozygote TT (Table 5). Considering the lackof effect on calpain activity, it is unlikely that the relationship betweengenotype and color depends on changes in proteolytic activity. Sincefresh meat color, and in particular redness, represents one of the mainfeatures affecting consumer acceptance (Norman, Berg, Heymann &Lorenzen, 2003), effect of this genetic marker on redness should beinvestigated further in larger samples.
4.4. Expression analysis of CAST, CAPN1 and CAPN3 genes
The expression of CAST, CAPN1 and CAPN3 genes was comparedbetween two groups of pigs with high and lowWarner–Bratzler shearforce at 6 days (144 h) post-mortem. The high and the low groupsconsisted of 4 and 6 pigs, respectively, and shear force mean and s.d.was 3.01±0.25 kg and 2.22±0.22 kg, respectively (Pb0.001).
Average CAST expression was 28.9% higher in muscle with highshear force (P=0.17, Fig. 2). The strong correlation betweencalpastatin activity and tenderness has been well described (Kempet al., 2010) but clear relationship between gene expression andtenderness has never been reported. CAST gene expression could beone factor regulating calpastatin activity, and further analysis at geneexpression level could provide information on tenderness in pork.
Average CAPN3 expression was 31.2% higher in muscle with highshear force (P=0.13, Fig. 2). Calpain 3 is not inhibited by calpastatin(Ono et al., 2004) and its role in meat tenderization has been matter ofdebate. In sheep, calpain 3 protein and mRNA level strongly correlatedwith tenderness (Ilian et al., 2004; 2001). Conversely, no associationwere detected between calpain 3 expression and 8 days shear force inporcine LDmuscle (Parr et al., 1999), and tenderization processes werenot affected in CAPN3 knock-out mice (Geesink et al., 2005). In thepresent study, the higher CAPN3 gene expression in less tendermusclesconfirmed the lack of a direct involvement of calpain 3 in meattenderization. In muscle wasting mechanisms in vivo, calpain 3 has theopposite expression pattern compared with the ubiquitous calpains,being under expressed in muscle depletion processes and in atrophicconditions (Bartoli & Richard, 2005). Also, inactivating mutations inhuman CAPN3 gene cause recessive limb girdle muscular dystrophytype 2 (Richard et al., 1995). These findings suggest that calpain 3 isnecessary for normal muscle homeostasis and function and exerts aprotective effect against atrophy, although its function is still largelyunknown (Bartoli & Richard, 2005). Recently, a novel CAPN3 structuralrole in the protein complex that regulates calcium release in skeletalmuscle has been reported (Kramerova, Kudryashova, Wu, Ottenheijm,Granzier & Spencer, 2008). Although the results obtained in this and in

Fig. 2. Longissimus dorsi CAST, CAPN1 and CAPN3 gene expression in groups of high andlow shear force at 144 h post-mortem.
Table 5Meat quality parameters at different times post-mortem for CAPN1 genotypes andeffect of CAPN1 genotype, time and genotype×time interaction.
Timepost-mortem
CAPN1 HM535412:g.157 TNC genotype(N)
P valueforCAPN1effect
P valuefortimeeffect
P value forCAPN1×time
TT (24) TC (16)
Muscle pH 0.719 b.0001 0.6621 h 6.41 6.323 h 6.04 5.996 h 5.78 5.8024 h 5.47 5.4772 h 5.36 5.40SEM 0.108 0.113
ColourLightness L* 0.108 b.0001 0.893
1 h 36.93 35.534.5 h 41.12 40.456 h 45.00 44.0224 h 53.20 50.6372 h 52.61 52.51144 h 54.40 53.45SEM 0.942 1.154
Redness a* 0.015 b.0001 0.3091 h 8.06 A 9.71 B
4.5 h 6.28 a 7.81 b
6 h 5.92 a 7.30 b
24 h 5.71 6.2172 h 6.01 6.16144 h 5.80 6.10SEM 1.055 1.093
Yellowness b* 0.407 b.0001 0.8021 h 5.58 6.254.5 h 5.86 6.016 h 6.94 6.7524 h 7.29 7.7472 h 9.89 10.14144 h 8.54 8.49SEM 0.311 0.381
Drip loss (%) 0.527 - -72 h 1.92 1.77SEM 0.1702 0.1914
Shear force (kg) 0.547 b.0001 0.57824 h 4.69 4.9272 h 3.09 3.17144 h 2.88 2.82SEM 0.366 0.376
Cooking loss (%) 0.505 b.0001 0.84124 h 22.68 22.0472 h 11.43 11.43144 h 17.40 15.88SEM 1.166 1.429
A, B Within rows, mean values without a common superscript differ (P≤0.01).a, b Within rows, mean values without a common superscript differ (P≤0.05).
699G. Gandolfi et al. / Meat Science 88 (2011) 694–700
previous studies excluded a direct effect of calpain 3 proteolytic activityon meat tenderization, unknown roles of calpain 3 in post-mortemmuscle could be investigated further.
5. Conclusions
This study represents the first investigation on the effect of porcineCAST and CAPN1 gene variants on post-mortem muscle calpainactivity. CAST EU137105:g.76,872GNA variant showed an effect ontime course activity of autolyzed calpain, and on drip loss, whileCAPN1 HM535412:g.157TNC showed effect on meat redness. Effectsof the proposed markers could provide information on the geneticcontrol of meat quality, after confirmation in a larger number ofanimals.
Acknowledgements
The authors thank the Italian pig farmer organization OPAS(Mantova, Italy).
References
Barendse, W., Harrison, B. E., Bunch, R. J., & Thomas, M. B. (2008). Variation at theCalpain 3 gene is associated with meat tenderness in zebu and composite breeds ofcattle. BMC Genetics, 9, 41.
Bartoli, M., & Richard, I. (2005). Calpains in muscle wasting. The International Journal ofBiochemistry & Cell Biology, 37(10), 2115−2133.
Bee, G., Anderson, A. L., Lonergan, S. M., & Huff-Lonergan, E. (2007). Rate and extent ofpH decline affect proteolysis of cytoskeletal proteins and water-holding capacity inpork. Meat Science, 76(2), 359−365.
Briley, G. P., Riggs, P. K., Womack, J. E., Hancock, D. L., & Bidwell, C. A. (1996).Chromosomal localization of porcine skeletal muscle calpain gene. MammalianGenome, 7, 226−228.
Bustin, S. A. (2004). A-Z of Quantitative PCR. La Jolla: International University Line Press.Camou, J. P., Marchello, J. A., Thompson, V. F., Mares, S. W., & Goll, D. E. (2007). Effect of
postmortem storage on activity of mu- and m-calpain in five bovine muscles.Journal of Animal Science, 85, 2670−2681.
Ciobanu, D. C., Bastiaansen, J.W., Lonergan, S.M., Thomsen, H., Dekkers, J. C., Plastow, G. S.,et al. (2004). New alleles in calpastatin gene are associated withmeat quality traits inpigs. Journal of Animal Science, 82, 2829−2839.
Claeys, E., De Smet, S., Demeyer, D., Geers, R., & Buys, N. (2001). Effect of rate of pHdecline on muscle enzyme activities in two pig lines. Meat Science, 57, 257−263.
Cong, J. Y., Goll, D. E., Peterson, A. M., & Kapprell, H. P. (1989). The role of autolysis inactivity of the Ca2+-dependent proteinases (μ-Calpain and m-Calpain). The Journalof Biological Chemistry, 264(17), 10096−10103.
Ernst, C. W., Robic, A., Yerle, M., Wang, L., & Rothschild, M. F. (1998). Mapping ofcalpastatin and three microsatellites to porcine chromosome 2q2.1–q2.4. AnimalGenetics, 29, 212−215.
Fridolfsson, A. K., Hori, T., Wintero, A. K., Fredholm, M., Yerle, M., Robic, A., et al. (1997).Expansion of the pig comparative map by expressed sequence tags (EST) mapping.Mammalian Genome, 8, 907−912.
Geesink, G. H., & Koohmaraie, M. (1999). Effect of calpastatin on degradation ofmyofibrillar proteins by mu-calpain under postmortem conditions. Journal ofAnimal Science, 77, 2685−2692.
Geesink, G. H., Kuchay, S., Chishti, A. H., & Koohmaraie, M. (2006). Micro-calpain isessential for postmortem proteolysis of muscle proteins. Journal of Animal Science,84, 2834−2840.
Geesink, G. H., Taylor, R. G., & Koohmaraie, M. (2005). Calpain 3/p94 is not involved inpostmortem proteolysis. Journal of Animal Science, 83(7), 1646−1652.
Goll, D. E., Thompson, V. F., Li, H., Wei, W., & Cong, J. (2003). The calpain system.Physiological Reviews, 83, 731−801.
Honikel, K. O. (1998). Reference methods for the assessment of physical characteristicsof meat. Meat Science, 49(4), 447−457.
Huff-Lonergan, E., & Lonergan, S. M. (2005). Mechanisms of water-holding capacity ofmeat: The role of postmortem biochemical and structural changes. Meat Science,71(1), 194−204.
Hwang, H., & Thompson, J. M. (2001). The interaction between pH and temperaturedecline early postmortem on the calpain system and objective tenderness inelectrically stimulated beef longissimus dorsi muscle. Meat Science, 58(2),167−174.
Ilian, M. A., Bekhit, A. E. D., & Bickerstaffe, R. (2004). The relationship between meattenderization, myofibril fragmentation and autolysis of calpain 3 during postmortemaging.Meat Science, 66, 387−397.
Ilian, M. A., Morton, J. D., Kent, M. P., Le Couteur, C. E., Hickford, J., Cowley, R., et al.(2001). Intermuscular variation in tenderness: Association with the ubiquitous andmuscle-specific calpains. Journal of Animal Science, 79, 122−132.
Kemp, C. M., Sensky, P. L., Bardsley, R. G., Buttery, P. J., & Parr, T. (2010). Tenderness—Anenzymatic view. Meat Science, 84(22), 248−256.
Kent, M. P., Spencer, M. J., & Koohmaraie, M. (2004). Postmortem proteolysis is reduced intransgenic mice overexpressing calpastatin. Journal of Animal Science, 82, 794−801.

700 G. Gandolfi et al. / Meat Science 88 (2011) 694–700
Koohmaraie, M. (1992). The role of Ca2+-dependent proteases (calpains) in postmortem proteolysis and meat tenderness. Biochimie, 74(3), 239−245.
Kramerova, I., Kudryashova, E., Wu, B., Ottenheijm, C., Granzier, H., & Spencer, M. J.(2008). Novel role of calpain-3 in the triad-associated protein complex regulatingcalcium release in skeletal muscle. Human Molecular Genetics, 17(21), 3271−3280.
Lindholm-Perry, A. K., Rohrer, G. A., Holl, J. W., Shackelford, S. D., Wheeler, T. L.,Koohmaraie, M., et al. (2009). Relationships among calpastatin single nucleotidepolymorphisms, calpastatin expression and tenderness in pork longissimus. AnimalGenetics, 40(5), 713−721.
Melody, J. L., Lonergan, S. M., Rowe, L. J., Huiatt, T. W., Mayes, M. S., & Huff-Lonergan, E.(2004). Early post-mortem biochemical factors influence tenderness and water-holding capacity of three porcine muscles. Journal of Animal Science, 82(4),1195−1205.
Meyers, S. N., & Beever, J. E. (2008). Investigating the genetic basis of pork tenderness:genomic analysis of porcine CAST. Animal Genetics, 39(5), 531−543.
Meyers, S. N., Rodriguez-Zas, S. L., & Beever, J. E. (2007). Fine-mapping of a QTLinfluencing pork tenderness on porcine chromosome 2. BMC Genetics, 8, 69.
Moeller, S. J., Miller, R. K., Edwards, K. K., Zerby, H. N., Logan, K. E., Aldredge, T. L., et al.(2010). Consumer perceptions of pork eating quality as affected by pork qualityattributes and end-point cooked temperature. Meat Science, 84, 14−22.
Norman, J. L., Berg, E. P., Heymann, H., & Lorenzen, C. L. (2003). Pork loin color relative tosensory and instrumental tenderness and consumer acceptance. Meat Science, 65,927−933.
Ono, Y., Kakinuma, K., Torii, F., Irie, A., Nakagawa, K., Labeit, S., et al. (2004). Possibleregulation of the conventional calpain system by skeletal muscle-specific calpain,p94/calpain 3. The Journal of Biological Chemistry, 23, 2761−2771.
Page, B. T., Casas, E., Heaton, M. P., Cullen, N. G., Hyndman, D. L., Morris, C. A., et al.(2002). Evaluation of single-nucleotide polymorphisms in CAPN1 for associationwith meat tenderness in cattle. Journal of Animal Science, 80, 3077−3085.
Parr, T., Sensky, P. L., Scothern, G. P., Bardsley, R. G., Buttery, P. J., Wood, J. D., et al.(1999). Skeletal muscle-specific calpain and variable postmortem tenderization inporcine longissimus muscle. Journal of Animal Science, 77, 661−668.
Pomponio, L., Ertbjerg, P., Karlsson, A. H., Nanni Costa, L., & Lametsch, R. (2010).Influence of early pH decline on calpain activity in porcine muscle. Meat Science,85(1), 110−114.
Pomponio, L., Lametsch, R., Karlsson, A. H., Nanni Costa, L., Grossi, A., & Ertbjerg, P.(2008). Evidence for post-mortem m-calpain autolysis in porcine muscle. MeatScience, 80(3), 761−764.
Richard, I., Broux, O., Allamand, V., Fougerousse, F., Chiannilkulchai, N., Bourg, N., et al.(1995). Mutations in the proteolytic enzyme calpain 3 cause limb-girdle musculardystrophy type 2A. Cell, 81(1), 27−40.
Sieczkowska, H., Zybert, A., Krze˛cio, E., Antosik, A., Koc´win-Podsiadła, M., Pierzchała,M., et al. (2010). The expression of genes PKM2 and CAST in the muscle tissue ofpigs differentiated by glycolytic potential and drip loss, with reference to thegenetic group. Meat Science, 84(1), 137−142.
Škrlep, M., Čandek-Potokar, M., Kavar, T., Žlender, B., Hortós, M., Gou, P., et al. (2010).Association of PRKAG3 and CAST genetic polymorphisms with traits of interest indry-cured ham production: Comparative study in France, Slovenia and Spain.Livestock Science, 128(1–3), 60−66.
Sorimachi, H., Imajoh-Ohmi, S., Emori, Y., Kawasaki, H., Ohno, S., Minami, Y., et al.(1989). Molecular cloning of a novel mammalian calcium-dependent proteasedistinct from both m- and mu-types. Specific expression of the mRNA in skeletalmuscle. The Journal of Biological Chemistry, 264, 20106−20111.
Stearns, T. M., Beever, J. E., Southey, B. R., Ellis, M., McKeith, F. K., & Rodriguez-Zas, S. L.(2005). Evaluation of approaches to detect quantitative trait loci for growth,carcass and meat quality on swine chromosomes 2, 6, 13 and 18. I. Univariateoutbred F2 and sib-pair analyses. Journal of Animal Science, 83, 1481−1493.
Vandesompele, J., De Preter, K., Pattyn, F., Poppe, B., Van Roy, N., De Paepe, A., et al.(2002). Accurate normalization of real-time quantitative RT-PCR data by geometricaveraging of multiple internal control genes.Genome Biology, 3 RESEARCH0034.
Veiseth, E., Shackelford, S. D., Wheeler, T. L., & Koohmaraie, M. (2001). Effect ofpostmortem storage on mu-calpain and m-calpain in ovine skeletal muscle. Journalof Animal Science, 79, 1502−1508.




















