
In vivo bicarbonate requirement for water oxidation by Photosystem II in the...
10
In vivo bicarbonate requirement for water oxidation by Photosystem II in the hypercarbonate-requiring cyanobacterium Arthrospira maxima Damian Carrieri, Gennady Ananyev, Tyler Brown, G. Charles Dismukes * Princeton University, Department of Chemistry and Princeton Environmental Institute, Princeton, NJ 08544, United States Received 20 April 2007; received in revised form 21 June 2007; accepted 26 June 2007 Available online 10 July 2007 Dedicated in fond memory of Edward I. Stiefel Abstract While the presence of inorganic carbon in the form of (bi)carbonate has been known to be important for activity of Photosystem II (PSII), the vast majority of studies on this ‘‘bicarbonate effect’’ have been limited to in vitro studies of isolated thylakoid membranes and PSII complexes. Here we report an in vivo requirement for bicarbonate that is both reversible and selective for this anion for efficient water oxidation activity in the hypercarbonate-requiring cyanobacterium Arthrospira (Spirulina) maxima, originally isolated from highly alkaline soda lakes. Using a non-invasive internal probe of PSII charge separation (variable fluorescence), primary electron acceptor (Q A =Q A ) reoxidation rate, and flash-induced oxygen yield, we report the largest reversible bicarbonate effect on PSII activity ever observed, which is due to the requirement for bicarbonate at the water-oxidizing complex. Temporal separation of this donor side bicar- bonate requirement from a smaller effect of bicarbonate on the Q A reoxidation rate was observed. We expect the atypical way in which Arthrospira manages intracellular pH, sodium, and inorganic carbon concentrations relative to other cyanobacteria is responsible for this strong in vivo bicarbonate requirement. Ó 2007 Published by Elsevier Inc. keywords: Arthrospira (Spirulina); Oxygenic photosynthesis; Bicarbonate; Photosystem II; Cyanobacteria 1. Introduction It was with a heavy heart that Princeton said goodbye to Ed Stiefel with his untimely passing last year. An academic advisor and mentor to the first author and to each of us a respected scientist and friend, Ed was an enthusiastic educa- tor. Discussing matters of science (and practically every- thing else) with him was a real joy. Ed’s encyclopedic knowledge of scientific information enabled him to make insightful connections between diverse subdisciplines of sci- ence, particularly inorganic chemistry and biology. We proudly dedicate this manuscript to his memory. In the spirit of his educational style, we present unique measure- ments that provide compelling evidence for a direct role for inorganic carbon in the water oxidation reaction of pho- tosynthesis in vivo. We discuss the implications of this find- ing in the context of a broad base of equivocal literature. In addition to its obvious involvement in photosynthesis as a substrate for CO 2 fixation by the Calvin–Benson–Bas- sham cycle, the importance of inorganic carbon (Ci, abbre- viation for CO 2 , HCO 3 or CO 2 3 ), for optimal function of the Photosystem II (PSII)-dependent O 2 evolution reaction (water ! plastoquinone electron transfer) has been known for nearly 50 years. In 1958 Warburg and Krippahl showed that CO 2 accelerates O 2 evolution from isolated thylakoids illuminated in the presence of an artificial electron acceptor [1]. The following five decades of research have led to the conclusion that bicarbonate affects both the electron donor and acceptor sides of PSII. These bicarbonate effects have been reviewed extensively [2–6], yet nearly all current knowledge relies on experiments carried out in vitro on 0162-0134/$ - see front matter Ó 2007 Published by Elsevier Inc. doi:10.1016/j.jinorgbio.2007.06.039 * Corresponding author. Tel.: +609 258 3949; fax: +609 258 1980. E-mail address: [email protected] (G.C. Dismukes). www.elsevier.com/locate/jinorgbio Available online at www.sciencedirect.com Journal of Inorganic Biochemistry 101 (2007) 1865–1874 JOURNAL OF Inorganic Biochemistry
Transcript of In vivo bicarbonate requirement for water oxidation by Photosystem II in the...

Available online at www.sciencedirect.comJOURNAL OF
www.elsevier.com/locate/jinorgbio
Journal of Inorganic Biochemistry 101 (2007) 1865–1874
InorganicBiochemistry
In vivo bicarbonate requirement for water oxidation by PhotosystemII in the hypercarbonate-requiring cyanobacterium Arthrospira maxima
Damian Carrieri, Gennady Ananyev, Tyler Brown, G. Charles Dismukes *
Princeton University, Department of Chemistry and Princeton Environmental Institute, Princeton, NJ 08544, United States
Received 20 April 2007; received in revised form 21 June 2007; accepted 26 June 2007Available online 10 July 2007
Dedicated in fond memory of Edward I. Stiefel
Abstract
While the presence of inorganic carbon in the form of (bi)carbonate has been known to be important for activity of Photosystem II(PSII), the vast majority of studies on this ‘‘bicarbonate effect’’ have been limited to in vitro studies of isolated thylakoid membranes andPSII complexes. Here we report an in vivo requirement for bicarbonate that is both reversible and selective for this anion for efficientwater oxidation activity in the hypercarbonate-requiring cyanobacterium Arthrospira (Spirulina) maxima, originally isolated from highlyalkaline soda lakes. Using a non-invasive internal probe of PSII charge separation (variable fluorescence), primary electron acceptor(Q�A=QA) reoxidation rate, and flash-induced oxygen yield, we report the largest reversible bicarbonate effect on PSII activity everobserved, which is due to the requirement for bicarbonate at the water-oxidizing complex. Temporal separation of this donor side bicar-bonate requirement from a smaller effect of bicarbonate on the Q�A reoxidation rate was observed. We expect the atypical way in whichArthrospira manages intracellular pH, sodium, and inorganic carbon concentrations relative to other cyanobacteria is responsible for thisstrong in vivo bicarbonate requirement.� 2007 Published by Elsevier Inc.
keywords: Arthrospira (Spirulina); Oxygenic photosynthesis; Bicarbonate; Photosystem II; Cyanobacteria
1. Introduction
It was with a heavy heart that Princeton said goodbye toEd Stiefel with his untimely passing last year. An academicadvisor and mentor to the first author and to each of us arespected scientist and friend, Ed was an enthusiastic educa-tor. Discussing matters of science (and practically every-thing else) with him was a real joy. Ed’s encyclopedicknowledge of scientific information enabled him to makeinsightful connections between diverse subdisciplines of sci-ence, particularly inorganic chemistry and biology. Weproudly dedicate this manuscript to his memory. In thespirit of his educational style, we present unique measure-ments that provide compelling evidence for a direct role
0162-0134/$ - see front matter � 2007 Published by Elsevier Inc.
doi:10.1016/j.jinorgbio.2007.06.039
* Corresponding author. Tel.: +609 258 3949; fax: +609 258 1980.E-mail address: [email protected] (G.C. Dismukes).
for inorganic carbon in the water oxidation reaction of pho-tosynthesis in vivo. We discuss the implications of this find-ing in the context of a broad base of equivocal literature.
In addition to its obvious involvement in photosynthesisas a substrate for CO2 fixation by the Calvin–Benson–Bas-sham cycle, the importance of inorganic carbon (Ci, abbre-viation for CO2, HCO�3 or CO2�
3 ), for optimal function ofthe Photosystem II (PSII)-dependent O2 evolution reaction(water! plastoquinone electron transfer) has been knownfor nearly 50 years. In 1958 Warburg and Krippahl showedthat CO2 accelerates O2 evolution from isolated thylakoidsilluminated in the presence of an artificial electron acceptor[1]. The following five decades of research have led to theconclusion that bicarbonate affects both the electron donorand acceptor sides of PSII. These bicarbonate effects havebeen reviewed extensively [2–6], yet nearly all currentknowledge relies on experiments carried out in vitro on

Scheme 1. Sites of bicarbonate involvement in photosynthetic electrontransport and damaging alternative electron transport routes that competewith carbon fixation.
1866 D. Carrieri et al. / Journal of Inorganic Biochemistry 101 (2007) 1865–1874
isolated thylakoid membranes or PSII enriched particles[4], with few exceptions [7–13]. This choice was driven bythe need to separate the specific effects of Ci within PSIIfrom the above-mentioned role of CO2 as a substrate forphotosynthesis and because many phototrophs have CO2
concentrating mechanisms that allow for storage of intra-cellular Ci.
Our research goals in oxygenic photosynthesis andbiological fuel production has motivated our interest incyanobacteria from diverse environments, including the fil-amentous nonheterocystous Arthrospira (Spirulina) max-
ima. This hypercarbonate-requiring cyanobacterium wasisolated from an alkaline soda lake in Chad, Africa, whichhouses extremely high concentrations of (bi)carbonate(0.2–1.2 M) at strongly alkaline pH (10–11). Previouslyour lab reported that A. maxima exhibits the fastest wateroxidation rate (by 3–5 fold) ever observed thus far in vivo
[14]. Herein we show that this cyanobacterium has a muchstronger requirement for bicarbonate to achieve optimalPSII turnover than is observed in other cyanobacteria,including the widely studied freshwater species Synechocys-
tis sp. PCC 6803 [15] (herein referred to as Synechocystis6803), and even isolated PSII complexes from higher plants[6]. We present three lines of evidence using intact cell fila-ments (denoted in vivo hereafter) that enable discriminationbetween donor side vs acceptor side electron transfer reac-tions of PSII: (1) PSII quantum efficiency monitored by theyield of variable fluorescence from chlorophyll, (2) the pri-mary electron acceptor Q�A reoxidation rate, and (3) the O2
evolution rate. Comparison of Ci replete and Ci depletedcells reveals a specific requirement for bicarbonate at thewater-oxidizing complex of A. maxima. Moreover, themuch lower sensitivity to bicarbonate removal in the fresh-water cyanobacterium Synechocystis 6803 is attributable atleast in part to a different system for transporting bicarbon-ate across the cytoplasmic membrane.
2. Experimental
2.1. Cell culturing
Whole cells of Arthrospira (Spirulina) maxima (CS-328)obtained from the Tasmanian CSIRO Collection of LivingMicroalgae were grown photoautrophically in completeZarrouk’s media [16], pH 9.5, sometimes with additionalNaHCO3 (final concentration 200–400 mM) under contin-uous illumination (40 lE m�2 s�1) to a cell density of about1 g dry weight L�1. Aliquots of growing cultures wereeither used for measurements (as control), or harvestedby centrifugation and resuspended in an equal volume ofa nearly isotonic buffer solution varying in componentsof interest before measuring.
2.2. Quantum efficiency
Chlorophyll-a (Chl a) variable fluorescence yield, whichis interpreted as a direct measure of PSII quantum efficiency
[17], was measured with a home-built laser-based fast repe-tition rate fluorometer (FRRF) described elsewhere [14].Briefly, a train of 30 laser pulses (wavelength 655 nm) each1 ls in duration and spaced 1 ls apart applied to a solutionof intact filaments of the cyanobacterium induces the fluo-rescence yield from Chl a to rise from a minimum dark-adapted level (Fo) to a saturated level (Fm) and is thereforecalled a single turnover flash (STF). These fluorescenceyields correspond to open and closed PSII centers, respec-tively. For measurements herein, these trains were repeatedat a frequency of 20 Hz (50 ms dark interval between eachSTF) for 50 turnovers. The difference in Fm and Fo givesthe variable fluorescence (Fv) of the sample, and the ratioFv/Fm is a measure of PSII quantum efficiency. Samplingof Fv/Fm behavior for 50 STFs is repeated every 2 min,leaving a 117.5 second dark window after each measure-ment to allow samples to return to a dark-adapted state.The average of 60–3000 recordings (over a 2–10 h period),depending on the need for time resolution, gives a precisequantum efficiency of PSII turnover. Previously we demon-strated that the kinetics of variable fluorescence yieldobserved with the FRRF corresponds directly with theevolved oxygen yield measured with a modified Clark-typeelectrode if illuminated by the same STF protocol [14], sup-porting the claim that Fv/Fm measurements are an accurateand reliable way to measure PSII turnover efficiency.
2.3. QA reoxidation kinetics
The primary electron acceptor after the water-oxidizingcomplex at PSII is plastoquinone, QA (Scheme 1), and thedecay of Fm fluorescence following a saturating light pulsemonitors reoxidation of Q�A (e.g., to QB and the PQ pool).The kinetics of reoxidation of Q�A were measured by prob-ing fluorescence signals following a long (2 ms) actinic flashthat reduces essentially all QA centers. Shorter laser pulses(0.75 ls), which are not sufficiently strong to further reducemost QA centers in the sample, were supplied at a samplingfrequency of 20 kHz and the resulting fluorescence signalwas recorded for 8 ms following the actinic flash. Theresulting decrease in yield from the saturated Fm fluores-cence level after actinic excitation to the lower fluorescencesignal observed at 8 ms provides a measure of the Q�A reox-
D. Carrieri et al. / Journal of Inorganic Biochemistry 101 (2007) 1865–1874 1867
idation efficiency in our work (Table 2). The kinetics of Q�Areoxidation in the 8 ms window are well described by twolifetime constants (s1 and s2), found by fitting the fluores-cence transients to a bi-exponential decay function. Minorlifetime components in the decay of Q�A that are longer than8 ms were not characterized further. Sampling of Q�A reox-idation is repeated at 1 second intervals, leaving a 990 msdark window after each measurement to allow samples toreturn to a dark-adapted state. The average of 7200 record-ings (over a 2 h period) gives a precise data set for findingQ�A reoxidation efficiency and lifetime.
2.4. Oxygen evolution
For oxygen evolution experiments, samples were illumi-nated with 30 ms light pulses at an intensity of 80 mW m�2
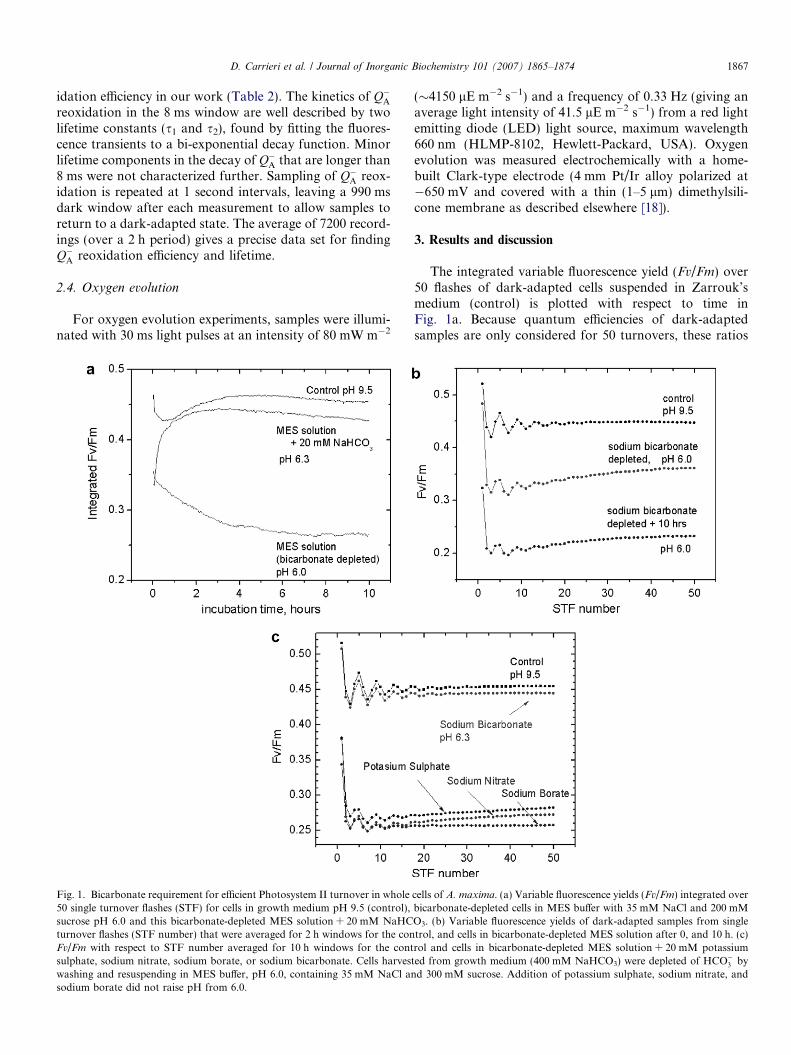
Fig. 1. Bicarbonate requirement for efficient Photosystem II turnover in whole50 single turnover flashes (STF) for cells in growth medium pH 9.5 (control),sucrose pH 6.0 and this bicarbonate-depleted MES solution + 20 mM NaHCturnover flashes (STF number) that were averaged for 2 h windows for the conFv/Fm with respect to STF number averaged for 10 h windows for the contsulphate, sodium nitrate, sodium borate, or sodium bicarbonate. Cells harveswashing and resuspending in MES buffer, pH 6.0, containing 35 mM NaCl ansodium borate did not raise pH from 6.0.
(�4150 lE m�2 s�1) and a frequency of 0.33 Hz (giving anaverage light intensity of 41.5 lE m�2 s�1) from a red lightemitting diode (LED) light source, maximum wavelength660 nm (HLMP-8102, Hewlett-Packard, USA). Oxygenevolution was measured electrochemically with a home-built Clark-type electrode (4 mm Pt/Ir alloy polarized at�650 mV and covered with a thin (1–5 lm) dimethylsili-cone membrane as described elsewhere [18]).
3. Results and discussion
The integrated variable fluorescence yield (Fv/Fm) over50 flashes of dark-adapted cells suspended in Zarrouk’smedium (control) is plotted with respect to time inFig. 1a. Because quantum efficiencies of dark-adaptedsamples are only considered for 50 turnovers, these ratios
cells of A. maxima. (a) Variable fluorescence yields (Fv/Fm) integrated overbicarbonate-depleted cells in MES buffer with 35 mM NaCl and 200 mMO3. (b) Variable fluorescence yields of dark-adapted samples from singletrol, and cells in bicarbonate-depleted MES solution after 0, and 10 h. (c)rol and cells in bicarbonate-depleted MES solution + 20 mM potassiumted from growth medium (400 mM NaHCO3) were depleted of HCO�3 by
d 300 mM sucrose. Addition of potassium sulphate, sodium nitrate, and

Table 1Miss (a), double Hit (b), goodness of fit (v2), and square residuals (R2) for extended Kok model fitted to variable fluorescence data with 95% confidenceintervals
Sample a b v2 R2
Control (Zarrouk medium) 0.0895 ± 0.006 0.0357 ± .007 3.1 · 10�6 0.947MES buffer (HCO�3 depleted) 0.175 ± .012 0.0330 ± .014 2.1 · 10�6 0.959MES buffer (HCO�3 depleted) + 10 h 0.183 ± .013 0.0390 ± .016 1.1 · 10�6 0.956MES buffer + 20 mM HCO�3 0.0853 ± 0.008 0.0316 ± .008 3.1 · 10�6 0.947
1868 D. Carrieri et al. / Journal of Inorganic Biochemistry 101 (2007) 1865–1874
represent efficiencies in harvesting photons to shuttle elec-trons from water to plastoquinones in the membrane (theso-called plastoquinone pool because they access a numberof PSII centers). These measurements are independent ofmore down-stream reactions in photosynthesis, those fol-lowing excitation by Photosystem I such as NADP+ reduc-tion and carbon fixation, which are only seen at a muchgreater number of turnovers. At these much higher turn-overs the fluorescence yield exhibit a complex inductioncurve, called the Kautsky induction curve, consisting of asequential series of distinct phases associated with thesedown-stream reactions [14,19].
To partially remove bicarbonate from A. maxima cellsthey are washed three times and resuspended in 50 mMMES buffer, pH 6.0, with 35 mM NaCl and 300 mMsucrose. The pKa of HCO�3 is 6.35, and the acidic condi-tions drive the dehydration of HCO�3 to form gaseousCO2, which escapes solution. Assuming a temperature of30̊C and an atmospheric concentration of CO2 of380 ppm, an air saturated solution at pH 6.0 is expectedto equilibrate at only 4 lM HCO�3 . Cells in this solu-tion show an immediate 27% decrease in the integratedFv/Fm, followed by a slower loss to a ratio that is 53%below the control after about 6 h incubation, as shown inFig. 1a. Also, addition of 20 mM NaHCO3 to suspensionsof filaments newly resuspended in MES buffer induces fullrecovery of variable fluorescence within 1 h.
The variable fluorescence yield from single turnoverflashes (STF) that were averaged over a 2 h window areshown for filaments in Zarrouk’s medium (control) andthe bicarbonate-depleted medium (Fig. 1b). Data areshown for filaments immediately after bicarbonate deple-tion and following 10 h of dark incubation in bicarbon-ate-free buffer. As can be observed in the control, Fv/Fm
values oscillate with a periodicity of four and dampen toa steady state ratio of 0.45. The period-four oscillationsare well understood and reflect the partial quenching ofChl fluorescence by the water oxidation enzyme as it cyclesthrough the S-states in PSII centers that progressively losesynchronization [14,20]. A significant loss of the amplitudeof the period-four oscillations (Fig. 1b) is observed in thetwo hour window following bicarbonate removal, reflectingloss in the quantum efficiency for stable charge separationby PSII. This loss will be further discussed below.
To examine whether the decrease in Fv/Fm concomitantwith depletion of bicarbonate is specific to this anion, andto rule out an effect of Na+, fluorescence yield measure-
ments were compared on bicarbonate-depleted samplesreconstituted with either 20 mM potassium sulphate,20 mM sodium nitrate, 20 mM sodium borate or 20 mMsodium bicarbonate (Fig. 1c). Sodium bicarbonate additioncauses full recovery of both period 4 oscillations (Table 1)and quantum efficiency within two hours of incubation(Fig. 1b) with minimal alkalization of the buffer (pHincreases from 6.0 to 6.3). Oscillations and quantum effi-ciency are not recovered upon addition of 20 mM potas-sium sulphate, 20 mM sodium nitrate, nor 20 mMsodium borate, and addition of these salts does not signif-icantly raise the pH of the buffer. An alkalization 0.3 pHunits upon addition of sodium bicarbonate to the buffercannot explain the recovery in PSII activity, as alkalizationof the 50 mM MES buffer to this pH using 6 M NaOHsolution does not result in significant recovery of Fv/Fm
values equal to the control (data not shown). We thereforeattribute the decrease in quantum efficiency and attenua-tion of period-four oscillations to a reversible loss of bicar-bonate function within PSII. To our knowledge thisbicarbonate effect on Fv/Fm upon bicarbonate depletionis the largest observed thus far in any native oxygenicphototroph.
The fluorescence yield oscillations from measurementsaveraged over a 2 h window were corrected with a splineand fitted to the classic Kok model [20] using the analyticalsolutions described by Shinkarev [21] to obtain parametersthat describe the probability of dephasing of the oscilla-tions. Fits were made for four data sets: untreated cells inbicarbonate medium (control), cells depleted of bicarbon-ate after 0 h and after 10 h (Fig. 1b), and bicarbonatereconstituted to bicarbonate-depleted cells (Fig. 1c). Thesevalues are listed in Table 1 along with goodness of fit (v2)and squared residuals (R2) describing the fit of the modelto the experimental data. The values a and b give the prob-ability ratios for ‘‘misses’’ and ‘‘double hits’’, respectively.A ‘‘miss’’ event describes the supply of a light flash thatdoes not result in the advancement of the photosystem toa further oxidized state. A ‘‘double hit’’ is an advancementof the photosystem through two oxidations with a singleflash.
The removal of bicarbonate does not cause a significantchange in the double-hit parameter, while the miss proba-bility for cells measured over the two hour window imme-diately following bicarbonate depletion doubles, reflectinga 2-fold increase in unsuccessful single turnovers of PSIIversus the control. There is no significant change in a fol-
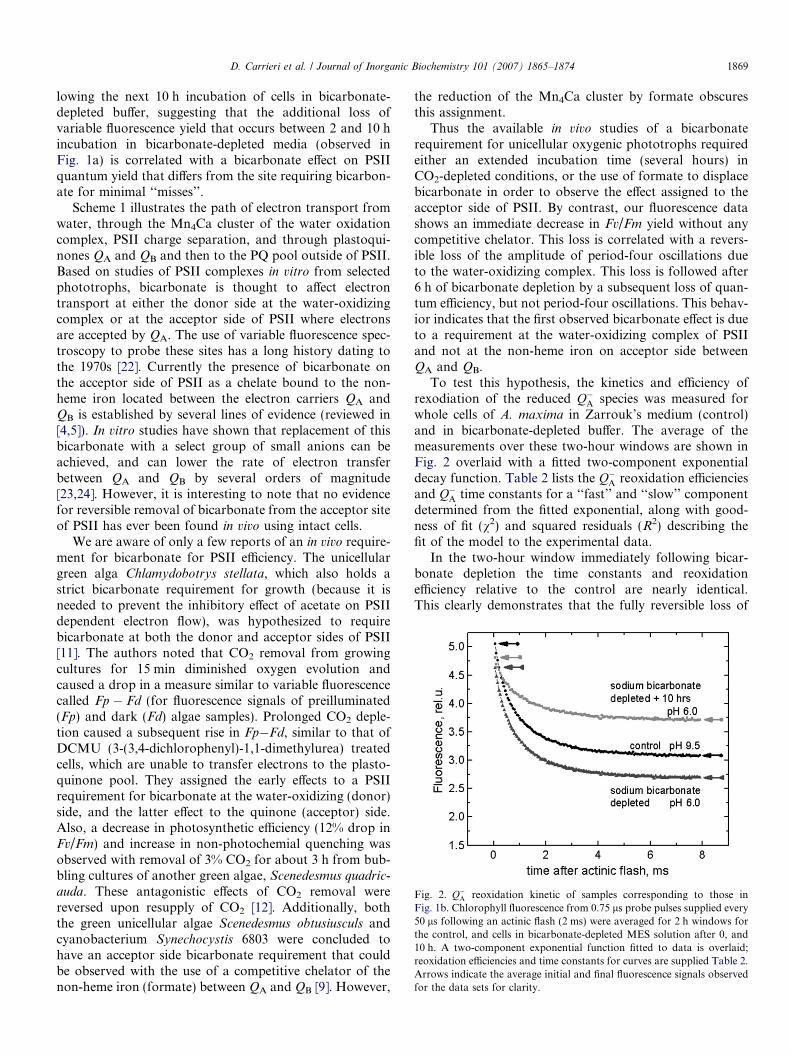
Fig. 2. Q�A reoxidation kinetic of samples corresponding to those inFig. 1b. Chlorophyll fluorescence from 0.75 ls probe pulses supplied every50 ls following an actinic flash (2 ms) were averaged for 2 h windows forthe control, and cells in bicarbonate-depleted MES solution after 0, and10 h. A two-component exponential function fitted to data is overlaid;reoxidation efficiencies and time constants for curves are supplied Table 2.Arrows indicate the average initial and final fluorescence signals observedfor the data sets for clarity.
D. Carrieri et al. / Journal of Inorganic Biochemistry 101 (2007) 1865–1874 1869
lowing the next 10 h incubation of cells in bicarbonate-depleted buffer, suggesting that the additional loss ofvariable fluorescence yield that occurs between 2 and 10 hincubation in bicarbonate-depleted media (observed inFig. 1a) is correlated with a bicarbonate effect on PSIIquantum yield that differs from the site requiring bicarbon-ate for minimal ‘‘misses’’.
Scheme 1 illustrates the path of electron transport fromwater, through the Mn4Ca cluster of the water oxidationcomplex, PSII charge separation, and through plastoqui-nones QA and QB and then to the PQ pool outside of PSII.Based on studies of PSII complexes in vitro from selectedphototrophs, bicarbonate is thought to affect electrontransport at either the donor side at the water-oxidizingcomplex or at the acceptor side of PSII where electronsare accepted by QA. The use of variable fluorescence spec-troscopy to probe these sites has a long history dating tothe 1970s [22]. Currently the presence of bicarbonate onthe acceptor side of PSII as a chelate bound to the non-heme iron located between the electron carriers QA andQB is established by several lines of evidence (reviewed in[4,5]). In vitro studies have shown that replacement of thisbicarbonate with a select group of small anions can beachieved, and can lower the rate of electron transferbetween QA and QB by several orders of magnitude[23,24]. However, it is interesting to note that no evidencefor reversible removal of bicarbonate from the acceptor siteof PSII has ever been found in vivo using intact cells.
We are aware of only a few reports of an in vivo require-ment for bicarbonate for PSII efficiency. The unicellulargreen alga Chlamydobotrys stellata, which also holds astrict bicarbonate requirement for growth (because it isneeded to prevent the inhibitory effect of acetate on PSIIdependent electron flow), was hypothesized to requirebicarbonate at both the donor and acceptor sides of PSII[11]. The authors noted that CO2 removal from growingcultures for 15 min diminished oxygen evolution andcaused a drop in a measure similar to variable fluorescencecalled Fp � Fd (for fluorescence signals of preilluminated(Fp) and dark (Fd) algae samples). Prolonged CO2 deple-tion caused a subsequent rise in Fp�Fd, similar to that ofDCMU (3-(3,4-dichlorophenyl)-1,1-dimethylurea) treatedcells, which are unable to transfer electrons to the plasto-quinone pool. They assigned the early effects to a PSIIrequirement for bicarbonate at the water-oxidizing (donor)side, and the latter effect to the quinone (acceptor) side.Also, a decrease in photosynthetic efficiency (12% drop inFv/Fm) and increase in non-photochemial quenching wasobserved with removal of 3% CO2 for about 3 h from bub-bling cultures of another green algae, Scenedesmus quadric-
auda. These antagonistic effects of CO2 removal werereversed upon resupply of CO2 [12]. Additionally, boththe green unicellular algae Scenedesmus obtusiusculs andcyanobacterium Synechocystis 6803 were concluded tohave an acceptor side bicarbonate requirement that couldbe observed with the use of a competitive chelator of thenon-heme iron (formate) between QA and QB [9]. However,
the reduction of the Mn4Ca cluster by formate obscuresthis assignment.
Thus the available in vivo studies of a bicarbonaterequirement for unicellular oxygenic phototrophs requiredeither an extended incubation time (several hours) inCO2-depleted conditions, or the use of formate to displacebicarbonate in order to observe the effect assigned to theacceptor side of PSII. By contrast, our fluorescence datashows an immediate decrease in Fv/Fm yield without anycompetitive chelator. This loss is correlated with a revers-ible loss of the amplitude of period-four oscillations dueto the water-oxidizing complex. This loss is followed after6 h of bicarbonate depletion by a subsequent loss of quan-tum efficiency, but not period-four oscillations. This behav-ior indicates that the first observed bicarbonate effect is dueto a requirement at the water-oxidizing complex of PSIIand not at the non-heme iron on acceptor side betweenQA and QB.
To test this hypothesis, the kinetics and efficiency ofrexodiation of the reduced Q�A species was measured forwhole cells of A. maxima in Zarrouk’s medium (control)and in bicarbonate-depleted buffer. The average of themeasurements over these two-hour windows are shown inFig. 2 overlaid with a fitted two-component exponentialdecay function. Table 2 lists the Q�A reoxidation efficienciesand Q�A time constants for a ‘‘fast’’ and ‘‘slow’’ componentdetermined from the fitted exponential, along with good-ness of fit (v2) and squared residuals (R2) describing thefit of the model to the experimental data.
In the two-hour window immediately following bicar-bonate depletion the time constants and reoxidationefficiency relative to the control are nearly identical.This clearly demonstrates that the fully reversible loss of
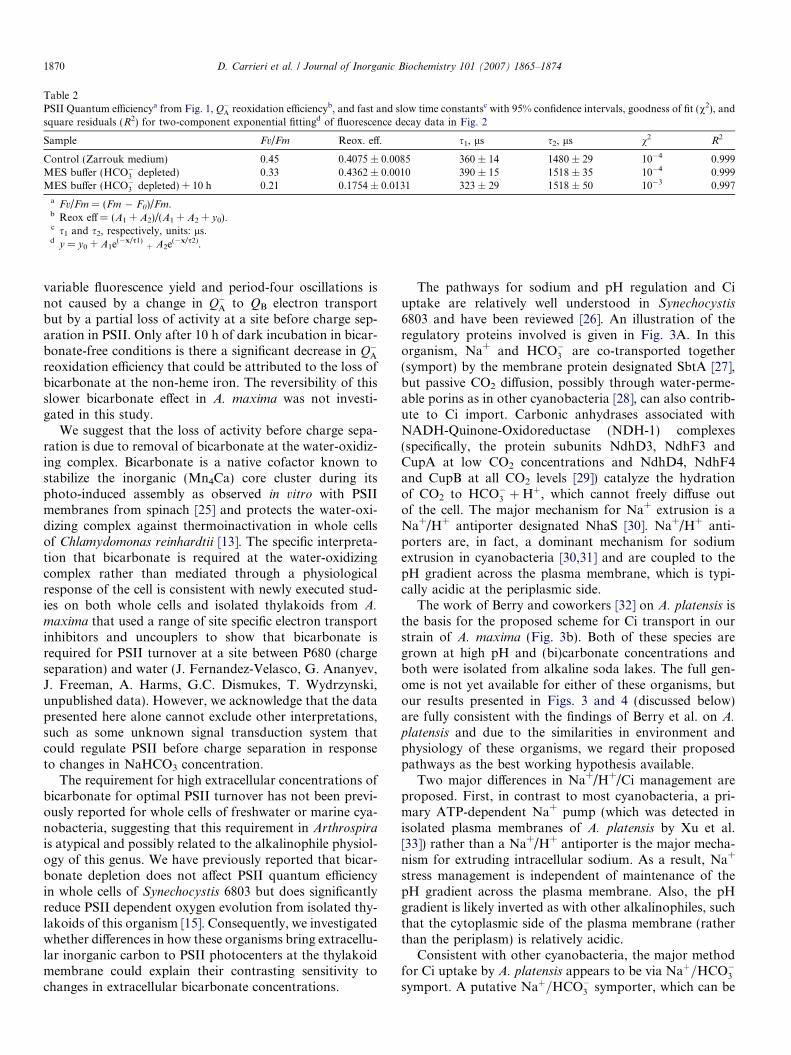
Table 2PSII Quantum efficiencya from Fig. 1, Q�A reoxidation efficiencyb, and fast and slow time constantsc with 95% confidence intervals, goodness of fit (v2), andsquare residuals (R2) for two-component exponential fittingd of fluorescence decay data in Fig. 2
Sample Fv/Fm Reox. eff. s1, ls s2, ls v2 R2
Control (Zarrouk medium) 0.45 0.4075 ± 0.0085 360 ± 14 1480 ± 29 10�4 0.999MES buffer (HCO�3 depleted) 0.33 0.4362 ± 0.0010 390 ± 15 1518 ± 35 10�4 0.999MES buffer (HCO�3 depleted) + 10 h 0.21 0.1754 ± 0.0131 323 ± 29 1518 ± 50 10�3 0.997
a Fv/Fm = (Fm � F0)/Fm.b Reox eff = (A1 + A2)/(A1 + A2 + y0).c s1 and s2, respectively, units: ls.d y = y0 + A1e(�x/s1)
+ A2e(�x/s2).
1870 D. Carrieri et al. / Journal of Inorganic Biochemistry 101 (2007) 1865–1874
variable fluorescence yield and period-four oscillations isnot caused by a change in Q�A to QB electron transportbut by a partial loss of activity at a site before charge sep-aration in PSII. Only after 10 h of dark incubation in bicar-bonate-free conditions is there a significant decrease in Q�Areoxidation efficiency that could be attributed to the loss ofbicarbonate at the non-heme iron. The reversibility of thisslower bicarbonate effect in A. maxima was not investi-gated in this study.
We suggest that the loss of activity before charge sepa-ration is due to removal of bicarbonate at the water-oxidiz-ing complex. Bicarbonate is a native cofactor known tostabilize the inorganic (Mn4Ca) core cluster during itsphoto-induced assembly as observed in vitro with PSIImembranes from spinach [25] and protects the water-oxi-dizing complex against thermoinactivation in whole cellsof Chlamydomonas reinhardtii [13]. The specific interpreta-tion that bicarbonate is required at the water-oxidizingcomplex rather than mediated through a physiologicalresponse of the cell is consistent with newly executed stud-ies on both whole cells and isolated thylakoids from A.
maxima that used a range of site specific electron transportinhibitors and uncouplers to show that bicarbonate isrequired for PSII turnover at a site between P680 (chargeseparation) and water (J. Fernandez-Velasco, G. Ananyev,J. Freeman, A. Harms, G.C. Dismukes, T. Wydrzynski,unpublished data). However, we acknowledge that the datapresented here alone cannot exclude other interpretations,such as some unknown signal transduction system thatcould regulate PSII before charge separation in responseto changes in NaHCO3 concentration.
The requirement for high extracellular concentrations ofbicarbonate for optimal PSII turnover has not been previ-ously reported for whole cells of freshwater or marine cya-nobacteria, suggesting that this requirement in Arthrospirais atypical and possibly related to the alkalinophile physiol-ogy of this genus. We have previously reported that bicar-bonate depletion does not affect PSII quantum efficiencyin whole cells of Synechocystis 6803 but does significantlyreduce PSII dependent oxygen evolution from isolated thy-lakoids of this organism [15]. Consequently, we investigatedwhether differences in how these organisms bring extracellu-lar inorganic carbon to PSII photocenters at the thylakoidmembrane could explain their contrasting sensitivity tochanges in extracellular bicarbonate concentrations.
The pathways for sodium and pH regulation and Ciuptake are relatively well understood in Synechocystis
6803 and have been reviewed [26]. An illustration of theregulatory proteins involved is given in Fig. 3A. In thisorganism, Na+ and HCO�3 are co-transported together(symport) by the membrane protein designated SbtA [27],but passive CO2 diffusion, possibly through water-perme-able porins as in other cyanobacteria [28], can also contrib-ute to Ci import. Carbonic anhydrases associated withNADH-Quinone-Oxidoreductase (NDH-1) complexes(specifically, the protein subunits NdhD3, NdhF3 andCupA at low CO2 concentrations and NdhD4, NdhF4and CupB at all CO2 levels [29]) catalyze the hydrationof CO2 to HCO�3 þHþ, which cannot freely diffuse outof the cell. The major mechanism for Na+ extrusion is aNa+/H+ antiporter designated NhaS [30]. Na+/H+ anti-porters are, in fact, a dominant mechanism for sodiumextrusion in cyanobacteria [30,31] and are coupled to thepH gradient across the plasma membrane, which is typi-cally acidic at the periplasmic side.
The work of Berry and coworkers [32] on A. platensis isthe basis for the proposed scheme for Ci transport in ourstrain of A. maxima (Fig. 3b). Both of these species aregrown at high pH and (bi)carbonate concentrations andboth were isolated from alkaline soda lakes. The full gen-ome is not yet available for either of these organisms, butour results presented in Figs. 3 and 4 (discussed below)are fully consistent with the findings of Berry et al. on A.
platensis and due to the similarities in environment andphysiology of these organisms, we regard their proposedpathways as the best working hypothesis available.
Two major differences in Na+/H+/Ci management areproposed. First, in contrast to most cyanobacteria, a pri-mary ATP-dependent Na+ pump (which was detected inisolated plasma membranes of A. platensis by Xu et al.[33]) rather than a Na+/H+ antiporter is the major mecha-nism for extruding intracellular sodium. As a result, Na+
stress management is independent of maintenance of thepH gradient across the plasma membrane. Also, the pHgradient is likely inverted as with other alkalinophiles, suchthat the cytoplasmic side of the plasma membrane (ratherthan the periplasm) is relatively acidic.
Consistent with other cyanobacteria, the major methodfor Ci uptake by A. platensis appears to be via Naþ=HCO�3symport. A putative Naþ=HCO�3 symporter, which can be
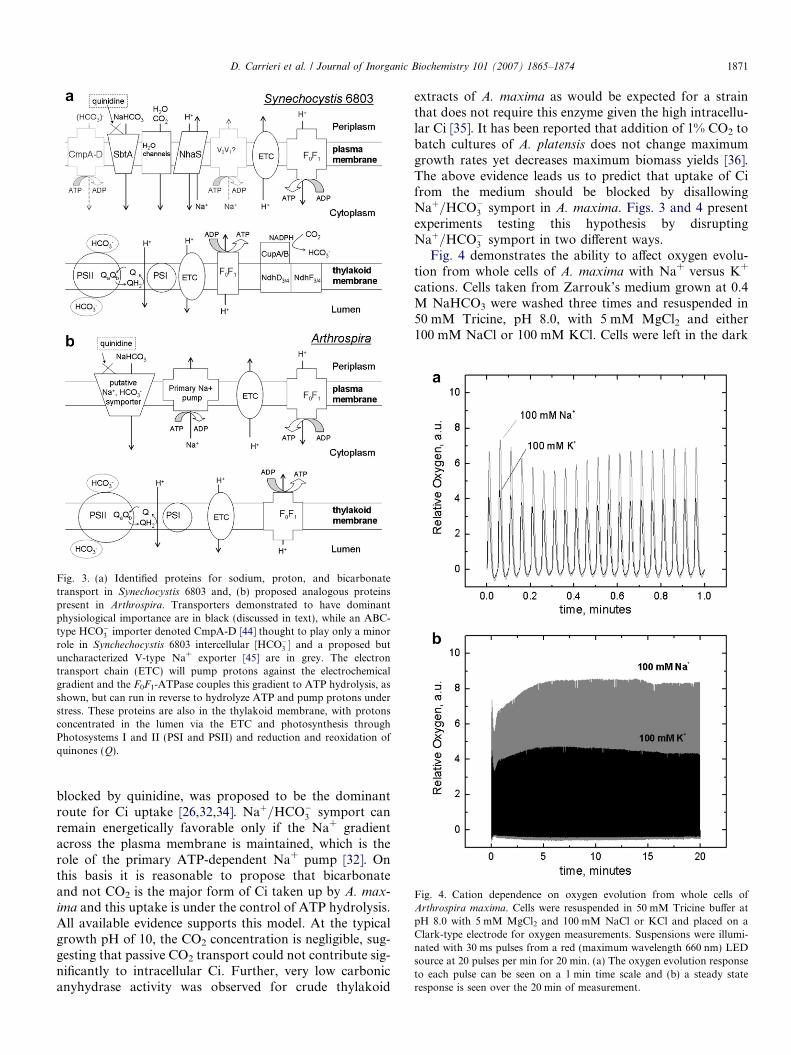
Fig. 3. (a) Identified proteins for sodium, proton, and bicarbonatetransport in Synechocystis 6803 and, (b) proposed analogous proteinspresent in Arthrospira. Transporters demonstrated to have dominantphysiological importance are in black (discussed in text), while an ABC-type HCO�3 importer denoted CmpA-D [44] thought to play only a minorrole in Synchechocystis 6803 intercellular [HCO�3 � and a proposed butuncharacterized V-type Na+ exporter [45] are in grey. The electrontransport chain (ETC) will pump protons against the electrochemicalgradient and the F0F1-ATPase couples this gradient to ATP hydrolysis, asshown, but can run in reverse to hydrolyze ATP and pump protons understress. These proteins are also in the thylakoid membrane, with protonsconcentrated in the lumen via the ETC and photosynthesis throughPhotosystems I and II (PSI and PSII) and reduction and reoxidation ofquinones (Q).
Fig. 4. Cation dependence on oxygen evolution from whole cells ofArthrospira maxima. Cells were resuspended in 50 mM Tricine buffer atpH 8.0 with 5 mM MgCl2 and 100 mM NaCl or KCl and placed on aClark-type electrode for oxygen measurements. Suspensions were illumi-nated with 30 ms pulses from a red (maximum wavelength 660 nm) LEDsource at 20 pulses per min for 20 min. (a) The oxygen evolution responseto each pulse can be seen on a 1 min time scale and (b) a steady stateresponse is seen over the 20 min of measurement.
D. Carrieri et al. / Journal of Inorganic Biochemistry 101 (2007) 1865–1874 1871
blocked by quinidine, was proposed to be the dominantroute for Ci uptake [26,32,34]. Naþ=HCO�3 symport canremain energetically favorable only if the Na+ gradientacross the plasma membrane is maintained, which is therole of the primary ATP-dependent Na+ pump [32]. Onthis basis it is reasonable to propose that bicarbonateand not CO2 is the major form of Ci taken up by A. max-ima and this uptake is under the control of ATP hydrolysis.All available evidence supports this model. At the typicalgrowth pH of 10, the CO2 concentration is negligible, sug-gesting that passive CO2 transport could not contribute sig-nificantly to intracellular Ci. Further, very low carbonicanyhydrase activity was observed for crude thylakoid
extracts of A. maxima as would be expected for a strainthat does not require this enzyme given the high intracellu-lar Ci [35]. It has been reported that addition of 1% CO2 tobatch cultures of A. platensis does not change maximumgrowth rates yet decreases maximum biomass yields [36].The above evidence leads us to predict that uptake of Cifrom the medium should be blocked by disallowingNaþ=HCO�3 symport in A. maxima. Figs. 3 and 4 presentexperiments testing this hypothesis by disruptingNaþ=HCO�3 symport in two different ways.
Fig. 4 demonstrates the ability to affect oxygen evolu-tion from whole cells of A. maxima with Na+ versus K+
cations. Cells taken from Zarrouk’s medium grown at 0.4M NaHCO3 were washed three times and resuspended in50 mM Tricine, pH 8.0, with 5 mM MgCl2 and either100 mM NaCl or 100 mM KCl. Cells were left in the dark
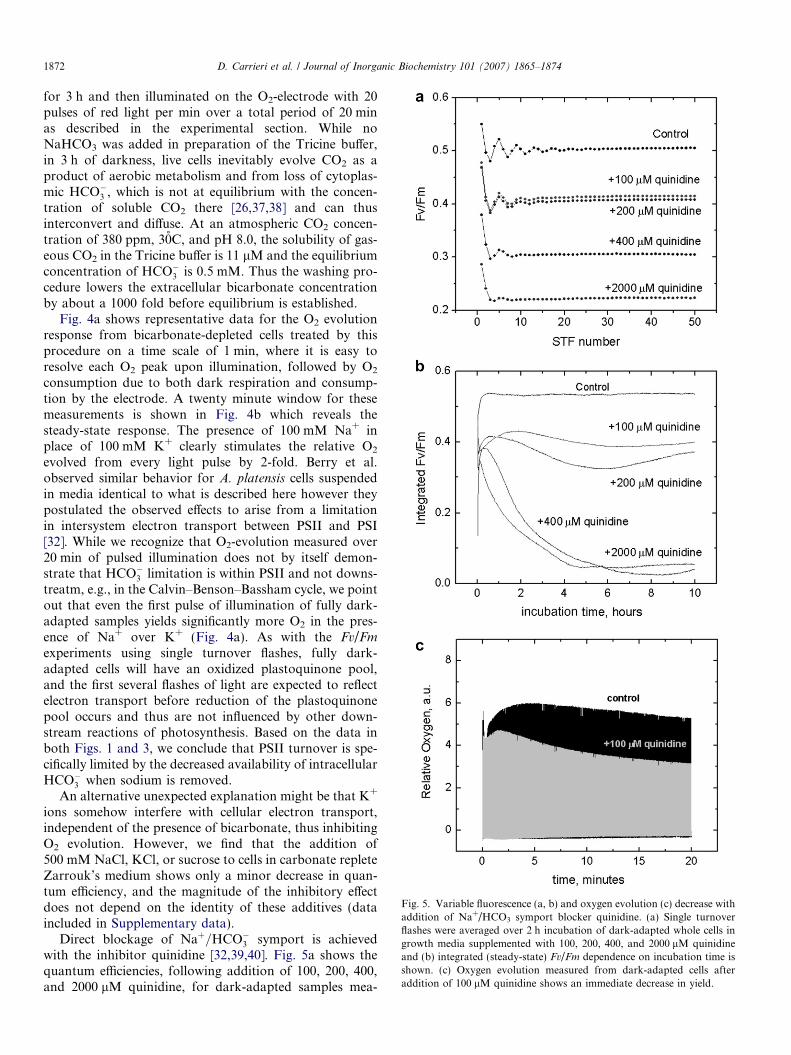
Fig. 5. Variable fluorescence (a, b) and oxygen evolution (c) decrease withaddition of Na+/HCO3 symport blocker quinidine. (a) Single turnoverflashes were averaged over 2 h incubation of dark-adapted whole cells ingrowth media supplemented with 100, 200, 400, and 2000 lM quinidineand (b) integrated (steady-state) Fv/Fm dependence on incubation time isshown. (c) Oxygen evolution measured from dark-adapted cells afteraddition of 100 lM quinidine shows an immediate decrease in yield.
1872 D. Carrieri et al. / Journal of Inorganic Biochemistry 101 (2007) 1865–1874
for 3 h and then illuminated on the O2-electrode with 20pulses of red light per min over a total period of 20 minas described in the experimental section. While noNaHCO3 was added in preparation of the Tricine buffer,in 3 h of darkness, live cells inevitably evolve CO2 as aproduct of aerobic metabolism and from loss of cytoplas-mic HCO�3 , which is not at equilibrium with the concen-tration of soluble CO2 there [26,37,38] and can thusinterconvert and diffuse. At an atmospheric CO2 concen-tration of 380 ppm, 30̊C, and pH 8.0, the solubility of gas-eous CO2 in the Tricine buffer is 11 lM and the equilibriumconcentration of HCO�3 is 0.5 mM. Thus the washing pro-cedure lowers the extracellular bicarbonate concentrationby about a 1000 fold before equilibrium is established.
Fig. 4a shows representative data for the O2 evolutionresponse from bicarbonate-depleted cells treated by thisprocedure on a time scale of 1 min, where it is easy toresolve each O2 peak upon illumination, followed by O2
consumption due to both dark respiration and consump-tion by the electrode. A twenty minute window for thesemeasurements is shown in Fig. 4b which reveals thesteady-state response. The presence of 100 mM Na+ inplace of 100 mM K+ clearly stimulates the relative O2
evolved from every light pulse by 2-fold. Berry et al.observed similar behavior for A. platensis cells suspendedin media identical to what is described here however theypostulated the observed effects to arise from a limitationin intersystem electron transport between PSII and PSI[32]. While we recognize that O2-evolution measured over20 min of pulsed illumination does not by itself demon-strate that HCO�3 limitation is within PSII and not downs-treatm, e.g., in the Calvin–Benson–Bassham cycle, we pointout that even the first pulse of illumination of fully dark-adapted samples yields significantly more O2 in the pres-ence of Na+ over K+ (Fig. 4a). As with the Fv/Fm
experiments using single turnover flashes, fully dark-adapted cells will have an oxidized plastoquinone pool,and the first several flashes of light are expected to reflectelectron transport before reduction of the plastoquinonepool occurs and thus are not influenced by other down-stream reactions of photosynthesis. Based on the data inboth Figs. 1 and 3, we conclude that PSII turnover is spe-cifically limited by the decreased availability of intracellularHCO�3 when sodium is removed.
An alternative unexpected explanation might be that K+
ions somehow interfere with cellular electron transport,independent of the presence of bicarbonate, thus inhibitingO2 evolution. However, we find that the addition of500 mM NaCl, KCl, or sucrose to cells in carbonate repleteZarrouk’s medium shows only a minor decrease in quan-tum efficiency, and the magnitude of the inhibitory effectdoes not depend on the identity of these additives (dataincluded in Supplementary data).
Direct blockage of Naþ=HCO�3 symport is achievedwith the inhibitor quinidine [32,39,40]. Fig. 5a shows thequantum efficiencies, following addition of 100, 200, 400,and 2000 lM quinidine, for dark-adapted samples mea-

D. Carrieri et al. / Journal of Inorganic Biochemistry 101 (2007) 1865–1874 1873
sured as in Fig. 1 using a series of 50 single turnover flasheswith averaging of STF trains over the first 2 h. Fig. 5b plotsthe steady state quantum efficiencies from these samples forup to 10 h after addition of the Naþ=HCO�3 symportblocker. Addition of 100 lM quinidine to cells in Zarrouk’smedium (control) shows a 20% decrease in quantum effi-ciency when averaged over a 2 h period (Fig. 5a) and asmall additional loss (total 26%) over the next 8 h(Fig. 5b). The oxygen evolution yield makes an immediatedrop over the first 20 min and produces a 20% decrease rel-ative to untreated control (Fig. 5c). The Kok parametersdescribing the damping of the period-four oscillations donot change as Fv/Fm decreases at 100 and 200 lM quini-dine. At concentrations of 400 lM quinidine and above afurther decrease in Fv/Fm occurs, and the onset of cell lysisis observed approximately 1 h after quinidine addition.This is correlated with a sharp loss in the amplitude ofthe period-four oscillations and change in their damping(Fig. 5a), and sharp decrease in integrated quantum effi-ciencies after 1 h (Fig. 5b). The immediate decrease of O2
evolution upon addition of quinidine is followed by asteady decline in evolution after several minutes of pulsedlight (Fig. 5c). Presumably, HCO�3 is consumed by the Cal-vin–Benson–Bassham cycle after about 2 min of pulsedlight, and intracellular HCO�3 is not replaced because ofthe presence of quinidine. Note that in Fig. 5b, averagequantum efficiencies decline only weakly beyond the initialloss observed upon addition of 100 and 200 lM quinidinebecause, in contrast to conditions for oxygen evolutionmeasurements, 50 STFs spaced by 2 min of dark incuba-tion are not sufficient to drive the Calvin–Benson–Basshamcycle that consumes intracellular HCO�3 .
Also, in contrast to the experimental conditions used forFig. 4, samples in Fig. 5 were not resuspended in a low-car-bonate buffer, but instead remained in growth media withhigh NaHCO3 concentrations. The effect of quinidine onFv/Fm values and initial differences in O2 evolution havetwo possible interpretations. Either cells experienced an ini-tial loss of intracellular HCO�3 by Calvin–Benson–Basshamactivity during dark-adaptation, which could be recoveredin the control but not in samples with quinidine, or alterna-tively, quinidine directly inhibits electron transport some-where between water oxidation and CO2 reduction. Whilewe favor the former explanation, Berry et al. noted thatfurther testing to exclude inhibitory action on photosyn-thetic complexes is needed when they reported that quini-dine simulated the inhibitory effect observed whensodium is replaced with potassium in buffered suspensionsof A. platensis [32].
We expect that bicarbonate’s role at the water-oxidizingcomplex in A. maxima is observable because Arthrospira’sonly major pathway for Ci uptake is through aNaþ=HCO�3 symport protein in the plasma membrane,which requires Na+ and not K+ for transport and is inde-pendent of the DpH across the plasma membrane because aDNa+ gradient is maintained by ATP-dependent Na+-extrusion [32,33].
The specific role of bicarbonate in A. maxima at thewater-oxidizing complex is unclear. The postulated roleof bicarbonate as a possible alternative substrate to waterhas been convincingly shown to not be the case in a fewselect cyanobacteria and isolated spinach PSII complexes[35,41], but other roles have been proposed. For example,mutagenesis of CP43-arginine-357 to serine in Synechocys-
tis 6803 suggests that Arginine R357 functions in binding a(bi)carbonate ion essential to normal catalytic turnover ofthe water-oxidizing complex [15], perhaps by involvementof hydrogen bonds with R357 which abstracts protonsfrom oxidized water molecules [42]. Alternatively, bicar-bonate may act as the endogenous base for protonsreleased into the lumen upon water oxidation [43].
There has been speculation in the literature that thebicarbonate effect on PSII charge separation seen previ-ously by in vitro experiments on isolated thylakoid mem-branes and PSII particles could indicate an importantphysiological role for bicarbonate in limiting overreductionof the PQ pool [2,12]. This results in damaging side reac-tions such as the Mehler reaction at PSI and increasedreduced ratios of soluble electron carriers via PQ-oxidore-ductases under low intracellular Ci levels when CO2 fixa-tion is slowed or shut down (Scheme 1). However, thispostulated mechanism of protection could not be verified,since until now, there were no reports demonstrating anappreciable and reversible bicarbonate effect on intact cellsor leafs (eg. in vivo) that would give support for a physio-logical role in PSII turnover. The present results thus pro-vide compelling support for this postulated but previouslyunestablished hypothesis for regulation of electron dona-tion in response to the reduced availability of the terminalelectron acceptor of photosynthesis (CO2). For high(bi)carbonate-requiring strains like A. maxima one canimagine a need for such regulation for withstanding acutedecreases in bicarbonate concentrations in shallow sodalakes resulting, for example, from heavy rains.
Acknowledgements
This work was supported by the Human Frontiers Sci-ence Program (RGP 29/2002) the National Institute ofHealth (GM-39932) the National Science Foundation(DBI-0138012) and the Air Force Office of Scientific Re-search (MURI Grant FA9550-05-1-0365) and is gratefullyacknowledged. The authors thank Derrick Kolling forinsightful advice and acknowledge Ed Stiefel’s otherPrinceton graduate students Madeli Castruita, MelissaCody, and Kelsey McNeely and research undergraduatesMariamawit Tamerat, Joey Contruvo, Abhinav Jha, andEddie Burgess.
Appendix A. Supplementary data
Supplementary data associated with this article can befound, in the online version, at doi:10.1016/j.jinorgbio.2007.06.039.

1874 D. Carrieri et al. / Journal of Inorganic Biochemistry 101 (2007) 1865–1874
References
[1] O. Warburg, G. Krippahl, Zeitschrift Fur Naturforschung Part B-Chemie Biochemie Biophysik Biologie Und Verwandten Gebiete 13(1958) 509–514.
[2] D.J. Blubaugh, Govindjee, Photosynth. Res. 19 (1988) 85–128.[3] A.J. Stemler, Photosynth. Res. 73 (2002) 177–183.[4] J.J.S. Van Rensen, Photosynth. Res. 73 (2002) 185–192.[5] J.J.S. Van Rensen, C.H. Xu, Govindjee, Physiol. Plant. 105 (1999)
585–592.[6] V.V. Klimov, S.V. Baranov, Bba-Bioenergetics 1503 (2001) 187–196.[7] F. Elshintinawy, Govindjee, Photosynth. Res. 24 (1990) 189–200.[8] G. Garab, Z. Rozsa, Govindjee, Naturwissenschaften 75 (1988) 517–
519.[9] Govindjee, J.F.H. Snel, O.J. Devos, J.J.S. Vanrensen, Physiol. Plant.
89 (1993) 143–148.[10] C.R. Ireland, N.R. Baker, S.P. Long, Biochim. Biophys. Acta 893
(1987) 434–443.[11] D. Mende, W. Wiessner, J. Plant Physiol. 118 (1985) 259–266.[12] J. Knoppova, J. Masojidek, J. Pokorny, Photosynthetica 28 (1992)
541–547.[13] S.K. Zharmukhamedov, G.N. Shirshikova, Z.V. Maevskaya, T.M.
Antropova, V.V. Klimov, Russ. J. Plant Phys. 54 (2007) 302–308.[14] G. Ananyev, G.C. Dismukes, Photosynth. Res. 84 (2005) 355–365.[15] G. Ananyev, T. Nguyen, C. Putnam-Evans, G.C. Dismukes, Photo-
chem. Photobiol. Sci. 4 (2005) 991–998.[16] C. Zarrouk, Contribution a l’etude d’une Cyanophycee. Influence de
divers facteurs physiques et chimiques sur la croissance et laphotosynthase de Spirulina maxima, Ph.D. thesis, University ofParis, Paris, 1966.
[17] Z.S. Kolber, O. Prasil, P.G. Falkowski, Bba-Bioenergetics 1367(1998) 88–106.
[18] G.M. Ananyev, G.C. Dismukes, Biochemistry-Us 35 (1996) 4102–4109.
[19] D. Lazar, Bba-Bioenergetics 1412 (1999) 1–28.[20] B. Kok, B. Forbush, M. Mcgloin, Photochem. Photobiol. 11 (1970)
457–475.[21] V.P. Shinkarev, Biophys. J. 85 (2003) 435–441.[22] T. Wydrzynski, Govindjee, Biochim. Biophys. Acta 387 (1975) 403–
408.[23] Y. Deligiannakis, V. Petrouleas, B.A. Diner, Bba-Bioenergetics 1188
(1994) 260–270.
[24] V. Petrouleas, Y. Deligiannakis, B.A. Diner, Bba-Bioenergetics 1188(1994) 271–277.
[25] S.V. Baranov, A.M. Tyryshkin, D. Katz, G.C. Dismukes, G.M.Ananyev, V.V. Klimov, Biochemistry-Us 43 (2004) 2070–2079.
[26] T. Ogawa, A. Kaplan, Photosynth. Res. 77 (2003) 105–115.[27] M. Shibata, H. Katoh, M. Sonoda, H. Ohkawa, M. Shimoyama, H.
Fukuzawa, A. Kaplan, T. Ogawa, J. Biol. Chem. 277 (2002) 18658–18664.
[28] D. Tchernov, Y. Helman, N. Keren, B. Luz, I. Ohad, L. Reinhold, T.Ogawa, A. Kaplan, J. Biol. Chem. 276 (2001) 23450–23455.
[29] M. Shibata, H. Ohkawa, T. Kaneko, H. Fukuzawa, S. Tabata, A.Kaplan, T. Ogawa, Proc. Natl. Acad. Sci. USA 98 (2001) 11789–11794.
[30] M. Inaba, A. Sakamoto, N. Murata, J. Bacteriol. 183 (2001) 1376–1384.
[31] R. Waditee, T. Hibino, T. Nakamura, A. Incharoensakdi, T. Takabe,Proc. Natl. Acad. Sci. USA 99 (2002) 4109–4114.
[32] S. Berry, Y.V. Bolychevtseva, M. Rogner, N.V. Karapetyan, Photo-synth. Res. 78 (2003) 67–76.
[33] C.H. Xu, A. Nejidat, S. Belkin, S. Boussiba, Plant Cell Physiol. 35(1994) 737–741.
[34] M. Shibata, H. Ohkawa, H. Katoh, M. Shimoyama, T. Ogawa,Funct. Plant Biol. 29 (2002) 123–129.
[35] W. Hillier, I. Mcconnell, M.R. Badger, A. Boussac, V.V. Klimov,G.C. Dismukes, T. Wydrzynski, Biochemistry-Us 45 (2006) 2094–2102.
[36] F.J.L. Gordillo, C. Jimenez, F.L. Figueroa, F.X. Niell, J. Appl.Phycol. 10 (1999) 461–469.
[37] G.D. Price, M.R. Badger, Plant Physiol. 91 (1989) 505–513.[38] L. Reinhold, R. Kosloff, A. Kaplan, Canadian Journal of Botany-
Revue Canadienne De Botanique 69 (1991) 984–988.[39] S.I. Allakhverdiev, A. Sakamoto, Y. Nishiyama, M. Inaba, N.
Murata, Plant Physiol. 123 (2000) 1047–1056.[40] S. Berry, B. Esper, I. Karandashova, M. Teuber, I. Elanskaya, M.
Rogner, M. Hagemann, FEBS Lett. 548 (2003) 53–58.[41] J. Clausen, K. Beckmann, W. Junge, J. Messinger, Plant Physiol. 139
(2005) 1444–1450.[42] J.P. Mcevoy, G.W. Brudvig, PCCP 6 (2004) 4754–4763.[43] A. Villarejo, T. Shutova, O. Moskvin, M. Forssen, V.V. Klimov, G.
Samuelsson, EMBO J. 21 (2002) 1930–1938.[44] T. Omata, G.D. Price, M.R. Badger, M. Okamura, S. Gohta, T.
Ogawa, Proc. Natl. Acad. Sci. USA 96 (1999) 13571–13576.[45] R.J. Ritchie, Can. J. Bot. 76 (1998) 1127–1145.




















