
Inadequate iron supply and high bicarbonate impair the symbiosis of peanuts ( Arachis hypogaea L.)...
10
Plant and Soil 138: 159-168, 1991 © 1991 Kluwer Academic Publishers. Printed in the Netherlands. PLSO 8948 Inadequate iron supply and high bicarbonate impair the symbiosis of peanuts (Arachis hypogaea L.) with different Bradyrhizobium strains CAIXIAN TANG 1'3, A.D. ROBSON l and M.J. DILWORTH 2 ~Soil Science and Plant Nutrition, School of Agriculture, The University of Western Australia, Nedlands, W.A. 6009, Australia, 2School of Biological and Environmental Sciences, Murdoch University, Murdoch, W.A. 6150, Australia and 3Soil Science and Agricultural Chemistry, Zhejiang Agricultural University, Hangzhou, China Received 26 October 1990. Revised July 1991 Key words: bicarbonate, Bradyrhizobium strains, iron deficiency, peanuts (Arachis hypogaea L.), symbiosis Abstract In the present study, we examined the effects of iron deficiency in an acid solution and in an alkaline solution containing bicarbonate on the growth and nodulation of peanuts inoculated with different bradyrhizobial strains or supplied with fertilizer nitrogen. Inadequate iron supply in acid solution decreased the number of nodule initials, nodule number and nodule mass. Alleviating the iron deficiency increased acetylene reduction but not bacteroid numbers in nodules. Nitrogen concentrations in shoots of inoculated plants increased as iron concentrations in solution increased when determined at day 30 but not at day 50. Higher iron concentrations in solution were required for maximum growth of plants reliant on symbiotic nitrogen fixation than for those receiving fertilizer nitrogen. Adding bicarbonate to the solution with 7.5/~M Fe markedly depressed nodule formation. This effect was much more severe than that of inadequate iron supply alone. Bicarbonate also decreased nitrogenase activity but did not decrease bacteroid concentrations in nodules. Both NC92 and TALl000 nodulated peanuts poorly when bicarbonate was present. However, an interaction between iron concentrations in acid solutions and Bradyrhizobium strains on nodulation of peanuts was observed. Alleviating iron deficiency increased the number of nodule initials and nodules to a much greater extent for plants inoculated with TALl000 than for plants inoculated with NC92. Introduction Iron deficiency occurs mainly on calcareous soils which have low iron availability and is usually associated with high concentrations of bicarbon- ate. It is an important limiting factor in legume production, particularly for peanut (A. hypogaea L.), which appears to be one of the most suscep- tible crops to lime-induced iron deficiency (Chen and Barak, 1982). The response of the peanut-bradyrhizobium symbiosis to iron application has generally been studied under alkaline conditions. Thus, O'Hara et al. (1988a) showed that foliar application of iron to peanuts grown in a calcareous soil in- creased nodule number and nodule mass, though they did not distinguish between the effect of iron deficiency per se and an effect of bicarbon- ate. The first experiment reported here tested the hypothesis that both low iron concentrations and high bicarbonate concentrations impair the symbiosis by limiting nodule initiation, nodule
Transcript of Inadequate iron supply and high bicarbonate impair the symbiosis of peanuts ( Arachis hypogaea L.)...

Plant and Soil 138: 159-168, 1991 © 1991 Kluwer Academic Publishers. Printed in the Netherlands. PLSO 8948
Inadequate iron supply and high bicarbonate impair the symbiosis of peanuts (Arachis hypogaea L.) with different Bradyrhizobium strains
CAIXIAN TANG 1'3, A.D. ROBSON l and M.J. DILWORTH 2 ~Soil Science and Plant Nutrition, School of Agriculture, The University of Western Australia, Nedlands, W.A. 6009, Australia, 2School o f Biological and Environmental Sciences, Murdoch University, Murdoch, W.A. 6150, Australia and 3Soil Science and Agricultural Chemistry, Zhejiang Agricultural University, Hangzhou, China
Received 26 October 1990. Revised July 1991
Key words: bicarbonate, Bradyrhizobium strains, iron deficiency, peanuts (Arachis hypogaea L.), symbiosis
Abstract
In the present study, we examined the effects of iron deficiency in an acid solution and in an alkaline solution containing bicarbonate on the growth and nodulation of peanuts inoculated with different bradyrhizobial strains or supplied with fertilizer nitrogen.
Inadequate iron supply in acid solution decreased the number of nodule initials, nodule number and nodule mass. Alleviating the iron deficiency increased acetylene reduction but not bacteroid numbers in nodules. Nitrogen concentrations in shoots of inoculated plants increased as iron concentrations in solution increased when determined at day 30 but not at day 50. Higher iron concentrations in solution were required for maximum growth of plants reliant on symbiotic nitrogen fixation than for those receiving fertilizer nitrogen.
Adding bicarbonate to the solution with 7.5/~M Fe markedly depressed nodule formation. This effect was much more severe than that of inadequate iron supply alone. Bicarbonate also decreased nitrogenase activity but did not decrease bacteroid concentrations in nodules.
Both NC92 and TALl000 nodulated peanuts poorly when bicarbonate was present. However, an interaction between iron concentrations in acid solutions and Bradyrhizobium strains on nodulation of peanuts was observed. Alleviating iron deficiency increased the number of nodule initials and nodules to a much greater extent for plants inoculated with TALl000 than for plants inoculated with NC92.
Introduction
Iron deficiency occurs mainly on calcareous soils which have low iron availability and is usually associated with high concentrations of bicarbon- ate. It is an important limiting factor in legume production, particularly for peanut (A. hypogaea L.), which appears to be one of the most suscep- tible crops to lime-induced iron deficiency (Chen and Barak, 1982).
The response of the peanut-bradyrhizobium
symbiosis to iron application has generally been studied under alkaline conditions. Thus, O'Hara et al. (1988a) showed that foliar application of iron to peanuts grown in a calcareous soil in- creased nodule number and nodule mass, though they did not distinguish between the effect of iron deficiency per se and an effect of bicarbon- ate. The first experiment reported here tested the hypothesis that both low iron concentrations and high bicarbonate concentrations impair the symbiosis by limiting nodule initiation, nodule

160 Tang et al.
development, nodule function or host plant growth.
Many gram-negative bacteria, including rhizobia and bradyrhizobia, respond to iron de- ficiency by producing high affinity iron-chelating compounds called siderophores to scavenge iron from their environments (Neilands, 1981). Vari- ous strains of bradyrhizobia differ in their ability to produce siderophores (Nambiar and Sivaramakrishnan, 1987). Recently, O'Hara et al. (1988b) reported from studies with high bicar- bonate concentrations at pH 8.6, that inoculation of peanuts with bradyrhizobium strain NC92 re- sulted in virtually normal growth of plants under iron stress while inoculation with strain TALl000 did not, and that NC92 was recognized as a catechol-type siderophore producer. Sidero- phores produced by bradyrhizobia could be im- portant in supplying iron in alkaline solutions with low iron concentration. The second experi- ment therefore attempted to compare the re- sponse of these two different bradyrhizobial strains in the symbiosis to both iron deficiency in acid solutions and to iron stress induced by high bicarbonate concentrations.
Materials and methods
Experiment 1
Plants were grown in a glasshouse in solution culture with roots maintained at 25°C. The de- sign of the experiment was a factorial combina- tion of four iron treatments (0.05, 0.4, 7.5 p.M Fe and 7.5/.tM Fe plus bicarbonate [7.5+ HCO3]) and three inoculation treatments (inoculation with Bradyrhizobium CB756, inocu- lation with Bradyrhizobium NC92 or combined nitrogen without inoculation [N]) with three re- plicates of each treatment.
Uniform sized seeds (mean Fe content 14/~g seed -1) of peanuts (A. hypogaea L.) cv. Tainan 9 were germinated on a stainless steel screen sitting over an aerated solution of 0.6 mM CaCI 2 and 10 ~ M H3BO 3 for 7 d. Seven seedlings were then transplanted into 5-L plastic pots containing continuously-aerated solutions with basal nu- trients in the following concentrations (/xM):
KHzPO4, 40; K2SO4, 1000; MgSO4, 400; CaC12, 1200; H3BO3, 10; Na2MoO4, 0.06; ZnSO4, 1.5; MnSO4, 2.0; COSO4, 0.4 and CuSO 4, 0.4. Iron was supplied as sequestrene-138Fe ® (Fem EDDHA). For the treatment containing HCO 3, 0.5 g L - l CaCO 3 and 5 mM NaHCO 3 were added. An inoculum of Bradyrhizobium NC92 or CB756 was grown on slopes of YGMA, cells suspended in deionized water (Tang et al., 1990a) and added to give a final concentration of 2 x 105 cells mL -~. The solution was unchanged for 4 d and replaced by a similar solution with added bradyrhizobia for another 4 d, after which solutions were changed every third day with no further additions of Bradyrhizobium. At day 34, combined nitrogen (1 mM N) with a ratio of 4 NO 3 (as Ca(NO3)2) to 1 NH 4 (as NHaNO3) was added to uninoculated plants at a time when it was estimated that N 2 fixation had commenced in inoculated plants. Solution pH was adjusted daily to 5.5 with KOH except in the HCO 3 treatment, where the solution pH was 8.6 to 8.7.
Plants were harvested at 20, 30 and 50 days after germination and separated into youngest fully expanded leaf blades (YEB), shoots, roots and nodules. To remove dust, entire plant shoots were washed in 0.1 N HCI and rinsed in deion- ized water. Roots were placed in 0.01 M CaC12 for 5 min and then rinsed in deionized water to remove exchangeable iron. One leaflet of the YEB was sampled for chlorophyll measurement. Other plant tissues and remaining nodules were oven-dried at 68°C for iron analysis.
Nodule initials were examined under a dissect- ing microscope as described by O'Hara et al. (1988a). Peanut nodules only occur at sites of emerging lateral roots (Chandler, 1978). Nodule initials are spherical and originate within the cortex of the roots. The initials represent an index of successful infection and initiation allow- ing this stage to be separated from subsequent development (Riley and Dilworth, 1985a). Acetylene reduction (Trinick et al., 1976) was assayed at 25°C. The method for bacteroid enumeration (haemocytometer counting in dark field microscopy) was as described by Riley and Dilworth (1985b). The methods for measure- ment of chlorophyl concentration, iron and ni- trogen were as previously described (Tang et al., 1990a).

Iron deficiency and nitrogen fixation in peanuts 161
Experiment 2
This experiment was a factorial combination of three iron treatments (0.05, 7.5 and 7.5/~M FeIUEDDHA plus bicarbonate [7.5 + HCO3] ) and two bradyrhizobial strains (NC92 and TALl000) with two replicates of each treatment.
Seven 6-day-old seedlings was transferred to each pot with 5-L nutrient solution. An inoculum of Bradyrhizobium NC92 or TALl000 was added at a rate of 2 × 105 cells mL-~ and the solution kept unchanged for 4d, after which solutions were changed every third day without further additions of bradyrhizobia. Plants were harves- ted at days 18, 22 and 35 after germination, and all other procedures were as in the first ex- periment.
Results
Experiment 1
roots was unaffected by iron concentrations in solution (Table 1). A significant response to iron had developed at the second harvest (day 30) except in uninoculated plants and was even greater by day 50. Chlorophyll concentrations were reduced by inadequate iron supply at all harvests.
Adding bicarbonate to the solution at 7.5/~M iron decreased the dry weight of shoots and roots at day 50, and chlorophyll concentration in the YEB at days 30 (except in uninoculated plants) and 50.
At day 30, inoculated plants had greater shoot weight than uninoculated plants without com- bined nitrogen at 0.4 and 7.5/~M Fe but not at 0.05 p~M Fe or where bicarbonate was added. By day 50, inoculated plants appeared to have high- er external iron requirements to achieve max- imum dry weight than did combined nitrogen-fed plants. In addition, by day 50, CB756-inoculated plants had greater shoot weight than those inocu- lated with NC92 in all treatments.
Symptoms Interveinal chlorosis of the young leaves was evident from day 18 on plants receiving 0.05/~M Fe and persisted throughout the experiment. The young leaves of plants receiving 0.4/~M Fe ap- peared mildly chlorotic. Roots exposed to 0.05/~M Fe were yellow-brown with new, white laterals. There were few symptoms of iron de- ficiency on plants supplied with 7.5/~M iron.
Adding bicarbonate at 7.5/~M iron induced iron deficiency. The plants showed the chlorosis of iron deficiency from day 22 but slightly re- covered by day 35. Plants treated with 5 mM HCO 3 had dark-brown taproots with short and prolific laterals. Root tips were black and the growth was stunted. The reduced growth and darkening of root tips were obvious 1 d after treatments were imposed. Leaves were necrotic at margins and tips.
All plants suffered slight nitrogen deficiency (general yellowing) about 3 weeks after germina- tion. Inoculated plants recovered from the de- ficiency on day 30 in bicarbonate-free treatments and on day 35 in the bicarbonate treatment.
Plant growth At the first harvest, dry weight of shoots and
Iron concentrations Iron concentrations in all parts of plants were clearly decreased by the decrease in iron concen- tration from 7.5 to 0.05/xM in solution (Table 2). Adding bicarbonate markedly decreased total iron content in tissues (Tables 1 and 2) but this decrease was much greater in shoots than in roots, implying an inhibition of iron transport to the shoots. Bicarbonate reduced the iron concen- tration in YEB and shoots by 45 to 65% and by 20 to 40% in roots and nodules respectively. Uninoculated plants supplied with combined ni- trogen generally had higher iron concentrations in tops than N2-fixing plants (day 50).
Nodulation Irrespective of bradyrhizobial strain, decreasing the iron concentration from 7.5 to 0.05/xM de- creased the number of nodule initials and visible nodules, and the nodule weight of inoculated plants (Table 3). Nodule size was not influenced by iron levels in the solution (data not shown), indicating that the effect of iron on nodule weight per plant is due to effects on nodule formation.
The presence of bicarbonate in the solution markedly decreased the number of nodule ini-

162 Tang et al.
Table 1. Effects of iron supply and bicarbonate on the dry weight of shoots and roots, and chlorophyll concentration in the youngest fully expanded leaf blade (YEB) of Arachis hypogaea with different nitrogen sources (Expt. 1)
Days to Iron harvest treatments
(p.M)
Shoots (g plant- ~ ) Roots (g plant - 1 ) Chlorophyll (mg g - ~ f. wt)
NC92 CB756 N a NC92 CB756 N NC92 CB756 N
20
30
50
0.05 0.25 0.22 0.18 0.17 0.11 0.11 0.96 0.98 0.87 0.4 0.26 0.19 0.23 0.14 0.09 0.12 1.29 1.26 1.18 7.5 0.30 0.26 0.20 0.19 0.12 0.11 1.25 1.21 1.33 7.5 + HCO 3 0.27 0.33 0.23 0.14 0.16 0.12 1.30 1.17 1.11
0.05 0.63 0.67 0.54 0.20 0.23 0.22 0.39 0.44 0.40 0.4 0.75 0.70 0.55 0.22 0.19 0.28 0.67 0.88 0.46 7.5 0.82 0.86 0.62 0.25 0.26 0.30 0.76 0.93 0.46 7.5 + HCO 3 0.74 0.67 0.65 0.27 0.25 0.27 0.42 0.42 0.49
0.05 0.73 0.82 1.62 0.24 0.27 0.48 0.24 0.09 0.53 0.4 1.54 1.98 2.68 0.49 0.54 0.89 1.35 1.45 1.13 7.5 2.05 3.19 2.91 0.52 0.75 0.94 1.44 1.53 1.21 7.5 + HCO 3 0.91 1.16 1.20 0.42 0.42 0.62 0.86 1.16 0.72
LSD ( p = 0.05) 20 NS b NS 0.23
30 0.17 0.07 0.15 50 0.53 0.13 0.27
a Uninoculated plants receiving combined nitrogen at day 35 after germination. Not significant at p = 0.05.
Table 2. Effects of iron supply and bicarbonate on iron concentration (p.g g-~ dry wt.) in youngest fully expanded leaf blades (YEB), shoots, roots and nodules of Arachis hypogaea inoculated with Bradyrhizobium strains or supplied with fertilizer nitrogen (Expt. 1)
Nodules Days to Iron YEB Shoots Roots harvest treatments
(p.M) NC92 CB756 N a NC92 CB756 N NC92 CB756 N NC92 CB756
20
30
50
0.05 25 f 27 ef 28 ef 23 ef 22 e 25 de n.d. ~ n.d. n.d. n.d. n.d. 0.4 42 cde 33 def 43 bcd 36 b 34 bc 30 bcd n.d. n.d. n.d. n.d. n.d. 7.5 65 ab 62 abc 69 a 74 a 57 a 52 ab n.d. n.d. n.d. n.d. n.d. 7.5 + HCO 3 36 def 37 def 24 f 26 cde 26 cde 18 f n.d. n.d. n.d. n.d. n.d.
0.05 17f 20f 18f 17d 17d 15d 62e 71e 49f 95d l15d 0.4 42 cd 57 bc 32 e 24 bc 27 b 23 bc 127 c 103 cd 92 d 304 bc 271 bc 7.5 66 ab 85 a 42 cd 63 a 68 a 54 a 399 a 353 ab 278 b 425 a 447 a 7.5 + HCO~ 33 de 30 e 28 e 18 cd 19 cd 19 cd 193 b 201 b 189 b 237 c 306 b
0.05 16 fg 14 g 25 e 14 g 18 fg 20 ef 91 f 95 ef 72 g 99 e 151 d 0.4 35c 35c 35c 31cd 28de 41bc 166d 149d l l 0 e 302c 344c 7.5 41 bc 49 ab 55 a 51 ab 59 a 65 a 417 a 316 b 309 b 483 a 433 ab 7.5 + HCO 3 20 ef 33 cd 26 de 21 ef 23 ef 21 ef 321 b 325 b 248 c 320 c 406 b
Data were analyzed as the natural logarithm. Values of each harvest followed by the same letter are not significantly different at p = 0.05. "Uninoculated plants receiving combined nitrogen at day 35. b Insufficient samples, not determined.

Iron deficiency and nitrogen fixation in peanuts 163
Table 3. Influence of iron and bicarbonate supply on the number of nodule initials and visible nodules, and fresh weight of
nodules of Arachis hypogaea inoculated with Bradyrhizobium strains (Expt. 1)
Days to Iron Number of nodule Nodule number Nodule weight harvest treatments initials (No plant-' ) (No plant ~) (g plant ~ )
(ktM) NC92 CB756 NC92 CB756 NC92 CB756
20
31)
50
0.05 71 b 62 b 14 b 13 b n.d? n.d. 0.4 84 ab 68 b 26 a 16 b n.d. n.d. 7.5 115 a 85 ab 4(I a 19 ab n.d. n.d. 7.5 + HCO3 9 c 7 c 2 c 1 c n.d. n.d.
0.05 133 b 140 b 0.28 b 0.26 b 0.4 208 a 166 ab 0.42 a 0.30 b 7.5 202 a 222 a 0.46 a 0.42 a 7.5 + HCO 3 26 c 8 d 0.12 c (I.05 c
0.05 172 b 172 b 0.46 c (I.39 cd 0.4 374 a 292 a 1.29 a (I.86 b 7.5 389 a 416 a 1.56 a 1.39 a 7.5 + HCO~ 101 c 60 c 0.20 e 0.25 de
Data were analyzed as log,v Values of each harvest followed by different letters are different at p = 0.05. ~' Insufficient samples, not determined.
tials, the number of nodules, and total nodule weight. Plant exposed to bicarbonate solution with 7.5/zM Fe produced only one tenth the number of nodule initials and nodules of plants receiving 0.05/xM Fe. A higher proportion of lateral nodules was observed in bicarbonate- treated plants at day 50 (data not shown). More- over, nodule size (data not shown) was increased where bicarbonate was present. Thus few nodules on taproots of bicarbonate-treated plants were compensated by increasing both the
number of lateral nodules and nodule size. No nodules were formed on uninoculated plants.
Acetylene reduction activity Plants receiving 7.5 and 0.4/zM iron had double (day 30) and five-fold (day 50) higher acetylene reduction activity on a fresh-weight basis than those given 0.05/.~M iron (Table 4). Total acetylene reduction activity per plant increased markedly with increase of iron supply with the maximum at 7.5/zM Fe. Bicarbonate addition
Table 4. Effects of iron and bicarbonate on acetylene reduction in nodules of Arachis hypogaea inoculated with Bradyrhizobium strains (Expt. i )
Days Iron Specific Total to treatments (nmolg ~ f.wt min ~) (nmolplant t min ~) harvest (/x M)
NC92 CB756 NC92 CB756
3(I
5(I
0.05 32 c 52 bc 26 bc 40 b I).4 79 ab 104 a 97 a 95 a 7.5 78 ab 108 a 105 a 139 a 7.5 + HCO 3 52 bc 47 bc 21 c 7 d
0.(15 18 b 21 b 25 d 25 d 0.4 109 a 101 a 422 ab 261 b 7.5 112 a 102 a 514 a 400 ab 7.5 + HCO 3 78 a 79 a 70 c 56 c
Data of total acetylene reduction were analyzed as log~.. Values of each harvest followed by different letters are different at p = 0.05.

164 Tang et al.
inhibited specific acetylene reduction activity about 25% and dramatically depressed total ac- tivity. While the possibility of differential effects of acetylene on the treatments cannot be ex- cluded, the general picture of the effects of increasing iron concentration and of bicarbonate/high pH is similar to that for shoot nitrogen content.
Bacteroid concentration Bacteroid concentrations in nodules (number per unit fresh weight) were not significantly affected by either iron deficiency or bicarbonate stress (data not shown). The increase of nodule bac- teroid content per plant by increasing iron supply and its decrease by bicarbonate addition was due to effects of these treatments on nodule mass.
Nitrogen in shoots Nitrogen concentrations and contents of shoots were affected by iron treatments and nitrogen sources (Table 5). At day 30, nitrogen concen- tration in uninoculated plants (receiving no nitro-
gen at this stage) decreased and nitrogen content remained unchanged as iron supply increased. In contrast, both nitrogen concentrations and con- tents of plants inoculated with CB756 were in- creased by increasing the iron supply. In NC92- inoculated plants, nitrogen concentration re- mained unaffected while the content was in- creased by increasing the iron concentration in the solution. By day 50, the three nitrogen sources exhibited similar effects of iron treat- ments on nitrogen concentration and content of shoots. NC92-inoculated plants had lower con- centrations and contents of nitrogen in shoots than CB756-inoculated plants.
Regardless of nitrogen sources, adding bicar- bonate clearly decreased nitrogen concentration and content of shoots. Under bicarbonate stress, the nitrogen contents and concentrations of inoculated plants were as low as in uninoculated plants receiving no nitrogen at day 30, indicating that no N 2 had been fixed. Nitrogen concen- trations of inoculated plants exposed to bicar- bonate were the same as, and the content was similar to, that of N-fed plants at day 50.
Table 5. Nitrogen concentration and content in shoots of Arachis hypogaea receiving Bradyrhizobium-fixed or fertilizer nitrogen as influenced by iron supply and bicarbonate (Expt. 1)
Days to Iron Concentration (% dry wt) Content (mg plant -~) harvest treatments
(/~M) NC92 CB756 Na NC92 CB756 N
30
50
0.05 1.95 2.18 1.81 12 14 I0 0.4 2.17 2.55 1.51 16 18 8 7.5 1.97 2.58 1.41 15 22 9 7.5 + HCO3 1.44 1.35 1.55 11 9 11
0.05 2.31 3.26 3.38 16 30 54 0.4 2.53 3.38 3.57 40 74 87 7.5 2.47 2.83 3.29 49 88 94 7.5 + HCO 3 2.05 2.02 2.02 19 25 26
LSD (p = 0.05) 30
50
Fe 0.23 NS ~ N 0.20 4 Fe × N 0.39 NS
Fe 0.20 15 N 0.17 13 Fe x N 0.34 NS
a Uninoculated plants receiving combined nitrogen at day 35. b Not significant at p = 0.05.

Experiment 2
Plant growth The symptoms of iron deficiency and bicarbonate toxicity shown on plants were similar to those observed in the first experiment.
Shoot and root weights, and chlorophyll con- centrations of plants inoculated with either bradyrhizobial strain were not significantly af- fected by iron treatments at days 18 and 22 although chlorosis of young leaves appeared in 0.05 p~M Fe and bicarbonate treatments from day 20. By day 35, root weight and chlorophyll concentration were reduced in 0.05 ~ M Fe and bicarbonate treatments while shoot weight re- mained unaffected (data not shown).
Iron concentration Iron concentrations in YEB, shoots, roots and nodules of plants grown in 7.5 ~ M Fe solution were four times greater than those of plants grown in 0.05/~M Fe solution (Table 6). Nodules had the highest iron concentration at both iron levels.
As in the first experiment, bicarbonate de- creased both iron absorption by roots and trans- location to shoots. The effect of bicarbonate in decreasing iron concentration in shoots was
Iron deficiency and nitrogen fixation in peanuts 165
greater than that on roots. The Bradyrhizobium strains used did not affect iron concentration in host tissues.
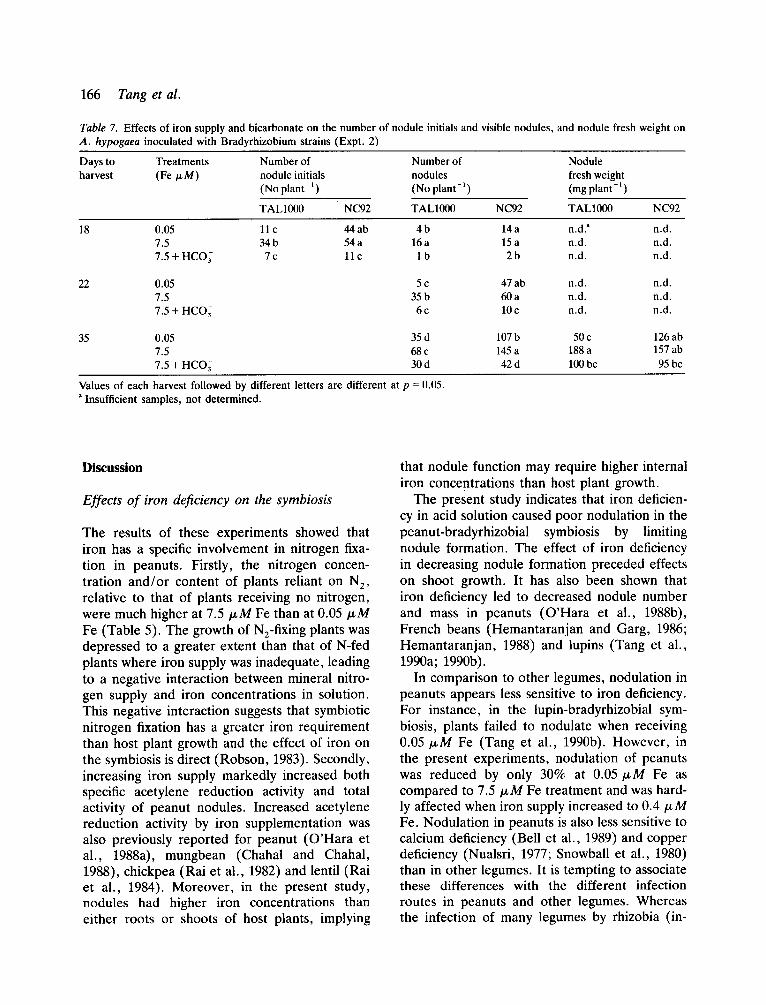
Nodulation Plants receiving 7.5 txM Fe had more nodule initials and nodules than those receiving 0.05/xM Fe (Table 7). Adding bicarbonate to the solution with 7.5/xM iron markedly depres- sed nodule formation.
There was a clear interaction between iron concentration in solution and bradyrhizobial strain on nodule formation (Table 7). TAL1000- inoculated plants given 0.05/xM Fe produced only one seventh to one third of the nodule initials and nodules of plants given 7.5/~M Fe. In comparison, NC92-inoculated plants given 0.05 ~ M Fe produced 80% of the initials and nodules of plants given 7.5 fzM Fe. Moreover, TAL1000-inoculated plants formed fewer and larger nodules at all treatments, and had a higher proportion of lateral nodules at 0.05/xM iron than NC92-inoculated plants.
The effect of treatments on bacteroid concen- tration in nodules was similar to the first experi- ment. Moreover, the effect of iron and bicarbon- ate concentrations on bacteroid concentration in nodules was similar for the two bradyrhizobial strains.
Table 6. Iron concentrations (~gg -~ dry wt.) in the youngest fully expanded leaf blade (YEB), shoots, roots and nodules of Bradyrhizobium inoculated peanuts in relation to iron and bicarbonate applications (Expt. 2)
Days to Treatments YEB Shoots Roots Nodules
harvest ( F e / x M ) TALl000 NC92 TALl000 NC92 TALl000 NC92 TALl000 NC92
18
22
35
(/.05 41 b 4(1 b 35 c 32 c 70 c 67 c n.d." n.d. 7.5 177a 200a 130a 101 b 270a 256a n.d. n.d. 7.5 + HCO 3 47b 45b 31c 28c 195b 22 lab n,d. n.d.
(1.05 38b 41 b 34b 35b 76c 79c n.d. n.d.
7.5 154 a 159 a 139 a 143 a 279 a 258 ab n.d. n.d. 7.5 + HCO~ 43 b 41 b 36 b 35 b 239 b 284 a n.d. n.d.
0.05 34b 32b 29c 30c 89b 86b n.d. 180b 7.5 103 a 108 a 129 a 141 a 372 a 350 a 374 a 429 a 7.5 + HCO3 41 b 38 b 42 b 44 b 3(12 a 326 a 321 a 361 a
Data were analyzed as the natural logarithm. Values of each harvest which do not have a common letter are different at p = 0.05. ~' Insufficient samples, not determined.
166 Tang et al.
Table 7. Effects of iron supply and bicarbonate on the number of nodule initials and visible nodules, and nodule fresh weight on A. hypogaea inoculated with Bradyrhizobium strains (Expt. 2)
Days to Treatments Number of Number of Nodule harvest ( Fe/z M) nodule initials nodules fresh weight
(No plant- t) (No plant -~ ) (mg plant-l)
TALl000 NC92 TALl000 NC92 TALl000 NC92
18
22
35
0.05 11 c 44 ab 4 b 14 a n.d. a n.d. 7.5 34 b 54 a 16 a 15 a n.d. n.d. 7.5 + HCO 3 7c 11 c 1 b 2b n.d. n.d.
0.05 5 c 47 ab n.d. n.d. 7.5 35 b 60 a n.d. n.d. 7.5 + HCO3 6 c 10 c n.d. n.d.
0.05 35 d 107 b 50 c 126 ab 7.5 68 c 145 a 188 a 157 ab 7.5 + HCO~ 30 d 42 d 100 bc 95 bc
Values of each harvest followed by different letters are different at p = 0.05. "Insufficient samples, not determined.
Discussion
Effects of iron deficiency on the symbiosis
The results of these experiments showed that iron has a specific involvement in nitrogen fixa- tion in peanuts. Firstly, the nitrogen concen- tration and/or content of plants reliant on N 2, relative to that of plants receiving no nitrogen, were much higher at 7.5/xM Fe than at 0.05/~M Fe (Table 5). The growth of N2-fixing plants was depressed to a greater extent than that of N-fed plants where iron supply was inadequate, leading to a negative interaction between mineral nitro- gen supply and iron concentrations in solution. This negative interaction suggests that symbiotic nitrogen fixation has a greater iron requirement than host plant growth and the effect of iron on the symbiosis is direct (Robson, 1983). Secondly, increasing iron supply markedly increased both specific acetylene reduction activity and total activity of peanut nodules. Increased acetylene reduction activity by iron supplementation was also previously reported for peanut (O'Hara et al., 1988a), mungbean (Chahal and Chahal, 1988), chickpea (Rai et al., 1982) and lentil (Rai et al., 1984). Moreover, in the present study, nodules had higher iron concentrations than either roots or shoots of host plants, implying
that nodule function may require higher internal iron concentrations than host plant growth.
The present study indicates that iron deficien- cy in acid solution caused poor nodulation in the peanut-bradyrhizobial symbiosis by limiting nodule formation. The effect of iron deficiency in decreasing nodule formation preceded effects on shoot growth. It has also been shown that iron deficiency led to decreased nodule number and mass in peanuts (O'Hara et al., 1988b), French beans (Hemantaranjan and Garg, 1986; Hemantaranjan, 1988) and lupins (Tang et al., 1990a; 1990b).
In comparison to other legumes, nodulation in peanuts appears less sensitive to iron deficiency, For instance, in the lupin-bradyrhizobial sym- biosis, plants failed to nodulate when receiving 0.05/zM Fe (Tang et al., 1990b). However, in the present experiments, nodulation of peanuts was reduced by only 30% at 0.05/xM Fe as compared to 7 .5/zM Fe treatment and was hard- ly affected when iron supply increased to 0.4/~M Fe. Nodulation in peanuts is also less sensitive to calcium deficiency (Bell et al., 1989) and copper deficiency (Nualsri, 1977; Snowball et al., 1980) than in other legumes. It is tempting to associate these differences with the different infection routes in peanuts and other legumes. Whereas the infection of many legumes by rhizobia (in-

Iron deficiency and nitrogen fixation in peanuts 167
cluding lupins, Robertson et al., 1978) is re- ported to involve the formation of infection threads (Sprent, 1984), rhizobia enter the peanut root intercellularly via the middle lamellae of cells (Chandler, 1978). If the deficiency of a nutrient inhibits the formation of infection threads, e.g. calcium deficiency (Lowther and Loneragan, 1968, 1970), the peanut-brady- rhizobium symbiosis may escape the effects of nutrient deficiencies which affect infection thread development. The most sensitive stage to iron deficiency may also be different in different sym- bioses. In peanuts, it was suggested that iron deficiency limited development of nodules al- ready initiated (O'Hara et al., 1988a). However, nodules could develop normally in lupins under iron deficiency once nodule initiation has occurred (Tang et al., 1990b). In addition, peanuts have larger seeds than most legumes. Higher seed iron content may also be beneficial for the nodulation. Moreover, iron transfer from the host root to the nodule bacteria in peanuts (O'Hara et al., 1988a) could be greater than that in other legumes such as lupins (Tang et al., 1990b).
Effects of bicarbonate~high pH on the symbiosis
Adding bicarbonate to the solution markedly depressed nodule formation and N 2 fixation in the peanut-bradyrhizobium symbiosis. It is un- likely that the effect of bicarbonate/high pH on decreasing nodule formation resulted solely from iron deficiency induced by bicarbonate since iron concentrations in nodules and other tissues were higher than those in the 0.05/xM Fe treatment. Moreover, this effect of bicarbonate was much more severe than the effect of inadequate supply of iron in an acid solution.
Bicarbonate/high pH impaired nodule func- tion as well as nodule formation. Nz-fixing plants growing in the bicarbonate solution contained the same low nitrogen concentration and content as non-inoculated plants growing in nitrogen-free solution. Those growing in bicarbonate-free solu- tion at the same level of iron contained much higher nitrogen concentrations and contents than non-inoculated plants at an early stage (day 30, Table 5). By day 50, N2-dependent plants ex- posed to 5 mM bicarbonate had the same nitro-
gen concentration as N-fed plants. These results suggest that bicarbonate/high pH may delay the onset of N 2 fixation. Bicarbonate/high pH also decreased specific acetylene reduction activity, unrelated to bacteroid proliferation in nodules.
The impaired symbiosis under bicarbonate stress in this study is not due to the deficiency of other nutrients such as copper, manganese and zinc which might be induced by high bicarbonate as the concentrations of these nutrients in the host plant (data not shown) were all in the adequate range (Nualsri, 1977; Reuter and Robinson, 1986).
Different responses of Bradyrhizobium strains to iron deficiency
Bradyrhizobium strains differ substantially in their ability to nodulate peanuts grown in an acid solution with low concentrations of iron. Where external supply of iron was limited, strain NC92 showed a greater ability to form nodules than TALl000. Similar results were obtained with peanuts grown on an iron-stressed soil (G W O'Hara, personal communication). Where no iron was applied to the soil, TAL1000-inoculated plants failed to form any nodules whereas NC92- inoculated plants produced more than 50% the nodule mass of those receiving FemEDDHA. The effectiveness of strains in establishing the symbiosis with their host plants could be related to their ability to produce siderophores. A catechol-type siderophore was detected in nu- trient solution for peanuts inoculated with NC92 but not for those with TALl000 (O'Hara et al., 1988b). Thus the siderophore producing strain may have been able to scavenge iron from the external medium or host roots, or may have had a lower iron requirement for its proliferation to initiate nodules under iron deficiency.
In contrast with the response to iron deficien- cy, the bradyrhizobial strains used in the present study had a similar response to bicarbonate/high pH. All strains nodulated peanuts poorly in the bicarbonate treatments. The results imply that the tolerances of bradyrhizobium strains to low iron and to bicarbonate/high pH are controlled differently.
The different ability of bradyrhizobial strains to establish the symbiosis under iron deficiency

168 Tang et al.
has important implications. For legumes to be grown successfully, strains which are tolerant of iron deficiency should be selected. In addition, as most iron-stressed soils are associated with high bicarbonate, selecting bicarbonate-tolerant strains may have some importance.
Acknowledgements
Grateful acknowledgements are made to Dr G W O'Hara, P Sanford and Dr R W Bell for their technical assistance.
References
Bell R W, Edwards D G and Asher C J 1989 External calcium requirements for growth and nodulation of six tropical food legumes grown in flowing solution culture. Aust. J. Agric. Res. 40, 85-96.
Chahal P P K and Chahal V P S 1988 Introduction of Rhizobium and Meloidogyne incognita on symbiotic nitro- gen fixation in mungbean under different concentrations of iron. J. Plant Nutri. 11, 821-828.
Chandler M R 1978 Some observations on infection of Arachis hypogaeaa L. by Rhizobium. J. Exp. Bot. 29, 749-755.
Chen Y and Barak P 1982 Iron nutrition of plants in calcare- ous soils. Adv. Agron. 35, 217-240.
Hemantaranjan A and Garg O K 1986 Introduction of nitrogen-fixing nodules through iron and zinc fertilization in the non nodule-forming French bean (Phaseolus vulgaris L.). J. Plant Nutr. 9, 281-288.
Lowther W L and Loneragan J F 1968 Calcium and nodula- tion in subterranean clover (Trifolium subterraneum L.). Plant Physiol. 43, 1362-1366.
Lowther W L and Loneragan J F 1970 Calcium in the nodulation of legumes. In Proceeding of Xlth International Grassland Congress, Surfers Paradise. Ed. M J T Norman. pp 446-50. The University of Queensland Press, St. Lucia, Queensland.
Nambiar P T C and Sivaramakrishnan S 1987 Detection and assay for siderophores in cowpea rhizobia (Brady- rhizobium) using radioactive Fe(~gFe). Lett. Appl. Mi- crobiol. 4, 37-40.
Neilands J B 1981 Microbial iron compounds. Annu. Rev. Biochemi. 50, 715-731.
Nualsri L 1977 Copper Nutrition in Peanuts. Ph.D. Thesis. The University of Western Australia.
O'Hara G W, Dilworth M J, Boonkerd N and Parkpian P 1988a Iron deficiency specifically limits nodule develop- ment in peanut inoculated with Bradyrhizobium sp. New Phytol. 108, 51-57.
O'Hara G W, Hartzook A, Bell R W and Loneragan J F 1988b Response to Bradyrhizobium strains of peanut cup tivars grow under iron stress. J. Plant Nutr. 11, 843-852.
Rai R, Prasad V, Choudhury S K and Sinha N P 1984 Iron nutrition and symbiotic N2-fixation of lentil (Lens culinaris) genotypes in calcareous soil. J. Plant Nutr. 7, 399-405.
Rai R, Singh S N and Prasad V 1982 Effect of pressmud amend pyrite on symbiotic N2-fixation, active iron contents of nodules, grain yield and quality of chickpea (Cicer arietinum Linn.) genotypes in calcareous soil. J. Plant Nutr. 5,905-913.
Reuter D J and Robinson J B 1986 Plant Analysis: An Interpretation Manual. Inkata Press, Melbourne and Sydney.
Riley I T and Dilworth M J 1985a Cobalt status and effects on soil population of Rhizobium lupini, rhizosphere coloni- zation and nodule initiation. Soil Biol. Biochem. 17, 81- 85.
Riley I T and Dilworth M J 1985b Cobalt requirement for nodule development and function in Lupinus angustifolius L. New Phytol. 100, 347-359.
Robertson J G, Lyttleton P, Bullivant S and Grayston G F 1978 Membranes in lupin root nodules. I. The role of Golgi bodies in the biogenesis of infection threads and peribacteroid membranes. J. Cell. Sci. 30, 129-149.
Robson A D 1983 Mineral nutrition. In Nitrogen Fixation, Vol. 3. Ed. W J Broughton. pp 36-55. Clarendon Press, Oxford.
Snowball K, Robson A D and Loneragan J F 1980 The effect of copper on nitrogen fixation in subterranean clover (Tri- folium subterraneum). New Phytol. 85, 63-72.
Sprent J I 1984 Nitrogen fixation. In Advanced Plant Physiol- ogy. Ed. M B Wilkins. pp 249-276. Pitman, London.
Tang C, Robson A D and Dilworth M J 1990a The role of iron in nodulation and nitrogen fixation in Lupinus angus- tifolius L. New Phytol. 114, 173-182.
Tang C, Robson A D and Dilworth M J 1990b A split-root experiment shows that iron is required for nodule initiation in Lupinus angustifolius L. New Phytol. 115, 61-67.
Trinick M J, Dilworth M J and Grounds M 1976 Factors affecting the reduction of acetylene by root nodules of Lupinus species. New Phytol. 77, 359-370.




















