
The characteristic changes of betung bamboo (Dendrocalamus asper) pretreated by fungal pretreatment

Pergamon Tooricnn, Vol. 34. No 5. pp. 567-577. 1996
Copyright <I 1996 Elrevier kence Ltd. All rqhts resewed Pruned in Great Britain
0041-0101(95)00162-X 0041 OlOl:96 $15.00 + 000
NEUTRALIZATION OF MYONECROSIS, HEMORRHAGE, AND EDEMA INDUCED BY
BOTHROPS ASPER SNAKE VENOM BY HOMOLOGOUS AND HETEROLOGOUS PRE-EXISTING ANTIBODIES IN
MICE
ALEXANDRA RUCAVADO and BRUNO LOMONTE* lnstituto Clodomiro Picado, Facultad de Microbiologia, Universidad de Costa Rica, San Jo&,
Costa Rica
(Rewired 19 September 1995; accepted 22 November 1995)
A. Rucavado and B. Lomonte. Neutralization of myonecrosis, hemorrhage, and edema induced by Bothrops asper snake venom by homologous and heterologous pre-existing antibodies in mice. Toxicon, 34, 567-577, 1996.-The ability of pre-existing antibodies to neutralize locally-acting toxins of Bothrops asper snake venom was investigated. Hemorrhage, myonecrosis, and edema were markedly reduced in actively immunized mice, although none of these effects was completely abolished. In mice passively immunized with equine antivenom, hemorrhage was prevented completely, while myonecrosis and edema were partially reduced. Pre-existing antibodies did not modify the early stage ( < 3 hr) of venom-induced edema, but significantly accelerated the normalization of this effect within 24 hr. Passive administration of antivenom either 5 or 120 min before venom injection gave similar results, suggesting that the presence of antibodies in the intravascular compartment may fully neutralize locally acting toxins, in this experimental animal model. Overall, the homologous or heterologous origin of antibodies was not a significant factor influencing their in uit,o neutralizing efficiency against local venom effects. Antibody titrations by enzyme-immunoassay using purified toxins and whole venom indicated that serum from actively-immunized mice had a higher proportion of anti-myotoxin antibodies than equine antivenom. Copyright 0 1996 Elsevier Science Ltd.
INTRODUCTION
In addition to systemic life-threatening effects, the venoms from many species of snakes cause striking tissue damage at the site of injection, classically defined as consisting of hemorrhage, myonecrosis, and edema (Ohsaka, 1979). In severe cases of envenoming, these local effects may lead to permanent tissue loss, disability, or amputation. Clinical studies have described a generally limited effectiveness of antivenom serotherapy in preventing the development of local tissue damage (Reid et al., 1963; Warrell et al., 1976; Nishioka and
*Author to whom correspondence should be addressed.
567

568 A. RUCAVADO and B. LOMONTE
Silveira, 1992; Cardoso et al., 1993). Several experimental animal studies are in agreement with this finding, attributing the low effectiveness to: (a) the lack (or scarcity) of antibodies to particular toxins involved in local tissue damage (Ownby et al., 1979, 1983; Lomonte et al., 1991); and/or (b) the fast action of these toxins, in comparison to the slow in vivo distribution of neutralizing antibodies, despite their presence in antivenoms (Homma and Tu, 1970; Russell et al., 1973; Ownby et al., 1984, 1986; Gutierrez et al., 1981, 1987).
In general, the ability of antivenoms to prevent local tissue damage, even when administered immediately after experimental envenomation, has been only partial. Little work has been done to determine to what extent locally acting toxins can be neutralized by pre-existing antibodies. This question is of interest for two reasons. First, it would provide a better reference frame for the evaluation of the neutralizing efficiency of antivenoms in viz~o, since it would represent the best possible situation for neutralization of toxins. Second, it would be of value in establishing a rationale for possible immunization strategies against venoms in the future.
In the present study, a previously characterized mouse model of Bothrops asper venom-induced local tissue damage (for a review, see Gutierrez and Lomonte, 1989) was utilized. Assuming that the homologous or heterologous origin of antibodies might have an influence on their in vivo distribution, we utilized both actively and passively immunized mice, to assess the extent by which edema, hemorrhage, and myonecrosis, could be prevented.
MATERIALS AND METHODS
Venom and antirenom Bothrops asper venom consisted of a pool from many ( > 50) specimens from the Pacific region of Costa Rica.
It was centrifuged, lyophilized and kept at - 20’C. The equine polyvalent antivenom (batch 2600295; expiry date February 1998) produced at Instituto Clodomiro Picado, University of Costa Rica (Bolaiios and Cerdas, 1980) was used throughout this study.
Active immunization of mice against Bothrops asper cenom Four groups of six Swiss female mice (1618 g body wt) were actively immunized against crude B. asper venom
according to the following schedule: on day 1, they received 20 pg of venom (s.c.) emulsified in Freund’s complete adjuvant (Sigma Chemical Co., St. Louis, MO, U.S.A.); all the subsequent booster doses were given in sodium alginate, via i.p. injection: day 15: 20 ng; day 22: 40 pg; day 29: 60 fig: day 36: 60 ng; and day 46: 100 pg. One week after the final booster dose, three groups of mice were challenged with venom for quantification of edema, hemorrhage, and myonecrosis. respectively, as described below. A group of six matched unimmunized mice served as a control in each of these three challenge tests.
The fourth group of immunized mice was anesthetized and bled retroorbitally, in order to obtain a serum pool that would be representative of the level of immunization achieved, in comparison to the equine antivenom.
Passire immwkation oJ‘ mice against Bothrops asper ccnom Six groups of five mice (18-20 g body wt) received an i.v. injection of 0.4 ml of equine polyvalent antivenom,
before being challenged with venom for quantification of edema, hemorrhage, and myonecrosis, as described below. Antivenom was given either 5 min (three groups) or 120 min (three groups) before the venom, in order to detect possible differences related to the time allowed for antibody distribution in viro. Three groups of five matched untreated mice served as controls in each of the three challenge tests (edema, hemorrhage, and myonecrosis).
Quantificarion of hemorrhage, edema, and myonecrosis in mice
Hemorrhage was quantified by the method of Kondo et al. (1960). Mice received an i.d. injection of 15 pg of venom. in 100 pl of 0.15 M phosphate-buffered saline (PBS), pH 7.2. This dose corresponds to approx. 10 times the ‘minimum hemorrhagic dose’ (dose resulting in a hemorrhagic spot of 10 mm dia in 2 hr). Mice were

Venom Neutralization by Pm-existing Antibodies 569
killed by ether inhalation 2 hr after injection, and the areas of hemorrrhage measured on the internal surface of the dissected skin.
The development of edema was quantified by measuring the increase in footpad thickness using a low-pressure spring caliper (Oditest, H.C. Kroplin, Germany), at 0.5, 1, 3, 6, 12, and 24 hr after the S.C. injection of 5 peg of venom, dissolved in 50 ~1 of PBS (Lomonte ef al., 1993). To control for vehicle-induced increase of footpad thickness, three mice in each experiment received an injection of PBS only.
Myonecrosis was evaluated by quantification of plasma creatine kinase (CK) activity, and by histological observation of muscle damage (Gutitrrez et al., 1981). Mice received an i.m. injection of 50 pg of venom (in 50 ~1 PBS) in the right gastrocnemius. After 3 hr, blood was collected from the tail, into heparizined capillary tubes, for plasma CK determination (Sigma No. 520). In addition, samples of the injected muscles were taken 3 and 24 hr after venom injection, fixed in Karnovsky solution (glutaraldehyde 2.5%, paraformaldehyde 2% in PBS) and 1% osmium tetroxide, dehydrated in ethanol, embedded in Spurr resin, sectioned and stained with Toluidine Blue for microscopic observation. Plasma CK levels of mice that received an injection of PBS only were included as a control.
Comparison of mouse immune seru with equine polyoalent antizwnom In order to assess the level of neutralizing antibodies obtained in mice after active immunization, a serum pool
obtained at the moment of the in airo challenge tests, as described above, was tested for anti-hemorrhagic, anti-myotoxic, and anti-edematigenous potencies, respectively, by preincubation-type neutralization experiments (Gutitrrez et al., 1990). Equine antivenom was assayed in parallel in each test, as a reference. In brief, venom and mouse serum (or equine antivenom) mixtures of different ratios were prepared, incubated 30 min at 37’ C, and then injected into mice. Venom doses and methods were those described above.
Since the limited amount of immune mouse serum obtained was not sufficient for testing the neutralization of lethal activity of the venom, an alternative test additionally performed was an indirect hemolysis neutralization, which has been previously shown to correlate significantly with lethality neutralization (Guticrrez et al., 1988a). In brief, venom and mouse serum (or equine antivenom) mixtures of different ratios were prepared, incubated 30 min at 37°C and then applied to wells in an agarose gel containing egg yolk and sheep erythrocytes, for the subsequent measurement of the hemolytic halos, as described by Gutitrrez er al., 1988a).
The relative content of antibodies to whole B. asper venom, hemorrhagic toxin BaPl (Gutierrez ef al., 1995) and myotoxin I1 (Lomonte and Gutierrez, 1989) in the serum of actively-immunized mice and in equine antivenom, was compared by enzyme-immunoassay, basically as described by Lomonte e/ al. (1991). Antigen concentrations used for microplate coating were I, 0.4, and 0.4 pg/lOO ~l/well, for venom, myotoxin-II, and BaPI, respectively. Samples were titrated in duplicate, with dilutions ranging from 1:200 to 1:204,800. Normal mouse and horse sera were included as negative controls, Murine or equine antibodies were detected with their corresponding anti-immunoglobulin-peroxidase conjugates (Sigma), using hydrogen peroxide and o-phenylendi- amine as chromogenic substrate. Absorbances were recorded at 492 nm using a Dynatech MRWOO reader.
RESULTS
Neutralizing ability of homologous pre-existing antibodies When actively-immunized mice were challenged with B. asper venom, the level of local
tissue damage was significantly reduced, although it was not completely abrogated, in comparison to unimmunized control mice (Fig. 1). The immediate phase ( < 1 hr) of footpad edema was not reduced by the pre-existing antibodies, but at subsequent time points this effect was roughly half of that recorded in unimmunized animals, resolving by 24 hr (Fig. 1A). Plasma CK activity was reduced by about 80% (Fig. lB), whereas hemorrhagic skin lesions were reduced by about 70% (Fig. 1C) in the immunized animals. Histologic evaluation of muscle sections showed a significant reduction of myonecrosis and hemorrhage in the immunized mice.
Neutralizing ability of heterologous pre-existing antibodies When mice passively immunized (with equine antivenom) were challenged with B. asper
venom, hemorrhage was prevented completely (data not shown). Groups of mice receiving

570 A. RUCAVADO and B. LOMONTE
the antivenom either 5 or 120 min before venom had a normal skin appearance after the venom challenge, with no macroscopic signs of erythrocyte extravasation.
Footpad edema at 0.5, 1, and 3 hr after venom injection was similar in the control and in the antivenom pre-treated mice, but at later time points there was a significant reduction of this effect (about 50% at 6 hr) with a tendency to resolve by 24 hr (Fig. 2A). There were no statistically significant differences between the animal groups pre-treated either 5 min or 120 min before venom injection (Fig. 2A).
Plasma CK activity was reduced by about 65% in the passively-immunized animals, with no difference between the groups receiving antivenom either 5 or 120 min before the venom challenge (Fig. 28).
3.0
2.5
” 1200
,E 2.1000
E .g 800
; 600
2 400 4 a 200
0
0 3 6 9 12 24
Time (hr)
: NI I PBS NI I
18 - -2 p -
& 14 - z E 72-
d IO-
$ 8-
e 6-
g 4- x
2-
n-
Fig. 1. EfTect of active immunization of mice on the development of edema, myonecrosis, and hemorrhage induced by the venom of B. asper. (A) Edema is estimated by the increase in footpad thickness after S.C. venom injection (5 pg). Each point represents the mean f S.D. of six mice. W, Unimmunized controls; l , immunized mice; A., PBS. The difference between immunized and unimmunized mice is statistically significant (P < 0.05) at all time points except at 0 and 0.5 hr. (B) Myonecrosis is estimated by the plasma CK activity, 3 hr after i.m. venom injection (50 pg). Bars represent the mean + S.D. of six mice. NI, nonimmunized animals; I, immunized animals: PBS, animals injected with PBS alone. The difference between NI and I is significant (P < 0.05). (C) Hemorrhage is estimated by the diameter of the skin lesion. 2 hr after i.d. venom injection (15 pg). Bars represent the mean f S.D. of six mice. NI, Nonimmunized animals; I. immunized
animals. The difference between NI and I is significant (P i 0.05).
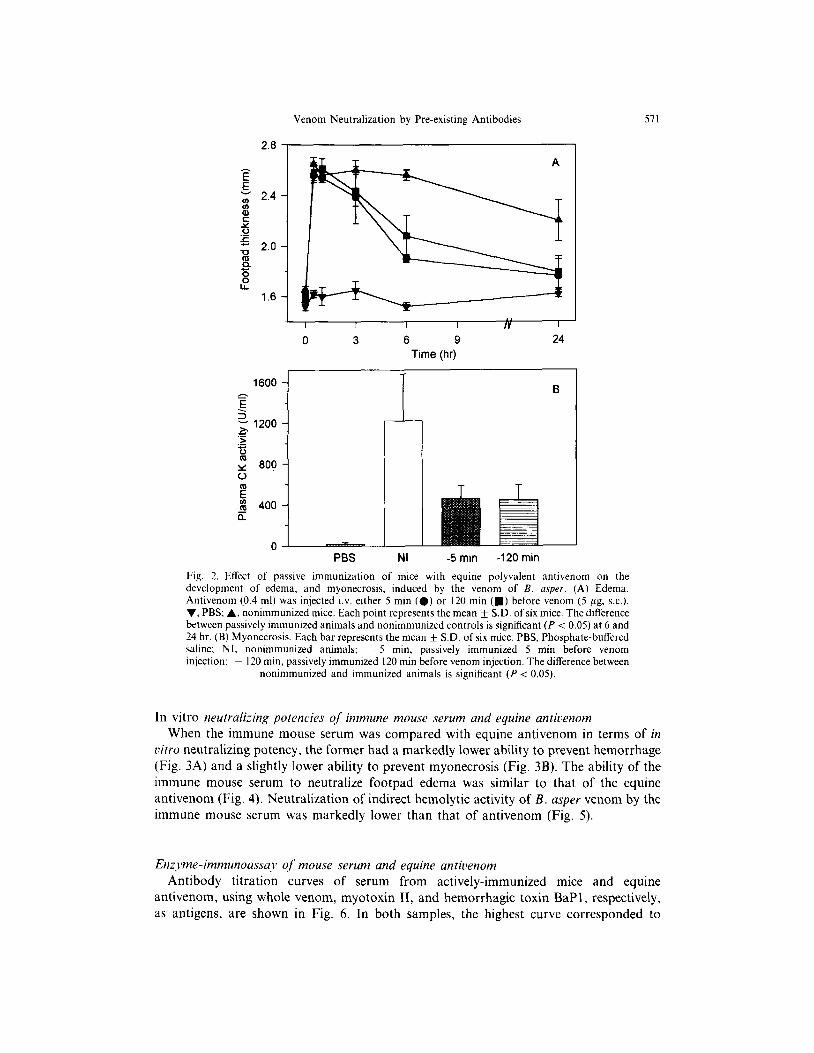
Venom Neutrahzation by Preexisting Antibodies 571
2.8 ,
A
E- k 3 2.4 -
2 a r g 2.0- lo a 5 2
1.6 -
1600 h E 3
s 1200 2 ‘ti cu SO!, t:
E 3 400 a
I I I I N I
r
0 3 6 9 Time (hr)
7
24
B
--
PBS -5 min -120 min
Fig. 2. EtTect of passive immunization of mice with equine polyvalent antivenom on the development of edema, and myonecrosis, induced by the venom of B. osper. (A) Edema. Antivenom (0.4 ml) was injected i.v. either 5 min (0) or 120 min (m) before venom (5 pg. s.c.). v, PBS; A, nonimmunized mice. Each point represents the mean f S.D. of six mice. The difference between passively immunized animals and nonimmunized controls is significant (P < 0.05) at 6 and 24 hr. (B) Myonecrosis. Each bar represents the mean + S.D. of six mice. PBS, Phosphate-buffered saline; NI, nonimmunized animals; - 5 min, passively immunized 5 min before venom injection: - 120 min. passively immunized 120 min before venom injection. The difference between
nonimmunized and immunized animals is significant (P i 0.05).
In vitro neutralizing potencies of immune mouse serum and equine antivenom When the immune mouse serum was compared with equine antivenom in terms of in
vitro neutralizing potency, the former had a markedly lower ability to prevent hemorrhage (Fig. 3A) and a slightly lower ability to prevent myonecrosis (Fig. 3B). The ability of the immune mouse serum to neutralize footpad edema was similar to that of the equine antivenom (Fig. 4). Neutralization of indirect hemolytic activity of B. asper venom by the immune mouse serum was markedly lower than that of antivenom (Fig. 5).
Enzyme-immunoassay of mouse serum and equine antivenom Antibody titration curves of serum from actively-immunized mice and equine
antivenom, using whole venom, myotoxin II, and hemorrhagic toxin BaPl, respectively, as antigens, are shown in Fig. 6. In both samples, the highest curve corresponded to

572 A. RUCAVADO and B. LOMONTE
anti-whole venom. Using this curve as an internal point of reference, the titration curve of anti-myotoxin in the mouse serum was significantly higher than the curve of anti-BaPl, at all dilutions above background (Fig. 6A). On the other hand, in the equine antivenom, the anti-myotoxin curve was lower, nearly superimposed to the anti-BaPl curve (Fig. 6B).
DISCUSSION
The ability of pre-existing antibodies to neutralize locally acting toxins of B. asper venom was investigated. Hemorrhage, myonecrosis, and edema induced by the venom were markedly reduced in mice that had been actively immunized (during approx. 2 months), although none of these effects was completely abolished.
The venom effect best neutralized in actively immunized mice was myotoxicity, followed by hemorrhage, and edema. Hemorrhage would have been expected to be neutralized more efficiently than myotoxicity, since several in vitro neutralization studies have shown a much higher potency of Costa Rican antivenoms against hemorrhage than against myonecrosis (Gutierrez et al., 1981, 1985, 1988b). In agreement with earlier results, the
= 600 E 2 500
f 400 ‘Ei y” 300 0 m El
200
$ 100
0
0.06 0.12 0.18 0.24
Serum/venom (mUmg)
B
I I I T 0.0 0.5 1.0 1.5 2.0
Serum/venom (mllmg)
Fig. 3. 1n ritm neutralizing potencies of immune mouse serum and equine antivenom against the hemorrhagic (A) and myotoxic (B) effects of B. asper venom in mice. Each point represents the mean + S.D. of five mice. n , Immune mouse serum; 0, equine antivenom. The difference between equine antivenom and immune mouse serum is significant (P < 0.05) at all points in (A), and at
0.5 and 2 ml/mg in (B).
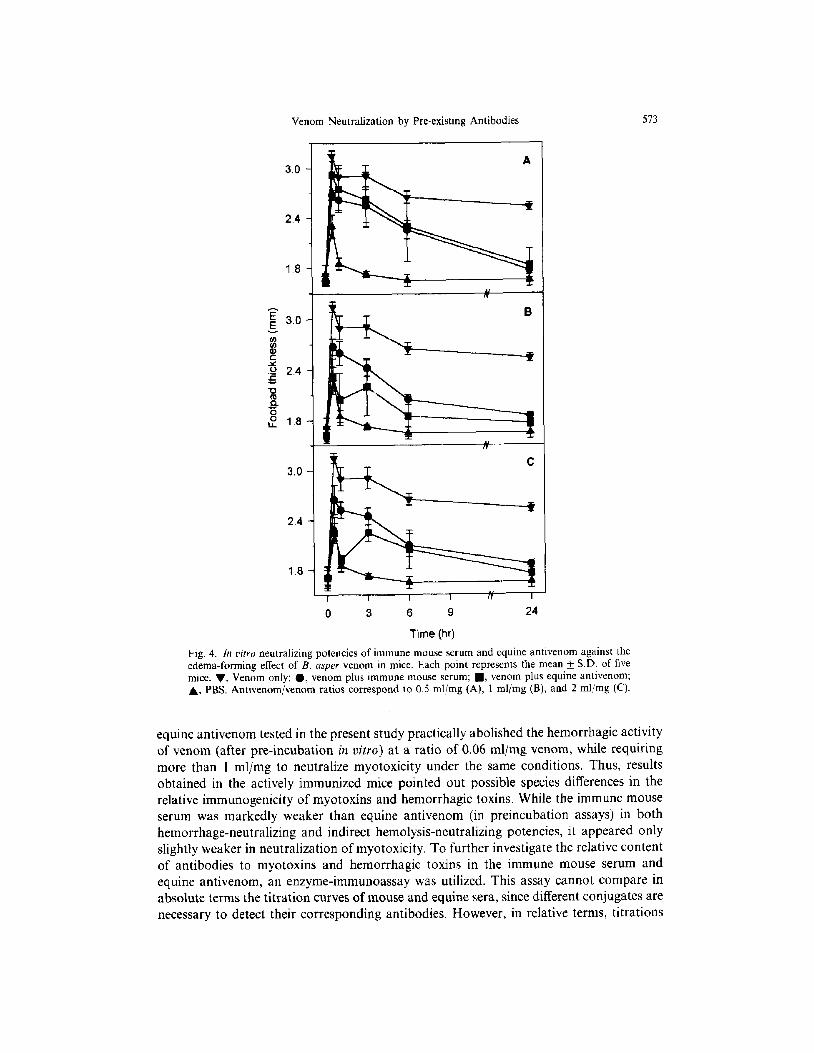
Venom Neutralization by Pre-existing Antibodies 573
1.8
2.4
1.8
I I I I n I
0 3 6 9 24
Time (hr)
Fig. 4. In vitro neutralizing potencies of immune mouse serum and equine antivenom against the edema-forming effect of B. asper venom in mice. Each point represents the mean k SD. of five mice. v, Venom only; 0, venom plus immune mouse serum; n , venom plus equine antivenom; A, PBS. Antivenom/venom ratios correspond to 0.5 ml/mg (A), I ml/mg (B), and 2 ml/mg (C).
equine antivenom tested in the present study practically abolished the hemorrhagic activity of venom (after pre-incubation in uitro) at a ratio of 0.06 ml/mg venom, while requiring more than 1 ml/mg to neutralize myotoxicity under the same conditions. Thus, results obtained in the actively immunized mice pointed out possible species differences in the relative immunogenicity of myotoxins and hemorrhagic toxins. While the immune mouse serum was markedly weaker than equine antivenom (in preincubation assays) in both hemorrhage-neutralizing and indirect hemolysis-neutralizing potencies, it appeared only slightly weaker in neutralization of myotoxicity. To further investigate the relative content of antibodies to myotoxins and hemorrhagic toxins in the immune mouse serum and equine antivenom, an enzyme-immunoassay was utilized. This assay cannot compare in absolute terms the titration curves of mouse and equine sera, since different conjugates are necessary to detect their corresponding antibodies. However, in relative terms, titrations
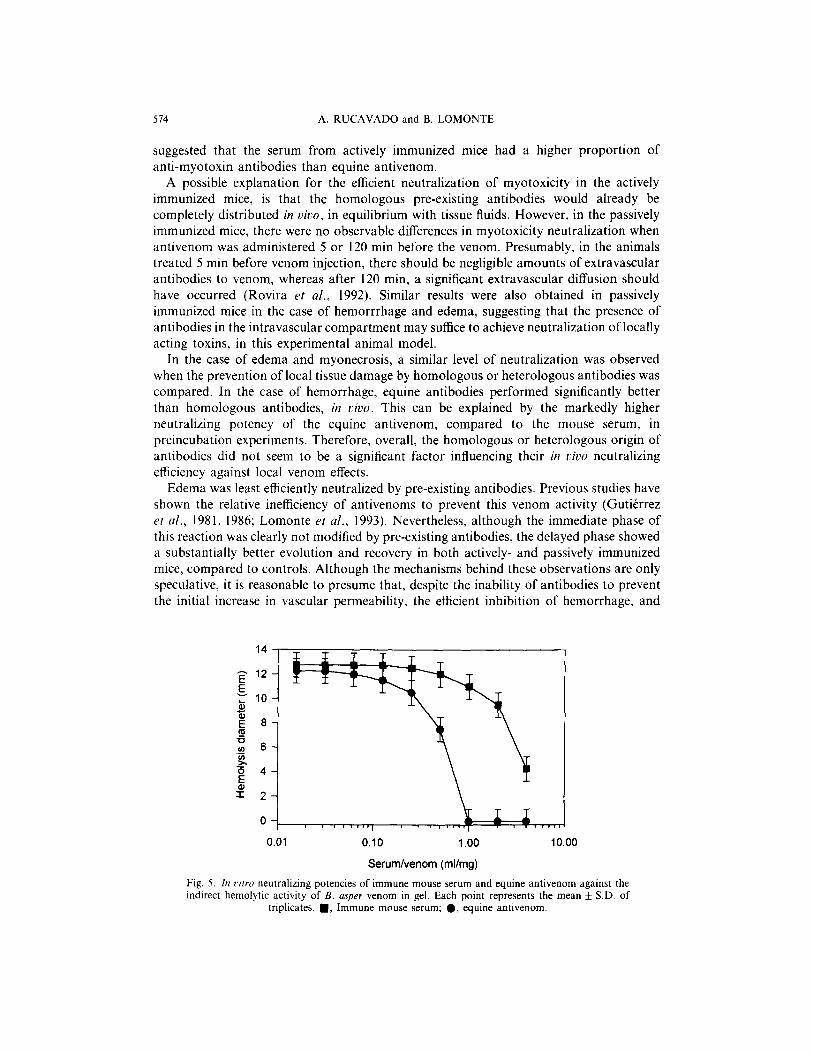
514 A. RUCAVADO and B. LOMONTE
suggested that the serum from actively immunized mice had a higher proportion of anti-myotoxin antibodies than equine antivenom.
A possible explanation for the efficient neutralization of myotoxicity in the actively immunized mice, is that the homologous pre-existing antibodies would already be completely distributed in duo, in equilibrium with tissue fluids. However, in the passively immunized mice, there were no observable differences in myotoxicity neutralization when antivenom was administered 5 or 120 min before the venom. Presumably, in the animals treated 5 min before venom injection, there should be negligible amounts of extravascular antibodies to venom, whereas after 120 min, a significant extravascular diffusion should have occurred (Rovira et al., 1992). Similar results were also obtained in passively immunized mice in the case of hemorrrhage and edema, suggesting that the presence of antibodies in the intravascular compartment may suffice to achieve neutralization of locally acting toxins, in this experimental animal model.
In the case of edema and myonecrosis, a similar level of neutralization was observed when the prevention of local tissue damage by homologous or heterologous antibodies was compared. In the case of hemorrhage, equine antibodies performed significantly better than homologous antibodies, in oit~o. This can be explained by the markedly higher neutralizing potency of the equine antivenom, compared to the mouse serum, in preincubation experiments. Therefore, overall, the homologous or heterologous origin of antibodies did not seem to be a significant factor influencing their in citlo neutralizing efficiency against local venom effects.
Edema was least efficiently neutralized by pre-existing antibodies. Previous studies have shown the relative inefficiency of antivenoms to prevent this venom activity (Gutitrrez et al., 1981, 1986; Lomonte et al., 1993). Nevertheless, although the immediate phase of this reaction was clearly not modified by pre-existing antibodies, the delayed phase showed a substantially better evolution and recovery in both actively- and passively immunized mice, compared to controls. Although the mechanisms behind these observations are only speculative, it is reasonable to presume that, despite the inability of antibodies to prevent the initial increase in vascular permeability, the efficient inhibition of hemorrhage, and
0.10 1.00
Serum/venom (ml/mg)
Fig. 5. In cirro neutralizing potencies of immune mouse serum and equine antivenom against the indirect hemolytic activity of B. asper venom in gel. Each point represents the mean _+ S.D. of
triplicates. n , Immune mouse serum; 0, equine antivenom.

Venom Neutralization by Pre-existing Antibodies 575
1.5
0.3
0.0
1.5
1.2
: p 0.9 Q:
0.6
0.3
0.0
A
AA venom 0 . Mt-It cl W BaP-1
102 103 104 105 106
SERUM DILUTION
Fig. 6. Enzyme-immunoassay titration curves of immune mouse serum (A) and equine antivenom (B) against whole 19. asprr venom (A), purified myotoxin II (a), or purified hemorrhagic toxin BaPl (m). Empty symbols represent the background signal obtained with normal equine or murine sera. Each point represents the mean absorbance at 492 nm + SD. of duplicate determinations.
hence, of microvascular damage (Lomonte et al., 1994), in the immunized animals, contributed significantly to their improved resolution of edema.
In conclusion, the present results are in agreement with the generally accepted view that local tissue damage induced by snake venoms is difficult to prevent by serotherapy, since it was shown that even in a situation where high amounts of antibodies are present before venom entry, protection may not necessarily be complete. This point should be taken into account when assessing the therapeutic efficiency of antivenoms. On the other hand. the remarkable reduction in tissue damage achieved by pre-existing antibodies motivates the search for adequate immunization strategies against venoms that would hopefully protect individuals at high risk of envenomation, in the future.
Arkno\&&ements-We are grateful to Dr Josi Maria GutiCrrez for valuable discussions and review of the manuscript. and to Javier Nlfiez, Ricardo Bolaiios. Marlen Herrera, Jorge Sanabria. Rodrigo Chaves, and Johnathan Molina for their assistance. The financial support from Vicerrectoria de Investigaci&, Universidad de Costa Rica (VI-741-95-264), and the International Foundation for Science (F/1388-3F) is gratefully acknowledged.

576 A. RUCAVADO and B. LOMONTE
REFERENCES
Bolaiios, R. and Cerdas, L. (1980) Production y control de sueros antioftdicos en Costa Rica. Boln. Of. Sanit. Punam. 88, 184196.
Cardoso, J. L. C., Fan, H. W., Franca, F. 0. S., Jorge, M. T., Leite, R. P., Nishioka, S. A., Avila, A., Sano-Martins, I. S., Tomy, S. C., Santoro, M. L., Chudzinski, A. M., Castro, S. C. B., Kamiguti, A. S., Kelen, E. M. A., Hirata, M. H., Mirandola, R. M. S., Theakston, R. D. G. and Warrell, D. A. (1993) Randomized comparative trial of three antivenoms in the treatment of envenoming by lance-headed vipers (Bofhrops jararaca) in Sao Paulo. Bra:il. Q. J. Med. 86, 315-325.
Gutierrez, J. M. and Lomonte, B. (1989) Local tissue damage induced by Bothrops snake venoms: a review. Memorias Inst. Butantan 51, 211-223.
Gutierrez, J. M., Chaves. F., Bolafios, R., Cerdas, L., Rojas, E., Arroyo, 0. and Portilla, E. (1981) Neutralization de 10s efectos locales de1 veneno de Bofhrops asper por un antiveneno polivalente. Toxicon 19, 493-500.
Gutierrez. J. M., Gene: J. A., Rojas, G. and Cerdas, L. (1985) Neutralization of proteolytic and hemorrhagic activities of Costa Rican snake venoms by a polyvalent antivenom. Toxicon 23, 887-893.
Gutierrez, J. M., Rojas. G., Lomonte, B.. Gene. J. A. and Cerdas, L. (1986) Comparative study of the edema-forming activity of Costa Rican snake venoms and its neutralization by a polyvalent antivenom. Camp. Biochem. Physiol. 85C, 171-175.
Gutierrez, J. M., Rojas, G. and Cerdas, L. (1987) Ability of a polyvalent antivenom to neutralize the venom of Lachesis muta melanocephala, a new Costa Rican subspecies of the bushmaster. Toxicon 25, 713-720.
Gutitrrez, J. M., Avila. C., Rojas, E. and Cerdas, L. (1988) An alternative in vitro method for testing the potency of the polyvalent antivenom produced in Costa Rica. Toxicon 26, 411413.
Gutitrrez, J. M., Chaves, F., Rojas, E., Elizondo, J., Avila, C. and Cerdas, L. (1988) Production of monovalent anti-Bofhrops asper antivenom: development of immune response in horses and neutralizing ability. Reu. Biol. Trap. 36, 51 t-517.
Gutierrez, J. M., Rojas. G., Lomonte, B., Gene, J. A., Chaves, F., Alvarado, J. and Rojas, E. (1990) Standardization of assays for testing the neutralizing ability of antivenoms (Letter). Toxicon 28, 1127-l 129.
Gutierrez, J. M., Romero. M., Diaz, C., Borkow, G. and Ovadia, M. (1995) Isolation and characterization of a metalloproteinase with weak hemorrhagic activity from the venom of the snake Bofhrops asper (terciopelo). To.uicon 33, 19-29.
Homma, M. and Tu. A. T. (1970) Antivenin for the treatment of local tissue damage due to envenomation by Southeast Asian snakes: ineffectiveness in the prevention of local tissue damage in mice after envenomation. Am. J. Trap. Med. Hyg. 19, 880-884.
Kondo, H., Kondo, S., Ikezawa. H., Murata, R. and Ohsaka, A. (1960) Studies on the quantitative method for determination of hemorrhagic activity of Habu snake venom. Jpn. J. Med. Sci. Biol. 13, 43-51.
Lomonte. B. and Gutierrez, J. M. (1989) A new muscle damaging toxin, myotoxin II, from the venom of the snake Bothrops asper (terciopelo). Toxicon 27, 725-733.
Lomonte, B., Gutierrez, J. M., Rojas, G. and Calderon, L. (1991) Quantitation by enzyme-immunoassay of antibodies against Bothrops myotoxins in four commercially-available antivenoms. Toxicon 29, 695-702.
Lomonte, B., Tarkowski, A. and Hanson, L. A. (1993) Host response to Bothrops asper snake venom: analysis of edema formation, inflammatory cells, and cytokine release in a mouse model. Inflammation 17, 93-10s.
Lomonte, B., Lundgren, J., Johansson, B. and Bagge, U. (1994) The dynamics of local tissue damage induced by Bothrops asper snake venom and myotoxin II on the mouse cremaster muscle: an intravital and electron microscopic study. Toxicon 32, 41-55.
Nishioka, S. A. and Silveira, P. V. P. (1992) A clinical and epidemiologic study of 292 cases of lance-headed viper bite in a Brazilian teaching hospital. Am. J. Trop. Med. Hyg. 47, 805-810.
Ohsaka, A. (1979) Hemorrhagic, necrotizing and edema-forming effects of snake venoms, In: Handbook of Experimenfal Pharmacology, Snake Venoms, Vol. 52, pp. 48&546 (Lee, C. Y., Ed). Berlin: Springer-Verlag.
Ownby, C. L., Woods, W. M. and Odell, G. V. (1979) Antiserum to myotoxin from prairie rattlesnake (Crotalus viridis uiridis) venom. Toxicon 17, 373-380.
Ownby, C. L., Odell, G. V., Woods, W. M. and Colberg, T. R. (1983) Ability of antiserum to myotoxin a from prairie rattlesnake (Crotalus tliridis Gridis) venom to neutralize local myotoxicity and lethal effects of myotoxin a and homologous crude venom. Toxicon 21, 3545.
Ownby. C. L., Colberg, T. R., Claypool, P. L. and Odell, G. V. (1984) In oivo test of the ability of antiserum to myotoxin a from prairie rattlesnake (Crotahu t?ridis uiridis) venom to neutralize local myonecrosis induced by myotoxin a and homologous crude venom. Toxicon 22, 99-105.
Ownby, C. L., Colberg, T. R. and Odell, G. V. (1986) In oioo ability of antimyotoxin a serum plus poly- valent (Crotalidae) antivenom to neutralize prairie rattlesnake (Crotalus Gridis Gridis) venom. Toxicon 24, 197-200.
Reid, H. A., Thean, P. C. and Martin, W. J. (1963) Specific antivenene and prednisone in viper-bite poisoning: controlled trial. Br. Med. J. II, 1378-1380.

Venom Neutralization by Preexisting Antibodies 577
Rovira. M. E., Carmona, E. and Lomonte, B. (1992) Immunoenzymatic quantitation of antibodies to Eothrops asper myotoxins after polyvalent antivenom administration in mice. Bruz. J. Med. Biol. Res. 25, 23-33.
Russell, F. E., Ruzic, N. and Gonzalez, H. (1973) Effectiveness of antivenin (Crotahdae) polyvalent following injection of Croralus venom. Toxicon 11, 461464.
Warrell, D. A., Greenwood, B. M., Davidson, N. McD.. Ormerod, L. D. and Prentice, C. R. M. (1976) Necrosis, haemorrhage and complement depletion following bites by the spitting cobra (Naja nigricollis). Q. J. Med. 45. I-22.
Copyright © 2022 FDOKUMEN




















