
Características biológicas e inmunológicas del veneno de Bothrops cotiara (Serpentes: Viperidae)
Upload
independentCategory
view
0download
0
ilable at ScienceDirect
Toxicon 54 (2009) 988–997
Contents lists ava
Toxicon
journal homepage: www.elsevier .com/locate/ toxicon
Inflammation induced by Bothrops asper venomq
Catarina Teixeira a,*, Yara Cury b, Vanessa Moreira a, Gisele Picolo b, Fernando Chaves c
a Laboratorio de Farmacologia, Instituto Butantan, Av. Vital Brazil, 1500, CEP 05503-900 Sao Paulo, SP, Brazilb Laboratorio de Fisiopatologia, Instituto Butantan, Sao Paulo, Brazilc Facultad de Microbiologıa, Instituto Clodomiro Picado, Universidad de Costa Rica, San Jose, Costa Rica
a r t i c l e i n f o
Article history:Available online 6 June 2009
KeywordsBothrops asperVenomInflammationPainToxins
DOI of original article: 10.1016/j.toxicon.2009.03q
Please note that this article was previously pregular issue [Toxicon 54 (1) pp. 67–76], while it shothis special issue, in which it is now published. Plarticle for citation. The Publisher regrets any inconvcause.
* Corresponding author. Tel.: þ55 11 3726 7222; faE-mail address: [email protected] (C. Teixeira).
0041-0101/$ – see front matter � 2009 Elsevier Ltddoi:10.1016/j.toxicon.2009.05.026
a b s t r a c t
Inflammation is a major characteristic of envenomation by snakes from viperine andcrotaline species. Bothrops asper snake venom elicits, among other alterations,a pronounced inflammatory response at the site of injection both in humans and exper-imental animals. This review describes the current status of our understanding of theinflammatory reaction, including pain, triggered by B. asper venom. The experimentalstudies on the action of this venom as well as the complex network of chemical mediatorsinvolved are summarized. Moreover, aspects of the molecular mechanisms orchestratingthis important response to envenomation by B. asper are presented. Considering thatisolated toxins are relevant tools for understanding the actions of the whole venom,studies dealing with the mechanisms of inflammatory and nociceptive properties ofphospholipases A2, a metalloproteinase and serine proteinases isolated from B. aspervenom are also described.
� 2009 Elsevier Ltd. All rights reserved.
1. Inflammation and inflammatory pain. Generalconcepts
Inflammation is an adaptive process whereby tissues ofthe body respond to injury. Regardless of the cause,inflammation is a stereotyped response for restoringhomeostasis, setting the stages for healing and reconsti-tution of damaged tissue. The acute inflammatory responseto injury involves functional alterations of microvessels,leading to the coordinated delivery of blood components(plasma and leukocytes) to the site of injury, followed byphagocytosis by competent cells, and local pain. Changes invascular flow and caliber begin early after injury and
.019.ublished in a recentuld have been part ofease use the originalenience that this may
x: þ55 11 3726 1505.
. All rights reserved.
develop at varying rates. After a transient vasoconstrictionof arterioles, vasodilation occurs. These events are followedby increased permeability of microvasculature, mainly thepostcapillary venules, leading to the escape of a protein-rich fluid into the extravascular tissue, which forms theoedema, accompanied by pain. After tissue injury,numerous inflammatory mediators, which include che-mokines, cytokines, vasoactive amines and eicosanoids, areproduced in the circulation or in close proximity to theinjury by endothelial cells and resident cells, mainly mastcells, tissue macrophages and dendritic cells. These medi-ators are key elements orchestrating all the inflammatoryevents in response to injury. In parallel, circulating leuko-cytes, initially neutrophils, migrate to the site of injuryunder the influence of chemotactic agents (Garcia-Lemeet al., 1973; Ryan and Majno, 1977; Granger and Kubes,1994). This process involves sequential capture on, rollingalong and firm adhesion to the microvascular endothelium,followed by transmigration through the vessel wall andfurther migration into extravascular tissue (Garcia-Lemeet al., 1973; Granger and Kubes, 1994). All the steps in therecruitment cascade are regulated by cell-adhesion

C. Teixeira et al. / Toxicon 54 (2009) 988–997 989
molecules (CAMs) on both leukocyte and endothelial cells,and the different subsets of CAMs are responsible for thedifferent steps in extravasation (Carlos and Harlan, 1994).The sequence of expression and function of CAMs is regu-lated by a range of inflammatory mediators (Hubbard andRothlein, 2000; Kaplanski et al., 2003). Thereafter, phago-cytosis of offending agents occurs, followed by the releaseof microbicidal products, like reactive oxygen species, intothe phagolysosome and the killing or degradation ofingested material (Rabinovitch and De Stefano, 1973).Resolution of inflammation and the repair phase involvedecay of the pro-inflammatory and release of anti-inflam-matory chemical mediators, mainly by tissue resident andrecruited macrophages as well as apoptosis of granulocytesand subsequent clearance by phagocytes. This episode iscompleted by their exit through the lymphatic system,which removes fluid and cell debris from the interstitialcompartment (Serhan and Savill, 2005). However, if theacute events do not succeed in neutralizing the injuriousstimulus, the inflammatory process persists and acquiresnew characteristics and a chronic inflammatory stateensues, involving the formation of granulomas and tertiarylymphoid tissues. The inflamed area is infiltrated by cellssuch as mononuclear phagocytes, lymphoid cells, fibro-blasts, and granulocytes in some circumstances, withchanges occurring in the structure of the surroundingconnective tissue (Medzhitov, 2008).
Inflammation is frequently associated with pain andhyperalgesia (McMahon et al., 2005). The physiologicalfunction of pain is to alert the brain of deregulatedhomeostasis providing information about the stimulicausing tissue injury, allowing the body to protect itselffrom greater damage. The pain phenomenon is originatedin peripheral sensory nerve fibers through activation ofnociceptors that relay information about noxious stimulifrom the periphery to the central nervous system (Tortoraand Grabowski, 2002; Cummins et al., 2007).
Peripheral pain pathways are activated by a range ofstimuli including inflammatory mediators that are involvedin the genesis, persistence, and severity of pain. The initialrecognition of injury leads to production and/or release ofmediators such as bradykinin, proteases, histamine, sero-tonine (5-HT), nitric oxide (NO), prostanoids, neuro-trophins, cytokines, ATP, protons, and other mediators thatmay act on sensory fibers by two different ways: a) elicitingpain by direct activation of nociceptors (e.g. protons, cyto-kines, and nerve growth factor), and/or b) sensitizingprimary afferent fibers to other key mediators and stimuli,reducing their thresholds so that they now respond tolower intensity stimuli (a phenomenon known as hyper-algesia) (e.g. prostaglandins, in particular prostaglandin E2-PGE2). In addition to those mediators, and as a consequenceof nerve terminal stimulation, the primary afferent neuronscontribute to the direct release of neuropeptides such assubstance P (SP) and calcitonin gene-related peptide(CGRP), or indirect (reflex) release of catecholaminesthrough the sympathetic efferent, leading to neurogenicinflammation (Haddad, 2007; Harvey and Dickenson,2008; Rittner et al., 2008a,b). Pain in response to lighttouch/pressure stimuli, which normally are perceived asinnocuous, may also occur in the presence of inflammation
and chronic pain states, a process known as allodynia(Woolf, 2004).
In addition to peripheral sensitization, many centralalterations can be observed after both inflammatory andneuropathic conditions. These alterations include changesin the expression of receptors (nociceptive plasticity),central spinal hyperexcitability (central sensitization) andalterations in descending control from the midbrain (Har-vey and Dickenson, 2008). The major excitatory neuro-transmitter involved in the processing of nociceptiveinformation in the dorsal horn of the spinal cord is gluta-mate (Schaible and Richter, 2004). During high frequencyfiring of C fibers, glutamate and its coreleased transmitterSP cooperate to allow NMDA receptor activation, resultingin the wind-up of dorsal horn (Harvey and Dickenson,2008). Increments in brain-derived neurotrophic factor(BDNF), a synaptic neuromodulator, activation of theMAPK/pCREB cascade and the induction of cyclo-oxygenase-2 (COX-2) in dorsal horn neurons alsocontribute to central sensitization (Woolf and Salter, 2000).Spinal glial cells (astrocytes/microglia), through the releaseof a variety of neuroactive substances, have been alsoimplicated in exaggerated pain states induced by periph-eral inflammation and nerve trauma (Wieseler-Frank et al.,2004).
Inflammatory mediators induce their effects by actingdirectly on ion channels or on pronociceptive receptorsthat, once activated, lead to activation of multiple proteinkinase pathways, inducing a rapid posttranslational regu-lation of ion channels (Cheng and Ji, 2008). The activationof these channels (e.g. acid-sensing ion channels – ASICsand the heat and capsaicin receptor – TRPV1) induces theiropening and the entrance of sodium and calcium ions onthe nociceptor, depolarizing their membranes. In addition,activation of a variety of protein kinases causes the phos-phorylation of voltage-gated ion channels. At specific Naþ
threshold, voltage-gated sodium channels are activated,initiating the propagation of action potentials (Woolf,2004; Giordano, 2005). Of particular importance to pain ininflammatory conditions are the sodium tetrodotoxin(TTX)-resistant channels NaV 1.8 and 1.9, that are expressedalmost exclusively in small, unmyelinated fibers, and TTX-sensitive NaV 1.7 channels, expressed in both sensory andsympathetic neurons (Cummins et al., 2007; Abrahamsenet al., 2008; Rittner et al., 2008a). In addition to sodiumchannels, high voltage-gated calcium channels CaV1 (L-type), CaV2.1 (P/Q type) and CaV2.2 (N-type) and lowvoltage-activated CaV3 (T-type) channels have been impli-cated in nociceptive transmission, including inflammatoryconditions, being involved in neuronal transmitter releaseand central sensitization (Vanegas and Schaible, 2000;Saegusa et al., 2001, 2002; Kim et al., 2003; Luvisetto et al.,2006).
2. Inflammation induced by Bothrops asper venom
2.1. Oedema
Inflammation is a major characteristic of envenomationby snakes from viperine and crotaline species (Rosenfeld,1971; Sawai, 1980). B. asper snake venom (BaV), among

C. Teixeira et al. / Toxicon 54 (2009) 988–997990
other alterations, elicits a pronounced inflammatoryresponse at the site of injection both in humans andexperimental animals (Rosenfeld, 1971; Bolanos, 1984;Gutierrez et al., 1986a). Prominent local oedema isa common clinical finding in snake-bitten victims (Ohsaka,1979; Otero et al., 2002), which frequently causes ischemiaand neural compression, leading to permanent tissue loss,disability or amputation (Ribeiro and Jorge, 1997; Jorgeet al., 1999). This is a relevant fact, since antivenom therapydoes not reverse the local manifestations of envenomation,and experimental studies have shown that oedemainduced by BaV is not well neutralized even if antivenom isincubated with venom prior to injection into animals(Gutierrez et al., 1981, 1986b).
The ability of BaV to experimentally generate inflam-mation was first reported by Gutierrez et al. (1980) whoshowed that injection of BaV into mouse foot paw causedoedema with a rapid onset but sustained up to 24 h, andleukocyte infiltration upon intramuscular injection intomice. Later, Lomonte et al. (1993) showed that injection ofa lower concentration of BaV into mouse foot paw induceda rapid and transient oedema. This local response was notrelated to the haemorrhagic activity of venom, since it wasobserved after injection of pre-neutralized venom, devoidof haemorrhagic activity. Subsequently, studies on thedynamics of local tissue damage induced by BaV on mousecremaster muscle showed plasma leakage from smallvenules and adjoining capillary segments after about 2 minof exposure to venom and stasis in the microcirculationwith dense packing of erythrocytes in capillaries and smallvenules (Lomonte et al.,1994). In addition, the ability of BaVto contract collecting lymphatic vessels of mouse mesen-tery associated with a reduction of vessel lumen anda halting in the flow of lymph was recently demonstrated(Mora et al., 2008). Since BaV was cytotoxic for smoothmuscle cells in culture, these authors suggested that thealterations observed in the lymphatics are due to venomeffect on smooth muscle cells of the lymphatic vessel wall.Thus, the effect of BaV on lymphatic system may be a rele-vant mechanism for the pathogenesis of oedema in enve-nomed tissues. Furthermore, a higher oedematogenicactivity of venom from newborn B. asper snake uponintramuscular injection, when compared with venomsfrom adults, was demonstrated (Chaves et al., 1992). Thehigher potency of venom from newborns was attributed tovariations between venom components from adult andnewborn snakes evidenced by electrophoresis and, morerecently, by proteomic analysis (Alape-Giron et al., 2008),although the components responsible for the oedemato-genic activity of venom have been identified only toa partial extent.
2.2. Inflammatory cell infiltrate
Besides oedema there is a considerable cellularcomponent in the inflammatory reaction induced by BaV.The time-course and composition of the inflammatoryleukocyte infiltrate induced by BaV was first described afterinjection of this venom into mouse gastrocnemius muscle(Gutierrez et al., 1986a). A scarce infiltrate by 6 h, whichmarkedly increased from 24 up to 72 h after venom
injection was demonstrated. The infiltrate was observedwithin necrotic muscle cells and in the interstitial space,and consisted predominantly of polymorphonuclearleukocytes at 6 and 24 h, but at 48 and 72 h, mononuclearcells were the major cell type involved. Similarly, in themouse footpad model, subcutaneously injected BaV evokedan early neutrophil influx followed by a late mononuclearcell infiltrate, accumulating in the muscle and subdermaltissue (Lomonte et al., 1993). In this experimental model,the venom also induced a rapid inversion in the ratio ofcirculating neutrophils to lymphocytes in peripheral blood,which normalized up to 12 h, suggesting that leukocyteinflux into tissues induced by this venom depends on itsearly effect on bone marrow compartment. Later, investi-gations on the mechanisms involved in leukocyte recruit-ment into the site of venom injection have revealed theability of BaV to stimulate neutrophils and endothelial cellsto express the adhesion molecules L-selectin, LFA-1, ICAM-1, PECAM-1 and CD-18 (Zamuner et al., 2005). Thesemolecules are responsible for rolling, firm adhesion andtransmigration events associated with neutrophil locomo-tion across endothelium towards the inflamed area (Fig. 1).This effect was attributed to the direct action of venom onvessel walls and leukocytes as well as to the action ofcytokines and eicosanoids released upon venom injection.However, despite the relevance of leukocytes in inflam-mation, the role of each cell type in development of theinflammatory response to venom needs to be betterunderstood.
Leukocytes play essential roles in the host defenseagainst offending agents (Sherwood and Toliver-Kinsky,2004). On exposure to a pathogen, these cells respond withphagocytosis followed by generation and release ofmicrobicidal products (Rohn et al., 1999). In this regard,experimental studies showed that BaV induces phagocy-tosis and triggers microbicidal functions of peritonealleukocytes in vivo with consequent increment in theproduction of hydrogen peroxide and nitric oxide bymacrophages followed by the generation of toxic perox-ynitrites (Zamuner et al., 2001) (Fig. 1). Moreover, a markedformation of nitrotyrosine-containing proteins was alsoobserved in macrophages at 12–48 h after BaV injection,leading the authors to suggest that locally generated reac-tive oxygen- and nitrogen-derived species could potentiatetissue damage caused by BaV (Zamuner et al., 2001).Subsequent studies, however, demonstrated that pretreat-ment of mice with antioxidant compounds and inhibitorsof nitric oxide synthase did not reduce myonecrosis andhemorrhage induced by BaV (Chaves et al., 2006).
Neutrophils are the first cells migrating into theinflamed area and play a critical role in elimination ofinjurious agents. The role of these cells in the local patho-logical effects induced by BaV was investigated byrendering mice neutropenic through the administration ofrat monoclonal antibodies against mouse granulocytes.Depletion of neutrophils did not affect the extent of myo-necrosis and hemorrhage, but reduced skeletal muscleregeneration after myonecrosis induced by BaV anda myotoxic phospholipase A2 (PLA2) (Teixeira et al., 2003).Neutrophil depletion may jeopardize the adequate removalof necrotic debris and the recruitment of macrophages
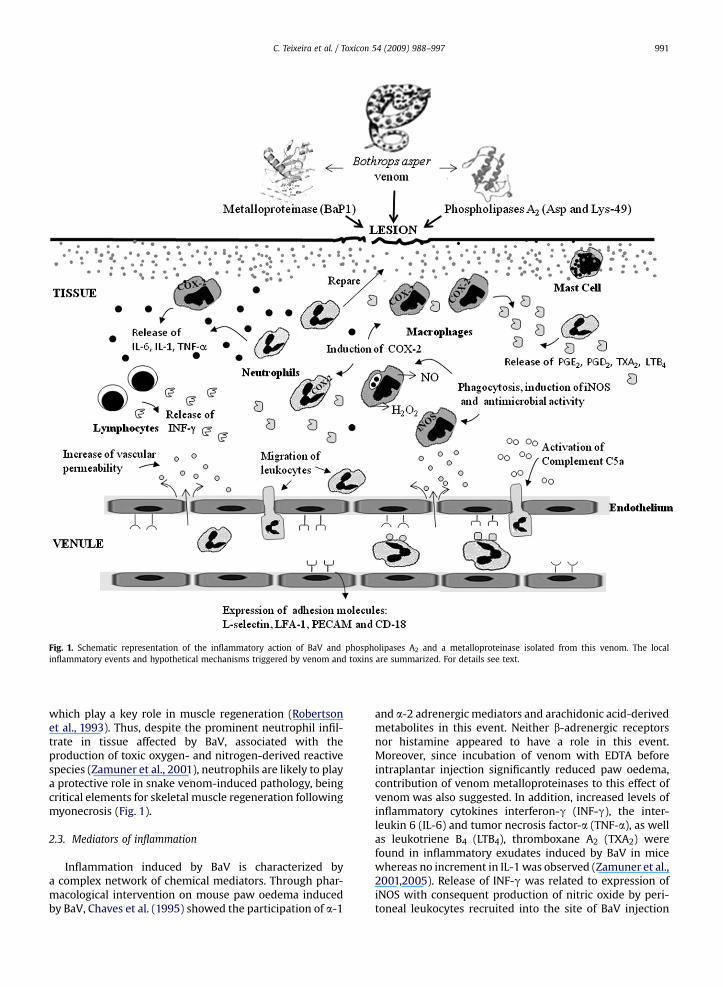
Fig. 1. Schematic representation of the inflammatory action of BaV and phospholipases A2 and a metalloproteinase isolated from this venom. The localinflammatory events and hypothetical mechanisms triggered by venom and toxins are summarized. For details see text.
C. Teixeira et al. / Toxicon 54 (2009) 988–997 991
which play a key role in muscle regeneration (Robertsonet al., 1993). Thus, despite the prominent neutrophil infil-trate in tissue affected by BaV, associated with theproduction of toxic oxygen- and nitrogen-derived reactivespecies (Zamuner et al., 2001), neutrophils are likely to playa protective role in snake venom-induced pathology, beingcritical elements for skeletal muscle regeneration followingmyonecrosis (Fig. 1).
2.3. Mediators of inflammation
Inflammation induced by BaV is characterized bya complex network of chemical mediators. Through phar-macological intervention on mouse paw oedema inducedby BaV, Chaves et al. (1995) showed the participation of a-1
and a-2 adrenergic mediators and arachidonic acid-derivedmetabolites in this event. Neither b-adrenergic receptorsnor histamine appeared to have a role in this event.Moreover, since incubation of venom with EDTA beforeintraplantar injection significantly reduced paw oedema,contribution of venom metalloproteinases to this effect ofvenom was also suggested. In addition, increased levels ofinflammatory cytokines interferon-g (INF-g), the inter-leukin 6 (IL-6) and tumor necrosis factor-a (TNF-a), as wellas leukotriene B4 (LTB4), thromboxane A2 (TXA2) werefound in inflammatory exudates induced by BaV in micewhereas no increment in IL-1 was observed (Zamuner et al.,2001,2005). Release of INF-g was related to expression ofiNOS with consequent production of nitric oxide by peri-toneal leukocytes recruited into the site of BaV injection

C. Teixeira et al. / Toxicon 54 (2009) 988–997992
(Zamuner et al., 2001) whereas the other cytokines andeicosanoids were associated with induction of leukocyterecruitment (Zamuner et al., 2005). In agreement withthese findings, increased levels of prostaglandin E2 andexpression of COX-2 were detected in mouse paw tissueinjected with BaV. Moreover, treatment of animals witha selective COX-2 inhibitor markedly reduced BaV-inducedoedema, indicating that arachidonic acid metabolites fromthe COX-2 pathway play a role in oedema-inducing activityof venom (Olivo et al., 2007). This is supported by recentfindings that BaV up regulated expression of COX-2 by bothmacrophage and neutrophil indicating that these cells arerelevant sources of eicosanoids in envenomed sites (Mor-eira et al., 2009) (Fig. 1). Systemic levels of inflammatorymediators are also altered during experimental localinflammatory reaction induced by BaV. Lomonte et al.(1993) have shown that injection of BaV into mouse pawalso evoked increased levels of serum IL-6, while levels ofserum TNF-a and IL-1 remained basal. The increased levelsof IL-6 in response to venom were associated with the onsetof muscle regeneration. The effect of this cytokine trig-gering neutrophil migration into tissues can also be sug-gested, since IL-6 activates maturation of neutrophils andup-regulates endothelial cell-adhesion molecules involvedin firm adhesion of leukocytes to endothelium (Hopkins,2003; Suwa et al., 2002). These findings agree with a clin-ical study reporting increased levels of circulating IL-6, IL-8and TNF-a, but not IL-1, in the first hours after B. aspersnake bite, although no significant clinical associationbetween severity of envenoming and serum cytokineconcentration was detected (Avila-Aguero et al., 2001). Inthis clinical study, serum levels of MIP-1b and Regulated onActivation Normal T cell Expressed and Secreted chemo-kine (RANTES) were also detected.
The ability of BaV to activate the complement systemwas also demonstrated. In vivo and in vitro studies showedthat BaV generates C5a fraction (Farsky et al., 2000), withconsequent role on leukocyte recruitment, since C5a andC5a des-arg fragments cause neutrophil activation, up-regulation of adhesion molecules, leukocyte emigrationand chemotaxis (Till, 1998) The importance of thecomplement system on leukocyte mobilization wasconfirmed in C5 deficient animals injected with BaV. Theseanimals exhibited reduction of about 50% on the number ofpolymorphonuclear cells in the peritoneal cavity after BaVinjection (Farsky et al., 2000). Although the mechanismsinvolved in this activation were not investigated, actions ofvenom on the complement cascade or directly on C5 frag-ment were suggested (Fig. 1). On the other hand, in vitroassays indicated that BaV per se is unable to evoke orientedcell locomotion and does not modify the intrinsic ability ofneutrophils to migrate in response to a chemotactic factor(Farsky et al., 2000).
Nitric oxide is also produced during the host responseevoked by BaV. This venom induced the synthesis of NOupon intramuscular and intraperitoneal injections in mice,mainly through induction of expression of the induciblenitric oxide synthase (iNOS) (Zamuner et al., 2001; Chaveset al., 2006) (Fig. 1). Reduction in both local footpad oedemaand the extent of leukocyte infiltrate was also observed inmice pretreated with L-NAME, an inhibitor of both
constitutive NOS and iNOS, and aminoguanidine, aninhibitor of iNOS, indicating that NO contributes to oedemaand inflammatory infiltrate induced by BaV. In addition,a protective role of NO against the lethal activity of thisvenom was demonstrated, whereas no significantinvolvement of NO in the venom-induced local pathologiceffects was observed (Chaves et al., 2006).
Additionally, it was demonstrated that BaV induces theexpression and activation of matrix metalloproteinase-2(MMP-2) in skeletal muscle in vivo, but promotes simulta-neous activation and degradation of already synthesizedproMMP-2 in cultured cells (Saravia-Otten et al., 2004). Theincreased synthesis of endogenous MMP-2 and its subse-quent activation after acute envenomation may contributeto the direct effect of venom in the affected tissues. Sincedegradation of the extracellular matrix by MMPs hasprofound effects on its function and on the behaviour ofneighbouring cells (Streuli, 1999; Sternlicht and Werb,2001), the effects of BaV on extracellular matrix may affectthe normal processes of tissue repair and muscle regener-ation and angiogenesis following envenomation. This is animportant subject to be further studied.
3. Pain induced by B. asper venom
Besides its characteristic inflammatory effects, injectionof BaV elicits pain in humans (Gutierrez, 1995; Arroyo et al.,1999). In an attempt to characterize the mechanismsinvolved in the nociceptive effects of BaV, experimentalstudies have been developed to identify the chemicalmediators and the cellular components responsible forsuch effects.
The first experimental evidence that BaV causeshyperalgesia was provided by Chacur et al. (2001). Theseauthors demonstrated that intraplantar injection of thevenom in rats significantly decreased the pain threshold ofthe animals. This characteristic hyperalgesic response wasdose- (5–15 mg/paw) and time-dependent, and was medi-ated by bradykinin, acting on B2 receptors, and leukotri-enes. In this study, intraplantar injection of BaV alsoinduced an oedematogenic response, however a directcorrelation between the inflammatory and hyper-nociceptive phenomena caused by the venom was notdetected.
Subsequently, the effectiveness of polyspecific anti-venom in neutralizing the nociceptive effects of BaV wasexperimentally investigated by Picolo et al. (2002). Poly-valent antivenom produced at Clodomiro Picado Institute,Costa Rica, did not interfere with hyperalgesia or oedemainduced by BaV, even when administered prior to enve-nomation. These findings are in accordance with previousstudies of Gutierrez et al. (1981,1986b). The lack ofneutralization of hyperalgesia and oedema induced by BaVis not attributable to the absence of neutralizing antibodiesin the antivenom, since neutralization was achieved inassays involving pre-incubation of venom and antivenom.These data confirm that the local effects, including pain,induced by BaV are only partially inhibited by commercialantivenoms. The data also point out the need to furtherinvestigate the mechanisms involved in the nociceptiveaction of the venom and also to characterize the

C. Teixeira et al. / Toxicon 54 (2009) 988–997 993
nociceptive components present in this venom in order todevelop therapeutic strategies aimed at controlling pain inenvenomation caused by BaV.
4. Inflammatory and nociceptive effects of purifiedvenom components
4.1. Phospholipases A2
Besides the role of stored and newly generated inflam-matory mediators, enzymes present in the venom, mainlyphospholipases A2 (PLA2s) and metalloproteinases, appearto play a role in inflammation and hyperalgesia induced byBaV. The study of isolated toxins has been an invaluableapproach to dissect and analyze the complex eventsleading to the development of inflammation induced bywhole venom.
Four myotoxins with phospholipase A2 structure wereisolated from BaV, named MT-I to MT-IV. Despite highhomology among these phospholipases, MT-II and MT-IVlack catalytic activity and present a Lys residue at position49, whereas MT-I and MT-III with high enzymatic activity,contain an Asp residue at 49 position (Lomonte and Car-mona, 1992; Gutierrez and Lomonte, 1995, 1997). Sincethese PLA2s comprise approximately 30% of BaV (Gutierrezand Lomonte, 1995), these toxins are likely to play a rele-vant role in the pro-inflammatory and hyperalgesic activi-ties that characterize this venom.
Regardless their catalytic activity, it has been demon-strated that PLA2s isolated from BaV induce a marked localinflammatory reaction. This reaction is characterized by anearly increase of plasma extravasation (Zuliani et al.,2005a), oedema (Gutierrez et al., 1986c; Chaves et al., 1998)and a conspicuous infiltrate of leukocytes (Zuliani et al.,2005a) followed by hyperalgesia (Chacur et al., 2003).Oedema induced by BaV PLA2s is multimediated, sincepharmacological approaches, using receptor antagonists orenzyme inhibitors, showed the involvement of serotonin,prostaglandins and alpha adrenergic receptors, similar tothe mediation observed for oedema induced by BaV.However, leukotrienes seem not to play a role in this event.In contrast to the whole venom, the oedema induced byMT-II and MT-III is mediated by histamine (Gutierrez et al.,1986c; Chaves et al., 1998). This discrepancy, observed indistinct experimental condition, might be due to differ-ences in the type of histamine antagonists used to analyzeoedema formation induced either by BaV (diphenidramine)and both PLA2s (cimetidine and pyrilamine). In addition,recent studies have demonstrated that upon local appli-cation MT-II reduced the lumen of lymphatic vessels andimpaired lymphatic flow in the mesentery of mice. Thisreproduced the effects of whole venom in the same system.Furthermore, complete elimination of the effect wasobserved when venom was preincubated with fucoidan, aninhibitor of myotoxic PLA2s (Mora et al., 2008). Therefore,the oedematogenic action of BaV myotoxic PLA2s mayresult from a combination of their effect on lymphaticvessels and their ability to release inflammatory mediatorsin the tissue.
The inflammatory effects evoked by catalytically-activeand -inactive PLA2 variants involve different signaling
pathways, since chemical modification of MT-III withp-bromophenacyl bromide, that promotes alkylation ofHis48 residue and abrogates enzymatic activity, reducedoedema (Chaves et al., 1998) and leukocyte influx into thesite of its injection (Zuliani et al., 2005a). Interestingly, theleukocyte adhesion molecules involved in MT-II and MT-III-induced leukocyte influx were similar to those recruited bythe whole venom, whereas participating endotheliumadhesion molecules differed among those PLA2s and wholevenom. In addition, despite few differences in kinetics ofrelease, MT-II (Lys49-PLA2) and MT-III (Asp49-PLA2) wereable to stimulate the production and release of inflamma-tory mediators such as IL-1 and IL-6, TNF-a, LTB4, TXA2,PGE2 and PGD2 at site of their injection as well as under invitro conditions (Zuliani et al., 2005a; Moreira et al., 2008).These effects are in accordance with those reported forwhole venom. Furthermore, up-regulation of expression ofactive COX-2 but not of COX-1, and release of arachidonicacid by MT-III were demonstrated in isolated macrophages.Hence, COX-2 is the major isoform involved in the synthesisof prostaglandins and thromboxane induced by venomPLA2s (Moreira et al., 2008). Studies have also shown thatMT-II and -III activate macrophages for microbicidal activ-ities, increasing the respiratory burst and up-regulatingphagocytosis (Zuliani et al., 2005b). Due to similarity withthe whole venom-induced effects on both COXs isoforms(Olivo et al., 2007) and microbicidal functions of macro-phages, it is likely that these PLA2s play a relevant role in thestimulatory effects of venom on inflammatory cells (Fig. 1).
The contribution of toxins present in the crude BaV tothe genesis and development of hyperalgesia was firstinvestigated by Chacur et al. (2003). These authors showedthat both MT-II and MT-III cause significant local hyper-algesia in the rat hind paw after intraplantar injection(5–20 mg/paw). The hyperalgesic response induced by bothmyotoxins was of rapid onset and similar time-course. Inaddition to hyperalgesia, the intraplantar injection of thecatalytically active MT-III caused mechanical allodynia,which was not observed after injection of the Lys49-PLA2
homologue MT-II. These results indicate that enzymaticactivity is not a strict requirement for the induction ofnociception, but is important for determination of theintensity of the nociceptive phenomenon. In addition,further studies demonstrated that enzymatic activityseems to be essential for hyperalgesia induced by thecatalytically active MT-III whereas the hyperalgesic activityof MT-II is due to the action of the C-terminal cationic/hydrophobic sequence 115–129 (Chacur et al., 2003),a molecular region also responsible for other toxic activitiesof Lys49-PLA2 homologues (Lomonte et al., 2003).
The hyperalgesic response induced by the Lys49-PLA2
seems to result from the release of several mediators,including biogenic amines, bradykinin, cytokines, prosta-glandins and sympathomimetic amines (Fig. 2), which maybe sequentially released and/or may interact for theinduction of hyperalgesia. On the other hand, only brady-kinin plays a major role in the generation of MT-III-inducedhyperalgesia. Studies on the spinal mechanisms involved inthe pain-enhancing effects of both myotoxins indicate thatspinal pro-inflammatory cytokines, nitric oxide, and pros-tanoids are involved in the pain-enhancing effects of the
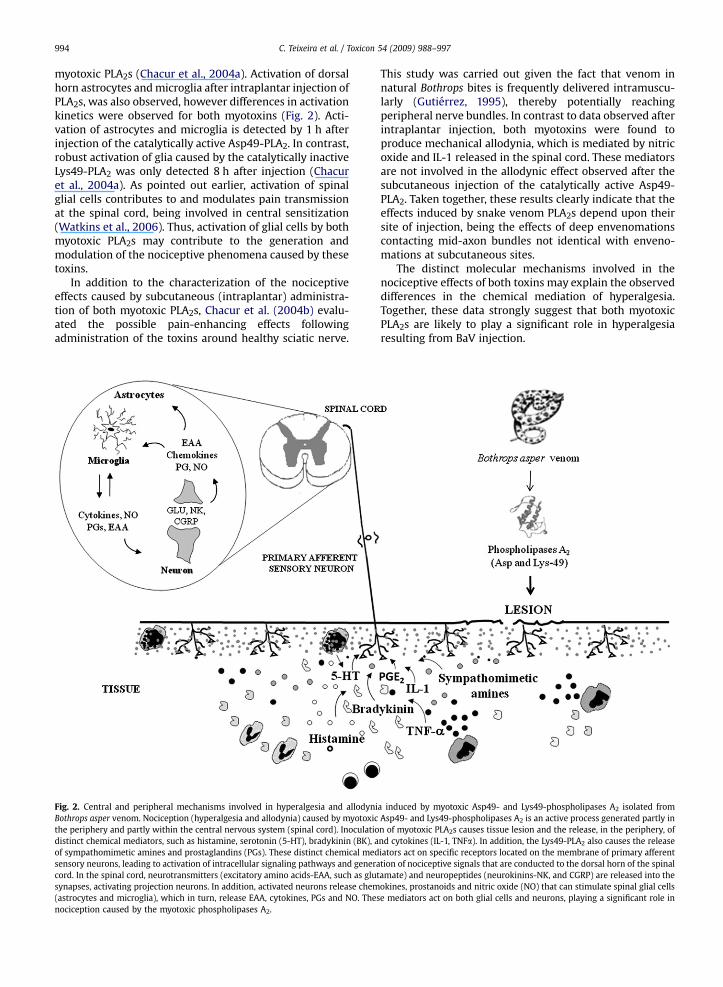
C. Teixeira et al. / Toxicon 54 (2009) 988–997994
myotoxic PLA2s (Chacur et al., 2004a). Activation of dorsalhorn astrocytes and microglia after intraplantar injection ofPLA2s, was also observed, however differences in activationkinetics were observed for both myotoxins (Fig. 2). Acti-vation of astrocytes and microglia is detected by 1 h afterinjection of the catalytically active Asp49-PLA2. In contrast,robust activation of glia caused by the catalytically inactiveLys49-PLA2 was only detected 8 h after injection (Chacuret al., 2004a). As pointed out earlier, activation of spinalglial cells contributes to and modulates pain transmissionat the spinal cord, being involved in central sensitization(Watkins et al., 2006). Thus, activation of glial cells by bothmyotoxic PLA2s may contribute to the generation andmodulation of the nociceptive phenomena caused by thesetoxins.
In addition to the characterization of the nociceptiveeffects caused by subcutaneous (intraplantar) administra-tion of both myotoxic PLA2s, Chacur et al. (2004b) evalu-ated the possible pain-enhancing effects followingadministration of the toxins around healthy sciatic nerve.
Fig. 2. Central and peripheral mechanisms involved in hyperalgesia and allodyniBothrops asper venom. Nociception (hyperalgesia and allodynia) caused by myotoxicthe periphery and partly within the central nervous system (spinal cord). Inoculatiodistinct chemical mediators, such as histamine, serotonin (5-HT), bradykinin (BK), aof sympathomimetic amines and prostaglandins (PGs). These distinct chemical medsensory neurons, leading to activation of intracellular signaling pathways and generacord. In the spinal cord, neurotransmitters (excitatory amino acids-EAA, such as glusynapses, activating projection neurons. In addition, activated neurons release chem(astrocytes and microglia), which in turn, release EAA, cytokines, PGs and NO. Thesnociception caused by the myotoxic phospholipases A2.
This study was carried out given the fact that venom innatural Bothrops bites is frequently delivered intramuscu-larly (Gutierrez, 1995), thereby potentially reachingperipheral nerve bundles. In contrast to data observed afterintraplantar injection, both myotoxins were found toproduce mechanical allodynia, which is mediated by nitricoxide and IL-1 released in the spinal cord. These mediatorsare not involved in the allodynic effect observed after thesubcutaneous injection of the catalytically active Asp49-PLA2. Taken together, these results clearly indicate that theeffects induced by snake venom PLA2s depend upon theirsite of injection, being the effects of deep envenomationscontacting mid-axon bundles not identical with enveno-mations at subcutaneous sites.
The distinct molecular mechanisms involved in thenociceptive effects of both toxins may explain the observeddifferences in the chemical mediation of hyperalgesia.Together, these data strongly suggest that both myotoxicPLA2s are likely to play a significant role in hyperalgesiaresulting from BaV injection.
a induced by myotoxic Asp49- and Lys49-phospholipases A2 isolated fromAsp49- and Lys49-phospholipases A2 is an active process generated partly inn of myotoxic PLA2s causes tissue lesion and the release, in the periphery, ofnd cytokines (IL-1, TNFa). In addition, the Lys49-PLA2 also causes the releaseiators act on specific receptors located on the membrane of primary afferenttion of nociceptive signals that are conducted to the dorsal horn of the spinal
tamate) and neuropeptides (neurokinins-NK, and CGRP) are released into theokines, prostanoids and nitric oxide (NO) that can stimulate spinal glial cellse mediators act on both glial cells and neurons, playing a significant role in

C. Teixeira et al. / Toxicon 54 (2009) 988–997 995
4.2. Metalloproteinases
A growing body of evidence points to a relevant role ofmetalloproteinases in the inflammatory reaction andhyperalgesia induced by BaV. BaP1 is the most abundantamong the five metalloproteinases isolated from BaV(Gutierrez et al., 1995). This toxin belongs to the class P-Iaccording to the classification of snake venom metal-loproteinases (Bjarnason and Fox, 1994). Besides inducingweak hemorrhage and moderate myonecrosis upon intra-muscular injection, this metalloproteinase induces oedemain the mouse footpad assay (Gutierrez et al., 1995). BaP1also induces inflammatory infiltrate composed mainly ofpolymorphonuclear leukocytes and macrophages as well asincrements in the cytokines IL-1b and IL-6 when injectedinto mouse gastrocnemius muscle (Rucavado et al., 1995,2002). Leukocyte margination along the venular endothe-lium after a few minutes of BaP1 exposure was observedthrough intravital microscopy analysis of mouse cremastermuscle (Rucavado et al., 1995). Subsequently, blisterformation and infiltration of leukocytes into the dermis wasdemonstrated after intramuscular injection of BaP1, thiseffect being associated with degranulation of mast cells andpresence of enlarged macrophages (Rucavado et al., 1998).
The ability of BaV metalloproteinases to recruit leuko-cytes was further dissected by Fernandes et al. (2006).Injection of BaP1 into mouse peritoneal cavity evokeda marked infiltration of neutrophils and mononuclearleukocytes into the peritoneum, in parallel to the release ofIL-1 and TNF-a. Leukocyte recruitment induced by BaP1wasregulated by LECAM, CD-18 and LFA-1 adhesion molecules,suggesting that BaP1 contributes to the inflammatoryevents induced by the whole venom (Fig. 1). Moreover, aneffect of BaP1 on bone marrow compartments leading toincreased numbers of leukocytes at inflamed tissue wassuggested by these authors, since a marked rise in theblood neutrophils, correlating with the peak increase ofleukocyte accumulation into peritoneum, was observedafter BaP1 injection. These data are in agreement withprevious studies showing the ability of this metal-loproteinase to activate the complement system, gener-ating chemotactic factors, such as C5a component, whichstimulates neutrophil chemotaxis (Farsky et al., 2000)(Fig. 1). These findings support a role for BaP1 and probablyother venom metalloproteinases in leukocyte recruitmentinduced by BaV.
In addition, recent data have shown that BaP1 causesjoint hypernociception, mediated by the local release ofPGE2 and TNF-a. (Fernandes et al., 2007). Therefore, inaddition to PLA2s, metalloproteinases could also contributeto the nociceptive effect induced by BaV.
4.3. Serine proteinases
Serine proteinases represent another group of venomcomponents potentially relevant for the inflammatoryresponse evoked by BaV. Studies with inhibitors and puri-fied enzymes demonstrated that venom serine proteinasesactivated proMMP-2 in human fibroblasts in culture, sug-gesting that these proteinases might play a predominantrole in the activation of endogenous metalloproteinases
(Saravia-Otten et al., 2004). These findings suggest theparticipation of serine proteinases in the local inflamma-tion induced by whole venom (Saravia-Otten et al., 2004).However, an in vivo study demonstrated that the localinjection of a thrombin-like serine proteinase from BaV inmice did not result in the activation of proMMPs (Perezet al., 2007).
Therefore, although the available evidence supporta predominant role for basic myotoxic phospholipases A2
and P-I class metalloproteinases in the pro-inflammatoryand nociceptive activities that characterize the action ofBaV, a role for other venom components, such as serineproteinases, P-II and P-III class metalloproteinases, acidicphospholipases A2 and other components acting on plasmainflammatory enzyme cascades or inducing mast celldegranulation, cannot be ruled out. In addition, it must beconsidered that a particular inflammatory event induced bythe venom may be the result of synergistic effects of two ormore components.
5. Concluding remarks
Despite the advances in our understanding of theinflammatory response to BaV, much remains to be learnedon the mechanisms involved in the initiation and devel-opment of the inflammatory events triggered by thisvenom, with special attention to the range of mediatorsinvolved, the regulatory steps associated with theirproduction and action, and the actual types and subtypes ofreceptors activated by the principal mediators. Within thisframe, the identification of mediators involved in thetransition from venom-induced inflammation to resolutionis one of the major questions to be clarified in the future.Moreover, the functionality of the physiological counter-part of the inflammatory state triggered by this venom,such as the hipotalamus–hypophysis–adrenal axis is stillunknown and represents another relevant aspect to beinvestigated. In addition, while there are much evidence forcontribution of PLA2s and metalloproteinases for inflam-mation induced by BaV, the participation of other venomtoxins needs to be addressed. On the other hand, at presentthe role of the inflammatory reaction in the developmentand resolution of venom-induced local pathology isunclear. Therefore, further studies on this subject arestressed. Finally, a deeper and comprehensive under-standing of the various aspects associated with BaV-induced inflammation and nociception should provideclues to the events and processes determining the resolu-tion of tissue damage and inflammation, thus paving theway for the development of novel therapeutic approachesaimed at counteracting the deleterious effects of localtissue damage and inflammation, and to foster thoseprocesses involved in tissue repair and regeneration.
Acknowledgments
We are grateful to Sergio A. de Lima and Marcio H.Matsubara for helpful discussions. The financial support ofCNPq (PQ: 301199/91-4) and Vicerrectoria de Investigacion(Universidad de Costa Rica) is acknowledged. VM is therecipient of FAPESP postdoctoral fellowship 07/0337-5.

C. Teixeira et al. / Toxicon 54 (2009) 988–997996
Conflict of interest
The authors declare that there are no conflicts ofinterest.
References
Abrahamsen, B., Zhao, J., Asante, C.O., Cendan, C.M., Marsh, S., Martinez-Barbera, J.P., Nassar, M.A., Dickenson, A.H., Wood, J.N., 2008. Cell andmolecular basis of mechanical, cold, and inflammatory pain. Science321, 702–705.
Alape-Giron, A., Sanz, L., Escolano, J., Flores-Dıaz, M., Madrigal, M.,Sasa, M., Calvete, J.J., 2008. Snake venomics of the lancead pitviperBothrops asper: geographic, individual, and ontogenetic variations.J. Prot. Res. 7, 3556–3571.
Arroyo, O., Rojas, G., Gutierrez, J.M., 1999. Envenenamiento por morde-dura de serpiente en Costa Rica en 1996: epidemiologıa y consid-eraciones clınicas. Acta Med. Costarric. 41, 23–29.
Avila-Aguero, M.L., Parıs, M.M., Hu, S., Peterson, P.K., Gutierrez, J.M.,Lomonte, B., Faingezicht, I., Snakebite Study Group, 2001. Systemiccytokine response in children bitten by snakes in Costa Rica. Pediatr.Emerg. Care 17, 425–429.
Bjarnason, J.B., Fox, J.W., 1994. Hemorrhagic metalloproteinases fromsnake venoms. Pharmacol. Ther. 62, 325–372.
Bolanos, R., 1984. Serpientes, Venenos y Ofidismo en Centroamerica.Editorial Universidad de Costa Rica, San Jose, p. 136.
Carlos, T.M., Harlan, J.M., 1994. Leukocyte-endothelial adhesion mole-cules. Blood 84, 2068–2101.
Chacur, M., Picolo, G., Gutierrez, J.M., Teixeira, C.F.P., Cury, Y., 2001.Pharmacological modulation of hyperalgesia induced by Bothropsasper (terciopelo) snake venom. Toxicon 39, 1173–1181.
Chacur, M., Longo, I., Picolo, G., Gutierrez, J.M., Lomonte, B., Guerra, J.L.,Teixeira, C.F.P., Cury, Y., 2003. Hyperalgesia induced by Asp49 andLys49 phospholipases A2 from Bothrops asper snake venom: phar-macological mediation and molecular determinants. Toxicon 41, 667–678.
Chacur, M., Gutierrez, J.M., Milligan, E.D., Wieseler-Frank, J., Britto, L.R.G.,Maier, S.F., Watkins, L.R., Cury, Y., 2004a. Snake venom componentsenhance pain upon subcutaneous injection: an initial examination ofspinal cord mediators. Pain 111, 65–76.
Chacur, M., Milligan, E.D., Sloan, E.M., Wieseler-Frank, J., Barrientos, R.,Martin, D., Poole, S., Gutierrez, J.M., Maier, S.F., Cury, Y., Watkins, L.R.,2004b. Snake venom phospholipase A2s (Asp49 and Lys49) inducemechanical allodynia upon peri-sciatic administration: involvementof spinal cord proinflammatory cytokines and nitric oxide. Pain 108,180–191.
Chaves, F., Gutierrez, J.M., Brenes, F., 1992. Pathological and biochemicalchanges induced in mice after intramuscular injection of venom fromnewborn specimens of the snake Bothrops asper (Terciopelo). Toxicon30, 1099–1109.
Chaves, F., Barboza, M., Gutierrez, J.M., 1995. Pharmacological study ofedema induced by venom of the snake Bothrops asper (terciopelo).Toxicon 33, 31–39.
Chaves, F., Leon, G., Alvarado, V.H., Gutierrez, J.M., 1998. Pharmacologicalmodulation of edema induced by Lys-49 and Asp-49 myotoxicphospholipases A2 isolated from the venom of snake Bothrops asper(terciopelo). Toxicon 36, 1861–1869.
Chaves, F., Teixeira, C.F., Gutierrez, J.M., 2006. Role of nitric oxide in thelocal and systemic pathophysiological effects induced by Bothropsasper snake venom in mice. Inflamm. Res. 55, 245–253.
Cheng, J.K., Ji, R.R., 2008. Intracellular signaling in primary sensoryneurons and persistent pain. Neurochem. Res. 33, 1970–1978.
Cummins, T.R., Sheets, P.L., Waxman, S.G., 2007. The roles of sodiumchannels in nociception: implications for mechanisms of pain. Pain131, 243–257.
Farsky, S.H., Gonçalves, L.R., Gutierrez, J.M., Correa, A.P., Rucavado, A.,Gasque, P., Tambourgi, D.V., 2000. Bothrops asper snake venom and itsmetalloproteinase BaP-1 activate the complement system. Role inleucocyte recruitment. Mediators Inflamm. 9, 213–221.
Fernandes, C.M., Zamuner, S.R., Zuliani, J.P., Rucavado, A., Gutierrez, J.M.,Teixeira, C.F.P., 2006. Inflammatory effects of BaP1 a metal-loproteinase isolated from Bothrops asper snake venom: leukocyterecruitment and release of cytokines. Toxicon 47, 549–559.
Fernandes, C.M., Pereira Teixeira, C.F., Leite, A.C., Gutierrez, J.M., Rocha, F.A., 2007. The snake venom metalloproteinase BaP1 induces jointhypernociception through TNF-alpha and PGE2-dependent mecha-nisms. Br. J. Pharmacol. 151, 1254–1261.
Garcia-Leme, J., Hammamura, L., Leite, M.P., Rocha e Silva, M., 1973.Pharmacological analysis of the acute inflammatory process inducedin the rat’s paw by local injection of carrageenin and by heating. Br. J.Pharmacol. 48, 88–96.
Giordano, J., 2005. The neurobiology of nociceptive and anti-nociceptivesystems. Pain Physician 8, 277–290.
Granger, D.N., Kubes, P., 1994. The microcirculation and inflammation:modulation of leukocyte-endothelial cell adhesion. J. Leukoc. Biol. 55,662–675.
Gutierrez, J.M., Arroyo, O., Bolanos, R., 1980. Myonecrosis, hemorrhageand edema induced by Bothrops asper venom in white mice. Toxicon18, 603–610.
Gutierrez, J.M., Chaves, F., Bolanos, R., Cerdas, L., Rojas, E., Arroyo, 0.,Portilla, E., 1981. Neutralization de los efectos locales de1 venenode Bothrops asper por un antiveneno polivalente. Toxicon 19,493–500.
Gutierrez, J.M., Chaves, F., Cerdas, L., 1986a. Inflammatory infiltrate in skeletalmuscle injected with Bothrops asper venom. Rev. Biol. Trop. 34, 209–214.
Gutierrez, J.M., Rojas, G., Lomonte, B., Gene, J.A., Cerdas, L., 1986b.Comparative study of the edema-forming activity of Costa Ricansnake venoms and its neutralization by polyvalent antivenom. Comp.Biochem. Physiol. C 85, 171–175.
Gutierrez, J.M., Lomonte, B., Chaves, F., Moreno, E., Cerdas, L., 1986c.Pharmacological activities of a toxic phospholipase A isolated fromthe venom of the snake Bothrops asper. Comp. Biochem. Physiol. 84,159–164.
Gutierrez, J.M., 1995. Clinical toxicology of snake bite in CentralAmerica. In: Meier, J., White, J. (Eds.), Handbook of Clinical Toxi-cology of Animal Venoms and Poisons. CRC Press, Boca Raton, FL,pp. 645–665.
Gutierrez, J.M., Lomonte, B., 1995. Phospholipase A2 myotoxins fromBothrops snake venoms. Toxicon 33, 1405–1424.
Gutierrez, J.M., Romero, M., Dıaz, C., Borkow, G., Ovadia, M., 1995. Isolationand characterization of a metalloproteinase with weak hemorrhagicactivity from the venom of the snake Bothrops asper (terciopelo).Toxicon 33, 19–29.
Gutierrez, J.M., Lomonte, B., 1997. Phospholipase A2 myotoxins fromBothrops snake venoms. In: Kini, R.M. (Ed.), Venom Phospholipases A2Enzymes. Structure, Function and Mechanism. Wiley, UnitedKingdom, pp. 321–352.
Haddad, J.J., 2007. On the enigma of pain and hyperalgesia: a molecularperspective. Biochem. Biophys. Res. Commun. 353, 217–224.
Harvey, V.L., Dickenson, A.H., 2008. Mechanisms of pain in nonmalignantdisease. Curr. Opin. Support. Palliat. Care 2, 133–139.
Hopkins, S.J., 2003. The pathophysiological role of cytokines. Leg. Med. 5,S45–S57.
Hubbard, A., Rothlein, R., 2000. Intercellular adhesion molecule-1(ICAM-1) expression and cell signaling cascades. Free Radic. Biol.Med. 28, 1379–1386.
Jorge, M.T., Ribeiro, L.A., O’Connell, J.L., 1999. Prognostic factors foramputation in the case of envenoming by snakes of the Bothropsgenus (Viperidae). Ann. Trop. Med. Parasitol. 93, 401–408.
Kaplanski, G., Marin, V., Montero-Julian, F., Mantovani, A., Farnarier, C.,2003. IL-6: a regulator of the transition from neutrophil to monocyterecruitment during inflammation. Trends Immunol. 24, 25–29.
Kim, D., Park, D., Choi, S., Lee, S., Sun, M., Kim, C., Shin, H.S., 2003.Thalamic control of visceral nociception mediated by T-Type Ca2þ
channels. Science 302, 117–119.Lomonte, B., Carmona, E., 1992. Individual expression patterns of myo-
toxin isoforms in the venom of the snake Bothrops asper. Comp.Biochem. Physiol. B 102, 325–329.
Lomonte, B., Lundgren, J., Johansson, B., Bagge, U., 1994. The dynamics oflocal tissue damage induced by Bothrops asper snake venom andmyotoxin II on the mouse cremaster muscle: an intravital and elec-tron microscopic study. Toxicon 32, 41–55.
Lomonte, B., Tarkowski, A., Hanson, L.A., 1993. Host response to Bothropsasper snake venom. Analysis of edema formation, inflammatory cells,and cytokine release in a mouse model. Inflammation 17, 93–105.
Lomonte, B., Angulo, Y., Calderon, L., 2003. An overview of lysine-49phospholipase A2 myotoxins from crotalid snake venoms and theirstructural determinants of myotoxic action. Toxicon 42, 885–901.
Luvisetto, S., Marinelli, S., Panasiti, M.S., D’Amato, F.R., Fletcher, C.F.,Pavone, F., Pietrobon, D., 2006. Pain sensitivity in mice lacking theCa(v)2.1alpha1 subunit of P/Q-type Ca2þ channels. Neuroscience 142,823–832.
McMahon, S.B., Cafferty, W.B., Marchand, F., 2005. Immune and glial cellfactors as pain mediators and modulators. Exp. Neurol. 192, 444–462.
Medzhitov, R., 2008. Origin and physiological roles of inflammation.Nature 454, 428–435.

C. Teixeira et al. / Toxicon 54 (2009) 988–997 997
Mora, J., Mora, R., Lomonte, B., Gutierrez, J.M., 2008. Effects of Bothropsasper snake venom on lymphatic vessels: insights into a hiddenaspect of envenomation. PLoS Negl. Trop. Dis. 2, e318.
Moreira, V., Gutierrez, J.M., Amaral, R.B., Zamuner, S.R., Teixeira, C.F.P.,2009. Effects of Bothrops asper snake venom on the expression ofcyclooxygenases and production of prostaglandins by peritonealleukocytes in vivo, and by isolated neutrophils and macrophages invitro. Prostaglandins Leukot. Essent. Fatty Acids 8, 107–114.
Moreira, V., Gutierrez, J.M., Soares, A.M., Zamuner, S.R., Purgatto, E.,Teixeira, C.F.P., 2008. Secretory phospholipases A2 isolated fromBothrops asper and from Crotalus durissus terrificus snake venomsinduce distinct mechanisms for biosynthesis of prostaglandins E(2)and D(2) and expression of cyclooxygenases. Toxicon 52, 428–439.
Ohsaka, A., 1979. Hemorrhagic, necrotizing and edema-forming effects ofsnake venoms. In: Lee, C.Y. (Ed.), Handbook of Experimental Phar-macology, Snake Venoms, vol. 52. Springer-Verlag, Berlin, pp.480–546.
Olivo, R.A., Teixeira, C.F.P., Wallace, J.L., Gutierrez, J.M., Zamuner, S.R.,2007. Role of cyclooxygenases in oedema-forming activity ofbothropic venoms. Toxicon 49, 670–677.
Otero, R., Gutierrez, J., Mesa, M.B., Duque, E., Rodrıguez, O., Luis Arango, J.,Gomez, F., Toro, A., Cano, F., Rodrıguez, L.M., Caro, E., Martınez, J.,Cornejo, W., Gomez, L.M., Uribe, F.L., Cardenas, S., Nunez, V., Dıaz, A.,2002. Complications of Bothrops, Porthidium, and Bothriechis snake-bites in Colombia. A clinical and epidemiological study of 39 casesattended in a university hospital. Toxicon 40, 1107–1114.
Perez, A.V., Saravia, P., Rucavado, A., Sant’Ana, C.D., Soares, A.M.,Gutierrez, J.M., 2007. Local and systemic pathophysiological alter-ations induced by a serine proteinase from the venom of the snakeBothrops jararacussu. Toxicon 49, 1063–1069.
Picolo, G., Chacur, M., Gutierrez, J.M., Teixeira, C.F.P., Cury, Y., 2002. Eval-uation of antivenoms in the neutralization of hyperalgesia and edemainduced by Bothrops jararaca and Bothrops asper snake venoms. Braz.J. Med. Biol. Res. 35, 1221–1228.
Rabinovitch, M., De Stefano, M.J., 1973. Macrophage spreading in vitro.I. Inducers of spreading. Exp. Cell Res. 77, 323–334.
Ribeiro, L.A., Jorge, M.T., 1997. Acidentes por serpentes do generoBothrops: serie de 3139 casos. Rev. Soc. Bras. Med. Trop. 30, 475–480.
Rittner, H.L., Brack, A., Stein, C., 2008a. Pain and the immune system. Br. J.Anaesth. 101, 40–44.
Rittner, H.L., Brack, A., Stein, C., 2008b. The other side of the medal: howchemokines promote analgesia. Neurosci. Lett. 437, 203–208.
Robertson, T.A., Maley, M.A.L., Grounds, M.D., Papadimitriou, J.M., 1993.The role of macrophages in skeletal muscle regeneration withparticular reference to chemotaxis. Exp. Cell Res. 207, 321–331.
Rohn, T.T., Nelson, L.K., Sipes, K.M., Swain, S.D., Jutila, K.L., Quinn, M.T.,1999. Priming of human neutrophils by peroxynitrite: potential rolein enhancement of the local inflammatory response. J. Leukoc. Biol.65, 59–70.
Rosenfeld, G., 1971. Symptomatology, pathology and treatment of snakebites in South America. In: Bucherl, W., Buckley, E.E. (Eds.), VenomousAnimals and their Venoms. Academic Press, New York, pp. 345–403.
Rucavado, A., Escalante, T., Teixeira, C.F.P., Fernandes, C.M., Diaz, C.,Gutierrez, J.M., 2002. Increments in cytokines and matrix metal-loproteinases in skeletal muscle after injection of tissue-damagingtoxins from the venom of the snake Bothrops asper. MediatorsInflamm. 11, 121–128.
Rucavado, A., Lomonte, B., Ovadia, M., Gutierrez, J.M., 1995. Local tissuedamage induced by BaP1, a metalloproteinase isolated from Bothropsasper (Terciopelo) snake venom. Exp. Mol. Pathol. 63, 186–199.
Rucavado, A., Nunez, J., Gutierrez, J.M., 1998. Blister formation and skindamage induced by BaP1, a haemorrhagic metalloproteinase from thevenom of the snake Bothrops asper. Int. J. Exp. Pathol. 79, 245–254.
Ryan, G.B., Majno, G., 1977. Antiinflammation. A review. Am. J. Pathol. 86,183–276.
Saegusa, H., Kurihara, T., Zong, S., Kazuno, A., Matsuda, Y., Nonaka, T.,Han, W., Toriyama, H., Tanabe, T., 2001. Suppression of inflammatoryand neuropathic pain symptoms in mice lacking the N-type Ca2þ
channel. EMBO J. 20, 2349–2356.Saegusa, H., Matsuda, Y., Tanabe, T., 2002. Effects of ablation of N- and
R-type Ca(2þ) channels on pain transmission. Neurosci. Res. 43,1–7.
Saravia-Otten, P., Frisan, T., Thelestam, M., Gutierrez, J.M., 2004.Membrane independent activation of fibroblast proMMP-2 by snakevenom: novel roles for venom proteinases. Toxicon 44, 749–764.
Sawai, Y., 1980. Studies on snakebites in the Asian areas. In: Eaker, D.,Wadstrom, T. (Eds.), Natural Toxins. Pergamon Press, Oxford, pp.25–32.
Schaible, H.G., Richter, F., 2004. Pathophysiology of pain. LangenbecksArch. Surg. 389, 237–243.
Serhan, C.N., Savill, J., 2005. Resolution of inflammation: the beginningprograms the end. Nat. Immunol. 6, 1191–1197.
Sherwood, E.R., Toliver-Kinsky, T., 2004. Mechanisms of the inflammatoryresponse. Best. Pract. Res. Clin. Anaesthesiol. 18, 385–405.
Sternlicht, M.D., Werb, Z., 2001. How matrix metalloproteinases regulatecell behavior. Annu. Rev. Cell Biol. 17, 463–516.
Streuli, C., 1999. Extracellular matrix remodeling and cellular differenti-ation. Curr. Opin. Cell Biol. 11, 634–640.
Suwa, T., Hogg, J.C., Quinlan, K.B., Van Eeden, S.F., 2002. The effect ofinterleukin-6 on L-selectin levels on polymorphonuclear leukocytes.Am. J. Physiol. Heart Circ. Physiol. 283, H879–H884.
Teixeira, C.F.P., Zamuner, S.R., Zuliani, J.P., Fernandes, C.M., Cruz-Hofling, M.A., Fernandes, I., Chaves, F., Gutierrez, J.M., 2003. Neutro-phils do not contribute to local tissue damage, but play a key role inskeletal muscle regeneration, in mice injected with Bothrops aspersnake venom. Muscle Nerve 28, 449–459.
Till, G.O., 1998. Chemotatic factors. In: Rother, K., Till, G.O. (Eds.), TheComplement System. Springer, Berlin, pp. 367–374.
Tortora, G.J., Grabowski, S.R., 2002. Principles of Anatomy and Physiology,10th ed. John Wiley, Hoboken.
Vanegas, H., Schaible, H., 2000. Effects of antagonists to high-thresholdcalcium channels upon spinal mechanisms of pain, hyperalgesia andallodynia. Pain 85, 9–18.
Watkins, L.R., Wieseler-Frank, J., Milligan, E.D., Johnston, I., Maier, S.F.,2006. Contribution of glia to pain processing in health and disease. In:Cervero, F., Jensen, T.S. (Eds.), Handbook of Clinical Neurology. Pain,vol. 81. Elsevier B.V, London, UK, pp. 309–323.
Wieseler-Frank, J., Maier, S.F., Watkins, L.R., 2004. Glial activation andpathological pain. Neurochem. Int. 45, 389–395.
Woolf, C.J., 2004. Pain: moving from symptom control toward mecha-nism-specific pharmacologic management. Ann. Intern. Med. 140,441–451.
Woolf, C.J., Salter, M.W., 2000. Neuronal plasticity: increasing the gain inpain. Science 288, 1765–1769.
Zamuner, S.R., Gutierrez, J.M., Muscara, M.N., Teixeira, S.A., Teixeira, C.F.P.,2001. Bothrops asper and Bothrops jararaca snake venoms triggermicrobicidal functions of peritoneal leukocytes in vivo. Toxicon 39,1505–1513.
Zamuner, S.R., Zuliani, J.P., Fernandes, C.M., Gutierrez, J.M., Teixeira, C.F.P.,2005. Inflammation induced by Bothrops asper venom: release ofproinflammatory cytokines and eicosanoids, and role of adhesionmolecules in leukocyte infiltration. Toxicon 46, 806–813.
Zuliani, J.P., Fernandes, C.M., Zamuner, S.R., Gutierrez, J.M., Teixeira, C.F.P.,2005a. Inflammatory events induced by Lys-49 and Asp-49 phos-pholipases A2 isolated from Bothrops asper snake venom: role ofcatalytic activity. Toxicon 45, 335–346.
Zuliani, J.P., Gutierrez, J.M., Casais e Silva, L.L., Sampaio, S.C., Lomonte, B.,Teixeira, C.F.P., 2005b. Activation of cellular functions in macrophagesby venom secretory Asp-49 and Lys-49 phospholipases A2. Toxicon46, 523–532.
Copyright © 2022 FDOKUMEN




















