
Bothrops jararaca venom gland transcriptome: Analysis of the gene expression pattern
25
Toxicon 48 (2006) 437–461 Bothrops jararaca venom gland transcriptome: Analysis of the gene expression pattern Daniela A.P. Cidade a, , Tatiana A. Sima˜o a , Alberto M.R. Da´vila b , Glauber Wagner b , Ina´cio de L.M. Junqueira-de-Azevedo c , Paulo Lee Ho c , Cassian Bon d , Russolina B. Zingali e , Rodolpho M. Albano a, a Departamento de Bioquı´mica, Universidade do Estado do Rio de Janeiro, CEP 20551 013 Rio de Janeiro, RJ, Brasil b Departamento de Bioquı´mica e Biologia Molecular, Instituto Oswaldo Cruz, FIOCRUZ, Av. Brasil, 4365 CEP 21045 900 Rio de Janeiro, RJ, Brasil c Centro de Biotecnologia, Instituto Butantan, CEP 05503-900 Sa˜o Paulo, SP, Brasil d Centre National de la Recherche Scientifique 3, rue Michel-Ange 75794, Paris, France e Instituto de Bioquı´mica Me´dica, Rede Proteoˆmica do Rio de Janeiro, Universidade Federal do Rio de Janeiro, CEP 21941-590 Rio de Janeiro, RJ, Brasil Available online 7 July 2006 Abstract Bothrops jararaca is a pit viper responsible for the majority of snake envenoming accidents in Brazil. As an attempt to describe the transcriptional activity of the venom gland, ESTs of a cDNA library constructed from B. jararaca venom gland were generated and submitted to bioinformatics analysis. The results showed a clear predominance of transcripts coding for toxins instead of transcripts coding for proteins involved in cellular functions. Among toxins, the most frequent transcripts were from metalloproteinases (52.6%), followed by serine-proteinases (28.5%), C-type lectins (8.3%) and bradykinin-potentiating peptides (BPPs) (6.2%). Results were similar to that obtained from the transcriptome analysis of B. insularis, a phylogenetically close sister of B. jararaca, though some differences were observed and are pointed out, such as a higher amount of the hypotensive BPPs in B. insularis transcriptome (19.7%). Another striking difference observed is that PIII and PII-classes of metalloproteinases are similarly represented in B. jararaca in contrast to B. insularis, in which a predominance of PIII-class metalloproteinase, which present a more intense hemorrhagic action, is observed. These features may, in part, explain the higher potency of B. insularis venom. The results obtained can help in proteome studies, and the clones can be used to directly probe the genetic material from other snake species or to investigate differences in gene expression pattern in response to factors such as diet, aging and geographic localization. r 2006 Elsevier Ltd. All rights reserved. Keywords: Snake venom; Viperidae; Toxins; cDNA library; Expressed sequence tags 1. Introduction Bothrops genus includes more than 30 species and subspecies distributed between Central and South America. Members of this genus are responsible for approximately 90% of venomous snakebite ARTICLE IN PRESS www.elsevier.com/locate/toxicon 0041-0101/$ - see front matter r 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.toxicon.2006.07.008 Corresponding author. Tel.: +55 21 2587 6428; fax: +55 21 25876136. E-mail addresses: [email protected] (D.A.P. Cidade), [email protected] (R.M. Albano).
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Bothrops jararaca venom gland transcriptome: Analysis of the gene expression pattern

ARTICLE IN PRESS
0041-0101/$ - see
doi:10.1016/j.tox
�Correspondifax: +5521 258
E-mail addre
(D.A.P. Cidade
Toxicon 48 (2006) 437–461
www.elsevier.com/locate/toxicon
Bothrops jararaca venom gland transcriptome:Analysis of the gene expression pattern
Daniela A.P. Cidadea,�, Tatiana A. Simaoa, Alberto M.R. Davilab,Glauber Wagnerb, Inacio de L.M. Junqueira-de-Azevedoc, Paulo Lee Hoc,
Cassian Bond, Russolina B. Zingalie, Rodolpho M. Albanoa,�
aDepartamento de Bioquımica, Universidade do Estado do Rio de Janeiro, CEP 20551 013 Rio de Janeiro, RJ, BrasilbDepartamento de Bioquımica e Biologia Molecular, Instituto Oswaldo Cruz, FIOCRUZ, Av. Brasil,
4365 CEP 21045 900 Rio de Janeiro, RJ, BrasilcCentro de Biotecnologia, Instituto Butantan, CEP 05503-900 Sao Paulo, SP, Brasil
dCentre National de la Recherche Scientifique 3, rue Michel-Ange 75794, Paris, FranceeInstituto de Bioquımica Medica, Rede Proteomica do Rio de Janeiro, Universidade Federal do Rio de Janeiro,
CEP 21941-590 Rio de Janeiro, RJ, Brasil
Available online 7 July 2006
Abstract
Bothrops jararaca is a pit viper responsible for the majority of snake envenoming accidents in Brazil. As an attempt to
describe the transcriptional activity of the venom gland, ESTs of a cDNA library constructed from B. jararaca venom
gland were generated and submitted to bioinformatics analysis. The results showed a clear predominance of transcripts
coding for toxins instead of transcripts coding for proteins involved in cellular functions. Among toxins, the most frequent
transcripts were from metalloproteinases (52.6%), followed by serine-proteinases (28.5%), C-type lectins (8.3%) and
bradykinin-potentiating peptides (BPPs) (6.2%). Results were similar to that obtained from the transcriptome analysis of
B. insularis, a phylogenetically close sister of B. jararaca, though some differences were observed and are pointed out, such
as a higher amount of the hypotensive BPPs in B. insularis transcriptome (19.7%). Another striking difference observed is
that PIII and PII-classes of metalloproteinases are similarly represented in B. jararaca in contrast to B. insularis, in which a
predominance of PIII-class metalloproteinase, which present a more intense hemorrhagic action, is observed. These
features may, in part, explain the higher potency of B. insularis venom. The results obtained can help in proteome studies,
and the clones can be used to directly probe the genetic material from other snake species or to investigate differences in
gene expression pattern in response to factors such as diet, aging and geographic localization.
r 2006 Elsevier Ltd. All rights reserved.
Keywords: Snake venom; Viperidae; Toxins; cDNA library; Expressed sequence tags
front matter r 2006 Elsevier Ltd. All rights reserved
icon.2006.07.008
ng author. Tel.: +5521 2587 6428;
76136.
sses: [email protected]
), [email protected] (R.M. Albano).
1. Introduction
Bothrops genus includes more than 30 species andsubspecies distributed between Central and SouthAmerica. Members of this genus are responsiblefor approximately 90% of venomous snakebite
.

ARTICLE IN PRESSD.A.P. Cidade et al. / Toxicon 48 (2006) 437–461438
accidents in Brazil. Bothrops jararaca is the mostcommon species that occurs in the southeast regionof Brazil, accounting for the majority of accidents(Franc-a and Malaque, 2003; Ribeiro and Jorge,1997). Clinically, patients bitten by B. jararaca
usually present edema, systemic bleeding, thrombo-cytopenia and prolongation of whole blood clottingtime (Sano-Martins et al., 1997; Santoro and Sano-Martins, 2004). These symptoms result from the 3main activities of bothropic venom: proteolytic,with local inflammatory edema at the snakebite site;hemorrhagic, with endothelium damage and sys-temic bleeding; and procoagulant, leading to theconsumption of coagulant factors and disruptingthe equilibrium of blood coagulation (Matsui et al.,2000; Varanda and Giannini, 1999). The principalclasses of toxins involved in these actions are lectins,prothrombin activating toxins, hemorrhagins, dis-integrins and serine-proteinases among others(Markland, 1998; White, 2005).
In this work we generated and analyzed 2318expressed sequence tags (ESTs) from a cDNAlibrary of the venom gland of B. jararaca, aimingto describe its protein contents. Similar transcrip-tome approach have been conducted with othersnakes such as B. insularis (Junqueira-de-Azevedoand Ho, 2002), Bitis gabonica (Franscischetti et al.,2004) and B. jararacussu (Kashima et al., 2004).This kind of study is a good source of informationabout venom composition, eventually leading tonew toxin discovery.
B. jararaca is a close sister taxa of B. insularis
(Werman, 1992), an endemic snake from Ilha daQueimada Grande, located in Sao Paulo State(Southeast of Brazil). The geographical isolationand special diet (based exclusively on birds andsome invertebrates available on the island) to whatB. insularis has been submitted for thousands ofyears may have caused some changes in its venomcomposition (Daltry et al., 1996). In this work,we adopted the bioinformatic parameters to con-struct the EST database as close as possible to thoseused for B. insularis transcriptome (Junqueira-de-Azevedo and Ho, 2002), in order to allowcomparative studies between these two pit vipers.Therefore, we were able to infer similarities anddifferences in their venom composition, and corre-late it with the potency and clinical symptoms ofboth venoms.
In our transcriptome analysis, since we havesequenced a larger number of sequence tags, wewere able to characterize even not abundant
transcripts besides the most frequent ones, provid-ing a set of sequences from a very specializedsecretory tissue that can be used for comparativestudies and homology searches, and are available atEST section of the NCBI GenBank (http://www.ncbi.nlm.nih.gov/dbEST). Moreover, an EST data-base can also be used as an auxiliary reference inproteome studies, leading to easier and fasterprotein identification (Mathesius et al., 2001).
2. Materials and methods
2.1. cDNA library construction and EST technology
The first procedure of an EST project is to obtainmRNA from the tissue of interest, transform it intodouble-stranded cDNA and clone it into a suitablevector to create a cDNA library. With this intent,poly(A)-rich RNA was prepared from total RNA ofthe venom gland by oligo(dT)-cellulose chromato-graphy. A directional cDNA library was con-structed using a plasmid cloning kit (Superscriptplasmid system, Life Technologies Inc.), as de-scribed previously (Arocas et al., 1997). The cDNAfragments, representing transcripts for the genesthat were expressed at the moment of RNAextraction, were ligated into the pT7T3D EcoRI/NotI/BAP phagemid vector (Pharmacia LKB Bio-technology Inc.) and used to transform Epicurian
coli KL1 Blue MRF supercompetent cells (Strata-gene), producing a library of about 2� 105 inde-pendent colonies.
An important quality assessment of the library isto determine the average length of the cDNAsthat were cloned in the vector and, for this, isolatedcolonies were randomly chosen and grown insmall-scale cultures for plasmid preparation. Plas-mid DNA extraction was performed accordingto Sambrook et al., 1989, and PCR reactionswere performed using a pair of primers that flankthe multiple cloning site of the vector (T7 upstreamand T3 downstream). Amplification conditions were95 1C for 5min and 30 cycles of 95 1C for 45 s, 50 1Cfor 45 s and 72 1C for 4min, with a final extensionstep of 72 1C for 5min. The length of cDNAs wasestimated by agarose gel electrophoresis analysis ofPCR amplified products.
The next step is the large-scale sequencing of thecDNAs and, for this, aliquots of the plasmid DNAlibrary were used to transform Escherichia coli
XL1-Blue by electroporation, and plated on CircleGrow medium (Bio 101 Systems, USA) containing

ARTICLE IN PRESSD.A.P. Cidade et al. / Toxicon 48 (2006) 437–461 439
ampicilin (100 mg/ml), isopropyl-b-D-thiogalacto-pyranoside (IPTG) (120 mg/ml) and 5-bromo-4-chloro-3-indolyl-b-D-galactopyranoside (X-Gal)(400 mg/ml) to allow blue/white selection of thecolonies. White colonies, which are the ones thatcontain vectors in which cDNA molecules havebeen cloned, were randomly selected and plasmidDNA was prepared by standard methods (Sam-brook et al., 1989). After quantification, 400 ng oftemplate DNA, corresponding to a single colony,were added to sequencing reactions based on thedideoxy-chain termination method (Sanger et al.,1977), with the Dyenamic ET dye terminators Kit(Amersham-Biosciences) and T7 primer. The se-quencing reactions were analyzed on the MEGA-BACE 1000 automated DNA sequencer (Amersham-Biosciences).
The next step is to analyze the sequencechromatograms and remove low quality bases andvector sequences. With this intent, the chromato-grams were analyzed with Phred/Phrap software(http://www.phrap.org/phredphrapconsed.html).With the aid of this software, nucleotide sequenceswere subjected to vector sequence removal and lowquality 50 and 30 ends were also trimmed. Only goodquality (Phred425) sequences, longer than 150 bp,were considered for annotation.
At this point we had a collection of goodquality sequences free of any vector contamination,which had to be organized into clusters. A clustercorresponds to a group of sequences that encode thesame transcript. The clusterization of the sequenceswas performed using CAP3 software (Huang andMadan, 1999). A minimum overlap of 50 bp, with atleast 98% identity, was required for any assembly,in order to avoid misassembly of paralogoussequences, considering the eventual presence ofmany isoforms of toxins in the venom gland.
Once the clusters have been formed, the next stepis their careful annotation. The annotation stepinvolves a thorough search for related sequences inseveral databases of DNA and protein sequences,protein motifs, etc. Our first action was to search theclustered sequences against the protein NR Gen-Bank NCBI database by the use of the Blast tool.The expectation values considered to allow aputative identification were e-values o10-05. Inthe cases where no matches were found, Blast wasperformed against the nucleotide NR GenBankNCBI database. The 3rd option in searching forhomologies was to perform Blast against the ESTsection of the NCBI nucleotide database, where
most of the B. insularis EST sequences have beendeposited (Junqueira-de-Azevedo and Ho, 2002).Conserved domain searches were also performedusing the CDD tool of NCBI. As all of theinformation generated by the ESTs and theirannotation had to be organized in an user-friendlyformat, the GARSA system (Davila et al., 2005) wasused to perform all above-mentioned bioinformaticsanalysis and to store all information as a data bank.
3. Results and discussion
3.1. Quality assessment of the cDNA library
The length of cloned cDNAs was estimated byagarose gel electrophoresis analysis of PCR ampli-fied products. The results showed a distributionbetween 400 and 1600 bp, with an average of 800 bp(data not shown). After sequencing, the 3327chromatograms were submitted to bioinformaticsanalysis to remove vector and poor qualitysequences, resulting in 2318 good quality sequences.After clusterization 1154 clusters were formed, fromwhich 855 were singlets. This high number ofclusters resulted from the very stringent parametersused for clusterization, in order to avoid misassem-bling of protein isoforms. All of the 1154 clusterswere deposited on the EST division of GenBank(http://www.ncbi.nlm.nih.gov/dbEST) under acces-sion numbers DW712660 to DW713813. Addition-ally, clusters, reads and similarity analysis resultsare available trough the GARSA system at http://www.biowebdb.org/garsa.
3.2. Cluster identification by homology searches
After performing the Blast searches against theprotein, nucleotide and EST databases of NCBIGenBank and considering a minimum e-value of10�05 for identification, we found 434 clusters (549reads) with no database match (Table 1). Thisaccounted for 23.7% of the reads, a value that isconsistent with other transcriptome studies, whichshow values that vary from 10% to 47% of no hits(Davey et al., 2001; Valenzuela et al., 2002;Francischetti et al., 2002 and 2004; Ribeiro et al.,2004). These sequences may represent a source ofnew information about venom composition, codingfor proteins not yet described. In addition to thesequences with no database match, there are 83clusters (115 reads) possessing homology withproteins that have been already described but with

ARTICLE IN PRESSD.A.P. Cidade et al. / Toxicon 48 (2006) 437–461440
no functional assessment, hereby named unknownproteins.
The identified proteins were separated in twogroups: toxins and cellular proteins. From thematching clones, toxins account for 77.6% of thetranscripts, while cellular proteins represent 22.4%(Table 1). In the analysis of the B. insularis
transcriptome, the toxin representation over match-ing clones was 67%, with the cellular proteins beingresponsible for 33% of the transcripts (Junqueira-de-
Table 1
Representation of the ESTs from B. jararaca venom glands
Category Clusters (% over total) Clones (% ove
No database match 434 (37.6) 549 (23.7)
Unknown proteins 83 (7.2) 115 (5.0)
Matching sequences 637 (55.2) 1654 (71.3)
Similar to toxins 409 (35.5) 1283 (55.3)
Similar to cellular
proteins
228 (19.7) 371 (16.0)
Table 2
Identification of putative toxin-encoding clusters from B. jararaca veno
Cluster N Putative identification
Metalloproteinases
JARO001001E01 137 Bothrops jararaca bothrostatin precursor
JARO001002D11 103 Bothropasin/jararhagin [Bothrops jararaca]
JARO001001E09 77 Bothropasin/jararhagin [Bothrops jararaca]
JARO001001C04 29 Bothropasin precursor [Bothrops jararaca]
JARO001001H12 15 Metalloproteinase/disintegrin ussurin precurso
JARO001003B09 14 Bothrops jararaca bothrostatin precursor
JARO001013A02 13 Hemorrhagic metalloproteinase HF3 [Bothrop
JARO001005A03 11 Bothropasin/jararhagin [Bothrops jararaca]
JARO001002H10 10 Bothrops jararaca bothrostatin precursor
JARO001006H07 10 Bothrops jararaca bothrostatin precursor
JARO001012C05 06 Metalloprotease BOJUMET II [Bothrops jara
JARO001016B04 06 Bothropasin precursor [Bothrops jararaca]
JARO001003D11 05 Metalloprotease BOJUMET II [Bothrops jara
JARO001006B09 05 Bothropasin precursor [Bothrops jararaca]
JARO001010B06 05 Bothrops jararaca bothrostatin precursor
JARO001010E11 05 Hemorrhagic metalloproteinase HF3 [Bothrop
JARO001012A02 05 Vascular apoptosis-inducing protein [western d
JARO001014A01 05 Bothrops jararaca bothrostatin precursor
JARO001003D12 04 Hemorrhagic metalloproteinase HF3 [Bothrop
JARO001004C08 04 Bothropasin/jararhagin [Bothrops jararaca]
JARO001013H03 04 Jararhagin precursor [Bothrops jararaca]
JARO001014H07 04 Crotalus atrox prepro-hemorrhagic toxin c, atr
JARO001015D06 04 Hemorrhagic metalloproteinase HF3 [Bothrop
JARO001023G05 04 Metalloproteinase-disintegrin-like protein [Agk
JARO001043B04 04 Bothrops jararaca bothrostatin precursor
JARO001001C06 03 Metalloproteinase precursor [Bothrops insulari
Azevedo and Ho, 2002). The same pattern wasobserved for B. jararaca, where 71.6% of matchingclones codes for toxins and 28.4% codes for cellularproteins (Kashima et al., 2004). It becomes clearthat cDNA for toxins are highly represented insnake venom gland transcriptomes, suggesting thatthese molecules are preferentially expressed overproteins related with cellular functions. The toxinclusters are listed on Table 2 and the cellular proteinclusters are listed on Table 3.
r total) Redundancy (clones/
clusters)
Representation over
matching clones (%)
1.3 —
1.4 —
2.6 100.0
3.1 77.6
1.6 22.4
m
e-value
e-131
0.0
3e-44
0.0
r [Gloydius ussuriensis] 1e-33
e-180
s jararaca] e-109
7e-14
e-137
6e-36
racussu] e-122
e-163
racussu] e-103
e-124
e-160
s jararaca] 2e-17
iamond back rattlesnake e-166
e-106
s jararaca] 6e-83
e-114
e-149
olysin c 4e-80
s jararaca] 7e-28
istrodon contortrix laticinctus] 1e-08
e-123
s] 5e-97

ARTICLE IN PRESS
Table 2 (continued )
Cluster N Putative identification e-value
JARO001001G09 03 Bothropasin precursor [Bothrops jararaca] 8e-97
JARO001002B05 03 Disintegrin jararacin [Bothrops jararaca] 1e-15
JARO001009G02 03 Bothropasin precursor [Bothrops jararaca] 0.0
JARO001010A02 03 Bothrops insularis cluster BITM02A (metalloproteinase precursor) e-163
JARO001010B05 03 Bothrops jararaca bothrostatin precursor e-117
JARO001010F02 03 Disintegrin jararacin [Bothrops jararaca] 1e-23
JARO001010G12 03 Bothropasin/jararhagin [Bothrops jararaca] 5e-08
JARO001013G05 03 Bothropasin precursor [Bothrops jararaca] e-146
JARO001023A06 03 Metalloproteinase-disintegrin-like protein[Agkistrodon contortrix laticinctus] 1e-39
JARO001041C11 03 Disintegrin jararacin [Bothrops jararaca] 3e-31
JARO001002A04 02 Deinagkistrodon acutus metalloproteinase MD1 precursor 5e-98
JARO001003B12 02 Bothropasin precursor [Bothrops jararaca] e-176
JARO001003C11 02 Bothrops jararaca bothrostatin precursor 1e-94
JARO001003F09 02 Metalloproteinase/disintegrin saxin precursor [Gloydius saxatilis] 7e-52
JARO001004D08 02 Metalloproteinase [Gloydius halys brevicaudus] 1e-54
JARO001008B04 02 Bothropasin/jararhagin [Bothrops jararaca] 3e-31
JARO001008E06 02 Disintegrin jararacin [Bothrops jararaca] 7e-06
JARO001010B03 02 Bothropasin precursor [Bothrops jararaca] 3e-59
JARO001010C06 02 Bothrops jararaca bothrostatin precursor 2e-93
JARO001010C10 02 Bothropasin precursor [Bothrops jararaca] 0.0
JARO001010E10 02 Berythractivase [Bothrops erythromelas] 1e-77
JARO001012B03 02 Zinc metalloproteinase jerdonitin precursor [Trimeresurus jerdonii] 1e-75
JARO001014G10 02 Bothropasin/jararhagin [Bothrops jararaca] 4e-48
JARO001016G03 02 Bothropasin precursor [Bothrops jararaca] 2e-74
JARO001032G05 02 Hemorrhagic metalloproteinase HF3 [Bothrops jararaca] 6e-06
JARO001034B08 02 Bothrops jararaca bothrostatin precursor e-108
JARO001035B06 02 Jararhagin precursor [Bothrops jararaca] e-137
JARO001040E08 02 Bothropasin precursor [Bothrops jararaca] e-127
JARO001041A06 02 Hemorrhagic metalloproteinase HF3 [Bothrops jararaca] 7e-37
JARO001041B03 02 Metalloprotease BOJUMET II [Bothrops jararacussu] 2e-92
JARO001045D04 02 Deinagkistrodon acutus metalloproteinase MD2 precursor e-114
JARO001052D06 02 Bothrops jararaca bothrostatin precursor e-130
JARO001053A12 02 Hemorrhagic metalloproteinase HF3 [Bothrops jararaca] 1e-47
JARO001001E05 01 Bothropasin precursor [Bothrops jararaca] 2e-80
JARO001002B06 01 Bothropasin precursor [Bothrops jararaca] 3e-44
JARO001004C09 01 Bothrops insularis cluster BITM02A (metalloproteinase precursor) 0.0
JARO001005A04 01 Bothropasin precursor [Bothrops jararaca] 0.0
JARO001005B03 01 Deinagkistrodon acutus metalloproteinase MD2 precursor 1e-112
JARO001005B07 01 Metalloproteinase [Gloydius halys brevicaudus] 7e-91
JARO001005E08 01 Metalloproteinase precursor [Bothrops insularis] 2e-82
JARO001005F05 01 Insularinase and insularin precursor [Bothrops insularis] 2e-24
JARO001005H04 01 Crotalus scutulatus scutulatus GP-IV metalloproteinase precursor-like protein 7e-06
JARO001005H11 01 Bothropasin precursor [Bothrops jararaca] 1e-156
JARO001006A12 01 Bothrops jararaca bothrostatin precursor 5e-69
JARO001006C09 01 Bothropasin/jararhagin [Bothrops jararaca] 7e-38
JARO001006H09 01 Bothrops jararaca bothrostatin precursor 3e-33
JARO001006H12 01 Crotalus atrox prepro-hemorrhagic toxin d, atrolysin d 1e-147
JARO001008A08 01 Bothrops jararacussu metalloprotease BOJUMET II 0.0
JARO001008D05 01 Bothropasin precursor [Bothrops jararaca] 1e-178
JARO001008F11 01 Bothrops jararaca bothrostatin precursor 2e-64
JARO001008G06 01 Bothropasin precursor [Bothrops jararaca] 4e-66
JARO001008H03 01 Bothrops jararaca bothrostatin precursor 1e-19
JARO001009H07 01 Bothrops jararaca bothrostatin precursor 9e-83
JARO001009H08 01 Bothrops insularis cluster BITM06A (metalloproteinase precursor) 0.0
JARO001010E04 01 Bothropasin precursor [Bothrops jararaca] 0.0
JARO001010F04 01 Bothrops jararacussu metalloprotease BOJUMET II 1e-107
JARO001010F09 01 Bothropasin/jararhagin [Bothrops jararaca] 4e-26
JARO001010H11 01 Disintegrin jararacin [Bothrops jararaca] 9e-28
D.A.P. Cidade et al. / Toxicon 48 (2006) 437–461 441

ARTICLE IN PRESS
Table 2 (continued )
Cluster N Putative identification e-value
JARO001012G06 01 Bothrops insularis cluster BITM02A (metalloproteinase precursor) 4e-06
JARO001013A11 01 Bothropasin/jararhagin [Bothrops jararaca] 7e-14
JARO001013C11 01 Bothrops jararaca bothrostatin precursor 1e-122
JARO001013F04 01 Hemorrhagic metalloproteinase HF3 [Bothrops jararaca] e-114
JARO001013H04 01 Bothrops insularis cluster BITM02A (metalloproteinase precursor) 8e-08
JARO001014A02 01 Vascular apoptosis-inducing protein [Trimeresurus flavoviridis] 1e-89
JARO001014A05 01 Bothrops jararaca bothrostatin precursor 1e-59
JARO001014A09 01 Bothropasin/jararhagin [Bothrops jararaca] 1e-63
JARO001014B12 01 Insularinase and insularin precursor [Bothrops insularis] 2e-21
JARO001014F08 01 Bothropasin precursor [Bothrops jararaca] 0.0
JARO001014H02 01 Bothropasin/jararhagin [Bothrops jararaca] 8e-27
JARO001015B03 01 Bothropasin/jararhagin [Bothrops jararaca] 5e-24
JARO001015F10 01 Bothropasin precursor [Bothrops jararaca] 7e-97
JARO001015G09 01 Bothropasin/jararhagin [Bothrops jararaca] 2e-11
JARO001017D01 01 Bothropasin precursor [Bothrops jararaca] 1e-109
JARO001017F10 01 Atrolysin A—western diamondback rattlesnake 4e-07
JARO001017F12 01 Bothropasin/jararhagin [Bothrops jararaca] 3e-73
JARO001023B11 01 Bothrops jararaca bothrostatin precursor 1e-110
JARO001023C01 01 Bothrops jararaca hemorrhagic metalloproteinase HF3 1e-174
JARO001023D12 01 Bothrops jararacussu metalloprotease BOJUMET III 2e-51
JARO001023F01 01 Bothrops jararaca bothrostatin precursor 1e-106
JARO001023H11 01 Bothrops jararaca bothrostatin precursor 1e-122
JARO001031A08 01 Metalloprotease [Gloydius halys] 4e-52
JARO001032H05 01 Bothrops insularis cluster BITM02A (metalloproteinase precursor) 1e-129
JARO001033H10 01 Bothropasin precursor [Bothrops jararaca] e-101
JARO001034A08 01 Jararhagin precursor [Bothrops jararaca] e-110
JARO001035H12 01 Bothrops jararaca bothrostatin precursor 3e-41
JARO001036A07 01 Bothropasin precursor [Bothrops jararaca] 2e-35
JARO001036B03 01 Hemorrhagic metalloproteinase HF3 [Bothrops jararaca] 1e-18
JARO001036D03 01 Bothrops jararaca bothrostatin precursor 1e-108
JARO001036F06 01 Bothrops jararaca bothrostatin precursor 4e-42
JARO001036F11 01 Bothropasin precursor [Bothrops jararaca] 0.0
JARO001036G01 01 Hemorrhagic metalloproteinase HF3 [Bothrops jararaca] 3e-74
JARO001036G11 01 Bothrops jararaca bothrostatin precursor 1e-106
JARO001036H01 01 Bothropasin precursor [Bothrops jararaca] 2e-39
JARO001040A03 01 Bothrops jararaca bothrostatin precursor 3e-89
JARO001040E12 01 Western diamondback rattlesnake metalloproteinase preprometalloproteinase;(atrolysin e) 1e-48
JARO001041A05 01 Bothrops insularis cluster BITM02A (metalloproteinase precursor) 1e-155
JARO001041C01 01 Bothropasin precursor [Bothrops jararaca] 3e-98
JARO001041E12 01 Bothropasin/jararhagin [Bothrops jararaca] 5e-34
JARO001041F03 01 Deinagkistrodon acutus acurhagin precursor 1e-162
JARO001042H12 01 Bothropasin/jararhagin [Bothrops jararaca] 4e-06
JARO001043D02 01 Bothropasin/jararhagin [Bothrops jararaca] 1e-59
JARO001043D09 01 Bothrops jararaca bothrostatin precursor 7e-49
JARO001044E02 01 Bothrops jararaca bothrostatin precursor 1e-85
JARO001044F06 01 Bothrops jararaca bothrostatin precursor 1e-65
JARO001045E08 01 Hemorrhagic metalloproteinase HF3 [Bothrops jararaca] 9e-76
JARO001046A07 01 Bothropasin/jararhagin [Bothrops jararaca] 7e-10
JARO001046E04 01 Bothropasin/jararhagin [Bothrops jararaca] 1e-18
JARO001047A01 01 Deinagkistrodon acutus acurhagin precursor 1e-35
JARO001047A11 01 Bothropasin/jararhagin [Bothrops jararaca] 4e-50
JARO001047B11 01 Bothrops jararaca bothrostatin precursor 1e-128
JARO001047G03 01 Bothropasin precursor [Bothrops jararaca] 5e-83
JARO001047H08 01 Bothropasin precursor [Bothrops jararaca] 1e-160
JARO001048A03 01 Bothropasin/jararhagin [Bothrops jararaca] 4e-19
JARO001048C06 01 Bothropasin precursor [Bothrops jararaca] 5e-61
JARO001048C08 01 Hemorrhagic toxin a [Crotalus atrox] 6e-15
JARO001048D02 01 Bothrops jararaca bothrostatin precursor 1e-68
D.A.P. Cidade et al. / Toxicon 48 (2006) 437–461442

ARTICLE IN PRESS
Table 2 (continued )
Cluster N Putative identification e-value
JARO001048E07 01 Bothropasin precursor [Bothrops jararaca] 1e-35
JARO001048F04 01 Bothrops jararaca bothrostatin precursor 1e-18
JARO001048G03 01 Bothropasin/jararhagin [Bothrops jararaca] 2e-27
JARO001049A12 01 Bothropasin precursor [Bothrops jararaca] 1e-137
JARO001049B08 01 Bothrops jararaca bothrostatin precursor 8e-24
JARO001049C10 01 Bothropasin precursor [Bothrops jararaca] 2e-39
JARO001049E07 01 Bothrops jararaca bothrostatin precursor 2e-63
JARO001050C06 01 Bothropasin precursor [Bothrops jararaca] 2e-83
JARO001050G02 01 Agkistrodon contortrix contortrix contortrostatin precursor 4e-48
JARO001051D01 01 Metalloproteinase precursor [Bothrops insularis] 9e-12
JARO001051G02 01 Bothropasin precursor [Bothrops jararaca] 1e-150
JARO001052B03 01 Hemorrhagic metalloproteinase HF3 [Bothrops jararaca] 1e-83
JARO001052B07 01 Bothrops jararaca bothrostatin precursor 2e-89
JARO001052C06 01 Bothropasin precursor [Bothrops jararaca] 1e-155
JARO001052D07 01 Bothropasin/jararhagin [Bothrops jararaca] 7e-10
JARO001052E12 01 Bothrops jararaca bothrostatin precursor 1e-117
JARO001052H10 01 Bothrops jararaca bothrostatin precursor 4e-86
JARO001053A09 01 Bothropasin precursor [Bothrops jararaca] 7e-89
JARO001053D01 01 Bothropasin/jararhagin [Bothrops jararaca] e-108
JARO001053D07 01 Bothropasin/jararhagin [Bothrops jararaca] 8e-14
JARO001053D09 01 Bothrops insularis cluster BITM06A (metalloproteinase precursor) 9e-96
JARO001054C08 01 Atrolysin A—western diamondback rattlesnake 1e-79
JARO001054C11 01 Bothrops jararaca bothrostatin precursor 6e-71
JARO001054D02 01 Bothrops jararaca bothrostatin precursor 1e-38
JARO001055D07 01 Metalloproteina-se precursor [Bothrops insularis] 5e-20
JARO001056C03 01 Bothropasin/jararhagin [Bothrops jararaca] 2e-34
JARO001056D07 01 Bothropasin precursor [Bothrops jararaca] 1e-118
JARO001056E06 01 Bothropasin/jararhagin [Bothrops jararaca] 5e-65
JARO001057A02 01 Bothropasin precursor [Bothrops jararaca] 2e-57
JARO001057B04 01 Bothrops jararaca bothrostatin precursor 2e-73
Serine-proteinases
JARO001002G10 32 Macrovipera lebetina serine alpha-fibrinogenase precursor 0.0
JARO001001F08 23 Macrovipera lebetina serine alpha-fibrinogenase precursor 2e-31
JARO001004E09 23 Bothrops jararaca mRNA for KN-BJ2 2e-78
JARO001002E07 13 Serine proteinase precursor [Bothrops insularis] e-111
JARO001001G01 11 Bothrops jararaca mRNA for KN-BJ2 1e-36
JARO001001E03 09 Bothrops jararaca mRNA for protease A 0.0
JARO001005B12 08 Macrovipera lebetina serine alpha-fibrinogenase precursor 2e-31
JARO001010A10 07 Macrovipera lebetina serine alpha-fibrinogenase precursor 4e-31
JARO001012H07 07 Trimeresurus gramineus mRNA for serine protease 0.0
JARO001002B10 06 Macrovipera lebetina serine alpha-fibrinogenase precursor 3e-07
JARO001005E06 06 Bothrops jararaca mRNA for KN-BJ2 e-151
JARO001006E08 06 Bothrops jararaca mRNA for KN-BJ2 8e-22
JARO001032H12 06 Macrovipera lebetina serine alpha-fibrinogenase precursor 3e-36
JARO001002C08 05 Macrovipera lebetina serine alpha-fibrinogenase precursor 2e-31
JARO001002C09 05 Bothrops jararaca mRNA for KN-BJ2 7e-28
JARO001004B09 05 Serine protease catroxase II precursor [Crotalus atrox] 1e-15
JARO001012D05 05 Macrovipera lebetina serine alpha-fibrinogenase precursor 2e-31
JARO001013E02 05 Bothrops jararaca mRNA for KN-BJ2 2e-62
JARO001041H08 05 Bothrops jararacussu serine protease 0.0
JARO001004A10 04 Macrovipera lebetina serine alpha-fibrinogenase precursor 1e-31
JARO001004C10 04 Bothrops jararacussu serine protease 0.0
JARO001006A11 04 Bothrops jararaca mRNA for KN-BJ2 2e-64
JARO001010A06 04 Bothrops jararaca mRNA for KN-BJ2 e-128
JARO001017E10 04 Macrovipera lebetina serine alpha-fibrinogenase precursor 9e-29
JARO001033G09 04 Bothrops jararaca mRNA for KN-BJ2 e-104
JARO001035B07 04 Macrovipera lebetina serine alpha-fibrinogenase precursor 2e-26
D.A.P. Cidade et al. / Toxicon 48 (2006) 437–461 443

ARTICLE IN PRESS
Table 2 (continued )
Cluster N Putative identification e-value
JARO001043F02 04 Macrovipera lebetina serine alpha-fibrinogenase precursor 1e-34
JARO001005A05 03 Macrovipera lebetina serine alpha-fibrinogenase precursor 1e-31
JARO001005H09 03 Bothrops jararaca mRNA for KN-BJ2 9e-15
JARO001008E04 03 Macrovipera lebetina serine alpha-fibrinogenase precursor 1e-31
JARO001008H09 03 Bothrops jararaca mRNA for KN-BJ2 3e-36
JARO001009A11 03 Macrovipera lebetina serine alpha-fibrinogenase precursor 1e-28
JARO001045G06 03 Acubin [Deinagkistrodon acutus] 5e-57
JARO001001C07 02 Macrovipera lebetina serine alpha-fibrinogenase precursor 1e-31
JARO001001F11 02 Bothrops atrox batroxobin gene 1e-24
JARO001001G04 02 Macrovipera lebetina serine alpha-fibrinogenase precursor 1e-34
JARO001002D06 02 Venom serine proteinase A precursor 7e-43
JARO001002D07 02 Serine protease [Bothrops jararacussu] 2e-80
JARO001003E11 02 Macrovipera lebetina serine alpha-fibrinogenase precursor 4e-34
JARO001005B11 02 Bothrops jararaca mRNA for KN-BJ2 1e-50
JARO001008F03 02 Bothrops jararaca mRNA for KN-BJ2 6e-09
JARO001009B02 02 Macrovipera lebetina serine alpha-fibrinogenase precursor 3e-22
JARO001009G08 02 Bothrops jararaca mRNA for KN-BJ2 e-106
JARO001010G07 02 Bothrops jararaca mRNA for KN-BJ2 5e-54
JARO001012F02 02 Macrovipera lebetina serine alpha-fibrinogenase precursor 1e-31
JARO001013E10 02 Macrovipera lebetina serine alpha-fibrinogenase precursor 3e-30
JARO001031B04 02 Acubin2 [Deinagkistrodon acutus] 5e-44
JARO001031B08 02 Bothrops jararaca mRNA for KN-BJ2 3e-10
JARO001034G10 02 Macrovipera lebetina serine alpha-fibrinogenase precursor 2e-09
JARO001040C05 02 Bothrops jararaca mRNA for KN-BJ2 2e-50
JARO001041G06 02 Macrovipera lebetina serine alpha-fibrinogenase precursor 2e-29
JARO001043E08 02 JH3C12F Snake Bothrops insularis serine proteinase 3e-23
JARO001044F02 02 Macrovipera lebetina serine alpha-fibrinogenase precursor 4e-18
JARO001046A06 02 Serine protease catroxase II precursor [Crotalus atrox] 2e-77
JARO001001B05 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 9e-07
JARO001001C08 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 1e-27
JARO001001E06 01 Bothrops jararaca mRNA for KN-BJ2 2e-10
JARO001001E12 01 Bothrops insularis cluster BITS01A serine proteinase precursor 1e-130
JARO001002B07 01 Bothrops jararaca mRNA for KN-BJ2 1e-104
JARO001002E08 01 JH3C12F Snake Bothrops insularis serine protease 2e-05
JARO001002F12 01 Bothrops jararaca mRNA for KN-BJ2 0.0
JARO001005C07 01 Bothrops jararaca mRNA for KN-BJ2 2e-14
JARO001005E03 01 Bothrops insularis cluster BITS01A serine proteinase precursor 1e-106
JARO001006A08 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 1e-31
JARO001006C08 01 Serine protease catroxase II precursor [Crotalus atrox] ] 4e-53
JARO001008B11 01 Bothrops insularis cluster BITS01A serine proteinase precursor 2e-94
JARO001008C06 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 4e-27
JARO001008H05 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 9e-23
JARO001009D11 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 3e-06
JARO001009G01 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 5e-20
JARO001010A07 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 1e-05
JARO001010B11 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 5e-11
JARO001012B07 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 1e-24
JARO001014A08 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 3e-26
JARO001014B07 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 4e-15
JARO001014C08 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 6e-39
JARO001015A05 01 Bothrops jararaca mRNA for protease A 1e-84
JARO001015A06 01 Venom serine proteinase A precursor 6e-36
JARO001015B06 01 Bothrops jararaca mRNA for KN-BJ2 3e-62
JARO001017C05 01 Trimeresurus flavoviridis mRNA for serine protease 1e-131
JARO001017D12 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 8e-32
JARO001023D03 01 Bothrops jararacussu serine protease 1e-139
JARO001023E07 01 Serine protease [Bothrops jararacussu] 3e-40
JARO001023F12 01 Bothrops jararaca mRNA for protease A 0.0
D.A.P. Cidade et al. / Toxicon 48 (2006) 437–461444

ARTICLE IN PRESS
Table 2 (continued )
Cluster N Putative identification e-value
JARO001023H05 01 Bothrops jararacussu serine protease 1e-178
JARO001031C05 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 2e-14
JARO001031D07 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 2e-16
JARO001031E02 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 6e-39
JARO001031H12 01 Bothrops jararaca mRNA for KN-BJ2 1e-134
JARO001033F12 01 Bothrops jararaca mRNA for KN-BJ2 0.0
JARO001034D01 01 Bothrops jararaca mRNA for protease A 5e-57
JARO001034G01 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 3e-16
JARO001035B09 01 Bothrops jararaca mRNA for protease A 0.0
JARO001035C09 01 Venom serine proteinase A precursor 4e-19
JARO001035G10 01 JH3C12F Snake Bothrops insularis serine protease 3e-25
JARO001036C12 01 Trimeresurus gramineus mRNA for serine protease 1e-177
JARO001036D01 01 Bothrops jararaca mRNA for KN-BJ2 2e-67
JARO001036F10 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 1e-14
JARO001040G05 01 Bothrops jararaca mRNA for KN-BJ2 3e-30
JARO001041C06 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 3e-31
JARO001041D12 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 1e-18
JARO001041H03 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 9e-11
JARO001042A12 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 3e-29
JARO001042D07 01 Bothrops insularis cluster BITS01A serine proteinase precursor 1e-152
JARO001042F12 01 Bothrops jararaca mRNA for KN-BJ2 1e-64
JARO001042H08 01 Bothrops jararaca mRNA for protease A 2e-80
JARO001043B07 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 5e-32
JARO001043E01 01 Bothrops jararaca mRNA for KN-BJ2 2e-44
JARO001043F06 01 Bothrops jararaca mRNA for KN-BJ2 8e-35
JARO001044E06 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 2e-16
JARO001044F05 01 Bothrops jararaca mRNA for protease A 0.0
JARO001045E05 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 1e-33
JARO001046A04 01 Bothrops jararaca mRNA for KN-BJ2 3e-35
JARO001046D10 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 1e-16
JARO001046E11 01 Bothrops jararaca mRNA for KN-BJ2 1e-121
JARO001046F04 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 8e-29
JARO001046H06 01 Bothrops jararaca mRNA for KN-BJ2 4e-39
JARO001047E10 01 Bothrops jararaca mRNA for KN-BJ2 0.0
JARO001048B03 01 Bothrops jararaca mRNA for KN-BJ2 1e-14
JARO001048D12 01 Bothrops jararaca mRNA for protease A 0.0
JARO001049A02 01 Bothrops jararacussu serine protease 1e-152
JARO001049C12 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 6e-27
JARO001050G01 01 Bothrops jararaca mRNA for KN-BJ2 2e-95
JARO001050G10 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 1e-21
JARO001051A10 01 JH3C12F Snake Bothrops insularis serine proteinase 1e-13
JARO001051B02 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 7e-11
JARO001051C04 01 Bothrops jararaca mRNA for KN-BJ2 5e-52
JARO001051C12 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 1e-15
JARO001051D05 01 Bothrops insularis cluster BITS01A serine proteinase precursor 1e-169
JARO001051E03 01 Bothrops jararaca mRNA for protease A 1e-104
JARO001051E11 01 Bothrops jararaca mRNA for KN-BJ2 2e-76
JARO001052F01 01 Bothrops jararaca mRNA for KN-BJ2 8e-81
JARO001052G04 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 4e-26
JARO001052G07 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 1e-21
JARO001053A05 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 1e-27
JARO001053G04 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 1e-31
JARO001054D11 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 2e-27
JARO001055E10 01 Trimeresurus gramineus mRNA for serine protease 1e-158
JARO001056B12 01 Trimeresurus gramineus mRNA for serine protease 0.0
JARO001056H07 01 Macrovipera lebetina serine alpha-fibrinogenase precursor 8e-11
JARO001057B12 01 Bothrops insularis cluster BITS01A serine proteinase precursor 1e-78
D.A.P. Cidade et al. / Toxicon 48 (2006) 437–461 445

ARTICLE IN PRESS
Table 2 (continued )
Cluster N Putative identification e-value
C-Type lectins
JARO001004F10 16 Platelet glycoprotein Ib-binding protein alpha subunit 1e-73
JARO001005B10 13 Platelet glycoprotein Ib-binding protein beta subunit 3e-69
JARO001009F12 09 Chain A Coagulation Factor Ix-Binding Protein 6e-55
JARO001015B10 08 ACF 1/2 B-chain [Deinagkistrodon acutus] 2e-61
JARO001005B08 07 bothojaracin chain A precursor [Bothrops jararaca] 8e-65
JARO001006G10 04 Platelet glycoprotein Ib-binding protein beta subunit 2e-66
JARO001002C10 03 Botrocetin beta chain [Bothrops jararaca] 1e-15
JARO001005F08 03 Bothojaracin chain B precursor [Bothrops jararaca] 1e-75
JARO001006B10 03 bothojaracin chain A precursor [Bothrops jararaca] 2e-78
JARO001012D04 03 bothojaracin chain B precursor [Bothrops jararaca] 6e-55
JARO001013F07 03 bothojaracin chain A precursor [Bothrops jararaca] 2e-78
JARO001017D09 03 Platelet glycoprotein Ib-binding protein beta subunit 3e-17
JARO001013E08 02 Mamushigin beta [Agkistrodon blomhoffi] 1e-50
JARO001044G03 02 Bothojaracin chain A precursor [Bothrops jararaca] 2e-27
JARO001001D10 01 Flavocetin-A beta chain [Trimeresurus flavoviridis] 4e-21
JARO001005G05 01 Botrocetin alpha chain [Bothrops jararaca] 7e-26
JARO001010D04 01 Bothojaracin chain B precursor [Bothrops jararaca] 3e-65
JARO001014B11 01 Bothojaracin chain A precursor [Bothrops jararaca] 6e-50
JARO001014E06 01 Bothojaracin chain A precursor [Bothrops jararaca] precursor 6e-25
JARO001015C03 01 Stejaggregin-A alpha chain [Trimeresurus stejnegeri] 2e-63
JARO001031D04 01 Bothojaracin chain B precursor [Bothrops jararaca] 1e-42
JARO001031H02 01 Bothojaracin chain A precursor [Bothrops jararaca] 8e-52
JARO001033A01 01 Bothojaracin chain B precursor [Bothrops jararaca] mucrosquamatus] 7e-06
JARO001035E08 01 Trimeresurus stejnegeri factor IX/X binding protein beta chain- 6e-43
JARO001036B08 01 Platelet glycoprotein Ib-binding protein beta subunit 2e-11
JARO001036D04 01 Coagulation factor IX/X-binding protein: A chain 1e-32
JARO001040C04 01 Platelet glycoprotein Ib-binding protein beta subunit 1e-10
JARO001040D10 01 Bothojaracin chain B precursor [Bothrops jararaca]] 8e-15
JARO001041G05 01 Coagulation factor IX-binding protein chain A [Gloydius halys] 6e-43
JARO001042E12 01 Botrocetin beta chain [Bothrops jararaca] 9e-18
JARO001043F04 01 Factor IX/factor X binding protein B chain [Trimeresurus flavoviridis] 5e-59
JARO001046G07 01 Bothojaracin chain A precursor [Bothrops jararaca] 3e-30
JARO001047B07 01 Bothojaracin chain A precursor [Bothrops jararaca] 1e-56
JARO001049E08 01 ACF 1/2 B-chain [Deinagkistrodon acutus] 8e-36
JARO001049F10 01 Trimeresurus stejnegeri factor IX/X binding protein alpha chain 8e-31
JARO001050C08 01 Platelet glycoprotein Ib-binding protein beta subunit 2e-61
JARO001050C09 01 Gloydius halys coagulation factor IX-binding protein chain B 7e-18
JARO001050F08 01 Trimeresurus flavoviridis flavocetin-A alpha chain 2e-29
JARO001054F10 01 C-type lectin [Deinagkistrodon acutus] 1e-42
JARO001054F12 01 Coagulation factor IX/X-binding protein: A chain 3e-29
JARO001056B10 01 Platelet glycoprotein Ib-binding protein alpha subunit 4e-28
Bradykinin-potentiating peptide precursors
JARO001009B06 16 B. insularis cluster BITB08 bradykinin-potentiating proteinprecursor 0.0
JARO001013A07 14 Bothrops jararaca bradykinin-potentiating/C-type natriuretic peptide 0.0
JARO001001A07 09 Bothrops jararaca bradykinin-potentiating/C-type natriuretic peptide e-119
JARO001014H12 09 Agkistrodon blomhoffi mRNA for BPP-CNP precursor e-110
JARO001003G09 06 Bothrops jararaca bradykinin-potentiating/C-type natriuretic peptide 1e-47
JARO001047D04 03 Bradykinin-potentiating peptides and C-type natriuretic peptide precursor [Bothrops jararaca] 3e-82
JARO001002E11 02 Bothrops jararaca bradykinin-potentiating/C-type natriuretic peptide e-104
JARO001010H03 02 Bothrops jararaca mRNA for bradykinin-potentiating peptide and C-type natriuretic peptide precursor 0.0
JARO001014E08 02 Bradykinin-potentiating peptides and C-type natriuretic peptide precursor [Bothrops jararaca] 6e-53
JARO001045B02 02 Bradykinin-potentiating peptides and C-type natriuretic peptide precursor [Bothrops jararaca] 2e-34
JARO001002C11 01 Bothrops jararaca bradykinin-potentiating/C-type natriuretic peptide 1e-94
JARO001006F09 01 Bothrops jararaca bradykinin-potentiating/C-type natriuretic peptide 5e-98
JARO001008A10 01 Bradykinin-potentiating/C-type natriuretic peptide [Bothrops jararaca] 6e-48
JARO001009H05 01 Bothrops insularis cluster BITB08 bradykinin-potentiating protein precursor 1e-108
D.A.P. Cidade et al. / Toxicon 48 (2006) 437–461446

ARTICLE IN PRESS
Table 2 (continued )
Cluster N Putative identification e-value
JARO001012E09 01 Bradykinin-potentiating peptides and C-type natriuretic peptide precursor [Bothrops jararaca] 3e-54
JARO001023A11 01 Bothrops insularis cluster BITB08 bradykinin-potentiating protein precursor 1e-116
JARO001023D07 01 Bothrops jararaca mRNA for bradykinin-potentiating peptide and C-type natriuretic peptide precursor 0.0
JARO001031F10 01 Bothrops jararaca bradykinin-potentiating/C-type natriuretic peptide 2e-99
JARO001031G01 01 Bothrops insularis cluster BITB08 bradykinin-potentiating protein precursor 1e-116
JARO001033G07 01 Bothrops jararacussu bradykinin-potentiating/C-type natriuretic peptide 1e-103
JARO001040E07 01 Bothrops insularis bradykinin-potentiating/c-type natriuretic protein precursor 1e-31
JARO001047G12 01 Bothrops jararaca bradykinin-potentiating/C-type natriuretic peptide 4e-73
JARO001052C04 01 Bradykinin-potentiating peptides and C-type natriuretic peptide precursor [Bothrops jararaca 1e-23
JARO001053F02 01 Bothrops jararaca bradykinin-potentiating/C-type natriuretic peptide 1e-178
JARO001054E08 01 Bothrops jararaca bradykinin-potentiating/C-type natriuretic peptide 8e-27
Phospholipases A2
JARO001034D12 02 Snake BP-II gene for phospholipase A2 3e-11
JARO001001G11 01 Snake BP-II gene for phospholipase A2 1e-90
JARO001006F07 01 Myotoxic phospholipase A2-like [Bothrops jararacussu] 2e-68
JARO001015A02 01 Vipera berus berus phospholipase A2 gene 5.0e-06
JARO001033F11 01 Gloydius shedaoensis acidic phospholipase A2 6e-65
JARO001045H10 01 Trimeresurus gramineus gene for phospholipase A2 2e-25
JARO001050F01 01 Vipera berus berus phospholipase A2 gene 5e-06
JARO001053F03 01 Phospholipase A2 precursor [Canis familiaris] 3e-11
Phospholipase A2 inhibitors
JARO001031A02 03 Phospholipase A2 inhibitor [Trimeresurus flavoviridis] 4e-07
JARO001009C01 02 Phospholipase A2 inhibitory protein Annexin A1 5e-34
JARO001005C10 01 Phospholipase A2 inhibitor [Trimeresurus flavoviridis] 2e-14
JARO001009A03 01 Phospholipase A2 inhibitor [Trimeresurus flavoviridis] 7e-11
JARO001010C09 01 Phospholipase A2 inhibitor [Trimeresurus flavoviridis] 6e-15
JARO001043A01 01 Phospholipase A2 inhibitor [Trimeresurus flavoviridis] 2e-26
JARO001045C10 01 Phospholipase A2 inhibitor [Trimeresurus flavoviridis] 2e-42
JARO001052F12 01 Phospholipase A2 inhibitor [Elaphe quadrivirgata] 2e-20
JARO001055F03 01 Phospholipase A2 inhibitor [Trimeresurus flavoviridis] 2e-31
Cysteine-rich venom protein precursors
JARO001006G08 08 Trimeresurus stejnegeri cysteine-rich secretory protein 0.0
JARO001006E12 05 Catrin [Crotalus atrox] 6e-99
JARO001009A05 01 Trimeresurus stejnegeri cysteine-rich secretory protein 3e-89
JARO001009B05 01 Trimeresurus stejnegeri cysteine-rich secretory protein 8e-96
JARO001036C10 01 Ablomin [Agkistrodon blomhoffi] 7e-16
JARO001045A07 01 Catrin [Crotalus atrox] 2e-33
JARO001046B09 01 Piscivorin [Agkistrodon piscivorus piscivorus] 1e-74
JARO001049G12 01 Trimeresurus mucrosquamatus prepro-cysteine-rich venom protein 4e-39
JARO001054F11 01 Catrin [Crotalus atrox] 8e-42
Other
JARO001034A06 01 Vascular endothelial growth factor [B. erythromelas] 2e-89
JARO001040H08 01 Vascular endothelial growth factor [B. insularis] 3e-47
JARO001034F04 04 L-amino acid oxidase [Trimeresurus stejnegeri] 1e-05
JARO001003E08 03 FAD-containing L-amino acid oxidase Apoxin 1 [Crotalus atrox] 6e-21
D.A.P. Cidade et al. / Toxicon 48 (2006) 437–461 447
3.3. Analysis of toxin clusters
The identification of some of the toxin-relatedclusters was not possible at the amino acid level, andcould only be accomplished by a Blast search
against the nucleotide database (Blast N). This isdue to the particularly long 30 untranslated region(UTR) of toxins, in particular of the serineproteinases, of which most of the clusters wereidentified by Blast N. The representation of each

ARTICLE IN PRESS
Table 3
Identification of putative cellular protein clusters from B. jararaca venom
Cluster N Putative identification e-value
General metabolism
JARO001015D08 03 Cytochrome oxidase subunit II [Dinodon semicarinatus] 3e-51
JARO001034A03 03 Mannosyl-oligosaccharide 1,2-alpha-mannosidase IC [Gallus gallus] e-104
JARO001043G05 03 UDP-N-acetylhexosamine pyrophosphorylase 2e-07
JARO001002G05 02 Asparagine synthase (glutamine-hydrolysing) [Gallus gallus] e-113
JARO001004D09 02 Cytochrome b [Crotalus viridis] e-140
JARO001010H05 02 Propionyl Coenzyme A carboxylase, beta polypeptide [Rattusnorvegicus] 5e-11
JARO001013H07 02 Cytochrome C oxidase copper chaperone [Ophiophagus hannah] 8e-25
JARO001032A10 02 Malate dehydrogenase, [Gallus gallus] 9e-41
JARO001033C09 02 Alpha 3 glucosyltransferase [Gallus gallus] 8e-59
JARO001002E09 01 Creatine kinase [Zaocys dhumnades] 5e-79
JARO001003D07 01 UDP-glucose pyrophosphorylase [Gallus gallus] e-104
JARO001003D08 01 Methionine aminopeptidase 2 4e-57
JARO001004B08 01 Cytochrome oxidase subunit III [Dinodon semicarinatus] 3e-70
JARO001005E12 01 Glutaminyl cyclase [Bothrops jararaca] 8e-21
JARO001009F07 01 ATPase subunit 6 [Crotalus viridis] 3e-14
JARO001010B02 01 Tyrosine 3/tryptophan 5 -monooxygenase activation protein [Homo sapiens] 2e-29
JARO001017A03 01 Cytochrome b [Crotalus viridis] 1e-117
JARO001023C02 01 Ornithine-oxo-acid aminotransferase [Gallus gallus] 2e-64
JARO001031A01 01 Cytochrome oxidase subunit I [Dinodon semicarinatus] 5e-26
JARO001031G07 01 Cytochrome c oxidase subunit VIb [Tarsius syrichta] 8e-31
JARO001031H09 01 Creatine kinase [Zaocys dhumnades] 1e-47
JARO001033E12 01 Glyceraldehyde-3-phosphate dehydrogenase [Pelodiscus sinensis] 3e-84
JARO001033G12 01 Glutaminyl cyclase [Bothrops jararaca] 5e-72
JARO001034B02 01 Creatine kinase [Zaocys dhumnades] 2e-47
JARO001035C06 01 GDP-mannose 4,6-dehydratase [Gallus gallus] 1e-13
JARO001042C12 01 ADP-ribosylation factor 1 [Homo sapiens] 7e-25
JARO001043D05 01 Glutathione peroxidase [Gallus gallus] 1e-36
JARO001043D08 01 Glyoxylate reductaseHomo sapiens] 6e-07
JARO001044B01 01 Inorganic pyrophosphatase (Pyrophosphate phospho-hydrolase) 5e-84
JARO001044B02 01 Aspartate aminotransferase, cytoplasmic (Transaminase A) 6e-93
JARO001046C04 01 Adenylate kinase 3 [Gallus gallus] 7e-59
JARO001048G12 01 Aldehyde dehydrogenase [Gallus gallus] 2e-30
JARO001050C10 01 Aldehyde dehydrogenase [Gallus gallus] 5e-35
JARO001052C02 01 Tyrosine phosphatase protein [Macaca fascicularis] 1e-10
JARO001052H05 01 Iron storage protein H-ferritin [Trichosurus vulpecula] 3e-53
JARO001053A11 01 Alpha 3 glucosyltransferase [Gallus gallus] 3e-59
JARO001053C05 01 Transitional endoplasmic reticulum ATPase 8e-60
JARO001053F11 01 ATPase subunit 9 1e-18
JARO001054D06 01 NADH dehydrogenase subunit 1 [Naja nivea] 1e-58
JARO001055B05 01 Tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein, theta polypeptide 1e-37
JARO001055C10 01 ATPase subunit 6 [Crotalus viridis] 6e-23
JARO001055E12 01 Dipeptidylpeptidase 4a [Gloydius blomhoffi brevicaudus] 4e-07
DNA transcription and RNA translation
JARO001002G08 05 Ribosomal protein L27a [Mus musculus] 3e-65
JARO001015G02 05 Ribosomal protein S3 [Xenopus tropicalis] e-119
JARO001002E10 04 Ribosomal protein 10 [Mus musculus] e-121
JARO001013E01 04 Ribosomal protein L45 [Mus musculus] 2e-41
JARO001023H03 04 Eukaryotic translation initiation factor 3 subunit 2 [Gallus gallus] e-133
JARO001001B02 03 Ribosomal protein S12 [Gallus gallus] 6e-71
JARO001003F12 03 Poly(A) binding protein [Homo sapiens] 3e-28
JARO001014C04 03 40S ribosomal protein S16 [Homo sapiens 1e-77
JARO001032G07 03 Heterogeneous nuclear ribonucleoprotein U isoform b [Gallus gallus] 2e-85
JARO001001D01 02 Eukaryotic translation elongation factor 1 alpha 1 [Gallus gallus] 6e-57
JARO001005G08 02 RNA helicase [Homo sapiens] 9e-74
JARO001010F03 02 Eukaryotic translation elongation factor 1 gamma [Homo sapiens] 6e-39
D.A.P. Cidade et al. / Toxicon 48 (2006) 437–461448

ARTICLE IN PRESS
Table 3 (continued )
Cluster N Putative identification e-value
JARO001012H09 02 40S ribosomal protein S8 [Gallus gallus] e-114
JARO001014G11 02 40S ribosomal protein S14 [Ictalurus punctatus] 2e-70
JARO001017E04 02 Ribosomal protein S3a [Ophiophagus hannah] 1e-88
JARO001017G06 02 Ribosomal protein L30 [Gallus gallus] 8e-53
JARO001032C04 02 60S acidic ribosomal protein P1 [Gallus gallus] 6e-31
JARO001040G03 02 Ophiophagus hannah ribosomal protein L30 2e-09
JARO001042D04 02 Eukaryotic translation initiation factor 3, subunit 7 (zeta) [Danio rerio] 2e-33
JARO001051A12 02 Poly-A binding protein [Gallus gallus] 9e-49
JARO001052G06 02 Ribosomal protein L19 [Macaca fascicularis] 1e-65
JARO001001A08 01 Ribosomal protein L4 [Gallus gallus] 7e-23
JARO001001D09 01 Eukaryotic translation elongation factor 1 alpha 1 [Gallus gallus] 1e-94
JARO001001G10 01 Ribosomal protein [Gallus gallus] 2e-29
JARO001001H07 01 40S ribosomal protein S3 [Gallus gallus] 6e-09
JARO001003E09 01 High mobility group protein B1[Gallus gallus] 3e-26
JARO001004B10 01 Ribosomal protein L23 [Pan troglodytes] 5e-21
JARO001006D09 01 Ribosomal protein L18a [Homo sapiens] 3e-58
JARO001009G04 01 TATA-box binding protein[Trimeresurus gramineus] 5e-06
JARO001010D06 01 Splicing factor U2AF 65 kDa subunit 7e-13
JARO001012B09 01 60S ribosomal protein L27 9e-31
JARO001012H01 01 60S acidic ribosomal protein P1 6e-25
JARO001014A04 01 Ribosomal protein L45 [Mus musculus] 3e-38
JARO001014D04 01 Ribosomal protein L23 [Pan troglodytes] 5e-62
JARO001017A06 01 Smu-1 suppressor of mec-8 and unc-52 homolog [Homo sapiens] 9e-50
JARO001017C07 01 Eukaryotic translation initiation factor 3 subunit 2 [Gallus gallus] 1e-15
JARO001031C12 01 60S ribosomal protein L10 [Homo sapiens] 4e-08
JARO001031D03 01 Eukaryotic translation initiation factor 3, subunit 9 [Homo sapiens] 2e-31
JARO001032A06 01 Ribosomal protein L10a [Rattus norvegicus] 4e-18
JARO001032B08 01 Gallus gallus ribonucleoprotein 1e-95
JARO001033B11 01 Ribosomal protein S27 [Pan troglodytes] 6e-38
JARO001033F02 01 60S ribosomal protein L28 [Hippocampus comes 5e-25
JARO001041A04 01 Helicase 1 (matrix associated, actin-dependent regulator of chromatin subfamily A) 1e-58
JARO001041H11 01 Eukaryotic translation initiation factor 3 [Danio rerio] 3e-33
JARO001042E10 01 Homo sapiens poly(A) binding protein 9e-18
JARO001043B03 01 Eukaryotic translation elongation factor 1 gamma [Homo sapiens] 2e-44
JARO001043D04 01 40S ribosomal protein S5 [Rattus norvegicus] 1e-80
JARO001043D11 01 Small nuclear ribonucleoprotein polypeptide A’ [Homo sapiens] 2e-19
JARO001043E03 01 DNA directed RNA polymerase II polypeptide K [Homo sapiens] 5e-25
JARO001043H10 01 Elongation factor 2 5e-91
JARO001044D12 01 Homo sapiens nucleolin 3e-07
JARO001045C08 01 H3 histone family, member I [Rattus norvegicus] 3e-41
JARO001045F04 01 60S ribosomal protein L36a 5e-52
JARO001046D07 01 Ribosomal protein S8 [Scyliorhinus canicula] 2e-35
JARO001046F06 01 Ribosomal protein S4 1e-20
JARO001050B05 01 Ribosomal protein S27 [Homo sapiens] 2e-38
JARO001051D08 01 Branchiostoma belcheri ribosomal protein L22 5e-06
JARO001053B05 01 60S ribosomal protein L13 [Pan troglodytes] 4e-41
JARO001055A08 01 Small nuclear ribonucleoprotein polypeptide G [Homo sapiens] 1e-29
JARO001055D01 01 Homo sapiens eukaryotic translation initiation factor 1A 1e-51
JARO001055F06 01 RNA binding protein (signal recognition particle 14kDa) 2e-26
JARO001055H10 01 Bcl-2-associated transcription factor shortform [Gallus gallus] 7e-95
JARO001056B08 01 Gallus gallus ribonucleoprotein 1e-106
JARO001056E08 01 Splicing factor [Mus musculus] e-110
JARO001057C01 01 Ribosomal protein S11 [Gallus gallus] 3e-78
JARO001057C03 01 DNA dependent ATPase and helicase [Homo sapiens] 6e-86
JARO001057G04 01 Ribosomal protein [Homo sapiens] 6e-70
JARO001057H04 01 HIV TAT specific factor 1 [Homo sapiens] 3e-28
D.A.P. Cidade et al. / Toxicon 48 (2006) 437–461 449

ARTICLE IN PRESS
Table 3 (continued )
Cluster N Putative identification e-value
Pos-translational processing and sorting
JARO001001E02 34 Protein disulfide-isomerase 4e-77
JARO001002H12 10 Protein disulfide isomerase A4 precursor [Gallus gallus] 2e-62
JARO001010B08 07 Protein disulfide-isomerase isomerase A6 precursor [Gallus gallus] 4e-60
JARO001005F10 05 JH4B01R Snake Bothrops insularis calreticulin precursor e-142
JARO001001A01 04 Protein disulfide-isomerase precursor 7e-87
JARO001014D01 03 JH4B01R Snake Bothrops insularis calreticulin precursor 1e-90
JARO001008F08 02 Protein disulfide isomerase 3e-44
JARO001010D07 02 Translocation protein SEC63 homolog [Gallus gallus] 8e-30
JARO001014G05 02 S100 calcium binding protein A6 (calcyclin) [Gallus gallus] 1e-18
JARO001014H04 02 Sec61 alpha subunit homolog [Gallus gallus] 7e-92
JARO001023B10 02 Protein disulfide-isomerase 4e-24
JARO001032C08 02 ER-associated dnaJ protein 3 [Gallus gallus] e-110
JARO001033B08 02 Protein disulfide isomerase A4 precursor [Gallus gallus] 1e-79
JARO001050D01 02 JH4B01R Snake Bothrops insularis calreticulin precursor e-129
JARO001001B04 01 Glucose regulated thiol oxido-reductase protein precursor [Gallus gallus] 1e-34
JARO001005G02 01 Protein disulfide-isomerase 4e-63
JARO001009F04 01 Chaperonin subunit 8 (theta) [Mus musculus] 5e-06
JARO001012D02 01 Clathrin-associated/assembly/adapter protein 2e-68
JARO001013A01 01 JH4B01R Snake Bothrops insularis calreticulin precursor 1e-116
JARO001023B05 01 GH053F Snake Bothrops insularis disulfide-isomerase 5e-11
JARO001023D06 01 Coatomer zeta-1 subunit [Rattus norvegicus] 7e-84
JARO001023G12 01 Protein disulfide isomerase A4 precursor [Gallus gallus] 3e-58
JARO001032D12 01 Calreticulin [Gallus gallus] 1e-19
JARO001034E09 01 Kinectin (kinesin-binding protein) 2e-59
JARO001035A08 01 GH053F Snake Bothrops insularis disulfide-isomerase 3e-44
JARO001035F07 01 GH053F Snake Bothrops insularis disulfide-isomerase 4e-09
JARO001042G01 01 GH053F Snake Bothrops insularis disulfide-isomerase 3e-48
JARO001045A02 01 GH011R Snake Bothrops insularis calcium binding protein 1e-128
JARO001046A08 01 Protein disulfide-isomerase [Cricetulus griseus] 4e-22
JARO001047D11 01 Protein disulfide-isomerase 4e-28
JARO001047G06 01 Protein disulfide-isomerase 7e-73
JARO001047G07 01 GH053F Snake Bothrops insularis disulfide-isomerase 3e-13
JARO001048H08 01 Plasma glutathione peroxidase precursor [Rattus norvegicus] 4e-52
JARO001049B07 01 GH053F Snake Bothrops insularis disulfide-isomerase 3e-84
JARO001051B11 01 Vesicle-fusing ATPase protein 5e-29
JARO001054B08 01 JH4D03F Snake Bothrops insularis Calreticulin precursor 2e-74
JARO001054H12 01 Transport protein SEC61 gamma subunit 1e-10
JARO001056E09 01 Glutathione reductase 1 [Mus musculus] 4e-41
Polypeptide degradation
JARO001009B03 03 SMT3 suppressor of mif two 3 homolog 2 [Homo sapiens] 6e-49
JARO001014G07 03 Proteasome beta 3 subunit [Homo sapiens] e-107
JARO001032A08 02 Dipeptidylpeptidase 4a [Gloydius blomhoffi brevicaudus] 2e-09
JARO001008G08 01 Ubiquitin carboxyl-terminal hydrolase [Gallus gallus] 1e-43
JARO001009B11 01 ubiquitin [Oncorhynchus mykiss] 1e-46
JARO001034F11 01 Ubiquitin-like protein/ribosomal protein S30 7e-21
JARO001042F08 01 Proteasome subunit alpha type 1 3e-20
JARO001043H09 01 Ubiquitin specific protease 7 [Mus musculus] 1e-09
JARO001052E04 01 Proteasome 26S subunit, ATPase 3 [Mus musculus] 4e-43
Structural functions
JARO001012H05 03 Beta-actin [Tupaia belangeri] 1e-25
JARO001003C08 02 Myosin light chain [Oxyuranus scutellatus scutellatus] 1e-92
JARO001003F07 02 Actin-related protein 6 8e-90
JARO001004G08 01 Mus musculus microtubule-actin crosslinking factor 1 4e-22
JARO001005B01 01 Gallus gallus alpha-actinin 5e-12
JARO001009C11 01 Daboia russellii vimentin 0.0
JARO001013C03 01 Gallus gallus myosin heavy chain 7e-05
D.A.P. Cidade et al. / Toxicon 48 (2006) 437–461450

ARTICLE IN PRESS
Table 3 (continued )
Cluster N Putative identification e-value
JARO001014B09 01 Homo sapiens capping protein (actin filament) 3e-10
JARO001033H11 01 Actin-related protein 6 3e-63
JARO001036E08 01 Spectrin alpha chain [Gallus gallus] 1e-23
JARO001042B07 01 Homo sapiens profilin 2 2e-71
JARO001047H09 01 phosphatase and actin regulator 2 [Gallus gallus] 7e-30
JARO001051B07 01 Homo sapiens spectrin SH3 3e-06
JARO001052D04 01 Myosin light chain smooth muscle isoform [Meleagris gallopavo] 2e-62
JARO001055C07 01 Keratin 18 [Homo sapiens] 5e-30
Cell regulation and other functions
JARO001010D11 07 Selenoprotein M precursor (SelM protein) 9e-31
JARO001001D11 06 ARMET protein precursor (Arginine-rich protein) 1e-32
JARO001002H09 03 Superoxide dismutase [Cu-Zn] 1e-30
JARO001023B02 03 Heat shock protein 108 [Gallus gallus] 1e-70
JARO001003B11 02 Transposase [Pleuronectes platessa] 8e-24
JARO001013B04 02 Homo sapiens calmodulin 1 (phosphorylase kinase, delta) 2e-79
JARO001013G12 02 Zinc transporter-similar to Dri 27/ZnT4 protein [Gallus gallus] 2e-14
JARO001033F01 02 Signal sequence receptor delta[Xenopus laevis] 5e-77
JARO001041F11 02 HA008F Snake Bothrops insularis retrotransposon 1e-16
JARO001002F11 01 Calcium/calmodulin-dependent protein kinase I [Pan troglodytes] 5e-73
JARO001005B02 01 Interleukin 25; lymphocyte antigen 6 complex, locus E ligand [Gallus gallus] 4e-53
JARO001005F01 01 Selenoprotein H 5e-13
JARO001006B12 01 Zinc finger protein 91 protein [Mus musculus] 5e-14
JARO001008D08 01 Selenoprotein [Xenopus laevis] 3e-07
JARO001009D08 01 Voltage-dependent anion-selective channel protein 2 (Outermitochondrial membrane protein porin) 9e-13
JARO001010F07 01 Proliferating cell nuclear antigen (PCNA) 2e-16
JARO001012D01 01 Transposase [Pleuronectes platessa] 7e-26
JARO001013B08 01 Adenine phosphoribosyl-transferase 7e-27
JARO001013B12 01 Zinc ion transporter 27/ZnT4 protein [Gallus gallus] 2e-13
JARO001014F02 01 Zinc finger protein- cellular nucleic acid binding protein 2e-29
JARO001015B12 01 Signal sequence receptor—delta subunit 2e-47
JARO001016D08 01 Translocation associated membrane protein 1 [Gallus gallus] 3e-81
JARO001023B08 01 Mus musculus RAS-related C3 botulinum substrate 1 1e-09
JARO001023E11 01 Iron storage protein H-ferritin [Trichosurus vulpecula] 2e-53
JARO001032B06 01 Caspase-7 [Gallus gallus] 3e-91
JARO001032D11 01 Heat shock 70 kDa protein 8 [Bos taurus] 3e-34
JARO001032E04 01 MAPHomo sapiens] 7e-31
JARO001032F04 01 Signal sequence receptor, beta precursor [Homo sapiens] 3e-26
JARO001034B07 01 Signal sequence receptor, beta precursor [Homo sapiens] 1e-21
JARO001034F03 01 Arginine-rich, mutated in early stage tumors [Homo sapiens] 5e-16
JARO001035D06 01 HA008F Snake Bothrops insularis retrotransposon 2e-08
JARO001035E12 01 Voltage-dependent anion channel 3 [Sus scrofa] 8e-64
JARO001040D03 01 glycolipid transfer protein—pig 1e-23
JARO001040F09 01 Translocation associated membrane protein 1 [Gallus gallus] 9e-52
JARO001041B07 01 Gallus gallus clone 9 MHC class II antigen 7e-23
JARO001041B10 01 Cell division control protein Septin 10, isoform 1 [Homo sapiens] 4e-38
JARO001042D03 01 Microvascular endothelial differentiation protein [Homo sapiens] 7e-17
JARO001043A05 01 Signal sequence receptor, delta [Mus musculus] 1e-35
JARO001043C11 01 Reverse transcriptase [schistosoma mansoni] 8e-06
JARO001043E05 01 THO complex subunit 3 (Tho3) [Gallus gallus] 1e-12
JARO001043F12 01 Endonuclease/reverse transcriptase [Mus musculus] 4e-19
JARO001044D01 01 DNA repair protein complementin G XP-A cells homolog 5e-29
JARO001045E01 01 Signal sequence receptor beta subunit [Canis familiaris] 8e-29
JARO001045E10 01 Heat shock cognate 70 protein [Spodoptera frugiperda] 8e-53
JARO001045F10 01 Pol (reverse transcriptase-RNase H-integrase) [Tricholoma matsutake] 2e-08
JARO001046G03 01 Cell division cycle 5 protein [Pan troglodytes] 7e-74
JARO001047E05 01 epithelial glycoprotein [Gallus gallus] 2e-34
JARO001048C09 01 Homo sapiens profilin 2 (PFN2) 7e-11
D.A.P. Cidade et al. / Toxicon 48 (2006) 437–461 451

ARTICLE IN PRESS
Table 3 (continued )
Cluster N Putative identification e-value
JARO001051E09 01 Nuclear protein Ytm1 [Mus musculus] 4e-15
JARO001051G05 01 Interleukin 17 receptor C isoform 1 precursor [Homo sapiens 2e-36
JARO001052H07 01 Troponin T3 [Xenopus tropicalis] 7e-59
JARO001053A02 01 Transmembrane protein precursor [Mus musculus] 5e-17
JARO001054A09 01 Ras-associated protein RAB8[Gallus gallus] 3e-08
JARO001054F04 01 Homo sapiens profilin 2 (PFN2) 3e-10
JARO001055A09 01 Thioredoxin-dependent peroxidereductase [Gallus gallus] 8e-44
JARO001055D03 01 Transmembrane 4 superfamily member 15 6e-59
Table 4
Representation of the putative toxin transcripts from B. jararaca
venom glands according to their structural types, as well as their
redundancy (clones/clusters)
Toxins Clusters (%) Clones (%) Redundancy
Metalloproteinase 172 (42.1) 681 (53.1) 4.0
Serine proteinase 141 (34.5) 366 (28.5) 2.6
C-type lectin 41 (10.0) 106 (8.3) 2.6
BPP precursor 25 (6.1) 80 (6.2) 3.2
PLA2 08 (1.9) 09 (0.7) 1.1
PLA2 inhibitor 09 (2.2) 12 (0.9) 1.3
Cysteine-rich protein 09 (2.2) 20 (1.6) 2.2
L-Amino acid oxidase 02 (0.5) 07 (0.5) 3.5
SvVEGF 02 (0.5) 02 (0.2) 1.0
BPP—bradykinin-potentiating peptide; PLA2-phospholipase A2;
svVEGF-snake venom vascular endothelium growth factor.
D.A.P. Cidade et al. / Toxicon 48 (2006) 437–461452
class of protein over the total number of toxintranscripts is shown in Table 4, as well as theirredundancy (clones/clusters).
3.3.1. Metalloproteinases/disintegrins
The most abundant toxins in our EST databaseare the metalloproteinases (53.1% of toxin tran-scripts). Most of these enzymes present hemorrhagicactivity through degradation of proteins of thebasement membrane in the blood vessel wall, andcan be divided into four classes (PI–PIV) dependingon the domain organization of their precursor thatwould be classified NI–NIV, as follows: NI com-prise precursors with only a metalloproteinasedomain, NII members have a metalloproteinasedomain followed by a disintegrin domain, NIIImembers present metalloproteinase, disintegrin-likeand cysteine-rich domains and the hypothetical NIVprecursor would contain an additional lectin-likedomain that remains linked by disulfide bonds tothe metalloproteinase domain on the venom(Takeya et al., 1992; Hite et al, 1994; Jia et al.,
1996; Markland, 1998). Concerning our database,198 clones (29.1%) could be characterized as NIImetalloproteinases. This class of metalloproteinasespresented the highest redundancy of all transcripts(14.1), including the most representative transcriptin the whole library (137 clones), identified as a B.
jararaca metalloproteinase bothrostatin homolog(Fernandez et al., 2005). In addition, 318 (46.7%)out of 681 metalloproteinase clones could beassigned as NIII precursors. These clones weregrouped in 49 clusters, resulting in a redundancy of6.5, which is lower than that observed for the NIIclass. The most abundant NIII clusters showedsimilarity with bothropasin (Mandelbaum et al.,1982) and/or jararhagin (Paine et al., 1992), whichare PIII metalloproteinases isolated from B. jarar-
aca venom presenting high-sequence homology.They show some differences in the metalloprotei-nase domain but are identical in their disintegrin-like and cysteine-rich domains. Thus, in some caseswhere the homology occurred at these domains, itwas not possible to determine to which of them theB. jararaca cluster is more similar, being thenclassified as a bothropasin/jararhagin homolog.These proteins act by cleaving fibrinogen whichleads to the enhancement of plasma fibrinolysis,resulting on local and systemic hemorrhage (Paineet al., 1992). The high expression of toxins similar tobothropasin/jararhagin observed in B. jararaca
venom gland is compatible with the abundantbleeding observed in patients bitten by this snake.In addition, 2 clusters (JARO001012A02 [5 reads]and JARO001014A02 [1 read]) presented homologywith the vascular apoptosis-inducing protein(VAIP), which is a PIII metalloproteinase present-ing the metalloproteinase, disintegrin-like and cy-steine-rich domains characteristic of this Zn2+-dependent enzyme. VAIP is able to degradefibrinogen and induce apoptosis in vascular en-dothelial cells (Masuda et al., 2000).

ARTICLE IN PRESSD.A.P. Cidade et al. / Toxicon 48 (2006) 437–461 453
Among the metalloproteinases, 24.2% of theclones could not be precisely identified throughtheir domain composition because the clusterscomprise only part of the metalloproteinase domainor because the homology occurred only at the30UTR. Concerning the metalloproteinases whichcould have their classes determined, in our study wefound molecules that belong to NII and NIII classesof precursors, which is similar to what was foundfor Bitis gabonica venom gland (Franscischetti et al.,2004). In B. jararaca (Kashima et al., 2004) andB. insularis (Junqueira-de-Azevedo and Ho, 2002),the NI class was also identified while the hypothe-tical NIV metalloproteinase precursor was neveridentified.
Looking at the clusters containing the disintegrinregion, we could find 34 clusters (348 reads) codingfor disintegrin (RGD-containing) or disintegrin-like(XCD-containing) peptides. Among the 348 reads,178 (51.1%) code for RGD disintegrins contained inNII metalloproteinase precursors and 165 (47.4%)code for ECD disintegrin-like peptides contained inNIII metalloproteinase precursors. The most repre-sentative disintegrin domain-containing cluster(JARO001001E01–137 reads) presents 100% iden-tity with the B. jararaca disintegrin jararacin(Scarborough et al., 1993), and the second one(JARO001002D11–103 reads) presents 100% iden-tity with the B. jararaca disintegrin-like domain ofthe bothropasin precursor (Mandelbaum et al.,1982).
Considering that the venom composition may belinked to adaptation to the environment (Okudaet al., 2001; Daltry et al., 1996), it is interesting tonote that B. jararaca transcriptome contains,proportionally, much more ‘‘true’’ (RGD-contain-ing) disintegrins, which are released from PIImetalloproteinases (Kini and Evans, 1992), thanthe transcriptome of its close sister, B. insularis, inwhich just one cluster (02 reads) presented a truedisintegrin motif, over a total of 33 metalloprotei-nases clusters (144 reads) (Junqueira-de-Azevedoand Ho, 2002). A dominance of NIII metallopro-teinases in B. insularis, in contrast to the similarproportions of NII and NIII precursors observed inB. jararaca, is consistent with the activity of theirvenoms. Hemorragins of the PIII class present themost potent activity (Gutierrez et al., 2005; Mark-land, 1998), which may be due to the presence of thecysteine-rich domain, known to enhance the inhibi-tion of collagen-induced platelet aggregation causedby PIII metalloproteinases through interaction with
the platelet integrin a2b1 (Kamiguti et al., 2003; Jiaet al., 2000). Thus, the presence of the cysteine-richdomain may function to synergistically increase thehemorragic effect of these toxins. The predominantexpression of NIII metalloproteinases in B. insularis
venom gland over other classes of metalloprotei-nases may contribute to its highly toxic effect,known as being much more potent than the venomof B. jararaca, with a faster lethal effect (Mosmann,2001; Melgarejo, 2003). Therefore, we can suggestthat the extremely competitive habitat to whichB. insularis has been submitted for thousands ofyears at Queimada Grande island, including highsnake density and little food availability, leads tothe selection of lineages expressing preferentiallymore potent toxins as PIII metalloproteinases.
More recently, a new gene structure of thedisintegrin family was identified in Agkistrodon
contortrix contortrix (acostatin) and Agkistrodon
piscivourus piscivourus (piscivostatin) venoms. Itconsists of a signal peptide, a pro-domain and adisintegrin domain, lacking the metalloproteinasedomain (Okuda et al., 2002). In Bitis gabonica ESTlibrary, two clusters presented homology with thiskind of structure, named gabonin 1 and 2, describedas new members of the short coding region family ofdisintegrins (Franscischetti et al., 2004). On theother hand, in B. jararaca transcriptome, we did notfind any disintegrin belonging to this group, asobserved for B. insularis and B. jararaca transcrip-tomes. Therefore, we may suggest that the absenceof such molecules is a shared character amongmembers of Bothrops genus.
3.3.2. Serine-proteinases
Snake venom serine-proteinases are enzymes thataffect hemostasis and thrombosis upon envenoma-tion, acting on a variety of components of thecoagulation cascade in order to cause an imbalanceof the haemostatic system of the prey (Matsui et al.,2000). They can act as procoagulants causing invivo activation of the coagulation system, butbecause of the excessive consumption of coagula-tion factors they frequently lead to clinical antic-oagulation (Markland, 1998). These enzymes aredefined by a common catalytic mechanism thatincludes a highly reactive serine residue that formsan active site within a histidine and aspartic acidresidues. A high number of isoforms have beendescribed for venom serine-proteinases, in whichsmall changes in sequence or structure have a greatimpact on activity (Saguchi et al., 2005; Serrano and

ARTICLE IN PRESSD.A.P. Cidade et al. / Toxicon 48 (2006) 437–461454
Maroun, 2005). Thus, venom gland serine-protei-nases are known to form a multigene family(Deshimaru et al., 1996).
The identification of most of the serine-proteinaseclusters was given by the 30UTR region, which isparticularly long, as noted in the B. insularis
transcriptome (Junqueira-de-Azevedo and Ho,2002). This class of toxin follows the metalloprotei-nases in abundancy of toxins present in B. jararaca
transcriptome (Fig. 1). Among them, the mostrepresentative molecules (62 clusters/173 reads) arehomologs of the serine a-fibrinogenase isolatedfrom Macrovipera lebetina, known to present antic-oagulant activity (NCBI accession number (acc)AF528193) (Table 2). The second most abundantgroup (38 clusters/107 reads) of serine-proteinases
Fig. 1. Graphics showing the representation of clones from each
toxin category over the total of toxin transcripts from B. jararaca
(A) and B. insularis (B) venom glands transcriptome (B was
reproduced with authorization from Junqueira-de-Azevedo and
Ho, 2002). BPP—bradykinin-potentiating peptide; LAO—
L-amino acid oxidase; svVEGF—snake venom vascular endothe-
lium growth factor; PLA2—phospholipase A2; NGF—nerve
growth factor.
presents homology to KN-BJ2 (acc AB004067),which is a serine-proteinase isolated from B.
jararaca venom that displays kinin-releasing andfibrinogen-clotting activity, being fully or largelyresponsible for bradykinin release (Serrano et al.,1998). The high amount of cDNAs coding forserine-proteinases found in B. jararaca venom glandis in accordance with the clinical effects observedupon accidents caused by this species, as the majoreffect of these toxins in the envenomation is theexcessive consumption of coagulation factors andformation of abnormal fibrin clots resulting in theabsence of blood coagulation (Matsui et al., 2000).
3.3.3. C-type lectins
There are two major C-type lectin classes ofproteins in snake venoms which belong to theC-type lectin superfamily: the ‘‘true’’ lectins, gen-erally homodimeric and presenting saccharide-bind-ing activity in a calcium-dependent manner (Xu etal., 1999) and the C-type lectin-like proteins,generally heterodimeric and that bind to proteinligands rather than saccharides (Wei et al., 2002).The main reported activity of the first class ishemagglutination (Ozeki et al., 1994; Xu et al.,1999; de Carvalho et al., 2002) while members of thesecond class interfere in the coagulation pathways invarious ways and exhibit anticoagulant or proag-gregating effects, which makes them promisingcandidates for the development of new drugs(Andrews and Berndt, 2000; Andrews et al., 2001).The C-type lectin-like proteins can be divided intofour main groups, according to their bindingcharacteristics: (1) inhibition of conversion ofprothrombin into thrombin through binding tocoagulation factors IX and X (Matsuzaki et al.,1996; Lee et al., 2003); (2) direct inhibition ofthrombin action, such as caused by bothrojaracin(Arocas et al., 1997); (3) inhibition of plateletaggregation through binding to GPIb present onplatelet surfaces (Shin et al., 2000) and (4) stimula-tion of platelet aggregation: as an example, the two-chain bothrocetin binds to von Willebrand factor,changing its conformation and activating its bindingto platelet glicoprotein (GP) Ib (Sen et al., 2001).
In our transcriptome analysis, we could identifyclusters presenting homology with all C-typelectin-like protein classes described before(Table 2). The most abundant C-type lectin-likeproteins are homologues of the glycoprotein Ib-binding protein, previously isolated from B. jarar-
aca venom (Kawasaki et al., 1996), followed by

ARTICLE IN PRESSD.A.P. Cidade et al. / Toxicon 48 (2006) 437–461 455
bothrojaracin homologues (Arocas et al., 1997),factor IX/X binding proteins (Matsuzaki et al.,1996) and botrocetin homologues (Sen et al., 2001).
In the B. jararaca venom gland transcriptome,there was only one C-type lectin cluster, whichcodes for a putative homodimeric C-type ‘‘true’’galactose-binding lectin. It was composed of 25reads, and corresponded to 7.4% of toxin-relatedsequences (Kashima et al., 2004). The presence oftrue lectins in B. jararaca venom has already beendescribed, and associated with strong hemaggluti-nation (de Carvalho et al., 2002). In fact, gumbleeding has been observed in only 6.9% ofBrazilian B. jararaca-caused accidents, as opposedto 24% of B. jararaca-caused accidents (Franc-a andMalaque, 2003). This feature may be, in part, due tothe predominance of hemagglutinating true-lectinsin B. jararaca venom, which are less abundant inB. jararaca. In the Bitis gabonica venom glandtranscriptome, 4 clusters of C-type lectins werefound, one of them being similar to galactose-binding lectin and the others similar to C-typelectin-like proteins (Franscischetti et al., 2004).
The cloning, characterization and structuralanalysis of a true-lectin (BiL) from B. insularis wasrecently reported (Guimaraes-Gomes et al., 2004).BiL displayed hemagglutinating activity that wasinhibited by galactose, lactose and EDTA, eviden-cing that it is a typical calcium-dependent galactose-binding lectin. In B. insularis venom gland tran-scriptome, among the 16 C-type lectin clusters, 2showed best similarities with galactose-binding true-lectins (Junqueira-de-Azevedo and Ho, 2002). Itseems that true lectins have a minor participation inthe B. insularis envenomation pathology, since theycorresponded to only 14% of the C-type lectin-related transcripts. No true lectin was found in ourtranscriptome analysis, even when the library wasscreened by PCR using specific primers, in spite of ithaving been previously described in B. jararaca
venom (Ozeki et al., 1994). This is probably due toindividual variation in venom composition.
3.3.4. Bradykinin-potentiating peptides (BPPs)
The BPPs are proline-rich oligopeptides of 5-14amino acid residues. They usually present apyroglutamyl residue at the N-terminus and anIle-Pro-Pro (IPP) sequence at the C-terminus. Theyare known to inhibit the endothelial metalloprotei-nase angiotensin-converting enzyme (ACE), whichproduces angiotensin II, a potent vasoconstrictor,through hydrolysis of angiotensin I (Hayashi and
Camargo, 2005). ACE is responsible for themaintenance of normal blood pressure levels, andis also able to inactivate the hypotensive peptidebradykinin, which is released in the blood ofenvenomed victims through the action of venomserine-proteinases. Therefore, BPPs are involvedwith the hypotension observed after envenomationby Bothrops species, and have been used as modelsfor the development of new antihypertensive drugs(Fernandez et al., 2004).
In our analysis, we found 25 clusters (80 reads)corresponding to BPP precursor transcripts, ac-counting for 6.2% of the toxin transcripts (Tables 2and 4). Most of them present only the 30UTR,known to be particularly long in BPP precursors(Murayama et al., 1997; Higuchi et al., 1999; Soareset al., 2005). These transcripts are known to containseveral BPP-coding regions tandemly arranged atthe N-region and a C-type natriuretic peptide at theC-region of the precursor (Hayashi and Camargo,2005). Cluster JARO001047D04 consists of apartial sequence that presented 100% identity withthe 7 BPPs located at the N-terminal region of aBPP precursor previously described in B. jararaca
(Murayama et al., 1997). In B. insularis transcrip-tome, BPP-related transcripts accounted for 19.7%of toxin transcripts. In addition, a higher redun-dancy of BPPs was observed, since 61 reads couldbe grouped in a same cluster (Junqueira-de-Azevedoand Ho, 2002). When we compare these two pitvipers, it seems that B. insularis became specializedin the production of high amounts of extremelyconserved BPPs while B. jararaca presents morevariability among BPP precursors. This high fre-quency of BPPs in B. insularis transcriptome isin accordance with the activity of its venom.Therefore, we could speculate that the action ofB. insularis venom, which is able to kill the preyimmediately, is fully or largely related to a BPP-caused hypotensive shock.
3.3.5. Phospholipases A2 and their inhibitors
Bothrops venoms contain numerous componentsthat could potentially affect neuromuscular re-sponses, including the phospholipases A2 (PLA2)myotoxins. They are the major components respon-sible for the intense local damage observed aftersome Bothropic accidents, being capable of inducinginflammatory events through the enhancement ofvascular permeability and the recruitment of neu-trophils (Zuliani et al., 2005). Some Bothrops
venoms (B. jararaca, B. moojeni and B. neuwiedi)

ARTICLE IN PRESSD.A.P. Cidade et al. / Toxicon 48 (2006) 437–461456
are known to cause high levels of muscle damagewhen compared to B. jararaca and B. erythromelas
venoms (Zamuner et al., 2004). These findings are inaccordance with our results, in which PLA2 seemsto have a minor participation in the envenomationprocess since it accounts for only 0.7% of the toxintranscripts, indicating that the local damage isless representative than the vascular disordersinduced by metalloproteinases, serine-proteinasesand C-type lectins (Fig. 1A). The same pattern wasobserved in the analysis of B. insularis transcrip-tome, in which the PLA2 accounted for 6.7% of thetoxin transcripts (Junqueira-de-Azevedo and Ho,2002). On the other hand, the high frequency ofPLA2 transcripts in B. jararacussu transcriptome(58% of the toxin transcripts) evidences the clearpredominance of the local action of this venom(Kashima et al., 2004). In Bitis gabonica transcrip-tome, a high number of transcripts coding for PLA2
was also found (32% of the toxin transcripts), PLA2
being the most expressed toxin (Franscischetti et al.,2004). This result indicates a predominance of localaction over systemic injury in both B. jararacussu
and Bitis gabonica envenoming accidents.Interestingly, we found some clusters coding for
PLA2 inhibitors, which are mostly present in snakesera. The most frequent ones, comprising only ofthe 30UTR, are homologous to a Trimeresurus
flavoviridis phospholipase A2 class A inhibitor(Nobuhisa et al., 1997) (Table 2). PLA2 inhibitorshave been purified from the sera of many venomoussnakes such as Notechis ater (Hains et al., 2000),Crotalus durissus terrificus (Faure et al., 2000) andTrimeresurus flavoviridis (Nobuhisa et al., 1997) andalso from nonvenomous snakes such as Elaphe
quadrivirgata (Okumura et al., 2002) and Pyton
reticulatus (Thwin et al., 2002). Several studiesshowed that the resistance of some animals tosnake venoms can be explained by the presence ofprotein factors in their blood which inhibit theactivity of important toxic compounds. Theseproteins are either metalloproteinase inhibitors(Perales et al., 2005) or PLA2 inhibitors (Lizanoet al., 2003). To our knowledge, PLA2 inhibitorshave never been described as venom components,although it is well known that some venoms containcompetitive components. B. jararaca venom, forinstance, contains not only botrocetin but alsoGPIb-BP which inhibits the botrocetin-inducedplatelet agglutination (Kawasaki et al., 1996). Ifthese kinds of PLA2 inhibitors are present in thevenom and are playing some specific role in the
envenomation caused by B. jararaca is a question tobe further elucidated.
Noteworthy, in our laboratory we have pre-viously detected the activity of PLA2 inhibitors inBothrops jararaca venom, although a molecularcharacterization of its structure was not performed(Sousa et al., 2001). There are very few reportsdescribing the presence of low molecular weightPLA2 inhibitors in the venom, in particular fromBothrops snakes (Vidal and Stoppani, 1971 and1980). These molecules are much smaller than thePLA2 inhibitors isolated from sera. The absence of acomplete characterization of these peptides, espe-cially regarding its amino acid sequence, makes itdifficult to fully appreciate the significance of thesemolecules.
3.3.6. Other toxins present in B. jararaca venom
gland transcriptome
In the EST database we found some venomcysteine-rich secretory proteins (CRISPs) (Table 2).These proteins are widely distributed in snakevenoms, but little information is available abouttheir action in the envenomation. Until now, twotypes of activities were found: the inhibition ofsmooth muscle contraction and the blocking ofcyclic-nucleotide-gated ion channels (Yamazaki andMorita, 2004; Oispov et al., 2005). We also foundsome toxin classes that seem to have minorparticipation in B. jararaca envenomation process,since they are present in small amounts in thedatabase. The L-amino acid oxidases, which are ableto induce apoptosis, affect platelets and havehemorragic effects (Aird, 2002), were found at afrequency of 0.5% of toxin-related reads (Table 4).In addition, the snake venom vascular endotheliumgrowth factor (svVEGF) accounts for 0.2% of thetoxin-related reads in B. jararaca transcriptome,which is less represented than in B. insularis
transcriptome (4.3%). B. insularis svVEGF wascloned, expressed and shown to be capable ofincreasing vascular permeability thus acting as atoxin dispersion agent and probably contributing tothe hypotensive action observed upon envenoma-tion (Junqueira-de-Azevedo et al., 2001).
3.4. Analysis of transcripts related to cellular
functions
Proteins related to cellular functions, includinghousekeeping proteins, comprise 22.4% of thematching transcripts. These transcripts were divided
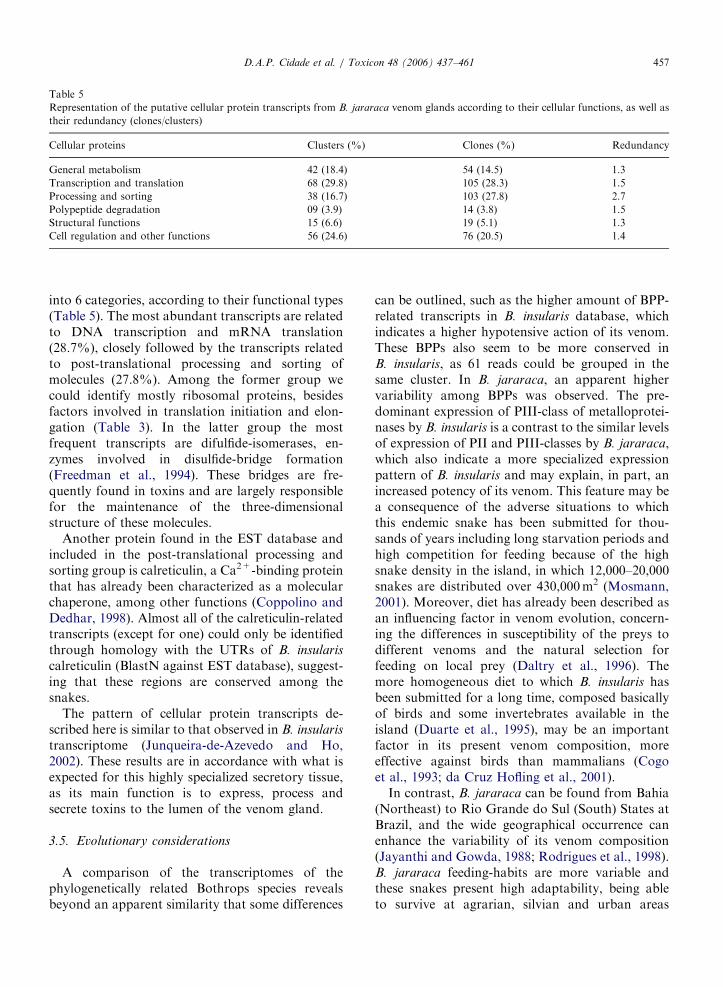
ARTICLE IN PRESS
Table 5
Representation of the putative cellular protein transcripts from B. jararaca venom glands according to their cellular functions, as well as
their redundancy (clones/clusters)
Cellular proteins Clusters (%) Clones (%) Redundancy
General metabolism 42 (18.4) 54 (14.5) 1.3
Transcription and translation 68 (29.8) 105 (28.3) 1.5
Processing and sorting 38 (16.7) 103 (27.8) 2.7
Polypeptide degradation 09 (3.9) 14 (3.8) 1.5
Structural functions 15 (6.6) 19 (5.1) 1.3
Cell regulation and other functions 56 (24.6) 76 (20.5) 1.4
D.A.P. Cidade et al. / Toxicon 48 (2006) 437–461 457
into 6 categories, according to their functional types(Table 5). The most abundant transcripts are relatedto DNA transcription and mRNA translation(28.7%), closely followed by the transcripts relatedto post-translational processing and sorting ofmolecules (27.8%). Among the former group wecould identify mostly ribosomal proteins, besidesfactors involved in translation initiation and elon-gation (Table 3). In the latter group the mostfrequent transcripts are difulfide-isomerases, en-zymes involved in disulfide-bridge formation(Freedman et al., 1994). These bridges are fre-quently found in toxins and are largely responsiblefor the maintenance of the three-dimensionalstructure of these molecules.
Another protein found in the EST database andincluded in the post-translational processing andsorting group is calreticulin, a Ca2+-binding proteinthat has already been characterized as a molecularchaperone, among other functions (Coppolino andDedhar, 1998). Almost all of the calreticulin-relatedtranscripts (except for one) could only be identifiedthrough homology with the UTRs of B. insularis
calreticulin (BlastN against EST database), suggest-ing that these regions are conserved among thesnakes.
The pattern of cellular protein transcripts de-scribed here is similar to that observed in B. insularis
transcriptome (Junqueira-de-Azevedo and Ho,2002). These results are in accordance with what isexpected for this highly specialized secretory tissue,as its main function is to express, process andsecrete toxins to the lumen of the venom gland.
3.5. Evolutionary considerations
A comparison of the transcriptomes of thephylogenetically related Bothrops species revealsbeyond an apparent similarity that some differences
can be outlined, such as the higher amount of BPP-related transcripts in B. insularis database, whichindicates a higher hypotensive action of its venom.These BPPs also seem to be more conserved inB. insularis, as 61 reads could be grouped in thesame cluster. In B. jararaca, an apparent highervariability among BPPs was observed. The pre-dominant expression of PIII-class of metalloprotei-nases by B. insularis is a contrast to the similar levelsof expression of PII and PIII-classes by B. jararaca,which also indicate a more specialized expressionpattern of B. insularis and may explain, in part, anincreased potency of its venom. This feature may bea consequence of the adverse situations to whichthis endemic snake has been submitted for thou-sands of years including long starvation periods andhigh competition for feeding because of the highsnake density in the island, in which 12,000–20,000snakes are distributed over 430,000m2 (Mosmann,2001). Moreover, diet has already been described asan influencing factor in venom evolution, concern-ing the differences in susceptibility of the preys todifferent venoms and the natural selection forfeeding on local prey (Daltry et al., 1996). Themore homogeneous diet to which B. insularis hasbeen submitted for a long time, composed basicallyof birds and some invertebrates available in theisland (Duarte et al., 1995), may be an importantfactor in its present venom composition, moreeffective against birds than mammalians (Cogoet al., 1993; da Cruz Hofling et al., 2001).
In contrast, B. jararaca can be found from Bahia(Northeast) to Rio Grande do Sul (South) States atBrazil, and the wide geographical occurrence canenhance the variability of its venom composition(Jayanthi and Gowda, 1988; Rodrigues et al., 1998).B. jararaca feeding-habits are more variable andthese snakes present high adaptability, being ableto survive at agrarian, silvian and urban areas

ARTICLE IN PRESSD.A.P. Cidade et al. / Toxicon 48 (2006) 437–461458
(Mosmann, 2001; Melgarejo, 2003). In spite of theapparent differences, we have to consider theseconclusions carefully, since the number of generatedESTs for both transcriptomes is different (610 forB. insularis and 2318 for B. jararaca) and this canlead to some bias in the analysis. Moreover,individual variation may have contributed for someof the observed differences.
3.6. Perspectives
Snake venom is a natural library for screeningvaluable bioactive substances for hemostasis andthrombosis, among others (Marsh and Williams,2005). This work provided a catalog of thetranscripts contained in B. jararaca venom gland,which has been used for supporting proteomicstudies of this species (unpublished). Moreover,the sequences can be used to directly probe thegenetic material from other snake species or toinvestigate differences in gene expression pattern inresponse to factors such as diet, aging andgeographic localization. Attempts for the cloningand expression of toxins from the database are incourse, aiming to elucidate some structure-activityrelationships.
Acknowledgments
This work was supported by grants from theBrazilian agencies Fundac- ao de Amparo a Pesquisado Estado do Rio de Janeiro (FAPERJ), ConselhoNacional de Desenvolvimento Cientıfico e Tecno-logico (CNPq), Rede Proteomica do Rio de Janeiroand Riogene.
References
Aird, S.D., 2002. Ophidian envenomation strategies and the role
of purines. Toxicon 40, 335–393.
Andrews, R.K., Berndt, M.C., 2000. Snake venom modulators of
platelet adhesion receptors and their ligands. Toxicon 38,
775–791.
Andrews, R.K., Kamiguti, A.S., Berlanga, O., Leduc, M.,
Theakston, R.D.G., Watson, S.P., 2001. The use of snake
venom toxins as tools to study platelet receptors for collagen
and von Willebrand factor. Haemostasis 31, 155–172.
Arocas, V., Castro, H.C., Zingali, R.B., Guillin, M.C., Jandrot-
Perrus, M., Bon, C., Wisner, A., 1997. Molecular cloning and
expression of bothrojaracin, a potent thrombin inhibitor from
snake venom. Eur. J. Biochem. 248, 550–557.
Cogo, J.C., Prado-Franceschi, J., Cruz-Hofling, M.A., Corrado,
A.P., Rodrigues-Simioni, L., 1993. Effect of Bothrops insularis
venom on the mouse and chick nerve-muscle preparation.
Toxicon 31, 1237–1247.
Coppolino, M.G., Dedhar, S., 1998. Calreticulin. Int. J. Biochem.
Cell Biol. 30, 553–558.
da Cruz Hofling, M.A., Paronetto, C.C., Cogo, J.C., Rodrigues-
Simioni, L., D‘Abreu, A.C., 2001. Histopathological changes
in avian kidney caused by Bothrops insularis (jararaca ilhoa)
venom and a phospholipase A2-containing fraction. Histol.
Histopathol. 16, 185–195.
Daltry, J.C., Wuster, W., Thorpe, R.S., 1996. Diet and snake
venom evolution. Lett. Nat. 379, 537–540.
Davey, G.C., Caplice, N.C., Martin, S.A., Powell, R., 2001. A
survey of genes in the Atlantic salmon (Salmo salar) as
identified by expressed sequence tags. Gene 263, 121–130.
Davila, A.M.R., Lorenzini, D.M., Mendes, P.N., Satake, T.S.,
Sousa, G.R., Campos, L.M., Mazzoni, C.J., Wagner, G.,
Pires, P.F., Grisard, E.C., Cavalcanti, M.C.R., Campos,
M.L.M., 2005. GARSA: Genomic Analysis Resources for
Sequence Annotation. Bioinformatics, October 6.
de Carvalho, D.D., Marangoni, S., Novello, J.C., 2002. Primary
structure characterization of Bothrops jararacussu snake
venom lectin. J. Prot. Chem. 21, 43–50.
Deshimaru, M., Ogawa, T., Nakashima, K.I., Nobuhisa, I.,
Chijiwa, T., Shimohigashi, Y., Fukumaki, Y., Niwa, M.,
Yamashina, I., Hattori, S., Ohno, M., 1996. Accelerated
evolution of crotalinae snake venom gland serine proteases.
FEBS Lett. 397, 83–88.
Duarte, M.R., Puorto, G., Franco, F.L., 1995. A biological
survey of the pit viper Bothrops insularis Amaral (Serpentes,
Viperidae): an endemic and threatened offshore island snake
of southeastern Brazil. Stud. Neotr. Fauna Environ. 30, 1–13.
Faure, G., Villela, C., Perales, J., Bon, C., 2000. Interaction of the
neurotoxic and nontoxic secretory phospholipases A2 with the
crotoxin inhibitor from Crotalus serum. Eur. J. Biochem. 267,
4799–4808.
Fernandez, J.H., Neshich, G., Camargo, A.C.M., 2004. Using
bradykinin-potentiating peptide structures to develop new
antihypertensive drugs. Genet. Mol. Res. 3, 554–563.
Fernandez, J.H., Silva, C.A., Assakura, M.T., Camargo, A.C.M.,
Serrano, S.M.T., 2005. Molecular cloning, functional expres-
sion, and molecular modeling of bothrostatin, a new highly
active disintegrin from Bothrops jararaca venom. Biochem.
Biophys. Res. Commun. 329, 457–464.
Franc-a, F.O.S., Malaque, C.M.S., 2003. Acidente botropico. In:
Cardoso, J.L.C., Franc-a, F.O.S., Wen, F.H., Malaque,
C.M.S., Haddad Jr. V. (Eds), Animais pec-onhentos no Brasil:
biologia, clınica e terapeutica dos acidentes, Sarvier, Sao
Paulo, pp. 72–86.
Francischetti, I.B.M., Valenzuela, J.G., Pham, V.M., Garfield,
M.K., Ribeiro, M.C., 2002. Toward a catalog for the
transcripts and proteins (sialome) from the salivary gland of
the malaria vector Anopheles gambiae. J. Exp. Biol. 205,
2429–2451.
Franscischetti, I.M.B., Pham, V.M., Harrison, J., Garfield, M.K.,
Ribeiro, J.M.C., 2004. Bitis gabonica (Gaboon viper) snake
venom gland: toward a catalog for the full-length transcripts
(cDNA) and proteins. Gene 337, 55–69.
Freedman, R.B., Hirst, T.R., Tuite, M.F., 1994. Protein
disulphide isomerase: building bridges in protein folding.
Trends Biochem. Sci. 19, 331–336.
Guimaraes-Gomes, V., Oliveira-Carvalho, A.L., Junqueira-de-
Azevedo, I.L.M., Dutra, D.L.S., Pujol-Luz, M., Castro, H.C.,

ARTICLE IN PRESSD.A.P. Cidade et al. / Toxicon 48 (2006) 437–461 459
Ho, P.L., Zingali, R.B., 2004. Cloning, characterization and
structural analysis of a C-type lectin from Bothrops insularis
(BiL) venom. Arch. Biochem. Biophys. 432, 1–11.
Gutierrez, J.M., Rucavado, A., Escalante, T., Dıaz, C., 2005.
Hemorrage induced by snake venom metalloproteinases:
biochemical and biophysical mechanisms involved in micro-
vessel damage. Toxicon 45, 997–1011.
Hains, P.G., Sung, K.L., Tseng, A., Broady, K.W., 2000.
Functional characteristics of a phospholipase A2 inhibitor
from Notechis ater serum. J. Biol. Chem. 275, 983–991.
Hayashi, M.A.F., Camargo, A.C.M., 2005. The bradykinin-
potentiating peptides from venom gland and brain of
Bothrops jararaca contain highly site specific inhibitors of
the somatic angiotensin-converting enzyme. Toxicon 45,
1163–1170.
Higuchi, S., Murayama, N., Saguchi, K., Ohi, H., Fujita, Y.,
Camargo, A.M.C., Ogawa, T., Deshimaru, M., Ohno, M.,
1999. Bradykinin-potentiating peptides and C-type natriuretic
peptides from snake venom. Immunopharmacology 44,
129–135.
Hite, L.A., Jia, L.G., Bjarnason, J.B., Fox, J.W., 1994. cDNA
sequences for four snake venom metalloproteinases: structure,
classification and their relationship to mammalian reproduc-
tive proteins. Arch. Biochem. Biophys. 308, 182–191.
Huang, X., Madan, A., 1999. CAP3: a DNA sequence assembly
program. Genome Res. 9, 868–877.
Jayanthi, G.P., Gowda, T.V., 1988. Geographical variation in
India in the composition and lethal potency of Russel’s viper
(Vipera russelli) venom. Toxicon 26, 257–264.
Jia, L.G., Shimokawa, K.I., Bjarnason, J.B., Fox, J.W., 1996.
Snake venom metalloproteinases: structure, function and
relationship to the ADAMs family of proteins. Toxicon 34,
1269–1276.
Jia, L.G., Wang, X.M., Shannon, J.D., Bjarnason, J.B., Fox,
J.W., 2000. Inhibition of platelet aggregation by the
recombinant cysteine-rich domain of the hemorragic snake
venom metalloproteinase, atrolysin A. Arch. Biochem.
Biophys. 373, 281–286.
Junqueira-de-Azevedo, I., Ho, P.L., 2002. A survey of gene
expression and diversity in the venom glands of the pit viper
snake Bothrops insularis through the generation of expressed
sequence tags (ESTs). Gene 299, 279–291.
Junqueira-de-Azevedo, I.L.M., Farsky, S.H.P., Oliveira, M.L.S.,
Ho, P.L., 2001. Molecular cloning and expression of a
functional snake venom vascular endothelium growth factor
(VEGF) from the Bothrops insularis, pit viper. J. Biol. Chem.
276, 39836–39842.
Kamiguti, A.S., Gallagher, P., Marcinkiewicz, C., Theakston,
R.D.G., Zuzel, M., Fox, J.W., 2003. Identification of sites in
the cysteine-rich domain of the class PIII snake venom
metallproteinases responsible for inhibition of platelet func-
tion. FEBS Lett. 549, 129–134.
Kashima, S., Roberto, P.G., Soares, A.M., Astolfi-Filho, S.,
Pereira, J.O., Giuliati, S., Faria Jr, M., Xavier, M.A.S.,
Fontes, M.R.M., Giglio, J.R., Franc-a, S.C., 2004. Analysis of
Bothrops jararacussu venomous gland transcriptome focusing
on structural and functional aspects: I-gene expression profile
of highly expressed phospholipases A2. Biochimie 86,
211–219.
Kawasaki, T., Fujimura, Y., Usami, Y., Suzuki, M., Miura, S.,
Sakurai, Y., Makita, K., Taniuchi, Y., Hirano, K., Titani, K.,
1996. Complete amino acid sequence and identification of the
platelet glycoprotein Ib-binding site of jararaca GPIb-BP, a
snake venom protein isolated from Bothrops jararaca. J. Biol.
Chem. 271, 10635–10639.
Kini, R.M., Evans, H.J., 1992. Structural domains in venom
proteins: evidence that metalloproteinases and nonenzymatic
platelet aggregation inhibitors (disintegrins) from snake
venoms are derived by proteolysis from a common precursor.
Toxicon 30, 265–293.
Lee, W.H., Zhuang, Q.Y., Zhang, Y., 2003. Cloning and
characterization of a blood coagulation factor IX-binding
protein from the venom of Trimeresurus stejnegeri. Toxicon
41, 765–772.
Lizano, S., Domont, G., Perales, J., 2003. Natural phospholipase
A2 myotoxin inhibitor proteins from snakes, mammals and
plants. Toxicon 42, 963–977.
Mandelbaum, F.R., Reichel, A.P., Assakura, M.T., 1982.
Isolation and characterization of a proteolytic enzyme from
the venom of the snake Bothrops jararaca (Jararaca). Toxicon
20, 955–972.
Markland, F.S., 1998. Snake venoms and the hemostatic system.
Toxicon 36, 1749–1800.
Marsh, N., Williams, V., 2005. Practical applications of snake
venom toxins in haemostasis. Toxicon 45, 1171–1181.
Masuda, S., Ohta, T., Kaji, K., Fox, J.W., Hayashi, H., Araki, S.,
2000. cDNA cloning and characterization of vascular
apoptosis-inducing protein. Biochem. Biophys. Res. Com-
mun. 278, 197–204.
Mathesius, U., Keijzers, G., Natera, S.H.A., Weinman, J.J.,
Djordjevic, M.A., Rolfe, B.G., 2001. Establishment of a root
proteome reference map for the model legume Medicago
truncatula using the expressed sequence tag database for
peptide mass fingerprinting. Proteomics 1, 1424–1440.
Matsui, T., Fujimura, Y., Titani, K., 2000. Snake venom
proteases affecting haemostasis and thrombosis. Biochim.
Biophys. Acta 1477, 146–156.
Matsuzaki, R., Yoshiara, E., Yamada, M., Shima, K., Atoda, H.,
Morita, T., 1996. cDNA cloning of IX/X-BP, a heterogeneous
two-chain anticoagulant protein from snake venom. Biochem.
Biophys. Res. Commun. 220, 382–387.
Melgarejo, A.R., 2003. Serpentes pec-onhentas do Brasil. In:
Cardoso, J.L.C., Franc-a, F.O.S., Wen, F.H., Malaque,
C.M.S., Haddad Jr, V. (Eds), Animais pec-onhentos no Brasil:
biologia, clınica e terapeutica dos acidentes, Sarvier, Sao
Paulo, pp. 72–86.
Mosmann, M.N., 2001. Guia das principais serpentes do mundo
(vol. 2), Ulbra, Canoas, Rio Grande do Sul, pp. 91–94.
Murayama, N., Hayashi, M.A.F., Ohi, H., Ferreira, L.A.F.,
Hermann, V.V., Saito, H., Fujita, Y., Higuchi, S., Fernandes,
B.L., Yamane, T., Camargo, A.C.M., 1997. Cloning and
sequence analysis of a Bothrops jararaca cDNA encoding a
precursor of seven bradykinin-potentiating peptides and a C-
type natriuretic peptide. Proc. Natl. Acad. Sci. USA 94,
1189–1193.
Nobuhisa, I., Inamasu, S., Nakai, M., Tatsui, A., Mimori, T.,
Ogawa, T., Shimohigashi, Y., Fukumaki, Y., Hattori, S.,
Kihara, H., Ohno, M., 1997. Characterization and evolution
of the gene encoding a Trimeresurus flavoviridis serum protein
that inhibits basic phospholipases A2 isozymes in the snake’s
venom. Eur. J. Biochem. 249, 838–845.
Oispov, A.V., Levashov, M.Y., Tsetlin, V.I., Utkin, Y.N., 2005.
Cobra venom contains a pool of cysteine-rich secretory
proteins. Biochem. Biophys. Res. Commun. 328, 177–182.

ARTICLE IN PRESSD.A.P. Cidade et al. / Toxicon 48 (2006) 437–461460
Okuda, D., Koike, H., Morita, T., 2002. A new gene structure of
the disintegrin family: a subunit of dimeric disintegrin has a
short coding region. Biochemistry 41, 14248–14254.
Okuda, D., Nozaki, C., Sekiya, F., Morita, T., 2001. Compara-
tive biochemistry of disintegrins isolated from snake venoms:
consideration of the taxonomy and geographical distribution
of snakes in the genus Echis. J. Biochem. 129, 615–620.
Okumura, K., Inoue, S., Ikeda, K., Hayashi, K., 2002.
Identification of b-type phospholipase A2 inhibitor in a
nonvenomous snake, Elaphe quadrivirgata. Arch. Biochem.
Biophys. 408, 124–130.
Ozeki, Y., Matsui, T., Hamako, J., Suzuki, M., Fujimura, Y.,
Yoshida, E., Sishida, S., Titani, K., 1994. C-type galactose-
binding lectin from Bothrops jararaca venom: comparison of
its structure and function with those of botrocetin. Arch.
Biochem. Biophys. 308, 306–310.
Paine, M.J., Desmond, H.P., Theakston, R.D., Crampton, J.M.,
1992. Purification, cloning, and molecular characterization of
a high molecular weight hemorragic metalloproteinase,
jararhagin, from Bothrops jararaca venom. Insights into the
disintegrin gene family. J. Biol. Chem. 267, 22869–22876.
Perales, J., Neves-Ferreira, A.G., Valente, R.H., Domont, G.B.,
2005. Natural inhibitors of snake venom hemorrhagic
metalloproteinases. Toxicon 45, 1013–1020.
Ribeiro, J.M.C., Charlab, R., Pham, V.M., Garfield, M.,
Valenzuela, J.G., 2004. An insight into the salivary tran-
scriptome and proteome of the adult female mosquito Culex
pipiens quinquefasciatus. Insect Biochem. Mol. Biol. 34,
543–563.
Ribeiro, L.A., Jorge, M.T., 1997. Acidente por serpentes do
genero Bothrops: serie de 3.139 casos. Rev. Soc. Bras. Med.
Trop. 30, 475–480.
Rodrigues, V.M., Soares, A.M., Mancin, A.C., Fontes, M.R.,
Homsi-Brandeburgo, M.I., Giglio, J.R., 1998. Geographic
variations in the composition of myotoxins from Bothrops
neuwiedi snake venoms: biochemical characterization and
biological activity. Comp. Biochem. Physiol. Part A Mol.
Integr. Physiol. 121, 215–222.
Saguchi, K.I., Hagiwara-Saguchi, Y., Murayama, N., Ohi, H.,
Fujita, Y., Camargo, A.C.M., Serrano, S.M.T., Higuchi, S.,
2005. Molecular cloning of serine proteinases from Bothrops
jararaca venom gland. Toxicon 46, 72–83.
Sambrook, J., Fritsch, E.F., Maniatis, T., 1989. Molecular
Cloning: A Laboratory Manual, second ed. Cold Spring
Harbor Laboratory, Cold Spring Harbor, NY.
Sanger, F., Nicklen, S., Calson, A.R., 1977. DNA sequencing
with chain terminating inhibitors. Proc. Natl. Acad. Sci. USA
74, 5463–5467.
Sano-Martins, I.S., Santoro, M.L., Castro, S.C.B., Fan, H.W.,
Cardoso, J.L.C., Theakston, D.G., 1997. Platelet aggregation
in patients bitten by the Brazilian snake Bothrops jararaca.
Thromb. Res. 87, 183–195.
Santoro, M.L., Sano-Martins, I.S., 2004. Platelet dysfunction
during Bothrops jararaca snake envenomation in rabbits.
Thromb. Haemost. 92, 369–383.
Scarborough, R.M., Rose, J.W., Naughton, M.A., Phillips, D.R.,
Nannizzi, L., Arfsten, A., Campbell, A.M., Charo, I.F., 1993.
Characterization of the integrin specificities of disintegrins
isolated from American pit viper venoms. J. Biol. Chem. 268,
1058–1065.
Sen, U., Vasudevan, S., Subbarao, G., McClintock, R.A.,
Celikel, R., Ruggeri, Z.M., Varughese, K.I., 2001. Crystal
structure of the von Willebrand factor modulator botrocetin.
Biochemistry 40, 345–352.
Serrano, S.M.T., Hagiwara, Y., Murayama, N., Higuchi, S.,
Mentele, R., Sampaio, C.A.M., Camargo, A.C.M., Fink, E.,
1998. Purification and characterization of a kinin-releasing
and fibrinogen-clotting serine proteinase (KN-BJ) from the
venom of Bothrops jararaca, and molecular cloning and
sequence analysis of its cDNA. Eur. J. Biochem. 251,
845–853.
Serrano, S.M.T., Maroun, R.C., 2005. Snake venom serine
proteinases: sequence homology vs. substrate specificity, a
paradox to be solved. Toxicon 45, 1115–1132.
Shin, Y., Okuyama, I., Hasegawa, J., Morita, T., 2000. Molecular
cloning of glycoprotein Ib-binding protein, flavocetin A,
which inhibits platelet aggregation. Thromb. Res. 99,
239–247.
Soares, M.R., Oliveira-Carvalho, A.L., Wermelinger, L.S.,
Zingali, R.B., Ho, P.L., Junqueira-de-Azevedo, I.L.M., Diniz,
M.R.V., 2005. Identification of novel bradykinin-potentiating
peptides and C-type natriuretic peptide from Lachesis muta
venom. Toxicon 46, 31–38.
Sousa, J.R.F., Monteiro, R.Q., Castro, H.C., Zingali, R.B., 2001.
Proteolytic action of Bothrops jararaca venom upon its own
constituents. Toxicon 39, 787–792.
Takeya, H., Nishida, S., Miyata, T., Kawada, S.I., Saisaka, Y.,
Morita, T., Iwanaga, S., 1992. Coagulation factor X
activating enzyme from Russell’s viper venom (RVV-X): A
novel metalloproteinase with disintegrin (platelet aggregation
inhibitor)-like and C-type lectin-like domains. J. Biol. Chem.
267, 14109–14117.
Thwin, M.M., Satish, R.L., Chan, S.T.F., Gopalakrishnakone,
P., 2002. Functional site of endogenous phospholipase
A2 inhibitor from python serum. Eur. J. Biochem. 269,
719–727.
Valenzuela, J.G., Pham, V.M., Garfield, M.K., Francischetti,
I.B.M., Ribeiro, J.M.C., 2002. Toward a description of the
sialome of the adult female mosquito Aedes aegypti. Insect
Biochem. Mol. Biol. 32, 1101–1122.
Varanda, E.A., Giannini, M.J.M., 1999. Bioquımica de venenos
de serpentes. In: Barraviera, B. (Ed.), Venenos: aspectos
clınicos e terapeuticos dos acidentes por animais pec-onhentos.
EPUB, Rio de Janeiro, pp. 205–223.
Vidal, J.C., Stoppani, A.O.M., 1971. Isolation and properties of
an inhibitor of phospholipase A from Bothrops neuwiedii
venom. Arch. Biochem. Biophys. 147, 66–76.
Vidal, J.C., Stoppani, A.O.M., 1980. Inhibition of phospholipase
A by a naturally occuring peptide in Bothrops venoms.
Experientia 26, 831–836.
Wei, Q., Lu, Q.M., Jin, Y., Li, R., Wei, J.F., Wang, W.Y., Xiong,
Y.L., 2002. Purification and cloning of a novel C-type lectin-
like protein with platelet aggregation activity from Trimer-
esurus mucrosquamatus venom. Toxicon 40, 1331–1338.
Werman, S.D., 1992. Phylogenetic relationships of Central and
South American pit vipers of the genus Bothrops (sensu lato):
cladistic analysis of biochemical and anatomical characters.
In: Campbell, J.A., Brodie Jr. E.D. (Eds.), Biology of Pit
vipers. Tyler, Selva, TX, pp. 21–40.
White, J., 2005. Snake venoms and coagulopathy. Toxicon 45,
951–967.
Xu, Q., Wu, X.F., Xia, Q.C., Wang, K.Y., 1999. Cloning of a
galactose-binding lectin from the venom of Trimeresurus
stejnegeri. Biochem. J. 341, 733–737.

ARTICLE IN PRESSD.A.P. Cidade et al. / Toxicon 48 (2006) 437–461 461
Yamazaki, Y., Morita, T., 2004. Structure and function of
snake venom cysteine-rich secretory proteins. Toxicon 44,
227–231.
Zamuner, S.R., Cruz-Hofling, M.A., Corrado, A.P., Hyslop, S.,
Rodrigues-Simioni, L., 2004. Comparison of the neurotoxic
and myotoxic effects of Brazilian Bothrops venoms and
their neutralization by commercial antivenom. Toxicon 44,
259–271.
Zuliani, J.P., Fernandes, C.M., Zamuner, S.R., Gutierrez, J.M.,
Teixeira, C.F.P., 2005. Inflammatory events induced by Lys-49
and Asp-49 phospholipases A2 isolated from Bothrops asper
snake venom: role of catalytic activity. Toxicon 45, 335–346.




















