
Neutralization of local tissue damage induced by (terciopelo) snake venom
Upload
independentCategory
view
4download
0
Comparative study on the ability of IgG andFab sheep antivenoms to neutralize local
hemorrhage, edema and myonecrosis inducedby Bothrops asper (terciopelo) snake venom
Guillermo Leo n, Juan Manuel Valverde, Gustavo Rojas,Bruno Lomonte, Jose Marõ a Gutie rrez*
Instituto Clodomiro Picado, Facultad de MicrobiologõÂa, Universidad de Costa Rica, San JoseÂ, Costa Rica
Received 3 March 1999; received in revised form 14 May 1999; accepted 14 May 1999
Abstract
The ability of sheep antivenoms, consisting of whole IgG molecules or Fab fragments, toneutralize local hemorrhage, edema and myonecrosis induced by Bothrops asper venom wascomparatively studied in mice. The two antivenoms were produced from the same batch of
hyperimmune plasma and were adjusted to the same neutralizing potency against thesee�ects in assays where venom and antivenoms were incubated prior to injection. Thus, ifdi�erences are observed in experiments involving independent injection of venom and
antivenoms, they would depend on the pharmacokinetic pro®les of the products. Despitethe observation that both antivenoms neutralized the three e�ects if preincubated withvenom, neutralization was only partial when antivenoms were administered i.v. at various
time intervals after envenomation. No signi®cant di�erences were observed between IgGand Fab antivenoms concerning neutralization of hemorrhagic and edema-formingactivities, whereas IgG antivenom was slightly more e�ective in neutralizing myotoxic
activity in experiments involving independent injection of venom and antivenom. Theseresults do not support the hypothesis that Fab fragments are more e�ective than whole IgGmolecules in the neutralization of locally-acting toxins from B. asper venom. # 1999Elsevier Science Ltd. All rights reserved.
0041-0101/99/$ - see front matter # 1999 Elsevier Science Ltd. All rights reserved.
PII: S0041 -0101 (99)00152 -X
Toxicon 38 (2000) 233±244
www.elsevier.com/locate/toxicon
* Corresponding author. Tel.: +506-229-0344; fax: +506-292-0485.
E-mail address: [email protected] (J.Marõ . Gutie rrez)

1. Introduction
Envenomations by pit vipers (family Viperidae, subfamily Crotalinae) arecharacterized by prominent local tissue alterations, i.e. hemorrhage, edema andnecrosis, which may result in permanent sequelae such as tissue loss ordysfunction (Ownby, 1982; Gutie rrez, 1995; Warrell, 1995). These e�ects developvery rapidly after envenomation, thereby making neutralization by antivenomsrather di�cult, especially if serotherapy is delayed due to either late access tomedical care or scarcity of antivenoms (Gutie rrez et al., 1998). In CentralAmerica, a large number of accidents are in¯icted by Bothrops asper, a speciesthat may induce prominent local e�ects (Gutie rrez and Lomonte, 1989; Hardy,1994; Gutie rrez, 1995). It is therefore relevant to search for new ways to improvethe neutralization of local tissue damage in envenomations induced by this andrelated species.
Traditionally, antivenoms in Latin America have been produced byfractionating horse hyperimmune plasma in order to obtain either whole IgG orF(ab ')2 fragments (BolanÄ os and Cerdas, 1980; Raw et al., 1991; Rojas et al.,1994). It has been postulated that sheep Fab fragments, obtained by papaindigestion of immunoglobulins, may constitute a more convenient therapeutic tool(Sullivan, 1987; Dart and Horowitz, 1996). Various Fab antivenoms have beenproduced (Smith et al., 1992; Laing et al., 1995; Landon and Smith, 1996) andsome have been evaluated in clinical trials (Karlson-Stiber et al., 1997; Meyer etal., 1997). Two theoretical advantages of Fab fragments are their larger volume ofdistribution, when compared to IgG and F(ab ')2 preparations, and their ability toreach tissue compartments at a faster rate (Covell et al., 1986; Scherrmann, 1994).These two pharmacokinetic characteristics would be of value in the neutralizationof locally-acting toxins, since Fab fragments might be able to reach and neutralizetoxins present in the tissues more readily than whole IgG and F(ab ')2. The presentstudy tested this hypothesis by comparing the ability of sheep IgG and Fabpolyvalent antivenoms to neutralize local hemorrhage, edema and myonecrosisinduced by B. asper venom in mice.
2. Materials and methods
2.1. Venom and antivenoms
Venom used was a pool obtained from more than 40 adult specimens of B.asper collected in both Atlantic and Paci®c regions of Costa Rica and kept at theSerpentarium of Instituto Clodomiro Picado. Venom was lyophilized and storedat ÿ208C. For antivenom production, four sheep were immunized with increasingdoses of a mixture of the venoms of B. asper, Lachesis muta and Crotalus durissusdurissus, using a similar immunization protocol as that described by Angulo et al.(1997). To obtain whole IgG, plasma was fractionated by caprylic acidprecipitation (Rojas et al., 1994). Brie¯y, plasma pH was adjusted to 7.0 and
G. LeoÂn et al. / Toxicon 38 (2000) 233±244234

caprylic acid was slowly added to a ®nal concentration of 5% (v/v), followed byvigorous stirring for 60 min. After precipitated proteins were separated by®ltration through Whatman 2V paper, the ®ltrate was dialyzed against phosphate-bu�ered saline solution, pH 7.2 (PBS) containing 0.25 g/0.1 L phenol. Then, thepreparations were sterilized by ®ltration through a 0.22 mm nitrocellulosemembrane and dispensed in 10 mL vials.
Fab antivenom was obtained by adjusting the pH of plasma to 7.0, followed bythe addition of 20 g/L papain (Sigma Chemical Co., St Louis, MO) and 1 g/Lcysteine. Digestion was performed at 22±258C for 2 h, after which caprylic acidwas added to attain a ®nal concentration of 5% (v:v). The mixture was vigorouslystirred during 24 h and then ®ltered. The ®ltrate was dialyzed against PBScontaining 0.25 g/0.1 L phenol and sterilized by ®ltration through 0.22 mmmembrane. Before ®nal dispensing of the product in 10 mL glass vials, bothantivenom preparations were adjusted to have the same neutralizing potencyagainst the venom of B. asper, as tested by the indirect hemolytic assay inerythrocyte±egg yolk agarose gels (Gutie rrez et al., 1988). This method correlateswith the neutralization of lethality (Gutie rrez et al., 1988). Antivenoms wereanalyzed by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS±PAGE) under nonreducing conditions, according to Laemmli (1970), using 10%polyacrylamide gels.
2.2. Neutralization studies
2.2.1. Neutralization of hemorrhageFor preincubation-type experiments, mixtures of a constant amount of venom
and various dilutions of antivenoms were prepared and incubated for 30 min at378C. Then, aliquots of 0.1 mL of the mixtures, containing 25 mg of venom, wereinjected intradermally in the abdominal region of groups of four Swiss±Webstermice (18±20 g), as described by Gutie rrez et al. (1985). 2 h after injection, micewere sacri®ced, their skin removed and the hemorrhagic area measured. Controlmice were injected with the same amount of venom without antivenom.Neutralizing ability was expressed as the e�ective dose 50% (ED50), de®ned as theratio mL antivenom/mg venom in which the diameter of hemorrhagic spot wasreduced by 50% when compared to the lesions induced by venom alone (Gutie rrezet al., 1985). In experiments with independent injection of venom and antivenoms,groups of four mice (18±20 g) were initially injected intradermally as describedwith 25 mg venom, dissolved in 0.1 mL PBS. Then, at various time intervals(immediately and at 10, 20 and 30 min), 0.2 mL of antivenom were administeredeither intravenously, in the caudal vein, or intramuscularly, in the rightgastrocnemius. Hemorrhage was assessed in the skin 2 h after venom injection, asdescribed.
2.2.2. Neutralization of edemaIn preincubation-type experiments, mixtures containing a constant amount of
venom and various dilutions of antivenoms were prepared and incubated for 30
G. LeoÂn et al. / Toxicon 38 (2000) 233±244 235

min at 378C. Then, groups of four mice (18±20 g) were injected subcutaneously inthe right foot pad with 50 mL of the mixtures, containing 10 mg venom, whereasthe left foot pad received 50 mL of PBS alone. Control mice were injected with 10mg venom in the right foot pad and 50 mL of PBS in the contralateral pad. 1 hafter injection edema was evaluated as described by Yamakawa et al. (1976).Edema was expressed as the percentage increase in weight of the right foot padcompared to the left foot pad. Neutralization was expressed as ED50, as describedabove. In other experiments, groups of four mice (18±20 g) were injected in theright foot pad with 10 mg venom, dissolved in 50 mL of PBS, whereas the left footpad received 50 mL of PBS alone. Then, 0.2 mL of either IgG or Fab antivenomswas administered i.v. either immediately or after 15 or 30 min. Control micereceived venom and no antivenom. Edema was assessed 1 h after envenomation,as described.
2.2.3. Neutralization of myonecrosisFor preincubation-type experiments, mixtures containing a constant amount of
venom and various dilutions of antivenoms were prepared and incubated for 30min at 378C. Then, 100 mL of the mixtures, containing 50 mg of venom, wereinjected intramuscularly in the right gastrocnemius of groups of four mice (18±20g). Control animals were injected with either 50 mg of venom alone or PBS alone.3 h after injection, a blood sample was collected from the tail into heparinizedcapillary tubes. Plasma creatine kinase (CK) activity was determined by using theSigma kit No. 520. CK activity was expressed as units/mL, one unit resulting inthe phosphorylation of one nanomole of creatine per min at 258C. Neutralizingability was expressed as ED50, as de®ned above. In experiments with independentinjection of venom and antivenoms, groups of four mice were injected i.m. with 50mg venom in the right gastrocnemius. Then, at di�erent time intervals(immediately and at 20 and 40 min), 0.2 mL of either IgG or Fab antivenomswere administered i.v. Control mice were injected with venom and did not receiveantivenom. Plasma CK activity was assessed 3 h after venom injection.
2.2.4. Statistical analysisResults are presented as mean2S.D. (n = 4). Student's t test was used to
determine the signi®cance of the di�erences between the mean values of twoexperimental groups.
3. Results
3.1. Characteristics of antivenoms
Papain digestion, followed by caprylic acid precipitation of nonimmunoglobulinproteins, gave a preparation consisting of highly puri®ed Fab fragments, asevidenced by electrophoretic analysis which showed one band of 50 kDa undernonreducing conditions (results not shown). IgG puri®ed by caprylic acid
G. LeoÂn et al. / Toxicon 38 (2000) 233±244236
precipitation (Rojas et al., 1994) showed a main band of 150 kDa. Bothantivenoms were diluted in order to have a similar neutralizing potency againstindirect hemolytic activity of B. asper venom. ED50s were 410222 and 438227mL antivenom/mg venom for IgG and Fab antivenoms, respectively. Thisdi�erence was not statistically signi®cant ( p > 0.05).
3.2. Neutralization studies
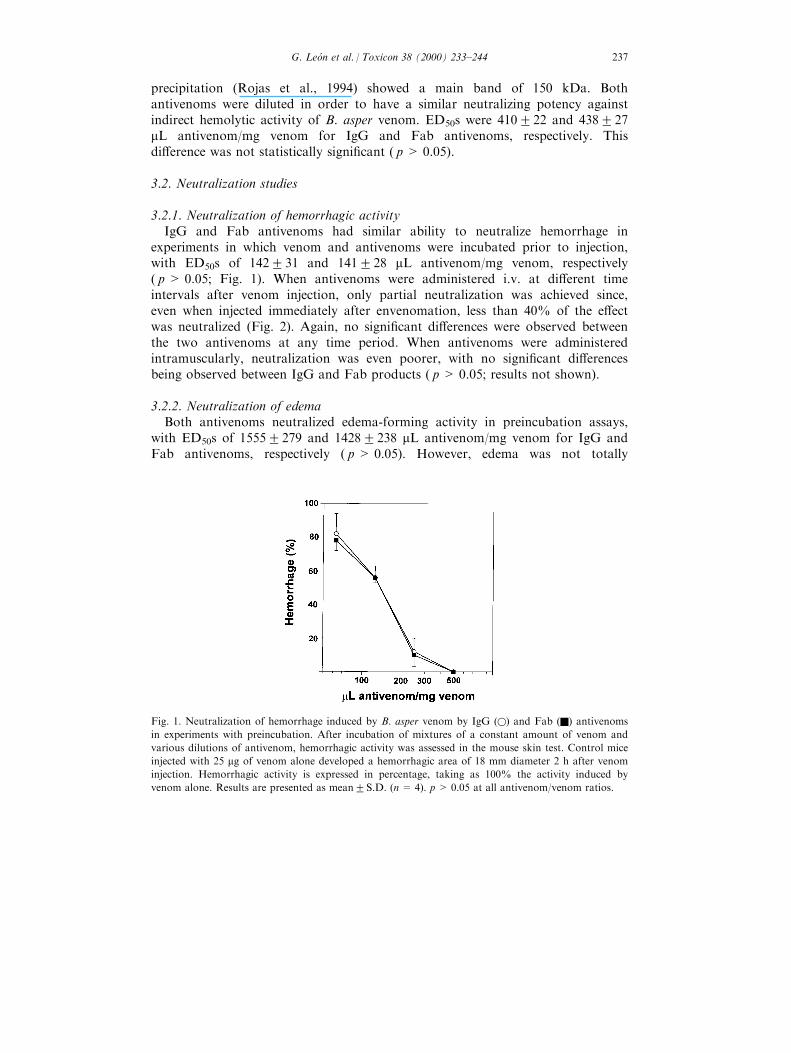
3.2.1. Neutralization of hemorrhagic activityIgG and Fab antivenoms had similar ability to neutralize hemorrhage in
experiments in which venom and antivenoms were incubated prior to injection,with ED50s of 142231 and 141228 mL antivenom/mg venom, respectively( p > 0.05; Fig. 1). When antivenoms were administered i.v. at di�erent timeintervals after venom injection, only partial neutralization was achieved since,even when injected immediately after envenomation, less than 40% of the e�ectwas neutralized (Fig. 2). Again, no signi®cant di�erences were observed betweenthe two antivenoms at any time period. When antivenoms were administeredintramuscularly, neutralization was even poorer, with no signi®cant di�erencesbeing observed between IgG and Fab products ( p> 0.05; results not shown).
3.2.2. Neutralization of edemaBoth antivenoms neutralized edema-forming activity in preincubation assays,
with ED50s of 15552279 and 14282238 mL antivenom/mg venom for IgG andFab antivenoms, respectively ( p > 0.05). However, edema was not totally
Fig. 1. Neutralization of hemorrhage induced by B. asper venom by IgG (w) and Fab (Q) antivenoms
in experiments with preincubation. After incubation of mixtures of a constant amount of venom and
various dilutions of antivenom, hemorrhagic activity was assessed in the mouse skin test. Control mice
injected with 25 mg of venom alone developed a hemorrhagic area of 18 mm diameter 2 h after venom
injection. Hemorrhagic activity is expressed in percentage, taking as 100% the activity induced by
venom alone. Results are presented as mean2S.D. (n= 4). p > 0.05 at all antivenom/venom ratios.
G. LeoÂn et al. / Toxicon 38 (2000) 233±244 237

abbrogated even at the highest antivenom/venom ratio used (Fig. 3). Whenantivenoms were administered i.v. after envenomation, edema neutralization wasrather poor, since even if treatment was performed immediately after venominjection, the e�ect was reduced only to 53% (Fig. 4). No signi®cant di�erenceswere observed in the neutralizing ability of the antivenoms.
Fig. 2. Neutralization of hemorrhage induced by B. asper venom by IgG (q) and Fab (Q) antivenoms
in experiments involving independent injection of venom and antivenoms. Venom (25 mg) was injected
i.d. and then, at various time intervals, 0.2 ml of antivenom were administered i.v. Hemorrhage was
assessed 2 h after envenomation and expressed in percentage, taking as 100% the e�ect induced by
venom alone. Results are presented as mean2S.D. (n= 4). p > 0.05 at all time intervals.
Fig. 3. Neutralization of edema induced by B. asper venom by IgG (w) and Fab (Q) antivenoms in
experiments with preincubation. Various mixtures of venom and antivenom were incubated and tested
in the foot pad assay. Edema was assessed 1 h after injection and expressed in percentage. Edema
induced in control mice, injected with 10 mg of venom alone, was considered as 100% activity. Results
are presented as mean2S.D. (n= 4). p> 0.05 at all antivenom/venom ratios.
G. LeoÂn et al. / Toxicon 38 (2000) 233±244238

3.2.3. Neutralization of myonecrosisMice injected with venom alone had plasma CK activity of 940286 units/mL,
whereas activity of plasma from mice receiving PBS alone was 62213 units/mL.Both products neutralized myotoxicity when preincubated with the venom, havingED50s of 432295 and 5672106 mL antivenom/mg venom for IgG and Fabantivenoms, respectively ( p > 0.05; Fig. 5). When antivenoms were injected aftervenom, neutralization of myotoxicity was only partial even in conditions whereimmunotherapy was performed immediately after envenomation (Fig. 6). IgGantivenom showed signi®cantly higher neutralizing ability at all times tested.
4. Discussion
Antivenoms constitute the mainstay in the treatment of snakebiteenvenomations (Warrell, 1992). However, despite their e�cacy in neutralizingsystemically-acting toxins, clinical and experimental evidence indicate that they areonly partially e�ective in the neutralization of venom-induced local tissue damage(Gutie rrez et al., 1981, 1985, 1998; Warrell, 1992; Theakston, 1996). This has beenrelated to the rapid development of local edema, myonecrosis and hemorrhageafter envenomation, as has been demonstrated in the case of B. asper venom(Gutie rrez et al., 1981, 1984; Moreira et al., 1992; Lomonte et al., 1994; Chaves etal., 1995). Moreover, even when antibodies are present in the circulation andtissues before envenomation, neutralization is only partial (Rucavado andLomonte, 1996). Therefore, on theoretical grounds, the use of antibody fragments
Fig. 4. Neutralization of edema induced by B. asper venom by IgG (q) and Fab (Q) antivenoms in
experiments with independent injection of venom and antivenoms. Venom (10 mg) was injected s.c. in
the right foot pad of mice. Then, at various time intervals, 0.2 ml of antivenom was administered i.v.
Edema was assessed 1 h after venom injection. Activity is expressed in percentage, taking as 100% the
e�ect induced by venom alone. Results are presented as mean2S.D. (n = 4). p> 0.05 at all time
intervals.
G. LeoÂn et al. / Toxicon 38 (2000) 233±244 239

such as Fab, which have a larger volume of distribution than IgG and F(ab ')2 andreach the tissue compartment at a faster rate, would be expected to circumvent
Fig. 5. Neutralization of myonecrosis induced by B. asper venom by IgG (w) and Fab (Q) antivenoms
in experiments involving preincubation of venom and antivenom. Various mixtures of venom and
antivenom were incubated and then injected i.m. in the right gastrocnemius. Myonecrosis was assessed
3 h after injection by determining plasma CK activity. CK activity in control mice, injected with 50 mgof venom alone, was 940286 units/mL. This was considered as 100% myotoxic activity and the rest of
the results were expressed in percentage. Results are presented as mean2S.D. (n= 4). p> 0.05 at all
antivenom/venom ratios.
Fig. 6. Neutralization of myonecrosis induced by B. asper venom by IgG (q) and Fab (Q) antivenoms
in experiments with independent injection of venom and antivenoms. Venom (50 mg) was injected i.m.
in the right gastrocnemius of mice. Then, at various time intervals, 0.2 mL of either IgG or Fab
antivenoms was administered i.v. Myonecrosis was assessed 3 h after envenomation by quantitating
plasma CK activity. Myonecrosis is expressed in percentage, taking as 100% the e�ect induced by
venom alone, which resulted in plasma CK activity of 940286 units/mL. Results are presented as
mean2S.D. (n= 4). IgG antivenom was more e�ective than Fab antivenom at all time intervals
( p< 0.05).
G. LeoÂn et al. / Toxicon 38 (2000) 233±244240

some of the problems involved in the neutralization of locally-acting venomcomponents. A new method to generate Fab fragments was adapted in this study,based on an initial digestion with papain, followed by precipitation ofnonimmunoglobulin proteins by the addition of caprylic acid. This procedureresulted in a highly pure Fab preparation.
In order to appropriately compare antivenoms and to minimize di�erencesbased on parameters other than the pharmacokinetics of antibodies and theirfragments, whole IgG and Fab antivenoms from the same pool of hyperimmunesheep plasma were prepared. Moreover, both antivenoms were standardized as tohave the same neutralizing potency against venom in experiments in which venomand antivenoms were incubated prior to injection. As proposed by Leo n et al.(1997), this would assure that if variations arise in experiments with independentinjection of venom and antivenoms, they would be due to the pharmacokineticpro®le of the products and not to di�erences in their ED50s.
Antivenoms were e�ective in neutralizing hemorrhage, edema and myonecrosisin preincubation-type experiments, con®rming previous observations with wholeIgG and F(ab ')2 equine antivenoms (Gutie rrez et al., 1985; Rojas et al., 1994;Leo n et al., 1997). In contrast, poor neutralization of these local e�ects wasobserved when antivenoms were administered after envenomation. Lack ofneutralization of local tissue damage in this type of experiment has beenpreviously demonstrated with equine antivenoms (Gutie rrez et al., 1981, 1985,1998). Administration of Fab fragments did not improve neutralization whencompared to whole IgG molecules, despite their more rapid di�usion toextravascular sites where locally-acting toxins are present. In the case ofmyotoxicity, IgG antivenom showed a slightly better neutralizing pro®le than Fabfragments.
The lack of improvement in neutralization of local e�ects by Fab fragments,despite their ability to reach the tissue compartment at a faster rate than IgGmolecules, may have to do with the extremely rapid development of local tissuedamage. Edema, myonecrosis and hemorrhage develop within minutes afterinjection of B. asper venom (Gutie rrez et al., 1980, 1984; Moreira et al., 1992;Lomonte et al., 1994; Chaves et al., 1995). On the other hand, the larger volumeof distribution of Fab fragments, together with the rapid renal clearance of thesemolecules (Scherrmann, 1994; Riviere et al., 1997), may decrease their actualconcentration in the tissues where toxins are present, thereby a�ecting theirneutralizing ability. Moreover, as proposed by Leo n et al. (1997) and Gutie rrez etal. (1998), local microvessel alteration caused by edema-forming and hemorrhagictoxins may induce a similar extravasation of IgG and Fab, suggesting thatantibody pharmacokinetics in envenomated and control limbs may di�ersubstantially, reducing the impact of di�erences in the molecular weight ofantibody fragments. Finally, it may be that neutralization of toxins in tissues isnegligible and the most important mechanism of neutralization is antibodybinding in the circulation, with the consequent redistribution of toxins from thetissues to the vascular compartment (Choumet et al., 1996; Riviere et al., 1997).
Independently of the possible mechanisms discussed above, our ®ndings
G. LeoÂn et al. / Toxicon 38 (2000) 233±244 241

demonstrate that an Fab sheep polyvalent antivenom is not superior to a wholeIgG antivenom in neutralizing local hemorrhage, edema and myonecrosis inducedby B. asper venom in mice. These results, taken together with previousobservations of Lomonte et al. (1996) and Leo n et al. (1997), evidence that the useof antibody fragments does not improve the ability of antivenoms to neutralizevenom-induced local e�ects. It is necessary to search for novel therapeutic agentsthat may complement antivenoms in the di�cult task of neutralizing locally-actingtoxins present in snake venoms.
Acknowledgements
The authors thank Ricardo Estrada, Javier Nu nÄ ez, Rodrigo Chaves and JorgeSanabria for their valuable collaboration in this work. This study was supportedby Vicerrectorõ a de Investigacio n, Universidad de Costa Rica (project 741-89-057).
References
Angulo, Y., Estrada, R., Gutie rrez, J.M., 1997. Clinical and laboratory alterations in horses during im-
munization with snake venoms for the production of polyvalent (Crotalinae) antivenom. Toxicon
35, 81±90.
BolanÄ os, R., Cerdas, L., 1980. Produccio n y control de sueros antiofõ dicos en Costa Rica. Boletõ n de la
O®cina Sanitaria Panamericana 88, 184±196.
Chaves, F., Barboza, M., Gutie rrez, J.M., 1995. Pharmacological study of edema induced by venom of
the snake Bothrops asper (terciopelo) in mice. Toxicon 33, 31±39.
Choumet, V., Audebert, F., Riviere, G., Sorkine, M., Urtizberea, M., Sabouraud, A., Scherrmann,
J.M., Bon, C., 1996. Toxicokinetics of Vipera aspis envenoming and antivenom therapy. In: Bon,
C., Goy�on, M. (Eds.), Envenomings and Their Treatments. Editions Fondation Marcel Me rieux,
Lyon, pp. 127±133.
Covell, D.G., Barbet, J., Holton, O.D., Black, C.D.V., Parker, R.J., Weinstein, J.N., 1986.
Pharmacokinetics of monoclonal immunoglobulin G1, F(ab ')2 and Fab ' in mice. Cancer Res. 46,
3969±3978.
Dart, R.C., Horowitz, R.S., 1996. Use of antibodies as antivenoms: a primitive solution for a complex
problem? In: Bon, C., Goy�on, M. (Eds.), Envenomings and Their Treatments. Editions Fondation
Marcel Me rieux, Lyon, pp. 83±94.
Gutie rrez, J.M., Arroyo, O., BolanÄ os, R., 1980. Mionecrosis, hemorragia y edema inducidos por el
veneno de Bothrops asper en rato n blanco. Toxicon 18, 603±610.
Gutie rrez, J.M., Chaves, F., BolanÄ os, R., Cerdas, L., Rojas, E., Arroyo, O., Portilla, E., 1981.
Neutralizacio n de los efectos locales del veneno de Bothrops asper por un antiveneno polivalente.
Toxicon 19, 493±500.
Gutie rrez, J.M., Ownby, C.L., Odell, G.V., 1984. Pathogenesis of myonecrosis induced by crude venom
and a myotoxin of Bothrops asper. Exp. Mol. Pathol. 40, 367±379.
Gutie rrez, J.M., Gene , J.A., Rojas, G., Cerdas, L., 1985. Neutralization of proteolytic and hemorrhagic
activities of Costa Rican snake venoms by a polyvalent antivenom. Toxicon 23, 887±893.
Gutie rrez, J.M., Avila, C., Rojas, E., Cerdas, L., 1988. An alternative in vitro method for testing the
potency of the polyvalent antivenom produced in Costa Rica. Toxicon 26, 411±413.
Gutie rrez, J.M., Lomonte, B., 1989. Local tissue damage induced by Bothrops snake venoms. Mem.
Inst. Butantan 51, 211±223.
Gutie rrez, J.M., 1995. Clinical toxicology of snakebite in Central America. In: Meier, J., White, J.
G. LeoÂn et al. / Toxicon 38 (2000) 233±244242

(Eds.), Handbook of Clinical Toxicology of Animal Venoms and Poisons. CRC Press, Boca Raton,
FL, pp. 645±665.
Gutie rrez, J.M., Leo n, G., Rojas, G., Lomonte, B., Rucavado, A., Chaves, F., 1998. Neutralization of
local tissue damage induced by Bothrops asper (terciopelo) snake venom. Toxicon 36, 1529±1538.
Hardy, D., 1994. Snakebite and ®eld biologists in Me xico and Central America: report of ten cases
with recommendations for ®eld management. Herpetol. Nat. Hist. 2, 67±82.
Karlson-Stiber, C., Persson, H., Smith, D., Al-Abdulla, I.H., Sjostrom, L., 1997. First clinical experi-
ences with speci®c sheep Fab fragments in snake bite. Report of a multicentre study of Vipera berus
envenoming. J. Int. Med. 241, 53±58.
Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage
T4. Nature 227, 680±685.
Laing, G.D., Lee, L., Smith, D.C., Landon, J., Theakston, R.G.D., 1995. Experimental assessment of a
new, low-cost antivenom for treatment of carpet viper (Echis ocellatus ) envenoming. Toxicon 33,
307±313.
Landon, J., Smith, D.C., 1996. Development of novel antivenoms based on speci®c ovine Fab. In: Bon,
C., Goy�on, M. (Eds.), Envenomings and Their Treatments. Editions Fondation Marcel Me rieux,
Lyon, pp. 173±180.
Leo n, G., Rojas, G., Lomonte, B., Gutie rrez, J.M., 1997. Immunoglobulin G and F(ab ')2 polyvalent
antivenoms do not di�er in their ability to neutralize hemorrhage, edema and myonecrosis induced
by Bothrops asper (terciopelo) snake venom. Toxicon 35, 1627±1637.
Lomonte, B., Lundgren, J., Johansson, B., Bagge, U., 1994. The dynamics of local tissue damage
induced by Bothrops asper snake venom and myotoxin II on the mouse cremaster muscle: an intravi-
tal and electron microscopic study. Toxicon 32, 41±55.
Lomonte, B., Leo n, G., Hanson, L.A., 1996. Similar e�ectiveness of Fab and F(ab ')2 antivenoms in the
neutralization of hemorrhagic activity of Vipera berus snake venom in mice. Toxicon 34, 1197±1202.
Meyer, W.P., Habib, A.G., Onayade, A.A., Yakubu, A., Smith, D.C., Naidi, A., Daudu, I.J., Warrell,
D.A., Theakston, R.D.G., 1997. First clinical experiences with a new ovine Fab Echis ocellatus
snake bite antivenom in Nigeria: randomized comparative trial with Institute Pasteur serum
(IPSER) Africa antivenom. Am. J. Trop. Med. Hyg. 56, 291±300.
Moreira, L., Gutie rrez, J.M., Borkow, G., Ovadia, M., 1992. Ultrastructural alterations in mouse capil-
lary blood vessels after experimental injection of venom from the snake Bothrops asper (terciopelo).
Exp. Mol. Pathol. 57, 124±133.
Ownby, C.L., 1982. Pathology of rattlesnake envenomation. In: Tu, A.T. (Ed.), Rattlesnake Venoms.
Their Actions and Treatment. Marcel Dekker, New York, pp. 163±209.
Raw, I., Guidolin, R., Higashi, H.G., Kelen, E.M.A., 1991. Antivenins in Brazil: preparation. In: Tu,
A.T. (Ed.), Handbook of Natural Toxins, vol. 5. Marcel Dekker, New York, pp. 557±581 (Reptile
Venoms and Toxins).
Riviere, G., Choumet, V., Audebert, F., Sabouraud, A., Debray, M., Scherrmann, J.M., Bon, C., 1997.
E�ect of antivenom on venom pharmacokinetics in experimentally envenomed rabbits: toward an
optimization of antivenom therapy. J. Pharmacol. Exp. Ther. 281, 1±8.
Rojas, G., Jime nez, J.M., Gutie rrez, J.M., 1994. Caprylic acid fractionation of hyperimmune horse
plasma: description of a rapid procedure for antivenom production. Toxicon 32, 351±363.
Rucavado, A., Lomonte, B., 1996. Neutralization of myonecrosis, hemorrhage and edema induced by
Bothrops asper snake venom by homologous and heterologous preexisting antibodies in mice.
Toxicon 34, 567±577.
Scherrmann, J.M., 1994. Antibody treatment of toxic poisoning, recent advances. Clin. Toxicol. 32,
363±375.
Smith, D.C., Reddi, K.R., Laing, G., Theakston, R.G.D., Landon, J., 1992. An a�nity puri®ed ovine
antivenom for the treatment of Vipera berus envenoming. Toxicon 30, 865±871.
Sullivan, J.B., 1987. Past, present and future of immunotherapy of snake venom poisoning. Ann.
Energ. Med. 16, 938±944.
Theakston, R.D.G., 1996. Snake bite: the kinetics of envenoming and therapy. In: Bon, C., Goy�on,
M. (Eds.), Envenomings and Their Treatments. Editions Fondation Marcel Me rieux, Lyon, pp.
117±126.
G. LeoÂn et al. / Toxicon 38 (2000) 233±244 243

Warrell, D.A., 1992. The global problem of snake bite: its prevention and treatment. In:
Gopalakrishnakone, P., Tan, C.R. (Eds.), Recent Advances in Toxicology Research, 1. National
University of Singapore, Singapore, pp. 121±153.
Warrell, D.A., 1995. Clinical toxicology of snakebite in Asia. In: Meier, J., White, J. (Eds.), Handbook
of Clinical Toxicology of Animal Venoms and Poisons. CRC Press, Boca Raton, FL, pp. 493±594.
Yamakawa, M., Nozaki, M., Hokama, Z., 1976. Fractionation of sakishima-habu (Trimeresurus ele-
gans ) venom and lethal, hemorrhagic and edema-forming activities of the fractions. In: Ohsaka, A.,
Hayashi, A., Sawai, Y. (Eds.), Animal, Plant and Microbial Toxins. Plenum Press, New York, pp.
97±109.
G. LeoÂn et al. / Toxicon 38 (2000) 233±244244
Copyright © 2022 FDOKUMEN










