
Dopaminergic neurotoxicity of HIV-1 gp120: Reactive oxygen species as signaling intermediates

ELSEVlER
Vaccine, Vol. 15, No. 11, pp. 1200-1208, 1997 0 1997 Elsevier Science Ltd. All rights reserved
Printed in Great Britain PII: SO264-410X(97)00012-1 0264-410X/97 $17+0.00
Broadly reactive antibodies against a gp120 V3 loop multi-epitope polypeptide neutralize different isolates of human immunodeficiency virus type 1 (HIV-l)
Marinieve Montero”, Alfred0 Menkndez”, Maria de1 Carmen Dominguez”, Leonor Navea?, Olga Lidia Vilarubia?, Di6genes Quintana”, Marisela Izquierdot, Victor Jimknez$, Osvaldo Reyes$, Leonor Lobaina?_, Enrique Noa-/- and Carlos A. Duarte*§
A gene encoding for a novel multi-epitope polypeptide (TAB4) was synthesized and expressed in Escherichia coli. The protein was composed of 15 amino acid fragments derived from the V3 loop of HIV-1 isolates MN, IIIB, Rl$ JYl, BRVA and LR1.50, joined by five-amino-acid linkers. Immunogenic@ of TAB4 in rabbits was studied, and the antibody response against individual peptides investigated. TAB4 was shown to be immunogenic in Complete Freund’s Adjuvant in a dose-dependent mannel; and was able to elicit a humoral response against all V3 epitopes included on the protein. Sera from some of the animals were able to neutralize the replication of viral strain MN, and in one case IIIB, with moderate titers. Some sera also neutralized several Cuban clinical strains, isolated in peripheral blood mononuclear cells, after one round of amplification in MT4 cells. 0 1997 Elsevier Science Ltd.
The third variable region (V3 loop), between cysteine residues 296 and 331 of the external glycoprotein gp120 is considered to play a key role in viral infec- tivity, tropism, and syncytium formation I-3. In this segment of the protein, the principal neutralizing domain (PND), which is able to stimulate the produc- tion of HIV-1 neutralizing antibodies in animals and humans”’ has been located. Different laboratories have also proved that this region contains epitopes recog- nized by cytotoxic T lymphocytes’&” and helper T cells”,‘4. Several groups have demonstrated protection against challenge with the homologous HIV-l strain by passive immunization with anti-V3 monoclonal antibodies either in the SCID mouseI or in chimpan- zees’7,‘8. Antibodies raised against synthetic peptides representing the V3 region are mainly type-specific’g-25,
*Vaccine Division, Center for Genetic Engineering and Biotechnology, P.O. Box 6162, Cubanackn, Havana 10600, Cuba. tNational Reference Center for AIDS, Havana, Cuba. *Chemical-Physics Division, Center for Genetic Engineer- ing and Biotechnology, P.O. Box 6162, CubanacAn, Havana 10600, Cuba. To whom correspondence should be addressed. Tel.: 53-7-218008. 218466: fax: 53-7- 218070; e-mail: [email protected] (Received 27 September 1996; revised 27 November 1996; accepted 17 December 1996)
although the generation of broadly reactive antibodies has been occasionally documented”-‘*. Some labora- tories have demonstrated that is possible to elicit antibodies against more than one V3 region by immunizin
Ii with a combination of two or more
peptides29.3 , or with hybrid peptides”‘. These facts suggested that a cocktail of V3 peptides could be an appropriate immunogen for the development of a protective response against HIV-l.
The authors have previously described the produc- tion of a recombinant HIV-l multi-epitope polypeptide (MEP) in Eschetichia coZiz3. That protein included the central part of V3 regions from three related HIV-l isolates (MN, SC and WMJ-II), and was shown to induce neutralizing monoclonal antibodies against HIV-lhlN. In the present work, a second MEP (TAB4) was designed, produced and evaluated, and it was demonstrated that presentation of more divergent V3 epitopes within a MEP format still allowed the genera- tion of a humoral immune response in rabbits, directed against all those peptides. It was also shown that anti- HIV-l neutralizing antibodies are present in anti- TAB4 sera. Furthermore, the authors characterized the anti-TAB4 antibody response for ELISA cross- reactivity with V3 peptides representing consensus sequences according to phenetic analysis.
1200 Vaccine 1997 Volume 15 Number 11

Reactive antibodies against gp120 neutralize HIV-7: CA. Duarte al.
MATERIALS AND METHODS
Bacterial strains and growth conditions E. co/i strain W3I 10 rrpA 905 was used for expres-
sion experiments. Cells harboring plasmids were grown overnight at 37°C in 5 ml LB broth supplemented with ampicillin (50 /lg ml-‘) and tryptophan (100 pg ml-‘). Cultures were transferred to 300 ml of LB medium supplemented with ampicillin (50 /lg ml-‘) at an initial OD,,,, between 0.01 and 0.1. The cultures were grown for 12 h at 37°C.
HIV-l isolates
HIV-1 MN and HIV-l LAI were obtained through the MRC AIDS Reagent Program, UK, donated by Robert Gallo and Luc Montagnier.
Genetic constructions
Synthesis of the gene was performed according to JimCnez et al.“‘. Twenty-five oligonucleotides were synthesized in an automated Gene Assembler (PharmaciaiBiotech, Sollentuna, Sweden) using P-cyanoethyl phosphoramidite. They were purified, phosphorylated, and ligated as described.
All genetic manipulations were carried out following the methods of Sambrook et al.““. Restriction and modification enzymes were obtained from Amersham International (Amersham, UK) or Biolabs (New England, USA) and used according to the manufac- turers’ recommendations.
Plasmid pI/IL2 has been described elsewhereX4; it contains the tryptophan promoter and the N-terminus of the human IL-2 gene. It was cleaved using the restriction enzymes XhaI-BumHI; the resulting large fragment was purified and ligated to the synthetic gene carrying cohesive ends using X&I and BarnHI. The resulting plasmid was named pTAB3.
Plasmid pTAB3 was digested with enzymes ScaI and HindIII. The short fragment was purified by 1% low gelling temperature agarose electrophoresis. It was then ligated to a vector derived from the pUC19 digested with the same enzymes. The final plasmid was called pTAB4. The DNA sequence of the gene cloned in plasmid pTAB4 was verified by the Sanger method”“.
Protein purification
Cells were harvested by centrifugation at 12OOOg at 4°C and suspended in Tris (10 mM)-EDTA (1 mM) pH 8.0 (TE) at 0.1 g biomass ml-‘. Lysozyme was added at a final concentration of 25 pg ml-‘, and incubated for 1 h with gentle, occasional mixing. After sonication at 4”C, the recovered pellet was washed with TE+8 M urea and then dissolved in TE+6 M guani- dium chloride. This material was centrifuged for 10 min at 10000 rev min-’ and the supernatant loaded into a 10 mm x250 mm C4 reverse-phase column (Vydac, Hesperia, CA, USA) for high-performance liquid chromatography (HPLC) (PharmaciaiBiotech). A continuous gradient from 10 to 80% acetonitrile in 0.1% trifluoroacetic acid was used for the elution. A flow rate of 1.5 ml min-’ was employed and absorbance at 226 nm monitored. The purity was assessed by sodium dodecyl sulphate-polycrylamide gel electro- phoresis (SDS-PAGE)36.
Peptide synthesis
Peptides from the V3 region were synthesized according to the method of simultaneous multiple peptide synthesis3’, and cleaved with liquid hydrogen fluoride. The composition was controlled by amino acid analysis and purity assessed by HPLC in a Cl8 reverse- phase chromatographic column (Vydac). The following peptides were used:
(a) Peptides representing the V3 epitopes of TAB4
Clade
SRGIRIGPGRAILAT LR150 B RKRITMGPGRWWT BRVA B RKSITKGPGRVIYAT RF B SIRIQRGPGRAFVT IIIIB B RQSTPIGLGQALY-IT JYl D RKRIHIGPGRAFYTT MN B
(b) V3 loop consensus peptides according to phenetic analysis
KSIHIGPGRAFYATG Cl KSIRIGPGQAFYATG c2 KSIPIGPGRAFYATG c3 KRlTlGPGRVFYlTG c4 RRRlHGPGRAFYlTK C5 TSITIGPGQVFYRTG C6 QSTHIGPGQALYTTK c7 RQRTSIGQGQALYTT C8 RQRTPIGLGQALYTT c9 RQSTPIGLGQALYTT Cl0
(c) Negative peptide from NEF
LDRAWKGFCYKMG
ELISA with anti-V3 monoclonal antibodies (mAbs)
Ninety-six well polystyrene microplates (High binding, Costar), were coated with 100 ~1 of recombi- nant proteins at 2 pg ml-‘, and incubated for 3 h at 37°C. After one wash with 0.05% Tween 20 in distilled water, plates were blocked with 0.5% bovine serum albumin in PBS (blocking solution) for 1 h at 37°C. Twofold serial dilutions of mAbs 2C4*’ and P4D1e8 in 0.05% Tween 20 in blocking solution were added, starting from 1 iig ml-‘. After three washes the reaction was developed with an anti-mouse peroxidase conjugated rabbit antibody for 1 h at 37”C, and ortho- phenylenediamine (OPD) substrate as previously described.
Preparation of immunogens
Purified recombinant proteins were adsorbed in aluminum hydroxide gel, at 60 /ig adjuvant pg-’ protein. The mixture was incubated for 1 h at 37°C. To evaluate the efficiency of the procedure the sample was centrifuged for 5 min at 1000 rev min-’ and the protein concentration in the supernatant determined by the method of Bradford”. Only samples with > 99% of protein adsorbed were used.
Other sets of immunogens were prepared by mixing the protein with an equivalent volume of either Complete (CFA) or Incomplete Freund’s Adjuvant (IFA) (Sigma, St Louis, MO, USA) and emulsified by strong agitation for 1 h at 2000 rev min-’ with a universal stirrer.
Vaccine 1997 Volume 15 Number 11 1201

Reactive antibodies against gp7.20 neutralize HIV-l: C.A. Duarte et al.
Immunization of rabbits
New Zealand White rabbits (2-2.5 kg body weight at the beginning of the experiment) were used. Three groups of rabbits were immunized with four subcutan- eous inoculations of TAB4 on days 0, 30, 64, and 92. Group A (n = 2) received 200 pg TAB4 in FCA, group B (n = 2) received 50 pg TAB4 in FCA, and group C (n = 4) received 200 /lg TAB4 in AIOH,. Animals were bled at days 64, 92, and 122.
ELISA for titration of rabbit sera
Plates were coated with the recombinant protein and blocked as described above. When synthetic peptides were used, the plates were allowed to dry at 37°C for ~20 h. Serum samples were serially diluted in 0.5% Tween 20 in blocking solution, from 1:lOO to 1:204800 for peptides, and from 1:12800 to 1:l.S x lo6 for proteins. Plates were washed five times and subse- quently incubated with a 1:4000 dilution of goat anti- rabbit Ig-biotin conjugate and a 1:4000 dilution of streptavidin-peroxidase (Amersham International). Plates were washed again five times and hydrogen peroxide and OPD 0.05% were used to detect bound antibodies. All sera were analyzed in duplicate. Cut-off values were calculated for each plate, as twice the absorbance value obtained for a pool of rabbit preim- mune sera. The last dilution that gave absorbance values higher than the cut-off was regarded as the serum titer.
Fine mapping with MN substituted peptides
A group of 12 substituted MN peptides (12-mer) were synthesized, each with a single alanine substitu- tion in any of the positions of the sequence KRIHIGP- GRAFY. Sera from animals of group A were used. A dilution that gave absorbance values between 1.0 and 1.5 in an ELISA against the MN peptide was selected for each serum. ELISA microplates were coated with 100 ~1 of 2 pg ml-’ solution of each peptide and incubated with the selected dilution of the sera for 1 h at 37°C. After completion of the ELISA as described above, the percent reactivity for each substituted peptide was calculated by the formula R = ( AbsSP - AbsNC)/(AbsMN - Abs( NC) x 100, where R is percent reactivity for the given peptide, AbsSP is absorbance for the substituted peptide, AbsNC is absorbance for the negative serum, and AbsMN is absorbance for the parental 12-mer MN peptide.
Classical neutralization assay
The neutralization assay was carried out as describedz4. Briefly, several dilutions of rabbit sera in RPM1 1640 medium with 10% fetal calf serum were incubated in triplicate together with 100 TCIDs,, of HIV-l LAI, MN, or Cuban isolates for 1 h at 37°C in 96-well flat bottom microplates. A total of 1 x lo-’ MT4 cells (or peripheral blood mononuclear cells (PBMC) for clinical isolates) was added to each well and incubated at 37°C. After 24 h, 150 {tl of medium were replaced. On the third day in culture the cells were suspended by gently pipetting, and 100 111 of the suspension were replaced by fresh medium. Viral production was estimated on day 7 by p24 ELISA (DAVIHLAB, San Jo&, La Habana, Cuba). Clinical
1202 Vaccine 1997 Volume 15 Number 11
isolates were obtained from seropositive patients by co-cultivation with PBMC stimulated with phytohemag- glutinin (PHA; DAVIHLAB), IL-2 (10 IU ml-‘) and Polybrene (Sigma). Before the neutralization assay these isolates underwent a single round of amplifica- tion in MT4 cells. Experiments were repeated three times.
Statistical procedures
Antibody titers against peptides were logarithmically transformed and means compared using Student’s t-test. The frequency of response against peptides between two groups was compared using 2 x 2 contin- gency tables. The CSS-Statistica program was used.
RESULTS
Synthesis and expression of TAB4
A DNA fragment encoding for 15 amino acid V3 epitopes, from six different HIV-l isolates, was designed and synthesized. The V3 peptides included in this gene represent the HIV-l isolates LR-150 (sequence listed with number 150 in 40), JYl, MN, RF, BRVA and IIIB. These segments are joined by the five-amino-acid linker AGGGA, as depicted in Figure I.
The gene tub3 was fused to a DNA fragment encoding for the N-terminus (26 amino acids) of the human IL-2 gene, resulting in an open reading frame encoding for a product of 143 amino acids, cloned under the control of the tryptophan promoter. The resulting gene was named tub4 and the plasmid pTAB4.
E. coli W3110 trp4 905 was transformed with pTAB4 and the cells were assayed for the expression of the protein. TAB4 was expressed to very high levels, as shown by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (Figure 2, lane 2). The protein was purified as described above and recovered with > 95% purity after a single chromatographic step, as shown in Figure 2, lane 3.
Reactivity of TAB4 with mAbs 2C4 and P4DlO
TAB4 was evaluated for its ability to react with two anti-HIV-l monoclonal antibodies in ELISA. The mAb
IL-2 LRlSO
MAPTSSSTAQTQLQLEHLLLDLQlFLDsRG~Rl
JYl
GPGRAILATAGGGARQSTPIGLGQALYT
TAGGGARKSI~KGPGRVIYATAGGGARKR
IHI&GRAFYTTAGGGARKR~GPGRV
YYTTAGGGASIRIQRGPGRAFVTI Figure 1 Amino acid sequence of the multi-epitope polypeptide TAB4

Reactive antibodies against gp120 neutralize HIV-l: CA. Duatte et al.
2C4 showed high affinity for V3 peptides from HIV-lMN and homologous isolates” but did not recog- nize HIV-I,,,, at all, whereas mAb P4DlO has the opposite properties. As shown in Figure 3, both antibodies were able to react with TAB4 with similar intensity. These results demonstrated that at least the MN and IIIB epitopes are properly exposed on the protein.
Immunogenicity of TAB4 in rabbits
After four inoculations with the recombinant protein, sera from immunized animals were tested in ELISA for antibody reactivity against TAB4 and the synthetic peptides from the V3 region (Table Z). Rabbits injected with TAB4 in FCA (groups A and B)
123 --
5=F
-67
-42
-25
-21
Figure 2 Expression of the TAB4 protein in E. co/i. Sodium dodecyl sulfate-polyacrylamide gel electrophoresis of: lane 1, W3110 trpA 905, negative control; lane 2. W3110 transformed with plasmid pTAB4; lane 3, purified TAB4
3.0
2.5
2 a 1 .o
developed a strong anti-TAB4 antibody response, with titers above 1:lO’. These were at least 1 log higher than those induced by the same protein formulated in AlOH (group C).
The antibody response against V3 peptides was also > 100 times stronger with FCA (group A) than AlOH
(group C) (PcO.05). Additionally, whereas in group A there was 100% reactivity against the six peptides presented in TAB4, animals in group C showed only 45% reactivity.
When the response against peptides was compared between groups A (geometric mean 65765) and B (geometric mean 13718) which differed only in the dose employed, there was a significant difference in the geometric mean of antibody titers (PcO.05). Each rabbit in the lower dose group failed to react against one of the V3 peptides.
The lowest antibody titers among V3 peptides corre- sponded to LR150 and BIB, while the response was stronger for the epitopes located at the central part of TAB4. More experimental data should be obtained to determine the statistical significance of these observations.
Classical neutralization assay Sera from animals in groups A and B with the
highest anti-V3 titers were selected to evaluate their capacity to neutralize the HIV-l isolates MN and LAI.
Three of four sera were able to neutralize > 90% of replication of the MN isolate (Table 2). All of them were able to neutralize 50% of replication (not shown). The titers were slightly higher in group A, where the highest dose of TAB4 was employed, and demon- strated the strongest reactivity in ELISA against this peptide. Only one of the four sera (no. 3) displayed neutralizing activity for the LA1 strain. The ELISA titer of this serum against the homologous BIB peptide was not higher than the rest.
Table 2 also shows the results of the neutralization of Cuban isolates. Four of the ten isolates tested were neutralized by anti-TAB4 sera. These four isolates were also neutralized by mAb 2C4 against the MN V3
I
P4Dl O-TAB1
- P4Dl O-TAB4
-A- 2C4-TAB1 -x- 2C4-TAB4
19 38 75
Mabs (ng/ml) Figure 3 ELISA reactivity of TAB4 with mAbs 2C4 (MN) and P4DlO (IIIB). TAB1 protein, used as control, only contains the MN epitope
Vaccine 1997 Volume 15 Number 11 1203
Reactive antibodies against gp720 neutralize HIV- 1: C.A. Duarte et al.
Table 1 Antibody titers against TAB4 and V3 peptides in rabbits immunized with TAB4”
Peptide
TAB4 LR-150 JYl RF MN BRVA IllB < vsp sp = “0.5”
Group A (n = 2) Group B (n = 2) Group C (n = 4)
Rabbit 1 Rabbit 2 Rabbit 3 Rabbit 4 Rabbit 5 Rabbit 6 Rabbit 7 Rabbit 8
8~10~ 204 800 102 400 102 400 819200 >2x106 25600
204 800 1038 400
819200 102 400
12800
>2x106 8~10~ 6400 <lOO
102 400 25600 102 400 204 800 204 800 102400
6400 25 600 12800 1600
200 <lOO <lOO <lOO 6400 <too 200 <lOO <lOO 12800
51200 6400 <lOO 3200 800 51200 200 <lOO <lOO 1600 12800 <lOO 6400 400 3200
1600 <lOO <lOO ilO0 <lOO
65765 13718 1837.9 100 83 45.8
“Rabbits were immunized with TAB4 in FCA (groups A and B) or AlOH (group C) with 2OOpg (groups A and C) or 5Oilg (group B); bgeometric mean of the inverse of the antibody titers of positive sera; “percent reactivity against V3 synthetic peptides
loop but none was neutralized by mAb P4DlO against the IIIB-LA1 V3 region.
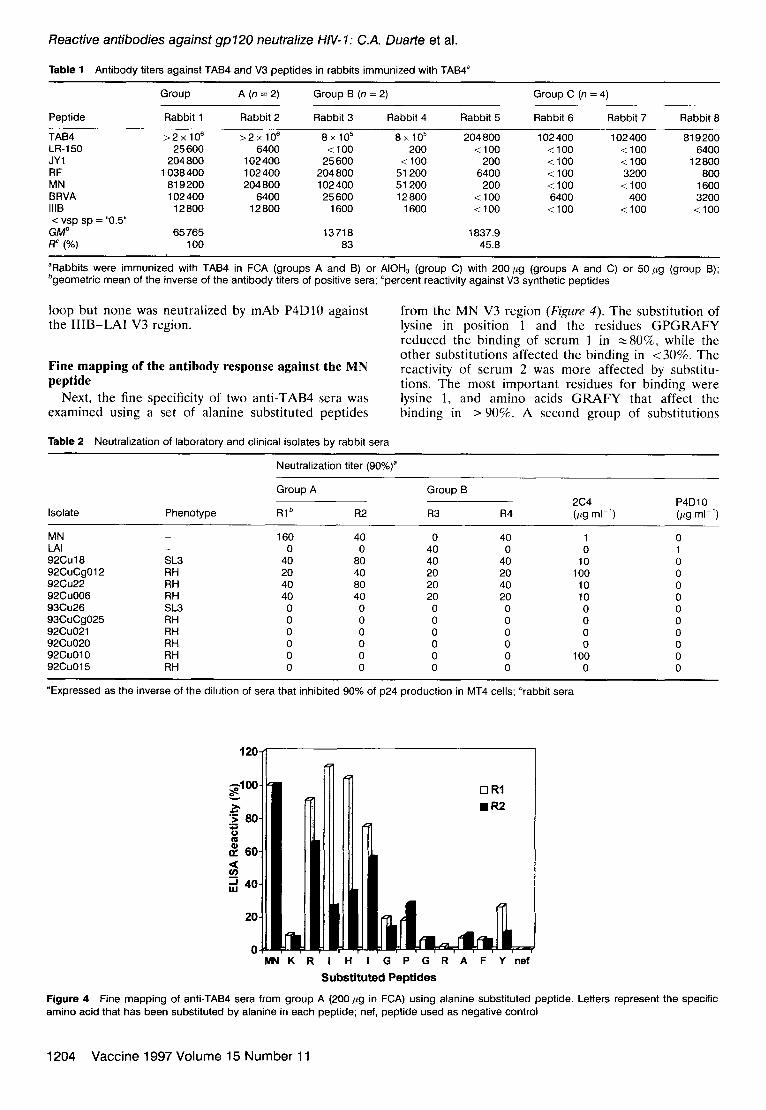
Fine mapping of the antibody response against the MN peptide
Next, the fine specificity of two anti-TAB4 sera was examined using a set of alanine substituted peptides
from the MN V3 region (Figure 4). The substitution of lysine in position 1 and the residues GPGRAFY reduced the binding of serum 1 in zgO%, while the other substitutions affected the binding in ~30%. The reactivity of serum 2 was more affected by substitu- tions. The most important residues for binding were lysine 1, and amino acids GRAFY that affect the binding in > 90%. A second group of substitutions
Table 2 Neutralization of laboratory and clinical isolates by rabbit sera
Neutralization titer (90%)a
Isolate Phenotype
Group A
Rib R2
Group B
R3 R4 2C4 P4DlO
kg ml-‘) (fig ml-‘)
MN _ 160 40 0 40 1 LAI 0 0 40 0 0 92Cu18 SL3 40 80 40 40 10 92cucgo12 RH 20 40 20 20 100 92cu22 RH 40 80 20 40 10 920006 RH 40 40 20 20 10 93Cu26 SL3 0 0 0 0 0 93cucgo25 RH 0 0 0 0 0 92cuo21 RH 0 0 0 0 0 92cuo20 RH 0 0 0 0 0 92CuOlO RH 0 0 0 0 100 92cuo15 RH 0 0 0 0 0
“Expressed as the inverse of the dilution of sera that inhibited 90% of p24 production in MT4 cells; ?abbit sera
0 1 0 0 0 0 0 0 0 0 0 0
120-
20-
03
f
OR1
n R2
MKRIHIGPGRAFYnef
Substituted Peptides
Figure 4 Fine mapping of anti-TAB4 sera from group A (200 pg in FCA) using alanine substituted peptide. Letters represent the specific amino acid that has been substituted by alanine in each peptide; nef, peptide used as negative control
1204 Vaccine 1997 Volume 15 Number 11

Reactive antibodies against gp720 neutralize HIV-l: CA. Duarte et al.
that affects binding from 60 to 90% includes isoleucine 3, histidine 4, glycine 6 and proline 7. Substitutions in arginine 2 and isoleucine 5 affected the binding in <50%.
Cross-reactivity with phenetic V3 consensus peptides
Finally, it was evaluated to what extent antibodies raised with TAB4 were able to recognize other diver- gent V3 peptides in ELISA. Ten peptides were studied, each representing the consensus sequence of a different group, according to the phenetic classification of the entire V3 region. The results in Figure 5 showed that sera of rabbits from group A reacted with nine of the ten peptides. Six peptides (nos. 1, 3, 4, 5, 9, 10) were strongly recognized (titer > 104), whereas titers against the rest were at least ten times lower (nos. 2, 7, and 8), or undetectable (no. 6).
DISCUSSION
In a previous paper*‘, the authors demonstrated that MEP TABl, which contains three central V3 regions from the related HIV-l isolates MN, SC and WMJ-2, was able to induce neutralizing mAbs against HIV-l(MN). In the present work, the spectrum of viral strains was expanded by the inclusion of more diver- gent V3 sequences in a single polypeptide. The selec- tion of those V3 sequences was done by analysis of the 245 DNA sequences reported in La Rosa et ~1.~“. They were grouped by amino acid homology following the criteria that two isolates would group together if they share a continuous sequence of seven or more amino acids. This presumes that such two isolates will probably share at least one common epitope. The central part of the V3 loop was considered because this is the region where the most important neutralizing epitopes are located. Six representative isolates were selected, whose corresponding groups represent 75.2% of the total number of sequences: MN (61.2%) LR150 (5.2%), BIB (4.4%) BRVA (3.2%), JYl (0.8%) and
RF (0.4%). Each of the remaining groups represented < 1% of the total.
According to the present classification of HIV-l isolates based on the gp120 DNA sequence4’, one isolate belongs to subgroup D (JYl) and the others to subgroup B, which is the most prevalent in Europe and North America. The MEP TAB4, composed of these epitopes, must be able to generate a broader neutral- izing antibody response than its predecessor TABl. Other differences between these proteins are:
(1)
(2)
the size of the N-terminal region from human IL-2 was reduced to only 26 amino acids in TAB4 (versus 62 in TABl); and the C155-65 region present in TAB1 was not included in TAB4.
TAB4 was produced in E. coli and its immuno- genic@ was evaluated in rabbits using two adjuvants. Higher mean antibody titers and better frequencies of response to individual epitopes were observed when TAB4 was administered in FCA than in aluminum hydroxide. Two doses of the immunogen were also studied. The mean antibody titer and the frequency of response after immunization with 50 pg (group B) were lower than with 200 pg (group A). The two rabbits from group A developed antibodies directed against all peptides included in TAB4.
It is known that V3 peptides used for immunization usually induce a neutralizing type-specific anti-HIV-l response; nevertheless, some degree of cross-neutral- ization between isolates with homologous V3 loop has been described. This cross-neutralization, supported by a close V3 loop-tip sequence similarity between the neutralized isolates, has been referred to as a ‘broadly neutralizing response’. Most of the viral strains included in these works belong to HIV-1 subgroup B and share the sequence GPGRAF28,“‘,42a.
However, several laboratories have shown the feasi- bility of inducing a truly broad anti-V3 loop antibody response by usin a cocktail of synthetic peptides from different isolates !Zl . A similar strategy with 15 V3 multi-
1 E+05
1 E+04
1 E+03
1 E+02
lE+Ol
q ic3 GPGR UC4
n C6 GPGQ WC7
IC6 GQGQ
TABC TABC RI R2
Sera
Figure 5 ELISA cross-reactivity against V3 consensus peptides (Cl-ClO) of anti-TAB4 sera from group A. The four letters represent the central tetrapeptide of the V3 present in the different groups
Vaccine 1997 Volume 15 Number 11 1205

Reactive antibodies against gp120 neutralize HIV-l: C.A. Duarte et al.
antigenic peptides (MAPS) from six clades of HIV-l has been evaluated in phase I clinical trials in humans4s. In this paper, it was demonstrated that a similarly broad response could be obtained using a MEP format. Antibody titers above 10’ were elicited against isolates as divergent as JYl (RQSTPIGLGQL- LYTT) and MN (RKRIHIGPGRAFYTT).
It is worth noting that although isolates LR1.50, IIIB and MN share the loop-tip sequence GPGRA, the titers against them differed markedly, by a factor of ten between MN and IIIB (GPGRAF in common). Javaherian et a[.“‘, reported a high degree of cross- reactivity between antibodies elicited in guinea pigs with V3 peptides from HIV-l MN and IIIB. They demonstrated not only ELISA cross-reactivity but also cross-neutralization, and inhibition of neutralization between these two isolates. The fraction of antibodies directed against the common sequence GPGRAF was the major one and responsible for these effects. In addition, Haynes et a1.46 were able to absorb the neutralizing activity in the sera of immunized animals, using a multimeric peptide of the GPGRAF sequence.
Although it seemed reasonable to expect the same results with anti-TAB4 sera, this was not the case. It was investigated whether the difference in recognition between MN and IIIB could be associated with a different degree of accessibility of the V3 epitopes in the surface of the protein bound to the plate. by their ELISA reactivity with mAbs 2C4 and P4D10, specific for the MN and IIIB V3 epitopes, respectively. However, the pattern of recognition with the two mAbs was very similar, which indicates that both epitopes are similarly exposed in TAB4. The differences found in immunogenicity between these two isolates should be then attributed to their intrinsic immunodominance. and suggest that other residues besides the GPGRAF region, which are not present in IIIB, are involved in the recognition of the MN peptide. Further experi- ments involving a larger number of animals and several species should be conducted to understand this phenomenon. Other authors have shown that antibodies against IIIB-V3 peptides do not necessarily cross-react with MN-V347.
To further characterize the nature of the antibody response against TAB4, fine mapping of the anti-MN response was performed in anti-TAB4 sera, using a set of alanine substituted peptides from the MN sequence. The results of this experiment demonstrate that although the GPGRAF region is important for antibody binding, the substitution of other residues like tyrosine could also abolish 80-90% of the binding, which indicates that the amino acids in these positions are also involved in the formation of reactive epitopes. The important contribution of tyrosine in the binding of the majority of the antibodies elicited against MN, and its substitution by threonine could explain why most anti-MN antibodies did not react with IIIB peptide.
To assess the range of cross-reactivity of anti-TAB4 antibodies, the binding of sera from group A against ten V3 consensus peptides was evaluated according to the phenetic classification of the V3 variability. High antibody titers against peptides exhibiting the central motif GPGR and GLGQ were found. This was coinci- dent with the presence of these two motifs in the V3
1206 Vaccine 1997 Volume 15 Number 11
regions included in TAB4. A lesser degree of reactivity was found for the rest of the peptides displaying the central tetrapeptides GPGQ or GQGQ, that were not included in TAB4. The results strongly suggest that sharing the central tetrapeptide in the V3 loop is critical for the binding of cross-reactive antibodies. In this case, unlike IIIB, consensus peptides Cl, C3, C4 and C.5 shared the sequence GPGRAFY with the MN. Taken together with previous results of fine mapping, it can be concluded that the central tetrapeptide is neces- sary but not sufficient to elicit cross-reacting antibodies.
Other authors have associated the ability to neutralize MN and LA1 isolates by several anti-V3 polyclonal antibodies raised against gp120 or V3
pcpt ides3 I .32.JX.JY, However, Excler et a1.5” and Pialoux et
al:’ reported a lack of MN-LA1 cross-neutralizing antibodies in human volunteers immunized with gp160 MNILAI. No cross-neutralization of HIV-l isolates from different clades by anti-V3 antibodies has been reported to date.
Anti-TAB4 sera from groups A and B were able to neutralize the MN isolate and one serum also neutral- ized the LA1 isolate. The anti-TAB4 titer correlated with the MN neutralization titer. In contrast, the LA1 neutralization correlated neither with anti-TAB4 nor with anti-IIIB titers. The predictive value of peptide binding assays is a subject of intense debate. As reviewed by Golding et al.“, while some authors have described a positive correlation between ELISA binding and neutralization titers in sera from either HIV-l infected patients or animals immunized with gp120 or synthetic peptides, others have failed to demonstrate this correspondence. The variability observed in both peptide binding and HIV neutral- ization assays among different laboratories may be the cause of such discrepancies.
Neutralization of the other isolates included in TAB4 could not be tested. However, sera from all rabbits neutralized four of the ten clinical isolates (Table 2). The neutralization of HIV-l field isolates is an important point to consider, since it has been reported that primary isolates seem to be more resistant to in vitro neutralization than laboratory strain?‘+. Our experiments were performed with Cuban isolates that were passed once through MT4 cells before the assay, so they cannot be defined as primary isolates. Characterization of the V3 sequence of these isolates is in progress but since they were also neutralized by mAb 2C4, which displays narrow specifi- city for the IHIGPXR epitopes5, they should be homol- ogous to the MN isolate. Additionally, in another set of experiments, the authors have found that rabbit anti- TAB4 sera is also able to neutralize the African primary isolate UG9246 previously adapted in MT4 cells (unpublished results). The reported sequence for the central part of the V3 loop of this isolate is identical to JYl, although the MT4 adapted strain suffered five amino acid changes in the relevant region.
These results warrant further evaluation of the MEP format as an AIDS vaccine candidate. MEPs, like synthetic peptides, are a flexible approach allowing the introduction of neutralizing targets from different clades to induce a broad humoral immune response or the addition of any relevant epitope from other viral

Reactive antibodies against gp720 neutralize HIV-7: C.A. Duarte et al.
proteins. In contrast to synthetic peptides, MEPs can be made highly immunogenic per se, voiding the use of carriers for immunizations. A further advantage is that the synthetic gene can be cloned in live vectors such as poxviruses or mycobacteria to induce a strong cytotoxic response against virus infected cells. Alternatively, it can be used for naked DNA immunization, a procedure that has been demonstrated to be simple and potent to elicit both humoral and cell mediated immunity.
ACKNOWLEDGEMENTS
The authors would like to thank Dr Harvey Holmes from the MRC AIDS Reagent Program for providing the viral isolates used in this work.
REFERENCES
1
2
3
4
5
6
7
0
9
10
11
Chiou, S.H., Freed, E.O., Paganiban, A.T. and Keneally, W.R. Studies on the role of the V3 region in Human Immunodefi- ciency virus type 1 envelope glycoprotein function. AIDS Research on Human Retroviruses 1992,8, 161 l-l 617. Ivanoff, L., Looney, D.J., McDanal, C., Morris, J.F., Wong- Staal, F., Langlois, A.J., Petteway, S.R. Jr. and Matthews, T.J. Alteration of HIV-l infectivity and neutralization by a single amino acid replacement in the V3 loop domain. AfDS Research on Human Retroviruses 1991, 7, 595-603. Shioda, T., Levy, J.A. and Chen-Mayer, C. Macrophage and T-cell line tropisms of HIV-1 are determined by specific regions of the envelope gpl20 gene. Nature 1991, 349, 167-169. Palker, T.J., Clark, M., Langloise, A.J., Mathews, T.J., Weinhold, K.J., Randall, RR., Bolognesi, D. and Haynes, B.F. Type-specific neutralization of the human immunodeficiency virus with antibodies to env-encoded synthetic peptides. Proceedings of the National Academy of Sciences of the United States of America 1988, 85, 1932-l 936. Rushe, J.R., Jahaverian, K., McDanal, C., Petro, J., Lynn, D.L., Grimaila, R., Langlois, A., Gallo, R.C., Arthur, L.O., Fischinger, P.J., Bolognesi, D.P., Putney, SD. and Mathews, T.J. Antibodies that inhibit fusion of human immunodeficiency virus-infected cells bind a 24-aminoacid sequence of viral envelope gp120. Proceedings of the National Academy of Sciences of the United States of America 1988, 85, 3192-3202. Goudsmit, J., Boucher, CA.. Meloen, R.H.. Epstein, L.G., Smit, L., van der Hoek, L. and Bakker, M. Human antibody response to a strain-specific HIV-l gpl20 epitope associated with cell fusion inhibition. A/LX 1988, 2, 157-164. Kennealy, W.R., Matthews, T.J., Ganfield, M.C., Langlois, A.J., Waselefsky, D.M. and Petteway, S.R. Jr Antibodies from Human lmmunodeficiency Virus-infected individuals bind to a short amino acid’sequence that elicits neutralizing antibodies in animals. AlDS Research on Human Retroviruses 1989, 5, 173-182. Jahaverian, K., Langlois, A.J., McDanal, C., Ross, K.L., Eckler, L.I., Cindy, L., Jellis, L., Profy, A.T., Rusche, J.R., Bolognesi, D.P., Putney. S.D. and Mathews, T.J. Principal neutralizing domain of the human immunodeficiency virus type 1 envelope protein. Proceedings of the National Academy of Sciences of the United States of America 1989, 86, 6768-6772. Putney, S., Javaherian, K., Profy, A., Emini, E., Langlois, A., Ross, K., et a/. In Human Retroviruses. Alan R. Liss, 1990, pp. 323-335. Takahashi, H., Cohen, J., Hosmalin, A., Cease, K.B., Houghten, R., Cornette, J.L., et a/. An immunodominant epitope of the human immunodeficiency virus envelope glycoprotein gpl60 recognized by class I major histocom- patibility complex molecule-restricted immune cytotoxic T lymphocytes. Proceedings of the National Academy of Sciences of the United States of America 1988, 85, 3105. Lasarte, J.J., Sarobe, P., Gullon, A., Prieto. J. and Borras- Cuesta, F. Induction of cytotoxic T lymphocytes in mice
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
against the principal neutralizing domain of HIV-l by immuniz- ation with an engineered T-cytotoxic-T-helper synthetic peptide construct. Cellular Immunology 1992, 141, 21 l-21 8. Takahashi, H., Nakagawa, Y., Pendleton, C., Hougten, R., Yokomuro, K., Germaine, R.N. and Berzofsky, J.A. Induction of broadly cross-reactive T cells recognizing an HIV-l envelope determinant. Science 1992, 255, 333-336. Moore, R.L. and Fox, B.S. Immunization of mice with human immunodeficiency virus glycoprotein gpl60 peptide 315-329 induce both class I and class II restricted T-cells: Not all cells can respond to whole molecule stimulation. AfDS Research on Human Retroviruses 1993, 9, 51-59. Takahashi, H., Germaine, R.N., Moss, B. and Berzofsky, J.A. An immunodominant class l-restricted cytotoxic T lympho- cytes determinant on human immunodeficiency virus type 1 induces CD 4 class II-restricted help for itself. Journal of Experimental Medicine 1990, 171, 571-576. Billaud, J.N., Yagello, M. and Gluckman, J.C. Primary in vitro sensitization of human T-helper lymphocytes by peptides derived from the V3 loop of human immunodeficiency virus type 1 (HIV-l) envelope glycoprotein. Vaccine 1994, 12, 46-55. Safrit, J.T., Fung, M.S.C., Andrews, CA., Braun, D.G., Sun, W.N.C., Chang, T.W. and Koup, R.A. hu-PBL-SCID mice can be protected from HIV-l infection by passive transfer of monoclonal antibody to the principal neutralizing determinant of envelope gpl20. A/LX 1993, 7, 15-21. Emini, E., Schleif, W.A., Numberg, J.H., Conley, A.J., Eda, Y., Tokiyoshi, S., Putney, SD., Matsushita, S., Cobb, K.E., Jett, C., Eichberg, J. and Murthy, K. Prevention of HIV-1 infection in chimpanzees by gp120 V3 domain specific monoclonal antibody. Nature 1992, 355, 728-730. Emini, E., Nara, P., Schleif, W., Lewis, J., Davide, J., Lee, D., Kessler, J., Conley, S., Matsusshita, S., Putney, S., Gerety, R. and Eichberg, J. Antibody mediated in vitro neutralization of human immunodeficiency virus type 1 abolishes infectivity for chimpanzees. Journal of Virology 1990, 64, 3674-3678. Matsushita, S., Robert-Guroff, M., Rusche, J., Koito, A., Hattori, T., Javaherian, K., Takatsuki, K. and Putney, S. Characterization of a Human lmmunodeficiency Virus neutral- izing monoclonal antibody and mapping of the neutralizing epitope. Journal of Virology 1988, 62, 2107-2114. Skinner, M., Ting, R., Langlois, A., Weinhold, K., Lyerly, H., Jahaverian, K. and Mathews, T.J. Characteristics of a neutral- izing monoclonal antibody to the HIV envelope glycoprotein. AIDS Research on Human Retroviruses 1989,4, 187- 197. Pal, R., di Marzo Veronese, F., Nair, B., Rahman, R., Hoke, G., Mumbauer, S. and Sarngadharan, M. Characterization of a neutralizing monoclonal antibody to the external glycoprotein of HIV-l. lntervirology 1992, 86, 86-93. Liou, R.S., Rosen, E., Fung, M., Sun, W., Sun, C., Gordon, W., Chang, N. and Chang, T. A chimeric mouse-human antibody that retains specificity for HIV gpl20 and mediates the lysis of HIV-infected cells. Journal of Immunology 1989, 143, 3967-3975. Duarte, C., Montero, M., Seralena, A., Valdes, R., Jimenez, V., Benitez, J., Narciandi, E., Madrazo, J., Padron, G., Sanchez, G., Girlund, G., Pearson, K., Dominguez, M., Wharen, B. and Menendez, A. Multi-epitope polypeptide containing epitopes of HIV-l envelope induces neutralizing monoclonal antibodies against V3 loop. AIDS Research on Human Retroviruses 1994, 19, 235. Scott, CF., Silver, S., Profy, A., Putney, S., Langlois, A., Weinhold, K. and Robinson, J. Human monoclonal antibody that recognized the V3 region of human immunodeficiency virus gpl20 and neutralizes the human T-lymphotropic virus type Ill MN strain. Proceedings of the National Academy of Sciences of the United States of America 1990, 8, 859668601. Durda, P., Bacheler, L., Clapham, P., Jenoski, A., Leece, B., Matthews, T., McKnight, A., Pomerantz, R., Rayner, M. and Weinhold, K. HIV-l neutralizing monoclonal antibodies induced by a synthetic peptide. AlDS Research on Human Retroviruses 1990, 6, 1115- 1213. Ohno, T., Terada, M., Yoneda, Y., Shea, K., Chambers, R., Stroka, D., Nakamura, M. and Kufe, D. A broadly neutralizing antibody that recognizes the V3 region of human immunodefi- ciency virus type 1 glycoprotein gpl20. Proceedings of the National Academy of Sciences of the United States of America 1991, 88,10726-l 0729.
Vaccine 1997 Volume 15 Number 11 1207

Reactive antibodies against gp720 neutralize HIV- 1: C.A. Duarte et al.
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
Buchbinder, A., Karwowska, S., Gorny, M., Burda, S. and Zolla-Pazner, S. Synergy between human monoclonal antibodies to HIV extends their effective biologic activity against homologous and divergent strains. AlDS Research on Human Retroviruses 1992, 8,425-427. Whitte-Scharf, M.E., Potts, B., Smith, L., Sokolowski, K., Rusche, J. and Silver, S. Broadly neutralizing monoclonal antibodies to the V3 region of HIV-l can be elicited by peptide immunization. Virology 1993, 192, 197-206. Palker, T., Matthews, T., Langloise, A., Tanner, M., Martin, M., Scearce, R., Kim, J., Berzofski, J., Bolognesi, D. and Haynes, B. Polyvalent human immunodeficiency virus synthetic immunogen comprised of envelope gp120 T helper cell sites and B cell neutralization epitopes. Journal of immunology 1989, 142,3612-3619. Neurath, Ft., Jiang, S., Strick, N., Kolbe, H., Kieny, M.P., Muchmore, E. and Girard, M. Antibody responses of chimpanzees immunized with synthetic peptides corre- sponding to full length V3 hypervariable loops of HIV-1 envelope glycoproteins. AIDS Research on Human Retro- viruses 1991, 7,ai3-823. Javaherian, K., Langlois, A., LaRosa, G., Profy, A., Bolognesi, D., Herlihy, W., Putney, S. and Mathews, T. Broadly neutral- izing antibodies elicited by the hypervariable neutralizing determinant of HIV-l. Science 1990, 250, 1590. Jimenez. V., Guimil, Ft., Ubieta, R., Ochagavia, M., Silva, A., Villegas, G. and Herrera, L. Sintesis quimico-enzimtitica del gen de la proinsulina humana. Biofecnologia Ap/icada 1990, 7,142-l 52. Sambrook, J., Fritsch, E.F. and Maniatis, T. Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989. Castellanos, L., Ubieta, R., Hardy, E., P6rez, G., Santiago, N., GonzBlez, M., Fernandez, C., Jimenez, V., Estrada, R., Silva, A. and Herrera, L. Expression of IL-2 proinsulin fusion protein. Folding of the fusion protein and conversion into insulin by one step removal of C-peptide and leader sequence. Protein Engineering 1993, 6, (S), 9. Sanger, F., Nicklen, S. and Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proceedings of the National Academy of Sciences of the United States of America 1977, 74, 5463-5467. Laemli, U.K. Cleavage of structural proteins during the assembly of the head of the bacteriophage T4. Nature 1970, 227,680-685. Houghten. R.A. General method for the rapid synthesis of large number of peptides. Specificity of antigen antibody interaction at the level of individual amino acids. Proceedings of the Nationai Academy of Sciences of the United States of America 1985, 82, 5131. Akerblom, L., Hinkula, J., Broliden, P., Mtikitalo, H., Fridbrger, T.. Rosen, J., Eriksson, M.V., Morein, 8. and Wahren, B. Neutralizing monoclonal antibodies to HIV-l gp120. AlDS 1990,4,953-960. Bradford, M.B. A rapid and sensitive method for the quantita- tion of microgram quantities of protein utilizing the principle of protein-dye binding. Annals of Biochemistry and Experimental Medicine 1976, 72, 248-254. La Rosa, G.J., Davide, J.P., Weinhold, K., Waterbury, J., Profy, A.T., Lewis, J.A., Langlois, A.J., Dreesman, G.R., Boswell, R.N., Shadduck, P., Halley, L.H., Karplus, M., Bolog- nesi. D.P.. Matthews, T.J.. Emini. E.A. and Putnev. S.D. Conserved sequence and structural elements in the HIV-l principal neutralizing determinant. Science 1990, 249, 932-935. Myers, G., Korber, B., Wain-Hobson, S., Smith, R. and Pavlakis, G.N. (Eds) Human Retroviruses and AIDS. A Compil- ation and Analysis of Nucleic Acid and Amino Acid Sequences. Theoretical Biology and Biophysics Group T-l 0, Los Alamos National Laboratory, Los Alamos, NM, 1994. Putney, S., LaRosa, G., Profy, A., Silver, S., Scott, C., Javaherian, K., et a/. HIV-1 principal neutralization determinant elicits broadly neutralizing antibodies. In Vaccines ‘97, eds
43
44
45
46
47
48
49
50
51
52
53
54
55
Brown, F., Chanock, R.M. and Lerner, R.A. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1991. Lake, F., Kawamura, T.. Tomiyama, T., Robinson, W.E., Matsumoto, Y., Masuno, Y. and Hersh, E. Generation and characterization of a human monoclonal antibody that neutral- izes diverse HIV-l isolates in vitro. AfDS 1992, 6, 17-24. Gorny, M., Conley, A., Karwoska, S., Buchbinder, A., Xu, J., Emini, E., Koening, S. and Zolla-Pazner, S. Neutralization of diverse human immunodeficiency virus type 1 variants by an anti-V3 human monoclonal antibody. Journal of Virology 1992, 66,7538-7542. O’Hagan, D.T., McGee, J.P., Wang, C.Y., et al. The UBI multi- component HIVR vaccine: the advantages of controlled release microparticles. In Retroviruses of Human AlDS and Related Animal Diseases. 8th Colloque des ‘Cent Gardes’, 25-27 October 1993. eds Girard. M. and Valeette. L. Founda- tion Marcel Merieuxl Marnes-La-Coquette, Paris, 1994, pp. 309-313. Haynes, B., Torres, J.V., Langlois, A.J., Bolognesi, D.P., Gardner, M.B., Palker, T.J., Scearce, R.M., Jones, D.M., Moody, M.A., McDanal, C. and Matthews, T.J. Induction of HIVMN neutralizing antibodies in primates using a prime- boost regimen of hybrid synthetic gp120 envelope peptides. Journal of Immunology 1993, 151,1646- 1653. Warren, R., Wolf, H., Stunz, G. and Kennedy, R. Antibody reactivity to synthetic peptides representing the principal neutralizing determinant of HIV-l in mouse strain following repeated immunization with recombinant gpl60. Experimental and Clinical Immunogenetics 1993, 10, 168 175. Laman, J., Schellekens, M., Abacioglu, Y., Lewis, G., Tersm- ette, M., Fouchier, R., Langedijk, J., Claassen, E. and Boersma, W. Variant-specific monoclonal and group-specific polyclonal human immunodeficiency virus type 1 neutralizing antibodies raised with synthetic peptides from the gp120 third variable domain. Journal of Virology 1992, 66, 1823-l 831. Belshe, R.B., Graham, M.S. and Keefer, M.C. et a/. Neutral- izing antibodies to HIV-1 in seronegative volunteers immunized with recombinant gp120 from the MN strain of HIV-l. Journal of the American Medical Association 1994, 272, 475-480. Excler, J., Salmon, D., Pialoux, G., et a/. Combined antigen regimes for vaccination against HIV-l. In Retroviruses of Human AIDS and Related Animal Diseases. 8th Colloque des ‘Cent Gardes’, 25-27 October 7993, eds Girard, M. and Valeette, L. Foundation Marcel Merieux, Marnes-La-Coquette, Paris, 1994, pp. 281-284. Pialoux, G., Excler, J.L., Riviere, Y., Gonzalez-Canali. G., Feuillie, V., Coulaud, P., Gluckman, J., Matthews, T., Meignier, B., Kreny, M.P., Gonnet, P., Diaz, I., Merit, C., Paoletti, E., Tartaglia, J.. Salomon, H. and Plotkin, S. The Agis Group and L’Agence Nationale de Recherche sur le SIDA. A prime boost approach to HIV preventive vaccine using a recombinant canarypox virus expressing gp160 (MN) followed by a recombinant glycoprotein 160 (MN/LAI). AlDS Research on Human Retroviruses 1995, 11, 373-381. Golding, H., D’Souza, P., Bradac, J., Mathieson, B. and Fast, P. Neutralization of HIV-l. AlDS Research on Human Retro- viruses 1994, 10, (6). 633-643. Francis, D., Fast, P., Harkonen, S., McElrath, M., Belshe, R., Berman, P., Gregory, T. and the AIDS Vaccine Evaluation Group. MN rgp120 (Genentech) vaccine safe and immuno- genic-But will it protect humans? In 10th international Conference on AIDS, Yokohama, Japan. Abstract No. 314A, Vol. 1, 1994, p. 90. Graham, B.S. Serological responses to candidate AIDS vaccines. AlDS Research on Human Retroviruses 1994. 10. (2) si45-si48.
,
Duarte, C.A., Perez, L., Vazquez, J., Duerias, M., Vilarrubia, O.L.. Navea. L.. Valdes, R.. Reves, O., Avala, M. and Gavilondo, J: Epitope mapping, V-region DNA sequence, and neutralizing Fab fragments of two monoclonal antibodies against the HIV-1 V3 loop. lmmunofechnology 1996, 2, 11-20.
1208 Vaccine 1997 Volume 15 Number 11
Copyright © 2022 FDOKUMEN




















