
N-Methyl-d-aspartate receptors mediating hippocampal noradrenaline and striatal dopamine release...
10
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 139–148 139 Journal of Neurochemistry, 2001, 76, 139–148 N-Methyl-d-aspartate receptors mediating hippocampal noradrenaline and striatal dopamine release display differential sensitivity to quinolinic acid, the HIV-1 envelope protein gp120, external pH and protein kinase C inhibition Anna Pittaluga, Roberto Pattarini, 1 Marco Feligioni and Maurizio Raiteri Dipartimento di Medicina Sperimentale, Sezione di Farmacologia e Tossicologia, Universita ` di Genova, Italy Abstract NMDA receptors regulating hippocampal noradrenaline (NA) and striatal dopamine (DA) release have been compared using superfused synaptosomes prelabelled with the [ 3 H]cate- cholamines. Both receptors mediated release augmentation when exposed to NMDA plus glycine. Quinolinic acid (100 mM or 1 mM) plus glycine (1 mM)-elicited [ 3 H]NA, but not [ 3 H]DA release. The NMDA (100 mM)-evoked release of [ 3 H]NA and [ 3 H]DA was similar and concentration-dependently enhanced by glycine or D-serine (0.1–1 mM); in contrast, the HIV-1 envel- ope protein gp120 potently (30–100 pM) enhanced the NMDA- evoked release of [ 3 H]NA, but not that of [ 3 H]DA. Gp120 also potentiated quinolinate-evoked [ 3 H]NA release. Ifenprodil (0.1– 0.5 mM) or CP-101,606 (0.1–10 mM) inhibited the NMDA plus glycine-evoked release of both [ 3 H]catecholamines. Zinc (0.1–1 mM) was ineffective. Lowering external pH from 7.4 to 6.6 strongly inhibited the release of [ 3 H]NA elicited by NMDA plus glycine, whereas the release of [ 3 H]DA was unaffected. The protein kinase C inhibitors GF 109203X (0.1 mM) or H7 (10 mM) selectively prevented the effect of NMDA plus glycine on the release of [ 3 H]NA. GF 109203X also blocked the release of [ 3 H]NA induced by NMDA or quinolinate plus gp120. It is concluded that the hippocampal NMDA receptor and the striatal NMDA receptor are pharmacologically distinct native subtypes, possibly containing NR2B subunits but different splice variants of the NR1 subunit. Keywords: gp120, NMDA receptor subtypes, NR1 subunit variants, pH, protein kinase C, quinolinic acid. J. Neurochem. (2001) 76, 139–148. A number of neurological and neuropsychiatric symptoms, including cognitive deficits, are present in a high percentage of AIDS patients. In some cases these symptoms are likely to reflect overt neurodegenerative events; in others they may be a consequence of more subtle effects caused by exogenous compounds originated from the HIV-1 virus or by endogenous molecules whose concentration becomes abnormally elevated (Giulian et al. 1996; Lipton 1998; Schroeder et al. 1998). The HIV-1 glycoprotein gp120 is an envelope protein that is shed by the virus and appears to exert a number of deleterious actions, some probably leading to neuronal death. The coat protein is thought to act in concert with agonists at glutamate receptors of the NMDA type, including glutamate itself, aspartate and quinolinic acid. Quinolinate is a product of tryptophan catabolism, usually present in the extracellular fluid of the mammalian CNS at relatively low concentrations (Stone 1993 and references therein), but the levels of which are reported to increase greatly during pathological conditions, including AIDS (Achim et al. 1993; Martin et al. 1993; Beagles et al. 1998). In previous works we found that gp120 can imitate glycine or d-serine as a glutamate coagonist at NMDA Received May 24, 2000; revised manuscript received August 3, 2000; accepted August 11, 2000. Address correspondence and reprint requests to Dr Anna Pittaluga, Dipartimento di Medicina Sperimentale, Sezione di Farmacologia e Tossicologia, Viale Cembrano 4, 16148 Genova, Italy. E-mail: [email protected] Abbreviations used: CP-101,606, (1S,2S)-1-(4-hydroxyphenyl)-2-(4- hydroxy-4-phenylpiperidino)-1-propanol; DA, dopamine; GF 109203X, dihydrochloride-3-{1-[3-(dimethylamino)propyl]-1H-indol-3-yl}-4-(1H- indol-3-yl)-1H-pyrrole-2,5-dione; H7, 1-(5-isoquinolinylsulphonyl)-2- methylpiperazine; NA, noradrenaline; PKC, protein kinase C. 1 The present address of Dr Roberto Pattarini is Department of Pharmacology, University of Manitoba, Faculty of Medicine, 753 McDermot Avenue, Winnipeg, MB R3E OT6, Canada.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of N-Methyl-d-aspartate receptors mediating hippocampal noradrenaline and striatal dopamine release...

q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 139±148 139
Journal of Neurochemistry, 2001, 76, 139±148
N-Methyl-d-aspartate receptors mediating hippocampal
noradrenaline and striatal dopamine release display differential
sensitivity to quinolinic acid, the HIV-1 envelope protein gp120,
external pH and protein kinase C inhibition
Anna Pittaluga, Roberto Pattarini,1 Marco Feligioni and Maurizio Raiteri
Dipartimento di Medicina Sperimentale, Sezione di Farmacologia e Tossicologia, UniversitaÁ di Genova, Italy
Abstract
NMDA receptors regulating hippocampal noradrenaline (NA)
and striatal dopamine (DA) release have been compared
using superfused synaptosomes prelabelled with the [3H]cate-
cholamines. Both receptors mediated release augmentation
when exposed to NMDA plus glycine. Quinolinic acid (100 mM
or 1 mM) plus glycine (1 mM)-elicited [3H]NA, but not [3H]DA
release. The NMDA (100 mM)-evoked release of [3H]NA and
[3H]DA was similar and concentration-dependently enhanced
by glycine or D-serine (0.1±1 mM); in contrast, the HIV-1 envel-
ope protein gp120 potently (30±100 pM) enhanced the NMDA-
evoked release of [3H]NA, but not that of [3H]DA. Gp120 also
potentiated quinolinate-evoked [3H]NA release. Ifenprodil (0.1±
0.5 mM) or CP-101,606 (0.1±10 mM) inhibited the NMDA plus
glycine-evoked release of both [3H]catecholamines. Zinc
(0.1±1 mM) was ineffective. Lowering external pH from 7.4 to
6.6 strongly inhibited the release of [3H]NA elicited by NMDA
plus glycine, whereas the release of [3H]DA was unaffected.
The protein kinase C inhibitors GF 109203X (0.1 mM) or H7
(10 mM) selectively prevented the effect of NMDA plus glycine
on the release of [3H]NA. GF 109203X also blocked the
release of [3H]NA induced by NMDA or quinolinate plus
gp120. It is concluded that the hippocampal NMDA receptor
and the striatal NMDA receptor are pharmacologically distinct
native subtypes, possibly containing NR2B subunits but
different splice variants of the NR1 subunit.
Keywords: gp120, NMDA receptor subtypes, NR1 subunit
variants, pH, protein kinase C, quinolinic acid.
J. Neurochem. (2001) 76, 139±148.
A number of neurological and neuropsychiatric symptoms,
including cognitive de®cits, are present in a high percentage
of AIDS patients. In some cases these symptoms are likely
to re¯ect overt neurodegenerative events; in others they
may be a consequence of more subtle effects caused by
exogenous compounds originated from the HIV-1 virus or
by endogenous molecules whose concentration becomes
abnormally elevated (Giulian et al. 1996; Lipton 1998;
Schroeder et al. 1998).
The HIV-1 glycoprotein gp120 is an envelope protein that
is shed by the virus and appears to exert a number of
deleterious actions, some probably leading to neuronal
death. The coat protein is thought to act in concert with
agonists at glutamate receptors of the NMDA type,
including glutamate itself, aspartate and quinolinic acid.
Quinolinate is a product of tryptophan catabolism, usually
present in the extracellular ¯uid of the mammalian CNS at
relatively low concentrations (Stone 1993 and references
therein), but the levels of which are reported to increase
greatly during pathological conditions, including AIDS
(Achim et al. 1993; Martin et al. 1993; Beagles et al. 1998).
In previous works we found that gp120 can imitate
glycine or d-serine as a glutamate coagonist at NMDA
Received May 24, 2000; revised manuscript received August 3, 2000;
accepted August 11, 2000.
Address correspondence and reprint requests to Dr Anna Pittaluga,
Dipartimento di Medicina Sperimentale, Sezione di Farmacologia
e Tossicologia, Viale Cembrano 4, 16148 Genova, Italy.
E-mail: [email protected]
Abbreviations used: CP-101,606, (1S,2S)-1-(4-hydroxyphenyl)-2-(4-
hydroxy-4-phenylpiperidino)-1-propanol; DA, dopamine; GF 109203X,
dihydrochloride-3-{1-[3-(dimethylamino)propyl]-1H-indol-3-yl}-4-(1H-
indol-3-yl)-1H-pyrrole-2,5-dione; H7, 1-(5-isoquinolinylsulphonyl)-2-
methylpiperazine; NA, noradrenaline; PKC, protein kinase C.1The present address of Dr Roberto Pattarini is Department of
Pharmacology, University of Manitoba, Faculty of Medicine, 753
McDermot Avenue, Winnipeg, MB R3E OT6, Canada.

receptors localized on noradrenergic neurones in the hippo-
campus and neocortex of rats (Pittaluga and Raiteri 1994;
Pattarini et al. 1998) and humans (Pittaluga et al. 1996).
These NMDA receptors are known to mediate the increase
of noradrenaline (NA) release (Fink et al. 1990; Pittaluga
and Raiteri 1990), and gp120, as well as its V3 loop peptide,
can mimic glycine with impressive potency, i.e. four orders
of magnitude higher than that of the natural glutamate
coagonist (Pattarini et al. 1998).
Quinolinic acid is an NMDA receptor agonist (Stone and
Perkins 1981; Stone 1993; Moroni 1999). The af®nity of
quinolinate for the receptor is much weaker than that of
NMDA; however, as an excitotoxin in the adult rat brain in
vivo, quinolinic acid was found to be as potent as NMDA
(Foster and Schwarcz 1988). Of note, quinolinate-induced
lesions differ qualitatively from those produced by NMDA
(see Guidetti and Schwarcz 1999 for references). One
possible reason is that quinolinic acid preferentially acti-
vates subtypes of the NMDA receptor (Monaghan and
Beaton 1991; Prado de Carvalho et al. 1996).
Release-regulating receptors of the NMDA type are
located not only on noradrenergic axon terminals in the
hippocampus, but also on striatal dopaminergic axon
terminals where they mediate enhancement of dopamine
(DA) release (Roberts and Anderson 1979; Krebs et al.
1991). The two presynaptic NMDA receptors regulating
catecholamine release have not been compared in terms of
functional properties and structural characteristics. In the
present work we studied the release of NA from rat hippo-
campal synaptosomes and of DA from striatal synaptosomes
and compared the effects of agonists at the glutamate
recognition site (including quinolinic acid) and at the
glycine site (including gp120). We found differences
between the two NMDA receptors and tried to clarify the
reasons for these differences by investigating the NR2
subunits present and the possible involvement of splice
variants of the NR1 subunit.
Materials and methods
Drugs
[3H]NA (speci®c activity 30 Ci/mmol) and [3H]DA (speci®c
activity 43 Ci/mmol) were purchased from Amersham Radio-
chemical Centre (Buckinghamshire, UK). Glycine, l-aspartate,
quinolinic acid, 1-(5-isoquinolinylsulphonyl)-2-methylpiperazine
(H7) and d-serine were obtained from Sigma Chemical Co.
(St Louis, MO, USA); NMDA was from Tocris Cookson (Bristol,
UK). Recombinant HIV-1 IIIB glycoprotein gp120 was obtained
from AMS Raggio Italgene (Milan, Italy). Sphingosine and
dihydrochloride-3-{1-[3-(dimethylamino)propyl]-1H-indol-3-yl}-4-
(1H-indol-3-yl)-1H-pyrrole-2,5-dione (GF 109203X) were pur-
chased from RBI (Sigma Aldrich, Milan, Italy). The following
substances were generous gifts from the indicated companies:
6-nitroquipazine maleate (Duphar, Amsterdam, the Netherlands)
(1S,2S)-1-(4-hydroxyphenyl)-2-(4-hydroxy-4-phenylpiperidino)-1-
propanol (CP-101,606; P®zer, Groton, CT, USA), memantine
(Merz Co., Frankfurt, Germany) and ifenprodil (Synthelabo
Recherche, Bagneaux, France).
Animals
The experimental procedures were approved by the Ethical
Committee of the Pharmacology and Toxicology Section, Depart-
ment of Experimental Medicine, in accordance with the European
legislation (European Communities Council Directive of 24
November 1986, 86/609/EEC). Adult male rats (Sprague±Dawley;
200±250 g) were housed at constant temperature (22 ^ 18C) and
relative humidity (50%) under a regular light±dark schedule (lights
on 7.00 a.m. to 7.00 p.m.). Food and water were freely available.
The animals were killed by decapitation, and hippocampi and
striata were rapidly dissected.
Preparation of synaptosomes
Crude synaptosomes were prepared as previously described (Raiteri
et al. 1984). Brie¯y, the tissue was homogenized in 40 volumes of
0.32 m sucrose buffered at pH 7.4 with phosphate (®nal concentra-
tion 0.04 m). The homogenate was centrifuged (5 min, 1000 g) to
remove nuclei and cellular debris, and synaptosomes were isolated
from the supernatant by centrifugation (12 000 g for 20 min). The
synaptosomal pellet was then resuspended in a physiological
medium having the following composition (mm): NaCl, 125; KCl,
3; MgSO4, 1.2; CaCl2, 1.2; NaH2PO4, 1; NaHCO3, 22; glucose, 10
(aeration with 95% O2 and 5% CO2 at 378C); pH 7.2±7.4.
Release experiments
Hippocampal synaptosomes were labelled with [3H]NA (®nal
concentration: 30 nm) in the presence of 0.1 mm 6-nitroquipazine to
avoid false labelling of serotonergic terminals. Striatal synapto-
somes were labelled with [3H]DA (®nal concentration: 23 nm).
Both preparations were incubated with the radioactive label at
378C, for 15 min, in a rotary water bath and in an atmosphere of
95% O2 and 5% CO2.
After the labelling period, identical portions of the synaptosomal
suspension were introduced in parallel superfusion chambers
maintained at 378C (Raiteri et al. 1974; Raiteri and Raiteri 2000).
Superfusion was then carried out at a rate of 0.5 mL/min, using
media aerated with 95% O2 and 5% CO2, for a total period of
48 min. Starting at t � 36 min, four 3-min fractions were collected.
At t � 20 min the medium was replaced with a physiological
solution from which Mg21 ions were omitted. Agonists were added
at t � 39 min, corresponding to the end of the ®rst fraction
collected, and were present till the end of superfusion. Antagonists
were added 8 min before agonists and maintained throughout the
superfusion period. Protein kinase C (PKC) inhibitors were added
at t � 39 min, together with agonists and were present till the end
of the superfusion.
In a set of experiments, performed to evaluate the effect of pH on
the evoked neurotransmitter release, the physiological medium
containing bicarbonate was replaced at t � 20 min by a medium
having the following composition (mm): NaCl, 140; KCl, 3; CaCl2,
1.2; NaH2PO4, 1; NaHCO3, 5; glucose, 10; HEPES, 10. The pH of
this medium was adjusted to the desired value (pH 6.6, pH 7.4 and
pH 8.0) with NaOH.
140 A. Pittaluga et al.
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 139±148

Fractions collected and solubilized superfused synaptosomes
were counted for radioactivity by liquid scintillation.
Calculation and statistics
The amounts of [3H]NA and [3H]DA released into each superfusate
fraction were expressed as percentages of the total synaptosomal
content at the start of the fraction collected. The effect of the drugs
was evaluated by calculating the ratio between the percent ef¯ux in
the fraction corresponding to the maximal effect (the third fraction)
and the ef¯ux in the ®rst fraction collected. This ratio was
compared to the corresponding ratio obtained under control
conditions. When studying the effect of NMDA antagonists, results
are expressed as percentages of the NMDA plus glycine-evoked
release. A two-tailed Student's t-test was used to analyse the
signi®cance of the difference between two means; multiple
comparisons were made by applying the Dunnett's test.
Results
Effects of NMDA, l-aspartate and quinolinic acid on
the release of [3H]NA from hippocampal synaptosomes
and of [3H]DA from striatal synaptosomes
Rat hippocampal or striatal synaptosomes were prelabelled
with [3H]NA or [3H]DA, respectively, and exposed during
superfusion to NMDA, l-aspartate or quinolinic acid alone
or in the presence of glycine. l-Aspartate was used because
previous results showed that this endogenous aminoacid,
differently from l-glutamate, is a selective agonist at NMDA
receptors (Roberts and Anderson 1979). The results with
NMDA con®rm previous data obtained with cortical (Fink
et al. 1990) or hippocampal (Pittaluga and Raiteri 1990)
synaptosomes and with striatal synaptosomes (Krebs et al.
1991) showing that the release of [3H]NA and [3H]DA could
be enhanced by NMDA alone, possibly due to the presence
of glycine contamination in the solutions (see Paudice et al.
1998 for details) and, more effectively, by NMDA plus
glycine (Table 1). l-Aspartate (100 mm) behaved very
similarly to 100 mm NMDA. Quinolinic acid signi®cantly
enhanced the release of tritium from hippocampal synapto-
somes labelled with [3H]NA only when added at 1 mm; this
effect was doubled by 1 mm glycine. Quinolinate, at 100 mm,
elicited [3H]NA release only in presence of 1 mm glycine. In
contrast, 1 mm quinolinic acid, either alone or in presence of
glycine, failed to signi®cantly elevate tritium release from
striatal synaptosomes labelled with [3H]DA (Table 1).
NMDA-evoked [3H]NA and [3H]DA release:
effects of glycine, dd-serine or gp120
The results with quinolinic acid suggest possible differences
between the NMDA receptors under study. Considering the
reciprocal in¯uences occurring between coagonist sites of
the NMDA receptor, we then compared the sensitivity of the
hippocampal and the striatal NMDA receptors to glycine
and d-serine, the endogenous coagonists of glutamate and to
gp120, the HIV-1 envelope protein previously found to
potently mimic glycine/d-serine at the NMDA receptor
regulating NA release from hippocampal axon terminals
(Pattarini et al. 1998). As shown in Table 2, glycine and d-
serine, added at 0.1 or 1 mm, concentration-dependently
potentiated the effects of 100 mm NMDA on the release
of [3H]NA or [3H]DA. In contrast, gp120 (30±1000 pm)
selectively potentiated the NMDA-induced release of
[3H]NA, but was ineffective on the NMDA-induced release
of [3H]DA. The potency of gp120 at the noradrenergic
terminals was about four orders of magnitude higher than
that of glycine and d-serine, respectively, con®rming pre-
vious data from this laboratory (Pattarini et al. 1998).
Enhancement of the quinolinate effect on [3H]NA
release by gp120
It was important to test if the effect of quinolinic acid
also could be affected by gp120. Figure 1 shows that the
quinolinate-evoked release of tritium from hippocampal
Table 1 Effect of different glutamatergic agonists on the release of [3H]NA from hippocampal synaptosomes and of [3H]DA from striatal
synaptosomes
[3H]NA [3H]DA
Control Glycine (1 mM) Control Glycine (1 mM)
NMDA (100 mM) 37.6 �̂ 3.1 75.5 �^ 4.7** 14.3 �^ 1.9 41.1 �̂ 2.4**
L-aspartate (100 mM) 55.1 �^ 4.6 81.4 �^ 8.4* 17.9 �^ 1.5 44.2 �̂ 2.7**
Quinolinate (100 mM) 2.5 �^ 3.1 18.7 �^ 4.3* 1.3 �^ 2.7 3.2 �̂ 2.8
(1 mM) 15.3 �^ 1.5 34.5 �^ 2.7* 0.05 �^ 2.5 5.5 �̂ 1.8
Synaptosomes from the two rat brain areas were exposed to different glutamatergic agonists (concentration as indicated) alone or in presence of
glycine. Results are expressed as percentage increase over basal release. Data are means ^ SEM of ®ve to seven experiments run in triplicate
(three superfusion chambers for each experimental condition).
*p , 0.05 vs. respective control; **p , 0.01 vs. respective control.
Native NMDA receptor subtypes 141
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 139±148
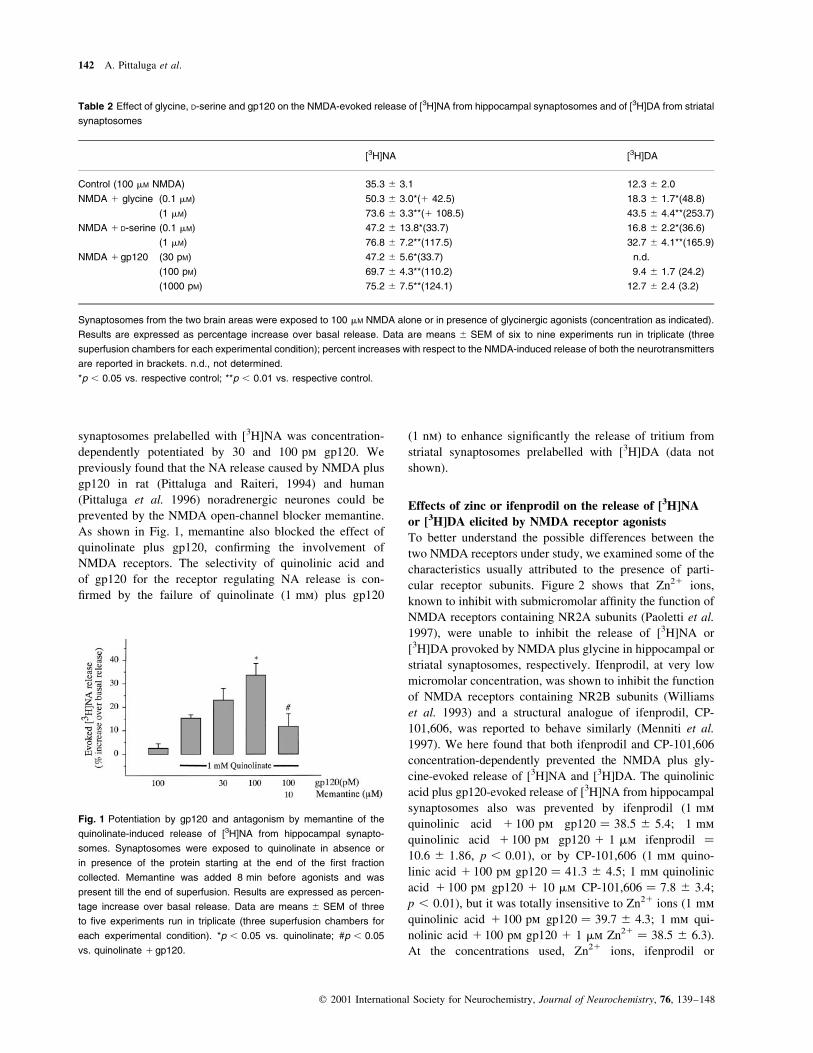
synaptosomes prelabelled with [3H]NA was concentration-
dependently potentiated by 30 and 100 pm gp120. We
previously found that the NA release caused by NMDA plus
gp120 in rat (Pittaluga and Raiteri, 1994) and human
(Pittaluga et al. 1996) noradrenergic neurones could be
prevented by the NMDA open-channel blocker memantine.
As shown in Fig. 1, memantine also blocked the effect of
quinolinate plus gp120, con®rming the involvement of
NMDA receptors. The selectivity of quinolinic acid and
of gp120 for the receptor regulating NA release is con-
®rmed by the failure of quinolinate (1 mm) plus gp120
(1 nm) to enhance signi®cantly the release of tritium from
striatal synaptosomes prelabelled with [3H]DA (data not
shown).
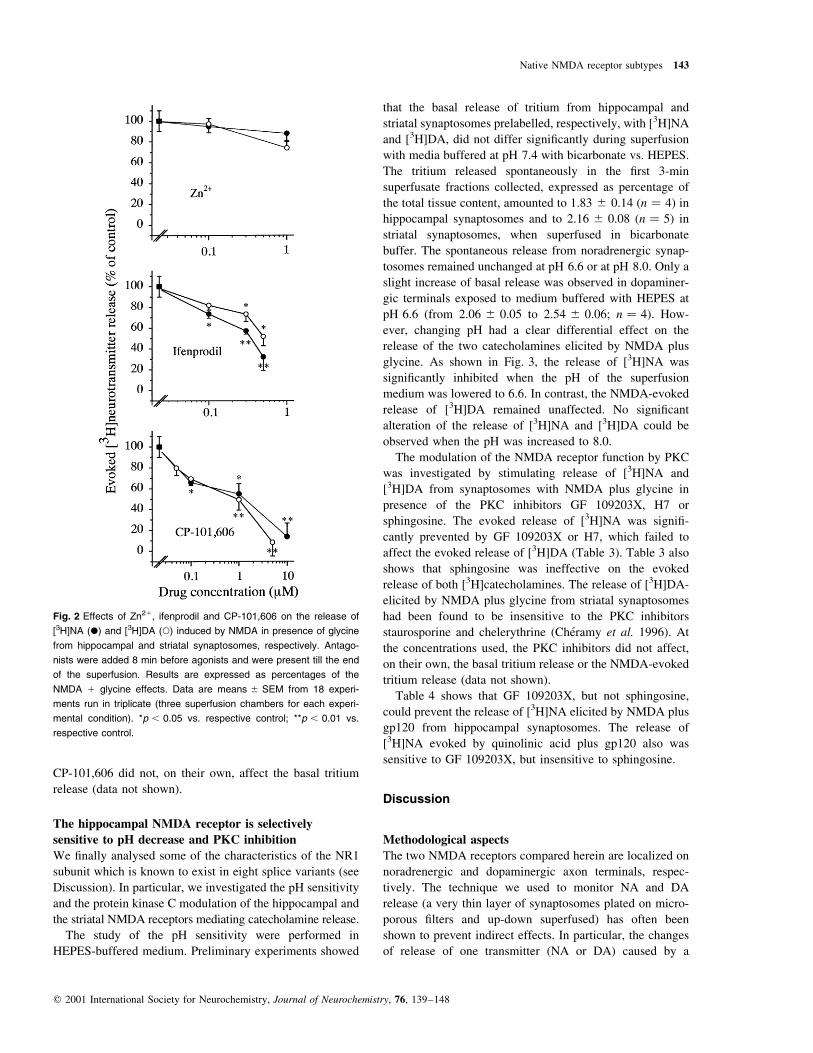
Effects of zinc or ifenprodil on the release of [3H]NA
or [3H]DA elicited by NMDA receptor agonists
To better understand the possible differences between the
two NMDA receptors under study, we examined some of the
characteristics usually attributed to the presence of parti-
cular receptor subunits. Figure 2 shows that Zn21 ions,
known to inhibit with submicromolar af®nity the function of
NMDA receptors containing NR2A subunits (Paoletti et al.
1997), were unable to inhibit the release of [3H]NA or
[3H]DA provoked by NMDA plus glycine in hippocampal or
striatal synaptosomes, respectively. Ifenprodil, at very low
micromolar concentration, was shown to inhibit the function
of NMDA receptors containing NR2B subunits (Williams
et al. 1993) and a structural analogue of ifenprodil, CP-
101,606, was reported to behave similarly (Menniti et al.
1997). We here found that both ifenprodil and CP-101,606
concentration-dependently prevented the NMDA plus gly-
cine-evoked release of [3H]NA and [3H]DA. The quinolinic
acid plus gp120-evoked release of [3H]NA from hippocampal
synaptosomes also was prevented by ifenprodil (1 mm
quinolinic acid 1 100 pm gp120 � 38.5 ^ 5.4; 1 mm
quinolinic acid 1 100 pm gp120 1 1 mm ifenprodil �10.6 ^ 1.86, p , 0.01), or by CP-101,606 (1 mm quino-
linic acid 1 100 pm gp120 � 41.3 ^ 4.5; 1 mm quinolinic
acid 1 100 pm gp120 1 10 mm CP-101,606 � 7.8 ^ 3.4;
p , 0.01), but it was totally insensitive to Zn21 ions (1 mm
quinolinic acid 1 100 pm gp120 � 39.7 ^ 4.3; 1 mm qui-
nolinic acid 1 100 pm gp120 1 1 mm Zn21 � 38.5 ^ 6.3).
At the concentrations used, Zn21 ions, ifenprodil or
Table 2 Effect of glycine, D-serine and gp120 on the NMDA-evoked release of [3H]NA from hippocampal synaptosomes and of [3H]DA from striatal
synaptosomes
[3H]NA [3H]DA
Control (100 mM NMDA) 35.3 �̂ 3.1 12.3 �^ 2.0
NMDA 1 glycine (0�.1 mM) 50.3 �^ 3.0*(1 42.5) 18.3 �̂ 1.7*(48.8)
(1 mM) 73.6 �^ 3.3**(1 108.5) 43.5 �^ 4.4**(253.7)
NMDA 1 D-serine (0�.1 mM) 47.2 �̂ 13.8*(33.7) 16.8 �̂ 2.2*(36.6)
(1 mM) 76.8 �^ 7.2**(117.5) 32.7 �̂ 4.1**(165.9)
NMDA 1 gp120 (30 pM) 47.2 �^ 5.6*(33.7) n.d.
(100 pM) 69.7 �̂ 4.3**(110.2) 9.4 �̂ 1.7 (24.2)
(1000 pM) 75.2 �̂ 7.5**(124.1) 12.7 �̂ 2.4 (3.2)
Synaptosomes from the two brain areas were exposed to 100 mM NMDA alone or in presence of glycinergic agonists (concentration as indicated).
Results are expressed as percentage increase over basal release. Data are means ^ SEM of six to nine experiments run in triplicate (three
superfusion chambers for each experimental condition); percent increases with respect to the NMDA-induced release of both the neurotransmitters
are reported in brackets. n.d., not determined.
*p , 0.05 vs. respective control; **p , 0.01 vs. respective control.
Fig. 1 Potentiation by gp120 and antagonism by memantine of the
quinolinate-induced release of [3H]NA from hippocampal synapto-
somes. Synaptosomes were exposed to quinolinate in absence or
in presence of the protein starting at the end of the ®rst fraction
collected. Memantine was added 8 min before agonists and was
present till the end of superfusion. Results are expressed as percen-
tage increase over basal release. Data are means ^ SEM of three
to ®ve experiments run in triplicate (three superfusion chambers for
each experimental condition). *p , 0.05 vs. quinolinate; #p , 0.05
vs. quinolinate 1 gp120.
142 A. Pittaluga et al.
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 139±148
CP-101,606 did not, on their own, affect the basal tritium
release (data not shown).
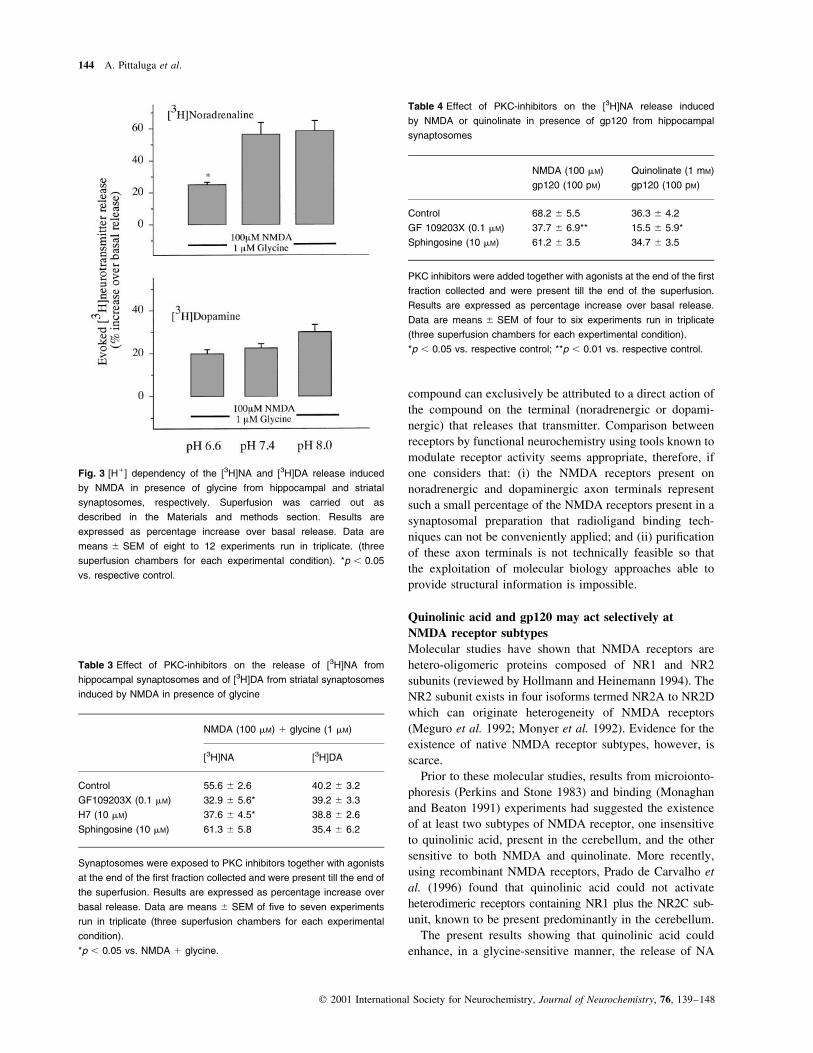
The hippocampal NMDA receptor is selectively
sensitive to pH decrease and PKC inhibition
We ®nally analysed some of the characteristics of the NR1
subunit which is known to exist in eight splice variants (see
Discussion). In particular, we investigated the pH sensitivity
and the protein kinase C modulation of the hippocampal and
the striatal NMDA receptors mediating catecholamine release.
The study of the pH sensitivity were performed in
HEPES-buffered medium. Preliminary experiments showed
that the basal release of tritium from hippocampal and
striatal synaptosomes prelabelled, respectively, with [3H]NA
and [3H]DA, did not differ signi®cantly during superfusion
with media buffered at pH 7.4 with bicarbonate vs. HEPES.
The tritium released spontaneously in the ®rst 3-min
superfusate fractions collected, expressed as percentage of
the total tissue content, amounted to 1.83 ^ 0.14 (n � 4) in
hippocampal synaptosomes and to 2.16 ^ 0.08 (n � 5) in
striatal synaptosomes, when superfused in bicarbonate
buffer. The spontaneous release from noradrenergic synap-
tosomes remained unchanged at pH 6.6 or at pH 8.0. Only a
slight increase of basal release was observed in dopaminer-
gic terminals exposed to medium buffered with HEPES at
pH 6.6 (from 2.06 ^ 0.05 to 2.54 ^ 0.06; n � 4). How-
ever, changing pH had a clear differential effect on the
release of the two catecholamines elicited by NMDA plus
glycine. As shown in Fig. 3, the release of [3H]NA was
signi®cantly inhibited when the pH of the superfusion
medium was lowered to 6.6. In contrast, the NMDA-evoked
release of [3H]DA remained unaffected. No signi®cant
alteration of the release of [3H]NA and [3H]DA could be
observed when the pH was increased to 8.0.
The modulation of the NMDA receptor function by PKC
was investigated by stimulating release of [3H]NA and
[3H]DA from synaptosomes with NMDA plus glycine in
presence of the PKC inhibitors GF 109203X, H7 or
sphingosine. The evoked release of [3H]NA was signi®-
cantly prevented by GF 109203X or H7, which failed to
affect the evoked release of [3H]DA (Table 3). Table 3 also
shows that sphingosine was ineffective on the evoked
release of both [3H]catecholamines. The release of [3H]DA-
elicited by NMDA plus glycine from striatal synaptosomes
had been found to be insensitive to the PKC inhibitors
staurosporine and chelerythrine (CheÂramy et al. 1996). At
the concentrations used, the PKC inhibitors did not affect,
on their own, the basal tritium release or the NMDA-evoked
tritium release (data not shown).
Table 4 shows that GF 109203X, but not sphingosine,
could prevent the release of [3H]NA elicited by NMDA plus
gp120 from hippocampal synaptosomes. The release of
[3H]NA evoked by quinolinic acid plus gp120 also was
sensitive to GF 109203X, but insensitive to sphingosine.
Discussion
Methodological aspects
The two NMDA receptors compared herein are localized on
noradrenergic and dopaminergic axon terminals, respec-
tively. The technique we used to monitor NA and DA
release (a very thin layer of synaptosomes plated on micro-
porous ®lters and up-down superfused) has often been
shown to prevent indirect effects. In particular, the changes
of release of one transmitter (NA or DA) caused by a
Fig. 2 Effects of Zn21, ifenprodil and CP-101,606 on the release of
[3H]NA (X) and [3H]DA (W) induced by NMDA in presence of glycine
from hippocampal and striatal synaptosomes, respectively. Antago-
nists were added 8 min before agonists and were present till the end
of the superfusion. Results are expressed as percentages of the
NMDA 1 glycine effects. Data are means ^ SEM from 18 experi-
ments run in triplicate (three superfusion chambers for each experi-
mental condition). *p , 0.05 vs. respective control; **p , 0.01 vs.
respective control.
Native NMDA receptor subtypes 143
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 139±148
compound can exclusively be attributed to a direct action of
the compound on the terminal (noradrenergic or dopami-
nergic) that releases that transmitter. Comparison between
receptors by functional neurochemistry using tools known to
modulate receptor activity seems appropriate, therefore, if
one considers that: (i) the NMDA receptors present on
noradrenergic and dopaminergic axon terminals represent
such a small percentage of the NMDA receptors present in a
synaptosomal preparation that radioligand binding tech-
niques can not be conveniently applied; and (ii) puri®cation
of these axon terminals is not technically feasible so that
the exploitation of molecular biology approaches able to
provide structural information is impossible.
Quinolinic acid and gp120 may act selectively at
NMDA receptor subtypes
Molecular studies have shown that NMDA receptors are
hetero-oligomeric proteins composed of NR1 and NR2
subunits (reviewed by Hollmann and Heinemann 1994). The
NR2 subunit exists in four isoforms termed NR2A to NR2D
which can originate heterogeneity of NMDA receptors
(Meguro et al. 1992; Monyer et al. 1992). Evidence for the
existence of native NMDA receptor subtypes, however, is
scarce.
Prior to these molecular studies, results from microionto-
phoresis (Perkins and Stone 1983) and binding (Monaghan
and Beaton 1991) experiments had suggested the existence
of at least two subtypes of NMDA receptor, one insensitive
to quinolinic acid, present in the cerebellum, and the other
sensitive to both NMDA and quinolinate. More recently,
using recombinant NMDA receptors, Prado de Carvalho et
al. (1996) found that quinolinic acid could not activate
heterodimeric receptors containing NR1 plus the NR2C sub-
unit, known to be present predominantly in the cerebellum.
The present results showing that quinolinic acid could
enhance, in a glycine-sensitive manner, the release of NA
Fig. 3 [H1] dependency of the [3H]NA and [3H]DA release induced
by NMDA in presence of glycine from hippocampal and striatal
synaptosomes, respectively. Superfusion was carried out as
described in the Materials and methods section. Results are
expressed as percentage increase over basal release. Data are
means ^ SEM of eight to 12 experiments run in triplicate. (three
superfusion chambers for each experimental condition). *p , 0.05
vs. respective control.
Table 3 Effect of PKC-inhibitors on the release of [3H]NA from
hippocampal synaptosomes and of [3H]DA from striatal synaptosomes
induced by NMDA in presence of glycine
NMDA (100 mM) 1 glycine (1 mM)
[3H]NA [3H]DA
Control 55.6 �^ 2.6 40.2 �^ 3.2
GF109203X (0�.1 mM) 32.9 �^ 5.6* 39.2 �^ 3.3
H7 (10 mM) 37.6 �̂ 4.5* 38.8 �^ 2.6
Sphingosine (10 mM) 61.3 �^ 5.8 35.4 �^ 6.2
Synaptosomes were exposed to PKC inhibitors together with agonists
at the end of the ®rst fraction collected and were present till the end of
the superfusion. Results are expressed as percentage increase over
basal release. Data are means ^ SEM of ®ve to seven experiments
run in triplicate (three superfusion chambers for each experimental
condition).
*p , 0.05 vs. NMDA 1 glycine.
Table 4 Effect of PKC-inhibitors on the [3H]NA release induced
by NMDA or quinolinate in presence of gp120 from hippocampal
synaptosomes
NMDA (100 mM)
gp120 (100 pM)
Quinolinate (1 mM)
gp120 (100 pM)
Control 68.2 �̂ 5.5 36.3 �̂ 4.2
GF 109203X (0�.1 mM) 37.7 �̂ 6.9** 15.5 �^ 5.9*
Sphingosine (10 mM) 61.2 �̂ 3.5 34.7 �̂ 3.5
PKC inhibitors were added together with agonists at the end of the ®rst
fraction collected and were present till the end of the superfusion.
Results are expressed as percentage increase over basal release.
Data are means ^ SEM of four to six experiments run in triplicate
(three superfusion chambers for each expertimental condition).
*p , 0.05 vs. respective control; **p , 0.01 vs. respective control.
144 A. Pittaluga et al.
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 139±148

but not that of DA suggest possible structural differences
between the NMDA receptors present, respectively, on
hippocampal noradrenergic and striatal dopaminergic axon
terminals (see below). The insensitivity to quinolinate is
therefore not restricted to cerebellar NMDA receptors, but it
also seems to be a characteristic of NMDA receptors present
on striatal dopaminergic axon terminals. Subtype selectivity
of quinolinate would be especially signi®cant, since the
compound, although a low af®nity agent, is endogenous and
its action on subpopulations of the NMDA receptor could
be of clinical interest. Certainly quinolinate seems to be
an interesting experimental tool to investigate receptor
subtypes.
Glycine and d-serine are endogenous glutamate coagonists
at NMDA receptors. As shown previously, and here con-
®rmed (see Table 2), the NMDA-evoked release of NA from
hippocampal synaptosomes can be potentiated by the HIV-1
envelope protein gp120. The protein seems to act through
its V3-loop at the glycine coagonist site of the NMDA
receptors localized on noradrenergic terminals, mimicking
glycine with a potency of approximately four orders of
magnitude higher than that of the endogenous agonists
(Pattarini et al. 1998). In contrast, gp120 was unable to
enhance the effect of NMDA alone on the release of DA
from striatal synaptosomes, whereas this effect could be
trebled by glycine (cf. Tables 1 and 2). These results with
gp120 also suggest involvement of two NMDA receptor
subtypes (see below).
Gp120 has been implicated in various neurological and
neuropsychiatric effects associated with AIDS. The selec-
tivity of action of the protein as a possible exogenous
glutamate coagonist at some NMDA receptors may there-
fore be of pathological interest. Also noteworthy, gp120 has
been proposed to act in concert with quinolinate and
glutamate/aspartate in the brain of AIDS patients (Lipton
1998) and quinolinate levels were shown to be largely
increased during HIV-1 infection and CNS in¯ammation
(Achim et al. 1993; Martin et al. 1993; Beagles et al. 1998).
Interestingly, low picomolar concentrations of gp120 could
potentiate the NA release brought about by quinolinate, an
effect prevented by the NMDA open channel antagonist
memantine (see Fig. 1). Considering the involvement of NA
in cognitive processes (McGaugh 1989; Izquierdo et al.
1993), the selectivity of quinolinic acid and gp120 for the
NMDA receptor-mediating release of NA in the hippo-
campus may play important role in AIDS dementia.
NR2B subunits in the hippocampal and striatal
NMDA receptors
Recent evidence indicates that NR2 subunits contain the
glutamate recognition site (Laube et al. 1997). The selec-
tivity for quinolinate of the receptor mediating NA release
could therefore be due to the presence, in the hippocampal
receptor, of NR2 subunits different from those in the striatal
receptor. Zinc ions at submicromolar concentrations are
known to preferentially affect NMDA receptors containing
NR2A subunits (Paoletti et al. 1997). The failure of 100 nm
Zn21 to inhibit the NMDA-evoked release of NA and that of
DA suggests that neither receptor contains NR2A subunits.
Instead, both receptors appear to contain subunits of the
NR2B isoform, in as much as the effects of NMDA plus
glycine on NA and DA release could be blocked by
ifenprodil or CP-101,606, two preferential inhibitors of
NMDA receptors containing NR2B subunits (Williams et al.
1993; Menniti et al. 1997). All together our results seem not
consistent with binding data showing that homoquinolinic
and quinolinic acid are relatively selective ligands for native
NR2B-containing receptors (Brown et al. 1998). Actually,
there seem to exist NR2B-containing receptors that do not
bind quinolinate. We considered the possibility that the
differential binding of quinolinic acid may not exclusively
originate from different NR2 subunits and that the NR1
subunit might play a role.
The NR1 subunit, which contains the glycine binding
domain (Kuryatov et al. 1994), is expressed in eight distinct
splice variants throughout the central nervous system. The
gene encoding the subunit contains three exons (5, 21 and
22) that are alternatively spliced to form these eight variants.
Exon-5 encodes a splice cassette of 21 amino acids (termed
N1) inserted in the extracellular amino-terminus domain.
Exons 21 and 22 encode two cassettes of 37 amino acids
(C1) and 38 amino acids (C2), respectively, located in the
intracellular carboxy-terminus domain (Nakanishi et al.
1992; Durand et al. 1993; Hollmann and Heinemann 1994;
Zukin and Bennett 1995). Since the NR1 splice variant can
affect the pharmacological and physiological properties of
heteromeric NR1-NR2 receptors (Hollmann et al. 1993; see
review by Zukin and Bennett 1995), the possibility exists
that the differential effects of quinolinate, as well as of
gp120, originate from the presence of different splice
variants of the NR1 subunit in the two NR2B-containing
NMDA receptors under study.
Different NR1 splice variants are present in the
hippocampal and striatal receptors
Protons can directly inhibit NMDA receptor function
(Traynelis and Cull-Candy 1990). The NR1 subunit contains
the pH sensor. The effect of protons is reduced by NR1
variants containing the exon-5 insert N1 (Traynelis et al.
1995). Our results suggest that the proton-sensitive NMDA
receptor sited on hippocampal noradrenergic nerve terminals
contain an N1-lacking NR1 subunit; in contrast, the proton-
insensitive NMDA receptor sited on striatal dopaminergic
nerve terminals may comprise an NR1 variant containing
the exon-5 insert.
The involvement of an exon-5 N1 insert may represent
one explanation for the differential effect of gp120. In
fact, the presence of the 21 amino acid cassette encoded by
Native NMDA receptor subtypes 145
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 139±148

exon-5 in the NR1 subunit has been proposed to in¯uence
the downstream S1 and S2 domains that constitute the
glycine binding pocket (Masuko et al. 1999). These
conformational changes could selectively affect the binding
of gp120, a molecule much larger than glycine/d-serine, and
prevent its action at the NMDA receptor regulating DA
release.
NMDA receptors can mediate calcium in¯ux with
consequent elevations in cytosolic calcium concentration
(Grimwood et al. 1996; Schousboe et al. 1997) and
activation of various enzymes, including PKC. This enzyme
can positively modulate the function of NMDA receptors
(Ben-Ari et al. 1992; Zukin and Bennett 1995; Bi et al.
1998; for reviews). Studies with recombinant receptors show
that the PKC potentiation correlates with the presence or
absence of the C1 sequence in the NR1 subunit, where the
presence of C1 leads to decrease in PKC potentiation.
Neither the N1 nor C2 exons appear to participate in the
potentiation (Logan et al. 1999).
Our results show that the effects of NMDA plus glycine
on NA release, but not on DA release, were blocked by
GF 109203X or H7 (Table 3). The selectivity of the
PKC blockers towards the NA release would be compatible
with the presence of the exon-21 C1 cassette in the striatal
NMDA receptor regulating DA release, but not in the
hippocampal NMDA receptor regulating the release of NA.
The PKC involved may be a Ca21-dependent diacyl-
glycerol-independent form, in as much as the NMDA
responses on NA release were sensitive to compounds
inhibiting PKC at the ATP binding site, but not by
sphingosine, which blocks PKC at the diacylglycerol
binding site.
The effects of NMDA plus gp120 or of quinolinate plus
gp120 also were prevented by GF 109203X, but not by
sphingosine (Table 4). Interestingly, NMDA agonists plus
gp120 applied to primary neuronal cultures were found to
cause a rise in the intracellular calcium level followed by
translocation of PKC from the cytosol to the membrane and
consequent sensitization of NMDA receptors (Ushijima et al.
1993). This also occurred in C6 astrocytoma cells exposed
to gp120 (Wyss-Coray et al. 1996). These authors also
found that PKC activity was upmodulated in the CNS of
gp120 transgenic mice and that brain tissue from patients
with HIV-1 showed increased PKC immunoreactivity.
Conclusions
We have compared two NMDA receptors located on
noradrenergic axon terminals of the hippocampus, where
they mediate release of noradrenaline, and on dopaminergic
axon terminals of the striatum, where they mediate release
of dopamine. The receptors appear to represent two subtypes
of the NMDA receptor because: (i) only the receptor
mediating NA release can be activated by quinolinate plus
glycine, NMDA plus gp120 or quinolinate plus gp120; and
(ii) the effect of NMDA plus glycine on the release of NA,
but not that on the release of DA, can be inhibited by
decreasing pH and by blocking PKC. These differential
responses may re¯ect structural differences. One possibility
is that the receptors, which both contain NR2B subunits,
display different splice variants of the NR1 subunit. In
particular, the NMDA receptor on noradrenergic neurones
might contain an NR1 subunit lacking both the N1 and the
C1 cassettes.
Quinolinic acid is present in normal postmortem human
brain (Moroni et al. 1984) at levels similar to those of rats
(Stone 1993 and references therein). The concentrations
found to be active here and in many other in vitro studies are
clearly much higher than those existing in normal human
brain tissue. However, during CNS in¯ammation and HIV-1
infection, the brain levels of quinolinate were found to be
largely increased (Achim et al. 1993; Martin et al. 1993;
Beagles et al. 1998). The extraordinarily high potency of
gp120 in enhancing selectively the effects of NMDA
receptor agonists, including quinolinate, at noradrenergic
terminals, and the rises of quinolinate levels during patho-
logical conditions, together with the absence of any speci®c
removal system for both quinolinic acid and gp120, could
play a role in the symptomatology of AIDS dementia.
Acknowledgements
This work was supported by the Italian Ministry of Health
(Contributo ISS, Programma Nazionale di Ricerca sull'AIDS,
Progetto Patologia Clinica e Terapia dell'AIDS 1998±99). The
authors thank Maura Agate for excellent assistance in preparing
the manuscript.
References
Achim C. L., Heyes M. P. and Wiley C. A. (1993) Quantitation of
human immunode®ciency virus, immune activation factors, and
quinolinic acid in AIDS brains. J. Clin. Invest. 91, 2769±2775.
Beagles K. E., Morrison P. F. and Heyes M. P. (1998) Quinolinic acid in
vivo synthesis rates, extracellular concentrations, and intercom-
partmental distributions in normal and immune-activated brain as
determined by multiple-isotope microdialysis. J. Neurochem. 70,
281±291.
Ben-Ari Y., Aniksztejn L. and Bregestovski P. (1992) Protein kinase C
modulation of NMDA currents: an important link for LTP
induction. Trends Neurosci. 15, 333±339.
Bi X., Standley S. and Baudry M. (1998) Posttranslational regulation of
ionotropic glutamate receptors and synaptic plasticity. Int. Rev.
Neurobiol. 42, 227±284.
Brown J. C. III, Tse H. W., Skifter D. A., Christie J. M., Andaloro V. J.,
Kemp M. C., Watkins J. C., Jane D. E. and Monaghan D. T.
(1998) [3H]Homoquinolinate binds to a subpopulation of
NMDA receptors and to a novel binding site. J. Neurochem. 71,
1464±1470.
CheÂramy A., Godeheu G., L'Hirondel M. and Glowinski J. (1996)
Cooperative contributions of cholinergic and NMDA receptor in
146 A. Pittaluga et al.
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 139±148

the presynaptic control of dopamine release from synaptosomes of
the rat striatum. J. Pharmacol. Exp. Ther. 276, 616±625.
Durand G. M., Bennett M. V. and Zukin R. S. (1993) Splice variants of
the N-methyl-d-aspartate receptor NR1 identify domains involved
in regulation by polyamines and protein kinase C. Proc. Natl
Acad. Sci. USA 90, 6731±6735.
Fink K., BoÈnisch H. and GoÈthert M. (1990) Presynaptic NMDA
receptors stimulate noradrenaline release in the cerebral cortex.
Eur. J. Pharmacol. 185, 115±117.
Foster A. C. and Schwarcz R. (1988) Neurotoxic effects of quinolinic
acid in the central nervous system, in Quinolinic Acid and Other
Kynurenines, pp. 173±192. CRC Press, Boca Raton, USA.
Giulian D., Yu J., Li X., Tom D., Li J., Wendt E., Lin S. N., Schwarcz
R. and Noonan C. (1996) Study of receptor-mediated neurotoxins
released by HIV-1-infected mononuclear phagocytes found in
human brain. J. Neurosci. 16, 3139±3153.
Grimwood S., Gilbert E., Ragan C. I. and Hutson P. H. (1996) Modula-
tion of 45Ca21 in¯ux into cells stably expressing recombinant
human NMDA receptors by ligands acting at distinct recognition
sites. J. Neurochem. 66, 2589±2595.
Guidetti P. and Schwarcz R. (1999) 3-Hydroxykynurenine potentiates
quinolinate but not NMDA toxicity in the rat striatum. Eur. J.
Neurosci. 11, 3857±3863.
Hollmann M., Boulter J., Maron C., Beasley L., Sullivan J., Pecht G.
and Heinemann S. (1993) Zinc potentiates agonist-induced
currents at certain splice variants of the NMDA receptor. Neuron
10, 943±954.
Hollmann M. and Heinemann S. (1994) Cloned glutamate receptors.
Annu. Rev. Neurosci. 17, 31±108.
Izquierdo I., Medina J. H., Bianchin M., Walz R., Zanatta M. S., Da
Silva R. C., Silva M. B. E., Ruschel A. C. and Paczko N. (1993)
Memory processing by the limbic system: role of speci®c
neurotransmitter systems. Behav. Brain Res. 58, 91±98.
Krebs M. O., Desce J. M., Kemel M. L., Gauchy C., Godeheu G.,
Cheramy A. and Glowinski J. (1991) Glutamatergic control of
dopamine release in the rat striatum: evidence for presynaptic
N-methyl-d-aspartate receptors on dopaminergic nerve terminals.
J. Neurochem. 56, 81±85.
Kuryatov A., Laube B., Betz H. and Kuhse J. (1994) Mutational
analysis of the glycine-binding site of the NMDA receptor:
structural similarity with bacterial amino acid-binding proteins.
Neuron 12, 1291±1300.
Laube B., Hirai H., Sturgess M., Betz H. and Kuhse J. (1997) Molecular
determinants of agonist discrimination by NMDA receptor
subunits: analysis of the glutamate binding site on the NR2B
subunit. Neuron 18, 493±503.
Lipton S. A. (1998) Neuronal injury associated with HIV-1: approaches
to treatment. Annu. Rev. Pharmacol. Toxicol. 38, 159±177.
Logan S. M., Rivera F. E. and Leonard J. P. (1999) Protein kinase C
modulation of recombinant NMDA receptor currents: roles for the
C-terminal C1 exon and calcium ions. J. Neurosci. 19, 974±986.
Martin A., Heyes M. P., Salazar A. M., Law W. A. and Williams J.
(1993) Impaired motor-skill learning, slowed reaction time, and
elevated cerebrospinal ¯uid quinolinic acid in a sub-group of HIV-
infected individuals. Neuropsychology 7, 149±157.
Masuko T., Kashiwagi K., Kuno T., Nguyen N. D., Pahk A. J., Fukuchi
J., Igarashi K. and Williams K. (1999) A regulatory domain
(R1±R2) in the amino terminus of the N-methyl-d-aspartate
receptor: effects of spermine, protons, and ifenprodil, and
structural similarity to bacterial leucine/isoleucine/valine binding
protein. Mol. Pharmacol. 55, 957±969.
McGaugh J. (1989) Involvement of hormonal and neuromodulatory
systems in the regulation of memory storage. Annu. Rev. Neurosci.
12, 255±287.
Meguro H., Mori H., Araki K., Kushiya E., Kutsuwada T., Yamazaki
M., Kumanishi T., Arakawa M., Sakimura K. and Mishina M.
(1992) Functional characterization of a heteromeric NMDA
receptor channel expressed from cloned cDNAs. Nature 357,
70±74.
Menniti F., Chenard B., Collins M., Ducat M., Shalaby I. and White F.
(1997) CP-101,606, a potent neuroprotectant selective for
forebrain neurons. Eur. J. Pharmacol. 331, 117±126.
Monaghan D. T. and Beaton J. A. (1991) Quinolinate differentiates
between forebrain and cerebellar NMDA receptors. Eur. J.
Pharmacol. 194, 123±125.
Monyer H., Sprengel R., Schoepfer R., Herb A., Higuchi M., Lomeli H.,
Burnashev N., Sakmann B. and Seeburg P. H. (1992) Heteromeric
NMDA receptors: molecular and functional distinction of sub-
types. Science 256, 1217±1221.
Moroni F. (1999) Tryptophan metabolism and brain function: focus on
kynurenine and other indole metabolites. Eur. J. Pharmacol. 375,
87±100.
Moroni F., Lombardi G., Carla V. and Moneti G. (1984) The excito-
toxin quinolinic acid is present and unevenly distributed in the rat
brain. Brain Res. 285, 352±355.
Nakanishi N., Axel R. and Schneider N. A. (1992) Alternative splicing
generates functionally distinct N-methyl-d-aspartate receptors.
Proc. Natl Acad. Sci. USA 89, 8552±8556.
Paoletti P., Ascher P. and Neyton J. (1997) High-af®nity zinc inhibition
of NMDA NR1-NR2A receptors. J. Neurosci. 17, 5711±5725.
Pattarini R., Pittaluga A. and Raiteri M. (1998) The human immuno-
de®ciency virus-1 envelope protein gp120 binds through its V3
sequence to the glycine site of N-methyl-d-aspartate receptors
mediating noradrenaline release in the hippocampus. Neuro-
science 87, 147±157.
Paudice P., Gemignani A. and Raiteri M. (1998) Evidence for func-
tional native NMDA receptors activated by glycine or d-serine
alone in the absence of glutamatergic coagonist. Eur. J. Neurosci.
10, 2934±2944.
Perkins M. N. and Stone T. W. (1983) Quinolinic acid: regional
variations in neuronal sensitivity. Brain Res. 259, 172±176.
Pittaluga A., Pattarini R., Severi P. and Raiteri M. (1996) Human brain
N-methyl-d-aspartate receptors regulating noradrenaline release
are positively modulated by HIV-1 coat protein gp120. AIDS 10,
463±468.
Pittaluga A. and Raiteri M. (1990) Release-enhancing glycine-
dependent presynaptic NMDA receptors exist on noradrenergic
terminals of hippocampus. Eur. J. Pharmacol. 191, 231±234.
Pittaluga A. and Raiteri M. (1994) HIV-1 envelope protein gp120
potentiates NMDA-evoked noradrenaline release by a direct
action at rat hippocampal and cortical noradrenergic nerve
endings. Eur. J. Neurosci. 6, 1743±1749.
Prado de Carvalho L., Bochet P. and Rossier J. (1996) The endogenous
agonist quinolinic acid and the non endogenous homoquinolinic
acid discriminate between NMDAR2 receptor subunits. Neuro-
chem. Int. 28, 445±452.
Raiteri M., Angelini F. and Levi. G. (1974) A simple apparatus for
studying the release of neurotransmitters from synaptosomes. Eur.
J. Pharmacol. 25, 411±414.
Raiteri M., Bonanno G., Marchi M. and Maura G. (1984) Is there a
functional linkage between neurotransmitter uptake mech-
anisms and presynaptic receptors? J. Pharmacol. Exp. Ther. 231,
671±677.
Raiteri L. and Raiteri M. (2000) Synaptosomes still viable after 25 years
of superfusion. Neurochem. Res. 25, 1265±1274.
Roberts P. J. and Anderson S. D. (1979) Stimulatory effect of
l-glutamate and related amino acids on [3H]dopamine release
Native NMDA receptor subtypes 147
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 139±148

from rat striatum: an in vitro model for glutamate actions.
J. Neurochem. 32, 1539±1545.
Schousboe A., Belhage B. and Frandsen A. (1997) Role of Ca21 and
other second messengers in excitatory amino acid receptor
mediated neurodegeneration: clinical perspectives. Clin. Neurosci.
4, 194±198.
Schroeder H. C., Perovic S., Kavsan V., Ushijima H. and Muller W. E.
(1998) Mechanisms of prionSc- and HIV-1 gp120 induced
neuronal cell death. Neurotoxicology 19, 683±688.
Stone T. W. (1993) Neuropharmacology of quinolinic and kynurenic
acids. Pharmacol. Rev. 45, 309±379.
Stone T. W. and Perkins M. N. (1981) Quinolinic acid: a potent
endogenous excitant at amino acid receptors in CNS. Eur. J.
Pharmacol. 72, 411±412.
Traynelis S. F. and Cull-Candy S. G. (1990) Proton inhibition of
N-methyl-d-aspartate receptors in cerebellar neurons. Nature 345,
347±350.
Traynelis S. F., Hartley M. and Heinemann S. F. (1995) Control of
proton sensitivity of the NMDA receptor by RNA splicing and
polyamines. Science 268, 873±876.
Ushijima H., Unten S., Honma H., Tsuchie H., Kitamura T., Weiler
B. E. and Muller W. E. (1993) Effect of serum components on
syncytium formation and virus production by cells infected with
human immunode®ciency virus in vitro. AIDS Res. Hum. Retro-
viruses 8, 513±520.
Williams K., Russell S. L., Shen Y. M. and Molinoff P. B. (1993)
Developmental switch in the expression of NMDA receptors
occurs in vivo and in vitro. Neuron 10, 267±278.
Wyss-Coray T., Masliah E., Toggas S. M., Rockenstein E. M., Brooker
M. J., Lee H. S. and Mucke L. (1996) Dysregulation of signal
transduction pathways as a potential mechanism of nervous
system alterations in HIV-1 gp120 transgenic mice and humans
with HIV-1 encephalitis. J. Clin. Invest. 97, 789±798.
Zukin R. S. and Bennett M. V. L. (1995) Alternatively spliced isoforms
of the NMDAR1 receptor subunit. Trends Neurosci. 18, 306±313.
148 A. Pittaluga et al.
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 139±148













![4Aminopyridine stimulates B50 (GAP43) phosphorylation and [3H]-noradrenaline release in rat hippocampal slices](https://static.fdokumen.com/doc/165x107/6314ccec3ed465f0570b4d12/4aminopyridine-stimulates-b50-gap43-phosphorylation-and-3h-noradrenaline-release.jpg)






