
Unique Mutational Patterns in the Envelope 2 Amphipathic Helix and Acquisition of Length in gp120...
11
JOURNAL OF VIROLOGY, June 2007, p. 5658–5668 Vol. 81, No. 11 0022-538X/07/$08.000 doi:10.1128/JVI.00257-07 Copyright © 2007, American Society for Microbiology. All Rights Reserved. Unique Mutational Patterns in the Envelope 2 Amphipathic Helix and Acquisition of Length in gp120 Hypervariable Domains Are Associated with Resistance to Autologous Neutralization of Subtype C Human Immunodeficiency Virus Type 1 Rong Rong, 1 S. Gnanakaran, 5 Julie M. Decker, 6,7 Frederic Bibollet-Ruche, 6,7 Jesse Taylor, 5 † Jeffrey N. Sfakianos, 8 John L. Mokili, 5 ‡ Mark Muldoon, 9 Joseph Mulenga, 10 Susan Allen, 3 Beatrice H. Hahn, 6,7 George M. Shaw, 6,7 Jerry L. Blackwell, 2,4 Bette T. Korber, 5,11 Eric Hunter, 1,2 and Cynthia A. Derdeyn 1,2 * Department of Pathology and Laboratory Medicine, 1 Yerkes National Primate Research Center, 2 Department of International Health, 3 and Division of Infectious Diseases, 4 Emory University, Atlanta, Georgia; Theoretical Biology and Biophysics Group, Los Alamos National Laboratory, Los Alamos, New Mexico 5 ; Department of Medicine 6 and Department of Microbiology, University of Alabama at Birmingham, Birmingham, Alabama 7 ; Department of Cell Biology, Yale University School of Medicine, New Haven, Connecticut 8 ; Department of Mathematics, University of Manchester Institute of Science and Technology, Manchester, United Kingdom 9 ; Zambia Blood Transfusion Service, Lusaka, Zambia 10 ; and The Santa Fe Institute, Santa Fe, New Mexico 11 Received 6 February 2007/Accepted 7 March 2007 Autologous neutralizing antibodies (NAb) against human immunodeficiency virus type 1 generate viral escape variants; however, the mechanisms of escape are not clearly defined. In a previous study, we determined the susceptibilities of 48 donor and 25 recipient envelope (Env) glycoproteins from five subtype C heterosexual transmission pairs to NAb in donor plasma by using a virus pseudotyping assay, thereby providing an ideal setting to probe the determinants of susceptibility to neutralization. In the present study, acquisition of length in the Env gp120 hypervariable domains was shown to correlate with resistance to NAb in donor plasma (P 0.01; Kendall’s tau test) but not in heterologous plasma. Sequence divergence in the gp120 V1-to-V4 region also correlated with resistance to donor (P 0.0002) and heterologous (P 0.001) NAb. A mutual information analysis suggested possible associations of nine amino acid positions in V1 to V4 with NAb resistance to the donor’s antibodies, and five of these were located within an 18-residue amphipathic helix (2) located on the gp120 outer domain. High nonsynonymous-to-synonymous substitution (dN/dS) ratios, indicative of positive selection, were also found at these five positions in subtype C sequences in the database. Nevertheless, exchange of the entire 2 helix between resistant donor Envs and sensitive recipient Envs did not alter the NAb phenotype. The combined mutual information and dN/dS analyses suggest that unique mutational patterns in 2 and insertions in the V1-to-V4 region are associated with NAb resistance during subtype C infection but that the selected positions within the 2 helix must be linked to still other changes in Env to confer antibody escape. These findings suggest that subtype C viruses utilize mutations in the 2 helix for efficient viral replication and immune avoidance. Circulating human immunodeficiency virus type 1 (HIV-1) strains belong to one of nine subtypes (A, B, C, D, F, G, H, J, and K) and at least 28 circulating recombinant forms (52; see http://www.hiv.lanl.gov/content/hiv-db/CRFs/CRFs.html for a current listing). Because subtype B infections predominate in developed regions of the world, the majority of studies char- acterizing the structure of gp120, as well as the interplay be- tween HIV-1 and neutralizing antibodies (NAb), have been performed using samples collected from subtype B-infected subjects. More information is therefore needed about the se- rology of non-subtype B infections because of their pervasive- ness in the AIDS pandemic (52, 57) and the urgent need for a vaccine that can induce cross-reactive immune responses. Sub- type C viruses, which circulate predominantly in sub-Saharan Africa and India, are often refractory to neutralization by monoclonal antibodies raised from subtype B-infected subjects (4, 7, 27). Consequently, little is known about the mechanisms of autologous neutralization and escape in subtype C infection. The HIV-1 genome encodes two glycoproteins; the trans- membrane subunit gp41 anchors the Env complex in the viral membrane and mediates fusion with the host cell membrane, while the surface subunit gp120 facilitates interactions with receptor molecules (30). HIV-1 gp120 is heavily glycosylated and contains five “hypervariable” domains (V1 to V5) that tolerate sequence heterogeneity (47). The V1V2 and V4 do- mains, in particular, can also accommodate dramatic inser- tions, deletions, and variation in the pattern of N-linked gly- * Corresponding author. Mailing address: Emory Vaccine Center, Emory University, 954 Gatewood Rd., Suite 1024, Atlanta, GA 30329. Phone: (404) 727-8594. Fax: (404) 727-9316. E-mail: cynthia.derdeyn @emory.edu. † Present address: La Jolla Institute for Allergy and Immunology, La Jolla, California. ‡ Present address: Department of Statistics, Oxford University, Ox- ford, United Kingdom. Published ahead of print on 14 March 2007. 5658
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Unique Mutational Patterns in the Envelope 2 Amphipathic Helix and Acquisition of Length in gp120...

JOURNAL OF VIROLOGY, June 2007, p. 5658–5668 Vol. 81, No. 110022-538X/07/$08.00�0 doi:10.1128/JVI.00257-07Copyright © 2007, American Society for Microbiology. All Rights Reserved.
Unique Mutational Patterns in the Envelope �2 Amphipathic Helixand Acquisition of Length in gp120 Hypervariable Domains Are
Associated with Resistance to Autologous Neutralization ofSubtype C Human Immunodeficiency Virus Type 1�
Rong Rong,1 S. Gnanakaran,5 Julie M. Decker,6,7 Frederic Bibollet-Ruche,6,7 Jesse Taylor,5†Jeffrey N. Sfakianos,8 John L. Mokili,5‡ Mark Muldoon,9 Joseph Mulenga,10 Susan Allen,3
Beatrice H. Hahn,6,7 George M. Shaw,6,7 Jerry L. Blackwell,2,4 Bette T. Korber,5,11
Eric Hunter,1,2 and Cynthia A. Derdeyn1,2*Department of Pathology and Laboratory Medicine,1 Yerkes National Primate Research Center,2 Department of International Health,3
and Division of Infectious Diseases,4 Emory University, Atlanta, Georgia; Theoretical Biology and Biophysics Group,Los Alamos National Laboratory, Los Alamos, New Mexico5; Department of Medicine6 and Department of Microbiology,
University of Alabama at Birmingham, Birmingham, Alabama7; Department of Cell Biology, Yale University School ofMedicine, New Haven, Connecticut8; Department of Mathematics, University of Manchester Institute of Science and
Technology, Manchester, United Kingdom9; Zambia Blood Transfusion Service, Lusaka, Zambia10;and The Santa Fe Institute, Santa Fe, New Mexico11
Received 6 February 2007/Accepted 7 March 2007
Autologous neutralizing antibodies (NAb) against human immunodeficiency virus type 1 generate viralescape variants; however, the mechanisms of escape are not clearly defined. In a previous study, we determinedthe susceptibilities of 48 donor and 25 recipient envelope (Env) glycoproteins from five subtype C heterosexualtransmission pairs to NAb in donor plasma by using a virus pseudotyping assay, thereby providing an idealsetting to probe the determinants of susceptibility to neutralization. In the present study, acquisition of lengthin the Env gp120 hypervariable domains was shown to correlate with resistance to NAb in donor plasma (P �0.01; Kendall’s tau test) but not in heterologous plasma. Sequence divergence in the gp120 V1-to-V4 region alsocorrelated with resistance to donor (P � 0.0002) and heterologous (P � 0.001) NAb. A mutual informationanalysis suggested possible associations of nine amino acid positions in V1 to V4 with NAb resistance to thedonor’s antibodies, and five of these were located within an 18-residue amphipathic helix (�2) located on thegp120 outer domain. High nonsynonymous-to-synonymous substitution (dN/dS) ratios, indicative of positiveselection, were also found at these five positions in subtype C sequences in the database. Nevertheless,exchange of the entire �2 helix between resistant donor Envs and sensitive recipient Envs did not alter the NAbphenotype. The combined mutual information and dN/dS analyses suggest that unique mutational patterns in�2 and insertions in the V1-to-V4 region are associated with NAb resistance during subtype C infection but thatthe selected positions within the �2 helix must be linked to still other changes in Env to confer antibody escape.These findings suggest that subtype C viruses utilize mutations in the �2 helix for efficient viral replication andimmune avoidance.
Circulating human immunodeficiency virus type 1 (HIV-1)strains belong to one of nine subtypes (A, B, C, D, F, G, H, J,and K) and at least 28 circulating recombinant forms (52; seehttp://www.hiv.lanl.gov/content/hiv-db/CRFs/CRFs.html for acurrent listing). Because subtype B infections predominate indeveloped regions of the world, the majority of studies char-acterizing the structure of gp120, as well as the interplay be-tween HIV-1 and neutralizing antibodies (NAb), have beenperformed using samples collected from subtype B-infected
subjects. More information is therefore needed about the se-rology of non-subtype B infections because of their pervasive-ness in the AIDS pandemic (52, 57) and the urgent need for avaccine that can induce cross-reactive immune responses. Sub-type C viruses, which circulate predominantly in sub-SaharanAfrica and India, are often refractory to neutralization bymonoclonal antibodies raised from subtype B-infected subjects(4, 7, 27). Consequently, little is known about the mechanismsof autologous neutralization and escape in subtype C infection.
The HIV-1 genome encodes two glycoproteins; the trans-membrane subunit gp41 anchors the Env complex in the viralmembrane and mediates fusion with the host cell membrane,while the surface subunit gp120 facilitates interactions withreceptor molecules (30). HIV-1 gp120 is heavily glycosylatedand contains five “hypervariable” domains (V1 to V5) thattolerate sequence heterogeneity (47). The V1V2 and V4 do-mains, in particular, can also accommodate dramatic inser-tions, deletions, and variation in the pattern of N-linked gly-
* Corresponding author. Mailing address: Emory Vaccine Center,Emory University, 954 Gatewood Rd., Suite 1024, Atlanta, GA 30329.Phone: (404) 727-8594. Fax: (404) 727-9316. E-mail: [email protected].
† Present address: La Jolla Institute for Allergy and Immunology, LaJolla, California.
‡ Present address: Department of Statistics, Oxford University, Ox-ford, United Kingdom.
� Published ahead of print on 14 March 2007.
5658

cosylation (N-Gly) sites. The V1V2 domain has been shown toinfluence sensitivity to NAb in studies of subtype B viruses (6,13, 26, 33, 35, 36, 49–51, 54, 59, 63, 70, 71, 74) and, morerecently, subtype C viruses (66). The V1V2 domain is thoughtto regulate neutralization sensitivity by masking conservedneutralization targets (39, 40, 44, 59, 66, 76) and can alsopresent type-specific neutralization epitopes (20, 21, 23, 28, 32,53, 58, 73). The mechanism by which V4 could influence neu-tralization is less clear, although this region could modulateglycan packing on the outer surface of the Env trimer (74).
There is evidence that distinct selective pressures are activeagainst Env during infections with different subtypes (14, 22).In an analysis of subtype C sequences from the HIV Databases(http://www.hiv.lanl.gov), a high density of positions with non-synonymous-to-synonymous substitution (dN/dS) ratios of �1was located in the third conserved region (C3) of gp120, whichis downstream in the linear sequence from the third hypervari-able domain (V3) (22). High dN/dS ratios are indicative ofstrong diversifying selection (77). In contrast, high dN/dS ratiosin subtype B sequences were concentrated within the bound-aries of V3, which, in contrast, is relatively conserved in sub-type C viruses (14, 22, 38). These findings suggest that thenature of NAb targets in the V3/C3 region differs betweensubtypes B and C (7, 24, 25, 46, 48, 74). Furthermore, werecently demonstrated that an amphipathic helix (�2) encodedin C3 exhibits mutational and structural differences in subtypesB and C (22a). Thus, the distributions of amino acid residuesunder selective pressure in the V3/C3 region are dramaticallydifferent in subtypes B and C, but the relationships betweenmutational patterns, viral subtypes, and sensitivity to autolo-gous NAb have not been elucidated.
We previously demonstrated that Envs from chronically in-fected (donor) transmission partners exhibited a range of re-sistance against contemporaneous donor NAb in a pseudovirusassay, but the Envs from their newly infected recipient partnerswere uniformly sensitive to NAb in this plasma sample (18).Using sequence and NAb data generated in the previous studyfrom 48 donor and 25 recipient Envs, we applied two statisticalapproaches to probe the determinants of neutralization sensi-tivity to donor plasma in the present study and used a biolog-ical assay to evaluate specific contributions. The results suggestthat acquisition of length in the hypervariable domains andmutation of the �2 helix domain track with NAb resistance, butthey demonstrate that the positions within the �2 helix must belinked to still other domains in Env that confer antibody es-cape. These findings suggest that the Env trimers of subtype Cviruses utilize mutations in the �2 helix to escape NAb. Thiscould be a pathway unique to subtype C viruses and is there-fore important for development of a vaccine that induces po-tent and cross-reactive NAb responses.
MATERIALS AND METHODS
Sequence data. Env sequences encompassing the V1-to-V4 region of gp120(HXB2 residues 136 to 418) were generated from seven subtype C heterosexualtransmission pairs and one subtype G heterosexual transmission pair enrolled ina discordant couple cohort in Lusaka, Zambia (18, 72). Evaluation of sensitivityto neutralization by plasma from the donor partner was performed previously forfive of the subtype C transmission pairs (18, 72). Details of the cohort and samplecollection have also been described previously (18, 72). Sequences are availablefrom GenBank under accession numbers AY423908 to AY424198.
Determination of BL. A maximum likelihood (ML) tree was constructedpreviously for each transmission pair (18), with gapped regions (including se-quences in the first, second, and fourth hypervariable domains [V1V2 and V4])stripped out of the analysis. Modeltest was used to identify the optimal evolu-tionary model (60), and for each tree, the model selected included base frequen-cies, a general reversible model for base substitution, and differences in ratevariation at different sites assigned according to a gamma distribution with fourcategories of rates and invariant sites. A further comparison was made with aREV model with ML assigned rate variation at different sites, but the additionalparameters were not justified in a likelihood ratio test for these data, and thus theModeltest model was used. The specific value for each of the model parameterswas estimated for each of the eight trees separately. PAUP was then used toconstruct a likelihood tree for each transmission pair. BranchLength.pl (B.Korber [www.santafe.edu/�btk/science-paper/bette.html]; a web-based versionof this code is now available at http://hcv.lanl.gov/content/hcv-db/BRANCHLEN/branchlength.html) was used to calculate the branch length (BL) to the patient’sancestral node for each donor and recipient sequence.
Correlation analysis. A nonparametric Kendall tau rank correlation test wasapplied to previously described paired sequence and neutralization data (18) todetermine whether the 50% inhibitory concentration (IC50) of plasma was cor-related with the following different parameters: BL to the patient’s ancestralnode on the ML tree, amino acid length of the V1-to-V4 region, and the numberof N-Gly sites in V1-to-V4. P values of �0.05 were considered statisticallyinteresting for hypothesis forming as a foundation for further experimentalexploration. The correlation tests were performed with the donor plasma IC50
and pooled plasma IC50 against each sequence attribute.Mutual information analysis. Shannon entropy is a measure of uncertainty
used in communication theory (64). It can be applied to other problems wherecharacter states are considered, such as sequence analysis (37). For example, forposition i in a protein alignment, the Shannon entropy function H(i) is defined bythe probability of different amino acids, s, in that position, as follows: H(i) ��S � P(si) � log P(si). This is a measure of the uncertainty in the column of data;if only one amino acid appears in a column, it has zero entropy. Maximal entropyoccurs when the frequencies of all of the characters are equal. Here weconsidered j a character state assigned according to the level of neutralizationsensitivity associated with a V1-to-V4 sequence (a, IC50 of �50; b, IC50 of 50to 249; c, IC50 of 250 to 999; d, IC50 of 1,000 to 4,999; and e, IC50 of �5,000).We asked whether j covaries with amino acid patterns found in any givenposition in an alignment. Given the joint probability distribution P(si,sj), H(i,j) ��S � P(si,sj) � log P(si,sj) and the mutual information distribution M(i,j) � H(i) �H(j) � H(i,j) (37), we measured how much the data covaries, given that themutual information is 0 when either i and j are completely independent or thereis no variation. To test the significance of these results, 10,000 randomizations ofneutralization phenotype assignments to viral sequences were performed, andthe number of times the mutual information for the random data was greaterthan or equal to the original data was tallied and divided by 10,000 to estimatethe P value. This statistical approach is based on the assumption that the data aredrawn from a random pool; however, the sequence data have strong underlyingphylogenetic correlations (2). To partially compensate for this, the randomiza-tion of neutralization assignments to sequences was done within donor-recipientpairs; however, the associations identified must therefore still be considered withthis limitation in mind and thus viewed in a hypothesis-forming framework toguide experiments and for comparison with other results, such as the dN/dSratios. Hypervariable loops were omitted from this analysis because we could notexclude the possibility that alignment artifacts could occur in the associationpatterns.
Codon-specific dN/dS ratios. Previously described data regarding codon-spe-cific dN/dS ratios were adapted (22). Nonsynonymous mutations become fixedwith greater or lesser probability than synonymous mutations depending onwhether a site is subject to positive or purifying selection, and thus the dN/dSratio can be used to characterize specific codons and regions of proteins evolvingunder positive selection (77). In viruses, regions under strong positive selectionoccur where immune escape confers a selective advantage to new variants thatarise within a host. Accordingly, codon-based models of molecular evolution canbe used in conjunction with sequence data and ML parameter estimation toidentify sites of potential immunological interest. The dN/dS ratio is indicative ofthe selection pressure at the protein level, as follows: a dN/dS ratio of �1 isindicative of purifying selection and amino acid conservation because of struc-tural and functional constraints, and a dN/dS ratio of �1 is indicative of diver-sifying, positive selection where amino acid substitutions confer an advantage. Inthis study, we asked whether or not the precise positions under positive selectivepressure may be lineage specific for HIV-1, comparing subtype B and subtype C,as representatives of both lineages are under consideration as vaccine candidates
VOL. 81, 2007 UNIQUE MUTATIONAL PATTERNS IN SUBTYPE C HIV-1 Env 5659

in trial populations where the HIV-1 C clade dominates. The programCODEML (S8 [http://abacus.gene.ucl.ac.uk/software/paml.html]) was used todetermine the regional distributions of dN/dS ratios in the V3 loop and flankingregions for two sets of 25 HIV-1 sequences, one for subtype B and the second forsubtype C. The subtype C sequences used did not include those from the Zam-bian transmission pairs analyzed for neutralization sensitivity here. For eachsubtype, a three-rate prior with flexible locations and weights was fitted to thedata. Bayes’s formula was then used to calculate the posterior distribution of therate ratio at each codon. The posterior mean was taken as a site-specific estimateof the dN/dS ratio.
Three-dimensional mapping. The core structure used for mapping corre-sponds to the X-ray structure of CD4-bound YU2 gp120 (43) (PDB code 1RZK).Variable loops V1V2 and V3 were modeled for clarity as described previously(5). Discussions on an unliganded gp120 core structure were based on the fullyglycosylated simian immunodeficiency virus (SIV) structure (12) (PDB code2BF1). Signature positions were mapped onto this structure based on the align-ment of sequences. Modeling calculations were performed using a modifiedversion of AMBER (56). Three-dimensional images were generated using VMD(29).
Molecular dynamics simulations. Comparative all-atom molecular dynamicssimulations were carried out on the modeled gp120 to capture the local differ-ences in the V3-C3-V4 regions of gp120 between B and C subtypes. The follow-ing three sets of simulations were carried out under identical conditions: (i)gp120 from YU2 (subtype B), (ii) YU2 gp120 with the �2 helix consensussequence from subtype C, and (iii) gp120 with the subtype C consensus sequence.In all cases, the gp120 protein was solvated in �16,000 water molecules. Thestarting structure for subtype C gp120 was based on the YU2 model describedabove. The subtype C consensus was homology modeled from the subtype Bstructure, using MODELLER software (69). All molecular dynamics simulationswere performed using the force field described by Cornell et al. (15) and theAMBER 6.0 suite of programs (9). A rectangular box of TIP3P water molecules(31) was added to solvate the complex, with the final system containing �55,000atoms within a box dimension of �79 by 79 by 89 Å. Periodic boundary condi-tions were applied, and the particle mesh Ewald approach (16) was used toaccurately treat electrostatic interactions. Both systems were minimized andequilibrated with the same protocol. The temperature was maintained at 300 K,using the Berendsen temperature algorithm (3). A 2-fs time step was used, andall bonds involving hydrogen atoms were constrained using the SHAKE algo-rithm (68). Simulations were carried out for approximately 4 ns, and the last 2 nswere considered for analysis. Additional details of the simulations and the resultsare provided in a separate publication (22a). The relative solvation betweensubtypes was obtained by calculating the number of water molecules in the firstsolvent shell surrounding a secondary structure (e.g., the �2 helix) or a specificresidue. The contact distances between C-alpha carbons were used to identifyinteractions between regions or differences between the clades. A 10-Å cutoffbetween C-alpha carbons was used as a measure for contact.
Construction of chimeric Envs. PCR amplification and cloning of the donorand recipient Envs from pairs 55 and 135 have been described previously (18).The env genes are in the cytomegalovirus-driven expression vector pCR3.1 (In-vitrogen), which is used to generate viral pseudotypes. Each of the donor �2 helixdomains shown in Fig. 3A was inserted into the corresponding recipient Env byusing a strategy similar to that described previously (66). A DNA duplex corre-sponding to each donor �2 helix was synthesized as a double-stranded oligonu-cleotide by IDT Technologies (Coralville, IA) and gel purified to ensure thateach fragment was of the correct length. These fragments were each blunt endligated to an �8-kb PCR amplicon corresponding to the recipient Env, excludingthe �2 helix and including the pCR3.1 vector sequences. The single-strandedDNA sequences corresponding to the 135 F and 55 M donor �2 helices (HXB2nucleotides [nt] 7227 to 7279) were as follows: for pair 135, F42a (5�-AAAAAGCTATGGAATAACACTTTAGACGGGATAAAGGAAAAATTAGCAAGATAC-3�), F48a (5�-AAACAAAATTGGACTAAAACTTTAGAAGAGGTAAAGACAGAATTAAGAAAATAC-3�), and F67a (5�-AGACAGAAATGGAATAGCACTTTAGACGGGATAAAGGAAAAATTAGCAAGATAC-3�); and forpair 55, M7a (5�-GAACAAAAATGGAGTACAACTTTAAAAAGGGTAGAGAAAAAATTAAAAGAGCAC-3�), M4.1 (5�-GGAAAAGAATGGAATACAACTTTAACAAGGGTAAAGGAAAGATTAAAAAAGCAC-3�), and M2.13(5�-AGAAAAAAATGGAATATAACTTTAGAAAGGGTAAAGGAAAGATTAAAAGAGCAC-3�).
For PCR amplification of the recipient Env (minus the helix) and pCR3.1vector sequences, 5�-phosphorylated primers were synthesized and gel purifiedby IDT Technologies. Primer sequences for amplification of the recipient Envbackbone (plus vector sequences) and their HXB2 locations were as follows:for pair 135, forward primer 5�-P-CTTCCCTGATAAAATAATAAAATTT-3�
(HXB2 nt 7280 to 7304) and reverse primer 5�-P-ACTAATGTTACAATGTGCTTGTCT-3� (HXB2 nt 7203 to 7226), and for pair 55, forward primer 5�-P-TTCCCTAATAAAATAATAAAATTT-3� (HXB2 nt 7281 to 7304) and reverseprimer 5�-P-ACTAATGTTACAATGTGCTTGTCT-3� (HXB2 nt 7203 to 7226).
PCR amplification conditions for the recipient Env backbones were as follows:1 cycle of 95°C for 3 min; 35 cycles of 95°C for 1 min, 62°C for 30 s, and 72°C for10 min; 1 cycle of 72°C for 15 min; and storage at 4°C. Pfu Turbo DNApolymerase (Stratagene) was used to generate a blunt-ended PCR amplicon,which was gel purified from an agarose gel by using a QIAquick gel extraction kit(QIAGEN) prior to ligation. The 25-l PCR mix contained 50 ng of each primer,10 ng of the plasmid template, 2.5 mM MgCl2, a 0.2 mM concentration of eachdeoxynucleoside triphosphate, and 1 reaction buffer. Each synthesized �2 helixDNA fragment was ligated to the purified recipient env backbone by using T4DNA ligase (5 U/l; Roche) at 4°C overnight. A portion of the ligation reactionmix was transformed into maximum efficiency XL2-Blue supercompetent cells(1 109 CFU/g DNA; Stratagene). The entire transformation mix was platedonto LB-ampicillin agar plates, generally resulting in �50 colonies per ligationreaction. These colonies were screened by PCR to identify colonies in which thefragments ligated together in the correct orientation, using forward primer EnvAand a reverse primer that anneals to the 3� end of the helix. The colony screeningprimers were as follows: forward, 5�-CAGCACAGTACAATGTACACATGGAA-3� (HXB2 nt 6950 to 6975); and reverse, 5�-TTCCTTTATCCCGTCTAAAGT-3� (HXB2 nt 7254 to 7274) for 135 F42a or 135 F67a �2 helix screening,5�-TGTCTTTACCTCTTCTAAAGT-3� (HXB2 nt 7254 to 7274) for 135 F48a�2 helix screening, 5�-TTTTAATCTTTCCTTTACCCT-3� (HXB2 nt 7254 to7274) for 55 M4.1 or 55 M2.13 �2 helix screening, or 5�-TTTTAATTTTTTCTCTACCCT-3� (HXB2 nt 7254 to 7274) for 55 M7a �2 helix screening. Coloniesthat were positive by PCR screening were inoculated into LB-ampicillin over-night cultures, and the plasmids were prepared using a QIAprep Spin miniprepkit (QIAGEN). Plasmids were then screened by (i) restriction digestion, (ii)biological function assay (described below), and (iii) nucleotide sequencing ofthe �2 helix and flanking regions to confirm that they contained the correctsequences.
Neutralization assay. All neutralization assays were performed using apseudotyped virion assay as described previously (17, 18, 48, 66, 74). In this assay,the NAb activity of plasmas collected contemporaneously with the Envs fromdonors 135 F and 55 M was evaluated against virions pseudotyped with recipientEnvs containing chimeric �2 helix regions in parallel with the parental donor andrecipient Envs. For the present study, to evaluate the neutralization sensitivitiesof the donor, recipient, and chimeric Envs together, it was necessary to conservelimited amounts of donor plasma. Therefore, the highest plasma dilution testedwas 1:100 for donor 55 M and 1:20 for donor 135 F.
RESULTS
Sequence divergence that occurs during chronic infection iscorrelated with NAb resistance. In our previous study of het-erosexual transmission, phylogenetic trees of gp120 V1-to-V4sequences from donor-recipient pairs were created using MLto investigate the phylogenetic relationships between donorand recipient sequences (18). In the present study, we calcu-lated the BL from each patient’s ancestral node for 48 donorand 27 recipient Env sequences from five transmission pairs forwhich neutralization sensitivity had also been evaluated (18).Informative sites in regions lacking insertions and deletionsdrove the BL, since most sequences from the V1V2 and V4hypervariable domains were excluded from the tree. To inves-tigate the relationship between sequence evolution outside thehypervariable domains and neutralization resistance, Kendall’stau rank correlation test was applied to determine whether BLwas significantly correlated with sensitivity to NAb in the in-fecting partner’s (donor) plasma or in a heterologous plasmapool from subtype C-infected subjects in Zambia (Fig. 1A andB). BLs for sequences from the chronically infected donorsranged from 0.002 to 0.138 but were significantly shorter forthe newly infected recipients (ranging from 0.000 to 0.005; P �0.001). When the recipient and donor BLs from their respec-
5660 RONG ET AL. J. VIROL.
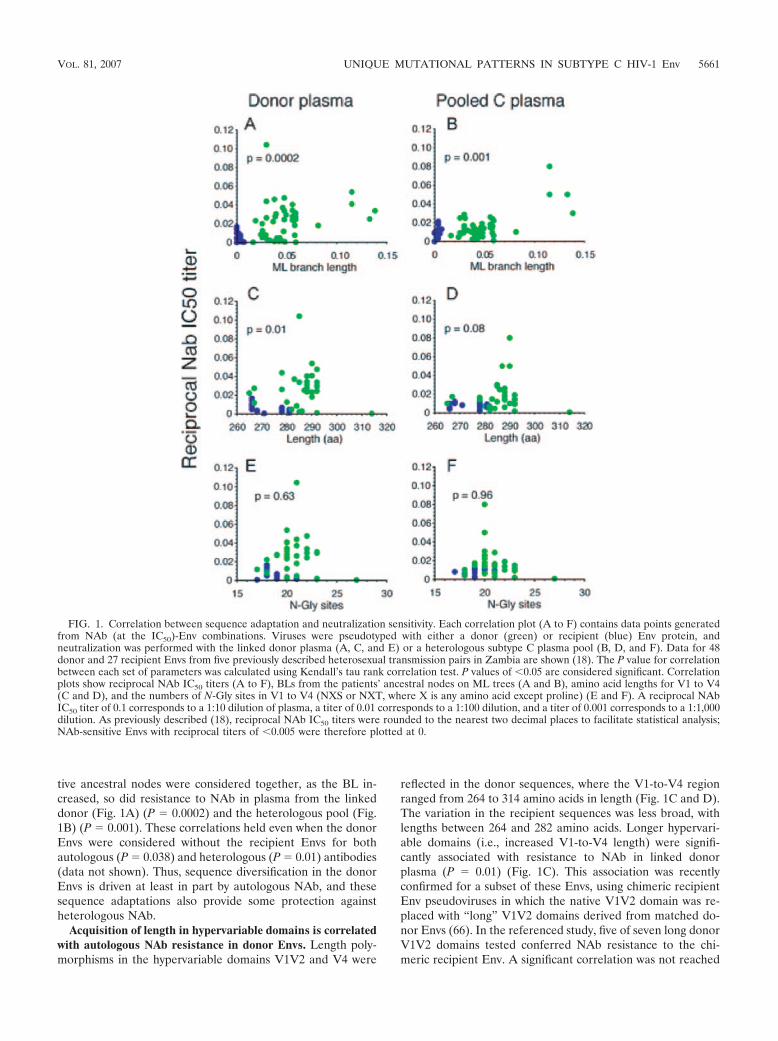
tive ancestral nodes were considered together, as the BL in-creased, so did resistance to NAb in plasma from the linkeddonor (Fig. 1A) (P � 0.0002) and the heterologous pool (Fig.1B) (P � 0.001). These correlations held even when the donorEnvs were considered without the recipient Envs for bothautologous (P � 0.038) and heterologous (P � 0.01) antibodies(data not shown). Thus, sequence diversification in the donorEnvs is driven at least in part by autologous NAb, and thesesequence adaptations also provide some protection againstheterologous NAb.
Acquisition of length in hypervariable domains is correlatedwith autologous NAb resistance in donor Envs. Length poly-morphisms in the hypervariable domains V1V2 and V4 were
reflected in the donor sequences, where the V1-to-V4 regionranged from 264 to 314 amino acids in length (Fig. 1C and D).The variation in the recipient sequences was less broad, withlengths between 264 and 282 amino acids. Longer hypervari-able domains (i.e., increased V1-to-V4 length) were signifi-cantly associated with resistance to NAb in linked donorplasma (P � 0.01) (Fig. 1C). This association was recentlyconfirmed for a subset of these Envs, using chimeric recipientEnv pseudoviruses in which the native V1V2 domain was re-placed with “long” V1V2 domains derived from matched do-nor Envs (66). In the referenced study, five of seven long donorV1V2 domains tested conferred NAb resistance to the chi-meric recipient Env. A significant correlation was not reached
FIG. 1. Correlation between sequence adaptation and neutralization sensitivity. Each correlation plot (A to F) contains data points generatedfrom NAb (at the IC50)-Env combinations. Viruses were pseudotyped with either a donor (green) or recipient (blue) Env protein, andneutralization was performed with the linked donor plasma (A, C, and E) or a heterologous subtype C plasma pool (B, D, and F). Data for 48donor and 27 recipient Envs from five previously described heterosexual transmission pairs in Zambia are shown (18). The P value for correlationbetween each set of parameters was calculated using Kendall’s tau rank correlation test. P values of �0.05 are considered significant. Correlationplots show reciprocal NAb IC50 titers (A to F), BLs from the patients’ ancestral nodes on ML trees (A and B), amino acid lengths for V1 to V4(C and D), and the numbers of N-Gly sites in V1 to V4 (NXS or NXT, where X is any amino acid except proline) (E and F). A reciprocal NAbIC50 titer of 0.1 corresponds to a 1:10 dilution of plasma, a titer of 0.01 corresponds to a 1:100 dilution, and a titer of 0.001 corresponds to a 1:1,000dilution. As previously described (18), reciprocal NAb IC50 titers were rounded to the nearest two decimal places to facilitate statistical analysis;NAb-sensitive Envs with reciprocal titers of �0.005 were therefore plotted at 0.
VOL. 81, 2007 UNIQUE MUTATIONAL PATTERNS IN SUBTYPE C HIV-1 Env 5661
between V1-to-V4 length and neutralization by heterologouspooled plasma (Fig. 1D), but there was a trend in this direction(P � 0.08). Thus, hypervariable domain insertions could havethe strongest effects on masking strain-specific epitopes, butsome protection against cross-neutralization could also be af-forded.
Accumulation of N-Gly sites is not correlated with NAbresistance. Tandem repeat sequences inserted within thelonger V1V2 and V4 domains often encoded N-Gly sites (66;data not shown), and in four of the five donors, the number ofN-Gly sites was correlated with V1-to-V4 length, using aSpearman rank correlation test (data not shown). Neverthe-less, the number of N-Gly sites was not significantly correlatedwith neutralization by either type of plasma (Fig. 1E and F).This finding could reflect a lack of statistical power for thenumber of samples analyzed, given the relatively small windowof N-Gly sites (17 to 23 sites for all but one Env). Alternatively,the relative positions and packing of the glycans could be moreimportant than net gains or losses for neutralization escape(74).
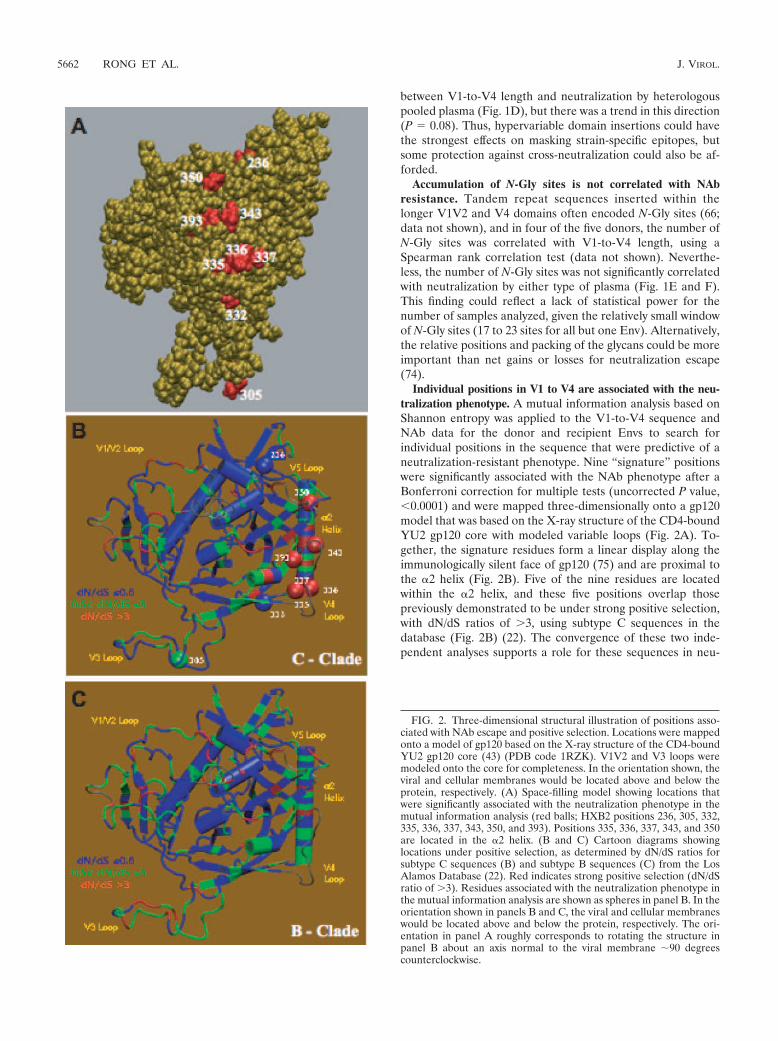
Individual positions in V1 to V4 are associated with the neu-tralization phenotype. A mutual information analysis based onShannon entropy was applied to the V1-to-V4 sequence andNAb data for the donor and recipient Envs to search forindividual positions in the sequence that were predictive of aneutralization-resistant phenotype. Nine “signature” positionswere significantly associated with the NAb phenotype after aBonferroni correction for multiple tests (uncorrected P value,�0.0001) and were mapped three-dimensionally onto a gp120model that was based on the X-ray structure of the CD4-boundYU2 gp120 core with modeled variable loops (Fig. 2A). To-gether, the signature residues form a linear display along theimmunologically silent face of gp120 (75) and are proximal tothe �2 helix (Fig. 2B). Five of the nine residues are locatedwithin the �2 helix, and these five positions overlap thosepreviously demonstrated to be under strong positive selection,with dN/dS ratios of �3, using subtype C sequences in thedatabase (Fig. 2B) (22). The convergence of these two inde-pendent analyses supports a role for these sequences in neu-
FIG. 2. Three-dimensional structural illustration of positions asso-ciated with NAb escape and positive selection. Locations were mappedonto a model of gp120 based on the X-ray structure of the CD4-boundYU2 gp120 core (43) (PDB code 1RZK). V1V2 and V3 loops weremodeled onto the core for completeness. In the orientation shown, theviral and cellular membranes would be located above and below theprotein, respectively. (A) Space-filling model showing locations thatwere significantly associated with the neutralization phenotype in themutual information analysis (red balls; HXB2 positions 236, 305, 332,335, 336, 337, 343, 350, and 393). Positions 335, 336, 337, 343, and 350are located in the �2 helix. (B and C) Cartoon diagrams showinglocations under positive selection, as determined by dN/dS ratios forsubtype C sequences (B) and subtype B sequences (C) from the LosAlamos Database (22). Red indicates strong positive selection (dN/dSratio of �3). Residues associated with the neutralization phenotype inthe mutual information analysis are shown as spheres in panel B. In theorientation shown in panels B and C, the viral and cellular membraneswould be located above and below the protein, respectively. The ori-entation in panel A roughly corresponds to rotating the structure inpanel B about an axis normal to the viral membrane �90 degreescounterclockwise.
5662 RONG ET AL. J. VIROL.
tralization escape. Also, since the database sequences weredrawn from a broader sampling that did not include the Envsstudied here, these findings could be generally relevant tosubtype C viruses and not limited to those circulating in Zam-bia. Importantly, the positions of the NAb escape signature
residues are not subject to the same degree of positive selec-tion in subtype B sequences, where the highest dN/dS ratiosare concentrated in V3 (Fig. 2C). Taken together, these datasuggest that substitutions in this region could be driven by theautologous NAb response and that changes in this region, in
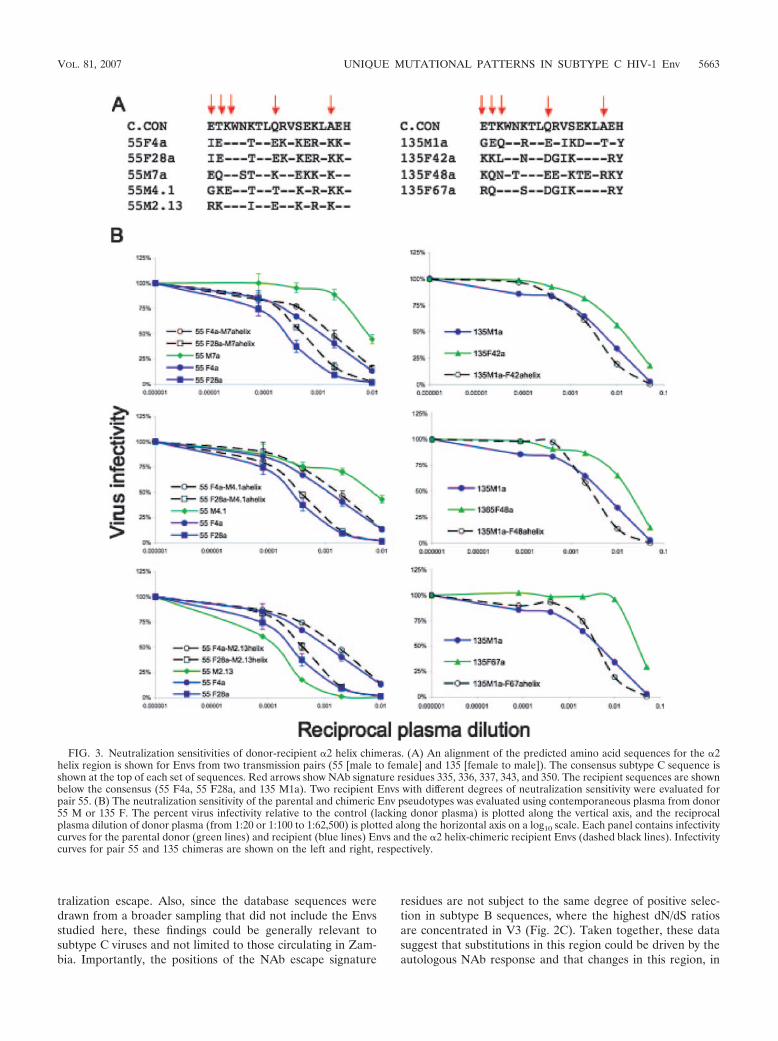
FIG. 3. Neutralization sensitivities of donor-recipient �2 helix chimeras. (A) An alignment of the predicted amino acid sequences for the �2helix region is shown for Envs from two transmission pairs (55 [male to female] and 135 [female to male]). The consensus subtype C sequence isshown at the top of each set of sequences. Red arrows show NAb signature residues 335, 336, 337, 343, and 350. The recipient sequences are shownbelow the consensus (55 F4a, 55 F28a, and 135 M1a). Two recipient Envs with different degrees of neutralization sensitivity were evaluated forpair 55. (B) The neutralization sensitivity of the parental and chimeric Env pseudotypes was evaluated using contemporaneous plasma from donor55 M or 135 F. The percent virus infectivity relative to the control (lacking donor plasma) is plotted along the vertical axis, and the reciprocalplasma dilution of donor plasma (from 1:20 or 1:100 to 1:62,500) is plotted along the horizontal axis on a log10 scale. Each panel contains infectivitycurves for the parental donor (green lines) and recipient (blue lines) Envs and the �2 helix-chimeric recipient Envs (dashed black lines). Infectivitycurves for pair 55 and 135 chimeras are shown on the left and right, respectively.
VOL. 81, 2007 UNIQUE MUTATIONAL PATTERNS IN SUBTYPE C HIV-1 Env 5663

conjunction with alterations in the hypervariable domains,could contribute to neutralization escape during subtype Cinfection.
Exchange of the �2 helix does not directly alter the neutral-ization phenotype. The �2 helix is located on the outer domainof gp120, and as an amphipathic helix, the outer face could bepartially exposed (Fig. 2A). Based on the mutual informationanalysis and dN/dS ratios from the database analysis, five po-sitions on the outer surface of this structure appear to be understrong positive selection in subtype C viruses. To investigatewhether determinants of neutralization resistance are actuallycontained within the �2 helix, we selected Envs from transmis-sion pairs 55 (male to female) and 135 (female to male) forfurther study (18). In pair 55, only 6 of the 18 positions in thehelix were completely conserved (Fig. 3A), and these werelocated on the inner surface facing the gp120 core (22a). Res-idues in all but one of the five NAb signature positions wereheterogeneous in the pair 55 Envs (Fig. 3A). Likewise, for pair135, the five internal positions of the helix were completelyconserved, and all NAb signature residues were heterogeneous(Fig. 3A). Chimeric Envs were created in which the entire helixof a recipient Env was replaced with the corresponding se-quences from three different donor Envs. In the case of pair 55,two recipient Envs with different degrees of neutralizationsensitivity determined by a charged residue in V2 were used(66). The entire �2 helix domain was exchanged instead ofperforming targeted site-directed mutagenesis due to the lim-ited quantities of donor plasma available and the extensivesequence heterogeneity in this region (Fig. 3A). Despite thestrong association between neutralization sensitivity and theamino acid residues in this region that emerged in the mutualinformation analysis, exchange of this region resulted in onlyslight changes in NAb sensitivity between the recipient andchimeric Envs for both transmission pairs (Fig. 3B). The re-ciprocal constructs were created and evaluated for pair 135(recipient helix into donor Envs), and again, only minimalchanges in neutralization phenotype were observed (data notshown). These results argue that despite the strong selectivepressure on the �2 helix and its association with NAb resis-tance, this structure does not directly modulate resistance toautologous NAb.
DISCUSSION
Longitudinal studies of humans and animal models havedemonstrated that the V1V2 and V4 regions are susceptible toinsertions, deletions, and alterations in the pattern of glycosyl-ation during chronic infection of HIV-1-infected individualsand experimentally SIV-infected macaques (8, 10, 11, 13, 34,55, 61, 62, 67, 71). Acquisition of length in V1V2 and/or V4 inthe current study was associated with decreased sensitivity toneutralization by linked donor plasma, but less so with heter-ologous plasma. Changes in this region may therefore bedriven by escape from NAb that target mainly strain-specificepitopes. Using a V1V2 domain exchange approach, we haveconfirmed that long V1V2 domains from chronically infecteddonors frequently mask determinants of sensitivity to autolo-gous NAb (66). The effect of V1V2, however, was highly de-pendent on the Env background. We also observed cases inwhich donor Envs had acquired NAb resistance to autologous
neutralization independently of V1V2, consistent with thepresence of neutralization determinants that are not containedwithin this domain.
Increasing sequence divergence, as measured by BL on MLtrees, showed a strong association with decreased sensitivity toneutralization by donor plasma and a heterologous plasmapool. This result suggests that point mutations in regions out-side the hypervariable domains V1V2 and V4, which wereexcluded from the trees, facilitate escape from antibodies thattarget some cross-neutralizing epitopes and is consistent withthe correlation observed previously between the IC50 titers forneutralization by donor and pooled plasmas (18) and V1V2-independent pathways to NAb resistance (66). Despite theinherent limitations, given that the analysis was based on mul-tiple sequences from five transmission pairs, a striking rela-tionship between signature sites was apparent, in that all nineneutralization “signature” residues identified in the mutualinformation analysis cluster together on the outer face of thegp120 molecule, nestled among glycans that were resolved onthe unliganded SIV gp120 structure (Fig. 4A and B). Variationin one residue identified in the mutual information analysis,i.e., residue 236 (based on HXB2 numbering) in C2, can alterglycosylation at N234, and its effect appeared to differ amongthe transmission pairs. For example, the presence of a glycan atN234 was associated with neutralization resistance in pair 135,while the loss of the same glycan was associated with resistancein pair 53; this sort of patient-specific gain or loss of a glycanillustrates why there may be no discernible association betweenthe net number of N-Gly sites and the resistance phenotype.Notably, glycosylation at the same position was previously as-sociated with cumulative escape from autologous neutraliza-tion in a subtype B-infected subject (74).
One potentially complicating factor for the mutual informa-tion analysis used here is that the recipient Envs were alwaysNAb sensitive and all NAb-resistant Envs were from the do-nors (18). It is therefore possible that the associations detectedby the mutual information analysis reflect positions in V1 toV4 that track with transmission or establishment of infection.This remains a formal possibility, as the transmission bottle-neck would constrict variation at all sites and thus could pro-duce an association with the neutralization phenotype simplydue to the overall sensitivity of the recipient Envs. However,the coincidence of high mutual information scores and highlevels of positive selection (high dN/dS ratios) at the popula-tion level for subtype C viruses for the same sites supports theirrole in neutralization escape. Moreover, an independent mu-tual information analysis was performed on 515 V1-to-V4 se-quences from an additional 13 transmission pairs from thesame cohort (including 502 donor sequences and 13 “consen-sus” sequences from each recipient) in the absence of NAbdata. In this analysis, four residues (two located in V2, one in�2, and one in V4) showed a trend of association with trans-mission status (P. Hawkins et al., unpublished data), and noneof these overlapped with the nine residues shown here to trackwith NAb resistance. Thus, the positions we have identified inthe current study as being associated with neutralization resis-tance appear to be independent of transmission status, giventhe sample size under consideration.
In the trimer model of gp120, based on the structure ofHXB2 (45), the 18-residue �2 helix is located on the outer
5664 RONG ET AL. J. VIROL.
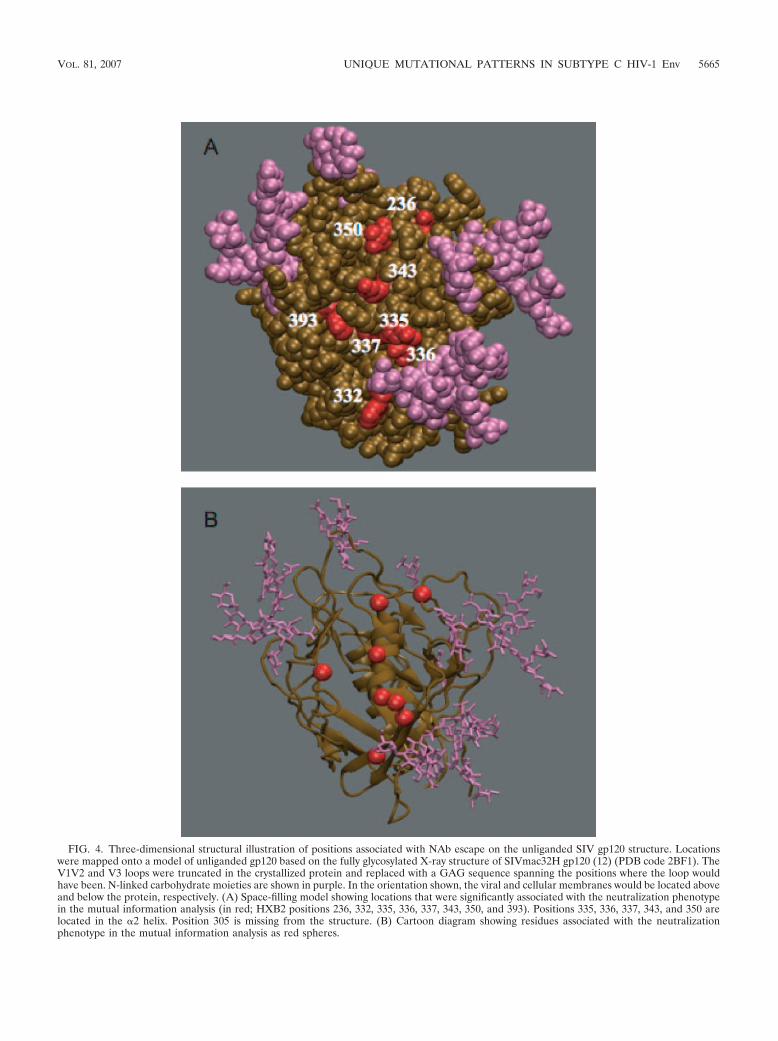
FIG. 4. Three-dimensional structural illustration of positions associated with NAb escape on the unliganded SIV gp120 structure. Locationswere mapped onto a model of unliganded gp120 based on the fully glycosylated X-ray structure of SIVmac32H gp120 (12) (PDB code 2BF1). TheV1V2 and V3 loops were truncated in the crystallized protein and replaced with a GAG sequence spanning the positions where the loop wouldhave been. N-linked carbohydrate moieties are shown in purple. In the orientation shown, the viral and cellular membranes would be located aboveand below the protein, respectively. (A) Space-filling model showing locations that were significantly associated with the neutralization phenotypein the mutual information analysis (in red; HXB2 positions 236, 332, 335, 336, 337, 343, 350, and 393). Positions 335, 336, 337, 343, and 350 arelocated in the �2 helix. Position 305 is missing from the structure. (B) Cartoon diagram showing residues associated with the neutralizationphenotype in the mutual information analysis as red spheres.
VOL. 81, 2007 UNIQUE MUTATIONAL PATTERNS IN SUBTYPE C HIV-1 Env 5665

surface of gp120. Even though this model is based on CD4-bound gp120, a comparison of liganded and unliganded gp120structures does not indicate major changes in the outer domainupon CD4 binding (12). Five of the nine NAb signature resi-dues were located within the �2 helix (Fig. 2B), and the sameresidues were under strong positive selection in subtype Cdatabase sequences (22); the coincidence of the two indepen-dent results raised the interesting hypothesis that substitutionsin this region could be related to antibody escape. Neverthe-less, exchange of the �2 helix between donor and recipientEnvs did not significantly alter the neutralization sensitivity ofthe related recipient Env, suggesting that it alone does notcontain direct determinants of NAb resistance. It is not sur-prising that an exchange of the �2 helix failed to produce directchanges in patterns of neutralization resistance, since this re-gion of Env is likely protected from antibody recognition by theglycan shield and thus, in itself, is unlikely to represent aneutralization epitope (74). More likely, this region of the Envglycoprotein is responsible for contributing to the three-dimen-sional structure of the protomer and the quaternary structureof the trimer that confers neutralization escape by mechanismsin addition to epitope variation (e.g., epitope masking, sterichindrance, conformation energetics, and glycan packing) (42,74). Molecular dynamics simulation studies provided evidencefor an extensive interaction between the �2 helix and a flexibleV4 loop in subtype C, with certain residues in the helix asso-ciated with restricting V4 loop length (22a). The nonpolar faceof the �2 helix is predicted to interact with structural domains�10, �11, �14, and �24, which could help to stabilize the gp120core structure. It is therefore possible that to see a phenotypiceffect of the �2 helix on neutralization sensitivity, other do-mains must be present. Perhaps the outer face of the �2 helixcompensates structurally for the adaptations that are directlyinvolved in NAb escape of subtype C viruses. If so, mutationsin the helix would track with a resistant phenotype withoutphysically defining or altering an epitope.
Sequence and structural studies revealed at least two majordifferences in the �2 helix between subtypes B and C. Eventhough the helix of subtype C exhibits high sequence entropy,the amphipathicity of the helix is preserved to a higher degreethan that of subtype B (22a). The helix of subtype C is alsomore solvated than that of subtype B (S. Gnanakaran, unpub-lished data), with high-sequence-entropy positions occurring atthe solvent-exposed face. All five signature residues identifiedhere occupy solvent-exposed positions of the helix, while res-idues that face toward the core are well conserved (22a). Thus,in subtype C viruses, the �2 helix may act as a flexible interfaceto maintain the integrity between NAb escape mutations oc-curring on the outer domain and the more highly conservedgp120 inner core. Alternatively, these mutations could accom-modate escape mechanisms occurring in the hypervariable re-gions. In subtype C, the N-terminal half of the helix interactsextensively with the V4 loop, while the C-terminal half inter-acts with the V5 domain.
Finally, mutations at some of the signature positions couldaffect the local stability of the �2 helix. Alpha helices representa common secondary structural motif in proteins and are sta-bilized by intramolecular hydrogen bonding between the i andi � 4 residues on parallel turns of the helix. However, the firstfour and last four residues of an � helix lack this intramolec-
ular hydrogen bonding and are often stabilized by alternativehydrogen bonding partners and hydrophobic interactions. Thisleads to differences in the propensities of amino acids found inthe N-terminal and C-terminal capping motifs. Several system-atic studies on the positional frequency of each of the aminoacids found in helices in protein structures show statisticaldeviations from a random distribution (1, 19, 41, 65). Forexample, asparagine has a higher preference of occurring atthe N-terminal cap position, whereas the structurally similarglutamic acid does not. Further studies have established arelationship between capping motifs and the stability of an �helix. Interestingly, five of the nine signature positions identi-fied here have the potential to play a structural role as cappingresidues of the �2 helix. Four of them coincide with the N-terminal capping positions, and one coincides with a C-termi-nal capping position in the �2 helix. Furthermore, the first turnof the N-terminal capping sequence is located adjacent to theantiparallel beta strand just downstream from the V3 domain.Thus, any local unfolding of the N-terminal region of the �2helix could potentially impact the projection of the V3 domainout from the core. Thus, the �2 helix of gp120 appears to be anovel and important source of structural adaptation duringsubtype C infection, and its structural role in the context of theentire gp120 molecule needs to be explored further.
The present study combines statistical, biological, and mo-lecular dynamics simulation analyses to elucidate the poorlyunderstood pathways to neutralization resistance in subtype Cvirus infection. NAb are clearly a strong evolutionary forceduring chronic infection with subtype C HIV-1, driving se-quence divergence in structural domains and also dramaticlength changes in the hypervariable regions of Env. Moreover,mutations in a purely structural domain (in a region that his-torically has been considered well conserved) track with NAbresistance, supporting a novel and potentially subtype-specificmechanism for neutralization escape. Further investigationswill be required to understand the selective forces directed atthe �2 helix and to determine why this region is differentiallytargeted depending on the viral subtype, as these questionshave direct implications for global vaccine design.
ACKNOWLEDGMENTS
We gratefully acknowledge the contributions of the staff, lab tech-nicians, participants, and project management group of the Lusakacohort. We thank Peter Kwong for providing us with the HXB2 trimercoordinates.
This work was supported by NIH grants R01-AI-58706 (C.D.) andR01-AI-51231 (E.H.), by The Bill & Melinda Gates Grand ChallengesProgram (G.M.S.), and by Yerkes National Primate Research Centerbase grant RR-00165, awarded by the National Center for ResearchResources of the National Institutes of Health.
REFERENCES
1. Aurora, R., and G. D. Rose. 1998. Helix capping. Protein Sci. 7:21–38.2. Ben-Naim, E., and A. S. Lapedes. 1999. Genetic correlations in mutation
processes. Phys. Rev. E 59:7000–7007.3. Berendsen, H. J. C., J. P. M. Postma, W. F. van Gunsteren, A. DiNola, and
J. R. Haak. 1984. Molecular dynamics with coupling to an external bath.J. Chem. Phys. 81:3684–3690.
4. Binley, J. M., T. Wrin, B. Korber, M. B. Zwick, M. Wang, C. Chappey, G.Stiegler, R. Kunert, S. Zolla-Pazner, H. Katinger, C. J. Petropoulos, andD. R. Burton. 2004. Comprehensive cross-clade neutralization analysis of apanel of anti-human immunodeficiency virus type 1 monoclonal antibodies.J. Virol. 78:13232–13252.
5. Blay, W. M., S. Gnanakaran, B. Foley, N. A. Doria-Rose, B. T. Korber, andN. L. Haigwood. 2006. Consistent patterns of change during the divergence
5666 RONG ET AL. J. VIROL.

of human immunodeficiency virus type 1 envelope from that of the inocu-lated virus in simian/human immunodeficiency virus-infected macaques.J. Virol. 80:999–1014.
6. Bouma, P., M. Leavitt, P. F. Zhang, I. A. Sidorov, D. S. Dimitrov, and G. V.Quinnan, Jr. 2003. Multiple interactions across the surface of the gp120 corestructure determine the global neutralization resistance phenotype of humanimmunodeficiency virus type 1. J. Virol. 77:8061–8071.
7. Bures, R., L. Morris, C. Williamson, G. Ramjee, M. Deers, S. A. Fiscus, S.Abdool-Karim, and D. C. Montefiori. 2002. Regional clustering of sharedneutralization determinants on primary isolates of clade C human immuno-deficiency virus type 1 from South Africa. J. Virol. 76:2233–2244.
8. Burns, D. P., and R. C. Desrosiers. 1994. Envelope sequence variation,neutralizing antibodies, and primate lentivirus persistence. Curr. Top. Mi-crobiol. Immunol. 188:185–219.
9. Case, D. A., D. A. Pearlman, J. W. Caldwell, T. E. Cheatham III, W. S. Ross,C. L. Simmerling, T. A. Darden, K. M. Merz, R. V. Stanton, A. L. Cheng, J. J.Vincent, M. Crowley, V. Tsui, R. J. Radmer, Y. Duan, J. Pitera, G. L. Seibel,U. C. Singh, P. K. Weiner, and P. A. Kollman. 1999. Amber 6 software.University of California, San Francisco.
10. Chackerian, B., W. R. Morton, and J. Overbaugh. 1994. Persistence ofsimian immunodeficiency virus Mne variants upon transmission. J. Virol.68:4080–4085.
11. Chackerian, B., L. M. Rudensey, and J. Overbaugh. 1997. Specific N-linkedand O-linked glycosylation modifications in the envelope V1 domain ofsimian immunodeficiency virus variants that evolve in the host alter recog-nition by neutralizing antibodies. J. Virol. 71:7719–7727.
12. Chen, B., E. M. Vogan, H. Gong, J. J. Skehel, D. C. Wiley, and S. C.Harrison. 2005. Structure of an unliganded simian immunodeficiency virusgp120 core. Nature 433:834–841.
13. Cheng-Mayer, C., A. Brown, J. Harouse, P. A. Luciw, and A. J. Mayer. 1999.Selection for neutralization resistance of the simian/human immunodefi-ciency virus SHIVSF33A variant in vivo by virtue of sequence changes in theextracellular envelope glycoprotein that modify N-linked glycosylation. J. Vi-rol. 73:5294–5300.
14. Choisy, M., C. H. Woelk, J. F. Guegan, and D. L. Robertson. 2004. Com-parative study of adaptive molecular evolution in different human immuno-deficiency virus groups and subtypes. J. Virol. 78:1962–1970.
15. Cornell, W. D., P. Cieplak, C. I. Bayly, I. R. Gould, K. M. Merz, D. M.Ferguson, D. C. Spellmeyer, T. Fox, J. W. Caldwell, and P. A. Kollman. 1995.A second generation force-field for the simulation of proteins; nucleic acids;and organic molecules. J. Am. Chem. Soc. 117:5179–5197.
16. Darden, T., D. York, and L. Pedersen. 1993. Particle mesh Ewald: an N log(N) method for Ewald sums in large systems. J. Chem. Phys. 98:10089–10092.
17. Decker, J. M., F. Bibollet-Ruche, X. P. Wei, S. Wang, D. N. Levy, W. Wang,E. Delaporte, M. Peeters, C. A. Derdeyn, S. Allen, E. Hunter, M. S. Saag,J. A. Hoxie, B. H. Hahn, P. D. Kwong, J. E. Robinson, and G. M. Shaw. 2005.Antigenic conservation and immunogenicity of the HIV coreceptor bindingsite. J. Exp. Med. 201:1–14.
18. Derdeyn, C. A., J. M. Decker, F. Bibollet-Ruche, J. L. Mokili, M. Muldoon,S. A. Denham, M. L. Heil, F. Kasolo, R. Musonda, B. H. Hahn, G. M. Shaw,B. T. Korber, S. Allen, and E. Hunter. 2004. Envelope-constrained neutral-ization-sensitive HIV-1 after heterosexual transmission. Science 303:2019–2022.
19. Engel, D. E., and W. F. DeGrado. 2004. Amino acid propensities are posi-tion-dependent throughout the length of alpha-helices. J. Mol. Biol. 337:1195–1205.
20. Etemad-Moghadam, B., G. B. Karlsson, M. Halloran, Y. Sun, D. Schenten,M. Fernandes, N. L. Letvin, and J. Sodroski. 1998. Characterization ofsimian-human immunodeficiency virus envelope glycoprotein epitopes rec-ognized by neutralizing antibodies from infected monkeys. J. Virol. 72:8437–8445.
21. Fung, M. S., C. R. Sun, W. L. Gordon, R. S. Liou, T. W. Chang, W. N. Sun,E. S. Daar, and D. D. Ho. 1992. Identification and characterization of aneutralization site within the second variable region of human immunode-ficiency virus type 1 gp120. J. Virol. 66:848–856.
22. Gaschen, B., J. Taylor, K. Yusim, B. Foley, F. Gao, D. Lang, V. Novitsky, B.Haynes, B. H. Hahn, T. Bhattacharya, and B. Korber. 2002. Diversity con-siderations in HIV-1 vaccine selection. Science 296:2354–2360.
22a.Gnanakaran, S., D. Lang, M. Daniels, T. Bhattacharya, C. A. Derdeyn, andB. Korber. 2007. Clade-specific differences between human immunodefi-ciency virus type 1 clades B and C: diversity and correlations in C3-V4regions of gp120. J. Virol. 81:4886–4891.
23. Gorny, M. K., J. P. Moore, A. J. Conley, S. Karwowska, J. Sodroski, C.Williams, S. Burda, L. J. Boots, and S. Zolla-Pazner. 1994. Human anti-V2monoclonal antibody that neutralizes primary but not laboratory isolates ofhuman immunodeficiency virus type 1. J. Virol. 68:8312–8320.
24. Gorny, M. K., K. Revesz, C. Williams, B. Volsky, M. K. Louder, C. A.Anyangwe, C. Krachmarov, S. C. Kayman, A. Pinter, A. Nadas, P. N. Nyambi,J. R. Mascola, and S. Zolla-Pazner. 2004. The V3 loop is accessible on thesurface of most human immunodeficiency virus type 1 primary isolates andserves as a neutralization epitope. J. Virol. 78:2394–2404.
25. Gorny, M. K., and S. Zolla-Pazner (ed.). 2003. Human monoclonal antibod-
ies that neutralize HIV-1. Los Alamos National Laboratory, TheoreticalBiology and Biophysics, Los Alamos, NM.
26. Gram, G. J., A. Hemming, A. Bolmstedt, B. Jansson, S. Olofsson, L. Aker-blom, J. O. Nielsen, and J. E. Hansen. 1994. Identification of an N-linkedglycan in the V1-loop of HIV-1 gp120 influencing neutralization by anti-V3antibodies and soluble CD4. Arch. Virol. 139:253–261.
27. Gray, E. S., T. Meyers, G. Gray, D. C. Montefiori, and L. Morris. 2006.Insensitivity of paediatric HIV-1 subtype C viruses to broadly neutralisingmonoclonal antibodies raised against subtype B. PLoS Med. 3:e255.
28. Ho, D. D., M. S. Fung, Y. Z. Cao, X. L. Li, C. Sun, T. W. Chang, and N. C.Sun. 1991. Another discontinuous epitope on glycoprotein gp120 that isimportant in human immunodeficiency virus type 1 neutralization is identi-fied by a monoclonal antibody. Proc. Natl. Acad. Sci. USA 88:8949–8952.
29. Humphrey, W., A. Dalke, and K. Schulten. 1996. VMD—Visual MolecularDynamics. J. Mol. Graphics 14:33–38.
30. Hunter, E. 1997. Viral entry and receptors, p. 71–121. In J. M. Coffin, S. H.Hughes, and H. E. Varmus (ed.), Retroviruses. Cold Spring Harbor Labo-ratory Press, Plainview, NY.
31. Jorgensen, W. L., J. Chandreshkar, J. D. Madura, R. W. Imprey, and M. L.Klein. 1983. Comparison of simple potential functions for simulating liquidwater. J. Chem. Phys. 79:926–935.
32. Kayman, S. C., Z. Wu, K. Revesz, H. Chen, R. Kopelman, and A. Pinter.1994. Presentation of native epitopes in the V1/V2 and V3 regions of humanimmunodeficiency virus type 1 gp120 by fusion glycoproteins containingisolated gp120 domains. J. Virol. 68:400–410.
33. Kinsey, N. E., M. G. Anderson, T. J. Unangst, S. V. Joag, O. Narayan, M. C.Zink, and J. E. Clements. 1996. Antigenic variation of SIV: mutations in V4alter the neutralization profile. Virology 221:14–21.
34. Kodama, T., K. Mori, T. Kawahara, D. J. Ringler, and R. C. Desrosiers.1993. Analysis of simian immunodeficiency virus sequence variation in tis-sues of rhesus macaques with simian AIDS. J. Virol. 67:6522–6534.
35. Kolchinsky, P., E. Kiprilov, P. Bartley, R. Rubinstein, and J. Sodroski. 2001.Loss of a single N-linked glycan allows CD4-independent human immuno-deficiency virus type 1 infection by altering the position of the gp120 V1/V2variable loops. J. Virol. 75:3435–3443.
36. Kolchinsky, P., E. Kiprilov, and J. Sodroski. 2001. Increased neutralizationsensitivity of CD4-independent human immunodeficiency virus variants.J. Virol. 75:2041–2050.
37. Korber, B. T., R. M. Farber, D. H. Wolpert, and A. S. Lapedes. 1993.Covariation of mutations in the V3 loop of human immunodeficiency virustype 1 envelope protein: an information theoretic analysis. Proc. Natl. Acad.Sci. USA 90:7176–7180.
38. Korber, B. T., K. MacInnes, R. F. Smith, and G. Myers. 1994. Mutationaltrends in V3 loop protein sequences observed in different genetic lineages ofhuman immunodeficiency virus type 1. J. Virol. 68:6730–6744.
39. Krachmarov, C., A. Pinter, W. J. Honnen, M. K. Gorny, P. N. Nyambi, S.Zolla-Pazner, and S. C. Kayman. 2005. Antibodies that are cross-reactive forhuman immunodeficiency virus type 1 clade A and clade B V3 domains arecommon in patient sera from Cameroon, but their neutralization activity isusually restricted by epitope masking. J. Virol. 79:780–790.
40. Krachmarov, C. P., W. J. Honnen, S. C. Kayman, M. K. Gorny, S. Zolla-Pazner, and A. Pinter. 2006. Factors determining the breadth and potency ofneutralization by V3-specific human monoclonal antibodies derived fromsubjects infected with clade A or clade B strains of human immunodeficiencyvirus type 1. J. Virol. 80:7127–7135.
41. Kumar, S., and M. Bansal. 1998. Dissecting alpha-helices: position-specificanalysis of alpha-helices in globular proteins. Proteins 31:460–476.
42. Kwong, P. D., M. L. Doyle, D. J. Casper, C. Cicala, S. A. Leavitt, S. Majeed,T. D. Steenbeke, M. Venturi, I. Chaiken, M. Fung, H. Katinger, P. W.Parren, J. Robinson, D. Van Ryk, L. Wang, D. R. Burton, E. Freire, R. Wyatt,J. Sodroski, W. A. Hendrickson, and J. Arthos. 2002. HIV-1 evades anti-body-mediated neutralization through conformational masking of receptor-binding sites. Nature 420:678–682.
43. Kwong, P. D., R. Wyatt, S. Majeed, J. Robinson, R. W. Sweet, J. Sodroski,and W. A. Hendrickson. 2000. Structures of HIV-1 gp120 envelope glyco-proteins from laboratory-adapted and primary isolates. Structure Fold Des.8:1329–1339.
44. Kwong, P. D., R. Wyatt, J. Robinson, R. W. Sweet, J. Sodroski, and W. A.Hendrickson. 1998. Structure of an HIV gp120 envelope glycoprotein incomplex with the CD4 receptor and a neutralizing human antibody. Nature393:648–659.
45. Kwong, P. D., R. Wyatt, Q. J. Sattentau, J. Sodroski, and W. A. Hendrickson.2000. Oligomeric modeling and electrostatic analysis of the gp120 envelopeglycoprotein of human immunodeficiency virus. J. Virol. 74:1961–1972.
46. LaRosa, G. J., J. P. Davide, K. Weinhold, J. A. Waterbury, A. T. Profy, J. A.Lewis, A. J. Langlois, G. R. Dreesman, R. N. Boswell, P. Shadduck, et al.1990. Conserved sequence and structural elements in the HIV-1 principalneutralizing determinant. Science 249:932–935.
47. Leitner, T., B. Foley, B. Hahn, P. Marx, F. McCutchan, J. Mellors, S.Wolinksy, and B. Korber. 2003. HIV sequence compendium. The Theoret-ical Biology and Biophysics Group, Los Alamos National Laboratory, LosAlamos, NM.
VOL. 81, 2007 UNIQUE MUTATIONAL PATTERNS IN SUBTYPE C HIV-1 Env 5667

48. Li, B., J. M. Decker, R. W. Johnson, F. Bibollet-Ruche, X. Wei, J. Mulenga,S. Allen, E. Hunter, B. H. Hahn, G. M. Shaw, J. L. Blackwell, and C. A.Derdeyn. 2006. Evidence for potent autologous neutralizing antibody titersand compact envelopes in early infection with subtype C human immuno-deficiency virus type 1. J. Virol. 80:5211–5218.
49. Losman, B., A. Bolmstedt, K. Schonning, A. Bjorndal, C. Westin, E. M.Fenyo, and S. Olofsson. 2001. Protection of neutralization epitopes in the V3loop of oligomeric human immunodeficiency virus type 1 glycoprotein 120 byN-linked oligosaccharides in the V1 region. AIDS Res. Hum. Retrovir.17:1067–1076.
50. Ly, A., and L. Stamatatos. 2000. V2 loop glycosylation of the human immu-nodeficiency virus type 1 SF162 envelope facilitates interaction of this pro-tein with CD4 and CCR5 receptors and protects the virus from neutraliza-tion by anti-V3 loop and anti-CD4 binding site antibodies. J. Virol. 74:6769–6776.
51. McCaffrey, R. A., C. Saunders, M. Hensel, and L. Stamatatos. 2004. N-linked glycosylation of the V3 loop and the immunologically silent face ofgp120 protects human immunodeficiency virus type 1 SF162 from neutral-ization by anti-gp120 and anti-gp41 antibodies. J. Virol. 78:3279–3295.
52. McCutchan, F. 2006. Global epidemiology of HIV. J. Med. Virol. 78:S7–S12.53. McKeating, J. A., C. Shotton, J. Cordell, S. Graham, P. Balfe, N. Sullivan,
M. Charles, M. Page, A. Bolmstedt, S. Olofsson, et al. 1993. Characterizationof neutralizing monoclonal antibodies to linear and conformation-dependentepitopes within the first and second variable domains of human immunode-ficiency virus type 1 gp120. J. Virol. 67:4932–4944.
54. Nabatov, A. A., G. Pollakis, T. Linnemann, A. Kliphius, M. I. Chalaby, andW. A. Paxton. 2004. Intrapatient alterations in the human immunodeficiencyvirus type 1 gp120 V1V2 and V3 regions differentially modulate coreceptorusage, virus inhibition by CC/CXC chemokines, soluble CD4, and the b12and 2G12 monoclonal antibodies. J. Virol. 78:524–530.
55. Palmer, C., P. Balfe, D. Fox, J. C. May, R. Frederiksson, E. M. Fenyo, andJ. A. McKeating. 1996. Functional characterization of the V1V2 region ofhuman immunodeficiency virus type 1. Virology 220:436–449.
56. Pearlman, D. A., D. A. Case, J. W. Caldwell, W. L. Ross, T. E. Cheatham,D. M. Ferguson, G. L. Seibel, U. C. Singh, P. K. Weiner, and P. A. Kollman.1995. AMBER 4.1 software. University of California, San Francisco.
57. Peeters, M. 2000. Recombinant HIV sequences: their role in the globalepidemic, p. 39–54. In F. B. Kuiken, B. Hahn, P. Marx, F. McCutchan, J.Mellors, J. Mullins, J. Sodroski, S. Wolinksy, and B. Korber (ed.), HIVsequence compendium 2000. Theoretical Biology and Biophysics Group, LosAlamos National Laboratory, Los Alamos, NM.
58. Pinter, A., W. J. Honnen, P. D’Agostino, M. K. Gorny, S. Zolla-Pazner, andS. C. Kayman. 2005. The C108g epitope in the V2 domain of gp120 functionsas a potent neutralization target when introduced into envelope proteinsderived from human immunodeficiency virus type 1 primary isolates. J. Virol.79:6909–6917.
59. Pinter, A., W. J. Honnen, Y. He, M. K. Gorny, S. Zolla-Pazner, and S. C.Kayman. 2004. The V1/V2 domain of gp120 is a global regulator of thesensitivity of primary human immunodeficiency virus type 1 isolates to neu-tralization by antibodies commonly induced upon infection. J. Virol. 78:5205–5215.
60. Posada, D., and K. A. Crandall. 1998. MODELTEST: testing the model ofDNA substitution. Bioinformatics 14:817–818.
61. Poss, M., and J. Overbaugh. 1999. Variants from the diverse virus popula-tion identified at seroconversion of a clade A human immunodeficiency virustype 1-infected woman have distinct biological properties. J. Virol. 73:5255–5264.
62. Poss, M., A. G. Rodrigo, J. J. Gosink, G. H. Learn, D. de Vange Panteleeff,H. L. Martin, Jr., J. Bwayo, J. K. Kreiss, and J. Overbaugh. 1998. Evolutionof envelope sequences from the genital tract and peripheral blood of womeninfected with clade A human immunodeficiency virus type 1. J. Virol. 72:8240–8251.
63. Pugach, P., S. E. Kuhmann, J. Taylor, A. J. Marozsan, A. Snyder, T. Ketas,S. M. Wolinsky, B. T. Korber, and J. P. Moore. 2004. The prolonged cultureof human immunodeficiency virus type 1 in primary lymphocytes increases itssensitivity to neutralization by soluble CD4. Virology 321:8–22.
64. Reza, F. 1994. Introduction to information theory. Dover, Mineola, NY.65. Richardson, J. S., and D. C. Richardson. 1988. Amino acid preferences for
specific locations at the ends of alpha helices. Science 240:1648–1652.66. Rong, R., F. Bibollet-Ruche, J. Mulenga, S. Allen, J. L. Blackwell, and C. A.
Derdeyn. 2007. Role of V1V2 and other human immunodeficiency virus type1 envelope domains in resistance to autologous neutralization during clade Cinfection. J. Virol. 81:1350–1359.
67. Rudensey, L. M., J. T. Kimata, E. M. Long, B. Chackerian, and J. Over-baugh. 1998. Changes in the extracellular envelope glycoprotein of variantsthat evolve during the course of simian immunodeficiency virus SIVMneinfection affect neutralizing antibody recognition, syncytium formation, andmacrophage tropism but not replication, cytopathicity, or CCR-5 coreceptorrecognition. J. Virol. 72:209–217.
68. Ryckaert, J. P., G. Ciccotti, and H. C. Berendsen. 1977. Numerical integra-tion of the Cartesian equations of motion of a system with constraints:molecular dynamics of n-alkanes. J. Comput. Phys. 23:327–341.
69. Sali, A., and T. L. Blundell. 1993. Comparative protein modelling by satis-faction of spatial restraints. J. Mol. Biol. 234:779–815.
70. Saunders, C. J., R. A. McCaffrey, I. Zharkikh, Z. Kraft, S. E. Malenbaum, B.Burke, C. Cheng-Mayer, and L. Stamatatos. 2005. The V1, V2, and V3regions of the human immunodeficiency virus type 1 envelope differentiallyaffect the viral phenotype in an isolate-dependent manner. J. Virol. 79:9069–9080.
71. Stamatatos, L., and C. Cheng-Mayer. 1998. An envelope modification thatrenders a primary, neutralization-resistant clade B human immunodeficiencyvirus type 1 isolate highly susceptible to neutralization by sera from otherclades. J. Virol. 72:7840–7845.
72. Trask, S., C. A. Derdeyn, F. Kasolo, R. Musonda, F. Gao, S. A. Allen, and B.Hahn. 2002. Epidemiological linkage analysis in a heterosexual transmissioncohort in Zambia. J. Virol. 76:397–405.
73. Warrier, S. V., A. Pinter, W. J. Honnen, M. Girard, E. Muchmore, and S. A.Tilley. 1994. A novel, glycan-dependent epitope in the V2 domain of humanimmunodeficiency virus type 1 gp120 is recognized by a highly potent, neu-tralizing chimpanzee monoclonal antibody. J. Virol. 68:4636–4642.
74. Wei, X., J. M. Decker, S. Wang, H. Hui, J. C. Kappes, X. Wu, J. F. Salazar,M. G. Salazar, J. M. Kilby, M. S. Saag, N. L. Komarova, M. A. Nowak, B. H.Hahn, P. D. Kwong, and G. M. Shaw. 2003. Antibody neutralization andescape by HIV-1. Nature 422:307–312.
75. Wyatt, R., P. D. Kwong, E. Desjardins, R. W. Sweet, J. Robinson, W. A.Hendrickson, and J. G. Sodroski. 1998. The antigenic structure of the HIVgp120 envelope glycoprotein. Nature 393:705–711.
76. Wyatt, R., J. Moore, M. Accola, E. Desjardin, J. Robinson, and J. Sodroski.1995. Involvement of the V1/V2 variable loop structure in the exposure ofhuman immunodeficiency virus type 1 gp120 epitopes induced by receptorbinding. J. Virol. 69:5723–5733.
77. Yang, W., J. P. Bielawski, and Z. Yang. 2003. Widespread adaptive evolutionin the human immunodeficiency virus type 1 genome. J. Mol. Evol. 57:212–221.
5668 RONG ET AL. J. VIROL.




















