
Genetic diversity and genomic distribution of homologs encoding NBS-LRR disease resistance proteins...
15
Mol Genet Genomics (2008) 280:111–125 DOI 10.1007/s00438-008-0346-1 123 ORIGINAL PAPER Genetic diversity and genomic distribution of homologs encoding NBS-LRR disease resistance proteins in sunXower Osman Radwan · Sonali Gandhi · Adam Heesacker · Brett Whitaker · Chris Taylor · Alex Plocik · Richard Kesseli · Alexander Kozik · Richard W. Michelmore · Steven J. Knapp Received: 26 March 2008 / Accepted: 21 April 2008 / Published online: 14 June 2008 © Springer-Verlag 2008 Abstract Three-fourths of the recognition-dependent dis- ease resistance genes (R-genes) identiWed in plants encode nucleotide binding site (NBS) leucine-rich repeat (LRR) proteins. NBS-LRR homologs have only been isolated on a limited scale from sunXower (Helianthus annuus L.), and most of the previously identiWed homologs are members of two large NBS-LRR clusters harboring downy mildew R-genes. We mined the sunXower EST database and used comparative genomics approaches to develop a deeper understanding of the diversity and distribution of NBS- LRR homologs in the sunXower genome. Collectively, 630 NBS-LRR homologs were identiWed, 88 by mining a data- base of 284,241 sunXower ESTs and 542 by sequencing 1,248 genomic DNA amplicons isolated from common and wild sunXower species. DNA markers were developed from 196 unique NBS-LRR sequences and facilitated genetic mapping of 167 NBS-LRR loci. The latter were distributed throughout the sunXower genome in 44 clusters or single- tons. Wild species ESTs were a particularly rich source of novel NBS-LRR homologs, many of which were tightly linked to previously mapped downy mildew, rust, and broomrape R-genes. The DNA sequence and mapping resources described here should facilitate the discovery and isolation of recognition-dependent R-genes guarding sun- Xower from a broad spectrum of economically important diseases. Keywords Helianthus · Lactuca · Compositae · Resistance genes Introduction Many recognition-dependent plant disease resistance (R) genes encode proteins comprised an amino-terminal domain, a nucleotide binding site (NBS) domain, and leu- cine-rich repeats (LRRs) (Bent 1996; Hammond-Kosack and Jones 1997; Hulbert et al. 2001). NBS-LRR encoding genes are often highly duplicated and evolutionarily diverse, with hundreds of family members in plant genomes (Meyers et al. 1999, 2003; Dangl and Jones 2001; GoV et al. 2002; Richly et al. 2002; McHale et al. 2006; Jones and Dangl 2006; Ameline-Torregrosa et al. 2007; Kohler et al. 2008). NBS-LRR resistance proteins directly or DDBJ/EMBL/GenBank sequence accession numbers. SunXower nucleotide and amino acid sequences have been deposited in DDBJ/ EMBL/GenBank under accession numbers EF560168-EF559378 and ABQ58077-ABQ57529. Communicated by K. Shirasu. O. Radwan · A. Heesacker · B. Whitaker · C. Taylor · S. J. Knapp (&) Center for Applied Genetic Technologies, The University of Georgia, 111 Riverbend Road, Athens, GA 30602, USA e-mail: [email protected] O. Radwan Plant Production Department, EYcient Productivity Institute, University of Zagazig, Sharkia, Egypt S. Gandhi Department of Crop and Soil Science, Oregon State University, Corvallis, OR 97331, USA A. Plocik · R. Kesseli Department of Biology, University of Massachusetts, Boston, MA 02125, USA A. Kozik · R. W. Michelmore Department of Plant Sciences and the Genome Center, University of California, Davis, CA 95616, USA
Transcript of Genetic diversity and genomic distribution of homologs encoding NBS-LRR disease resistance proteins...

Mol Genet Genomics (2008) 280:111–125
DOI 10.1007/s00438-008-0346-1ORIGINAL PAPER
Genetic diversity and genomic distribution of homologs encoding NBS-LRR disease resistance proteins in sunXower
Osman Radwan · Sonali Gandhi · Adam Heesacker · Brett Whitaker · Chris Taylor · Alex Plocik · Richard Kesseli · Alexander Kozik · Richard W. Michelmore · Steven J. Knapp
Received: 26 March 2008 / Accepted: 21 April 2008 / Published online: 14 June 2008© Springer-Verlag 2008
Abstract Three-fourths of the recognition-dependent dis-ease resistance genes (R-genes) identiWed in plants encodenucleotide binding site (NBS) leucine-rich repeat (LRR)proteins. NBS-LRR homologs have only been isolated on alimited scale from sunXower (Helianthus annuus L.), andmost of the previously identiWed homologs are members oftwo large NBS-LRR clusters harboring downy mildewR-genes. We mined the sunXower EST database and usedcomparative genomics approaches to develop a deeper
understanding of the diversity and distribution of NBS-LRR homologs in the sunXower genome. Collectively, 630NBS-LRR homologs were identiWed, 88 by mining a data-base of 284,241 sunXower ESTs and 542 by sequencing1,248 genomic DNA amplicons isolated from common andwild sunXower species. DNA markers were developed from196 unique NBS-LRR sequences and facilitated geneticmapping of 167 NBS-LRR loci. The latter were distributedthroughout the sunXower genome in 44 clusters or single-tons. Wild species ESTs were a particularly rich source ofnovel NBS-LRR homologs, many of which were tightlylinked to previously mapped downy mildew, rust, andbroomrape R-genes. The DNA sequence and mappingresources described here should facilitate the discovery andisolation of recognition-dependent R-genes guarding sun-Xower from a broad spectrum of economically importantdiseases.
Keywords Helianthus · Lactuca · Compositae · Resistance genes
Introduction
Many recognition-dependent plant disease resistance (R)genes encode proteins comprised an amino-terminaldomain, a nucleotide binding site (NBS) domain, and leu-cine-rich repeats (LRRs) (Bent 1996; Hammond-Kosackand Jones 1997; Hulbert et al. 2001). NBS-LRR encodinggenes are often highly duplicated and evolutionarilydiverse, with hundreds of family members in plantgenomes (Meyers et al. 1999, 2003; Dangl and Jones 2001;GoV et al. 2002; Richly et al. 2002; McHale et al. 2006; Jonesand Dangl 2006; Ameline-Torregrosa et al. 2007; Kohleret al. 2008). NBS-LRR resistance proteins directly or
DDBJ/EMBL/GenBank sequence accession numbers. SunXower nucleotide and amino acid sequences have been deposited in DDBJ/EMBL/GenBank under accession numbers EF560168-EF559378 and ABQ58077-ABQ57529.
Communicated by K. Shirasu.
O. Radwan · A. Heesacker · B. Whitaker · C. Taylor · S. J. Knapp (&)Center for Applied Genetic Technologies, The University of Georgia, 111 Riverbend Road, Athens, GA 30602, USAe-mail: [email protected]
O. RadwanPlant Production Department, EYcient Productivity Institute, University of Zagazig, Sharkia, Egypt
S. GandhiDepartment of Crop and Soil Science, Oregon State University, Corvallis, OR 97331, USA
A. Plocik · R. KesseliDepartment of Biology, University of Massachusetts, Boston, MA 02125, USA
A. Kozik · R. W. MichelmoreDepartment of Plant Sciences and the Genome Center, University of California, Davis, CA 95616, USA
123

112 Mol Genet Genomics (2008) 280:111–125
indirectly recognize pathogen avirulence factors, whichtrigger signal transduction cascades leading to rapiddefense responses and frequently to hypersensitive reac-tions and programmed cell death (Dangl and Jones 2001;Bent 1996; Hammond-Kosack and Jones 1997; Hulbertet al. 2001). Of the 40 or so R-genes known to confer resis-tance to bacterial, fungal, viral, and nematode pathogens inplants, 75% encode NBS-LRR proteins. LRRs are determi-nants of response speciWcity and are typically under strongdiversifying selection, whereas NBS domains are not(Parniske et al. 1997; McDowell et al. 1998; Meyers et al.1998a; Sun et al. 2001; Kuang et al. 2004).
Conserved amino acid sequence motifs in the NBSdomain have been widely used to isolate and classify NBS-LRR encoding genes (Meyers et al. 1999, 2002, 2003; Panet al. 2000; Bai et al. 2002; He et al. 2004; Cannon et al.2002). Two superfamilies have been described in dicots,one with an N-terminal structural domain homologous tothe intracellular signaling domains of the Drosophila Tolland mammalian interleukin1 receptor (TIR) and the otherlacking the TIR structural domain (non-TIR) (Meyers et al.1999; Pan et al. 2000). Coiled-coil (CC) motifs are com-monly found in the N-terminal structural domains of genesencoding non-TIR-NBS-LRR proteins, and members of thetwo superfamilies can be distinguished using conservedamino acid sequence motifs in the NBS domain (Meyerset al. 1999; Pan et al. 2000). Hundreds of NBS-LRR encod-ing genes have been identiWed in sequenced plant genomesand create a complex pathogen defense network—the Ara-bidopsis genome harbors ca. 150, the Populus trichocarpaand Medicago truncatula genomes harbor ca. 400, and therice (Orzya sativa L.) genome harbors ca. 600 NBS-LRRencoding genes (Meyers et al. 1999, 2003; Dangl and Jones2001; Cannon et al. 2002; GoV et al. 2002; Richly et al.2002; Holt et al. 2003; Jones and Dangl 2006; Ameline-Torregrosa et al. 2007; Kohler et al. 2008). The number anddiversity of NBS-LRR encoding genes in the sunXower(Helianthus annuus L.; 2n = 2x = 34) genome are notknown.
NBS-LRR encoding genes have been isolated fromnumerous species by cloning and sequencing genomicDNA fragments ampliWed by degenerate oligonucleotideprimers complementary to conserved amino acid sequencemotifs in the NBS domain, typically from the P-loop to theGLPALP motif (Kanazin et al. 1996; Leister et al. 1996;Yu et al. 1996; Shen et al. 1998). The spectrum of resis-tance gene candidates (RGCs) isolated from sunXowerusing the degenerate primer strategy has been limited, andonly a few of the previously identiWed RGCs have beengenetically mapped, primarily using RFLP markers (Gentz-bittel et al. 1998; Gedil et al. 2001). The latter facilitatedthe discovery and cloning of downy mildew (Plasmoparahalstedii (Farl.) Berl. & De Toni 1888) R-genes found in
two tightly linked families of NBS-LRR encoding genes(Gentzbittel et al. 1998; Bert et al. 2001; Gedil et al. 2001;Bouzidi et al. 2002; Radwan et al. 2003, 2004; Slabaughet al. 2003).
EST databases are another resource for identifying NBS-LRR encoding genes (Zhu et al. 2002; Rossi et al. 2003).The sunXower genome is physically large (3,500 Mbp;Baack et al. 2005) and has not been sequenced; however,326,421 ESTs have been produced from H. annuus, Jerusa-lem artichoke (Helianthus tuberosus L.; 2n = 6x = 102), sil-verleaf sunXower (Helianthus argophyllus Torr. and Gray;2n = 2x = 34), and other wild species (http://cgpdb.ucda-vis.edu/cgpdb2/). We identiWed several hundred RGCs bymining the sunXower EST database and sequencing geno-mic DNA fragments ampliWed from H. annuus, H. tubero-sus, and other wild sunXower species using degenerateprimers complementary to conserved amino acid sequencemotifs in the NBS domain. The newly isolated RGCs aredescribed here. Nearly one-fourth of the unique NBS-LRRencoding genes identiWed in the present study were geneti-cally mapped using insertion–deletion (INDEL) or single-strand conformational polymorphism (SSCP) markers(Orita et al. 1989; Kukita et al. 1997; Larsen et al. 1999,2007). The analyses described here build a more completepicture of the genetic diversity and genomic distribution ofNBS-LRR encoding genes in sunXower and supply a geno-mic framework for identifying and isolating recognition-dependent R-genes conferring resistance to a wide range ofpathogens of sunXower.
Materials and methods
Plant materials
Genomic DNAs were isolated from young leaves harvestedfrom greenhouse grown plants of an elite oilseed inbredline (RHA373) and a wild self-incompatible outbredpopulation (ANN1811; PI 494567) of H. annuus and wild,self-incompatible, outbred populations of H. argophyllus(ARG-1807, PI 494573), Helianthus deserticola (DES-2345, PI 649872), Helianthus paradoxus (PAR-Cibola),and H. tuberosus (TUB-49, PI 650095). Seeds of PAR-Cibola were supplied by Dr. Loren Rieseberg (IndianaUniversity, Bloomington, Indiana). Seeds of the othergermplasm accessions were supplied by the United StatesDepartment of Agriculture (USDA) Agricultural ResearchService (ARS) National Plant Germplasm System (http://www.ars-grin.gov/npgs/) or the USDA-ARS NorthernCrop Science Research Laboratory. DNA was isolated frommanually ground fresh or frozen leaf tissues using a modi-Wed CTAB (cetyltrimethylammonium bromide) method(Murray and Thompson 1980).
123

Mol Genet Genomics (2008) 280:111–125 113
NBS fragment isolation by PCR ampliWcation using degenerate primers
Genomic DNA fragments spanning conserved NBSsequences were isolated from common and wild sunXowerspecies using two pairs of degenerate forward and reverseoligonucleotide primers (NBS-F1/NBS-R1 and NBS-F2/NBS-R2) complementary to conserved P-loop andGLPLAL motifs in the NBS domain (Table 1; Leister et al.1996; He et al. 2004) and were predicted to amplify ca. 500bp DNA fragments. Standard PCR methods were used,amplicons were separated on 1.5% agarose gels (Sigma,USA), and bands of the predicted length were excised andpuriWed using the QIAquick method (Qiagen USA), clonedinto the pCR 4-TOPO vector (Invitrogen, USA), chemi-cally transformed into TOP10 E. coli cells (Invitrogen,USA), and plated on Luria broth agar containing 100 mg ofampicillin/l. We randomly selected 192 clones from theANN1811, ARG-1807, DES-2345, PAR-Cibola, and TUB-49 libraries and 288 clones from the RHA373 library forDNA sequencing. The selected clones were bidirectionallySanger sequenced on an ABI3730XL using ABI Big-Dyetechnology (Applied Biosystems, USA). Sequences ofclones harboring putative pseudogenes were substantiatedby resequencing.
Mining sunXower and lettuce EST databases for NBS-LRR encoding resistance gene homologs
SunXower and lettuce transcript assemblies (TAs) in theCompositae Genome Database (CGPdb; http://www.cgpdb.ucdavis.edu/) were screened to identify unigenes homolo-gous to NBS-LRR encoding genes using “NBS”, “LRR”,and “NBS-LRR” keyword searches (TAs were BLASTannotated). Homologs with BLASTX scores ·e¡0.15 wereselected, cDNA sequences were translated in six framesusing the CGPdb pipeline (http://cgpdb.ucdavis.edu/data-base/sms/translation.php), and BLASTX analyses were
performed against the NCBI Protein Database to substanti-ate the homology of the selected sunXower and lettuceunigenes to NBS-LRR encoding R-genes identiWed in otherplant species (Altschul et al. 1997; http://www.ncbi.nlm.nih.gov). SunXower NBS-LRR sequences were used tosearch for lettuce homologs in the CGPdb EST database.
Sequence and genetic diversity analyses
Nucleotide binding site nucleotide and amino acidsequences were aligned using Clustal_X (Thompson et al.1997). JALVIEW was used to display and identify redun-dant sequences (Clamp et al. 2004; http://www.jalview.org/).NBS amino acid sequences were screened for motifscharacteristic of proteins encoded by plant disease resis-tance genes and were classiWed as non-TIR or TIR usingamino acid sequence motifs characteristic of the twosuperfamilies (Bent 1996; Hammond-Kosack and Jones1997; Meyers et al. 1999, 2003; Pan et al. 2000). TIRsequences were identiWed by the presence of the RNBS-A TIR motif (FLENIRExSKKHGLEHLQKKLLSKLL)and aspartic acid (D) as the last amino acid residue in theKin-2 motif, whereas non-TIR sequences were identiWedby the presence of the RNBS-A non-TIR motif (FDLx-AWVCVSQxF) and tryptophan (W) as the last aminoacid residue in the Kin-2 motif (Meyers et al. 1999, 2003;Pan et al. 2000).
Because many of the newly isolated NBS sequences(n = 758) were closely related (>90% amino acid similarity),sequences with <90% amino acid similarity (n = 69) wereselected for in-depth analysis. The NBS nucleotidesequence alignment for the newly isolated RGCs (n = 758)and the amino acid sequence alignment for the selectedsunXower NBS sequences (n = 69) have been supplied asGeneDoc input Wles [Appendices (S1) and (S2)]; GeneDocis freeware and can be downloaded from http://www.nrbsc.org/gfx/genedoc/index.html or http://www.nrbsc.org/downloads/. Neighbor-joining (NJ) trees were pro-duced from analyses of NBS amino acid similarity matri-ces with bootstrap resampling (k = 1,000 permutations)(Saitou and Nei 1987; Thompson et al. 1997). Two treeswere produced, one from an alignment and analysis of the69 selected sunXower sequences with <90% amino acidsimilarity and 24 lettuce (Lactuca sativa L.) sequencesselected from previously identiWed families (Meyers et al.1998b; Kuang et al. 2004) and one from an alignment andanalysis of sunXower and lettuce sequences withsequences from diverse NBS-LRR encoding R-genes iso-lated from other plant species: RPS4 (AJ243468), RPP1(AF098962), ADR1 (AJ581996), RPS5 (AF074916),RPM1 (X87851), RPP13 (AF209730), RPS2 (ATU14158),RPP5 (ATRPP5LE1), and RPP8 (AF089711) from Ara-bidopsis; L6 (LUU27081) from Xax (Linum usitatissimum L.);
Table 1 Conserved NBS amino acid sequences targeted by degener-ate oligonucleotide primers for isolating genomic DNA fragmentsfrom sunXower
a I (Inosine); N (A,C,G,T); R (A,G); Y (C,T); S (G,C); H (A,T,C) andV (G,A,C)
Target amino acid sequence
Primer name
Oligonucleotide primer sequence (5�–3�)a
GGVGKTT NBS-F1 GGIGGIGTIGGIAAIACIAC
GLPLAL NBS-R1 IAGIGCIAGIGGIAGICC
GMGGVGKT NBS-F2 GGNATGGGNGGNGTNGGNAARAC
GLPLALKV NBS-R2 ACYTTNARNGCNARNGGNARNCC
123

114 Mol Genet Genomics (2008) 280:111–125
N (NGU15605) from tobacco (Nicotiana tabacum L.)and NRG1 (DQ054580) from Nicotiana benthamiana; Mi(AF039681) and Rx (AJ011801) from potato (Solanumtuberosum L.); I2 (AF004878) from tomato (Solanum lyc-opersicon L.); Xa1 (AB002266), Pi-ta (AF207842), andPib (AB013449) from rice (Oryza sativa L.); Rp1(AF107293) from maize (Zea mays L.); and Dm3(LSRGC2B1) from lettuce (GenBank accession numbersare shown in parentheses).
DNA marker development, genotyping, and genetic mapping
Single-strand conformational polymorphism and INDELmarkers were developed for genotyping NBS-LRR loci bydesigning Xanking oligonucleotide primers complemen-tary to cDNA and genomic DNA reference sequences.Whenever possible, nucleotide polymorphisms were usedto identify paralog-speciWc sequences for designing prim-ers for INDEL and SSCP marker genotyping. INDELmarkers were manually genotyped on 1.5% NuSieve aga-rose gels (FMC, USA). SSCP markers were manuallygenotyped by silver-staining polyacrylamide gels (Bas-sam et al. 1991; Slabaugh et al. 2003). PCRs were per-formed using 30 ng of DNA template, 0.65 U Taqpolymerase (Qiagen, USA), 1£ PCR reaction buVer, 2.5mM Mgcl2, 0.2 mM dNTPs, 0.16 �M of each primer (onepair/marker), and a ‘touchdown’ PCR protocol (Don et al.1991; Hecker and Roux 1996) with an initial denaturationtemperature of 94 C for 3 min, followed by 1 cycle of 94C for 30 s, 68 C for 30 s and 72 C for 60 s; annealing tem-peratures in subsequent cycles were decreased by 1 Cuntil reaching 58 C, products were ampliWed for 35 cyclesat 94 C for 30 s/cycle, 58 C for 30 s/cycle, and 72 C for 60s/cycle, and products were extended in the Wnal cycle at72 C for 15 min. Genomic DNA amplicons were sepa-rated on agarose to screen for INDEL polymorphismsand, when absent, were separated and genotyped usingSSCP analysis. Polymorphic INDEL and SSCP markerswere genotyped on 94 RHA280 £ RHA801 recombinantinbred lines (RILs) (Tang et al. 2002; Yu et al. 2003) or94 NMS373 £ (NMS373 £ ANN1238) BC1 progeny(Gandhi et al. 2005). NBS-LRR INDEL and SSCP markerloci were anchored, grouped, and ordered against a previ-ously mapped genome-wide backbone of SSR and INDELmarkers (Tang et al. 2002; Yu et al. 2003; Gandhi et al.2005). Genetic mapping analyses were done using MAP-MAKER (Lander et al. 1987) as previously described(Tang et al. 2002). DNA marker loci were ordered byallowing 0 or 2% genotyping errors (Lincoln and Lander1992), primarily for comparing likelihoods of locusorders in genomic regions densely populated with NBS-LRR marker loci.
Results
SunXower RGCs identiWed by targeting conserved NBS sequence motifs and mining the EST database
Genomic DNA fragments were ampliWed from H. annuus,H. argophyllus, desert sunXower (H. deserticola Heiser;2n = 2x = 34), Pecos sunXower (H. paradoxus Heiser; 2n =2x = 34), and H. tuberosus using two pairs of degenerateoligonucleotide primers complementary to highly conservedNBS sequence motifs (Leister et al. 1996; Meyers et al.1999; He et al. 2004, Tables 1, 2). Both primer pairs (NBS-F1/R1 and NBS-F2/R2) were predicted to amplify ca. 500bp fragments from TIR and non-TIR-NBS-LRR encodinggenes. Collectively, 1,248 amplicons were cloned andsequenced (196–288/genotype), 1,052 of the 1,248 rese-quenced amplicons (RSAs) were homologous to NBSsequences in NBS-LRR encoding genes previously isolatedfrom sunXower or other plant species, 784 RSAs had unin-terrupted ORFs from the P-loop to the GLPLAL motif, 542RSAs were unique and lacked stop codons or frame shiftmutations, and 242 RSAs harbored stop codons or frameshift mutations and were suspected to be pseudogenes[Table 2; GenBank Acc. No. EF559385-EF560169; Appen-dix (S3)]. Sequencing errors should have been minimalbecause each clone was bidirectionally sequenced and puta-tive pseudogenes were resequenced. The 542 unique NBSsequences ranged in length from 164 to 173 amino acids(GenBank Acc. No. ABQ58077-ABQ57529). Three-fourths of the newly isolated NBS sequences lacking stopcodons were TIR class (Meyers et al. 1999, 2003; Pan et al.2000). Nearly half of the NBS sequences isolated from H.annuus were TIR (53.6%) class, whereas 93.9% of the NBSsequences isolated from H. tuberosus were TIR class. Ofthe NBS sequences harboring stop codons, 95.3% were TIRclass [Appendix (S3)].
Eighty-eight unigenes homologous to NBS-LRR encod-ing R-genes were identiWed by screening transcript assem-blies (TAs) developed from 284,745 Helianthus ESTs inthe Compositae Genome Program Database (CGPdb; http://cgpdb.ucdavis.edu/cgpdb2/). The ESTs spanned pre-NBS,NBS, LRR-, or post-LRR domains or combinations thereof[Appendix (S3)]. ESTs spanning NBS domains werealigned with previously and newly isolated NBS domainsequences (Gedil et al. 2001; Radwan et al. 2003; Plociket al. 2004). Nucleotide identities ranged from 28 to 100%,while amino acid similarities ranged from 16 to 100%.
Diversity of sunXower homologs encoding NBS-LRR resistance proteins
SunXower NBS sequences (n = 758) spanning the P-loop tothe GLPLAL motif were translated and aligned to identify
123
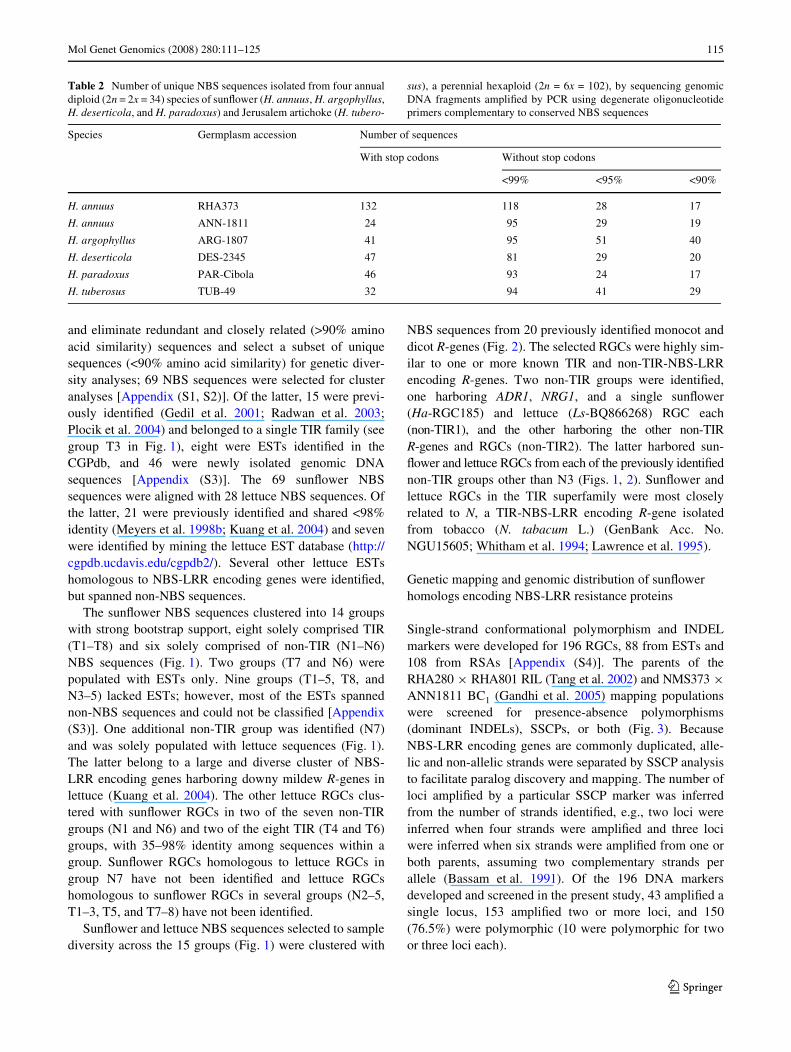
Mol Genet Genomics (2008) 280:111–125 115
and eliminate redundant and closely related (>90% aminoacid similarity) sequences and select a subset of uniquesequences (<90% amino acid similarity) for genetic diver-sity analyses; 69 NBS sequences were selected for clusteranalyses [Appendix (S1, S2)]. Of the latter, 15 were previ-ously identiWed (Gedil et al. 2001; Radwan et al. 2003;Plocik et al. 2004) and belonged to a single TIR family (seegroup T3 in Fig. 1), eight were ESTs identiWed in theCGPdb, and 46 were newly isolated genomic DNAsequences [Appendix (S3)]. The 69 sunXower NBSsequences were aligned with 28 lettuce NBS sequences. Ofthe latter, 21 were previously identiWed and shared <98%identity (Meyers et al. 1998b; Kuang et al. 2004) and sevenwere identiWed by mining the lettuce EST database (http://cgpdb.ucdavis.edu/cgpdb2/). Several other lettuce ESTshomologous to NBS-LRR encoding genes were identiWed,but spanned non-NBS sequences.
The sunXower NBS sequences clustered into 14 groupswith strong bootstrap support, eight solely comprised TIR(T1–T8) and six solely comprised of non-TIR (N1–N6)NBS sequences (Fig. 1). Two groups (T7 and N6) werepopulated with ESTs only. Nine groups (T1–5, T8, andN3–5) lacked ESTs; however, most of the ESTs spannednon-NBS sequences and could not be classiWed [Appendix(S3)]. One additional non-TIR group was identiWed (N7)and was solely populated with lettuce sequences (Fig. 1).The latter belong to a large and diverse cluster of NBS-LRR encoding genes harboring downy mildew R-genes inlettuce (Kuang et al. 2004). The other lettuce RGCs clus-tered with sunXower RGCs in two of the seven non-TIRgroups (N1 and N6) and two of the eight TIR (T4 and T6)groups, with 35–98% identity among sequences within agroup. SunXower RGCs homologous to lettuce RGCs ingroup N7 have not been identiWed and lettuce RGCshomologous to sunXower RGCs in several groups (N2–5,T1–3, T5, and T7–8) have not been identiWed.
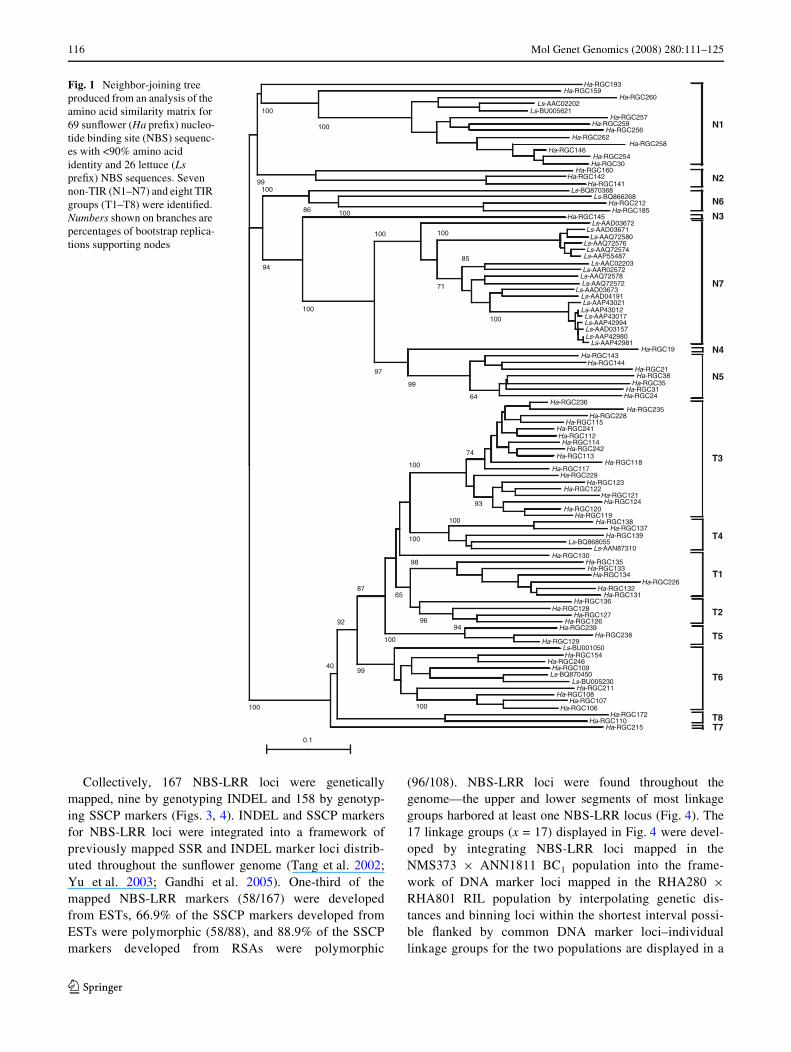
SunXower and lettuce NBS sequences selected to samplediversity across the 15 groups (Fig. 1) were clustered with
NBS sequences from 20 previously identiWed monocot anddicot R-genes (Fig. 2). The selected RGCs were highly sim-ilar to one or more known TIR and non-TIR-NBS-LRRencoding R-genes. Two non-TIR groups were identiWed,one harboring ADR1, NRG1, and a single sunXower(Ha-RGC185) and lettuce (Ls-BQ866268) RGC each(non-TIR1), and the other harboring the other non-TIRR-genes and RGCs (non-TIR2). The latter harbored sun-Xower and lettuce RGCs from each of the previously identiWednon-TIR groups other than N3 (Figs. 1, 2). SunXower andlettuce RGCs in the TIR superfamily were most closelyrelated to N, a TIR-NBS-LRR encoding R-gene isolatedfrom tobacco (N. tabacum L.) (GenBank Acc. No.NGU15605; Whitham et al. 1994; Lawrence et al. 1995).
Genetic mapping and genomic distribution of sunXower homologs encoding NBS-LRR resistance proteins
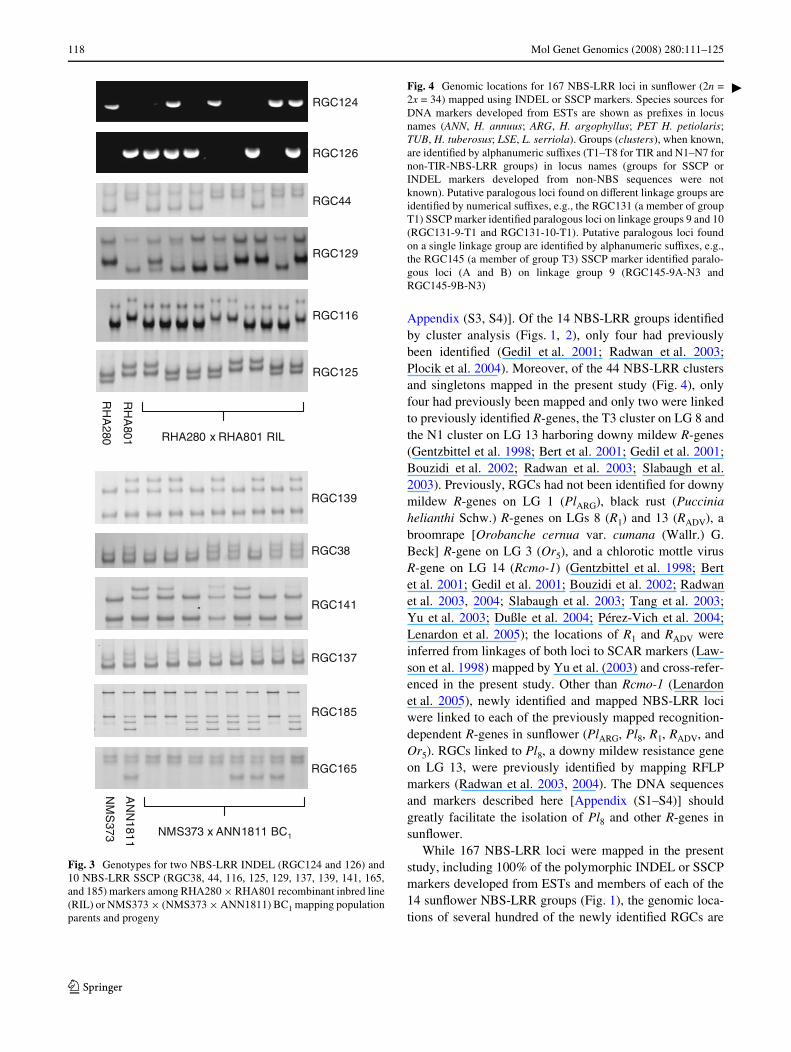
Single-strand conformational polymorphism and INDELmarkers were developed for 196 RGCs, 88 from ESTs and108 from RSAs [Appendix (S4)]. The parents of theRHA280 £ RHA801 RIL (Tang et al. 2002) and NMS373 £ANN1811 BC1 (Gandhi et al. 2005) mapping populationswere screened for presence-absence polymorphisms(dominant INDELs), SSCPs, or both (Fig. 3). BecauseNBS-LRR encoding genes are commonly duplicated, alle-lic and non-allelic strands were separated by SSCP analysisto facilitate paralog discovery and mapping. The number ofloci ampliWed by a particular SSCP marker was inferredfrom the number of strands identiWed, e.g., two loci wereinferred when four strands were ampliWed and three lociwere inferred when six strands were ampliWed from one orboth parents, assuming two complementary strands perallele (Bassam et al. 1991). Of the 196 DNA markersdeveloped and screened in the present study, 43 ampliWed asingle locus, 153 ampliWed two or more loci, and 150(76.5%) were polymorphic (10 were polymorphic for twoor three loci each).
Table 2 Number of unique NBS sequences isolated from four annualdiploid (2n = 2x = 34) species of sunXower (H. annuus, H. argophyllus,H. deserticola, and H. paradoxus) and Jerusalem artichoke (H. tubero-
sus), a perennial hexaploid (2n = 6x = 102), by sequencing genomicDNA fragments ampliWed by PCR using degenerate oligonucleotideprimers complementary to conserved NBS sequences
Species Germplasm accession Number of sequences
With stop codons Without stop codons
<99% <95% <90%
H. annuus RHA373 132 118 28 17
H. annuus ANN-1811 24 95 29 19
H. argophyllus ARG-1807 41 95 51 40
H. deserticola DES-2345 47 81 29 20
H. paradoxus PAR-Cibola 46 93 24 17
H. tuberosus TUB-49 32 94 41 29
123
116 Mol Genet Genomics (2008) 280:111–125
Collectively, 167 NBS-LRR loci were geneticallymapped, nine by genotyping INDEL and 158 by genotyp-ing SSCP markers (Figs. 3, 4). INDEL and SSCP markersfor NBS-LRR loci were integrated into a framework ofpreviously mapped SSR and INDEL marker loci distrib-uted throughout the sunXower genome (Tang et al. 2002;Yu et al. 2003; Gandhi et al. 2005). One-third of themapped NBS-LRR markers (58/167) were developedfrom ESTs, 66.9% of the SSCP markers developed fromESTs were polymorphic (58/88), and 88.9% of the SSCPmarkers developed from RSAs were polymorphic
(96/108). NBS-LRR loci were found throughout thegenome—the upper and lower segments of most linkagegroups harbored at least one NBS-LRR locus (Fig. 4). The17 linkage groups (x = 17) displayed in Fig. 4 were devel-oped by integrating NBS-LRR loci mapped in theNMS373 £ ANN1811 BC1 population into the frame-work of DNA marker loci mapped in the RHA280 £RHA801 RIL population by interpolating genetic dis-tances and binning loci within the shortest interval possi-ble Xanked by common DNA marker loci–individuallinkage groups for the two populations are displayed in a
Fig. 1 Neighbor-joining tree produced from an analysis of the amino acid similarity matrix for 69 sunXower (Ha preWx) nucleo-tide binding site (NBS) sequenc-es with <90% amino acid identity and 26 lettuce (Ls preWx) NBS sequences. Seven non-TIR (N1–N7) and eight TIR groups (T1–T8) were identiWed. Numbers shown on branches are percentages of bootstrap replica-tions supporting nodes
Ha-RGC215Ha-RGC110
Ha-RGC172Ha-RGC106
Ha-RGC107Ha-RGC108
Ha-RGC211
100
Ls-BU005230Ls-BQ870450Ha-RGC109
Ha-RGC246Ha-RGC154Ls-BU001050
Ha-RGC129Ha-RGC238
Ha-RGC239Ha-RGC126
Ha-RGC127Ha-RGC128
94
Ha-RGC136
96
Ha-RGC131Ha-RGC132
Ha-RGC226Ha-RGC134
Ha-RGC133Ha-RGC13598
Ha-RGC130Ls-AAN87310
Ls-BQ868055Ha-RGC139
Ha-RGC137Ha-RGC138100
Ha-RGC119Ha-RGC120
Ha-RGC124Ha-RGC121
Ha-RGC122Ha-RGC123
Ha-RGC229
93
Ha-RGC117Ha-RGC118
Ha-RGC113Ha-RGC242
Ha-RGC114Ha-RGC112Ha-RGC241
Ha-RGC115Ha-RGC228
Ha-RGC235Ha-RGC236
74
100
100
65
100
99
87
92
40
Ha-RGC24Ha-RGC31
Ha-RGC35Ha-RGC38Ha-RGC21
Ha-RGC144Ha-RGC143
64
Ha-RGC19
99
Ls-AAP42981Ls-AAP42980Ls-AAD03157Ls-AAP42994Ls-AAP43017
Ls-AAP43012Ls-AAP43021Ls-AAD04191
Ls-AAD03673
100
Ls-AAQ72572Ls-AAQ72578Ls-AAR02572
Ls-AAC0220385 Ls-AAP55487
Ls-AAQ72574Ls-AAQ72576
Ls-AAQ72580Ls-AAD03671
71
100Ls-AAD03672
97
100
Ha-RGC145
100
Ha-RGC185Ha-RGC212
Ls-BQ866268
100
Ls-BQ870368
86
94
100
100
Ha-RGC141Ha-RGC142
Ha-RGC160Ha-RGC30Ha-RGC254
Ha-RGC146Ha-RGC258
Ha-RGC262Ha-RGC256
Ha-RGC259Ha-RGC257
Ls-BU005621Ls-AAC02202
Ha-RGC260Ha-RGC159
100
Ha-RGC193
100
99
N1
N2
N6
N3
N7
N4
N5
T3
T4
T1
T2
T6
T5
T7T8
0.1
123

Mol Genet Genomics (2008) 280:111–125 117
comparative mapping database (http://www.sunflower.uga.edu/CMAP) developed using CMAP (http://www.gmod.org/wiki/index.php/CMAP; Gonzales et al. 2005).
Wild species ESTs were a particularly rich source ofnovel RGCs, with 31 originating from H. tuberosus, 20originating from domesticated and wild H. annuus, threeoriginating from prairie sunXower (H. petiolaris Nutt.;2n = 2x = 34), one originating from H. argophyllus, andone originating from prickly lettuce (L. serriola L.). ESTsfacilitated the discovery of 14 new NBS-LRR clusters orsingletons, which were found on the upper and lower seg-ments of linkage group (LG) 1, upper segments of linkagegroups 2, 3, 4, 5, 9, 10, and 14, and lower segments of link-age groups 6, 8, 12, 15, 16, and 17 (Fig. 4). Several NBS-LRR homologs identiWed from ESTs mapped to the twolargest and most complex and well known clusters in sun-Xower, a TIR cluster (T3) on the upper end of LG 8 and a
non-TIR cluster (N1) on the lower end of LG 13 (Gentzbit-tel et al. 1998; Bert et al. 2001; Gedil et al. 2001; Bouzidiet al. 2002; Radwan et al. 2003; Slabaugh et al. 2003).NBS-LRR encoding genes from each of the 14 groupsidentiWed from analyses of the NBS amino acid sequencealignment [Fig. 1; Appendix (S2)], other than N7, weregenetically mapped, and members of several groups werefound on two or more linkage groups. Collectively, 44NBS-LRR clusters or singletons were identiWed by geneticmapping (Fig. 4).
Discussion
The NBS-LRR markers mapped in the present study supplylandmarks for identifying and isolating R-genes conferringresistance to diverse pathogens in sunXower [Fig. 4;
Fig. 2 Neighbor-joining tree produced from an analysis of the amino acid similarity matrix for 26 sunXower (Ha preWx) and six lettuce (Ls preWx) NBS sequenc-es and NBS sequences for 20 plant R-genes (see text)
RPS4L6
RPP5RPP1
Ha-RGC215
95
Ha-RGC172Ha-RGC110
NLs-BU001050
Ha-RGC154Ha-RGC10954
Ha-RGC236Ha-RGC118
Ha-RGC119
97
Ha-RGC138Ls-AAN87310
100
95Ha-RGC239
Ha-RGC129Ha-RGC130
100
Ha-RGC135Ha-RGC131
Ha-RGC136Ha-RGC126
77
41
39
41
53
69
100
99
69
100
47
48
35
58
64
I2Ls-AAC02202Ls-BU005621
Ha-RGC30
100
Xa1
100
44Rp1
Ha-RGC159
55
55Pib
Pi-taRPM1
RXHa-RGC142Mi
78
RPP8RPP13
56
4962
36
30
89
Ha-RGC193RPS2
RPS5Ha-RGC2
Ls-AAP42981Ls-AAD03672
79
Ha-RGC143Ha-RGC24
Ha-RGC19
94
99
91Ha-RGC145
100
94
47
83
24
43
87
36
NRG1Ha-RGC185
ADR1Ls-BQ870368
100
100
35
97
APAF1
non-TIR1
non-TIR2
TIR
0.1
123
118 Mol Genet Genomics (2008) 280:111–125
Appendix (S3, S4)]. Of the 14 NBS-LRR groups identiWedby cluster analysis (Figs. 1, 2), only four had previouslybeen identiWed (Gedil et al. 2001; Radwan et al. 2003;Plocik et al. 2004). Moreover, of the 44 NBS-LRR clustersand singletons mapped in the present study (Fig. 4), onlyfour had previously been mapped and only two were linkedto previously identiWed R-genes, the T3 cluster on LG 8 andthe N1 cluster on LG 13 harboring downy mildew R-genes(Gentzbittel et al. 1998; Bert et al. 2001; Gedil et al. 2001;Bouzidi et al. 2002; Radwan et al. 2003; Slabaugh et al.2003). Previously, RGCs had not been identiWed for downymildew R-genes on LG 1 (PlARG), black rust (Pucciniahelianthi Schw.) R-genes on LGs 8 (R1) and 13 (RADV), abroomrape [Orobanche cernua var. cumana (Wallr.) G.Beck] R-gene on LG 3 (Or5), and a chlorotic mottle virusR-gene on LG 14 (Rcmo-1) (Gentzbittel et al. 1998; Bertet al. 2001; Gedil et al. 2001; Bouzidi et al. 2002; Radwanet al. 2003, 2004; Slabaugh et al. 2003; Tang et al. 2003;Yu et al. 2003; Dußle et al. 2004; Pérez-Vich et al. 2004;Lenardon et al. 2005); the locations of R1 and RADV wereinferred from linkages of both loci to SCAR markers (Law-son et al. 1998) mapped by Yu et al. (2003) and cross-refer-enced in the present study. Other than Rcmo-1 (Lenardonet al. 2005), newly identiWed and mapped NBS-LRR lociwere linked to each of the previously mapped recognition-dependent R-genes in sunXower (PlARG, Pl8, R1, RADV, andOr5). RGCs linked to Pl8, a downy mildew resistance geneon LG 13, were previously identiWed by mapping RFLPmarkers (Radwan et al. 2003, 2004). The DNA sequencesand markers described here [Appendix (S1–S4)] shouldgreatly facilitate the isolation of Pl8 and other R-genes insunXower.
While 167 NBS-LRR loci were mapped in the presentstudy, including 100% of the polymorphic INDEL or SSCPmarkers developed from ESTs and members of each of the14 sunXower NBS-LRR groups (Fig. 1), the genomic loca-tions of several hundred of the newly identiWed RGCs are
Fig. 3 Genotypes for two NBS-LRR INDEL (RGC124 and 126) and10 NBS-LRR SSCP (RGC38, 44, 116, 125, 129, 137, 139, 141, 165,and 185) markers among RHA280 £ RHA801 recombinant inbred line(RIL) or NMS373 £ (NMS373 £ ANN1811) BC1 mapping populationparents and progeny
RGC139
RGC38
RGC141
RGC137
RGC185
RGC165
AN
N181
1
NM
S373 NMS373 x ANN1811 BC1
RH
A801
RH
A280
RGC124
RGC126
RGC44
RGC129
RGC116
RGC125
RHA280 x RHA801 RIL
Fig. 4 Genomic locations for 167 NBS-LRR loci in sunXower (2n =2x = 34) mapped using INDEL or SSCP markers. Species sources forDNA markers developed from ESTs are shown as preWxes in locusnames (ANN, H. annuus; ARG, H. argophyllus; PET H. petiolaris;TUB, H. tuberosus; LSE, L. serriola). Groups (clusters), when known,are identiWed by alphanumeric suYxes (T1–T8 for TIR and N1–N7 fornon-TIR-NBS-LRR groups) in locus names (groups for SSCP orINDEL markers developed from non-NBS sequences were notknown). Putative paralogous loci found on diVerent linkage groups areidentiWed by numerical suYxes, e.g., the RGC131 (a member of groupT1) SSCP marker identiWed paralogous loci on linkage groups 9 and 10(RGC131-9-T1 and RGC131-10-T1). Putative paralogous loci foundon a single linkage group are identiWed by alphanumeric suYxes, e.g.,the RGC145 (a member of group T3) SSCP marker identiWed paralo-gous loci (A and B) on linkage group 9 (RGC145-9A-N3 andRGC145-9B-N3)
�
123

Mol Genet Genomics (2008) 280:111–125 119
123

120 Mol Genet Genomics (2008) 280:111–125
not known, although most of the unmapped RGCs werehighly similar (>91% amino acid similarity) to one or moreof the mapped RGCs [Appendix (S3); Fig. 4]. The develop-ment and genetic mapping of additional NBS-LRR markerscould be accelerated using multiplex capillary SSCP geno-typing (Larsen et al. 1999, 2007) or other high-throughputDNA fragment analysis methods (Slabaugh et al. 2003) andmight identify unique loci or clusters of loci, although oneor more members of nearly every group have been mapped.Until the sunXower genome has been sequenced, the dis-covery of RGCs will be limited to the sorts of forwardgenetic and comparative genomic approaches describedhere, as has been done in many other plant species(Kanazin et al. 1996; Leister et al. 1996; Yu et al. 1996;Collins et al. 1998; Shen et al. 1998; Pan et al. 2000; Wanget al. 2001; Wei et al. 2002; Zhu et al. 2002; Lee et al.2003; Madsen et al. 2003; Meyers et al. 2003; He et al.2004; Rossi et al. 2003). On the basis of analyses ofsequenced plant genomes (Meyers et al. 2003; Ameline-Torregrosa et al. 2007; Kohler et al. 2008), the full comple-ment of NBS-LRR encoding genes in the sunXowergenome is undoubtedly larger than the complement isolatedso far [Appendix (S1–S3)].
Wild species are rich sources of genetic diversity for dis-ease resistance and other traits in domesticated plants
(Tanksley and McCouch 1997; Michelmore 2003) and havebeen an important source of R-genes in sunXower (Pusto-voit and Kroknin 1978; Kkoric 1985; Miller and Gulya1988, 1991; Seiler 1991a, b; Tan et al. 1992; Quresh et al.1993; Degener et al. 1999; Viguié et al. 2000; Langar et al.2002; Miller et al. 2002; Vear et al. 2003; Slabaugh et al.2003; Dußle et al. 2004). Several novel sunXower NBS-LRR loci were identiWed in the present study by miningwild species ESTs and isolating NBS sequences from wildspecies. We sequenced more deeply than had been done inearlier studies and targeted wild species to increase thedepth and breath of sequences discovered by the degenerateprimer approach, particularly since wild species haveplayed an important role in disease resistance breeding insunXower, e.g., the downy mildew R-genes found on threechromosomes (PlARG on LG 1, Pl7 on LG 8, and Pl8 on LG13) were introgressed from wild species (Miller et al. 2002;Slabaugh et al. 2003; Dußle et al. 2004).
Between ESTs and RSAs, at least 918 unique NBS-LRR sequences have been identiWed in sunXower so far[Gentzbittel et al. 1998; Bert et al. 2001; Gedil et al. 2001;Bouzidi et al. 2002; Radwan et al. 2003, 2004; Slabaughet al. 2003; Plocik et al. 2004; Appendix (S3)]. Wild spe-cies ESTs were a particularly rich source of novel RGCs,were the sole source of NBS-LRR encoding genes in four
Fig. 4 continued
123

Mol Genet Genomics (2008) 280:111–125 121
of 10 newly identiWed groups (T2, T4, T8, and N5), andwere the sole source of RGCs linked to previously mappedR-genes for which RGCs have not been identiWed (Figs. 1,4). H. tuberosus ESTs supplied the largest number ofnovel RGCs, several of which were linked to R-genes pre-viously identiWed from resistance phenotypes. H. tubero-sus has been a historically important source of R-genestransferred across the diploid £ hexaploid bridge, e.g., theWrst dominant race-speciWc broomrape R-gene (Or1) wasdiscovered in H. tuberosus, perhaps as early as 1916, andwas introgressed into H. annuus through backcross breed-ing (Vranceanu et al.1980; Parker and Riches 1993). Sev-eral clusters of NBS-LRR loci identiWed from ESTs andRSAs are not linked to R-genes identiWed from resistancephenotypes, but could be the source of as yet undiscoveredR-genes.
The NBS-LRR copy numbers often vary widely withinspecies, and NBS-LRR loci frequently rearrange andevolve through recombination, unequal crossing-over,gene conversion, insertions–deletions, and point mutations(Leister et al. 1998; Michelmore and Meyers 1998; Meyerset al. 1998a; Noel et al. 1999; Chin et al. 2001; Noir et al.2001; Zhu et al. 2002; AshWeld et al. 2004; Kuang et al.2004). The copy number in a large cluster of NBS-LRRloci harboring Pl1 and other downy mildew R-genes onlinkage group 8 is highly variable in sunXower, e.g., 2 to24 copies/individual have been identiWed among elite andexotic individuals using INDEL Wngerprinting (Slabaughet al. 2003). The LG 8 cluster is the largest and most com-plex discovered so far in sunXower, with 54 NBS-LRRloci spanning a 36 cM segment in three subclusters(Fig. 4). One subcluster spans 12 cM, is primarily populated
Fig. 4 continued
123

122 Mol Genet Genomics (2008) 280:111–125
with T3 group members, has two T4 group members in themiddle of the cluster (RGC138 and RGC139), and isdemarcated on the upper end by two apparently telomericor near-telomeric loci (RGC230 and RGC237) and lowerend by additional T3 group NBS-LRR loci (e.g.,RGC244). The second subcluster is slightly downstream,spans 7 cM, and harbors four T3 group members. Finally,additional unclassiWed NBS-LRR loci were found on thelower end of LG 8, two of which were identiWed fromESTs and could not be classiWed (ANN-RGC198 and PET-RGC221).
The second largest NBS-LRR cluster identiWed so far insunXower is found on the lower end of LG 13, with 27 lociin two subclusters and a few singletons spanning a 31 cMsegment (Fig. 4). This cluster harbors TIR- and non-TIR-NBS-LRR encoding genes. Other than three TIR-NBS-LRR loci, two from the T3 group at the upper end of thesegment and one from the T1 group, loci in this clusterbelong to a single non-TIR group (N1). This segmentharbors downy mildew (Pl5 and Pl8) and black rust (RADV)R-genes (Lawson et al. 1998; Radwan et al. 2003; Slab-augh et al. 2003; Yu et al. 2003), and is now much moredensely populated with RGCs, which should facilitate theidentiWcation and cloning of R-genes for Pl8 and RADV
(Fig. 4). The presence of complex mixed clusters of NBS-LRR loci, such as the TIR and non-TIR cluster found onLG 13, is common in the genomes of higher plants (Meyerset al. 1999; Zhu et al. 2002).
While an unknown number of genes encoding NBS-LRR proteins have presumably not yet been discovered insunXower, a signiWcant number have and supply resourcesfor cataloging and cross-referencing phenotypic and quan-titative trait loci conferring resistance to diverse pathogensin sunXower and, when coupled with the steadily growingcollection of NBS-LRR sequences and mapped NBS-LRRloci, should greatly facilitate the discovery of additional R-genes and a greater understanding of R-gene diversity indomesticated and wild sunXower species (Michelmore andMeyers 1998; Noel et al. 1999; Ellis et al. 2000; Noir et al.2001; Cannon et al. 2002; Michelmore 2003). Wild spe-cies have been an important source of R-genes in sun-Xower and, not coincidentally, RGCs identiWed from wildspecies ESTs were linked to several important R-genes insunXower (PlARG, Or5, Pl8, R1, and RADV). Genetic analy-ses of RGCs for many of these R-genes are underway andshould lead to the discovery of functionally importantdeterminants of resistance and the development of addi-tional high-throughput DNA markers for accelerating dis-ease resistance breeding in sunXower through marker-assisted selection.
Acknowledgments This research was supported by grants from theUnited States Department of Agriculture National Research Initiative
Plant Genome Program (No. 2000-04292) and National Science Foun-dation Plant Genome Program (No. 0421630).
Appendix
S1. GeneDoc (http://www.nrbsc.org/gfx/genedoc/index.html) input Wle displaying a CLUSTAL-X alignment of 785sunXower NBS nucleotide sequences.S2. GeneDoc (http://www.nrbsc.org/gfx/genedoc/index.html) input Wle displaying a CLUSTAL-X alignment of 69sunXower NBS amino acid sequences sharing less than90% similarity.S3. SunXower NBS-LRR sequence database.S4. SunXower NBS-LRR SSCP and INDEL marker data-base.
References
Altschul SF, Madden TL, SchaVer AA, Zhang Z, Miller W, Lipman DJ(1997) Gapped BLAST and PSI-BLAST: a new generation ofprotein database search programs. Nucleic Acids Res 25:3389–3402
Ameline-Torregrosa C, Wang BB, O’Bleness MS, Deshpande S, ZhuH, Roe B, Young ND, Cannon SB (2007) IdentiWcation andcharacterization of nucleotide-binding site-leucine-rich repeatgenes in the model plant Medicago truncatula. Plant Physiol146:5–21
AshWeld T, Ong LE, Nobuta K, Schneider CM, Innes R (2004) Con-vergent evolution of disease resistance gene speciWcity in twoXowering plant families. Plant Cell 16:309–318
Baack EJ, Whitney KD, Rieseberg LH (2005) Hybridization andgenome size evolution: timing and magnitude of nuclear DNAcontent increases in Helianthus homoploid hybrid species. NewPhytol 167:623–630
Bai J, Pennill LA, Ning J, Lee SW, Ramalingam J, Webb CA, Zhao B,Sun Q, Nelson JC, Leach JE, Hulbert SH (2002) Diversity innucleotide binding site-leucine-rich repeat genes in cereals.Genome Res 12:1871–1884
Bassam BJ, Caetano-Anollés G, GresshoV PM (1991) Fast and sensi-tive silver staining of DNA in polyacrylamide gels. Anal Biochem196:80–83
Bent AF (1996) Plant disease resistance genes: function meets struc-ture. Plant Cell 8:1757–1771
Bert PF, Labrouhe DTD, Philippon J, Mouzeyar S, Jouan I, Nicolas P,Vear F (2001) IdentiWcation of a second linkage group carrying genescontrolling resistance to downy mildew (Plasmopara halstedii) insunXower (Helianthus annuus L.). Theor Appl Genet 103:992–997
Bouzidi MF, Badaoui S, Cambon F, Vear F, De Labrouhe DT, NicolasP, Mouzeyar S (2002) Molecular analysis of a major locus forresistance to downy mildew in sunXower with speciWc PCR-basedmarkers. Theor Appl Genet 104:592–600
Cannon SB, Shu H, Baumgarten AM, Spangler R, May G, Cook DR,Young ND (2002) Diversity, distribution, and ancient taxonomicrelationships within the TIR and non-TIR NBS-LRR resistancegene subfamilies. J Mol Evol 54:548–562
Chin DB, Arroyo-Garcia R, Ochoa OE, Kesseli RV, Lavelle DO,Michelmore RW (2001) Recombination and spontaneousmutation at the major cluster of resistance genes in lettuce(Lactuca sativa). Genetics 157:831–849
123

Mol Genet Genomics (2008) 280:111–125 123
Clamp M, CuV J, Searle SM, Barton GJ (2004) The Jalview Java align-ment editor. Bioinformatics 20:426–427
Collins NC, Webb CA, Seah S, Ellis JG, Hulbert SH, Pryor A (1998)The isolation and mapping of disease resistance gene analogs inmaize. Mol Plant Microbe Interact 11:968–978
Dangl JL, Jones JDG (2001) Plant pathogens and integrated defenseresponses to infection. Nature 411:826–833
Degener J, Melchinger AE, Hahn V (1999) InterspeciWc hybrids assource of resistance to Sclerotinia and Phomopsis in sunXowerbreeding. Helia 22:49–60
Don RH, Cox PT, Wainwright BJ, Baker K, Mattick JS (1991) ‘Touch-down’ PCR to circumvent spurious priming during gene ampliW-cation. Nucleic Acids Res 19:4008
Dußle CM, Hahn V, Knapp SJ, Bauer E (2004) PlArg from Helianthusargophyllus is unlinked to other known downy mildew resistancegenes in sunXower. Theor Appl Genet 109:1083–1086
Ellis J, Dodds P, Pryor T (2000) The generation of plant disease resis-tance gene speciWcities. Trends Plant Sci 5:373–379
Gandhi SD, Heesacker AF, Freeman CA, Argyris J, Bradford K,Knapp SJ (2005) The self-incompatibility locus (S) and quantita-tive trait loci for self-pollination and seed dormancy in sunXower.Theor Appl Genet 111:619–629
Gedil MA, Slabaugh MB, Berry S, Johnson R, Michelmore R, MillerJ, Gulya T, Knapp SJ (2001) Candidate disease resistance genesin sunXower cloned using conserved nucleotide-binding site mo-tifs: genetic mapping and linkage to the downy mildew resistancegene Pl1. Genome 44:205–212
Gentzbittel L, Mouzeyar S, Badaoui S, Mestries E, Vear F, Tourvieillede Labrouhe D, Nicolas P (1998) Cloning of molecular markersfor disease resistance in sunXower, Helianthus annuus L. TheorAppl Genet 96:519–525
GoV SA, Ricke D, Lan TH, Presting G, Wang R, Dunn M, GlazebrookJ, Sessions A, Oeller P, Varma H, Hadley D, Hutchison D, MartinC et al (2002) A draft sequence of the rice genome (Oryza sativaL. ssp. japonica). Science 296:79–92
Gonzales MD, Archuleta E, Farmer A, Gajendran K, Grant D, Shoe-maker R, Beavis WD, Waugh ME (2005) The Legume Informa-tion System (LIS): an integrated information resource forcomparative legume biology. Nucleic Acids Res 33:D660–D665
Hammond-Kosack KE, Jones JDG (1997) Plant disease resistancegenes. Ann Rev Plant Phys Plant Mol Biol 48:575–607
He L, Du C, Covaleda L, Xu Z, Robinson AF, Yu JZ, Kohel RJ, ZhangHB (2004) Cloning, characterization, and evolution of the NBS-LRR-encoding resistance gene analogue family in polyploid cot-ton (Gossypium hirsutum L.). Mol Plant Microbe Interact17:1234–1241
Hecker KH, Roux KH (1996) High and low annealing temperatures in-crease both speciWcity and yield in touchdown and stepdownPCR. Biotechniques 20:478–485
Holt BF, Hubert DA, Dangl JL (2003) Resistance gene signaling inplants: complex similarities to animal innate immunity. Curr OpinImmunol 15:20–25
Hulbert SH, Webb CA, Smith SM, Sun Q (2001) Resistance gene com-plexes: evolution and utilization. Ann Rev Phytopathol 39:285–312
Jones JD, Dangl JL (2006) The plant immune system. Nature 444:323–329
Kanazin V, Marek LF, Shoemaker RC (1996) Resistance gene analogsare conserved and clustered in soybean. Proc Natl Acad Sci USA93:11746–11750
Kohler A, Rinaldi C, Duplessis S, Baucher M, Geelen D, DuchaussoyF, Meyers BC, Boerjan W, Martin F (2008) Genome-wide identi-Wcation of NBS resistance genes in Populus trichocarpa. PlantMol Biol 66:619–636
Kuang H, Woo SS, Meyers BC, Nevo E, Michelmore RW (2004) Mul-tiple genetic processes result in heterogeneous rates of evolution
within the major cluster disease resistance genes in lettuce. PlantCell 16:2870–2894
Kukita Y, Tahira T, Sommer SS, Hayashi K (1997) SSCP analysis oflong DNA fragments in low pH gel. Hum Mutat 10:400–407
Lander ES, Green P, Abrahamson J, Barlow A, Daly MJ, Lincoln SE,Newburg L (1987) MAPMAKER: an interactive computer pack-age for constructing primary genetic linkage maps of experimen-tal and natural populations. Genomics 25:511–557
Langar K, Griveau Y, Kaan F, Serieys H, Varès D, Bervillé A (2002)Evaluation of parameters accounting for Phomopsis resistanceusing natural infection and artiWcial inoculation on recombinantinbred lines from a cross between susceptible and resistantsunXower. Eur J Plant Path 108:307–315
Larsen LA, Christiansen M, Vuust J, Andersen PS (1999) High-throughput single-strand conformation polymorphism analysis byautomated capillary electrophoresis: robust multiplex analysisand pattern-based identiWcation of allelic variants. Hum Mutat13:318–327
Larsen LA, Jespersgaard C, Andersen PS (2007) Single-strand confor-mation polymorphism analysis using capillary array electrophore-sis for large-scale mutation detection. Nat Protoc 2:1458–1466
Lawrence GJ, Finnegan EJ, AyliVe MA, Ellis JG (1995) The L6 genefor Xax rust resistance is related to the Arabidopsis bacterial resis-tance gene RPS2 and the tobacco viral resistance gene N. PlantCell 7:1195–1206
Lawson RR, Goulter KC, Henry RJ, Kong GA, Kochman JK (1998)Marker-assisted selection for two rust resistance genes in sun-Xower. Mol Breed 4:227–234
Lee SY, Seo JS, Rodriguez-Lanetty M (2003) Comprative analysis ofsuperfamilies of NBS-encoding disease resistance gene analogs incultivated and wild apple species. Mol Gen Genomics 269:101–108
Leister D, Ballvora A, Salamini F, Gebhardt C (1996) A PCR-basedapproach for isolating pathogen resistance genes from potato withpotential for wide application in plants. Nat Genet 14:421–429
Leister D, Kurth J, Laurie DA, Yano M, Sasaki T, Devos K, Graner A,Schulze-Lefert P (1998) Rapid reorganization of resistance genehomologues in cereal genomes. Proc Natl Acad Sci USA 95:370–375
Lenardon SL, Bazzalo ME, Abratti G, Cimmino CJ, Galella MT,Grondona M, Giolitti F, Leon AJ (2005) Screening sunXower forresistance to sunXower chlorotic mottle virus and mapping theRcmo-1 resistance gene. Crop Sci 45:735–739
Lincoln SE, Lander ES (1992) Systematic detection of errors in geneticlinkage data. Genomics 14:604–610
Madsen LH, Collins NC, Rakwalska M, Backes G, Sandal N, KrusellL, Jensen J, Waterman EH, Jahoor A, AyliVe M, Pryor AJ, Lang-ridge P, Schulze-Lefert P, Stougaard J (2003) Barley diseaseresistance gene analogs of the NBS-LRR class: identiWcation andmapping. Mol Genet Genomics 269:150–161
McDowell JM, Dhandaydham M, Long TA, Aarts MG, GoV S, HolubEB, Dangl JL (1998) Intragenic recombination and diversifyingselection contribute to the evolution of downy mildew resistanceat the RPP8 locus of Arabidopsis. Plant Cell 10:1861–1874
McHale L, Tan X, Koehl P, Michelmore RW (2006) Plant NBS-LRRproteins: adaptable guards. Genome Biol 7:212
Meyers BC, Shen KA, Rohani P, Gaut BS, Michelmore RW (1998a)Receptor-like genes in the major resistance locus of lettuce aresubject to divergent selection. Plant Cell 10:1833–1846
Meyers BC, Chin DB, Shen KA, Sivaramakrishnan S, Lavelle DO,Zhang Z, Michelmore RW (1998b) The major resistance genecluster in lettuce is highly duplicated and spans several megabas-es. Plant Cell 10:1817–1832
Meyers BC, Dickerman AW, Michelmore RW, Sivaramakrishnan S,Sobral BW, Young ND (1999) Plant disease resistance genesencode members of an ancient and diverse protein family withinthe nucleotide-binding superfamily. Plant J 20:317–332
123

124 Mol Genet Genomics (2008) 280:111–125
Meyers BC, Morgante M, Michelmore RW (2002) TIR-X andTIR-NBS proteins: two new families related to disease resistanceTIR-NBS-LRR proteins encoded in Arabidopsis and other plantgenomes. Plant J 32:77–92
Meyers BC, Kozik A, Griego A, Kuang H, Michelomore RW (2003)Genome-wide analysis of NBS-LRR-encoding genes in Arabid-opsis. Plant Cell 15:809–834
Michelmore RW (2003) The impact zone: genomics and breeding fordurable disease resistance. Curr Opin Plant Biol 6:397–404
Michelmore RW, Meyers BC (1998) Clusters of resistance genes inplants evolve by divergent selection and a birth- and- death pro-cess. Genome Res 8:1113–1130
Miller JF, Gulya TJ (1988) Registration of six downy mildew resistantsunXower germplasm lines. Crop Sci 28:1040–1041
Miller JF, Gulya TJ (1991) Inheritance of resistance to race 4 of downymildew derived from interspeciWc cross in sunXower. Crop Sci31:40–43
Miller JF, Gulya TJ, Seiler GJ (2002) Registration of Wve fertilityrestorer sunXower germplasms. Crop Sci 42:989
Murray MG, Thompson WR (1980) Rapid isolation of high molecularweight plant DNA. Nucleic Acids Res 8:4321–4325
Noel L, Moores TL, van Der Biezen EA, Parniske M, Daniels MJ,Parker JE, Jones JD (1999) Pronounced intraspeciWc haplotypedivergence at RPP5 complex disease resistance locus of Arabid-opsis. Plant Cell 11:2099–2112
Noir S, Combes MC, Anthony F, Lashermes P (2001) Origin, diversityand evolution of NBS-type disease-resistance gene homologuesin coVee trees (CoVea L.). Mol Gent Genomics 265:654–662
Orita M, Iwahana H, Kanazawa H, Hayashi K, Sekiya T (1989) Detec-tion of polymorphisms of human DNA by gel electrophoresis assingle-strand conformation polymorphisms. Proc Natl Acad SciUSA 86:2766–2770
Pan Q, Liu YS, Budai-Hadrian O, Sela M, Carmel-Goren L, Zamir D,Fluhr R (2000) Comparative genetics of nucleotide binding site-leucine rich repeat resistance gene homologues in the genomes oftwo dicotyledons: tomato and arabidopsis. Genetics 155:309–222
Parker C, Riches CR (1993) Parasitic weeds of the world: biology andcontrol. CAB Int, Wallingford
Parniske M, Hammond-Kosack KE, Golstein C, Thomas CM, JonesDA, Harrison K, WulV BB, Jones JD (1997) Novel diseaseresistance speciWcities result from sequence exchange betweentandemly repeated genes at the Cf-4/9 locus of tomato. Cell91:821–832
Pérez-Vich B, Akhtouch B, Knapp SJ, Leon AJ, Velasco L, Fernández-Martínez JM, Berry ST (2004) Quantitative trait loci for broom-rape (Orobanche cumana Wallr.) resistance in sunXower. TheorAppl Genet 109:92–102
Plocik A, Layden J, Kesseli R (2004) Comparative analysis of NBS do-main sequences of NBS-LRR disease resistance genes from sun-Xower, lettuce, and chicory. Mol Phylogen Evol 31:153–163
Pustovoit GV, Kroknin EY (1978) Inheritance of resistance in inter-speciWc hybrids of sunXower to downy mildew. Rev Plant Pathol57:209
Quresh Z, Jan CC, Gulya TJ (1993) Resistance of sunXower rust and itsinheritance in wild sunXower species. Plant Breed 110:297–306
Radwan O, Bouzidi MF, Vear F, Philippon J, Tourvieille de LabrouheD, Nicolas P, Mouzeyar S (2003) IdentiWcation of non-TIR-NBS-LRR markers linked to PL5/PL8 locus for resistance to downymildew in sunXower. Theor Appl Genet 106:1438–1446
Radwan O, Bouzidi MF, Nicolas P, Mouzeyar S (2004) Developmentof PCR markers for the Pl5/ Pl8 locus for resistance to Plasmo-para halstedii in sunXower, Helianthus annuus L. from completeCC-NBS-LRR sequences. Theor Appl Genet 109:176–185
Richly E, Kurth J, Leister D (2002) Mode of ampliWcation and reorga-nization of resistance genes during recent Arabidopsis thalianaevolution. Mol Biol Evol 19:76–84
Rossi M, Araujo PG, Paulet F, Garsmeur O, Dias VM, Chen H, VanSluys MA, D’Hont A (2003) Genomic distribution and character-ization of EST-derived resistance gene analogs (RGAs) in sugar-cane. Mol Genet Genomics 269:406–419
Saitou N, Nei M (1987) The neighbor-joining method: a new methodfor reconstructing phylogenetic trees. Mol Biol Evol 4:406–425
Seiler GJ (1991a) Registration of 15 interspeciWc sunXower germ-plasm lines derived from wild annual species. Crop Sci 31:1389–1390
Seiler GJ (1991b) Registration of 13 downy mildew tolerant interspe-ciWc sunXower germplasm lines derived from wild annual spe-cies. Crop Sci 31:1714–1716
Shen KA, Keyers BC, Islam-Faridi MN, Chin DB, Stelly DM, Michel-more RW (1998) Resistance gene candidate identiWed by PCRwith degenerate oligonucleotide primers map to clusters of resis-tance genes in lettuce. Mol Plant Microbe Interact 11:815–823
Kkoric D (1985) SunXower breeding for resistance to Diaporthe/Phomopsis helianthi. Helia 8:21–23
Slabaugh MB, Yu JK, Tang S, Heesacker A, Hu X, Lu G, Han F, Bid-ney D, Knapp SJ (2003) Haplotyping and mapping a large clusterof resistance gene candidates in sunXower using multilocus intronfragment length polymorphisms. Plant Biotech J 1:167–185
Sun Q, Collins NC, AyliVe M, Smith SM, Drake J, Pryor T, HulbertSH (2001) Recombination between paralogues at the Rp1 rustresistance locus in maize. Genetics 158:423–438
Tan AS, Jan CC, Gulya TJ (1992) Inheritance of resistance to race 4 ofsunXower downy mildew in wild sunXower accessions. Crop Sci32:949–952
Tang S, Yu JK, Slabaugh MB, Shintani DK, Knapp SJ (2002) Simplesequence repeat map of the sunXower genome. Theor Appl Genet105:1124–1136
Tang S, Heesacker A, Kishore VK, Fernandez A, Sadik E, Cole G,Knapp SJ (2003) Genetic mapping of the Or5 gene for resistanceto Orobanche race E in sunXower. Crop Sci 43:1021–1028
Tanksley SD, McCouch S (1997) Seed banks and molecular maps:unlocking the genetic potential from the wild. Science 277:1063–1066
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgens DG(1997) The CLUSTAL-X windows interface: Xexible strategiesfor multiple sequence alignment aided by quality analysis tools.Nucleic Acids Res 25:4876–4882
Vear F, De Labrouhe DT, Miller JF (2003) Inheritance of the wide-range downy mildew resistance in the sunXower line RHA 419.Helia 26:19–24
Vranceanu AV, Tudor VA, Stoenescu FM, Pirvu N (1980) Virulencegroups of Orobanche cumana Wallr., diVerent hosts and resistancesources and genes in sunXower. In: Proceedings of the 9th Interna-tional SunXower Conference, Torremolinos, Spain, pp 74–82
Viguié A, Labrouhe DTD, Vear F (2000) Inheritance of several sourc-es of resistance to Phomopsis stem canker (Diaporthe helianthiMunt.-Cvet.) in sunXower (Helianthus annuus L.). Euphytica116:167–179
Wang Z, Taramino G, Yang D, Liu G, Tingey SV, Miao GH, Wang GL(2001) Rice ESTs with disease-resistance gene or defense-re-sponse gene-like sequences mapped to regions containing majorresistance genes or QTLs. Mol Gen Genet 265:302–310
Wei FS, Wong RA, Wise RP (2002) Genome dynamics and evolutionof the Mla (powdery mildew) resistance locus in barley. Plant Cell14:1903–1917
Whitham S, Dinesh-Kumar SP, Choi D, Hehl R, Corr C, Baker B(1994) The product of tobacco mosaic virus resistance gene N:similarity to Toll and interleukin-1 receptor. Cell 78:1101–1115
Yu T, Buss GR, Saghai-Maroof MA (1996) Isolation of the super fam-ily of candidate disease-resistance genes in soybean based on aconserved nucleotide-binding site. Proc Natl Acad Sci USA93:11751–11756
123

Mol Genet Genomics (2008) 280:111–125 125
Yu JK, Tang S, Slabaugh MB, Heesacker AF, Cole G, Herring M, Sop-er J, Han F, Chu WC, Webb DM, Thompson L, Edwards KJ, Ber-ry ST, León AJ, Olungu C, Maes N, Knapp SJ (2003) Towards asaturated molecular genetic linkage map for cultivated sunXower.Crop Sci 43:367–387
Zhu H, Cannon SB, Young ND, Cook DR (2002) Phylogeny and geno-mic organization of the TIR and non-TIR NBS-LRR resistancegene family in Medicago truncatula. Mol Plant Microbe Interact15:529–539
123




















