
Novel homologs of the multiple resistance regulator marA in antibiotic-contaminated environments

Cockroach Homologs of Praying Mantis PeripheralAuditory System ComponentsDavid D. Yager*
Department of Psychology and Neuroscience and Cognitive Science Program, University of Maryland, College Park,Maryland 20742
ABSTRACT This study identifies the cuticular metatho-racic structures in earless cockroaches that are the ho-mologs to the peripheral auditory components in theirsister taxon, praying mantids, and defines the nature ofthe cuticular transition from earless to eared in the Dic-tyoptera. The single, midline ear of mantids comprises anauditory chamber with complex walls that contain thetympana and chordotonal transduction elements. The cor-responding area in cockroaches, between the furcaster-num and coxae, has many socketed hairs arranged indiscrete fields and the Nerve 7 chordotonal organ, thehomolog of the mantis tympanal organ. The Nerve 7 chor-dotonal organ attaches at the apex of the lateral ventro-pleurite (LVp), which has the same shape and generalstructure as an auditory chamber wall. High-speed videoshows that when the coxa moves toward the midline, theLVp rotates medially to stimulate socketed hairs, and alsomoves like a triangular hinge giving the chordotonal or-gan maximal in–out stimulation. Formation of the mantisauditory chamber from the LVp and adjacent structureswould involve only enlargement, a shift toward the mid-line, and a mild rotation. Almost all proprioceptive func-tion would be lost, which may constitute the major cost ofbuilding and maintaining the mantis ear. Isolation fromleg movement dictates the position of the mantis ear in themidline and the rigid frame, formed by the cuticularknobs, which protects the chordotonal organs. J. Morphol.000:000–000, 2005. © 2005 Wiley-Liss, Inc.
KEY WORDS: insect hearing; evolution; proprioceptor;insect ear; chordotonal organ; tympanum
In part because of their immense diversity, insectauditory systems offer an opportunity to understandthe basic processes that can give rise to “new” sen-sory systems. Such studies are hampered by a fossilrecord of insect ears that is sketchy at best, and evenwhere it exists (Rust et al., 1999), soft tissue compo-nents such as the nervous system are rarely pre-served. Comparative studies (Harvey and Pagel,1991) can help offset this deficit. A traditional strat-egy has been to identify and compare homologousanatomical components of the auditory system in arange of species and then integrate the results withinformation on the phylogenetic relationships of thespecies (acridids: Mason, 1968; Riede et al., 1990;ensifera: Otte, 1990; Gwynne, 1995; moths: Yack etal., 1999; Gopfert et al., 2001; Minet and Surlykke,
2003; tachinid flies: Edgecomb et al., 1995; Lakes-Harlan et al., 1999; mantids: Yager, 1990, 1999a).Choice of appropriate comparison species is impor-tant, and most useful is a sister taxon without anauditory system (sister groups share an ancestralspecies not shared by any other taxon; Wiley, 1981).Comparisons among eared and earless serially ho-mologous regions within the same species can alsoprovide compelling evidence (van Staaden et al.,2003; Yager, 1996a; Yack et al., 1999), especiallywhen coupled with the definition of developmentalpaths of ears and ear precursors (Meier andReichert, 1991). More recently, molecular geneticshas provided a window on the evolution of the pri-mary sensory receptors of insect ears, chordotonalsensilla, and provides yet another level of compari-son (Merritt, 1997; Okabe and Okano, 1997; Eberl etal., 2000).
Praying mantids (Mantodea; Dictyoptera) havesensitive ultrasonic hearing mediated by a uniqueauditory system (reviewed in Yager, 1999a). An an-atomically and functionally single ear comprisingtwo tympana in a deep, cuticular auditory chamberis located in the ventral midline of the caudal met-athorax. Auditory transduction takes place in a pairof chordotonal organs attached on or near each tym-panum (Yager and Hoy, 1987; Yager and Michelsen,2001). These are a tympanal organ (TO) and bifidsensilla (BS) that have �45 and exactly two chor-dotonal sensilla, respectively. Their bipolar sensoryneurons send axons through the tympanal nerve(Nerve Root 7; N.7) to the metathoracic ganglion.Nerve 7 is completely afferent and bifunctional. Inaddition to the chordotonal component, branches ofthe nerve innervate cuticular sensilla (mostly sock-
Contract grant sponsor: National Institutes of Health, NationalInstitute on Deafness and Other Communication Disorders (NIH/NIDCD); Contract grant number: DC01382.
*Correspondence to: Dr. David D. Yager, Department of Psychologyand Neuroscience and Cognitive Science Program, University ofMaryland, College Park, MD 20742. E-mail: [email protected]
Published online inWiley InterScience (www.interscience.wiley.com)DOI: 10.1002/jmor.10343
JOURNAL OF MORPHOLOGY 000:000–000 (2005)
© 2005 WILEY-LISS, INC.

eted hairs) of the caudal and ventral metathorax(Yager and Scaffidi, 1993). One small lineage ofmantids is especially interesting because they haveevolved a second auditory system that responds to2–4 kHz, but not ultrasound (Yager, 1996a). Lo-cated in the mesothorax, its components are seriallyhomologous to those of the metathoracic auditorysystem and it has the same basic structure. Of atleast six bilaterally symmetrical pairs of auditoryinterneurons in the metathoracic ganglion, one ofthem, 501-T3, has been studied extensively in termsof its physiology (Yager and Hoy, 1989) and its in-volvement in triggering and/or controlling a com-plex, coordinated, but nondirectional evasive behav-ior (Triblehorn and Yager, 2002). The behaviorprotects flying mantids from capture by echolocat-ing, insectivorous bats (Yager et al., 1990; Trible-horn, 2003).
The unusual auditory system of the mantis posesa range of questions relevant to the evolution ofinsect auditory systems, e.g., Why at that specificlocation? Is location determined by specific precur-sors of the structural and/or neural components ofthe auditory periphery? Why a midline ear? Thefossil record for the Mantodea is sparse (Gratshevand Zherikhin, 1993; Grimaldi, 1997; Vrsansky,2002), and the ventral thorax is visible in only one ortwo relatively recent specimens in amber (Grimaldi,pers. commun.). These questions must therefore beaddressed through comparative studies. The mostimportant comparison, and the focus of this study, iswith cockroaches (Blattodea; Dictyoptera). The closeanatomical similarity between cockroaches andmantids is well established (Kristensen, 1995;Klass, 2001)—they differ largely in adaptations fortheir contrasting feeding styles. Cockroaches, how-ever, have no known acoustic behaviors requiringsensitive hearing above 5 kHz, and there is no ana-tomical evidence of a tympanate auditory system.The neurophysiological evidence in this study sub-stantiates the absence of tympanate hearing in cock-roaches. (The subgenual organs of cockroaches areextremely sensitive substrate vibration sensors thatcan also detect airborne sound at frequencies below5 kHz [Nelson, 1980; Shaw, 1994] over short dis-tances. This sensory system, however, is not compa-rable in location, structure, or function to the tym-panate ear of mantids.) The uncertainty about theexact phylogenetic relationships among the subor-ders within the Dictyoptera is ongoing (summarizedin Eggleton, 2001). The two best-supported hypoth-eses are ((Mantodea � Blattaria) � Isoptera)[Thorne and Carpenter, 1992; Grimaldi, 1997;Wheeler et al., 2001] and (Mantodea � (Blattaria �Isoptera)) with the placement of the primitive familyCryptocercidae in debate (Grandcolas, 1999; Nalepaand Bandi, 1999; Lo et al., 2000; Klass, 2001). Manystudies (Grandcolas, 1996; Klass, 2001) place theBlattidae, which contains our focal species, Peripla-
neta americana, at or near the base of cockroachphylogeny.
Thus, the core goal of this study is to determinewhat cuticular structures, if any, in Periplanetaamericana are homologous to the peripheral audi-tory system of mantids. A similar study focusing onthe mantis peripheral auditory neural structuresand their cockroach homologs will be reported sep-arately. This study includes comparisons with man-tis nymphs throughout their development (Yager,1996b), with serially homologous anatomy, and withmantids that are primarily or secondarily earless(Yager, 1990, 1992). The results show that P. ameri-cana has clear homologs on each side of the midlinewith the same shape, basic structure, and innerva-tion as the mantis auditory chamber wall. Thetransformation from cockroach structures to themantis ear appears to be very simple in the periph-ery: an enlargement, medial shift, and mild rotationto form the auditory chamber. This closely parallelsthe ontogenetic changes that lead from earless man-tis nymphs to eared adults.
MATERIALS AND METHODSAnimals
Mantids: focal species. Most of the mantis data come fromHierodula membranacea, Sphodromantis lineola, and S. aurea.These closely related (Mantidae; Paramantinae) and very similarspecies were selected because of their large size and ease ofrearing. In addition, the auditory system of H. membranacea hasbeen studied previously (Yager, 1990, 1996b). Male body lengthsare �65 mm. Because extensive physiological and behavioralinformation on the species is available, Parasphendale agrionina(Mantidae; Miomantinae) was included in some parts of thestudy. The four species have no significant differences in auditorystructure and function from Mantis religiosa (Yager and Hoy,1987) or from each other. Hierodula membranacea, S. lineola, andS. aurea have only very minor sexual dimorphism in their audi-tory physiology and in the structure of the metathorax. Paras-phendale agrionina has strong sexual dimorphism: males havenormal ears and hearing, but females have markedly reducedears and lowest thresholds �90 dB.
The mantids came from long-standing laboratory colonies. Forthe ontogeny studies, nymphs were isolated within 7 days ofhatching and were reared individually. Daily checks for moltedskins provided stage information. The mantids were misted withwater once or twice daily and fed size-appropriate food (fruit flies,houseflies, crickets of various sizes) twice each week. The colonyroom was maintained at 30°C during the day (13 h) and 22°C atnight (11 h) with 40–60% relative humidity.
Mantids: comparative species. Data for mantids other thanthe focal species come from three sources: 1) The metathorax of�60 species that were studied anatomically and physiologicallyin the laboratory over the last 20 years was photographed whilethe animal was alive. 2) These animals are part of a collection of�80 species (�500 specimens) preserved in 70% alcohol andstored in the laboratory. 3) Pinned museum specimens also pro-vided extensive anatomical data for 329 of the �430 mantisgenera (Yager, 1990). The taxonomy follows Ehrmann and Roy(2002).
Cockroaches: focal species. Our focal cockroach, Periplanetaamericana (Blattidae; Blattinae), is a medium-size species (bodylengths 30–40 mm) with minimal sexual dimorphism. They werereared at moderate density in 20-gallon garbage barrels contain-ing egg crating for shelters. Both water and food (Purina CatChow supplemented with flake fish food) were continuously avail-
2 D.D. YAGER
able. They were housed in the same room as the mantids, but themaximum daytime temperature was �30°C.
Cockroaches: comparative species. The other cockroachspecies used in the physiological portion of this study (Table 1)were acquired from commercial sources, from hobbyists, and fromthe cockroach facility at Virginia Polytechnic University (Blacks-burg, VA). They were housed under the same conditions as theother cockroaches, but were not kept in long-term culture.
PhysiologySurgery. After the mantis or cockroach was fixed ventral-
side-up to a wax/plastic block, the ventral nerve cord was exposedby removing a rectangle of cuticle just rostral (cockroach) orcaudal (mantis) to the prothoracic legs. The more rostral record-ing site in cockroaches was chosen to include any auditory infor-mation ascending to the brain from the prothoracic segment aswell as from the rest of the body. The surgical field was sur-rounded by a well built from dental wax that was kept filled withsaline (Strausfeld et al., 1983; Pichon, 1970; Adams and Wilcox,1972). The legs were removed from mantids at mid-coxa to elim-inate low-frequency subgenual responses.
Procedures. Physiological experiments were carried out in abox (2.0 � 0.8 � 0.8 m) lined with acoustic foam to suppress soundreflections. The preparation was positioned at the center of thebox, 0.7 m from speakers at both ends.
Ascending neural activity was recorded using a suction elec-trode placed over the caudal cut end of one of the connectives andstandard extracellular recording electronics. Responses weremonitored both visually on an oscilloscope screen and aurallythrough headphones.
Standard stimuli were 70 ms sound pulses with 3–5 ms rise/falltimes (Stanford Research DS345 function generator; Grass S11stimulator; Hewlett-Packard 350D attenuator; custom rise/fallgenerator). The interval between stimuli was kept long (3–5 sec)to minimize habituation. Playback in the range of 8–100 kHz wasthrough Panasonic EAS10TH400B speakers; for 1–8 kHz weused Radio Shack 40-1289A or 40-1011 speakers. Calibration ofthe stimulus presentation system used a Bruel & Kjaer 2231
sound level meter with a 4135 6.25 mm microphone (protectivegrid off) positioned above the wax/plastic block at the locationoccupied by the caudal metathorax during experiments. Calibra-tion tones were �300 ms long to assure adequate meter responsetime. The frequency response of the calibration system was flat�1 dB up to 70 kHz. For frequencies above 70 kHz, a correctioncurve was derived from the frequency responses of the sound levelmeter and the microphone. Speaker output was linear with re-spect to input up to 95 dB over the frequency range of interest. AStanford Research Systems SR760 FFT spectrum analyzer con-firmed that harmonics of the stimulus tones were �30 dB belowthe fundamental at all frequencies up to 50 kHz.
Data collection. Each animal received stimuli at 10 frequen-cies between 1 kHz and 100 kHz. The presentation order wassemirandom to prevent any effects of ascending or descendingfrequency sequences. For initial screening, stimuli were pre-sented at 80, then 90, and then 100 dB SPL. We determined athreshold (two responses out of three stimuli) for every frequencywhere there was a response.
If there was no consistent response to sound, both connectiveswere checked to be sure that the negative results were not theresult of damage on one side.
AnatomyGross anatomy. Several dyes including Janus green (Yack,
1993) facilitated dissections of the metathorax by providing im-proved contrast and some selectivity for nerves. An Auto-Montage image capture and analysis system (Syncroscopy)proved valuable for defining spatial relationships among smallerexternal structures. Digestion with a 1% trypsin solution followedby storage in glycerol yielded cleared specimens (Periplanetaamericana: n � 6; Sphodromantis sp.: n � 6) in which the par-tially transparent cuticular structure could be examined fromboth inside and outside. Counts of trichoid sensilla were donewith a dissecting microscope (Zeiss SV-11), and because some ofthe sensilla are small, may underestimate slightly the numbers ofthe smallest socketed hairs, especially from the dorsal, inner-
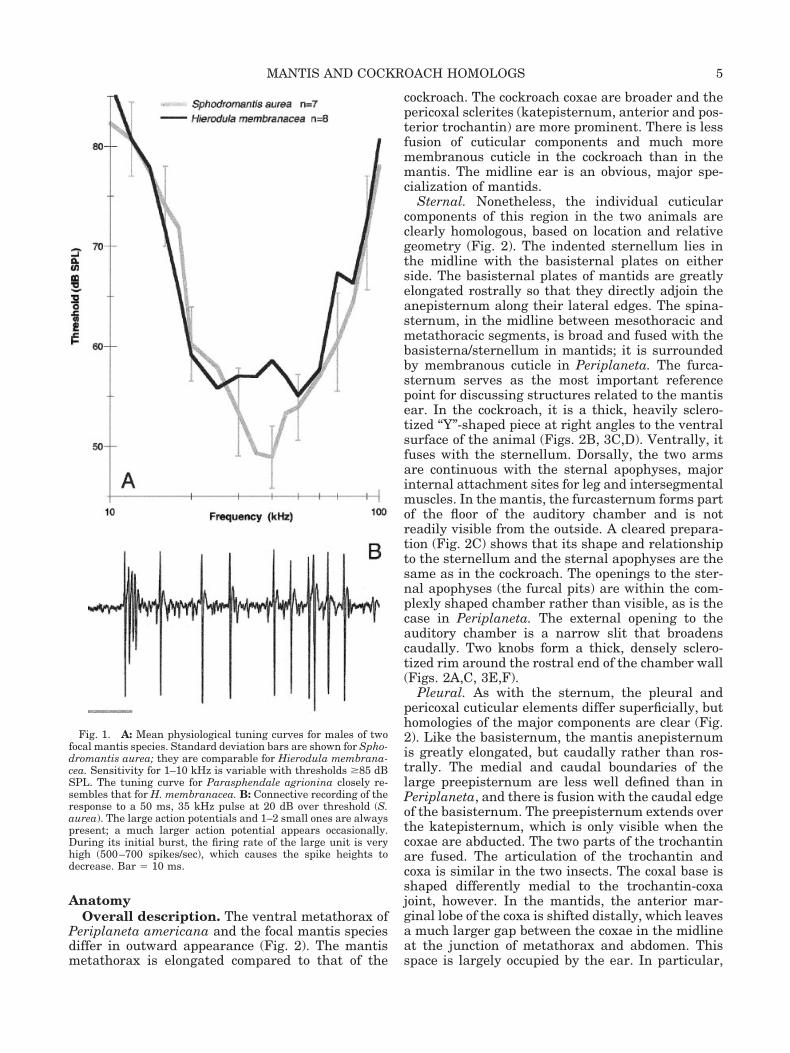
TABLE 1. Results of neurophysiological recordings to detect sound pressure reception in two mantidsand various species of cockroaches
Family Subfamily Genus/species
Number tested WingsResponse at
� 1 kHmale female male female
MantidaeMantinae Sphodromantis sp. �10 �10 ** ** **
Hierodula sp. �10 �10 ** ** **Polyphagidae
Polyphaginae Therea petiveriana 1 0 0 0 0Blattidae
Blattinae Periplaneta americana 5 1 ** ** 0Deropeltis sp. 2 0 ** 0 0
Polyzosteriinae Eurycotis floridana 6 2 0 0 0Melanozosteria soror 3 0 0 0 0
BlatellidaePlecopterinae Supella longipalps 2 0 ** * 0Blattellinae Blattella germanica 5 2 ** ** 0
Symploce hospice 2 0 ** 0 0Nyctiborinae Nyctibora lutzi 7 0 ** ** 0
BlaberidaeBlaberinae Blaberus sp. 6 3 ** ** 0Pycnoscelinae Pycnoscelis indicus 0 4 ** ** 0Diplopterinae Diploptera punctata 6 0 * * 0Panchlorinae Panchlora nivea 1 0 ** ** 0Oxyhaloinae Gromphadorhina portentosa 2 0 0 0 0
Nauphoeta cinerea 3 2 ** * 0
The legs were removed to eliminate subgenual responses. Data on the presence of wings are included because of the close associationof hearing with wings and lack of hearing with reduced wings in mantids (Yager, 1990).
3MANTIS AND COCKROACH HOMOLOGS

slanting cuticle. The nomenclature for the cuticular elementsprimarily follows Matsuda (1956, 1960, 1970).
Scanning electron microscopy. Preparation for scanning elec-tron microscopy included fixation (4% formaldehyde and 1% glutar-aldehyde in phosphate buffer), dehydration from ethanol, and coat-ing with gold/palladium. Tissue distortion was reduced by a finaldehydration from hexamethyldisilazane (HMDS) rather than theusual critical-point drying (Nation, 1983; Botes et al., 2002). TheAmray 1820 SEM provided calibrated digital and Polaroid images.
Because both the mantis and cockroach caudal metathoraxcontains areas of very thin cuticle, inconsistent folding artifactsand tissue distortion compromised interpretation of the SEMimages, especially for early instar mantis nymphs. Resin casts(Mercox, Ladd Industries, Burlington, VT) of the region allevi-ated the problem. This ultralow-viscosity resin shrinks minimallywith hardening (�2%) and provides extreme detail, even of struc-tures �1 � in size (Aharinejad et al., 1993). A dental wax wellsurrounding the caudal metathorax of a live animal was filledwith Mercox and the preparation was briefly placed under vac-uum before curing to free trapped air bubbles. SEM of the gold/palladium-coated casts showed no folding of the thin cuticle andalmost no distortion.
Histology. Serial and semiserial sections helped correlate thelocations of internal and external structures in both mantids andcockroaches. For paraffin sections (8 �m; Microm HM330 mic-rotome), the isolated caudal metathorax of freshly molted ani-mals was fixed in alcoholic Bouin’s for 1–3 days followed bydehydration. Slow dehydration in butanol (Lee, 1950; Barros-Pita, 1971) rather than the usual ethanol proved effective inkeeping sclerotized cuticle relatively soft. The Baker modificationof the Masson trichrome stain (Pantin, 1946) yielded excellentcolor contrast of different tissue types and selectively stains scolo-pale rods and caps a bright, cherry red. For plastic sections (4–6�m; Leica RM2165 microtome), the tissue was fixed as for SEM,embedded under vacuum in JB-4 (Polysciences, Warrington, PA),sectioned, and stained with toluidine blue, which colored scolo-pale caps a distinct robin’s egg blue.
Movement Studies
The structures of the cockroach and mantis caudal metathoraxare not normally static. High-speed video records of the regionaround the furcasternum during active and passive movement ofthe coxa showed the transient folds of the membranous cuticleand movements of the cuticular elements. The cockroach or man-tis was immobilized ventral-side-up with one leg fixed well awayfrom the midline. This positioning exposed most of the contralat-eral area between the furcasternum and coxa and allowed freemotion of the contralateral leg. In some cases, part of the anteriormarginal lobe of the free coxa that did not contact the membra-nous cuticle was removed to broaden the field of view. Illumina-tion was by high-intensity fiber optic light (Schott DCR III). Thecamera (Fastcam-X 1280 PCI, Photron), mounted on a ZeissSV-11 dissecting microscope, ran at 250 or 500 frames/sec. Thesespeeds gave a good compromise between temporal resolution andthe high illumination demands of the microscope/camera systemat adequate magnification (usually 50�). The free leg wasgripped with forceps or fingers and moved passively in a broadrange of directions. The animal actively moved its leg in responseto wind or tactile stimuli to the abdomen.
RESULTSNeurophysiology: Can Cockroaches Hear?
Summary of taxa tested. The mantis data comefrom two focal species in this study, Hierodula mem-branacea and Sphodromantis aurea, which arebroadly representative of at least 65% of all mantisspecies with respect to their auditory range andsensitivity (Yager, 1990).
The cockroach data come from 15 species represent-ing 11 of the 20 subfamilies, which in turn representall of the cockroach families except the Cryptocercidae(following McKittrick, 1964). The recordings focusedpreferentially on animals with long hindwings becauseof the strong correlation across several insect taxabetween flight and tympanate hearing (Mason, 1968;Otte, 1990; Yager, 1990). Sexual dimorphism of winglength is frequent among cockroach species, and there-fore the data is biased toward males, the winged sex inthose cases. One species in our sample, Pycnoscelusindicus, is parthenogenetic; the females have long,well-developed wings.
Cockroach and mantis displacement sensi-tivity. Both mantids and cockroaches have exquis-itely sensitive subgenual organs in the tibiae. Whenthe legs were intact, all species of both groupsshowed very robust multiunit responses to frequen-cies below 5 kHz with latencies as short as 10 ms.Thresholds rose progressively with increasing fre-quency above 1 kHz. In some species, weak respon-siveness extended up to 6–8 kHz before disappear-ing entirely. Removing the legs completely abolishedthe responses in almost all cases. Occasionally (par-ticularly in Blaberus sp.), low-amplitude “whoosh”responses to stimuli below 5 kHz and at �90 dB SPLremained after leg removal. These disappeared whenthe cerci were submerged in saline or removed.
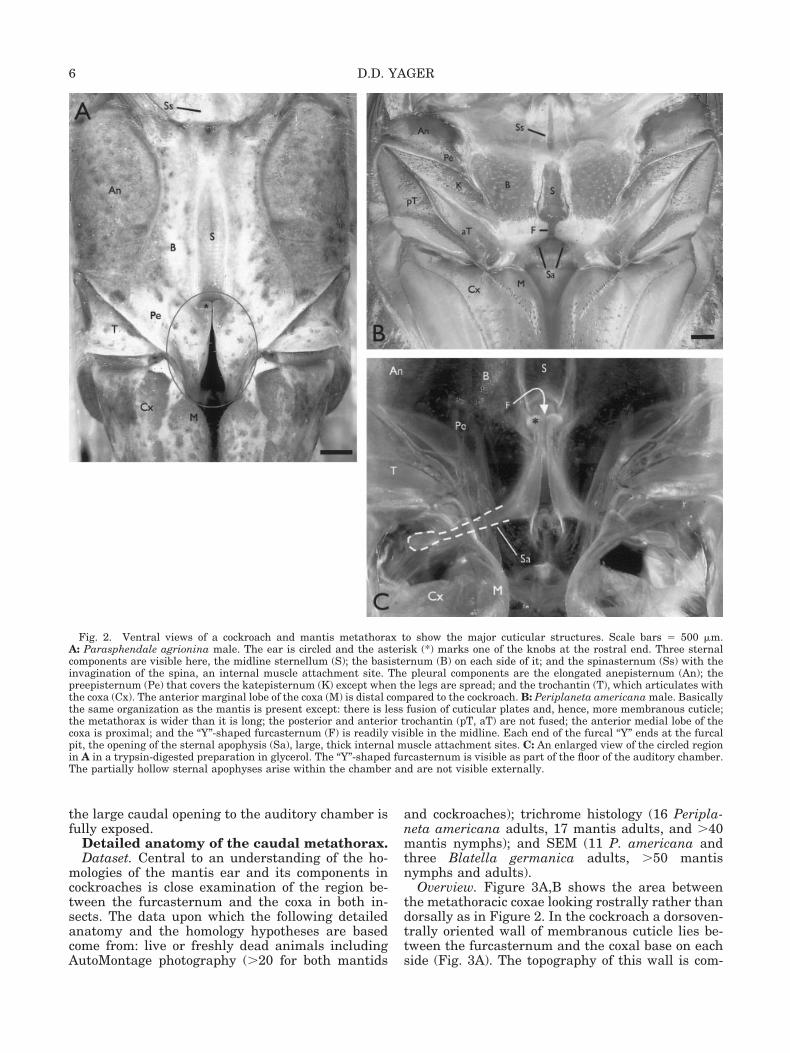
Pressure sensitivity.Mantis. Figure 1A shows mean tuning curves for
males of two focal species. Frequencies with highestsensitivity are typically 25–50 kHz and thresholdswere 40–60 dB SPL, depending on the individual.Females were often a few dB more sensitive withslightly lower best frequency range. The neural re-sponses (Fig. 1B) were always robust and dominatedby a single large unit with a distinctive phasic-tonicfiring pattern. There were consistently 1–3 addi-tional auditory units visible that had distinct actionpotential sizes and firing patterns. Shortest laten-cies were 12–15 ms. Removal of the legs and cercidid not alter the response at frequencies �10 kHz.Nonsubgenual hearing, i.e., legs off, below 10 kHzwas insensitive and highly variable among individ-uals. Transection of the connectives between themesothoracic and metathoracic ganglia eliminatedthe responses at �10 kHz, but not at lower frequen-cies.
Cockroach. The neural recordings made with legsremoved yielded uniform results across all of thecockroach species. There was no evidence of soundreception at frequencies of 1–100 kHz at SPLs �95dB (Table 1). No action potentials locked to the stim-uli, nor were there any audible “whooshes” thatwould indicate low-amplitude responses difficult todistinguish visually from baseline. There was theusual nonauditory activity in the connectives, andthe signal:noise ratio for the recordings was nor-mally �10:1, and always at least 5:1.
4 D.D. YAGER
AnatomyOverall description. The ventral metathorax of
Periplaneta americana and the focal mantis speciesdiffer in outward appearance (Fig. 2). The mantismetathorax is elongated compared to that of the
cockroach. The cockroach coxae are broader and thepericoxal sclerites (katepisternum, anterior and pos-terior trochantin) are more prominent. There is lessfusion of cuticular components and much moremembranous cuticle in the cockroach than in themantis. The midline ear is an obvious, major spe-cialization of mantids.
Sternal. Nonetheless, the individual cuticularcomponents of this region in the two animals areclearly homologous, based on location and relativegeometry (Fig. 2). The indented sternellum lies inthe midline with the basisternal plates on eitherside. The basisternal plates of mantids are greatlyelongated rostrally so that they directly adjoin theanepisternum along their lateral edges. The spina-sternum, in the midline between mesothoracic andmetathoracic segments, is broad and fused with thebasisterna/sternellum in mantids; it is surroundedby membranous cuticle in Periplaneta. The furca-sternum serves as the most important referencepoint for discussing structures related to the mantisear. In the cockroach, it is a thick, heavily sclero-tized “Y”-shaped piece at right angles to the ventralsurface of the animal (Figs. 2B, 3C,D). Ventrally, itfuses with the sternellum. Dorsally, the two armsare continuous with the sternal apophyses, majorinternal attachment sites for leg and intersegmentalmuscles. In the mantis, the furcasternum forms partof the floor of the auditory chamber and is notreadily visible from the outside. A cleared prepara-tion (Fig. 2C) shows that its shape and relationshipto the sternellum and the sternal apophyses are thesame as in the cockroach. The openings to the ster-nal apophyses (the furcal pits) are within the com-plexly shaped chamber rather than visible, as is thecase in Periplaneta. The external opening to theauditory chamber is a narrow slit that broadenscaudally. Two knobs form a thick, densely sclero-tized rim around the rostral end of the chamber wall(Figs. 2A,C, 3E,F).
Pleural. As with the sternum, the pleural andpericoxal cuticular elements differ superficially, buthomologies of the major components are clear (Fig.2). Like the basisternum, the mantis anepisternumis greatly elongated, but caudally rather than ros-trally. The medial and caudal boundaries of thelarge preepisternum are less well defined than inPeriplaneta, and there is fusion with the caudal edgeof the basisternum. The preepisternum extends overthe katepisternum, which is only visible when thecoxae are abducted. The two parts of the trochantinare fused. The articulation of the trochantin andcoxa is similar in the two insects. The coxal base isshaped differently medial to the trochantin-coxajoint, however. In the mantids, the anterior mar-ginal lobe of the coxa is shifted distally, which leavesa much larger gap between the coxae in the midlineat the junction of metathorax and abdomen. Thisspace is largely occupied by the ear. In particular,
Fig. 1. A: Mean physiological tuning curves for males of twofocal mantis species. Standard deviation bars are shown for Spho-dromantis aurea; they are comparable for Hierodula membrana-cea. Sensitivity for 1–10 kHz is variable with thresholds �85 dBSPL. The tuning curve for Parasphendale agrionina closely re-sembles that for H. membranacea. B: Connective recording of theresponse to a 50 ms, 35 kHz pulse at 20 dB over threshold (S.aurea). The large action potentials and 1–2 small ones are alwayspresent; a much larger action potential appears occasionally.During its initial burst, the firing rate of the large unit is veryhigh (500–700 spikes/sec), which causes the spike heights todecrease. Bar � 10 ms.
5MANTIS AND COCKROACH HOMOLOGS
the large caudal opening to the auditory chamber isfully exposed.
Detailed anatomy of the caudal metathorax.Dataset. Central to an understanding of the ho-
mologies of the mantis ear and its components incockroaches is close examination of the region be-tween the furcasternum and the coxa in both in-sects. The data upon which the following detailedanatomy and the homology hypotheses are basedcome from: live or freshly dead animals includingAutoMontage photography (�20 for both mantids
and cockroaches); trichrome histology (16 Peripla-neta americana adults, 17 mantis adults, and �40mantis nymphs); and SEM (11 P. americana andthree Blatella germanica adults, �50 mantisnymphs and adults).
Overview. Figure 3A,B shows the area betweenthe metathoracic coxae looking rostrally rather thandorsally as in Figure 2. In the cockroach a dorsoven-trally oriented wall of membranous cuticle lies be-tween the furcasternum and the coxal base on eachside (Fig. 3A). The topography of this wall is com-
Fig. 2. Ventral views of a cockroach and mantis metathorax to show the major cuticular structures. Scale bars � 500 �m.A: Parasphendale agrionina male. The ear is circled and the asterisk (*) marks one of the knobs at the rostral end. Three sternalcomponents are visible here, the midline sternellum (S); the basisternum (B) on each side of it; and the spinasternum (Ss) with theinvagination of the spina, an internal muscle attachment site. The pleural components are the elongated anepisternum (An); thepreepisternum (Pe) that covers the katepisternum (K) except when the legs are spread; and the trochantin (T), which articulates withthe coxa (Cx). The anterior marginal lobe of the coxa (M) is distal compared to the cockroach. B: Periplaneta americana male. Basicallythe same organization as the mantis is present except: there is less fusion of cuticular plates and, hence, more membranous cuticle;the metathorax is wider than it is long; the posterior and anterior trochantin (pT, aT) are not fused; the anterior medial lobe of thecoxa is proximal; and the “Y”-shaped furcasternum (F) is readily visible in the midline. Each end of the furcal “Y” ends at the furcalpit, the opening of the sternal apophysis (Sa), large, thick internal muscle attachment sites. C: An enlarged view of the circled regionin A in a trypsin-digested preparation in glycerol. The “Y”-shaped furcasternum is visible as part of the floor of the auditory chamber.The partially hollow sternal apophyses arise within the chamber and are not visible externally.
6 D.D. YAGER
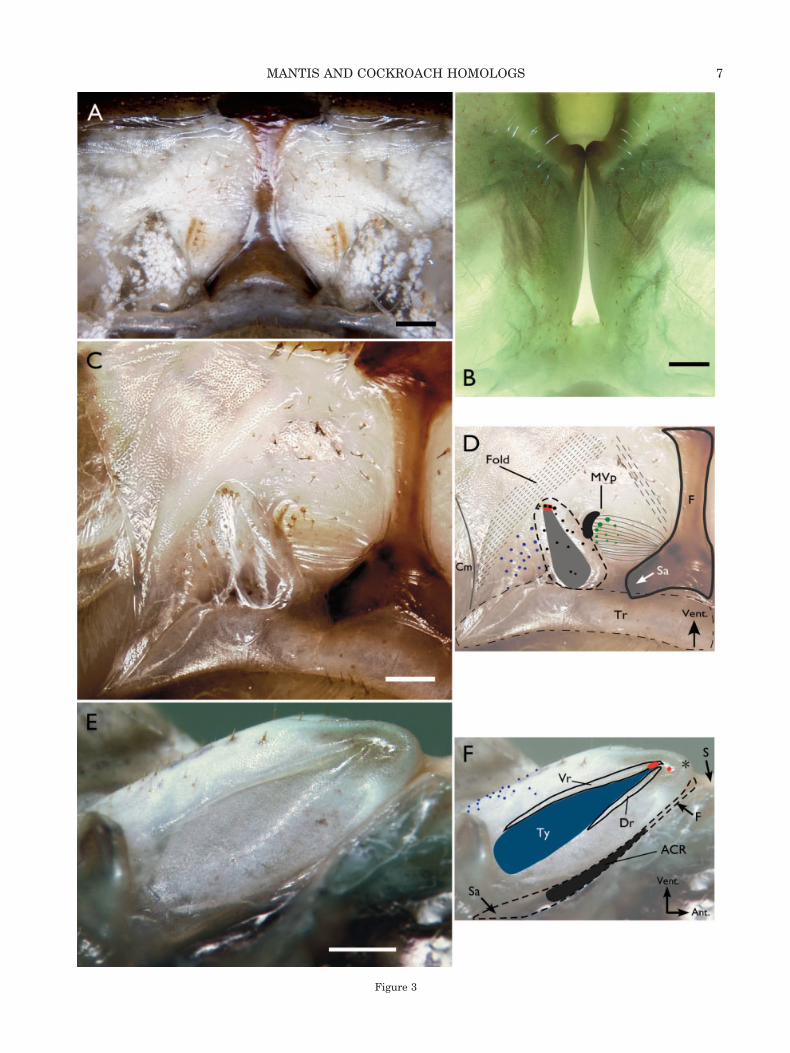
Figure 3
7MANTIS AND COCKROACH HOMOLOGS

plex, with several fields of socketed hairs (trichoidsensilla), discrete areas of cuticle of varying thick-ness, degree of sclerotization, and “bumpiness.” Fur-thermore, it is not in a single dorsoventral plane. Inthe mantis (Fig. 3B) the area between the coxal baseand the ear on each side consists almost entirely ofthin, smooth, transparent cuticle. The cuticle thick-ens somewhat just lateral to the opening of the au-ditory chamber. Rostrally, near the knobs there is amixture of stout and thin socketed hairs on rela-tively thick cuticle. Along the caudal border of thechamber opening on each side there is a field ofsmaller socketed hairs.
Cockroach. A more detailed examination of thecockroach’s dorsoventral wall shows six distinct re-gions (Figs. 3C,D, 4A).
1) A large ventromedial area just lateral to thefurcasternum: it is a roughly planar surface with aslight ventral tilt. The cuticle is thin with an aver-age of 24.2 (�2.6 SD; n � 9) socketed hairs of varioussizes. As in Figure 3C, the hairs in this region wereoften broken and/or bent even in specimens thatotherwise showed little wear. The “bumpy” appear-ance of the cuticle laterally in this region (and oth-ers) is caused by a large number of small, conicalcuticular spines (microtrichea). These are not hollowand do not appear to be innervated. In some areasthe microtrichea have a tiny hair-like projectionfrom their tips so that in an edge view the cuticlelooks like a very fine brush.
2) A very large ventrolateral area of thin, bumpycuticle usually separated from the previous area bya large flexion line: there may be a few socketedhairs near the flexion line (included in the count for
Region 1), but otherwise there are no distinctivefeatures. In some leg positions, Regions 1 and 2 arecontinuous.
3) An area of thin, smooth, transparent cuticle atthe base of the coxa: this is the cuticle of the coxaljoint, and is quite strong. When the coxa movestoward the midline (adducts) or elevates, this cuticlecompresses into folds. The boundary between Re-gion 3 and the adjacent areas is particularly distinctbecause of the clear flexion line and the very differ-ent cuticle appearance.
4) An area of thin cuticle that has a field of me-dium and small-size socketed hairs: its ventral por-tion is bumpy, but is smooth in its dorsal-medialportion.
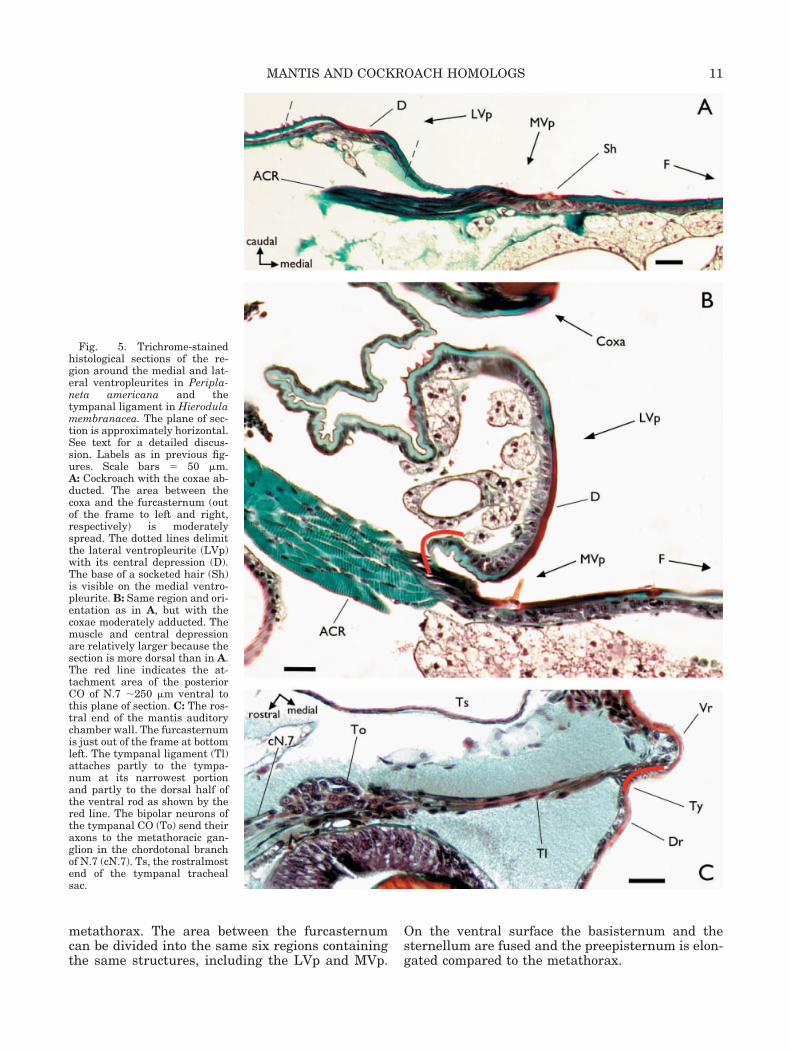
5) An oval region of complex structure, the lateralventropleurite (LVp): in a resting cockroach, it facesapproximately medially and somewhat caudally,and it bulges out from the surrounding area (Fig.2A,C). The length averages 716 � 44 �m (mean �SD; n � 5). The width is 198 � 16 �m at its greatestand 51.4 � 6.6 �m at the narrow apex. There is acentral, teardrop-shaped depression that is largelytransparent and completely smooth with the excep-tion of 3–5 trichoid sensilla (Figs. 3C,D, 7B). Thethickness of the cuticle in the depression is 35.6 �4.7 �m (12.3 � 5.4 �m without the epidermis). Lat-erally, the depression is bounded by a ridge ofbumpy cuticle that often has a row of socketed hairsalong its ventral half. The ridge falls off and mergeswith Region 4. Medially, the depression is boundedby a well-demarcated ridge. It has a few ventralsocketed hairs, not in a row, and some bumpiness onthe ventral half. The cuticle of this ridge adjacent toRegion 6 resembles that in Region 1 ventrally, and issmooth cuticle dorsally. The medial ridge is roundedinto a half-tube over most if its length, and theentire Region 5, especially the dorsal half, takes ona tube-like shape when the coxa is adducted (Fig.5A,B). At the ventral apex of the depression thereare always two or three large trichoid sensilla in atransverse row accompanied by two or three smallhairs. Regions 4 and 5 together have an average of23.8 � 3.9 (n � 9) socketed hairs of which 8–12 arearound or in the depression (Fig. 7B). The cuticleand epidermis around the apical hairs is thickenedto form a “pad” which is especially important be-cause it is the posterior attachment site for the chor-dotonal component of N.7, the tympanal nerve ho-molog (Yager and Scaffidi, 1993). Figure 5 showsdetails of Region 5’s fine structure from horizontaltrichrome sections. The cuticle of the depression islightly sclerotized; in some animals the ridges arealso sclerotized.
6) An area adjacent to the furcasternum that con-tains a large, organized hair plate, and a rectangle ofsclerotized cuticle: it is separated from Region 5 by astrip of cuticle �2 �m thick. The sclerotized cuticleis the ventropleurite discussed by Matsuda (1960). Itis not clear, however, whether he refers to only the
Fig. 3. The caudal end of the ventral metathorax lookingrostrally from the abdomen in a cockroach and a mantis. Labelingas in Figure 2. Scale bars � 300 �m. A: Overview of the region inPeriplaneta americana with the coxae abducted. The coxae arejust out of the photograph to right and left. The dominant land-mark is the central furcasternum. Lateral to it on each side is themedial ventropleurite and further lateral is the bulging lateralventropleurite. B: The same view as in A of the mantis Hierodulamembranacea emphasizing the overall difference between cock-roach and mantid in this region. The ear, with its two rostralknobs, dominates the picture. The furcasternum is deep withinthe auditory chamber and not visible. There is a field of small,socketed hairs on each side of the caudal end of the ear. Thesmooth cuticle extends from the base of the coxa to the ear.Trachea can be seen through the thin, almost transparent cuticlein this area. C: A close-up of the area between the cockroachfurcasternum and the coxa on one side. D: Schematic of thephotograph in C with labels. The light gray region is the centraldepression of the lateral ventropleurite. The red oval is the in-sertion site of the N.7 CO. The attachment of the anterior coxalrotator muscle is at the dark gray area, part of the medial ven-tropleurite (MVp). The dots are socketed hairs. Tr, tracheal tube;Cm, coxal membranous cuticle. E: The wall of the auditory cham-ber of a male Parasphendale agrionina. F: Schematic of thephotograph in E with labels. The red oval is the attachment siteof the tympanal CO and the red dot is the bifid sensillum. Vr,ventral rod; Dr, dorsal rod; Ty, tympanum; ACR, origin of ante-rior coxal rotator muscle.
8 D.D. YAGER

Fig. 4. Schematic diagrams based on the photo-graphs in Figure 3 with labeling as in that figure.A: Division of the area between the coxa and fur-casternum of the cockroach, Periplaneta americana,into six anatomical regions as described in the text.Region 6 (and Region 5, depending on leg position)face somewhat dorsally, whereas the other regionsface ventrocaudally. The flexion lines vary depend-ing on coxal position. The dashed arrows indicatecompression that forms folds of cuticle like an ac-cordion. The solid arrows indicate that movementcauses an enlarging, “rolling” fold of thin cuticle atthe flexion line. As the fold enlarges and moves, itbends strategically located socketed hairs. The solidwhite arrows indicate rotation. Region 5 normallyfaces medially, but with coxal adduction rotates toface rostrally. Region 4 initially faces mediocau-dally and faces medially with adduction. The blackoval is the attachment site for the N.7 CO. B: As inA, but for Hierodula membranacea. The attachmentsites for the tympanal CO (black oval) and the bifidsensilla (black dot) are not visible, but their relativepositions are indicated. C,D: Two hypotheses forthe homology of medial and lateral ventropleuritesand adjacent structures of the cockroach with com-ponents of the mantis’ auditory chamber wall usingthe six regions defined in A. See text for a detaileddiscussion.
9MANTIS AND COCKROACH HOMOLOGS

sclerotized rectangle and its associated hair plate, oralso includes Region 5. For this study the former willbe called the medial ventropleurite (MVp) to distin-guish it from the LVp. The rectangle of cuticle is theinsertion point of a long, thin muscle that originatesat the anterior rim of the coxa (ACR; Fig. 5A,B). Thismuscle is innervated at its coxal end by a smallbranch of Nerve 4. There are no other muscles in theimmediate area, and specifically none originatingfrom the furcasternum. The cuticle between theMVp and the furcasternum is deeply corrugated andfan-like. The hair plate comprises 4–5 tall trichoidsensilla in a curved row just medial to the muscleinsertion. The row is sometimes extended dorsallywith 2–3 much shorter sensilla. Medial to the row oftall hairs is a less organized field of shorter socketedhairs. The total number of hairs in Region 6 aver-ages 12.7 � 1.52 (n � 11).
The posterior branch of N.7 innervates the regionbetween the coxa and furcasternum. The branchtravels slightly dorsal and medial to N.7’s posteriorchordotonal ligament. It bifurcates just beforereaching the cuticle into a medial branch that inner-vates the hair plate and other medial sensilla and alateral branch that innervates the sensilla of Re-gions 4 and 5.
The area surrounding the base of the coxa and thefurcasternum is richly supplied with trachea. Thevery large transverse tracheal tube connecting thefirst abdominal spiracles runs along the base of thecaudal wall (Fig. 3A–D). It has several ventralbranches, including one to two consistently near theLVP.
Mantis. The anatomy of the caudal ventral met-athorax has been described previously in relation-ship to hearing (Yager and Hoy, 1987; Yager andScaffidi, 1993). For this study the basic structureand components will be described here, and the is-sue of homology hypotheses dealt with in detail be-low.
The cuticle between the base of the coxa and theear is thin and transparent (Fig. 3B). Its boundarieswith the thicker cuticle of the basisternum/preepisternum and the ear are distinct. As the coxaeadduct and/or elevate, this cuticle compresses intofolds. A prominent field of socketed hairs is alwayspresent medial to and sometimes extending onto thetransparent cuticle, near the caudal opening of theauditory chamber (Figs. 3B, 4B). The number ofsensilla varies somewhat by species. For Hierodulamembranacea males it averages 18.4 � 6.6 (n � 7)on each side.
All of the remaining area lateral to the furcaster-num faces medially and is tilted somewhat dorsally;it makes up the wall of the auditory chamber (Fig.3B,E,F). Two ridges bound the central teardrop-shaped depression (the tympanum). Most promi-nent, the ventral ridge is tube-like with thick cuticlewalls. The posterior ligament of the chordotonalcomponent of N.7 attaches on its inner surface at the
rostral end (Figs. 5C, 7A). The dorsal ridge is lesstube-like and generally less well defined, especiallyalong its caudal two-thirds. The central depressioncomprises thin, smooth cuticle devoid of hairs. Therostral end becomes very narrow (30–50 �m) andthen merges into the flat, bumpy cuticle within thecurve of the knob (Fig. 7A). The attachment site ofthe chordotonal ligament includes part of the centraldepression where it is narrowest. The areas ventralto the ventral rod and dorsal to the dorsal rod are“bumpy” cuticle of varying thickness, but alwaysmuch thicker than in the depression. The dorsalcuticle is without hairs and merges with the furca-sternum. The openings to the sternal apophyses(Figs. 2C, 4C) are in the floor of the chamber at thelevel of the caudal end of the central depression. Theventral cuticle is gently rounded to merge with theouter border of the ear. It has the caudal field ofsmall hairs described above, and a few longer,coarser hairs rostrally.
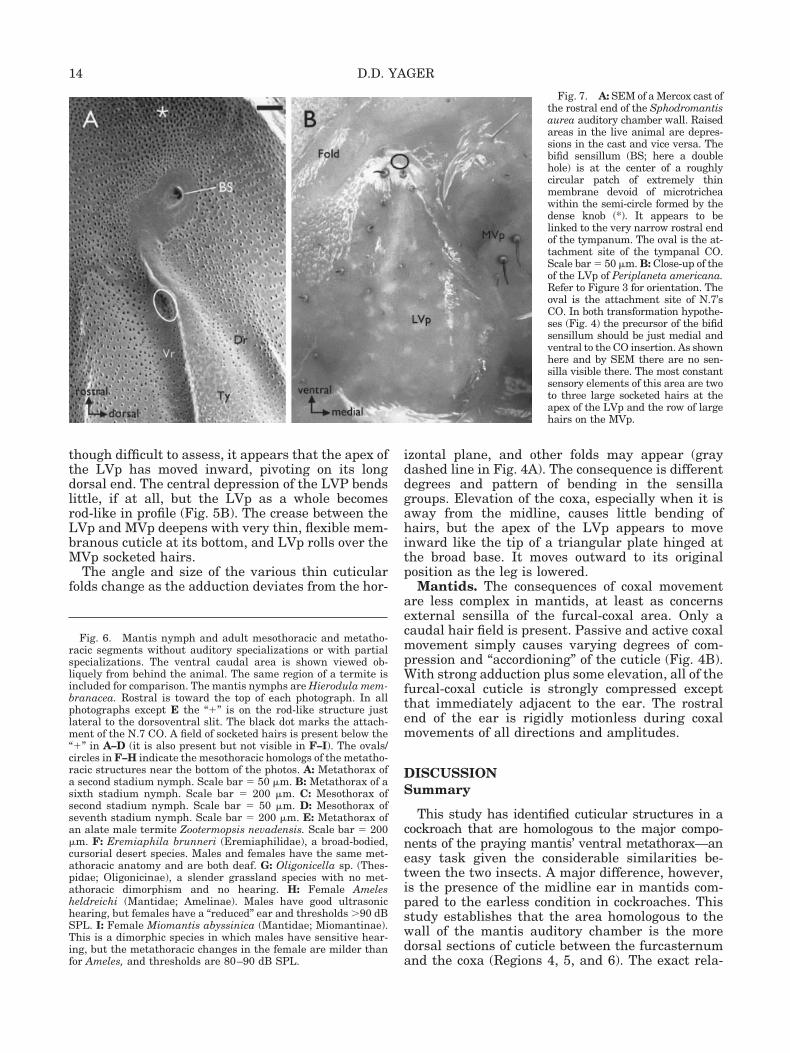
The rostralmost portion of the wall has a complexstructure (Figs. 3B,E,F, 7A). The extremely thick,highly sclerotized cuticle of the knob forms a semi-circle that shelters thin cuticle with large micro-trichea. Dorsal and slightly rostral to the ends of therods is a 30–50-�m diameter circle of extremely thin(�1 �m) cuticle. Approximately at the center of thecircle is the bifid sensillum described in greater de-tail below. The knobs themselves appear, at least inpart, to be a caudal continuation of the lateral wallsof the sternellum.
The lateral border of the furcasternum is the ori-gin for a long, thin muscle that inserts onto thecaudal rim of the coxa just medial to the trochantin-coxa joint (Figs. 3F, 4C). Levereault (1936) identifiedit as the anterior rotator of the coxa (muscle 98). Themuscle appears to have two heads with slightly off-set insertions onto the coxa. It is innervated near itscoxal end by a small branch of Nerve 4. There are noother muscles with origins on the furcasternum orattaching to the furcal-coxal cuticle.
A tracheal air sac is tightly apposed to the centraldepression of the chamber wall and adjacent dorsaland ventral cuticle over the caudal three-quarters ofits length. The air sac is an expansion of a ventralbranch from the large transverse trachea runningbetween the first abdominal spiracles.
The posterior branches of N.7 provide innervationto the sensilla in the region. These axons travel withthe chordotonal portion of N.7 before splitting off asone to three branches headed to the ipsilateral ven-tral cuticle. Like cockroaches, mantis species varywidely in the number and length of sensory hairs onthe sternum.
Comparisons. Establishing homology of cock-roach structures with components of the mantis earrequires comparisons across body segments, ontoge-netic stages, and taxa.
Cockroaches. For Periplaneta americana the anat-omy of the ventral mesothorax is organized like the
10 D.D. YAGER
metathorax. The area between the furcasternumcan be divided into the same six regions containingthe same structures, including the LVp and MVp.
On the ventral surface the basisternum and thesternellum are fused and the preepisternum is elon-gated compared to the metathorax.
Fig. 5. Trichrome-stainedhistological sections of the re-gion around the medial and lat-eral ventropleurites in Peripla-neta americana and thetympanal ligament in Hierodulamembranacea. The plane of sec-tion is approximately horizontal.See text for a detailed discus-sion. Labels as in previous fig-ures. Scale bars � 50 �m.A: Cockroach with the coxae ab-ducted. The area between thecoxa and the furcasternum (outof the frame to left and right,respectively) is moderatelyspread. The dotted lines delimitthe lateral ventropleurite (LVp)with its central depression (D).The base of a socketed hair (Sh)is visible on the medial ventro-pleurite. B: Same region and ori-entation as in A, but with thecoxae moderately adducted. Themuscle and central depressionare relatively larger because thesection is more dorsal than in A.The red line indicates the at-tachment area of the posteriorCO of N.7 �250 �m ventral tothis plane of section. C: The ros-tral end of the mantis auditorychamber wall. The furcasternumis just out of the frame at bottomleft. The tympanal ligament (Tl)attaches partly to the tympa-num at its narrowest portionand partly to the dorsal half ofthe ventral rod as shown by thered line. The bipolar neurons ofthe tympanal CO (To) send theiraxons to the metathoracic gan-glion in the chordotonal branchof N.7 (cN.7). Ts, the rostralmostend of the tympanal trachealsac.
11MANTIS AND COCKROACH HOMOLOGS

Periplaneta americana nymphs (0.3–0.5 adultsize; n � 4) differ from adults only in having some-what fewer sensory hairs in the furcal areas of thetwo thoracic segments. LVp and MVp are presentand oriented as in adults.
All the species of cockroach in which the caudalmetathorax was examined closely, Deropeltis sp.,Blatella germanica, Blaptica dubia, Panchloranivea, and Therea petiveriana, had the same basicorganization as Periplaneta americana. The relativesizes of MVp and LVp to the rest of the furcal-coxalarea were not the same, however: T. petiveriana hadthe smallest and P. nivea the largest. The number ofsocketed hairs varied widely. Therea petiverianaand B. germanica had few; B. dubia had 10–20interspersed with at least two other hair types ofvarious sizes.
Mantids. The mesothoracic and metathoracicanatomy across the full range of earless mantidsexamined (representatives in �149 genera) uni-formly resembles that in first and second stadiumnymphs, which, in turn, mirrors the cockroach pat-tern (Fig. 6). In the youngest nymphs this patterncomprises (lateral to medial): smooth, transparentpericoxal cuticle; a prominent dorsal field of sock-eted hairs bounded ventrally by thin, “bumpy” cuti-cle; a dorsoventrally oriented rod-like component; anarrow depression that looks like a slit when viewedfrom the caudal or ventral direction; smooth, thickcuticle that merges with the furcasternum. The at-tachment site of the N.7 chordotonal organ (CO) is atthe ventral edge of the rod. Direct examination,trichrome histology, and SEM show no external sen-silla in the region other than the caudal, dorsal hairfield.
The transitions from the anatomy shown in Fig-ure 6A to that in Figure 6B for the metathorax andfrom the anatomy shown in Figure 6C to that inFigure 6D for the mesothorax demonstrate that thedevelopmental starting point for both segments isthe same. In eared species, the metathoracic wallsfold medially and rotate over the remaining 6–8stadia to form the auditory chamber (Fig. 6B). Themesothoracic anatomy does not change appreciablyduring nymphal development (Fig. 6D).
The major difference between eared and earlessmantids is that in the latter both the mesothoracic(Fig. 6C,D) and the metathoracic region retain theearly structure throughout development. Examplesof earless mantids with no auditory sensitivity areshown in Figure 6F–H. The Miomantis abyssinicafemale (Fig. 6I) has auditory thresholds of 80–90 dBSPL, but even in this mild hearing reduction andaccompanying structural change, the nymphal (andcockroach) pattern is clear. The museum studiescovering 329 mantis genera indicate that the speciesof Figure 6F–I are secondarily earless (Yager, 1990).The largely plesiomorphic genera Chaeteesa andMantoida are most probably primitively earless.Specimens are not common, but 5–10 pinned indi-
viduals of each genus showed metathoracic anatomyessentially identical to the mesothoracic anatomy ofOligonicella sp. and female Ameles heldreichi (Fig.6G,H).
Bifid sensillum. The bifid sensilla are located atthe rostral ends of the auditory chamber walls (Figs.3E,F, 4C,D, 7A) dorsal and slightly rostral to the endof the ventral rod and the tympanum. Two scolopo-phorous sensilla innervate each bifid sensillum. Inthird and fourth stadium nymphs, SEMs andtrichrome sections show two discrete, adjacent sen-sory structures at the appropriate location.Trichrome and JB-4 histology confirms the presenceof two scolopophorous sensilla attaching to the innersurface of the cuticle at that site in first instar Spho-dromantis nymphs.
If a corresponding sensory structure exists inadult cockroaches, both transition models (Fig.4C,D, and Discussion) predict that it would be lo-cated medial and slightly ventral to the insertionsite of the N.7 CO. Direct observation of live speci-mens (n � 10), SEM examination (n � 8), andtrichrome sections (n � 9) all failed to find anycandidate surface or internal structure in that posi-tion. The nearest sensilla are the apical hairs of theLVp and small socketed hairs in Regions 1 and 2.
Function
Although it is possible to infer from the anatomygeneral movements that might stimulate the differ-ent sensilla groups between the furcasternum andthe coxae, the situation during normal behavior islikely to be complex. High-speed video observation ofthe region during passive and active motion of thecoxae provides a first step in understanding thefunction of the sensilla and, from this, the nature ofthe transition to auditory transduction. The intentat this stage was a descriptive rather than quanti-tative analysis.
Cockroaches. Abduction of the coxa stretchesand flattens the furcal-coxal cuticle of Regions 3–6(Fig. 5A). Bending is eliminated in all but a few ofthe most ventral socketed hairs nearest the majorcuticular fold (Figs. 3C, 4A, 7B). Movement towardthe midline causes compression in Region 3, result-ing in some displacement of hairs in Region 4. Themajor cuticular fold progressively rolls down overthe apex of the LVp. For intermediate adduction, afew of the LVp apical hairs are bent; Regions 4 and5 have rotated so that the latter faces medial andslightly rostral. Bending of the hairs of the MVp isminimal. With extreme adduction of the coxa, espe-cially with the medial border moving ventrally aswell, the major fold mostly covers the LVp apex.Regions 5 and 6 rotate to face each other and evenpress together. A small, discrete fold that formsabove the ventral end of the muscle insertion alsostimulates the hairs of MVp. Most or all of the sen-silla in Regions 4–6 are bent in that position. Al-
12 D.D. YAGER
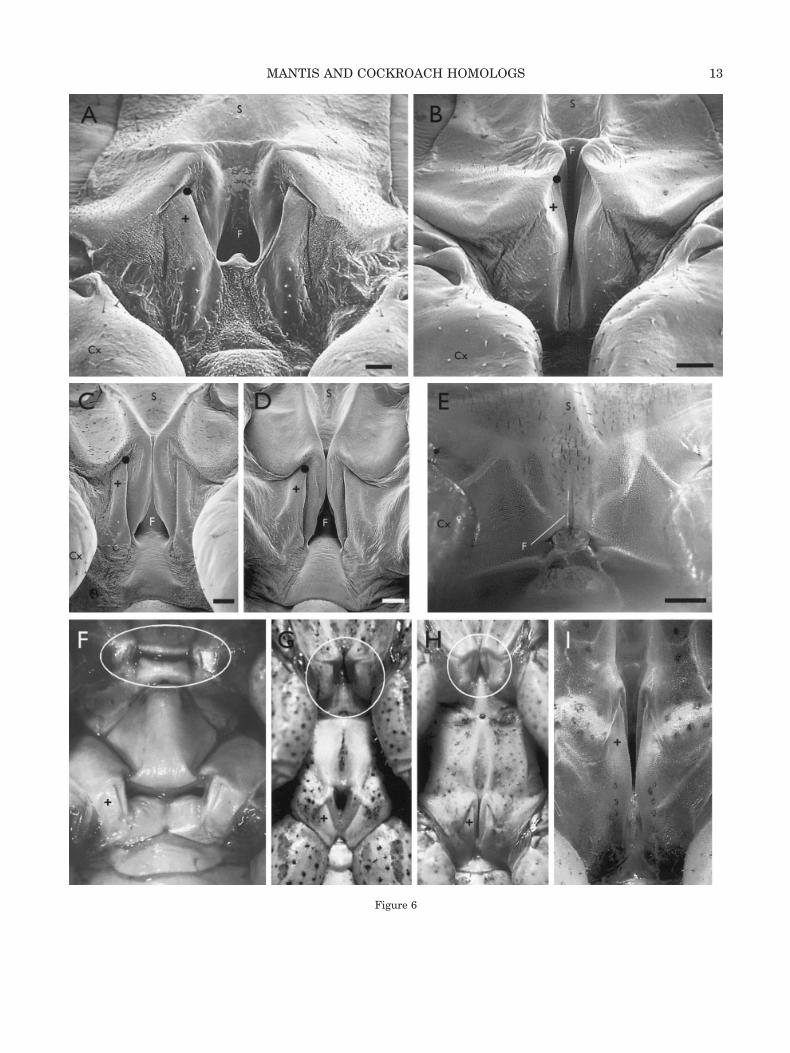
Figure 6
13MANTIS AND COCKROACH HOMOLOGS
though difficult to assess, it appears that the apex ofthe LVp has moved inward, pivoting on its longdorsal end. The central depression of the LVP bendslittle, if at all, but the LVp as a whole becomesrod-like in profile (Fig. 5B). The crease between theLVp and MVp deepens with very thin, flexible mem-branous cuticle at its bottom, and LVp rolls over theMVp socketed hairs.
The angle and size of the various thin cuticularfolds change as the adduction deviates from the hor-
izontal plane, and other folds may appear (graydashed line in Fig. 4A). The consequence is differentdegrees and pattern of bending in the sensillagroups. Elevation of the coxa, especially when it isaway from the midline, causes little bending ofhairs, but the apex of the LVp appears to moveinward like the tip of a triangular plate hinged atthe broad base. It moves outward to its originalposition as the leg is lowered.
Mantids. The consequences of coxal movementare less complex in mantids, at least as concernsexternal sensilla of the furcal-coxal area. Only acaudal hair field is present. Passive and active coxalmovement simply causes varying degrees of com-pression and “accordioning” of the cuticle (Fig. 4B).With strong adduction plus some elevation, all of thefurcal-coxal cuticle is strongly compressed exceptthat immediately adjacent to the ear. The rostralend of the ear is rigidly motionless during coxalmovements of all directions and amplitudes.
DISCUSSIONSummary
This study has identified cuticular structures in acockroach that are homologous to the major compo-nents of the praying mantis’ ventral metathorax—aneasy task given the considerable similarities be-tween the two insects. A major difference, however,is the presence of the midline ear in mantids com-pared to the earless condition in cockroaches. Thisstudy establishes that the area homologous to thewall of the mantis auditory chamber is the moredorsal sections of cuticle between the furcasternumand the coxa (Regions 4, 5, and 6). The exact rela-
Fig. 6. Mantis nymph and adult mesothoracic and metatho-racic segments without auditory specializations or with partialspecializations. The ventral caudal area is shown viewed ob-liquely from behind the animal. The same region of a termite isincluded for comparison. The mantis nymphs are Hierodula mem-branacea. Rostral is toward the top of each photograph. In allphotographs except E the “�” is on the rod-like structure justlateral to the dorsoventral slit. The black dot marks the attach-ment of the N.7 CO. A field of socketed hairs is present below the“�” in A–D (it is also present but not visible in F–I). The ovals/circles in F–H indicate the mesothoracic homologs of the metatho-racic structures near the bottom of the photos. A: Metathorax ofa second stadium nymph. Scale bar � 50 �m. B: Metathorax of asixth stadium nymph. Scale bar � 200 �m. C: Mesothorax ofsecond stadium nymph. Scale bar � 50 �m. D: Mesothorax ofseventh stadium nymph. Scale bar � 200 �m. E: Metathorax ofan alate male termite Zootermopsis nevadensis. Scale bar � 200�m. F: Eremiaphila brunneri (Eremiaphilidae), a broad-bodied,cursorial desert species. Males and females have the same met-athoracic anatomy and are both deaf. G: Oligonicella sp. (Thes-pidae; Oligonicinae), a slender grassland species with no met-athoracic dimorphism and no hearing. H: Female Amelesheldreichi (Mantidae; Amelinae). Males have good ultrasonichearing, but females have a “reduced” ear and thresholds �90 dBSPL. I: Female Miomantis abyssinica (Mantidae; Miomantinae).This is a dimorphic species in which males have sensitive hear-ing, but the metathoracic changes in the female are milder thanfor Ameles, and thresholds are 80–90 dB SPL.
Fig. 7. A: SEM of a Mercox cast ofthe rostral end of the Sphodromantisaurea auditory chamber wall. Raisedareas in the live animal are depres-sions in the cast and vice versa. Thebifid sensillum (BS; here a doublehole) is at the center of a roughlycircular patch of extremely thinmembrane devoid of microtricheawithin the semi-circle formed by thedense knob (*). It appears to belinked to the very narrow rostral endof the tympanum. The oval is the at-tachment site of the tympanal CO.Scale bar � 50 �m. B: Close-up of theof the LVp of Periplaneta americana.Refer to Figure 3 for orientation. Theoval is the attachment site of N.7’sCO. In both transformation hypothe-ses (Fig. 4) the precursor of the bifidsensillum should be just medial andventral to the CO insertion. As shownhere and by SEM there are no sen-silla visible there. The most constantsensory elements of this area are twoto three large socketed hairs at theapex of the LVp and the row of largehairs on the MVp.
14 D.D. YAGER

tionships of the MVp and LVp to components of thewall are less clear, although ontogenetic and phylo-genetic comparisons provide useful clues. The com-plex proprioceptive structures and functions of thecockroach furcal-coxal region have largely been lostin mantids. The one remaining component, the N.7CO, is the auditory transducer of the mantis ear.
Do Cockroaches Hear?
The answer is “Yes and No.” Some cockroaches inthe blaberoid subfamily Oxyhaloinae use acousticcommunication systems during intraspecific inter-actions (Fraser and Nelson, 1984; Clark and Moore,1995). Hissing produces temporally patterned sig-nals with broad frequency ranges (0.5–5 kHz). Thesesounds are effective over only short distances. Re-ception has been localized to the subgenual organs(Nelson, 1980) that, because of their extreme sensi-tivity, respond to near-field particle displacementstransferred to the legs either directly or indirectlythrough the tracheal system (Shaw, 1994).
The data here provide physiological confirmationthat cockroaches do not hear above a few kHz andthat they do not have any nonsubgenual sound pres-sure detection. Behavioral experiments (Ballard etal., 1984; Gold et al., 1984) and limited physiologicalstudies (Decker et al., 1989) in the past contradictoccasional claims to the contrary made by scientists(Mistal et al., 2000) and by commercial firms (elec-tronic roach repellents such as Pest A Cator Plus).The absence of higher frequency hearing across abroad taxonomic range demonstrated in this studylargely eliminates the possibility that some specieshear and others do not. There are two interpretiveconsiderations. First, it is possible that suction elec-trode recording from the whole connectives did notdetect audition either because the relevant axonsare very small or because they do not travel rostralto the prothoracic ganglion. Extracellular connectiverecordings in this lab and others (Casaday and Hoy,1977, for instance) from a range of insects (crickets,tiger beetles, mantids, moths) that show stimulus-locked responses, even from axons known to besmall, make this very unlikely. Second, this andearlier studies would not have detected hearing orig-inating in the head or cervical region. The findingmost relevant to the origins of the praying mantisear is that cockroaches have no auditory sensitivityoriginating from the metathorax.
Homologies
The homologies between the larger metathoracicsternal and pleural structures in cockroaches andmantids are obvious based on structure and relativeposition, and the results here parallel previous stud-ies (Crampton, 1909; Snodgrass, 1935; Levereault,1936; LaGreca, 1949; LaGreca and Rauchi, 1949).Matsuda (1960, 1970) reviewed the earlier literature
and unified much of the terminology. The organiza-tion of the ventral metathorax is fundamentally thesame in these two insects except for the caudal areabetween the furcasternum and the coxae.
The key features of the furcal-coxal area for un-derstanding the origins of the mantis ear are theMVp and LVp. The former was recognized in severalstudies (Crampton, 1914; Weber, 1928; Snodgrass,1935) that noted its highly variable occurrenceacross insect orders and, in some cases, across fam-ilies within orders. Matsuda (1960) notes that itappears to merge with other structures in the re-gion, e.g., the basisternum and furcasternum, or beabsent altogether. The LVp has not previously beendescribed as a separate structure. It undoubtedlyescaped notice because it is transparent and canlook like a simple fold of cuticle as it rotates duringcoxal motion.
The MVp and LVp are two components of a classictype of insect external proprioceptive unit (Chap-man, 1982) that consists of a hair plate (MVp) heldin position by a thin muscle (ACR) inserting alongits edge plus a fold of cuticle (LVp) that rolls over theadjacent hair plate as a limb moves. The hair plateand the cuticle fold must be separated by thin, flex-ible membrane to allow relative motion of the twoparts. The sensitivity of this sensory unit can beadjusted by contraction or relaxation of the muscle.Thus, the cockroach furcal-coxal region actually con-tains three types of sensory organs: fields of socketedhairs, the MVp-LVp complex, and the N.7 CO.
The N.7 CO that attaches to the apex of the LVp isnot part of the usual external proprioceptive unit,and turns the LVp into a dual-function structure.The high-speed videos show that the relatively stiffcentral depression allows the LVp to move like a stiffplate or rod, hinged at one end, with a sensor forin/out movement at the other.
The early literature disagrees about derivation ofthe MVp (and by extension, the LVp). Crampton(1914) identified its origin as the sternum, butothers felt it had a pleural origin even though itwas termed the “laterosternite” (Weber, 1928;Snodgrass, 1935; Matsuda, 1960). Assuming thatthe MVp and LVp are homologs of the auditorychamber wall (see below), the disagreement could betranslated as the difference between an ear derivedfrom a pleural versus a sternal chordotonal organgroup. The issue can be resolved in favor of thesternal origin by examining the innervation (Yagerand Scaffidi, 1993). The hairs and CO of the MVpand LVp are innervated by the most ventral nerveroot, N.7, of the metathoracic ganglion. N.7 hasbranches to most of the sternum, but none to thepleural region. In terms of “auditory clusters” (Yagerand Spangler, 1995; Yager, 1999b), this confirms theplace of mantids in the sternal cluster along withtachinid and sarcophagid flies rather than with in-sects with pleural ears like locusts, tiger beetles, andmany moths.
15MANTIS AND COCKROACH HOMOLOGS

Two Hypotheses
The homology of the dorsal furcal-coxal cuticle(Regions 4, 5, and 6) in cockroaches to the wall of themantis auditory chamber is supported by severalobservations: 1) it is in the right place relative to thefurcasternum; 2) it has the attachment of the N.7CO; 3) the anterior coxal rotator of mantids and the“sensory muscle” of cockroaches have the same coxalattachment, the same shape, the same innervation,and are the only muscles in that immediate area;and 4) the pericoxal cuticle (Region 3) and caudalhair field have the same spatial relationships. Theprimary aspects to the contrary are the absence ofsocketed hairs on the chamber wall itself and therelated absence of any obvious mantis counterpartsto Regions 1 and 2. Given the extreme variability innumbers of sternal socketed hairs across species ofboth insects (Yager, unpubl. obs.), the counterargu-ments are not compelling, especially in comparisonwith the supporting observations. It is thus reason-able to suggest that Regions 4, 5, and 6 togethercomprise the area homologous to the mantis audi-tory chamber wall.
Drawing more exact parallels is trickier. Thereare at least two possible hypotheses (Fig. 4D). Bothwould involve a medial shift and rotation, and bothwould require an overall enlargement (the auditorychamber walls are 1.5–2.0 mm long for larger man-tis species versus 0.7 mm for the LVp). The twohypotheses differ most importantly in the role of theLVp: 1) The LVp forms almost the entire auditorychamber wall (Fig. 4C). This would arise from asimple medial shift so that the edge of the LVp isclose to the furcasternum, a move strongly sup-ported by the medial shift of the ACR muscle attach-ment to the furcasternum. The muscle position im-plies that MVp would be lost. With thinner (12.3 �mvs. 4.9 �m for mantids; Yager, 1996b) but still stiffcuticle, the central depression of the LVp wouldbecome the tympanum. The ventral and dorsal rodswould arise from the lateral and medial borders ofthe LVp, respectively. The distinct hair field of Re-gion 4 (Figs. 4B, 6A) is retained in the correct spatialrelationship in both hypotheses. The very similarconfigurations of the LVp and the auditory chamberwall make this an appealing hypothesis. In disagree-ment is the location of the N.7 CO attachment siteon the medial edge of the LVp apex, which wouldbecome the dorsal rod rather than the ventral rodwhere it attaches in the mantis.
2) The LVp forms the ventral rod and the ventral-most cuticle of the chamber wall and the MVp be-comes the dorsal rod and cuticle. The very thin cu-ticle between the LVp and MVp would broaden andstiffen to form the tympanum. In contrast to therequirement posited for the evolution of tympana inmost other insects (Michelsen and Larsen, 1985;Yager, 1999b; Yack, 2004), no thinning would berequired because this cuticle in thinner than the
mantis tympanum (1–2 �m vs. 5 �m). The perfectcorrespondence of the N.7 CO attachment sites,along with the exact attachment configuration (com-pare Fig. 5B,C) argues for this scheme. The MVpwould not need to disappear. However, the ACRmuscle attachment on the furcasternum would be inthe wrong position relative to the MVp.
Comparisons with earless mantids lend support tothe second hypothesis. Early stadium nymphs, bothprimarily and secondarily earless species, and themesothoracic serial homologs all have the same ba-sic anatomical configuration (Fig. 6); it is only in theeared adults that the metathoracic segment differs.The distinctive hair field like Region 4 is uniformlypresent. The dorsoventral rod with the N.7 CO atthe apex overlies a groove that is separated from thefurcasternum by thicker cuticle. Further supportcomes from a developmental study (Yager, 1996b)that showed that the dorsoventral rod of nymphsbecomes the ventral rod of adults and the groovebroadens to form the tympanum. The thicker cuticleadjacent to the furcasternum becomes less promi-nent with age.
The bifid sensilla (BS) remain a puzzle. Judgingby their vibration (Yager and Michelsen, 2001), theyare highly responsive to ultrasound. Structurally,they look like mini-tympana with very thin mem-branes and COs attaching at their centers, but with-out impedance-matching air spaces. They have noobvious homolog in cockroaches, but there are atleast two possible origins. The BS structure suggeststhe fusion of two socketed hairs, possibly the veryprominent hairs at the apex of the LVp. The receptorelements are different for hair sensilla and chor-dotonal sensilla, but several lines of evidence showthe close interrelationship of the two types (Slifer1985; Field and Matheson, 1998). For instance, asingle gene mutation at the cut locus in Drosophilacan transform hair sensilla into chordotonal sensilla(Bodmer et al., 1987). Either sensilla type can arisefrom the same precursor cell, depending of the stateof the cut gene. Alternatively, the two sensilla of theBS could be a subgroup that has split off from theprimary chordotonal organ. A clear example of sucha division can be found in pneumorid grasshoppers(van Staaden et al., 2003). Another, more tantalizingexample is the tympanal organ of locusts that hasfour subgroups specialized for detection of differentsound frequencies by their separate locations on thetympanum (Michelsen and Larsen, 1985). Thismechanism, if true, combined with the observationof somewhat different tuning between the BS andthe tympanum (Yager and Michelsen, 2001) mightbe the first small hint of a basis for frequency dis-crimination in mantis hearing.
Transitions
The cuticular revisions necessary to build an in-sect ear vary considerably in their complexity. At its
16 D.D. YAGER

simplest, a thinned region of cuticle with a largetrachea or tracheal sac nearby is sufficient (crickets:Ball et al., 1989; butterflies and some moths: Minetand Surlykke, 2003; beetles: Yager and Spangler,1995; Forrest et al., 1997; locusts and grasshoppers:Michelsen and Larsen, 1985). This is the more com-mon pattern. In one unusual case (van Staaden andRomer, 1998; van Staaden et al., 2003), bladdergrasshoppers have sensitive hearing, but little ex-ternal modification. Some hawkmoths also lack truetympana (Gopfert et al., 2001), but there are othercuticular specializations that serve the same func-tion. At the opposite extreme that includes cicadas(Young and Hill, 1977; Doolan and Young, 1981),water boatmen (Prager, 1976), tettigoniids (Bailey,1990), and some moths (Fullard, 1984; Minet andSurlykke, 2003) are complex external and/or inter-nal cuticular changes including auditory chambers,hoods, tympanal protuberances, and cuticular inter-mediates between the tympanum and tympanal or-gan. Mantids fall somewhere toward the more com-plex end of this spectrum. Despite the complexalteration in the cockroach metathorax to accommo-date the ear, the major change, formation of theauditory chamber, appears quite simple becauseprecursor elements are present and in place. TheLVp normally rotates medially during coxal motion,so it need only be enlarged and fixed in its medialposition. Both hypotheses include a medial shift to-ward the furcasternum, although the extent differs.
The neural transition required to form the mantisperipheral auditory system differs from most otherinsects where it has been studied in that there isessentially none. The number of N.7 CO sensilla andtheir arrangement is the same in cockroaches andadult mantids (Yager and Scaffidi, 1993; Yager, inprep.) as well as in newly hatched mantis nymphs(Yager, 1996b). In moths (Yack et al., 1999) there isa trend toward reduction of the number of chor-dotonal sensilla in the ear. In cicadas (Doolan andYoung, 1981), tachinid flies (Robert and Willi, 2000),and pneumorid grasshoppers (van Staaden et al.,2003) there has been a massive elaboration yieldingup to 2,000 sensilla. Functional grouping of sensillaas in locusts (Breckow and Sippel, 1985), cicadas,and possibly tachinid flies may be a byproduct ofincreased CO size, but may also reflect the normaloccurrence of COs in groups (Slifer, 1936; Finlayson,1976).
In many respects the most significant transitioninherent in forming the mantis ear involves func-tion. Chordotonal organs in insects and other ar-thropods are internal proprioceptors, although theirroles have sometimes been extended or coopted toinclude gravity, wind, vibration, and surface wavedetection (Field and Matheson, 1998). Thus, theprimitive function of tympanal COs is, by definition,proprioception or a derivative. This has been con-firmed by comparisons of tympanal and atympanalCOs across life stages, across body segments, and
across species (Meier and Reichert, 1990; Yack andFullard, 1990; Lakes-Harlan et al., 1991, 1999;Rossler, 1992; Pfluger and Field, 1999; van Staadenet al., 2003). The furcal-coxal area of cockroaches isrichly appointed with socketed hairs that the high-speed videos show bending in various combinationsduring the range of coxal motions. The N.7 CO cancontribute proprioceptive information about motionin a different plane. Except for the Region 4 hairfield, all local proprioceptive function available tocockroaches is lost in the transition to the mantisear. The socketed hairs are simply gone, and the N.7CO monitors ultrasonic tympanal movement. Formantids and other insects without massive prolifer-ation of CO sensilla or extensive cuticular special-izations, it is likely that the primary—if not sole—cost of forming and maintaining an auditory systemis the loss of the proprioceptive information origi-nally supplied by the auditory precursors. Given theperfectly adequate capabilities of the metathoraciclegs in meeting the locomotor demands of the man-tis, the presence of the ear obviously does not com-promise motor effectiveness.
Why Is the Mantis Ear Where It Is?
The high-speed videos provide a compelling andsatisfying explanation for the central location of themantis ear. In that area, the only way to have astable structure that is not deformed during legmovement is to put it in the midline. Much smallermovements, including ventilation, are known to af-fect hearing in locusts (Hedwig et al., 1988). Theneed for stability can also explain the dense cuticu-lar knobs: they form an immovable rim that isolatesthe N.7 CO and the bifid sensillum from leg andbody movement. An added benefit comes from theformation of the auditory chamber that contributesto sensitivity and frequency response (Yager andHoy, 1989; Yager, unpubl. obs.).
A more fundamental question asks why the man-tis ear is in the caudal metathoracic sternum. Theappearance of the low-frequency ear of hymenopodidmantids in the serially homologous location (Yager,1996a) attests to the importance of this site for man-tids, but does not help with an explanation. Basedon suitable CO clusters (Slifer, 1936; Finlayson,1976; McIver, 1985) and precedents from othereared insects (Yack and Fullard, 1993; Yager,1999b), several candidate locations exist (only theprothoracic legs, mouthparts, and possibly wingswould be excluded in this case). It might be that theproprioceptors of the furcal-coxal region are themost expendable for mantids. The reasons might bestructural, such as a competition for space (Nijhoutand Emlen, 1998; Bailey and Kamien, 2001), ormore biomechanical, having to do with flight or lo-comotion. Alternatively, the location could be a mat-ter of chance (Gould, 1989). In the mantis case, thedual sensory functions of the LVp would seem to
17MANTIS AND COCKROACH HOMOLOGS

predispose or preadapt it for transformation into asound receiver. It would be a small step to go from atriangular hinge with an in/out sensor at the apex toa teardrop-shaped auditory chamber wall with asensor at the narrow end to monitor in/out vibra-tions caused by sound.
ACKNOWLEDGMENTS
The following provided valuable assistance duringthis project: Melinda Byrns, Kirsten Gaffke, Timo-thy Maugel, Sangeeta Pandya, Alex Pettyjohn, andDavid Scaffidi. Phillip Grandcolas and the Depart-ment of Entomology, Virginia Polytechnic Univer-sity, and other colleagues generously contributedcockroaches.
LITERATURE CITED
Adams JR, Wilcox TA. 1972. Determination of osmolalities ofinsect hemolymph from several species. Ann Entomol Soc Am66:575–577.
Aharinejad S, MacDonald IC, MacKay CE, Mason-Savas A. 1993.New aspects of microvascular corrosion casting: a scanning,transmission electron, and high-resolution intravital video mi-croscopic study. Microsc Res Tech 26:473–488.
Bailey WJ. 1990. The ear of the bushcricket. In: Bailey WJ, RentzDCF, editors. The Tettigoniidae: biology, systematics and evo-lution. Bathurst, NSW Australia: Crawford House Press. p217–247.
Bailey WJ, Kamien D. 2001. Hearing dimorphism, trait variation,and conflicts over space in the thorax of the bushcricket Re-quena verticalis (Listroscelisinae: Tettigoniidae: Orthoptera).J Comp Physiol A 187:647–652.
Ball EE, Oldfield BP, Michel Rudolph K. 1989. Auditory organstructure, development, and function. In: Huber F, Moore TE,Loher W, editors. Cricket behavior and neurobiology. Ithaca,NY: Cornell University Press. p 391–422.
Ballard JB, Gold RE, Decker TN. 1984. Response of Germancockroach (Orthoptera: Blattellidae) populations to a frequencysweeping ultrasound-emitting device [Blattella germanica]. JEcon Entomol 77:976–979.
Barros-Pita JC. 1971. Protective paraffin infiltration of soft tis-sues in insects to facilitate softening of hard exoskeletons. StainTech 46:171–175.
Bodmer R, Barbel S, Shepard S, Jack JW, Jan LY, Jan YN. 1987.Transformations of sensory organs by mutation of the cut locusof D. melanogaster. Cell 51:293–307.
Botes L, Price B, Waldron M, Pitcher GC. 2002. A simple andrapid scanning electron microscope preparative technique fordelicate “gymnodinioid” dinoflagellates. Microsc Res Tech 59:128–130.
Breckow J, Sippel M. 1985. Mechanics of the transduction ofsound in the tympanal organ of adults and larvae of locusts.J Comp Physiol A 157:619–629.
Casaday GB, Hoy RR. 1977. Auditory interneurons in the cricketTeleogryllus oceanicus: physiological and anatomical proper-ties. J Comp Physiol 121:1–13.
Chapman RF. 1982. The insects. Structure and function. Cam-bridge, MA: Harvard University Press.
Clark DC, Moore AJ. 1995. Social communication in the Mada-gascar hissing cockroach: features of male courtship hisses anda comparison of courtship and agonistic hisses. Behaviour 132:401–417.
Crampton GC. 1909. A contribution to the comparative morphol-ogy of the thoracic sclerites of insects. Proc Acad Nat Sci Phila1909:3–54 (and plates).
Crampton GC. 1914. Notes on the thoracic sclerites of wingedinsects. Entomol News 25:15–25 (and 11 plates).
Decker TN, Jones TA, Gold RE. 1989. Auditory thresholds in theAmerican cockroach (Orthoptera: Blattidae): estimates usingsingle-unit and compound-action potential recordings. J EconEntomol 82:687–691.
Doolan JM, Young D. 1981. The organization of the auditoryorgan of the bladder cicada, Cystosoma saudersii. Philos TransR Soc Lond 291:525–540.
Eberl DF, Hardy RW, Kernan MJ. 2000. Genetically similartransduction mechanisms for touch and hearing in Drosophila.J Neurosci 20:5981–5988.
Edgecomb RS, Robert D, Read MP, Hoy RR. 1995. The tympanalhearing organ of a fly: phylogenetic analysis of its morphologi-cal origin. Cell Tissue Res 282:251–268.
Eggleton P. 2001. Termite and trees: a review of recent advancesin termite phylogenetics. Insectes Sociaux 48:187–193.
Ehrmann R, Roy R. 2002. Systematische Aufstellung der Gattun-gen. In: Ehrmann R, editor. Mantodea. Gottesanbeterinnen derWelt. Munich: Natur Teil. p 374–378.
Field LH, Matheson T. 1998. Chordotonal organs of insects. AdvInsect Physiol 27:1–228.
Finlayson LH. 1976. Abdominal and thoracic receptors in insects,centipedes and scorpions. In: Mill PJ, editor. Structure andfunction of proprioceptors in the invertebrates. London: Chap-man and Hall. p 153–211.
Forrest TG, Read MP, Farris HE, Hoy RR. 1997. A tympanalhearing organ in scarab beetles. J Exp Biol 200:601–606.
Fraser JM, Nelson MC. 1984. Communication in the courtship ofa Madagascan hissing cockroach. I. Normal courtship. AnimBehav 32:194–203.
Fullard JH. 1984. External auditory structures in two species ofneotropical notodontid moths. J Comp Physiol A 155:625–632.
Gold RE, Decker TN, Vance AD. 1984. Acoustical characteriza-tion and efficacy evaluation of ultrasonic pest control devicesmarketed for control of German cockroaches (Orthoptera: Blat-tellidae). J Econ Entomol 77:1507–1512.
Gopfert M, Surlykke A, Wasserthal L. 2001. Tympanal and atym-panal ’mouth-ears’ in hawkmoths (Sphingidae). Proc R SocLond B 269:89–95.
Gould SJ. 1989. Wonderful life: the Burgess Shale and the natureof history. New York: WW Norton.
Grandcolas P. 1996. The phylogeny of cockroach families: a cla-distic appraisal of morpho-anatomical data. Can J Zool 74:508–527.
Grandcolas P. 1999. Reconstructing the past of Cryptocercus(Blattaria: Polyphagidae): phylogenetic histories and stories.Ann Entomol Soc Am 92:303–307.
Gratshev VG, Zherikhin VV. 1993. New fossil mantids (Insecta,Mantida [sic]). Paleontol J 27.
Grimaldi D. 1997. A fossil mantis (Insecta: Montodea) in Creta-ceous amber of New Jersey, with comments on the early historyof the Dictyoptera. Am Mus Novit 3204:1–11.
Gwynne DT. 1995. Phylogeny of the Ensifera (Orthoptera): ahypothesis supporting multiple origins of acoustical signalling,complex spermatophores and maternal care in crickets, katy-dids, and weta. J Orthop Res 4:203–218.
Harvey PH, Pagel MD. 1991. The comparative method in evolu-tionary biology. Oxford: Oxford University Press.
Hedwig B, Lang F, Elsner N. 1988. The interference of sound andmovement stimuli in tympanal receptors of Locusta migratoria.J Comp Physiol A 163:243–252.
Klass K-D. 2001. Morphological evidence on blatarian phylogeny:“phylogenetic histories and stories” (Insecta, Dictyoptera). MittMus Nat kd Berl, Dtsch Entomol Z 48:223–265.
Kristensen NP. 1995. Forty years’ insect phylogenetic systemat-ics. Zool Beitr 36:83–124.
LaGreca M. 1949. L’evoluzione della pleure pterotoraciche degliInsetti Blattotteroidi. Boll Zool, Torino 16:119–129.
LaGreca M, Rauchi A. 1949. Il dermascheletro e la muscola-tura del torace di Mantis religiosa. Ann Ist Mus Zool Napoli1:1– 41.
Lakes-Harlan R, Bailey WJ, Schikorski T. 1991. The auditorysystem of an atympanate bushcricket Phasmodes ranatriformes
18 D.D. YAGER

(Westwood) (Tettigoniidae: Orthoptera). J Exp Biol 158:307–324.
Lakes-Harlan R, Stolting H, Stumpner A. 1999. Convergent evo-lution of insect hearing organs from a preadaptive structure.Proc R Soc Lond B 266:1161–1167.
Lee B. 1950. The microtomists’s vade-mecum. Philadelphia:Blakiston.
Levereault P. 1936. The morphology of the Carolina mantis. UnivKansas Sci Bull 24:205–259.
Lo N, Tokuda G, Watanabe H, Rose H, Slaytor M, Maekawa K,Bandi C, Noda H. 2000. Evidence from multiple gene sequencesindicates that termites evolved from wood-feeding cockroaches.Curr Biol 10:801–804.
Mason JB. 1968. The tympanal organ of Acridomorpha. Eos 44:267–355.
Matsuda R. 1956. The comparative morphology of the thorax oftwo species of insects. Microentomology 21:1–65.
Matsuda R. 1960. Morphology of the pleurosternal region of thepterothorax in insects. Ann Entomol Soc Am 53:712–731.
Matsuda R. 1970. Morphology and evolution of the insect thorax.Mem Entomol Soc Can 76:1–431.
McIver SB. 1985. Mechanoreception. In: Kerkut GA, Gilbert LI,editors. Comprehensive insect physiology biochemistry andpharmacology. Oxford: Pergamon Press. p 71–132.
McKittrick FA. 1964. Evolutionary study of cockroaches. MemCornell Agric Exp Sta 389:1–123.
Meier T, Reichert H. 1990. Embryonic development and evolu-tionary origin of the orthopteran auditory organs. J Neurobiol21:592–610.
Merritt DJ. 1997. Transformation of external sensilla to chor-dotonal sensilla in the cut mutant of Drosophila assessed bysingle-cell marking in the embryo and larva. Microsc Res Tech39:492–505.
Michelsen A, Larsen ON. 1985. Hearing and sound. In: KerkutGA, Gilbert LI, editors. Comprehensive insect physiology, bio-chemistry, and pharmacology. Oxford: Pergamon Press. p 495–556.
Minet J, Surlykke A. 2003. Auditory and sound producing organs.In: Kristensen NP, editor. Lepidoptera, moths and butterflies.New York: Walter de Gruyter. p 290–323.
Mistal C, Takacs S, Gries G. 2000. Evidence for sonic communi-cation in the German cockroach (Dictyoptera: Blatellidae). CanEntomol 132:867–876.
Nalepa CA, Bandi C. 1999. Phylogenetic status, distribution, andbiogeography of Cryptocercus (Dictyoptera:Cryptocercidae).Ann Entomol Soc Am 92:292–302.
Nation JE. 1983. A new method using hexamethyldisilazane forpreparation of soft insect tissues for scanning electron micros-copy. Stain Tech 58:347–351.
Nelson MC. 1980. Are subgenual organs “ears” for hissing cock-roaches? Soc Neurosci Abstr 6:198.195.
Nijhout HF, Emlen DJ. 1998. Competition among body parts inthe development and evolution of insect morphology. Proc NatlAcad Sci U S A 95:3685–3689.
Okabe M, Okano H. 1997. Two-step induction of chordotonalorgan precursors in Drosophila embryogenesis. Development124:1045–1053.
Otte D. 1990. The relation between hearing and flying in crickets.Entomol News 101:29034.
Pantin CFA. 1946. Notes on microscopical techniques for zoolo-gists. Cambridge, UK: Cambridge University Press.
Pfluger H-J, Field LH. 1999. A locust chordotonal organ codingfor proprioceptive and acoustic stimuli. J Comp Physiol A 184:169–183.
Pichon Y. 1970. Ionic content of haemolymph in the cockroach,Periplaneta americana. A critical analysis. J Exp Biol 53:195–209.
Pollack AJ, Ritzmann RE, Watson JT. 1994. Dual pathways fortactile sensory information to thoracic interneurons in the cock-roach. J Neurobiol 26:33–46.
Prager J. 1976. Das mesothorakale Tympanalorgan von Corixapunctata Ill. (Heteroptera, Corixidae). J Comp Physiol 110:33–50.
Riede K, Kamper G, Hofler I. 1990. Tympana, auditory thresholdsand projection areas of tympanal nerves in singing and silentgrasshoppers (Insecta, Acridoidea). Zoomorphology 109:223–230.
Robert D, Willi U. 2000. The histological architecture of theauditory organs in the parasitoid fly Ormia ochracae. Cell Tis-sue Res 301:447–457.
Rossler W. 1992. Functional morphology and development oftibial organs in the legs I, II and III of the bushcricket Ephip-piger ephippiger (Insecta, Ensifera). Zoomorphology 112:181–188.
Rust J, Stumpner A, Gottwald J. 1999. Singing and hearing in aTertiary bushcricket. Nature 399:650.
Shaw SR. 1994. Re-evaluation of the absolute threshold andresponse mode of the most sensitive known “vibration” detector,the cockroach’s subgenual organ: a cochlea-like displacementthreshold and a direct response to sound. J Neurobiol 25:1167–1185.
Slifer EH. 1936. The scoloparia of Melanoplus differtialis (Or-thoptera Acrididae). Entomol News 47:174–180.
Snodgrass RE. 1935. Principles of insect morphology. Ithaca, NY:Cornell University Press.
Strausfeld NJ, Seyan HS, Wohlers D, Bacon JP. 1983. Luciferyellow histology. In: Strausfeld NJ, editor. Functional neuro-anatomy. Berlin: Springer. p 132–155.
Thorne BL, Carpenter JM. 1992. Phylogeny of the Dictyoptera.Syst Entomol 17:253–268.
Triblehorn JD. 2003. Multisensory integration in the ultrasound-triggered escape response of the praying mantis, Parasphen-dale agrionina. Doctoral dissertation. University of Maryland,College Park.
Triblehorn JD, Yager DD. 2002. Implanted electrode recordingsfrom a praying mantis auditory interneuron during flying batattacks. J Exp Biol 205:307–320.
van Staaden MJ, Romer H. 1998. Evolutionary transition fromstretch to hearing organs in ancient grasshoppers. Nature 394:773–776.
van Staaden MJ, Rieser M, Ott SR, Pabst MA, Romer H. 2003.Serial hearing organs in the atympanate grasshopper Bullacrismembracioides (Orthoptera, Pneumoridae). J Comp Neurol465:579–592.
Vrsansky P. 2002. Origin and early evolution of mantises. AMBAProjekty 6:1–16.
Weber H. 1928. Die Gleiderung der Sternopleuralregion desLepidopteran-Thorax. Eine vergleichende morphologische Stu-dien zur Subcoxaltheorie. Zeitschr Wiss Zool 131:181–254.
Wheeler WC, Whiting MF, Wheeler QD, Carpenter JM. 2001.The phylogeny of the extant hexapod orders. Cladistics 17:113–169.
Wiley EO. 1981. Phylogenetics. The theory and practice of phy-logenetic systematics. New York: John Wiley & Sons.
Yack JE. 1993. Janus Green B as a rapid, vital stain for periph-eral nerves and chordotonal organs in insects. J Neurosci Meth-ods 49:17–22.
Yack JE. 2004. The structure and function of auditory chor-dotonal organs in insects. Microsc Res Tech 63:315–337.
Yack JE, Fullard JH. 1990. The mechanoreceptive origin of insecttympanal organs: a comparative study of similar nerves intympanate and atympanate moths. J Comp Neurol 300:523–534.
Yack JE, Fullard JH. 1993. What is an insect ear? Ann EntomolSoc Am 86:677–682.
Yack JE, Scudder GGE, Fullard JH. 1999. Evolution of the met-athoracic tympanal ear and its mesothoracic homologue in theMacrolepidoptera (Insecta). Zoomorphology 119:93–103.
Yager DD. 1990. Sexual dimorphism of auditory function andstructure in praying mantises (Mantodea; Dictyoptera). J ZoolLond 221:517–537.
Yager DD. 1992. Ontogeny and phylogeny of the cyclopean man-tis ear. J Acoust Soc Am 92:2421.
Yager DD. 1996a. Serially homologous ears perform frequencyrange fractionation in the praying mantis, Creobroter (Man-todea, Hymenopodidae). J Comp Physiol A 178:463–475.
19MANTIS AND COCKROACH HOMOLOGS

Yager DD. 1996b. Nymphal development of the auditory systemof the praying mantis Hierodula membranacea Burmeister(Dictyoptera, Mantidae). J Comp Neurol 364:199–210.
Yager DD. 1999a. Sensory processes. Hearing. In: Prete FR,Wells H, Wells PH, Hurd LE, editors. The praying mantids:research perspectives. Baltimore: Johns Hopkins UniversityPress. p 93–113.
Yager DD. 1999b. Structure, development, and evolution of insectauditory systems. Microsc Res Tech 47:380–400.
Yager DD, Hoy RR. 1987. The midline metathoracic ear of thepraying mantis, Mantis religiosa. Cell Tissue Res 250:531–541.
Yager DD, Hoy RR. 1989. Audition in the praying mantis, Mantisreligiosa L.: identification of an interneuron mediating ultra-sonic hearing. J Comp Physiol A 165:471–493.
Yager DD, Michelsen A. 2001. A traveling wave in the curious earof the praying mantis. In: Sixth International Congress of Neu-roethology, Bonn, Germany.
Yager DD, Scaffidi DJ. 1993. Cockroach homolog of the mantistympanal nerve. Soc Neurosci Abstr 19:340.
Yager DD, Spangler HG. 1995. Characterization of auditory af-ferents in the tiger beetle, Cicindela marutha Dow. J CompPhysiol A 176:587–600.
Yager DD, May ML, Fenton MB. 1990. Ultrasound-triggered,flight-gated evasive maneuvers in the praying mantis, Paras-phendale agrionina (Gerst.). I. Free-flight. J Exp Biol 152:17–39.
Young D, Hill KG. 1977. Structure and function of the auditorysystem of the cicada, Cystosoma saundersii. J Comp Physiolll7:23–45.
20 D.D. YAGER
Copyright © 2022 FDOKUMEN


















