
Patterns of praying mantis auditory system evolution based on ...
28
Patterns of praying mantis auditory system evolution based on morphological, molecular, neurophysiological, and behavioural data DAVID D. YAGER 1 * and GAVIN J. SVENSON 2 1 Department of Psychology and Neuroscience and Cognitive Science Program, University of Maryland, College Park, Maryland 20742, USA 2 Department of Biology, Brigham Young University, Provo, Utah 84602, USA Received 12 March 2007; accepted for publication 02 October 2007 Some praying mantids have sensitive ultrasonic hearing arising from a unique ‘cyclopean’ ear located in the ventral metathorax. The present study explores the evolutionary history of the mantis auditory system by integrating large anatomical, neurophysiological, behavioural, and molecular databases. Using an ‘auditory phylogeny’ based on 13 morphological characters, we identified a primitively earless form of metathoracic anatomy in several extant taxa. In addition, there are five distinct mantis auditory systems. Three of these can be identified anatomically, and the other two can only be detected neurophysiologically. Superimposing these results onto a phylogenetic tree derived from molecular data from seven genes shows that the cyclopean mantis ear evolved once approximately 120 Mya. All the other auditory system types are either varying degrees of secondary loss, or are recent innovations that each occurred independently multiple times. The neurophysiological response to ultrasound is remarkably consistent across all taxa tested, as is the multicomponent, in-flight behaviour triggered by ultrasound. Thus, mantids have an ancient, highly conserved auditory neural–behavioural system. Although ultrasonic hearing in several insect groups evolved in response to bat predation, mantis hearing predates the appearance of bats (approximately 63 Mya) and must originally have functioned in communication, prey detection, or avoidance of nonbat preda- tors. © 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568. ADDITIONAL KEYWORDS: acoustic startle – bat evasion – cyclopean ear – insect ear – insect hearing – molecular phylogeny – tympanum. INTRODUCTION In part because of their immense diversity, insect tympanate auditory systems offer an opportunity to understand the evolutionary patterns and processes that lead to ‘new’ sensory systems. A traditional strategy has been to identify and compare auditory systems, or the homologous anatomical components of the auditory system, in a broad range of species (acridids: Mason, 1968; ensifera: Gwynne, 1995; moths: Yack, Scudder & Fullard, 1999; Hasenfuss, 2000; Minet & Surlykke, 2003; flies: Edgecomb et al., 1995; Lakes-Harlan, Stölting & Stumpner, 1999; mantids: Yager, 1999a, 2005). An even more informa- tive approach is to integrate comparative anatomical data with phylogenetic data, and this has been con- ducted primarily for moths, ensifera, and flies. It is clear from these studies that tympanate hearing has evolved independently more than 20 times among eight orders (Yager, 1999b; Yack & Hoy, 2003). Praying mantids (Dictyoptera; Mantodea) offer an interesting and useful counterpoint to other tympan- ate insect groups for several reasons: (1) the auditory system is unique (Yager, 1999a) and consists of an anatomically and functionally single ear comprising two tympana in a deep, cuticular auditory chamber, located in the ventral midline of the caudal metatho- rax (the vast majority of mantids can hear only ultra- sound); (2) the suborder Mantodea is small by insect *Corresponding author. E-mail: [email protected] Biological Journal of the Linnean Society, 2008, 94, 541–568. With 6 figures © 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568 541 Downloaded from https://academic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Patterns of praying mantis auditory system evolution based on ...

Patterns of praying mantis auditory system evolutionbased on morphological, molecular, neurophysiological,and behavioural data
DAVID D. YAGER1* and GAVIN J. SVENSON2
1Department of Psychology and Neuroscience and Cognitive Science Program, University ofMaryland, College Park, Maryland 20742, USA2Department of Biology, Brigham Young University, Provo, Utah 84602, USA
Received 12 March 2007; accepted for publication 02 October 2007
Some praying mantids have sensitive ultrasonic hearing arising from a unique ‘cyclopean’ ear located in the ventralmetathorax. The present study explores the evolutionary history of the mantis auditory system by integrating largeanatomical, neurophysiological, behavioural, and molecular databases. Using an ‘auditory phylogeny’ based on 13morphological characters, we identified a primitively earless form of metathoracic anatomy in several extant taxa.In addition, there are five distinct mantis auditory systems. Three of these can be identified anatomically, and theother two can only be detected neurophysiologically. Superimposing these results onto a phylogenetic tree derivedfrom molecular data from seven genes shows that the cyclopean mantis ear evolved once approximately 120 Mya.All the other auditory system types are either varying degrees of secondary loss, or are recent innovations that eachoccurred independently multiple times. The neurophysiological response to ultrasound is remarkably consistentacross all taxa tested, as is the multicomponent, in-flight behaviour triggered by ultrasound. Thus, mantids havean ancient, highly conserved auditory neural–behavioural system. Although ultrasonic hearing in several insectgroups evolved in response to bat predation, mantis hearing predates the appearance of bats (approximately63 Mya) and must originally have functioned in communication, prey detection, or avoidance of nonbat preda-tors. © 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568.
ADDITIONAL KEYWORDS: acoustic startle – bat evasion – cyclopean ear – insect ear – insect hearing –molecular phylogeny – tympanum.
INTRODUCTION
In part because of their immense diversity, insecttympanate auditory systems offer an opportunity tounderstand the evolutionary patterns and processesthat lead to ‘new’ sensory systems. A traditionalstrategy has been to identify and compare auditorysystems, or the homologous anatomical componentsof the auditory system, in a broad range of species(acridids: Mason, 1968; ensifera: Gwynne, 1995;moths: Yack, Scudder & Fullard, 1999; Hasenfuss,2000; Minet & Surlykke, 2003; flies: Edgecomb et al.,1995; Lakes-Harlan, Stölting & Stumpner, 1999;
mantids: Yager, 1999a, 2005). An even more informa-tive approach is to integrate comparative anatomicaldata with phylogenetic data, and this has been con-ducted primarily for moths, ensifera, and flies. It isclear from these studies that tympanate hearing hasevolved independently more than 20 times amongeight orders (Yager, 1999b; Yack & Hoy, 2003).
Praying mantids (Dictyoptera; Mantodea) offer aninteresting and useful counterpoint to other tympan-ate insect groups for several reasons: (1) the auditorysystem is unique (Yager, 1999a) and consists of ananatomically and functionally single ear comprisingtwo tympana in a deep, cuticular auditory chamber,located in the ventral midline of the caudal metatho-rax (the vast majority of mantids can hear only ultra-sound); (2) the suborder Mantodea is small by insect*Corresponding author. E-mail: [email protected]
Biological Journal of the Linnean Society, 2008, 94, 541–568. With 6 figures
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568 541
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022

standards with only approximately 2400 species(Ehrmann & Roy, 2002; Otte & Spearman, 2005);(3) neither of the two other groups within the Dicty-optera, the cockroaches and the termites, has tympa-nate hearing (Yager, 2005); (4) despite its small size,the Mantodea is rich in diversity (Beier, 1968; Preteet al., 1999), including adult body size (6 mm–155 mm), habitat (from desert to cloud forest andmost habitats in between, with complementarybehavioural, ecological, and anatomical specializa-tions), chromosome number and sex chromosomesystem (Hughes-Schrader, 1950; White, 1965); (5) theontogeny of the mantis ear is known (Yager, 1996b);(6) the cockroach homologs of the mantis auditorysystem components have been identified (Yager,2005); and (7) although the fossil record of mantids isscant, it has grown recently to include several well-preserved specimens from the Cretaceous (Gratschev& Zherikhin, 1993; Grimaldi, 1997, 2003; Vrsansky,2002).
Ultimately, audition must subserve adaptive behav-iours, and the evolution of those behaviours is acrucial part of the overall story. For mantids, batdetection and evasion is a major function of hearing.Trains of ultrasonic pulses trigger a complex, multi-component behaviour that converts a straight flightpath into a power dive toward the ground (Yager &May, 1990; Yager, May & Fenton, 1990; Triblehorn,2003). Hearing mantids escape capture by flying batsin 76% of attacks, whereas deafened mantids escapeonly 34% of the time. The evolution of such a behav-iour requires alterations in the central nervoussystem to link the ‘new’ sensory information to aneffective behaviour. Some of these neural underpin-nings are known for mantids (Yager & Hoy, 1989;Triblehorn & Yager, 2002, 2005).
The major drawback to using mantids in compara-tive studies of ear evolution has been the lack of aphylogeny for the suborder. The standard taxonomicscheme for mantids (Beier, 1968) and its recent revi-sion (Roy, 1999; Ehrmann & Roy, 2002) are basedsolely on a traditional taxonomic interpretation ofphysical characters. Two recent studies have ad-dressed the question of mantis phylogeny. Grimaldi(2003) created a phylogentic hypothesis for the fossiltaxa and the three extant basal families of mantids.Svenson & Whiting (2004) generated a molecularphylogeny of the suborder based on analysis of mul-tiple genes, both nuclear and mitochondrial, from 63species. The molecular phylogeny found that only oneof five families examined and five of 14 subfamilies ofthe revision by Ehrmann & Roy (2002) were poten-tially monophyletic.
The present study reports the results of an exten-sive anatomical, physiological, and behaviouralsurvey of mantis auditory systems. It complements
and extends several previous studies (Yager & Hoy,1989; Yager & May, 1990; Yager, 1990, 1996a; Trible-horn & Yager, 2001) with new data and new analyses,and then integrates the overall results using phylo-genetic techniques. The results show six major audi-tory types among the praying mantids, although onlyfour of these can be recognized anatomically. Wesuperimposed the ear types onto a newly-createdmolecular phylogeny of the suborder. The anatomical/physiological/behavioural patterns are highly congru-ent with the phylogenetic tree derived from moleculardata. The metathoracic ear arose just once appro-ximately 120 Mya, and there are several extantlineages of primitively earless mantids. The neuro-physiological and behavioural characteristics arehighly conserved throughout the suborder, suggestinga defensive system as old as the ear itself.
MATERIAL AND METHODSANIMALS
Preserved specimensThe anatomical data came from examination ofpinned specimens in the collections at the BritishMuseum (Natural History), the PhiladelphiaAcademy of Sciences, the US National Museum ofNatural History, and the American Museum ofNatural History.
Live specimensA total of 76 species of live mantids were availableduring the project. Sources included commercial sup-pliers, hobbyists, other scientists, and field collection.In some cases, we tested wild-caught adults and, inthe remainder, the mantids were raised in our colonyfrom eggs and nymphs. Although identification at thegenus level was always clear, the species was some-times not. Voucher specimens are stored at theDepartment of Psychology, University of Maryland,College Park.
Specimens for molecular studiesAll genetic data were taken from the GenBank sub-missions of Svenson & Whiting (2004). The originaltissue samples for that data came almost exclusivelyfrom freshly preserved specimens, ensuring highestquality DNA sequencing. However, in a few cases, theanimals had been preserved in 70% ethyl alcohol andstored for more than 5 years at room temperature. Inone case (Metallyticus), muscle was removed from arecently pinned specimen and, in another (Chaeteesa),from a museum specimen approximately 10 years old.
Table 1 shows the breadth of the data sets in thisstudy relative to the taxonomic scheme of Ehrmann &Roy (2002) for the entire suborder.
542 D. D. YAGER and G. J. SVENSON
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022
METATHORACIC ANATOMY
Examination of each specimen focused on 13 anato-mical characters of the caudal midline of themetathoracic sternum (Fig. 1). Because of its strongcorrelation with ear anatomy, we also recorded winglength (full winged, mildly brachypterous, stronglybrachypterous, apterous). Classification above thespecies level was conducted sensu Ehrmann & Roy(2002).
Of the species included in the final data set, typespecimens accounted for 28% of the data. An addi-tional 57% was identified by established authorities(primarily M. Beier, E. Giglio-Tos, M. Hebard, A.Kaltenbach, J. Marshall, J. A. G. Rehn, R. Roy, B.Uvarov). The remaining 15% were determined withreference to identified specimens in the collections. Inmost cases, several individuals of each species andseveral species from each genus were examined.Examination of live mantids and animals recentlypreserved in alcohol helped overcome the issue ofdistortion due to drying.
PHYSIOLOGY
ProceduresPhysiological experiments were carried out using theprocedures described in Triblehorn & Yager (2001).Briefly, after surgical exposure of the ventral nervecord in the prothorax, the animal was positioned atthe centre of an anechoic box. The legs were removedto eliminate subgenual responses. Stimuli (50 or70-ms sound pulses with 3–5 ms rise/fall times overthe range of 1–150 kHz) were played from PanasonicEAS10TH400B (for > 10 kHz) and Radio Shack40–1289A speakers at the end of the box. Ascendingneural activity was recorded using a suction electrodeplaced over the caudal cut end of one of the connec-tives and standard recording electronics.
Calibration of the stimulus presentation systemused a Brüel & Kjaer 2231 sound level meter with a4135 6.25-mm microphone positioned above the wax/plastic block at the location occupied by the caudalmetathorax during experiments. The frequencyresponse of the calibration system in the range
1–150 kHz was derived from the frequency responsesof the sound level meter itself and the microphone.
Data collectionA physiological tuning curve was determined for eachanimal using stimuli at � 25 frequencies in the range1–150 kHz. Threshold was defined as the lowestsound pressure level that elicited a response in twoout of three stimuli. Stimulus levels are reported asdB re: 20 mPa (dB SPL). Sample sizes for individualspecies were in the range from 1 to > 50 depending onavailability, and in most cases included both sexes.
Animals that showed no auditory responses in aninitial scan were nevertheless tested at a minimum of15 frequencies in the range 1–100 kHz with threeintensities, including 95 dB SPL. We normally did notknow the age of wild-caught animals, but extensivestudies with several genera (Mantis, Sphodromantis,Taumantis, Hierodula, Miomantis, Creobroter, Paras-phendale) over several years has shown no decrementin auditory sensitivity with age.
BEHAVIOUR
Mantids suspended by a wire attached to their prono-tum will fly readily, especially when a gentle wind isdirected onto their head from the front. Using theset-up and stimulus producing system detailed above,we assessed the behavioural responses of flyingmantids. The stimuli were 300-ms trains of 10-mspulses with 50% duty cycle 20 dB over threshold atthe individual animal’s most sensitive frequency.
The evasive response comprises five distinct andseparate components. For each species, we recordedthe occurrence, strength, and variation of each com-ponent. In addition, we noted any qualitative varia-tions and overall variability of the responses. Formore details of the behavioural techniques refer toTriblehorn & Yager (2001).
PHYLOGENY OF METATHORACIC ANATOMY
To assess the possible evolutionary relationshipsamong the types of ventral metathorax, we created
Table 1. Taxonomic coverage of the various data sets used in this study compared to the scheme of Ehrmann & Roy(2002)
Ehrmann/roy Anantomy Physiology Phylogeny Behaviour
Families 15 15 10 15 5Subfamilies 48 48 21 43 15Tribes 46 46 23 45 15Genera 434 344 54 146 23Species ª 2300 > 500 76 158 30
MANTIS EAR PHYLOGENY 543
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022
a phylogenetic reconstruction using the charactersstudied in the anatomy data set (Fig. 1; see characterdescriptions below and Appendix 1). Male (197genera) and female (175 genera) data sets were analy-sed separately. The outgroups were cockroaches:Periplaneta americana, Gyna lurida, Blaptica dubia,
and Panchlora nivea. We also included second instarnymphs of the DK-eared mantis, Sphodromantisaurea. The data sets included both ordered and unor-dered characters with three weight categories (1, 3,and 6). To determine the influence of character order-ing and weighting, we analysed the two data sets
544 D. D. YAGER and G. J. SVENSON
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022

under three additional parsimony strategies: (1) noordering, no weighting; (2) ordering, no weighting;and (3) no ordering, characters weighted. Strict con-sensus trees were calculated to asses the recovery ofcertain nodes under each strategy.
Parsimony analysis was performed on each strategyfor each data set using TNT (Goloboff, Farris & Nixon,2003). We performed 5000 random additions using the‘New Technology Search’, which employed severaladditional search algorithms (Goloboff, 1999; Nixon,1999). Strict consensus trees were calculated for eachanalysis to assess the recovery of specific nodes.
MOLECULAR PHYLOGENY
A molecular phylogeny was reconstructed with DNAsequence data for seven nuclear and mitochondrialgenes downloaded from GenBank (for accessionnumbers, see Appendix 2). The specific genes haveserved to resolve effectively Mantodean and otherinsect relationships (Caterino, Cho & Sperling, 2000;Terry & Whiting, 2005). Outgroups came from theBlattodea (two taxa) and Isoptera (three taxa).
We assembled the seven genes using SEQUENCHER,version 4.2.2 (Genecodes, 1991–2003). We performedmanual alignment on protein coding genes to matchtheir conserved codon reading frame. Length variableribosomal genes were manually aligned according toconserved and nonconserved regions to create mul-tiple partitions for algorithmic alignment. The result-ing sequence partitions for 28s rDNA (13 partitions),18S rDNA (five partitions), 12S rDNA (three parti-tions), 16S rDNA (ten partitions) were aligned usingMAFFT, version 5.7 (Katoh et al., 2002; Katoh et al.,2005) under the L-INS-i parameter. All genes wereconcatenated using MacClade, version 4.06 (Maddi-son & Maddison, 2003) to include the gene loci:12S rDNA, 477 bp; 16S rDNA, 657 bp; 18S rDNA,1965 bp; 28S rDNA, 2609 bp; cytochrome oxidasesubunit I, 1414 bp; cytochrome oxidase subunit II,642 bp; and histone 3, 376 bp. We calculated the best-fit model for each of the loci using the Akaike Infor-
mation Criterion implemented in MODELTEST,version 3.06 (Posada & Crandall, 1998) using PAUP4.0b10 (Swofford, 2002).
Using TREEFINDER (Jobb, 2006), we performedpartitioned likelihood analysis on the entire data set.We specified separate models for each partition basedon results from MODELTEST, although model param-eters for each model were estimated by TREEFINDER.Four identical runs of 5000 replicates, ensured findingthe best tree across independent analyses. We calcu-lated nodal support using partitioned bootstrap analy-sis within TREEFINDER (500 replicates). PartitionedBayesian methods coupled with Markov chain MonteCarlo were implemented in the program MrBAYES,version 3.04b (Ronquist & Huelsenbeck, 2003). Weemployed six parameter models for each of the sevenpartitions. Four independent analyses using fourchains (three cold and one hot) for 10 million genera-tions ran in parallel. Generations prior to reachingstability were discarded as the burn-in. The finalposterior probability tree was a 50% majority ruleconsensus tree (Huelsenbeck & Imennov, 2002;Huelsenbeck et al., 2002).
We calculated divergence times for the most likelytree using the program R8S, version 1.70 (Sanderson,2004a). Age constraints placed on nodes within theingroup were based on fossil Mantodea presented byGratschev & Zherikhin (1993), Grimaldi (1997, 2002,2003), Nel & Roy (1996), and Sharov (1962). Fossilsreferable to extant Mantodea were Choeradodinae:Prochaeradodis enigmaticus (shale, 60 Mya) andMantoididae: Mantoida sp. (amber, 20 Mya). Addi-tionally, we placed minimum and maximum ageconstraints on Blattodea following the timeline ofVrsansky (2002) and Vrsansky, Vishniakova & Ras-nitsyn (2002), which postulates that the ancestor tomodern Blattaria emerged during the middle Triassicapproximately 235 Mya. Further, a minimum age con-straint for the outgroup was based on the oldestknown fossils representing the modern Blattodeafamilies of Polyphagidae and Blattellidae, which arefrom the Early Cretaceous approximately 130 Mya.
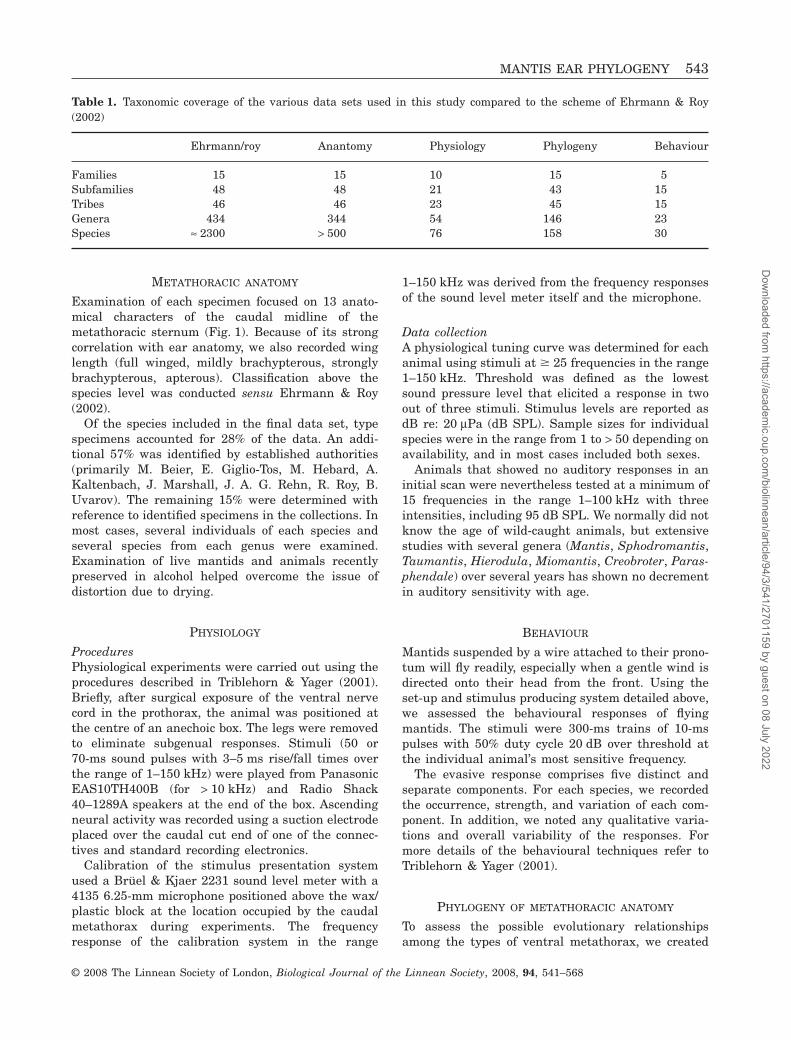
Figure 1. Anatomy of the ultrasound-sensitive DK (deep groove and prominent knobs) ear. A. ventral thorax of thehymenopodid mantis Pseudocreobotra ocellata showing the locations of the DK ear (lower grey oval) and the MESO ear(upper grey oval) between the bases of the metathoracic and mesothoracic coxae, respectively. B. The DK ear of P. ocellata.The asterisk is on one of the knobs that mark the rostral end of the ear. The knob angle is approximately 90°. The greenarrows indicate the openings to the auditory chamber. The blue arrow points to the very narrow wall separation typicalof this anatomical type. The tympanal tracheal sacs create the bulges just lateral to the groove (+). CX, coxa. C, D,midsagittal view of one wall of the auditory chamber in Parasphendale agrionina. The blue colour comes from a dye(Janus green) injected to enhance contrast. The thick dashed line outlines the ventral border of the wall and the thin lineindicates the inner extent of the heavy cuticle that makes up the knob (*). The ventral rod somewhat overhangs thetympanal depression (a true tympanum in this case). The bifid sensillum is at the centre of a circle of very thin cuticle(yellow). Although the structures of the chamber wall are difficult to see in live animals, the shrinkage typical of pinnedspecimens make them visible in museum specimens.�
MANTIS EAR PHYLOGENY 545
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022

Therefore, the age constraints are: Mantoididae -minimum age = 20 Mya; Choeradodinae - minimumage = 60 Mya; and outgroup - minimum age = 130Mya and maximum age = 235 Mya.
To obtain 95% confidence intervals (CI) for theestimated nodes we performed bootstrap divergencetime estimation using a package of Perl scriptswritten and distributed by Eriksson (2002). Followingthe procedure originally described by Sanderson(2004b), we created 100 bootstrap replicate data setsfrom our total combined data set using PHYLIP,version 3.6 (Felsenstein, 2005), and estimated branchlengths for the previously chosen topology using eachof the 100 replicate data sets. All branch lengthestimation was carried out in PAUP, simultaneouslyestimating parameters for the GTR + G + I model.Subsequently, all 100 optimized branch length treeswere analysed in R8S to profile the selected nodes forcalculation of the 95% CI.
RESULTSANATOMICAL PATTERNS
DescriptionsWe took as a starting reference the structure of theultrasound-sensitive ear described by Yager & Hoy(1987) and Yager (2005). The primary structures andnomenclature are shown in Figure 1. In the museumspecimens, ventral metathoracic anatomy revealedseveral clear categories, most with one or more vari-ants. The variants primarily differed in relative sizesof the components, rather than their organization.The most important differentiating characteristics aresummarized in Table 2.
The DK form: This is the ultrasound-sensitive ear.‘DK’ denotes a deep groove and prominent knobs that
are obvious to even casual inspection of the regionbetween the metathoracic legs (Figs 1 and 2A). Theear proper comprises an auditory chamber demar-cated at its anterior end by two, well-defined, thickcuticular knobs. The rostral border of the knobapproximately forms a right angle with the long axisof the thorax. Viewed from the outside, two lateralwalls of the chamber almost touch for the rostralthird of their length and then progressively diverge sothat at the posterior end of the auditory chamberthe walls are often widely separated. The floor of theauditory chamber is primarily the furcasternum. Thetwo furcal pits (origins of two internal muscle attach-ment structures) lie deep within the chamber nearits caudal end and are not visible from the outside.The DK ear length (mean ± SD) for 180 genera is1.47 ± 0.46 mm.
The lateral walls of the auditory chamber facemedially and incorporate three distinctive structures.A prominent ridge of cuticle (the ventral rod; VR)runs in a rostral–caudal direction roughly in themiddle of the wall. Dorsal to it is a broad, flat depres-sion with a teardrop shape (tympanal depression;TD). This is the tympanum. Its dorsal edge mergeswith the furcasternum. Rostral to the ends of theventral rod and the tympanum is a conical sensorystructure, the bifid sensillum (BS) situated in a circleof thin cuticle, approximately 50 mm in diameter. Inliving and alcohol preserved specimens, all of 74 DKspecies (male and female treated separately) hadlarge bifid sensilla.
The basic DK form is consistent across taxa, butthere are several minor variations. For example, knobshape and size are variable, although the knob angleis always close to 90°. Because ear length does notscale linearly with body length, there is a high appar-ent variability in ear size: large, elongated mantids
Table 2. Summary of the most important distinguishing anatomical characteristics of the four metathoracic forms
DK DNK DO MSMT
Knobs Strong; well-defined Reduced; moderatelywell-defined
Almost none No knobs
Groove configuration Deep; walls close Moderately deep; wallsslightly separated
Very shallow No groove or grooverostral to walls
TD width Wide; flat floor Narrowed; still has flatfloor
Narrow andgroove-like
Slit
Wall orientation Medial Medial or slightventral tilt
Strong ventral tilt Faces posteriorly orpost-eromedially
Wall re: furcal pits Mostly anterior Mostly anterior Mostly or partlyanterior
Entirely posterior toor level with pits
DK, deep groove and prominent knobs; DNK, deep groove, no knobs; DO, knobs are barely visible or absent; MSMT,mesothoracic and metathoracic segments similar; TD, tympanal depression.For detailed descriptions of the characters, see text and Figure 1.
546 D. D. YAGER and G. J. SVENSON
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022
appear to have tiny ears (minimum = 18% of metatho-rax length), and smaller, chunky animals can haveears up to 53% the length of the metathorax.However, animals in the tribe Rivetinini as well asgenera such as Compsothespis and Euchomena dohave ears with an elongated shape. It is characteristicof some Amorphoscelidae, Iridopteryginae, and Caliri-dinae that the areas over the tympanal tracheal sacsbulge and look like big, semitransparent bubbles; theknobs appear sunken in contrast.
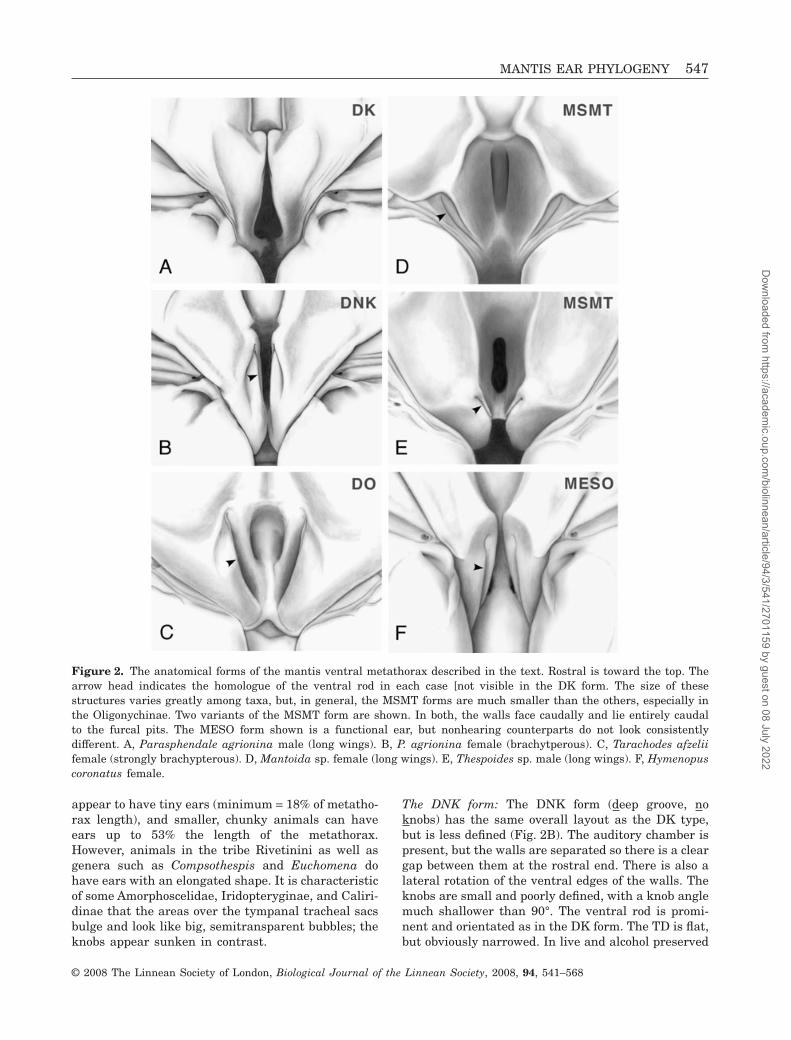
The DNK form: The DNK form (deep groove, noknobs) has the same overall layout as the DK type,but is less defined (Fig. 2B). The auditory chamber ispresent, but the walls are separated so there is a cleargap between them at the rostral end. There is also alateral rotation of the ventral edges of the walls. Theknobs are small and poorly defined, with a knob anglemuch shallower than 90°. The ventral rod is promi-nent and orientated as in the DK form. The TD is flat,but obviously narrowed. In live and alcohol preserved
Figure 2. The anatomical forms of the mantis ventral metathorax described in the text. Rostral is toward the top. Thearrow head indicates the homologue of the ventral rod in each case [not visible in the DK form. The size of thesestructures varies greatly among taxa, but, in general, the MSMT forms are much smaller than the others, especially inthe Oligonychinae. Two variants of the MSMT form are shown. In both, the walls face caudally and lie entirely caudalto the furcal pits. The MESO form shown is a functional ear, but nonhearing counterparts do not look consistentlydifferent. A, Parasphendale agrionina male (long wings). B, P. agrionina female (brachytperous). C, Tarachodes afzeliifemale (strongly brachypterous). D, Mantoida sp. female (long wings). E, Thespoides sp. male (long wings). F, Hymenopuscoronatus female.
MANTIS EAR PHYLOGENY 547
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022

specimens, the bifid sensilla were large in six cases,reduced in eight cases, and absent in nine cases. Thechamber walls are largely rostral to the furcal pits.
The DNK form varies by degree among genera. Insome genera, the anatomy is almost DK (although theneurophysiology shows significantly elevated thresh-olds), whereas in others it is almost DO.
The DO form: In this form, the knobs are barelyvisible or absent (Fig. 2C). The acoustic chamber isopened up by wall separation and lateral rotation(hence DO: deep groove opened). The walls face moreventrally than medially. Bifid sensilla are absent.The ventral rod remains prominent and orientatedpredominantly rostrocaudally, but the TD is a verynarrow space, almost a slit, beneath the rod. Most ofthe ventral rod and tympanal depression are rostralto the furcal pits.
In its mildest form, this configuration is essentiallyand extreme DNK. In the most extreme form, thereis no auditory chamber at all, and the walls faceventrally.
The MSMT form: Although the same components arepresent, the MSMT form (mesothoracic and metatho-racic segments similar) differs radically and qualita-tively from the other types (Fig. 2D, E). The onlypossible similarity would be with the few mostextreme DO cases.
In the MSMT form there is no auditory chamber. Aversion of the ventral rod is present, but orientateddorsoventrally, rather than rostrocaudally. It has aslit-like space beneath its medial edge that corre-sponds to the tympanal depression. The regionaround the rod (the wall) is narrow and faces pre-dominantly posteriorly. The walls, rods, and TDs arealways caudal to the furcal pits. In many cases, thereis a large, ventral opening with the furcal pits at thebottom. However, the wall and TD are outside andcaudal to it, so it is essentially a ‘false auditorychamber.’ The bifid sensilla absent in all cases, includ-ing 12 live and alcohol preserved species. In severalMSMT genera (but not in other forms), there aresocketed hairs along the medial edge of the walls, andin the false auditory chamber. These mechanosensorystructures were not organized in any pattern as theyare in cockroaches (Yager, 2005).
Variants of the MSMT form can look quite differentfrom one another at first glance. In the classic MSMTform found in Chaeteesa, Mantoida, Metallyticus, andthe Thespidae, the entire ‘ear’ region is very small.However, in the aconthopids, the walls are broad andflat, they face straight back toward the abdomen, andcan be partially fused in the midline, which creates alarge, open pit on the rostral side. In some liturgusidssuch as Liturgusa and Hagiomantis and some
angelids such as Angela and Thespoides, the walls arevery small and shifted rostrally, but are nonethelesscaudal to the furcal pits. The Photininae have typicalMSMT characteristics but, in some genera, there is aventral tilt to the wall that creates a resemblance tothe DO configuration.
The Eremiaphilidae are not like any other group,although they have the general characteristics of theMSMT form. The walls are broad, face backward, andare far from the midline, so they appear completelydissociated from the furcal region.
The MESO form: This is the ear in the mesothoracicsegment, which is sensitive to low frequencies ratherthan ultrasound. A functional mesothoracic ear canreliably be confirmed by neurophysiological recording,but not solely by external features.
OccurrenceAs shown in Table 3, the prevalence of the variousanatomical forms in the suborder Mantodea variesconsiderably and differs between the sexes. Strongsexual dimorphism is present in approximately 34%of mantis genera, and Yager (1990) established thatthe dimorphism extends to the auditory system. Inthose cases, males are always DK, and the femaleshave other anatomical forms and reduced hearing.Therefore, we have treated males and females sepa-rately for this analysis, operationally as different‘genera.’ Because there were a number of cases inwhich specimens of only one sex were available for agenus, the sample size is less than twice the numberof genera of the anatomical (552 instead of 688) andphysiological (84 instead of 108) data sets (Table 1).
Seventy-eight percent of mantis species have atleast one sex (male) with a DK ear. The MSMT form,never associated with sexual dimorphism of theventral metathorax, accounts for all other generaexcept the seven (of 344) in which both males andfemales have DNK or DO anatomy (i.e. 97.7% ofmales are DK or MSMT). The seven genera are alsothe only ones with flightless males.
Strong evidence points to a current function ofultrasonic hearing in protecting flying mantids fromcapture by echolocating bats (Yager et al., 1990;Triblehorn, 2003). Because mantids that do not fly arenot vulnerable to predation by echolocating bats, weinvestigated whether there is a correlation betweentypes of ventral metathorax anatomy, and winglength (Table 4; again treating males and females asseparate ‘genera’). In the majority of cases, mantidsthat can fly (FW + MBF) have either the DK (80.5%)or the MSMT (15.7%) forms. Animals with stronglyreduced wings (SB + A) rarely have a DK ear (2.2%).The overall pattern is that the shorter the wings, thelower the probability of a DK ear and the higher the
548 D. D. YAGER and G. J. SVENSON
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022
incidence of the DNK and DO forms. The MSMTform, however, is not associated with a particularwing length.
Physiological patternsOverall description: The approximately 700 physi-ological experiments on 76 species of mantids evalu-ated the firing characteristics of ascending auditoryinterneurones, sensitivity to acoustic stimuli, and thefrequency tuning. The results show a high degree ofuniformity within each anatomical form, and largedifferences among forms. The physiological resultsalso uncovered two auditory system types not evidentfrom external examination alone.
Ascending auditory interneurone activity: Theneurophysiological recordings from the prothoracic-mesothoracic connectives consistently show action
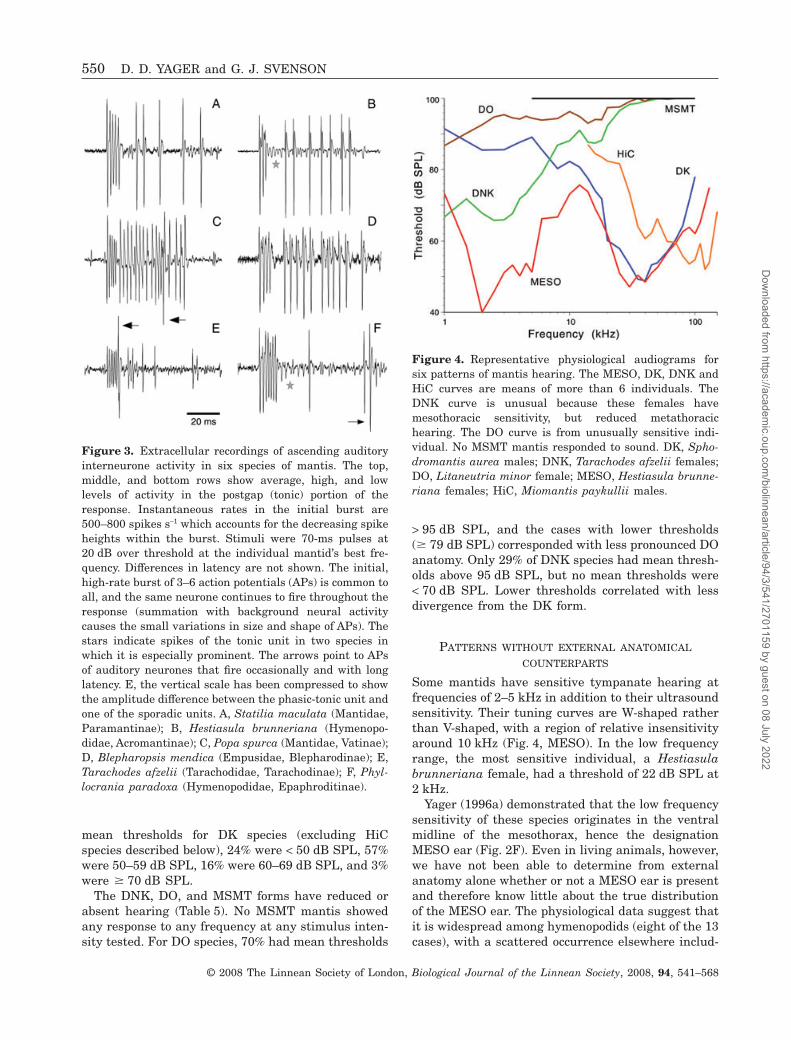
potentials (spikes) from two auditory interneurones(Fig. 3). One neurone with large spikes in the extra-cellular record fires in a very distinctive phasic-tonicpattern: an initial burst of 2–8 spikes with instanta-neous rates of 500–800 s-1; a pause in firing that lasts5–15 ms; resumption of firing, but at lower rates(100–200 s-1) in an irregular tonic pattern. Minimumlatencies were typically 8–13 ms. This firing patternoccurred in every DK species and in DNK animalswith thresholds low enough for testing.
A second auditory neurone, encountered in most, ifnot all, DK species, has small action potentials firedin a strictly tonic pattern. This neurone typically has3–5 dB lower thresholds than the phasic-tonic unit. Insome species, spikes of this neurone were barelydetectable and, in others (hymenopodids, in particu-lar), they were up to half the height of the phasic-tonic neurone’s spikes, which suggests that the axondiameter varies among species.
The extracellular records from the low-frequencymesothoracic ear (see below) also uniformly show twounits, one with very large spikes plus a tonic unitwith small spikes.
Auditory sensitivity and tuning: By contrast to theconsistency of the auditory interneurones and theirfiring patterns, the tuning and sensitivity of the audi-tory system falls into several categories, four of whichcorrespond to anatomical forms (Fig. 4, Table 5).
PATTERNS WITH ANATOMICAL COUNTERPARTS
For the ‘standard’ DK ear, the audiograms areV-shaped with best sensitivity at frequencies in therange 25–50 kHz. The lowest best frequency weencountered was 18 kHz. The most sensitive animalin the study was a female Sphodromantis lineola witha threshold of 12 dB SPL at 30 kHz. Considering
Table 3. Occurrence of the morphological types in the anatomical and physiological data sets expressed as percentagesof the total number of genera for each sex and for the full data set
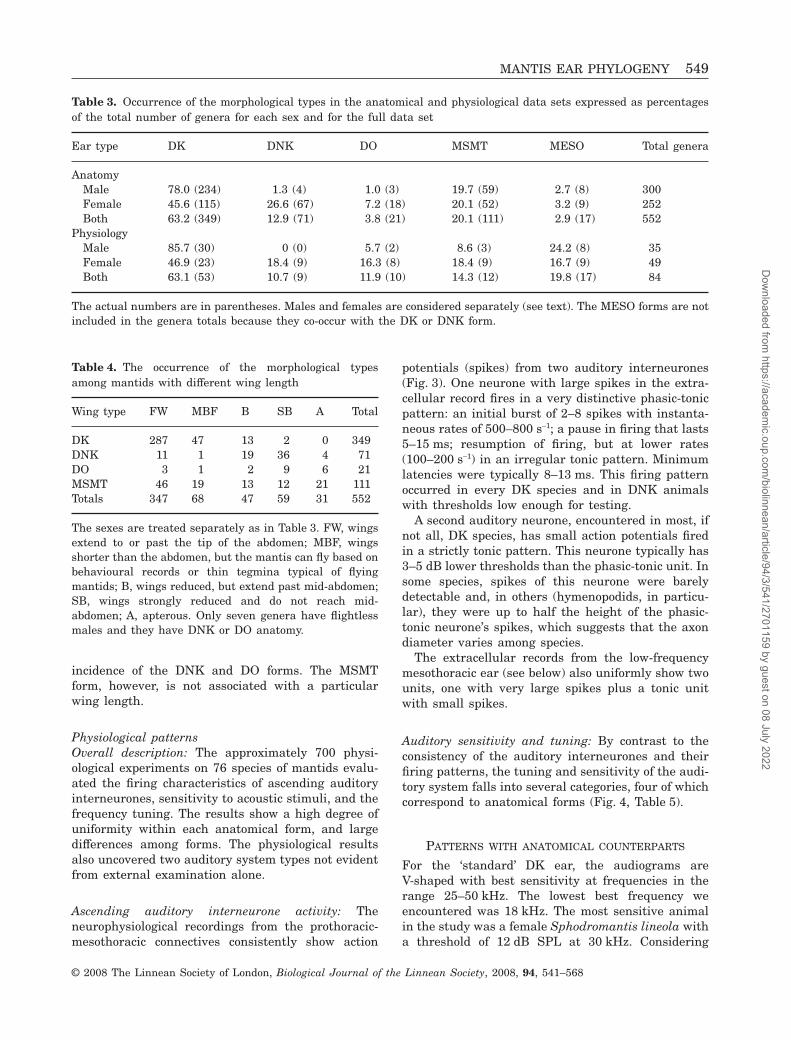
Ear type DK DNK DO MSMT MESO Total genera
AnatomyMale 78.0 (234) 1.3 (4) 1.0 (3) 19.7 (59) 2.7 (8) 300Female 45.6 (115) 26.6 (67) 7.2 (18) 20.1 (52) 3.2 (9) 252Both 63.2 (349) 12.9 (71) 3.8 (21) 20.1 (111) 2.9 (17) 552
PhysiologyMale 85.7 (30) 0 (0) 5.7 (2) 8.6 (3) 24.2 (8) 35Female 46.9 (23) 18.4 (9) 16.3 (8) 18.4 (9) 16.7 (9) 49Both 63.1 (53) 10.7 (9) 11.9 (10) 14.3 (12) 19.8 (17) 84
The actual numbers are in parentheses. Males and females are considered separately (see text). The MESO forms are notincluded in the genera totals because they co-occur with the DK or DNK form.
Table 4. The occurrence of the morphological typesamong mantids with different wing length
Wing type FW MBF B SB A Total
DK 287 47 13 2 0 349DNK 11 1 19 36 4 71DO 3 1 2 9 6 21MSMT 46 19 13 12 21 111Totals 347 68 47 59 31 552
The sexes are treated separately as in Table 3. FW, wingsextend to or past the tip of the abdomen; MBF, wingsshorter than the abdomen, but the mantis can fly based onbehavioural records or thin tegmina typical of flyingmantids; B, wings reduced, but extend past mid-abdomen;SB, wings strongly reduced and do not reach mid-abdomen; A, apterous. Only seven genera have flightlessmales and they have DNK or DO anatomy.
MANTIS EAR PHYLOGENY 549
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022
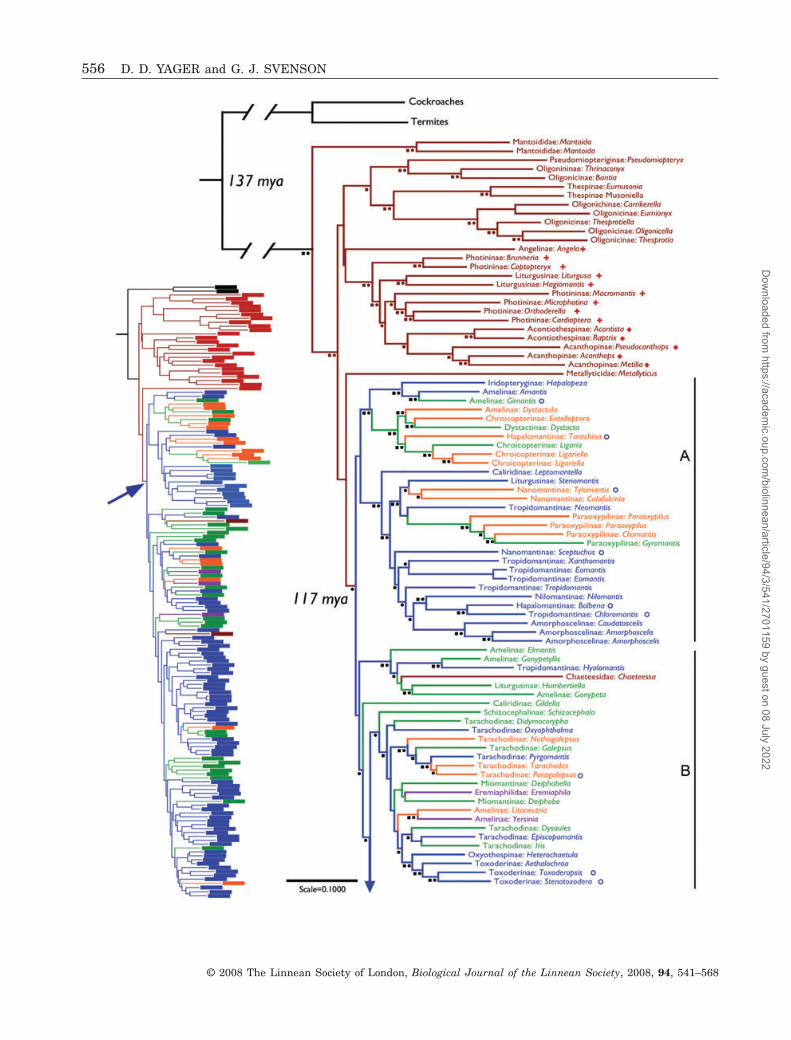
mean thresholds for DK species (excluding HiCspecies described below), 24% were < 50 dB SPL, 57%were 50–59 dB SPL, 16% were 60–69 dB SPL, and 3%were � 70 dB SPL.
The DNK, DO, and MSMT forms have reduced orabsent hearing (Table 5). No MSMT mantis showedany response to any frequency at any stimulus inten-sity tested. For DO species, 70% had mean thresholds
> 95 dB SPL, and the cases with lower thresholds(� 79 dB SPL) corresponded with less pronounced DOanatomy. Only 29% of DNK species had mean thresh-olds above 95 dB SPL, but no mean thresholds were< 70 dB SPL. Lower thresholds correlated with lessdivergence from the DK form.
PATTERNS WITHOUT EXTERNAL ANATOMICAL
COUNTERPARTS
Some mantids have sensitive tympanate hearing atfrequencies of 2–5 kHz in addition to their ultrasoundsensitivity. Their tuning curves are W-shaped ratherthan V-shaped, with a region of relative insensitivityaround 10 kHz (Fig. 4, MESO). In the low frequencyrange, the most sensitive individual, a Hestiasulabrunneriana female, had a threshold of 22 dB SPL at2 kHz.
Yager (1996a) demonstrated that the low frequencysensitivity of these species originates in the ventralmidline of the mesothorax, hence the designationMESO ear (Fig. 2F). Even in living animals, however,we have not been able to determine from externalanatomy alone whether or not a MESO ear is presentand therefore know little about the true distributionof the MESO ear. The physiological data suggest thatit is widespread among hymenopodids (eight of the 13cases), with a scattered occurrence elsewhere includ-
Figure 3. Extracellular recordings of ascending auditoryinterneurone activity in six species of mantis. The top,middle, and bottom rows show average, high, and lowlevels of activity in the postgap (tonic) portion of theresponse. Instantaneous rates in the initial burst are500–800 spikes s-1 which accounts for the decreasing spikeheights within the burst. Stimuli were 70-ms pulses at20 dB over threshold at the individual mantid’s best fre-quency. Differences in latency are not shown. The initial,high-rate burst of 3–6 action potentials (APs) is common toall, and the same neurone continues to fire throughout theresponse (summation with background neural activitycauses the small variations in size and shape of APs). Thestars indicate spikes of the tonic unit in two species inwhich it is especially prominent. The arrows point to APsof auditory neurones that fire occasionally and with longlatency. E, the vertical scale has been compressed to showthe amplitude difference between the phasic-tonic unit andone of the sporadic units. A, Statilia maculata (Mantidae,Paramantinae); B, Hestiasula brunneriana (Hymenopo-didae, Acromantinae); C, Popa spurca (Mantidae, Vatinae);D, Blepharopsis mendica (Empusidae, Blepharodinae); E,Tarachodes afzelii (Tarachodidae, Tarachodinae); F, Phyl-locrania paradoxa (Hymenopodidae, Epaphroditinae).
Figure 4. Representative physiological audiograms forsix patterns of mantis hearing. The MESO, DK, DNK andHiC curves are means of more than 6 individuals. TheDNK curve is unusual because these females havemesothoracic sensitivity, but reduced metathoracichearing. The DO curve is from unusually sensitive indi-vidual. No MSMT mantis responded to sound. DK, Spho-dromantis aurea males; DNK, Tarachodes afzelii females;DO, Litaneutria minor female; MESO, Hestiasula brunne-riana females; HiC, Miomantis paykullii males.
550 D. D. YAGER and G. J. SVENSON
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022

ing the Orthoderinae, Paramantinae, the Deroplatyi-nae, and the Tarachodidae. The last two MESO taxaare especially interesting because the females haveDNK anatomy with ultrasound thresholds > 85 dBSPL and therefore have a V-shaped tuning curve, butcentred at low frequencies.
A second pattern only detectable physiologically(and behaviourally; Triblehorn & Yager, 2001)involves best frequencies of 60–130 kHz, unusuallybroad tuning curves, and generally higher thresholdsthan those of other DK mantids (Table 5). We use thedesignation HiC (high cyclops) for this pattern. Themean tuning curve shown in Figure 4 is fromMiomantis paykullii, the species with both the lowestmean threshold (53 dB SPL) and the highest bestfrequency (130 kHz). Three of the 11 HiC species inthe data set have best frequencies � 100 kHz. Thebreadth of the tuning curves arises primarily from asecondary sensitivity region in the range 25–45 kHz.
As with the MESO ear, the HiC pattern may bemuch more prevalent than our data indicate. Thenine genera so far discovered (Ameles, Empusa,Euchomenella, Hymenopus, Miomantis, Pseudem-pusa, Stagmomantis, Taumantis, and Tropidomantis)come from three families and six subfamilies. Stag-momantis centralis fits least well with the other HiCspecies. It is the only New World representative, itsmean threshold is 10 dB higher than the next leastsensitive species, and several other species in thegenus have conventional DK hearing.
BEHAVIOURAL PATTERNS
Responses to ultrasound only occurred when themantis was stimulated during flight. The data comepredominantly from males. because females of manyspecies were reluctant fliers or had reduced hearing.Behavioural tuning agreed with the physiologicaltuning. The behavioural results from 30 speciesrevealed three patterns.
By far, the most common pattern (18 species) com-prises four components: prothoracic leg extension,head roll, wing beat changes (wing ‘hitch’), andabdomen dorsiflexion. Ultrasound caused flight cessa-tion in � 1% of trials. In 80–85% of responses, all fourcomponents were present and strong; in the remain-der one or more components were partial. Meanlatencies were 72, 87, 92, and 137 ms, respectively,and the response was fully developed within 200–250 ms poststimulus. The greatest variation amongspecies was in the relative robustness of the fourcomponents. For example, Hierodula grandis usedonly weak, sporadic head rolls. Animals with broad orplump abdomens produced small abdominal dorsiflex-ions, presumably for purely mechanical reasons. Theprothoracic leg extension and the wing hitch were theleast variable components.
Species in the HiC physiological group displayed adifferent behavioural pattern. First, these were theonly species that included flight cessation in theirresponses. Second, the behaviour of individuals wasunpredictabe both among and within individuals.These mantids used random combinations of the fivecomponents so that, for example, response #1 might bearms and abdomen, #2 cessation, #3 head, arms andwings, #4 abdomen and wings. All four usual compo-nents occurred together in only 25–35% of trialsdepending on the species. Flight cessation followed byresumed flight was present in 20–25% of trials, andcomplete cessation occurrent in an additional 15–20%of trials.
The third pattern occurred in only two species.Mantis religiosa and Tenodera aridifolia sinensishave never responded behaviourally to ultrasounddespite normal DK anatomy and sensitive ultrasonichearing. We tried animals from different locationsusing a range of stimulus parameters and times ofday. These two species are not native to the USA, sowe obtained eggs from within their natural ranges.The resulting adults had normal hearing, but did not
Table 5. Summary of the physiological characteristics of the six auditory patterns taken from the mean tuning curves
Ear type
DK HiC DNK DO MSMT MESO
TH BF TH BF TH BF TH BF TH BF TH BF
Mean 55.1 34.9 65.1 86 > 82 30 > 88.9 25.0 > 95 NA 58.1 2.97SD 8.5 8.0 9.6 24.1 12.2 14.3 9.6 5.0 0 NA 8.7 0.5N 67 67 10 10 16 16 8 8 13 13 22 22Maximum 79 50 85 130 > 95 70 > 95 30 NA NA 69 4Minimum 29 20 53 60 58 15 70 20 NA NA 36 2
NA, not applicable.The number of individuals tested for each curve ranged from 1 to > 60. The two sexes are treated separately as in previoustables. TH, lowest threshold in dB SPL. BF, frequency (kHz) with the lowest threshold.
MANTIS EAR PHYLOGENY 551
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022

respond to ultrasound. In contrast, Tenodera super-stitiosa from South Africa does respond with a normalfour-component behaviour.
Despite testing in a variety of behavioural contextswith many different stimulus types, we have as yetfound no behavioural counterpart to the low-frequency hearing of the MESO ear.
PHYLOGENY OF THE ANATOMICAL TYPES
To assess the evolutionary relationships among thepatterns of ventral metathoracic structure, we gen-erated morphological phylogenetic trees for male andfemale mantids using 13 anatomical characters. Apriori character weighting was included to increaseresolution in the consensus trees for both sexes. Theweighting was determined by the strong correlationof some metathoracic characters (knobs, bifid sen-silla) with low auditory thresholds. Appendix 1 givesthe complete character matrix for all genera in theanalysis.
CHARACTER DESCRIPTIONS
The character descriptions below can be referenced toFigures 1 and 2 and the morphological descriptionsabove.
GrooveThe groove is the invagination between the meta-thoracic coxae at the caudal end of the ventralmetathorax.
1. Groove: absent (0) or present (1); weight = 12. Groove length relative to metathorax: < 0.25 (1),
0.25–0.5 (2), or > 0.5 (3); unordered; weight = 13. Groove shape: circular (1), slightly rounded (2), or
straight (3); unordered; weight = 14. Groove depth: deep (1) or shallow (2); weight = 1
KnobsWhen present, we measured the angle formedbetween the anterior edges of the knobs and thelongitudinal body axis.
5. Knobs: absent (0) or present (1); weight = 66. Knob angle: < 60° (1), 60–90° (2), or � 90° (3);
ordered; weight = 3
WallsThese are a bilaterally symmetrical pair of flat areasbetween the coxae that contain the longitudinaldepression. Walls are always present.
7. Wall: outside (0) or inside groove (1); weight = 68. Wall orientation: strong caudal component (0),
medial (1), ventromedial (2), or ventral (3);ordered; weight = 6
9. Wall separation: wide (1), narrow (2), or almostnone (3); ordered; weight = 3
Bifid sensilla10. Sensillum: absent (0) or present (1); weight = 6
Longitudinal (tympanal) depressionsAlways present, these are straight depressions ofvarying width located in the walls near the midline.
11. TD shape: slit-like (0), groove-like (1), narrowwith flat floor (2), or wide with flat floor (3);unordered; weight = 6
12. TD orientation: predominantly dorsal-ventral (0),rostral-caudal with dorsal-ventral tilt (1), orrostral-caudal (2); ordered; weight = 3
13. TD location: caudal to furcal pits (0), at leastpartly rostral to furcal pits (1); weight = 6
EVOLUTIONARY PATTERNS
Parsimony analysis of the 13 weighted and orderedmale morphology characters resulted in 34 trees ofequal length (tree length = 221; CI = 0.56; RI = 0.98).For the female data set, there were 74 trees of equallength (tree length = 398; CI = 0.46; RI = 0.98). Wecalculated a strict consensus tree for each sex (Fig. 5).There are many resolved nodes, but relatively lowdiscrimination among lineages, probably because ofthe relatively low number of characters relative to thenumber of genera. We mapped selected characterstate changes on deep level, resolved nodes usingACCTRAN optimization in lieu of Bremer and boot-strap values.
The overall tree topologies for both males andfemales are similar (Fig. 5). They show three basicmetathoracic configurations: (1) a form like MSMT(defined in Table 2) at the base of the trees. Theoutgroups and the second instar nymphs clustertogether in the most basal lineage in each case. Thephysiology confirms that these genera do not hear;(2) the combination of characters that defines the DKmorphological form clearly evolved just once (nodes Cand F in females and males, respectively). After thatdivergence, males have a single, uniform clade withDK anatomy and sensitive hearing. The correspond-ing female clade combines DK and sexually dimorphicforms, and auditory thresholds span the full range.The analysis largely did not resolve differences be-tween the DK and DNK forms, but the genera withDNK females are all at the distal end of the tree. Thetwo MSMT genera in the female DK clade (a litur-gusid from Madagascar and a Photininae from Brazil)had anatomy that strongly resembled the DO form insome respects; (3) there are combinations of charac-ters intermediate between the most basal and most
552 D. D. YAGER and G. J. SVENSON
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022

distal groups that most closely correspond to the DOmetathorax. However, some previously defined DOgenera cluster in the MSMT groups, and others, inthe female tree, cluster with DK genera. This sug-gests that the DO form as defined by the morphologyis not ‘monophyletic’ and makes it impossible to con-sider the entire type basal or derived.
The trees reflect similar sequences of charactertransitions leading to the morphology linked tohearing. Earliest is the infolding of the walls to formthe groove, the precursor to the auditory chamber(character 1; nodes A). The tympanal depressions(and the walls in males) then rotate rostrally, whichputs them at least partly in front of the furcal pits(12 and 13; nodes B). Finally, the knobs (5 and 6; nodeC in females and D/E in males) and bifid sensilla(10; nodes C and F) make their appearance.
The effects of ordering and weighting the charac-ters on nodal recovery differed between the sexes. Formales, the analysis with no character ordering orweighting recovered three of the six nodes inFigure 5D, E, F, and ordering without weightingrecovered five of the six nodes (A, C, D, E, and F). Thenumber of equally parsimonious trees was 642 and1659, respectively, with lengths of 65 and 70, respec-tively. For females, only the analysis with orderedand weighted characters yielded any of the threenodes indicated in Figure 5. The very few nodes thatwere recovered in the other analyses were moreterminal.
EAR PHYLOGENY AND MANTODEAN PHYLOGENY
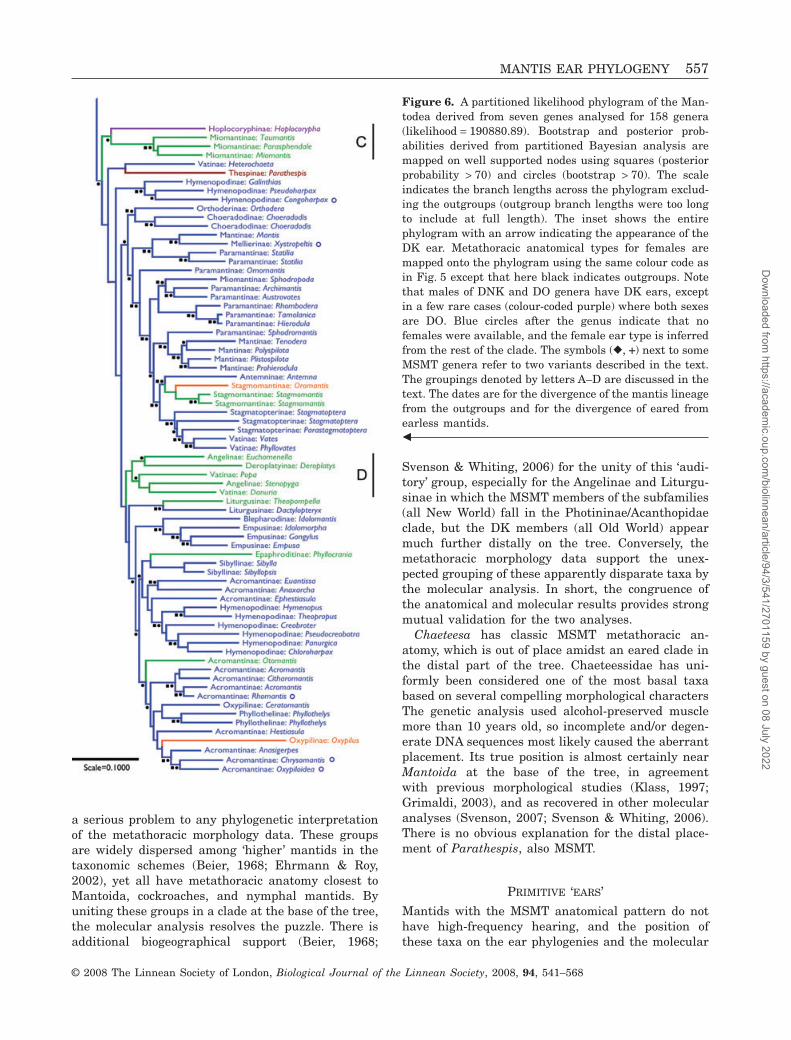
Phylogenetic treeThe Mantodea phylogenetic tree derived from analy-sis of the seven genes is shown in Figure 6.
The GTR + G + I model was the best fit for the sevengene loci, although 16S rDNA, 12S rDNA, and cyto-chrome oxidase subunit II required restricted ver-sions. Partitioned phylogenetic reconstruction usingTREEFINDER recovered a tree with a likelihoodscore of -190880.89 (Fig. 6). Nonparametric bootstrapvalues demonstrate good support for most of thehigher clades, but lower values for the majority ofdeeper nodes (Fig. 6), which is consistent with previ-ous studies (Svenson & Whiting, 2004).
Partitioned Bayesian analysis recovered a topologythat was largely congruent with the likelihood tree.We considered only generations after the 1000 000thin the four independent runs. The compiled majorityrule consensus tree demonstrated high posterior prob-abilities for most of the nodes recovered by likelihoodanalysis (Fig. 6). A total of 88 out of 154 nodes showeda posterior probability greater than 0.70. As with thebootstrap values, high posterior probabilities (70% orabove) were primarily clustered in higher clades, and
deeper level nodes were not recovered in the Bayesianconsensus tree.
Metathoracic anatomy and phylogenySuperimposing anatomical types as defined by themorphology onto the phylogenetic tree for the subor-der revealed several clear patterns (Fig. 6).
The MSMT form clusters with the genera at thebase of the tree (Fig. 6A). There are five clades withinthis overall group. Mantoida and the Thespidae allhave the classic MSMT form. The genus Angela andthe clade containing the Photininae, the Acontistinae,the Acanthopidae, and two genera from the Litur-gusidae all have MSMT subtypes described above.Metallyticus has metathoracic anatomy similar toMantoida, and is the only MSMT genus outside theNew World. Two subfamilies (Liturgusinae and Ange-linae) represented by genera in the early branchingMSMT clades are notable because they also containgenera located more distally on the tree and that haveDK ears. There are two exceptions to the basalcorrespondence of the MSMT form, Chaeteesa andParathespis.
The phylogeny indicates a single evolution of theDK ear, which is consistent with the anatomical,physiological, cladistic, and behavioural data. Exceptfor a very small number of special cases, all taxa afterthat point have DK anatomy, taking into account thatthe DNK and DO forms are limited to sexually dimor-phic taxa where the males have DK ears. If the DOform were primitive (i.e. a MSMT variant; Fig. 5), itsdistribution would require, at absolute minimum,nine independent evolutions of the DK ear. The nodeat which the DK ear appeared maintained moderatebootstrap support (75%), which indicates a true sepa-ration of mantids with functional ears from mantidswith primitive metathoracic morphology.
Among the DK taxa, the DNK and DO forms occurin discrete clades distributed widely at the distal endof the lineage. A conservative count yields > 15appearances of DNK/DO lineages. For discussion, wehave denoted four groups of taxa in Figure 6 thatcontain DNK and/or DO taxa. Group A is most diversewith a mix of DK and DNK/DO clades. It contains theTropidomantinae and Amorphoscelinae that share aDK subtype with large, recessed knobs and hugebulges over the tympanal tracheal sacs. It also con-tains some of the Amelinae, a subfamily highly vari-able in ear type. The Amelinae contains four genera(out of seven in the entire data set) in which bothmales and females have strongly reduced wings andDNK or DO anatomy.
Groups B and C are largely dimorphic in ear typeand wing length with the exception of the Toxode-ridae. Based on its position embedded in group B, theunique metathoracic morphology of Eremiaphila (see
MANTIS EAR PHYLOGENY 553
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022

554 D. D. YAGER and G. J. SVENSON
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022

above) appears to be derived rather than a variant ofthe MSMT form. The Hoplocoryphinae in group C arefar from the rest of the Thespidae and also have anunusual ventral metathoracic anatomy that is clearlynot MSMT, but may be an extreme variation of theDO form. Neither Eremiaphila nor the Hoplocoryphi-nae are sexually dimorphic for metathoracic anatomy.
Whereas the middle third of the tree is dominated bydimorphic taxa, the distal third is solidly DK with onlyoccasional, isolated clades of dimorphic genera (e.g.group D). Even in those, the auditory dimorphismtends to be mild, and the DO form is uncommon.Occurrences of isolated DNK/DO genera reflect incom-plete sampling in most cases (e.g. all Epaphroditinaein the larger anatomical data set are DNK like Phyl-locrania; all the Stagmomantinae are DNK or DO).
DatesComputation of divergence dates for two nodes ofinterest in the molecular tree (Fig. 6) yielded a date of117.33 Mya for the lineage in which the DK earevolved (95% CI = 109.2–130.9 Mya). The divergenceof mantids from the cockroach and termite outgroupsoccurred at the beginning of the Cretaceous,136.8 Mya (95% CI = 123.9–149.6 Mya).
DISCUSSIONSUMMARY
The present study combines large anatomical, neuro-physiological, and behavioural databases to identify aprimitively earless form of metathoracic anatomy(MSMT) found in several extant mantis taxa, and todefine five different mantis auditory system types.Three of these systems can be identified by externalanatomy and two can so far be detected only byneurophysiology. By superimposing the results ofthese studies onto a phylogenetic tree derived frommolecular data, we have shown that the cyclopeanmantis ear (DK) evolved once approximately120 Mya, and all the other auditory system types areeither varying degrees of loss associated with wingreduction (DNK and DO), or are much more recentinnovations that occurred independently multipletimes (MESO and HiC). The mutually reinforcingcombination of anatomical, behavioural, and neuro-
physiological data with the molecular phylogeny pro-vides an extensive picture of praying mantis auditorysystem evolutionary patterns.
MATCHING ANATOMICAL AND MOLECULAR
PHYLOGENIES
The attempt to make sense of the metathoracic mor-phological types using cladistic techniques met withmoderate success (Fig. 5). Most significantly, thesimilarity of this tree with that of the phylogeneticanalysis (Fig. 6) confirms the MSMT form as mostprimitive and strongly supports a single appearanceof the DK ear. The distribution of physiologicalthresholds mirrors the anatomical patterns. The mor-phological analysis also revealed a sequence of tran-sitions (infolding, rotation, knobs/bifid sensilla) thatmatches the ontogenetic sequence of ear formation(Yager, 1996b) in which a gradual infolding and rota-tion of the TD and walls takes place over the last halfof nymphal development. As would be expected, thesequence matches the evolutionary transition hypoth-esized from a study of the homologous components incockroaches, one of the outgroups (Yager, 2005).
The analysis left the relationship between thesimilar DK and DNK forms unresolved in females. Inaddition, the DO form grouped entirely basal to theDK ear in males, but was scattered throughout thefemale tree (Fig. 5). To some extent, these resultsreflect similarities between extreme DO and MSMTforms as well as the exclusive occurrence of DOanatomy in sexually dimorphic females. The maletree proved robust, returning the same basic topologyin all variations of the analysis. However, the depen-dence of the female tree on character ordering andweighting suggests that more characters would berequired to yield clear resolution of morphologicaltypes in females.
Superimposing metathoracic anatomy onto generain the molecular tree (Fig. 6) shows a remarkablecongruence between two entirely different, indepen-dently collected data sets. The match is not only inlarger groupings, but also even extends to small, butwell-defined clades of DNK/DO genera. Prior to thisstudy, the Photininae, Acanthopidae, and some rep-resentatives of the Liturgusidae and Angelinae posed
Figure 5. Strict consensus phylogenies of male and female ventral metathoracic types derived from 13 anatomicalcharacters. Female anatomical analysis resulted in 74 trees of equal length (tree length = 398; CI = 0.46; RI = 0.98). Maleanatomical analysis resulted in 34 trees of equal length (tree length = 221; CI = 0.56; RI = 0.98). The anatomical types asdefined in Table 2 are colour-coded for each genus: red, MSMT; blue, DK; green, DNK; orange, DO (knobs are barelyvisible or absent); purple, outgroups; black, mantis nymph. The blue stars indicate the appearance of the DK ear. Todemonstrate which characters support the deep-level, resolved nodes, a selection of character state changes are shown tothe left of each node (Character: State A → State B). Nodes are labelled with letters for referencing within text. Meanphysiological thresholds (dB SPL) to ultrasound are indicated for tested genera. X, > 95; �, 70–90; �, < 70.�
MANTIS EAR PHYLOGENY 555
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022
556 D. D. YAGER and G. J. SVENSON
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022
a serious problem to any phylogenetic interpretationof the metathoracic morphology data. These groupsare widely dispersed among ‘higher’ mantids in thetaxonomic schemes (Beier, 1968; Ehrmann & Roy,2002), yet all have metathoracic anatomy closest toMantoida, cockroaches, and nymphal mantids. Byuniting these groups in a clade at the base of the tree,the molecular analysis resolves the puzzle. There isadditional biogeographical support (Beier, 1968;
Svenson & Whiting, 2006) for the unity of this ‘audi-tory’ group, especially for the Angelinae and Liturgu-sinae in which the MSMT members of the subfamilies(all New World) fall in the Photininae/Acanthopidaeclade, but the DK members (all Old World) appearmuch further distally on the tree. Conversely, themetathoracic morphology data support the unex-pected grouping of these apparently disparate taxa bythe molecular analysis. In short, the congruence ofthe anatomical and molecular results provides strongmutual validation for the two analyses.
Chaeteesa has classic MSMT metathoracic an-atomy, which is out of place amidst an eared clade inthe distal part of the tree. Chaeteessidae has uni-formly been considered one of the most basal taxabased on several compelling morphological charactersThe genetic analysis used alcohol-preserved musclemore than 10 years old, so incomplete and/or degen-erate DNA sequences most likely caused the aberrantplacement. Its true position is almost certainly nearMantoida at the base of the tree, in agreementwith previous morphological studies (Klass, 1997;Grimaldi, 2003), and as recovered in other molecularanalyses (Svenson, 2007; Svenson & Whiting, 2006).There is no obvious explanation for the distal place-ment of Parathespis, also MSMT.
PRIMITIVE ‘EARS’
Mantids with the MSMT anatomical pattern do nothave high-frequency hearing, and the position ofthese taxa on the ear phylogenies and the molecular
Figure 6. A partitioned likelihood phylogram of the Man-todea derived from seven genes analysed for 158 genera(likelihood = 190880.89). Bootstrap and posterior prob-abilities derived from partitioned Bayesian analysis aremapped on well supported nodes using squares (posteriorprobability > 70) and circles (bootstrap > 70). The scaleindicates the branch lengths across the phylogram exclud-ing the outgroups (outgroup branch lengths were too longto include at full length). The inset shows the entirephylogram with an arrow indicating the appearance of theDK ear. Metathoracic anatomical types for females aremapped onto the phylogram using the same colour code asin Fig. 5 except that here black indicates outgroups. Notethat males of DNK and DO genera have DK ears, exceptin a few rare cases (colour-coded purple) where both sexesare DO. Blue circles after the genus indicate that nofemales were available, and the female ear type is inferredfrom the rest of the clade. The symbols (�, +) next to someMSMT genera refer to two variants described in the text.The groupings denoted by letters A–D are discussed in thetext. The dates are for the divergence of the mantis lineagefrom the outgroups and for the divergence of eared fromearless mantids.�
MANTIS EAR PHYLOGENY 557
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022

tree establishes that their earlessness is primitiveand not a secondary loss. Both the second-instarnymph metathorax and the homologous region incockroaches (Yager, 2005) cluster with the MSMTform. In cockroaches, however, the region has manysensory structures, including fields of socketed hairs,a chordotonal organ, and a proprioceptive complexcomprising ranks of socketed hairs, thinned cuticle,and a tensioning muscle. This is consistent withseveral studies showing that the function of auditoryorgan precursors is proprioception (Meier & Reichert,1990; Yack & Fullard, 1990; Lakes-Harlan et al.,1999; Yack, 2004). In DK-eared mantids, only thechordotonal organ and thinned cuticle remain. Amajor cost of transition from earless (cockroach) toeared (mantis) may be loss of proprioceptive feedbackfrom that area.
The fossil mantis Santanmantis axelrodi, dated tothe early Cretaceous (120 Mya), has features of theventral metathorax even more similar to those of P.americana (D. D. Yager, unpubl. data). A broad wall ispresent lateral to the furcasternum, and the impres-sion of what may be the lateral ventropleurite, theputative precursor to the ventral rod (Yager, 2005), isvisible.
The genus Eremiaphila has often been consideredamong the more primitive mantids based partly onits almost square pronotum (Beier, 1968; Grimaldi,2003). Its distinctive metathoracic anatomy and itsposition on the molecular tree argue against this.What appears to be a very extreme secondary loss ofthe ear resulted in a metathoracic anatomy differentfrom both the MSMT and DO forms, although moreakin to the primitive state. The thoracic anatomy ofEremiaphila undoubtedly reflects an overall bodymorphology specialized for a highly cursorial lifestylein its desert habitat.
LOST EARS
The superposition of anatomical forms on the molecu-lar tree confirms that the DNK and DO anatomies aresecondary losses of the DK ear, the only reasonableconclusion because males in the same genera haveDK ears. Both anatomical and physiological data indi-cate a continuum of ear reduction and hearing loss,DK to extreme DO. Even mild anatomical DNK-nessresults in elevated thresholds (> 70 dB SPL), distin-guishing it from the DK ear.
The middle third of the phylogenetic tree is distinc-tive for its high density of DNK/DO taxa, whichindicates a high incidence of sexual dimorphism withflight loss in females. Under appropriate ecologicalconditions, sacrifice of flight/wings for increasedfecundity is advantageous (Roff et al., 1999). Thus,the high DNK/DO incidence undoubtedly reflects
patterns of Mantodean evolution rather than auditoryevolution per se.
Secondary reduction or loss of hearing is not unusualand has been well documented in locusts and grass-hoppers (Mason, 1968; Riede, Kämper & Höfler, 1990),crickets (Otte, 1990), and moths (Fullard, 1998; Muma& Fullard, 2004). A common feature of all these groupsis many independent appearances of auditory reduc-tion. As in mantids, these changes are most frequentlylinked to loss of flight ability, usually accompanied byreduced wings. On the behavioural side, the linkbetween flight and ultrasonic hearing is predationpressure from bats. Flightlessness removes the preda-tion risk, therefore ultrasonic hearing confers noadvantage and is reduced or lost. The exceptions occurwhen hearing is used for intraspecific communicationas well as predator avoidance, as in some moths(Connor, 1999) and in crickets (Otte, 1990).
NEW EARS
When first documented (Yager, 1996a), the MESO earappeared limited to two subfamilies of the Hymenopo-didae. Its actual distribution in several unrelatedlineages raises two points. First, the transformationfrom earless to eared in the mesothorax must follow asimple path. Given the conservative nature of nervoussystems (Dumont & Robertson, 1986; Edwards &Palka, 1991; Paul, 1991; Tierney, 1995), the keychanges are most likely biomechanical, such as broad-ening of thinned cuticular areas and apposition ofenlarged tracheal sacs to that cuticle. As graphicallydemonstrated in pneumorid grasshoppers (vanStaaden & Römer, 1998; van Staaden et al., 2003),serially homologous regions can have a shared poten-tial for transformation to an auditory organ. Althoughindependently evolved, such segmental ears areactually the same in an evolutionary–developmentalsense (Raff, 1996; Yager, 1999a). By similar argu-ments, we expect to find serially homologous interneu-rones subserving auditory function in the mesothorax(Prier & Boyan, 2000). Second, there must be somecompelling selective advantage to the low frequencyhearing that all MESO taxa share. So little is knownabout the natural history of most mantids, that themost profitable next step to understanding MESO earevolution may be intensive field studies.
The modifications producing the broad tuningcurves and very high best frequencies of HiC mantids,are also most likely to be bioacoustic. The concurrentbehavioural differences, however, make it clear thatthe entire auditory system, from input to output, hasevolved new patterns. The HiC evasive behaviouruses the same components as do DK mantids, so thecentral nervous system circuitry changes need not beelaborate (e.g. changes in relative synaptic strengths,
558 D. D. YAGER and G. J. SVENSON
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022

decoupling of behavioural modules through inhibitionor desynchronization). It would be especially interest-ing to know the circuit design that generates therandom behavioural component combinations. Unpre-dictable behaviour can be a powerful escape strategy[‘evitability’ sensu Roeder (1971)] and increases theeffectiveness in bat evasion of the erratic flight ofnoctuid moths, the wing flicks of green lacewings(Miller & Olesen, 1979), and the nondirectional divesof DK-eared mantids (Triblehorn, 2003).
HiC hearing has the same type of patchy distribu-tion as the MESO ear. There is even one case, Hymeno-pus, in which both innovations occur. A plausibleexplanation is that HiC mantids are especially vulner-able to rhinolophid, hipposiderid, and/or other batsthat use echolocation frequencies > 60 kHz (Fentonet al., 1998; Fullard, 1998). Cumming (1996) showed,in a field study, that one HiC species, Miomantisnatalica, successfully evaded capture by a rhinolophidbat echolocating at 80–85 kHz. The puzzle remains asto why a few Old World mantids have very high-frequency hearing and modified evasive behaviourwhile others in the same geographical regions do not.
METATHORACIC ANATOMY AS
A TAXONOMIC CHARACTER
The anatomy of the ventral metathorax can be helpfulin identifying mantis taxa at the subfamily and some-times genus level. The DK ear anatomy is very stablecompared to the high degrees of convergence found inmany of the traditional taxonomic characters (Beier,1968; Ehrmann & Roy, 2002; Svenson & Whiting,2006; Svenson, 2007). The contrast between MSMTand DK forms is most useful because the DNK or DOforms are so tightly correlated to wing length, a moreobvious character. For example, some clear examplessupport changes of Beier’s 1968 taxonomic schemethat were proposed by Roy (1999) and by Ehrmann &Roy (2002):
• the Thespinae of Beier (all MSMT) do not belongwithin the Mantidae (predominantly DK) and havebeen elevated to family status
• Beier placed Stenophylla in the Toxoderinae (all DK)and Callibia in the Hymenopodinae (all DK), butboth have the same MSMT subtype and now have abetter home among the MSMT Acanthopidae
• the Epaphroditini was a mix of MSMT and DKforms and the former are now with the MSMTAcanthopidae
On the other hand, the metathoracic anatomysuggests several further opportunities for revisionincluding:
• none of the Old World Thespidae have the MSMTanatomy typical of the New World genera (Sinomi-
opteryx is DK; the Hoplocoryphinae are apparentlyan extreme DO; the Haaniinae are DK; see abovefor Parathespis)
• the genus Angela (MSMT; Neotropics) differs fromall other Angelinae (DK; Old World)
• the family Liturgusidae contains both MSMT andDK genera and is unlikely to be monophyletic
• the Photininae with their MSMT variant do not fitamong the Mantidae (DK)
• both sexes of the genus Callimantis are fullywinged with DO morphology, a very rare combina-tion that does not match any other Miomantini(Beier, 1968) or Stagmomantinae (Ehrmann & Roy,2002).
AUDITORY DISTRIBUTION
The anatomical data, with confirmation from thephysiological data, show that ultrasonic hearing isvery common among mantids. Every volant male inthe Old World has a DK ear, with the exceptions ofMetallyticus, the Hoplocoryphinae, Majanga, and avery few other unusual cases. Most flying males inthe New World also hear well, except animals of thebasal groups and rare, odd cases such as Callimantis.The situation for females is more complex, but, basi-cally, all nonflying mantids of both sexes world-widehave reduced or absent hearing.
The contrast between mantis ear distributions inthe Old and New Worlds is strong and pervasive. Theonly unequivocal MSMT genus outside the NewWorld is Metallyticus. Insectivorous, echolocating batsare very common in tropics and subtropics throughoutthe world (Nowak, 1991), including the habitats of theexcellent flyers Mantoida, Chaeteesa, and the Thespi-dae, eliminating low predation pressure as a probableexplanation for the MSMT distribution. Despite theirbroad and patchy distribution, we found no cases ofMESO or HiC (except for the aberrant Stagmomantiscentralis) hearing in New World taxa.
CONSTANCY AND ANCIENT FUNCTIONS
Although the present study shows a range of morpholo-gical and physiological types associated with mantisaudition, the dominant underlying theme is constancy.
Once the DK ear appeared in the early Cretaceous,every genus in the lineage maintained it in funda-mentally unchanged form. The variations such as thesunken knobs and huge lateral bulges of the Amor-phoscelinae and Iridopteryginae, are alterations ofproportion, not structure. Grimaldi (2003) points tothe DK ear as a defining feature of modern mantids(Mantoidea). The situation in the Dytrisia (mostmoths and butterflies), a taxon very much larger thanthe Mantodea, and the other insect group for which
MANTIS EAR PHYLOGENY 559
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022

there is broad comparative data on ears (Scoble, 1995;Minet & Surlykke, 2003) and some phylogeneticinformation (Hasenfuss, 1997, 2000; Yack et al.,1999), provides both contrast and similarity. Auditorysystems have evolved independently many times inthe Dytrisia with different ancillary structures, dif-ferent numbers of sensory neurones, and at differentlocations on the body. In some families, a few generahave ears and others do not, the Pyraloidea have twodistinct ear types, and at least two superfamilies(Geometroidea and Noctuoidea) have a consistentbasic ear anatomy throughout.
The constancy in mantis audition extends to boththe physiological and behavioural realms. All of the91 hearing ‘species’ tested (males and females consid-ered separately and including DNK/DO animals withhigh-threshold hearing) had the same basic neuralresponse pattern to ultrasound: a neurone with ashort-latency, phasic-tonic firing pattern and a largeaxon plus a neurone with a short-latency, strictlytonic firing and a smaller axon. In M. religiosa andStagmomantis carolina, the phasic-tonic interneuronehas been identified with intracellular recording(501-T3; Yager & Hoy, 1989), and it is reasonable thatthe homologous interneurone accounts for the equiva-lent responses in other species. This is also true forthe smaller, tonic unit, which is also an identifiedinterneurone, 401-T3, in M. religiosa (D. D. Yager,unpublished observations). This uniformity has twoimportant implications. These two neurones and theirinvolvement with hearing must be as ancient as theear itself and just as stable. An interneurone with thesame anatomy as 501-T3 occurs in the cockroach P.americana, and appears to play a role in processingvibratory information (Pollack, Ritzmann & Watson,1994), which suggests that the transition in functionmay be relatively simple. The evolutionary stability ofmantis auditory interneurones is consistent with thehighly conserved components of the central nervoussystem in general, and of escape systems in particular(Dumont & Robertson, 1986; Edwards & Palka, 1991;Paul, 1991; Tierney, 1995). Second, the phasic-tonicfiring pattern with very high instantaneous rates inthe phasic portion must play a strongly adaptive rolelinked to behaviour. Ultrasound-sensitive auditoryinterneurones in several other insects fire with asimilar pattern (Boyan & Fullard, 1986; Nolan & Hoy,1987; Stumpner & Lakes-Harlan, 1996; Faure & Hoy,2000), so an initial phasic burst is likely to be a keyfeature, especially in predator avoidance (Nabatiyanet al., 2003; Fullard, Ratcliffe & Guignion, 2005). In anespecially well-studied case, Nolan & Hoy (1984) dem-onstrated that a firing rate of 220 spikes s-1 in a cricketinterneurone was both necessary and sufficient totrigger escape turning. Strong evidence supports thisin mantids (Triblehorn & Yager, 2002, 2005).
The same evolutionary arguments hold for thehighly conserved ultrasound-triggered behaviour inmantids. In the face of strong, constant pressure frompredators that use ultrasound, for example, a success-ful evasive behaviour would persist unchanged. Modi-fication would occur only if the most relevant predatorchanged, and Fullard et al. (2005) has provided anexample with moths on Tahiti, a bat-free environ-ment. This also appears to be the case with the HiCmantis genera that have a much extended frequencyrange and modified behaviour, probably in response toincreased vulnerability to bats whose calls are abovethe useful hearing range of most mantids (Fentonet al., 1998; Fullard, 1998; Schoeman & Jacobs, 2003).Hearing insects have a range of responses to attack-ing bats (Hoy, Nolen & Brodfuehrer, 1989; Miller &Surlykke, 2001): directional turns (moths, crickets,locusts), erratic flight (moths, possibly crickets),passive drops (green lacewings, tettigoniids), ultra-sonic clicking (arctiid moths, tiger beetles), and powerdives (mantids, some moths, possibly tiger beetles).As far as is known, the specific evasive behaviours ineach group are also conserved, although very littlecomparative and phylogenetic information is avail-able for most. A downside to the constancy of mantisauditory behaviour for the researcher is that itreveals nothing of the evolution of the behaviouritself. Some data support the possibility that theevasive behaviour is the same defensive display usedon the ground, adapted to a different context and witha different trigger stimulus (Yager et al., 1990; Cooket al., 2005; Yager, Ottinger & Benedick, 2006), anespecially important observation because mantishearing significantly predates echolocating bats.
WHEN AND WHY
The origin of the mantis DK ear in the early Creta-ceous is consistent with biogeographical patterns inthe evolution of the Mantodea (Svenson, 2007). Thevery broad distribution of hearing among mantidssuggests that it must have evolved before the sepa-ration of Gondwanaland continents. Only the forelegear of crickets, which appeared > 200 Mya (Otte,1992), is known to be older than the mantis ear.
Although cricket hearing probably first functionedin communication (Hoy & Robert, 1996), ultrasonichearing in that and several other insects probablyevolved as protection from predators, specificallyecholocating bats (Hoy, 1992; Connor, 1999). Theappearance of the DK ear approximately 120 Myaconsiderably predates the divergence of the modernfamilies of bats approximately 63 Mya (Eick, Jacobs& Matthee, 2005; Teeling et al., 2005). There are twopossible interpretations of the disparity. First, as incrickets, mantis hearing originally served another
560 D. D. YAGER and G. J. SVENSON
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022

function such as intraspecific communication or preydetection that was lost or reduced when the auditorysystem was co-opted for bat evasion. Second, mantisultrasonic hearing was originally an adaptation toavoid capture by predators other than bats. The ultra-sound may have been produced passively by predatormovement or actively as it is by a number of contem-porary animals ranging from frogs to insectivores toseveral types of birds (Sales & Pye, 1974; Narinset al., 2004; Pytte et al., 2004). Birds were present inthe Cretaceous, and some may have been crepuscularhunters that used ultrasound as do oilbirds and greyswiftlets today (Suthers & Hector, 1982, 1985).
The sensitive low frequency hearing of the MESOear implies an important undiscovered function ofaudition in mantids. The lack of directionality arguesagainst its use in hunting, although the detection ofincoming prey by their wing beat sounds is a possi-bility. Intraspecific communication is an appealingexplanation, but there are no observations of mantidsproducing sounds in nondefensive contexts. In short,this is one of many remaining puzzles about mantishearing.
ACKNOWLEDGEMENTS
Many individuals made significant contributions tothis project over the years and heartfelt thanks areextended to all of them. In particular, the help ofDan Otte and his staff at the Philadelphia Academy ofNatural Sciences and of Judith Marshall at theBritish Museum, Natural History was indispensable.Dr Ron Hoy inspired the first studies of mantishearing and very generously supported some of theearliest data collection. Thanks also go to DavidGrimaldi who provided access to mantis fossils at theAmerican Museum of Natural History. Biologicalillustrator John Norton put in extremely long hoursgrappling with the task of drawing tiny, complex,highly 3D structures. Several generations of techni-cians and animals care folk capably handled the innu-merable and crucial, day-to-day details of the project.Many of the mantids for the physiological and behav-ioural studies came through the generosity of Euro-pean insect hobbyists. We also thank a number ofcolleagues, including Michael Whiting, Kelly Miller,Stephen Cameron, Marcos Perez-Losada, and Kim-berly Baer, who provided analytical advice and assis-tance for the molecular portion of the study. We wouldalso like to recognize the additional support andassistance of Michael Whiting throughout this study.He was generous in providing some molecular dataprior to its publication elsewhere, and in providingweb site space to post this study’s data files, align-ments, and other materials. Supported by the Eppley
Foundation for Research, and NIH/NIDCD grant#DC01382 to D.D.Y.
NOTE ADDED IN PROOF
A recent phylogram for the Mantodea incorporatingtissue from a fresh specimen of Chaeteesa valida placesthe Chaeteesidae as the most basal group in thesuborder. (Svenson GJ & Whiting MF, unpubl. data).
REFERENCES
Beier M. 1968. Mantodea (Fangheuschrecken). Handbook derZoologie Band 4, Hälfte 2, Teil 2. Berlin: de Gruyter, 1–47.
Boyan GS, Fullard JH. 1986. Interneurones responding tosound in the tobacco budworm moth Heliothis virescens(Noctuidae): morphological and physiological characteris-tics. Journal of Comparative Physiology A 158: 391–404.
Caterino MS, Cho S, Sperling FAH. 2000. The currentstate of insect molecular systematics: a thriving Tower ofBabel. Annual Review of Entomology 45: 1–54.
Connor WE. 1999. ‘Un chant d’appel amoureux’: acousticcommunication in moths. Journal of Experimental Biology202: 1711–1723.
Cook AP, Triblehorn JD, Lorsong A, Lenon M, YagerDD. 2005. Praying mantis evasive response: descendingcontrol and context gating. Integrative and ComparativeBiology 44: 684.
Cumming GS. 1996. Mantis movements by night and theinteractions of sympatric bats and mantises. CanadianJournal of Zoology 74: 1771–1774.
Dumont JPC, Robertson RM. 1986. Neuronal circuits: anevolutionary perspective. Science 233: 849–853.
Edgecomb RS, Robert D, Read MP, Hoy RR. 1995. Thetympanal hearing organ of a fly: phylogenetic analysis of itsmorphological origin. Cell and Tissue Research 282: 251–268.
Edwards JS, Palka J. 1991. Insect neural evolution – afugue or an opera? Seminars in the Neurosciences 3: 391–398.
Ehrmann R, Roy R. 2002. Systematische Aufstellung derGattungen. In: Ehrmann R, ed. Mantodea. Gottesanbeterin-nen der Welt. Munich: Natur und Teil – Verlag, 374–378.
Eick GN, Jacobs DS, Matthee CA. 2005. A nuclear DNAphylogenetic perspective on the evolution of echolocationand historical biogeography of extant bats (Chiroptera).Molecular Biology and Evolution 22: 1869–1886.
Eriksson T. 2002. The r8s bootstrap kit (package distributedby the author). Stockholm: Bergius Foundation, RoyalSwedish Academy of Sciences.
Faure PA, Hoy RR. 2000. Neuroethology of the katydidT-cell II. Responses to acoustic playback of conspecific andpredatory signals. Journal of Experimental Biology 203:3243–3254.
Felsenstein J. 2005. PHYLIP (Phylogeny Inference Package),version 3.6. Distributed by the Author. Seattle, WA: Depart-ment of Genome Sciences, University of Washington.
MANTIS EAR PHYLOGENY 561
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022

Fenton MB, Portfors CV, Rautenbach IL, Waterman JM.1998. Compromises: sound frequencies used in echolocationby aerial-feeding bats. Canadian Journal of Zoology 76:1174–1182.
Fullard JH. 1998. The sensory coevolution of moths and bats.In: Hoy RR, Popper AN, Fay RR, eds. Comparative hearing:insects. New York, NY: Springer-Verlag, 279–326.
Fullard JH, Ratcliffe JM, Guignion C. 2005. Sensoryecology of predator–prey interactions: responses of the AN2interneuron in the field cricket, Teleogryllus oceanicus to theecholocation calls of sympatric bats. Journal of ComparativePhysiology A 191: 605–618.
Genecodes. 1991–2003. Sequencher. Ann Arbor, MI: Gen-codes Company.
Goloboff P. 1999. Analyzing large data sets in reasonabletimes: solutions for composite optima. Cladistics 15: 415–428.
Goloboff P, Farris SJ, Nixon KC. 2003. TNT: tree analysisusing new technology, Version 1.0. Tucumán, Argentina.Published by the author.
Gratschev VG, Zherikhin VV. 1993. New fossil mantids(Insecta, Mantida [sic]). Paleontology Journal 27: 148–165.
Grimaldi D. 1997. A fossil mantis (Insecta: Mantodea) inCretaceous amber of New Jersey, with comments on theearly history of the Dictyoptera. American Museum Novi-tates 3204: 1–11.
Grimaldi D. 2002. Fossiliferous Cretaceous amber fromBurma (Myanmar): its rediscovery, biotic diversity, andpaleontological significance. American Museum Novitates3361: 1–71.
Grimaldi D. 2003. A revision of Cretaceous mantises andtheir relationships, including new taxa (Insecta: Dicty-optera: Mantodea). American Museum Novitates 3412: 1–47.
Gwynne DT. 1995. Phylogeny of the Ensifera (Orthoptera): ahypothesis supporting multiple origins of acoustical signal-ing, complex spermatophores and maternal care in crickets,katydids, and weta. Journal of the Orthopterist’s Society 4:203–218.
Hasenfuss I. 1997. Precursor structures and evolution oftympanal organs in Lepidoptera (Insecta, Pterygota).Zoomorphology 117: 155–164.
Hasenfuss I. 2000. Evolutionary pathways of truncal tympa-nal organs in Lepidoptera (Insecta: Holometabola). Zoolo-gischer Anzeiger 239: 27–44.
Hoy RR. 1992. Evolution of ultrasound hearing in insects. In:Webster DB, Fay RR, Popper AP, eds. The evolutionarybiology of hearing. New York, NY: Springer-Verlag, 115–129.
Hoy RR, Nolen TG, Brodfuehrer PD. 1989. The neuroet-hology of acoustic startle and escape in flying insects.Journal of Experimental Biology 146: 287–306.
Hoy RR, Robert D. 1996. Tympanal hearing in insects.Annual Reviews of Entomology 41: 433–450.
Huelsenbeck JP, Imennov NS. 2002. Geographic origin ofhuman mitochondrial DNA: accommodating phylogeneticuncertainty and model comparison. Systematic Biology 51:155–165.
Huelsenbeck JP, Larget B, Miller RE, Ronquist F. 2002.
Potential applications and pitfalls of Bayesian inferencephylogeny. Systematic Biology 51: 673–688.
Hughes-Schrader S. 1950. The chromosomes of mantids(Orthoptera: Manteidae) in relation to taxonomy. Chromo-soma 4: 1–55.
Jobb G. 2006. TREEFINDER, May 2006. Munich, Germany.Distributed by the author. Available at http://www.treefinder.de
Katoh K, Kuma K, Toh H, Miyata T. 2005. MAFFT version5: improvement in accuracy of multiple sequence alignment.Nucleic Acids Research 33: 511–518.
Katoh K, Misawa K, Kuma K, Miyata T. 2002. MAFFT: anovel method for rapid multiple sequence alignment basedon fast Fourier transform. Nucleic Acids Research 30: 3059–3066.
Klass K-D. 1997. The external male genitalia and the phy-logeny of Blattaria and Mantodea. Bonner ZoologischeMonographien 42: 1–341.
Lakes-Harlan R, Stölting H, Stumpner A. 1999. Conver-gent evolution of insect hearing organs from a preadaptivestructure. Proceedings of the Royal Society of London SeriesB, Biological Sciences 266: 1161–1167.
Maddison DR, Maddison WP. 2003. MacClade, Version4.06. Sunderland, MA: Sinauer Associates.
Mason JB. 1968. The tympanal organ of Acridomorpha. Eos44: 267–355.
Meier T, Reichert H. 1990. Embryonic development andevolutionary origin of the orthopteran auditory organs.Journal of Neurobiology 21: 592–610.
Miller LA, Olesen J. 1979. Avoidance behavior in greenlacewings I. Behavior of free flying green lacewings tohunting bats and ultrasound. Journal of ComparativePhysiology 131: 113–120.
Miller LA, Surlykke A. 2001. How some insects detect andavoid being eaten by bats: tactics and countertactics of preyand predator. BioScience 51: 570–581.
Minet J, Surlykke A. 2003. Auditory and sound producingorgans. In: Kristensen NP, ed. Lepidoptera, moths andbutterflies. Berlin: Walter de Gruyter, 290–323.
Muma KE, Fullard JH. 2004. Persistence and regression ofhearing in the exclusively diurnal moths, Trichodezia albo-vittata (Geometridae) and Lycomorpha pholus (Arctiidae).Ecological Entomology 29: 718–726.
Nabatiyan A, Poulet JFA, de Polavieja GG, Hedwig B.2003. Temporal pattern recognition based on instantaneousspike rate coding in a simple auditory system. Journal ofNeurophysiology 90: 2484–2493.
Narins PM, Feng AS, Lin W, Schnitzler H-U, DenzingerA, Suthers RA, Xu C. 2004. Old world frog and birdvocalizations contain prominent ultrasonic harmonics.Journal of the Acoustical Society of America 115: 910–913.
Nel A, Roy R. 1996. Revision of the fossil ‘mantid’ and‘ephemerid’ species described by Piton from the Paleoceneof Menat (France) (Mantodea: Chaeteesidae, Mantidae;Ensifera: Tittigonioidea). European Journal of Entomology93: 223–234.
Nixon KC. 1999. The parsimony ratchet, a new method forrapid parsimony analysis. Cladistics 15: 407–414.
562 D. D. YAGER and G. J. SVENSON
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022

Nolan TG, Hoy RR. 1984. Initiation of behavior by singleneurons: the role of behavioral context. Science 226: 992–994.
Nolan TG, Hoy RR. 1987. Postsynaptic inhibition mediateshigh-frequency selectivity in the cricket Teleogryllus oceani-cus: implications for flight phonotaxis behavior. Journal ofNeuroscience 7: 2081–2096.
Nowak RM. 1991. Walker’s mammals of the world, Vol. 1, 5thedn. Baltimore, MD: Johns Hopkins University Press.
Otte D. 1990. The relation between hearing and flying incrickets. Entomology News 101: 29–34.
Otte D. 1992. Evolution of cricket songs. Journal of Ortho-ptera Research 1: 25–49.
Otte D, Spearman L. 2005. Mantida species file. Catalog ofthe mantids of the world. Philadelphia, PA: Association ofInsect Diversity.
Paul DH. 1991. Pedigrees of neurobehavioral circuits: tracingthe evolution of novel behaviors by comparing motor pat-terns, muscles, and neurons in members of related taxa.Brian, Behavior and Evolution 38: 226–239.
Pollack AJ, Ritzmann RE, Watson JT. 1994. Dual pathwaysfor tactile sensory information to thoracic interneurons in thecockroach. Journal of Neurobiology 26: 33–46.
Posada D, Crandall KA. 1998. Modeltest: testing the modelof DNA substitution. Bioinformatics 14: 817–818.
Prete FR, Wells H, Wells PH, Hurd LE. 1999. The prayingmantids. Baltimore, MD: Johns Hopkins Press.
Prier KR, Boyan GS. 2000. Synaptic input from serialchordotonal organs onto segmentally homologous interneu-rons in the grasshopper Schistocerca gregaria. Journal ofInsect Physiology 46: 297–312.
Pytte CL, Ficken MS, Moiseff A. 2004. Ultrasonic singingby the blue-throated hummingbird: a comparison betweenproduction and perception. Journal of Comparative Physi-ology A 190: 665–673.
Raff RA. 1996. The shape of life. Genes, development, and theevolution of animal form. Chicago, IL: University of ChicagoPress.
Riede K, Kämper G, Höfler I. 1990. Tympana, auditorythresholds and projection areas of tympanal nerves insinging and silent grasshoppers (Insecta, Acridoidea).Zoomorphology 109: 223–230.
Roeder KD. 1971. Neural factors and evitability in insectbehavior. Journal of Experimental Zoology 194: 75–88.
Roff DA, Tucker J, Stirling G, Fairbairn DJ. 1999. Theevolution of threshold traits: effects of selection on fecundityand correlated response in wing dimorphism in the sandcricket. Journal of Evolutionary Biology 12: 535–548.
Ronquist F, Huelsenbeck JP. 2003. MrBayes 3: Bayesianphylogenetic inference under mixed models. Bioinformatics19: 1572–1574.
Roy R. 1999. Morphology and taxonomy. In: Prete FR, WellsH, Wells PH, Hurd LE, eds. The praying mantids. Balti-more, MD: Johns Hopkins University Press, 19–40.
Sales GD, Pye JD. 1974. Ultrasonic communication byanimals. London: Chapman & Hall.
Sanderson MJ. 2004a. r8s; inferring absolute rates of evo-lution and divergence times in the absence of a molecularclock. Bioinformatics 19: 301–302.
Sanderson MJ. 2004b. r8s, user’s manual. version 1.70,December 2004. http://loco.biosci.arizona.edu/r85
Schoeman MC, Jacobs DS. 2003. Support for the allotonicfrequency hypothesis in an insectivorous bat community.Oecologia 134: 154–162.
Scoble MJ. 1995. The Lepidoptera. Form, function and diver-sity. Oxford: Oxford University Press.
Sharov AG. 1962. Redescription of Lithophotina floccosaCock. (Mantodea) with some notes on the manteod wingvenation. Psyche 69: 102–106.
van Staaden MJ, Rieser M, Ott SR, Pabst MA, Römer H.2003. Serial hearing organs in the atympanate grasshopperBullacris membracioides (Orthoptera, Pneumoridae).Journal of Comparative Neurology 465: 579–592.
van Staaden MJ, Römer H. 1998. Evolutionary transitionfrom stretch to hearing organs in ancient grasshoppers.Nature 394: 773–776.
Stumpner A, Lakes-Harlan R. 1996. Auditory interneuronsin a hearing fly (Therobia leonidei, Ormiini, Tachinidae,Diptera). Journal of Comparative Physiology A 178: 227–233.
Suthers RA, Hector DH. 1982. Mechanism for the produc-tion of echolocating clicks by the grey swiftlet, Collocaliaspodiopygia. Journal of Comparative Physiology A 148:457–470.
Suthers RA, Hector DH. 1985. The physiology of vocaliza-tion by the echolocating oilbird, Steatornis capirensis.Journal of Comparative Physiology A 156: 243–266.
Svenson GJ. 2007. The origins, evolution, and phylogeny ofthe praying mantises (Dictyoptera; Mantodea). PhD Thesis,Brigham Young University.
Svenson GJ, Whiting MF. 2004. Phylogeny of Mantodeabased on molecular data: evolution of a charismatic preda-tor. Systematic Entomology 29: 359–370.
Svenson GJ, Whiting MF. 2006. Phylogeography and con-vergent ecomorphic evolution of the praying mantids(Dictyoptera: Mantodea). Entomological Society of AmericaAbstracts 54: 239.
Swofford DL. 2002. PAUP*. Phylogenetic analysis using par-simony (*and other methods), version 4.0b10. Sunderlands,MA: Sinauer Associates.
Teeling EC, Scally M, Kao DJ, Romagnoll ML, SpringerMS, Stanhope MJ. 2005. Molecular evidence regarding theorigin of echolocation and flight in bats. Nature 403: 188–192.
Terry MD, Whiting M. 2005. Mantophasmatodea and phy-logeny of the lower neopterous insects. Cladistics 21: 240–257.
Tierney AJ. 1995. Evolutionary implications of neural cir-cuitry structure and function. Behavioral Processes 35: 173–182.
Triblehorn JD. 2003. Multisensory integration in theultrasound-triggered escape response of the praying mantis,Parasphendale agrionina. PhD Thesis, University of Mary-land, College Park.
Triblehorn JD, Yager DD. 2001. Broad vs. narrow auditorytuning and corresponding bat-evasive behaviors in prayingmantids. Journal of Zoology, London 254: 27–41.
Triblehorn JD, Yager DD. 2002. Implanted electrode
MANTIS EAR PHYLOGENY 563
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022

recordings from a praying mantis auditory interneuronduring flying bat attacks. Journal of Experimental Biology205: 307–320.
Triblehorn JD, Yager DD. 2005. Timing of praying mantisevasive responses during simulated bat attack sequences.When does the mantis dive? Journal of ExperimentalBiology 208: 1867–1876.
Vrsansky P. 2002. Origin and early evolution of mantises.AMBA Projekty 6: 1–16.
Vrsansky P, Vishniakova VN, Rasnitsyn AP. 2002. OrderBlattida LATREILLE, 1810. The cockroaches (= BlattodeaBrunner von Wattenvill, 1882). In: Rasnitsyn AP, QuickeDLJ, eds. History of insects. Dordrecht: Kluwer AcademicPublishers, 145–176.
White MJD. 1965. Sex chromosomes and meiotic mecha-nisms in some African and Australian mantids. Chromo-soma 16: 521–547.
Yack JE. 2004. The structure and function of auditory chor-dotonal organs in insects. Microscopy Research and Tech-nique 63: 315–337.
Yack JE, Fullard JH. 1990. The mechanoreceptive origin ofinsect tympanal organs: a comparative study of similarnerves in tympanate and atympanate moths. Journal ofComparative Neurology 300: 523–534.
Yack JE, Hoy RR. 2003. Hearing. In: Resh VH, Cardé RT,eds. Encyclopedia of insects. San Diego, CA: AcademicPress, 498–505.
Yack JE, Scudder GGE, Fullard JH. 1999. Evolution of themetathoracic tympanal ear and its mesothoracic homologuein the Macrolepidoptera (Insecta). Zoomorphology 119:93–103.
Yager DD. 1990. Sexual dimorphism of auditory function andstructure in praying mantises (Mantodea; Dictyoptera).Journal of Zoology, London 221: 517–537.
Yager DD. 1996a. Serially homologous ears perform
frequency range fractionation in the praying mantis, Creo-broter (Mantodea, Hymenopodidae). Journal of ComparativePhysiology A 178: 463–475.
Yager DD. 1996b. Nymphal development of the auditorysystem of the praying mantis Hierodula membranaceaBurmeister (Dictyoptera, Mantidae). Journal of Compara-tive Neurology 364: 199–210.
Yager DD. 1999a. Sensory processes. Hearing. In: Prete FR,Wells H, Wells PH, Hurd LE, eds. The praying mantids.Baltimore, MD: Johns Hopkins University Press, 93–113.
Yager DD. 1999b. Structure, development, and evolution ofinsect auditory systems. Microscopy Research and Tech-nique 47: 380–400.
Yager DD. 2005. Cockroach homologues of praying mantisperipheral auditory system components. Journal of Mor-phology 205: 120–139.
Yager DD, Hoy RR. 1987. The midline metathoracic ear ofthe praying mantis, Mantis religiosa. Cell and TissueResearch 250: 531–541.
Yager DD, Hoy RR. 1989. Audition in the praying mantis,Mantis religiosa L.: identification of an interneuron medi-ating ultrasonic hearing. Journal of Comparative PhysiologyA 165: 471–493.
Yager DD, May ML. 1990. Ultrasound-triggered, flight-gatedevasive maneuvers in the praying mantis, Parasphendaleagrionina (Gerst.). II: Tethered flight. Journal of Experi-mental Biology 152: 41–58.
Yager DD, May ML, Fenton MB. 1990. Ultrasound-triggered, flight-gated evasive maneuvers in the prayingmantis, Parasphendale agrionina (Gerst.). I: free-flight.Journal of Experimental Biology 152: 17–39.
Yager DD, Ottinger S, Benedick M. 2006. Evidence sup-porting the evolution of aerial bat-evasion maneuvers fromthe terrestrial deimatic display in praying mantids. Ento-mological Society of America Abstracts 54: 239.
564 D. D. YAGER and G. J. SVENSON
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022

AP
PE
ND
IX1
MANTIS EAR PHYLOGENY 565
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022
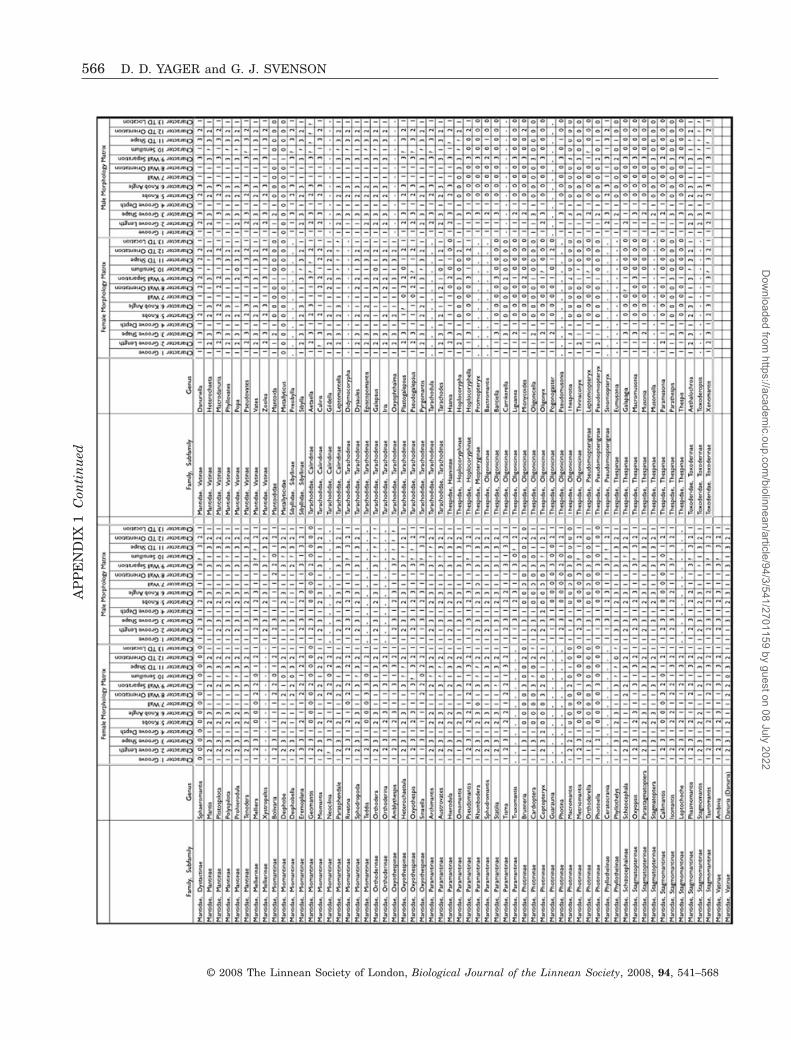
AP
PE
ND
IX1
Con
tin
ued
566 D. D. YAGER and G. J. SVENSON
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022

AP
PE
ND
IX2
MANTIS EAR PHYLOGENY 567
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022

AP
PE
ND
IX2
Con
tin
ued
568 D. D. YAGER and G. J. SVENSON
© 2008 The Linnean Society of London, Biological Journal of the Linnean Society, 2008, 94, 541–568
Dow
nloaded from https://academ
ic.oup.com/biolinnean/article/94/3/541/2701159 by guest on 08 July 2022




















