
E xtensive Ethnogenomic Diversity of Endothelial Nitric Oxide Synthase (eNOS) Polymorphisms
Upload
independentCategory
view
2download
0
JAP-00061-2003-R1
Gender, Exercise Training, and eNOS expression
in Porcine Skeletal Muscle Arteries.
by
M. Harold Laughlin1,2, Wade V. Welshons1, Michael Sturek3,
James W. E. Rush4, James R. Turk1, Julia A. Taylor1, Barbara M. Judy1,
Kyle K. Henderson1, and V. K. Ganjam1.
1Department of Biomedical Sciences,
2Dalton Cardiovascular Research Center, and
3Departments of Medical Pharmacology and Physiology and Internal Medicine,
University of Missouri
Columbia, MO 65211
and
4Department of Kinesiology, University of Waterloo,
Waterloo, ON Canada N2L 3G1
Running Head: Training and endothelial dependent responses.
Address all correspondence to:
M. H. Laughlin, Ph.D.
Professor of Physiology
E-102 Vet. Med. Bldg.
University of Missouri
Copyright (c) 2003 by the American Physiological Society.
Articles in PresS. J Appl Physiol (March 7, 2003). 10.1152/japplphysiol.00061.2003

JAP-00061-2003-R1
Columbia, MO 65211.

JAP-00061-2003-R1
1
Abstract
The purpose of the present study was to determine the effects of gender and exercise training
on endothelial nitric oxide synthase (eNOS) and superoxide dismutase protein (SOD) protein content
of porcine skeletal muscle arteries and to evaluate the role of 17ß-estradiol (E2) in these effects. We
tested the hypothesis that endothelial cells of female arteries express more eNOS and/or SOD than
male endothelial cells by measuring protein content with immunoblots and immunohistochemistry
in femoral and brachial arteries of male (%) and female (&) pigs. Second, we measured eNOS and
SOD content of these arteries from trained % and & pigs to test the hypothesis that exercise training
increases eNOS and/or SOD content of these arteries. We also measured estrogen receptor (ERα)
mRNA in aortic endothelial cells and ERα and ERβ protein in aortas of % and & pigs. Results
indicate that & arteries contain more eNOS than % arteries and that exercise training increases eNOS
content independent of gender. % and & pigs expressed similar levels of ERα mRNA and protein
and similar amounts ERβ protein in their arteries. E2 concentrations, as measured by
radioimmunoassay (RIA) were 180 ± 34 pg/ml in % sera, and approximately 5 pg/ml in & sera and
neither was changed by training. However, bioassay indicated that biologically active estrogen
equivalent to an E2 concentration of only 35 ± 5 pg/ml was present in % sera. E2 in & pigs, whether
measured by RIA or bioassay, was approximately 24 pg/ml at peak estrous, and approximately 2
pg/ml on day 5 diestrus. Free serum E2 was lower in % than in & pigs and exercise training
increased free E2 in %s and decreased it in &s. We conclude that: 1) Gender has significant influence
on eNOS and SOD content of porcine skeletal muscle arteries., 2) The effects of gender and
exercise training vary among arteries of different anatomic origin, 3) That % sera contains

JAP-00061-2003-R1
2
compounds that cause RIA to overestimate circulating estrogenic activity., and 4) Relative to the
human %, the % pig is not biologically estrogenized by high levels of E2 reported by RIA, while in
& pig E2 levels are lower than in the blood of human & s.
INDEX WORDS: skeletal muscle blood flow, vascular smooth muscle, nitric oxide, estradiol free
fraction, estrogen, RIA, biological activity.

JAP-00061-2003-R1
3
INTRODUCTION
Men have a higher incidence of atherosclerosis and coronary heart disease (CHD) than do
premenopausal women of similar age (40). After menopause the incidence of cardiovascular disease
increases in women so that there is no longer a gender difference (30, 43). Current evidence
suggests that female hormones mediate these beneficial effects but which hormones are most
important has not been established. These beneficial effects of the female gender are associated with
modification of reactivity of arteries due to differences in vascular smooth muscle, endothelial and
adventitial cells in the arteries (34). It has been proposed that the influence of estrogens on
endothelial cell phenotype and function plays an important role in the protective effects of female
gender on vascular disease (18, 40). Taddei et al. (53) proposed that protective female gender
effects may abate with menopause because endothelium dysfunction is produced by endogenous
estrogen deficiency (53). Consistent with the proposal of a role for estrogen in normal endothelial
phenotype are recent reports that estrogen increases endothelial cell nitric oxide synthase (eNOS)
gene expression (5, 24, 32), basal release of nitric oxide (NO) by vascular cells (18, 19), and
endothelium-dependent vasodilation (5, 18). Also, the hypothesis that plasma estrogen can influence
endothelial function is supported by recent reports of improved vascular function (39) and
acetylcholine-induced, endothelium-mediated vasodilation in male to female transsexuals treated
with estrogen therapy (38).
In view of these results, we were intrigued by an observation that brachial arteries of female
pigs were less responsive to endothelium-dependent relaxation effects of bradykinin (BK) and
acetylcholine (ACH) than brachial arteries from male pigs (29). These responses were indeed

JAP-00061-2003-R1
4
endothelium-dependent since removal of the endothelium abolished relaxation responses of the
brachial arteries to both BK and ACH. Another interesting observation in these experiments was
that inhibition of NOS by treatment with arginine analogs (L-NAME) revealed that NOS contributes
fractionally more to endothelium-dependent relaxation in brachial arteries from female pigs than in
male brachial arteries. Finally, we reported that male pigs had greater plasma concentrations of 17ß-
estradiol (E2) than female pigs (29). These concentrations of E2 suggest the possibility that brachial
arteries of male pigs exhibit greater endothelium-dependent relaxation than female brachial arteries
because males have higher plasma concentrations of E2. However, two other sets of data argue
against this postulate. First, male pigs were phenotypic boars and exhibited no evidence of being
estrogenized such as mammary development. Second, endothelium-dependent relaxations are
greater in coronary arteries (2) and femoral arteries (29) of female pigs than in males. The purpose
of the study reported herein was to further investigate determinants of gender differences in
endothelium-dependent relaxation of porcine femoral and brachial arteries and interactions of gender
with exercise training effects on expression of eNOS in these arteries. Because superoxide
dismutase can increase bio-availability of NO and because previous studies indicate that exercise
training modulates SOD protein levels in aorta and coronary arterioles of pigs (46, 47), we also
examined effects of gender and exercise on SOD levels of femoral and brachial arteries.
We conducted five experiments. First, we measured eNOS and SOD protein content of
femoral and brachial arteries isolated from male and female pigs using immunoblots and
immunohistochemistry. Our hypothesis was that endothelial cells of female arteries express more
eNOS and/or SOD than male endothelial cells. Second, we measured estrogen receptor (ER) mRNA

JAP-00061-2003-R1
5
in aortic endothelial cells of male and female pigs and ER protein in aortas of male and female pigs
to test the hypothesis that male arteries are less responsive to circulating E2 because they express
fewer or no ER receptors. Third, we measured eNOS and SOD protein content in femoral and
brachial arteries of trained male and female pigs to determine the role of altered eNOS and/or SOD
expression in the training-induced changes. Fourth, we tested the hypothesis that only a small
fraction of E2 detected by RIA in the blood of male pigs is free, unbound to serum binding proteins.
Finally, because we found that free fractions of E2 did not explain how the male pigs could show
high levels of E2 in their blood without overt estrogenization of phenotype, we used a sensitive
bioassay to measure biological estrogenic activity in male and female blood to test the hypothesis
that E2 content, as measured by RIA overestimates estrogenic activity in male pig blood. Our results
appeared to confirm this hypothesis and suggested that not all of the E2 measured in male pig serum
by RIA is biologically active.
Results indicate that conduit arteries of female pigs have a greater eNOS content than do the
same arteries of male pigs and that exercise training increased eNOS protein content in both male
and female arteries. ER receptor content of aortic tissues appear to be similar in samples from males
and females. Interestingly, we observed that total estrogenic activity present in male pig serum was
lower, and sometimes much lower, than the E2 content indicated by RIA, while E2 free fractions in
male and female blood were similar in magnitude though statistically significantly different.

JAP-00061-2003-R1
6
METHODS
Experimental Animals
Experiments were completed on adult male (n = 73) and female (n = 72) Yucatan miniature
swine (Charles River) weighing 25-40 kg obtained from the breeder. Animals were procured in lots
of 8 or 16 animals and familiarized with treadmill exercise over a 1-2 wk period of time. Treadmill
performance tests were administered to each animal to evaluate exercise tolerance. Each lot of pigs
was then randomly divided into two groups: exercise-trained (EX) or sedentary (SED). EX pigs
underwent a progressive treadmill training program, lasting 13 - 21 weeks that in our hands
produces adaptations in miniature swine that are classically associated with the exercise trained state
in all mammals (26-29). SED pigs were restricted to their pens (6 x 12 feet) for 13 - 21 wks. Pigs
were procured at 9-10 months of age and were 13-14 months of age at the termination of
experiments. All experimental procedures involving animals were approved by the University of
Missouri Animal Care and Use Committee in accordance with the “Principles for the utilization and
care of vertebrate animals used in testing, research and training”.
Training Procedures
Training Program: EX pigs trained on the treadmill 5 days/wk and were given positive
reinforcement for exercise by feeding them after each training bout. The speed and duration of daily
running were progressively increased over the first 8-12 wks at a rate dependent upon the tolerance
of each pig. A typical training session during the 12th wk consisting of the following 85 min
workout: 1) 5 min warm up run at 2.5 mph, 2) 15 min sprint at speeds of 5 to 8 mph, 3) 60 min
endurance run at 4 - 5 mph and 4) 5 min warm-down run at 2 mph. Ranges of running speed are

JAP-00061-2003-R1
7
presented because the EX program was customized to each pig's exercise ability. Treadmill
performance tests were administered to the SED and EX pigs before initiation of training and at the
completion of SED or EX periods.
Treadmill Performance Test: The performance test consisted of 4 stages of exercise (26-29).
During stage 1 pigs ran at 3.1 mph, 0 % grade for 5 min. Pigs ran for 10 min at stage 2 (speed = 3.1
mph and grade = 10 %) and then for 10 min at stage 3 (speed = 4.3 mph and grade = 10 %). Finally,
pigs ran at stage 4 (speed = 6 mph, grade = 10 %) until the pigs could not maintain treadmill speed.
Heart rates were recorded throughout the treadmill performance test along with total duration of
exercise.
Efficacy of training: The effectiveness of the training program was determined by
comparing the exercise tolerance (as reflected in the treadmill performance test), heart weight/body
weights, and skeletal muscle oxidative capacity of trained and control groups. At the time of
sacrifice, samples were taken from and stored at -70 oC until processed. Citrate synthase activity
of the lateral and long heads of triceps brachii and the deltoid muscles as described by Srere (52).
Estrous Cycle
Estrous cycles were measured by heat checking 10 female pigs (gilts) on a daily basis with
standard procedures using a male pig to determine when gilts were receptive. Gilts were exposed
individually to a Yucatan boar daily and considered in heat when they responded to the presence of
the boar (4). After establishing estrous cycle duration in each of the 10 gilts they were instrumented
with vascular access ports so that we could obtain blood samples at 8 defined times during the estrus

JAP-00061-2003-R1
8
cycle (1, 41).
Implanting Vascular Access Port (VAP): Pigs were sedated with an intramuscular injection
of Telazol® (5.0 mg/kg), Xylazine® (2.2 mg/kg) and atropine (0.05 mg/kg), intubated, and placed
under isoflurane anesthesia. Using aseptic techniques the right jugular vein was catheterized and
the tip of the catheter positioned cranial to the right atrium with the aid of a fluoroscope (1, 41).
Catheters were routed subcutaneously to the left side of the neck where a accessed with a vertical
incision in the jugular furrow of the neck, where a subcutaneous pocket was made for the vascular
access port. After implantation access ports and catheters were filled with a 50 % dextrose/saline
solution (2.8 ml) containing 200 units/ml of heparin and 1mg/ml Vancomycin to maintain patency
and prevent infection (Vancomycin). Penicillin G Procaine (600K Units) was given intramuscular
once per day for three days post surgery.
Previous heat checking demonstrated an average 20 day estrus cycle for these pigs, therefore
blood samples were collected on days: 17, 18, 19, 20, 0, 1, 2, 3, 5 (day 20/day 0 = day of peak serum
estradiol levels) to measure changing levels of estrogen. Ten milliliters of blood were collected,
centrifuged and stored (-80 Co) prior to analysis.
Blood Sampling Regimen: In the male and female pigs not instrumented with vascular
access ports, jugular vein blood samples were obtained via venipuncture and collected in vacutainers
containing EDTA at the conclusion of training or cage confinement. Samples were collected after
a 12 hour fast. Plasma was separated by centrifugation (Beckman TJ-6R centrifuge, Palo Alto, CA)
at 4 degrees C for 15 minutes at 3750 rev/min. Plasma was stored at –70 degrees C until assay.
Tissue sampling: After completion of exercise training or sedentary confinement and

JAP-00061-2003-R1
9
approximately 24 hrs after the last exercise bout, pigs were sedated with ketamine (Fort Dodge)(35
mg/kg) and Rompun (Bayer)(2.25 mg/kg), anesthetized with Thiopental (Abbott Labs)( 10 mg/kg),
and euthanized by removal of the heart in full compliance with American Veterinary Medical
Association Panel on Euthanasia guidelines. Segments of femoral and brachial artery of
approximately 2-3 mm outer diameter (OD) and 3 to 5 mm in length were carefully removed and
trimmed of fat and connective tissue, frozen in liquid N2 and stored at -80°C until processed. Artery
samples were taken from the same locations of male and female SED and EX pigs. In addition, a 15
cm segment of thoracic aorta was isolated starting approximately 3 cm distal to the end of the arch
of the aorta, and placed in Krebs bicarbonate buffer (4°C), previously aerated with 95%O2-5%CO2
gas mixture. Aortic endothelial cell lysates were collected as previously described (47).
RT-PCR using Aortic Endothelial Cell samples: First-strand cDNA synthesis was initiated
using 1 µg of total aortic endothelial cell RNA and an oligo(dT)12-18 primer (SuperScript
preamplification system, GIBCO-BRL) in a total volume of 20 µl. A 5 µl aliquot of cDNA product
was used in PCR in a total reaction volume of 50 µl containing (in mM) 50 KCL, 20 Tris!HCl (pH
8.4), 4 MgCl2, 0.2 dNTP, and 0.2 forward and reverse primers as well as 2.5 U Taq DNA
polymerase (Promega). Estrogen receptor alpha (ERα) primers were based on a reported sequence
for porcine ERα (6), and were designed to amplify a 325-bp product. The primers were: sense 5’-
CATGTTGCTGGCTACATCATCTCG-3’; anti-sense 5’-CACCACGTTCTTGCACTTCATG-3’.
PCR was initiated with a 5-min denaturation step (94°C) and a 5-min annealing step (63°C)
followed by cycles of 72°C (elongation, 2 min), 94°C (1 min), and 63°C (1 min). The final step was
a 10 min 72°C elongation. The PCR products were separated by electrophoresis (1.5% agarose

JAP-00061-2003-R1
10
gels), and visualized under UV light via ethidium bromide staining. Semi-quantitative analysis was
performed by scanning densitometry of photographic images (Polaroid) of ethidium bromide-stained
gels. Preliminary experiments confirmed that under these conditions, 30 cycles of PCR produced
results that were in the linear range for product accumulation. The relationship between volume of
the PCR preparation loaded on the agarose gel and the densitometric signal was determined, and an
optimal loading volume of 3 µl was found for the conditions outlined above. In subsequent analyses
3 µl of PCR preparation was loaded for all samples. To validate the densitometric method, we also
performed semi-quantitative analyses by radiolabelling experiments using α-(32P)dCTP in PCR
(data not shown) as we have detailed previously (46). The two methods yielded similar qualitative
and quantitative results for group differences. Only the densitometric analysis data are illustrated
in this manuscript.
Immunoblots: Aortic endothelial cell samples were prepared for immunoblots as previously
described (47). Artery (brachial and femoral) samples were homogenized in protein solubilization
buffer consisting of 50 mM Tris-HCl, pH 7.4, 6 M Urea and 2% SDS using a ground glass
homogenizer. After a 2 hour incubation at 45°C and centrifugation (10 000g, 1 min) protein
concentration of the supernatants was determined using the bicinchoninic acid assay (Pierce).
Samples were processed for immunoblot analysis as described previously (47). For gender, artery
location, and exercise comparisons, equal numbers of samples from the treatment groups were
loaded in the same gel for comparison.
Immunohistochemistry: Samples of aorta were dissected and immersed in 10% formalin for
a minimum of 24 hours. Rings 3 mm in length were processed routinely to paraffin embedment.

JAP-00061-2003-R1
11
Five micron sections were cut with an automated microtome (Microm), floated onto positively
charged slides (Fischer), and deparaffinized. The slides were steamed in citrate buffer at pH 6.0
(Dako target retrieval solution S1699) for 30 minutes to achieve antigen retrieval and then cooled
for 30 minutes. The slides were stained manually with sequential Tris buffer and water wash steps
performed after each protocol step. Sections were incubated with avidin biotin two-step blocking
solution (Vector SP-2001) to inhibit background staining and in 3% hydrogen peroxide to inhibit
endogenous peroxidase. Nonserum protein block (Dako X909) was applied to inhibit nonspecific
protein binding. The primary antibodies utilized were mouse monoclonal antibody for eNOS (BD
Transduction Laboratories) (21) and rabbit polyclonal antibodies for SOD1, SOD2 (Stressgen
Biotechnology Corporation) (46), ERα and ERβ (Santa Cruz Biotechnology). All primary
antibodies were diluted 1:800 and incubated with the tissue sections overnight at 4< C. After
appropriate washing steps were completed the sections were incubated with biotinylated anti-mouse
or rabbit link secondary antibody in PBS containing 15 mM sodium azide and peroxidase-labeled
streptavidin (Dako LSAB+ kit, peroxidase, K0690). Diaminobenzidine (Dako) applied for 5
minutes allowed visualization of primary antibody staining. Sections were counterstained with
Mayer’s hematoxylin stain for 1 minute, dehydrated, and coverslipped. For negative controls,
histological sections were prepared as described above, but incubation in primary antibody was
excluded from the protocol. Sections were examined and photographed using an Olympus BX40
photomicroscope.
Measurement of E2 and biologically active E2.
Serum E2 has traditionally been measured by radioimmunoassays (RIA), but because of

JAP-00061-2003-R1
12
reports of extremely high E2 concentrations in male pig serum, we chose to use a variety of methods
to determine not only total E2 present in serum but also the free, biologically active fraction of E2,
and the effective concentration that was biologically active in bioassay.
Radioimmunoassay: We measured E2 in serum and plasma by RIA according to the method
of Kirby et al. (23), validated for porcine serum by Laughlin et al. (29). Some samples were brought
to pH 9 by adding NaHCO3 (60 mM) to inhibit solvolysis prior to ether extraction. Aliquots of 50-
200 µl of serum or plasma were extracted twice with 3 ml methyl tertiary butyl ether. Radiolabeled
estradiol and the estradiol antibody were obtained from ICN Pharmaceuticals (Costa Mesa, CA).
The intra-assay and inter-assay coefficients of variation were 4.9% and 13.4%, respectively.
Sensitivity of the assay was 0.5 pg/ml, and recoveries from a spectrum of differing amounts of
estradiol-17β were approximately 88%. Parallelism between the standard curve and different
volumes of a porcine serum quality control standard was not different (slopes = -1.99 0.07 for
standard curve and –2.22 0.34 for porcine serum). Measurements of E2 in a test set of comparisons
from 8 female pigs, from which both plasma and serum were obtained at the same time, indicated
statistically similar values for E2. E2 in serum was 4.4 + 1.2 pg/ml and in plasma was 5.0 + 1.0
pg/ml.
LH-20 Chromatography-RIA: To ensure that high E2 concentrations measured by RIA were
not due to contaminating estrone, which can cross-react in E2 RIAs, we first separated extracted
serum or plasma on LH-20 columns (ISOLAB, Akron, OH), as described by Nagel et al. (36), using
85% benzene:15% methanol as the elution solvent. Fractions collected from the column were dried
before being assayed by RIA.

JAP-00061-2003-R1
13
Ultrafiltration Dialysis: The free fraction of E2 was measured in serum or plasma after
addition of tritiated E2 by centrifugal ultrafiltration dialysis, according to the method of Montano
et al. (35). The percent free hormone was calculated from relative amounts of labeled hormone
present in the ultrafiltrate and in the total, non-dialyzed samples. Measurements of the free fraction
of E2 in a test set of comparisons from 18 animals from which both plasma and serum were obtained
at the same time gave identical results. The free fraction in serum was 3.59 % + 0.10 % and in
plasma was 3.64% + 0.17 % (mean + SEM, n = 18).
Bioassay of Estrogenic Activity: Serum was extracted with methyl-tertiary butyl ether as
for RIA, and an aliquot of the same extract was also tested for estrogenic activity by bioassay using
estrogen-responsive MCF-7 human breast cancer cells, with modifications to Grady et al. (16).
Plating the cells, preparing dose-response series by serial dilution, feeding of the cells, and parts of
the final DNA assay were all performed by robotics with a Tomtec Quadra 96. Cells were plated
on day 0 in estrogen-free medium in 96-well plates. On day 1 the medium was replaced with fresh
estrogen-free medium. Serum extracts were dried and resuspended in estrogen-free tissue culture
medium, and then serially diluted to yield a series of serum equivalent concentrations. Cells were
fed with this medium (180 µl /well) for 4 days with daily medium changes, days 3 to 6. On day 7
cell growth was determined by DNA content/well, using the diphenylamine assay of Natarajan et
al. (37), and absorbance was read on a Bio-Tek PowerWave-x 96 well plate reader. Serum
estrogenic activity was determined against a standard dose-response curve of purified 17ß-estradiol
(Sigma). HPLC/Bioassay separation of estrogenic activities was performed as described previously
(16).

JAP-00061-2003-R1
14
Data Analysis: All values are means + SE. Between group differences were assessed using
repeated measures ANOVA or Student’s T tests where appropriate. Differences with p < 0.05 were
considered significant. Where between-group variances were not homogeneous, the data were log-
transformed before ANOVA and post-hoc tests: in each case variance was homogeneous after the
log transform.

JAP-00061-2003-R1
15
RESULTS
GENDER EFFECTS ON ENDOTHELIAL PROTEINS IN SEDENTARY ANIMALS.
Experimental Design. To determine whether endothelial cells of female arteries express
more eNOS and/or SOD than male endothelial cells, we measured eNOS and SOD protein content
of femoral and brachial arteries isolated from male and female pigs. Then, because previous results
indicated that exercise training resulted in increased endothelium-dependent relaxation in female,
but not male, brachial arteries (29), we also measured eNOS and SOD protein content in femoral
and brachial arteries of trained male and female pigs to determine the role of altered eNOS and SOD
expression in the training-induced changes. In both of these experiments, protein content was
assessed with immunoblots and immunohistochemistry. Results indicated that endothelial cells in
arteries from female pigs appeared to have greater content of eNOS and SOD protein relative to
arteries of male pigs just as do arteries of other female mammals described above. Of interest,
anatomical location also appears to influence expression of these proteins.
Gender and anatomical location effects on arterial eNOS, SOD-1, SOD-2 proteins. A
single 130 kDa band was evident in immunoblots for eNOS. As illustrated in Figure 1 the eNOS
content of brachial and femoral arteries from female pigs was 1.9- and 2.4-times greater than in the
respective arteries from male pigs (P = 0.006, and P = 0.004, respectively). Aortic endothelial cells
from female pigs also had significantly higher eNOS protein levels compared to males (5.9-times,
P = 0.01; data not shown). Greater eNOS content of endothelial cells of female arteries was also
apparent in immunohistochemistry results (Figure 2).
A single 17 kDa band was evident in immunoblots for SOD-1. SOD-1 content was greater

JAP-00061-2003-R1
16
in female femoral arteries (2.41-times male; P = 0.005) but male and female brachial arteries had
similar amounts of SOD-1 (P = 0.434; Fig. 1). A single 22 kDa band was evident in immunoblots
for SOD-2. Female pigs had significantly greater SOD-2 content than males in both brachial (1.48-
times, P = 0.013) and femoral (1.57-times, P = 0.008) arteries (Fig. 1). Figure 3 presents
immunohistochemical results demonstrating that the endothelium and smooth muscle exhibited
immunoreactivity for SOD-1 in the arteries of both males and females. Immunohistochemical
staining for SOD-2 revealed punctuate staining (consistent with SOD-2 being most concentrated in
mitochondria) in the endothelium and smooth muscle in arteries of both males and female pigs (data
not shown).
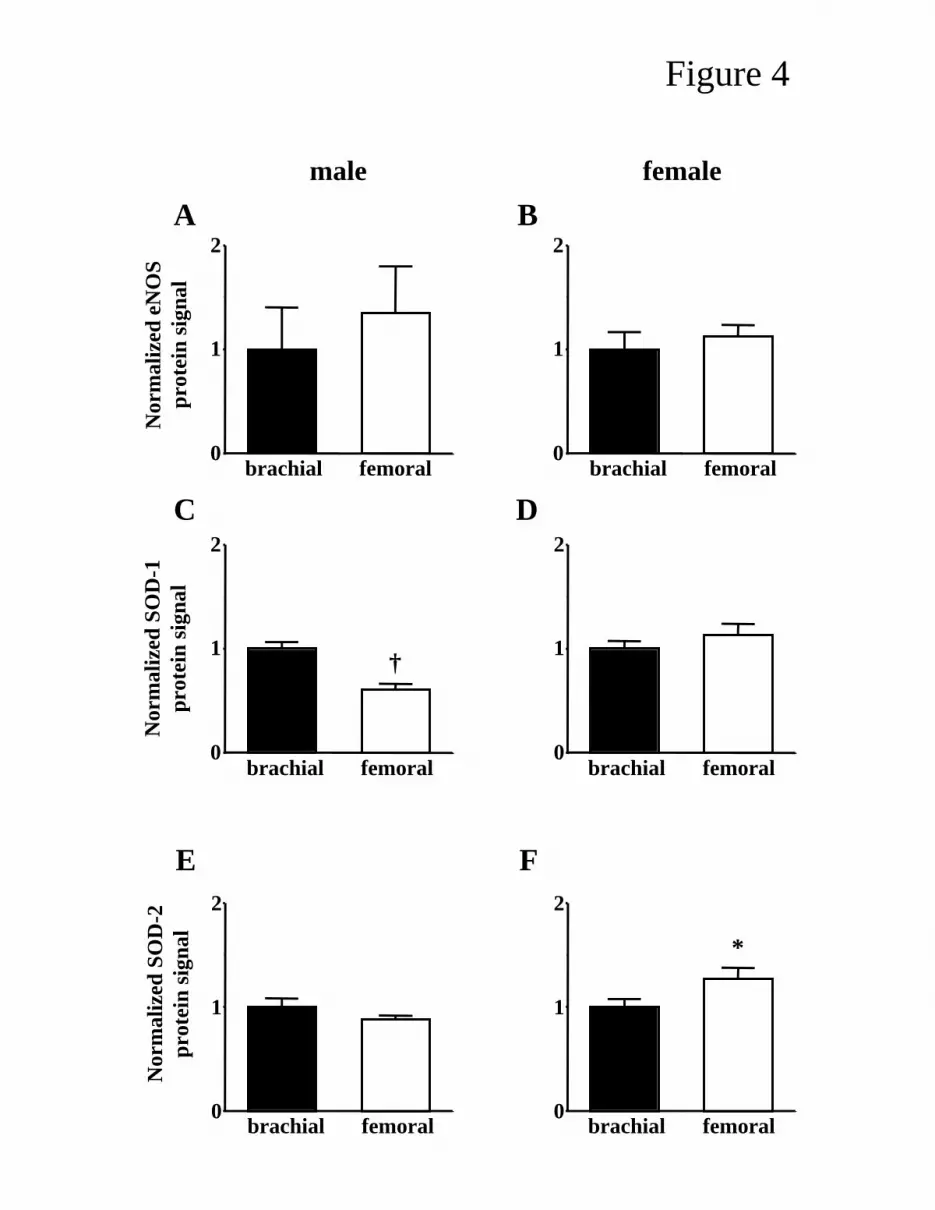
Figure 4 illustrates the relative levels of eNOS, SOD-1 and SOD-2 proteins in brachial versus
femoral arteries within a given gender. There are only two significant differences in protein content
between brachial and femoral artery: SOD-1 was significantly lower in femoral compared to brachial
arteries in male pigs (0.61-times brachial levels, P = 0.001); and SOD-2 was significantly higher in
femoral compared to brachial arteries in female pigs (1.26-times brachial levels, P = 0.048).
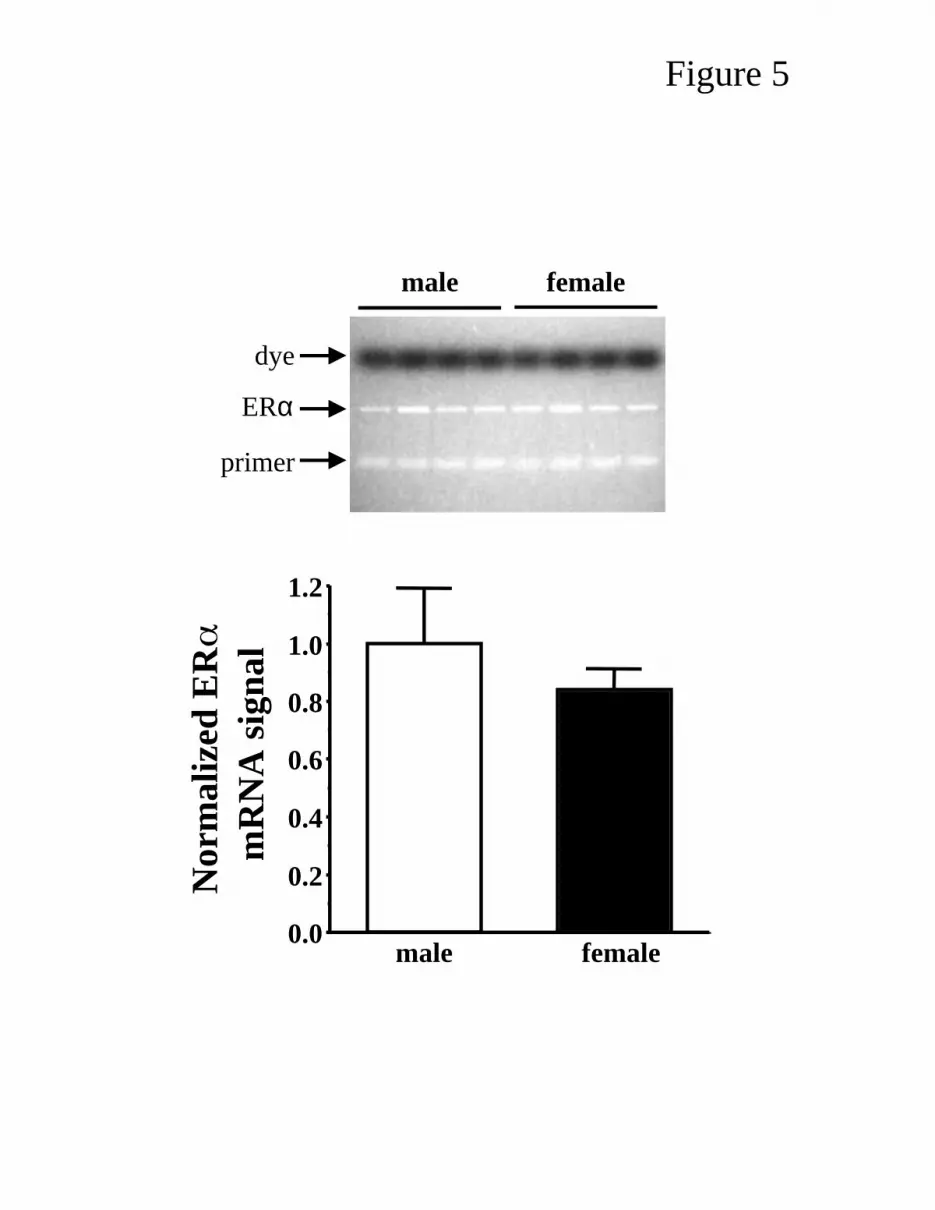
Gender effects on ERα mRNA levels in Aortic Endothelial Cells. Figure 5 illustrates the
data from RT-PCR experiments designed to evaluate relative levels of ERα mRNA in aortic
endothelial cells from male and female pigs. A single PCR product of 325 bp was evident. This is
consistent with the predicted product size based on the primers designed. Identification of this PCR
product was confirmed as ERα by direct sequencing after band excision from an agarose gel. The
gel-purified band shared 100% homology with the pig ERα gene (6) over 300 sequenced bases.
Negative control experiments leaving either reverse transcriptase enzyme or cDNA out of the

JAP-00061-2003-R1
17
reactions yielded no detectable PCR products (data not shown). There was no significant effect of
gender on ERα mRNA levels (female 0.84-times male levels, P = 0.189) under conditions optimized
for product formation and densitometric analysis (Fig. 5).
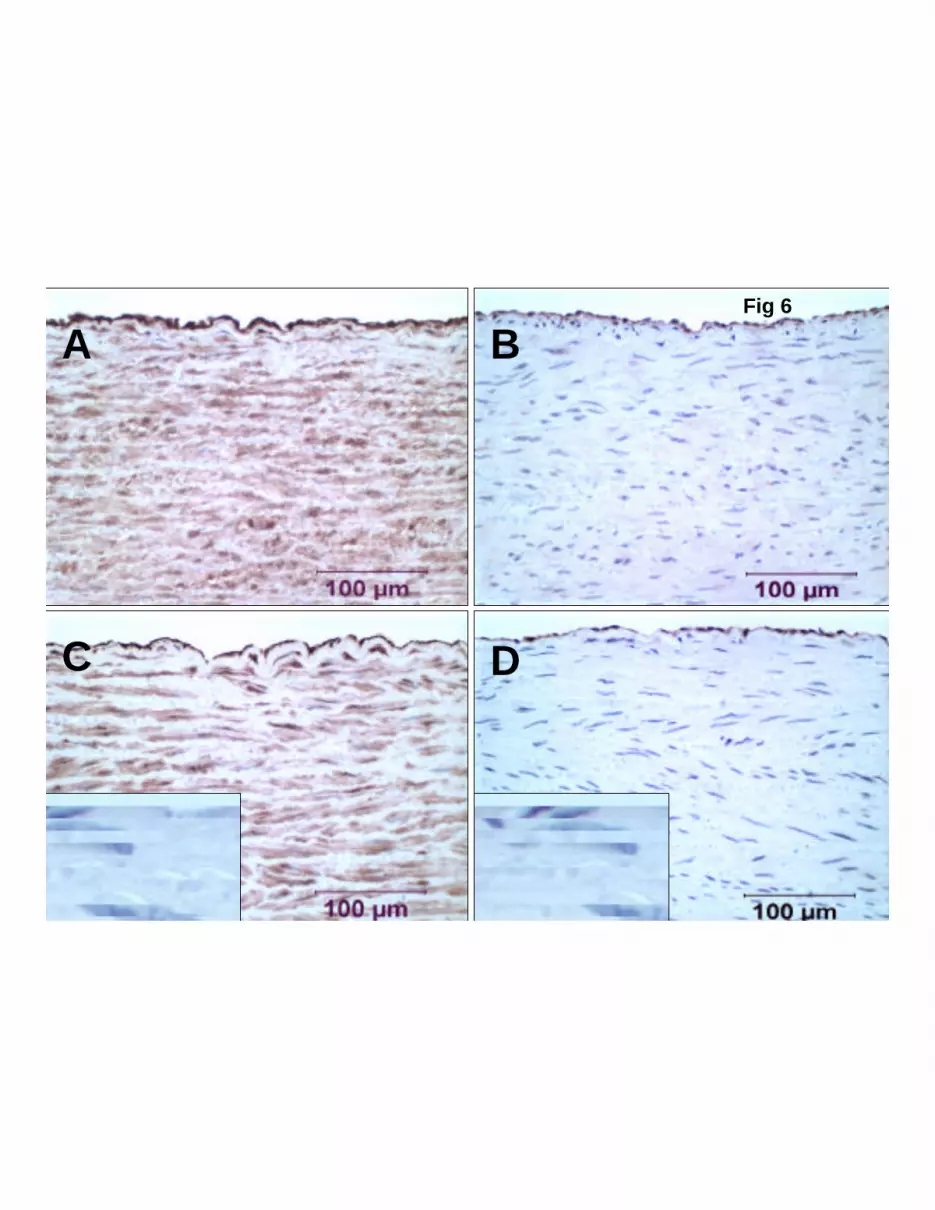
Gender effects on ER protein levels in aorta. Representative panels of
immunohistochemistry for ERα and ERβ protein of abdominal aorta are presented in figure 6. These
results show similar staining (immunoreactivity) in both endothelial and smooth muscle cells for
ERα protein (Fig 6 A & C) and for ERβ protein in endothelial cells (Fig 6 B & D) in aorta from
both females and males.
EXERCISE TRAINING EFFECTS.
Exercise training produced the expected adaptations in exercise endurance, heart weight and
skeletal muscle oxidative capacity in pigs of both genders. Thus in both male and female pigs,
average heart weight and heart weight-to-body weight ratio was greater in EX than in SED pigs
resulting from greater heart weights in EX animals (Table 1). The heart weight body weight ratio
of SED male pigs was greater than that for SED female pigs. Treadmill performance tests revealed
that endurance times (total run time) were significantly longer in EX pigs (Table 1). Citrate
synthase activity of the long and lateral heads of the triceps brachii muscle and the deltoid muscle
of EX pigs was 20 to 40 % greater than SED values in male and female pigs confirming the shift in
skeletal muscle oxidative capacity that characterizes effective exercise training. Finally, as shown
in the last two columns of Table 1, exercise training did not alter E2 levels in the blood of male or
female pigs.
Figure 7 illustrates the effects of gender and exercise training on eNOS, SOD-1 and SOD-2

JAP-00061-2003-R1
18
protein levels in both brachial and femoral arteries. EX increased eNOS protein levels in both the
brachial and femoral arteries of male (1.80- and 2.08-times SED males, P = 0.021 and P = 0.030,
respectively) and female (1.67- and 1.84-times SED females, P = 0.013 and P = 0.001, respectively)
pigs. EX did not significantly affect SOD-1 protein levels in brachial or femoral arteries of either
males or females (Fig. 7). Levels of SOD-2 protein were not significantly affected by EX in either
artery type in males, nor in brachial arteries of female pigs. However, femoral arteries of EX female
pigs had significantly higher levels of SOD-2 protein than their SED counterparts (EX 1.39-times
SED levels, P = 0.0002; Fig. 7).
CIRCULATING ESTRADIOL AND ESTROGENIC ACTIVITY IN MALE AND FEMALE PIGS.
Experimental Rationale. Results of the experiments above indicated that endothelial cells
in arteries from female pigs appeared to have greater content of eNOS and SOD protein relative to
arteries of male pigs just as do arteries of other female mammals described above. These results
raised the question: why does endothelial phenotype of male pigs not appear to reflect the high
levels of E2 in their serum? To test the hypothesis that male arteries do not reflect the high levels
of circulating E2 because they express fewer or no estrogen receptors (ER) we measured ER mRNA
in aortic endothelial cells and ER protein in aortas of male and female pigs. We reasoned that the
level of ER expression in the male must be one tenth or less that of female arteries to explain our
results. However, our results (above) revealed indistinguishable levels of ER" mRNA (RT-PCR)
and levels of both ERα and ERß protein by immunocytochemistry in arteries of male and female
pigs.
We then tested the hypothesis that only a small fraction of E2 detected by RIA in the blood

JAP-00061-2003-R1
19
of male pigs is free (unbound to serum binding proteins). In previous studies, E2 content of serum
of male and female pigs was measured with radioimmunoassay (RIA), which measures total E2 in
the serum (including free E2 and E2 bound to glycoproteins and serum albumin in the blood) (2,
29, 34). It is widely accepted that the biologically active form of E2 is the free, non-protein-bound,
fraction (54). If the free fraction for E2 in male pig serum were much lower than the free fraction
in female serum, then our results would be explained. Free fraction of E2, determined by centrifugal
ultrafiltration dialysis, indicated that the free fraction of E2 in males was only slightly lower than in
females, 3.5% versus 3.7%, respectively. Finally, we used a sensitive bioassay to test the hypothesis
that total estrogenic activity present in male pig serum was lower than the E2 content indicated by
RIA. In a subset of animals we measured estrogenic activity and E2 content in the same blood
samples of male and female pigs. E2 content was measured with standard RIA and with RIA after
running serum on LH-20 columns to evaluate any influence of circulating estrone in our samples that
could have been released by solvolysis.
Biologically active estrogenic activity in the blood was determined with a bioassay of
estrogenic activity in a subset of male and female sera. Bioassay results indicated that total
estrogenic activity in the blood of our male pigs (35 pg E2/ml serum) was similar to human male
concentrations of E2 reported in the literature ( 10-50 pg/ml)(54) in stark contrast to published
values of 100's to 1500 pg E2/ml as detected by RIA in male pig sera. Finally, we also measured
serum E2 concentrations (both total by RIA and biologically active by bioassay) throughout the
estrus cycle in 10 female pigs to establish the peak concentration of E2 in Yucatan females just prior
to ovulation for comparison to the values of E2 in males and human females.

JAP-00061-2003-R1
20
Free fraction of E2 in serum: Determinations of % free estradiol in pig blood revealed that
there were significant differences (p = 0.021) between free estradiol levels in males and females
(Table 2). However, the free fraction of E2 in males would have had to be one tenth or less of the
level in females to explain the fact that the males were not apparently estrogenized by the high total
E2 levels in their serum measured with RIA. Therefore, the differences in total E2 do not appear to
be counteracted by a much lower free fraction in males. Interestingly, exercise produced significant
increases in free fraction in males but significant decreases in females (Table 2).
Biologically active estrogen by bioassay in male and female blood: In blood samples from
a subset of pigs used in the exercise experiments described above, total estrogenic activity was
measured by bioassay in the same extracts used for RIA, to compare biologically active estrogen to
the RIA value for E2 measured in the serum. We used HPLC/bioassay to fractionate the serum and
measured estrogenic activity in each fraction with a tissue culture bioassay. Figure 8 presents results
for two male pigs that did not exhibit significant levels of estrone. As shown in Figure 8 (pig 1284)
the level of estrogenic activity, corresponding to where E2 ran in the HPLC, was only 42 pg E2
equivalents/ml (7.5 min sample) whereas RIA indicated that E2 was 100 pg/ml. Also, in a sample
chosen for its low measured E2 to provide a low background, almost no estrogenic activity of any
other kind was observed in the serum (Figure 8 pig 1343, bottom), indicating that most or all of the
biologically significant estrogenic activity, at least in these two serum samples, migrated with E2.
Analysis of group data reveals that in female pigs, estrogen determination as E2 by RIA and
by RIA after LH-20 were indistinguishable from estrogenic activity measured by bioactivity (Table
3). In contrast, in males, all groups were statistically different, with estrogenic activity by bioassay

JAP-00061-2003-R1
21
less than half of the level of E2 determined by RIA and the value by RIA after LH-20
chromatography was intermediate (Table 3). The highest E2 we observed in a male serum sample,
as measured by RIA was 645 pg/ml. In this sample, the total estrogenic activity was equivalent to
only 66 pg E2/ml (data not shown). So consistently in these samples, the biologically active
estrogenic activity in male blood was lower and sometimes much lower than the E2 level indicated
by RIA. In contrast to this pattern, in the unstaged females, biologically active estrogen content was
identical to the E2 measured by RIA (Table 3).
Estradiol in female blood across full estrous cycles: The changes in serum estradiol
observed in the 10 female pigs are presented as a function of days relative to the day of peak
estradiol levels in Figure 9. These results reveal that the peak value for these cycling female pigs
was approximately 24 pg/ml, while diestrous values were 1-5 pg/ml as determined by RIA. Table
4 presents results demonstrating that E2 in the serum of these 10 pigs was similar at peak and during
diestrous when measured by RIA and by bioassay of estrogen equivalents in the circulation.

JAP-00061-2003-R1
22
DISCUSSION
The purpose of this study was to test the hypothesis that female gender and exercise training
increase eNOS and SOD-1 protein content of porcine skeletal muscle arteries and to evaluate the
role of serum17ß-estradiol (E2) in these effects. This hypothesis was stimulated by previous
observations that gender and exercise training have important effects on endothelium-dependent
relaxation in these arteries. Results provide six important findings. First, femoral and brachial
arteries as well as aortic endothelial cells isolated from female pigs have significantly greater eNOS
content than arteries from male pigs, consistent with gender related effects in arteries of other
mammals (5, 18, 19, 24, 32). Second, although female femoral arteries contained greater SOD-1
content than the femoral artery of male pigs, there was no gender difference between SOD-1 content
in brachial arteries. Third, SOD-2 content was greater in brachial and femoral arteries of female
pigs. Fourth, biologically active estrogen concentration in male pigs is similar to E2 values reported
for human males while female pigs exhibit estradiol concentrations during diestrous (2 pg/ml) and
estrus (24 pg/ml) that are less than those of women (22, 54). Fifth, there was no effect of gender on
ERα mRNA levels in aortic endothelial cells or on ERα or ERβ protein in smooth muscle or
endothelial cells in aortas of male and female pigs. Sixth, although gender has important influences
on endothelium-dependent dilator responses and eNOS protein content of femoral and brachial
arteries, gender did not seem to interact with the effects of exercise training, as training increased
eNOS content in both male and female arteries. As discussed below, our results combined with
current literature indicate that mechanisms of gender differences in arteries perfusing skeletal muscle
and of exercise training-induced adaptations in these arteries are not uniform across agonists,

JAP-00061-2003-R1
23
species, or anatomic origin of the artery.
Effects of Gender on expression of eNOS, SOD-1, and SOD-1 in Skeletal Muscle Arteries.
Barber and Miller reported that endothelium-dependent relaxations of coronary arteries to
bradykinin and UK-14304(α2-adrenergic analogue) were greater and/or shifted leftward in arterial
rings from female pigs compared to males (2). Similarly we found that femoral arteries of female
pigs exhibited greater endothelium-mediated relaxation (bradykinin- and acetylcholine-induced)
than did femoral arteries from male pigs (29). Thus, there is substantial evidence that conduit
arteries of female pigs exhibit greater endothelium-dependent responses than do males. However,
in contrast to these observations, we found that male brachial arteries exhibited greater bradykinin-
induced relaxation and greater acetylcholine-induced relaxation than did brachial arteries from
female pigs (29). Blockade of the cyclo-oxygenase (COX) pathway and NOS pathways revealed
that COX seems to be relatively un-important in porcine brachial arteries and the NOS pathway
contributes more to endothelium-dependent relaxation in female brachial arteries than in males (29).
Because present results (Figs. 1 & 2) indicate that female femoral arteries contain more than twice
as much eNOS protein as male femoral arteries, it is reasonable to propose that greater release of
NO contributes to the greater relaxation seen in female arteries (29). On the other hand, the greater
endothelium-dependent relaxation responses in male brachial arteries can not be explained by
differences in eNOS content since female brachial arteries also had higher eNOS content than male
(Fig. 1). These observations suggest that gender has different effects on vasomotor reactivity of
porcine femoral and brachial arteries.
We are intrigued by the fact that the effects of gender on vasomotor reactivity (29) and on

JAP-00061-2003-R1
24
eNOS and SOD expression (figs 1 and 2) of femoral and brachial arteries do not appear to fully
correspond. Ours is not the only observation of such differing effects of gender among peripheral
arteries. For example, Vagnoni et al. reported that estradiol stimulated an increased expression of
eNOS protein in the sheep uterine artery but not in systemic arteries (55). Thus, our results provide
further evidence supporting the conclusion of Barber and Miller that the mechanisms of gender
differences in endothelial function differ by agonist, species, and anatomic origin of the artery (2).
Role of Estrogen in the Porcine Gender Differences.
Barber and Miller reported that the gender differences in porcine coronary endothelium-
dependent relaxation may not be related to estrogen levels because male pigs had higher plasma
concentrations of estrogen than female pigs, perhaps due to metabolism of testosterone by aromatase
in the adipose tissue (2). It has been know for some time that boars appear to have high
concentrations of estrogens in their blood. Indeed, domestic boars as well as stallions appear to be
unique among mammals in having high blood and urinary concentration of estrogens (7-9, 51, 57).
Paradoxically, these levels of estrogens in the boars are many fold higher compared to either
preovulatory females of the same species (20), or the males of other species (44, 45). The interstitial
compartment of the testis is the site of expression of steroidogenic enzymes in the boar (11, 48) and
therefore the major cellular site of both androgen and estrogen biosynthesis. The dynamic nature
of the secretory patterns of free androgens (10), estrogens (14), and conjugated steroids (49) from
the boar testis, especially the predominance of ∆5-C19-steroid production (3) and other conjugated
estrogens (56) has been well documented. Our results (Table 1) initially seemed to confirm these
observations since RIA data indicated plasma concentrations of 17ß-estradiol were greater in male

JAP-00061-2003-R1
25
pigs than in female pigs and these levels of plasma 17ß-estradiol are in the range of values reported
in the literature (2, 44, 45, 60).
Nevertheless as the experiments described herein proceeded we used several assays of E2 to
allow evaluation of the possibility that E2-RIA, as used by us and others, provides high values of
circulating estrogens ( > 1000 pg/ml) in male pigs because of a lack of specificity of the estrogen
antibody used in our E2-RIA. For example, in male pig blood the antibody may react with estrone,
or ether extraction of samples may cause solvolysis of estrogen conjugates, or the antibody may
cross-react with a closely-related ∆5-C19-steroid in the blood. We obtained similar values for E2 with
RIA, LH-20/RIA, and bioassay in female samples (Table 3). In contrast, our results indicate that
the levels of biologically active estrogen (estradiol and other estrogens) in male blood were much
lower than indicated by RIA measures of E2 (Table 3). This activity, expressed as estradiol
equivalents, was similar to the total amount of E2 (10 - 50 pg/ml) reported to circulate in human
males (54) and in the range of the peak levels (estrus) of both E2 and total estrogenic activity
measured in female blood (Table 4). These results indicate that one reason male pigs show no
evidence of being estrogenized, in spite of the apparently elevated levels of E2, is that boars do not
have excessive amounts of biologically active E2 in their blood. Thus, present results indicate that
male pigs appear to have E2 levels equal to or greater than female pigs and that endothelial cells
from male and female pigs express similar amounts of ERα mRNA and protein. These results
suggest that male pigs are likely as sensitive to E2 as are female pigs and that other (non-estrogen)
sex hormones and/or other gender-related processes play a role in establishing the greater eNOS
content in arteries of female pigs.

JAP-00061-2003-R1
26
Our male pig serum E2 results are even more interesting in light of the fact that estradiol as
well as testosterone is required for full reproductive development in the male pig, while only
estradiol is required in the female (15, 31). Whether or not human males also require estrogen for
development of a normal phenotype is not known. However, it is intriguing that development of the
male phenotype in pigs requires both estradiol and testosterone and the biologically active levels of
estrogen are similar in males and females.
It is also interesting that the paradox presented by the male blood E2 content which was high
when measured by RIA but consistently lower when measured by bioassay of actual estrogenic
activity, was not seen in the females. In the female pig the two measures, RIA and bioassay,
corresponded closely (Tables 3 & 4). Thus, it appears that most or all of the estrogenic activity
present in female serum was accounted for by the circulating E2 as measured by RIA. Dietary
estrogens did not appear to contribute to circulation of biologically significant levels of
phytoestrogens in the male or female pigs even though the feeds used in these studies were soy
based and contained detectable estrogenic activity (data not shown). Clearly, our results indicate
that in male pig blood, RIA-measured E2 values did not reflect estrogenic activity whereas, in
female pig blood, there was little contribution to estrogenic activity by any other circulating
estrogens (including any potential dietary estrogens).
There have been previous observations where biologically active estrogen levels are less than
estimates of total estrogen in the blood of mammals. For example, high levels of total E2 measured
by RIA in the rodent fetus are counterbalanced by very high levels of the estradiol-binding protein
in alphafetofetal serum. This leads to a free (unbound) bioavailable level of E2 that is biologically

JAP-00061-2003-R1
27
low, in the range of the male or diestrous female (35, 58). To determine whether differences in
binding protein in the blood contributed to our findings, we examined the free fraction of E2 in our
male and female blood samples. Results reveal that the free fraction of E2 in male pigs was very
similar to that of the female pigs (Table 2).
There are at least two potential explanations of the observation that biologically active
estrogen in male pig blood, as measured by a standard in vitro bioassay, was lower than E2 measured
by RIA, and the apparent absence of physiological estrogenic effects in male pigs. First, it is
possible that a molecule is present in the male pig blood that cross-reacts in the RIA for estradiol
but which exhibits little estrogenic activity. Secondly, it is possible that male pig serum contains
an inhibitor of estrogenic activity that reduces the effective estrogenic activity of the E2 present in
the serum, perhaps acting as a natural antiestrogen. Further work is required to determine an
explanation for why biologically active estrogen in male pig blood, as measured by a standard in
vitro bioassay, is lower than E2 measured by RIA, and the apparent absence of physiological
estrogenic effects in male pigs.
Effects of Exercise Training
Exercise training has been reported to result in enhanced endothelium-dependent relaxation
in rat aorta (12, 13), in rat skeletal muscle arteries (25), in brachial arteries of pigs (33), in peripheral
arteries of humans (17), in dog aorta and coronary arteries (50, 59), and porcine coronary arteries
(28). Present results (Figure 7) indicate that exercise training increased eNOS expression by similar
amounts in both male and female femoral and brachial arteries. The results provide no evidence for
an interaction of gender with the effects of exercise training on eNOS expression. Exercise training

JAP-00061-2003-R1
28
did not alter content of SOD-1 protein in femoral or brachial arteries in either gender.
The fact that exercise training increased eNOS content of femoral and brachial arteries in
both male and female pigs is interesting since previous results indicated that exercise training did
not alter net endothelium-dependent relaxation of male femoral or brachial arteries or of female
femoral arteries but increased endothelium-dependent relaxation of female brachial arteries (29).
One purpose of this study was to determine the role of changes in eNOS expression in these events.
It is not clear at this time why exercise training increases eNOS expression in both femoral and
brachial arteries of female and male pigs but endothelium-dependent relaxation is only enhanced
by exercise training in the brachial artery of female pigs.
Exercise training did not alter estradiol levels in either male or female pigs. Training did
produce slight changes in the binding of E2 in serum, as it increased free fraction in males but
lowered it in females. Since the source of E2 binding proteins is believed to be the liver, these
results suggest that training may have gender specific effects on production of serum proteins by the
liver.
CONCLUSION
The results of this study indicate that conduit arteries of female pigs have greater eNOS
content than do the same arteries of male pigs. Although male pigs have high concentrations of RIA
measured E2 (estrogen like molecules) in their blood, the levels of biologically active estrogen are
similar to those in male humans. These results combined with the observation that there is no
gender effect on ERα mRNA or ERα or ERβ protein levels suggest that eNOS expression in
endothelial cells of arteries may be modulated by other female hormones instead of estrogen.

JAP-00061-2003-R1
29
These data stimulate us to propose that, based on circulating estrogenic activity, the male pig
is not hyperestrogenized relative to the human male. Further, our results indicate that female pigs
may be hypoestrogenized relative to the human since E2 levels at peak estrus are substantially lower
than values of human females at peak estrus. However, it is important to emphasize that these levels
of estradiol or bioassayable estrogenic activity in serum of male pigs are not sufficiently high to
interfere with spermatogenesis and male reproduction, and that the levels of estradiol and estrogenic
activity we measured in the female are not so low as to fail to generate the cyclic changes to the
uterus and to sustain pregnancy, lactation and female reproduction. Therefore both genders in the
pig may represent appropriate models of cardiovascular function and gender differences relative to
the human.

JAP-00061-2003-R1
30
ACKNOWLEDGMENTS
The authors wish to thank Pam Thorne, Denise Holliman, Tammy Strawn, and Lisa Code
for technical contributions to this work. This work was supported by NIH grants HL-36088 and HL-
52490 (MHL), RR-13223 (MS) and Heart and Stroke Foundation of Ontario Grant NA-4604
(JWER), and the University of Missouri Food for the 21st Century VMFC-0018 (WVW).

JAP-00061-2003-R1
31
Table 1. Efficacy of Exercise Training in Female and Male Pigs
BW HW HW/BW Endurance Citrate Synthase E2 -RIA
(kg) (g) g/kg (minutes) Activity (µmol.min-1.gwet wt-1) (pg E2/ml)
Long Lat Delt
Female
SED 36 ± 1 161 ± 3 4.5 ± 0.1 23 ± 1 14 ± 1 12 ± 1 17 ± 1 5.7 + 1.0 (8)
EX 36 ± 1 *191 ± 5 *5.4 ± 0.1 *32 ± 1 *18 ± 1 *15 ± 1 *21 ± 1 4.4 + 1.1 (6)
MALE
SED 36 ± 2 &183 ± 6 &5.2 ± 0.1 23 ± 1 13 ± 1 13 ± 1 16 ± 1 180 + 34 (19)
EX 37 ± 1 *208 ± 7 *5.7 ± 0.1 *31 ± 1 *16 ± 1 *19 ± 2 *24 ± 1 168 + 28 (21)
Means ± SEs are presented. BW = body weight. HW = heart weight. Endurance = duration of total exercise time on the treadmill during
the performance test. Long = long head of triceps brachii muscle. Lat = lateral head of triceps brachii. Delt = deltoid muscle. SED =
sedentary. EX = exercise trained. N = 55 SED and EX female and 30 SED and EX male pigs for the BW, HW, Endurance and Citrate
synthase data. Numbers in () present ns for the E2 data. *indicates that the EX value is significantly different from SED (p<0.05).

JAP-00061-2003-R1
32
&indicates that the male value is significantly different from the female value (p<0.05).

JAP-00061-2003-R1
33
TABLE 2
Free Fraction of Estradiol in Serum of Sedentary (SED) and Exercise trained (EX) Pigs.
Males Females
SED 3.48 + 0.059 % (16) 3.66 + 0.071 % (11)
EX 3.69* ± 0.046 % (10) 3.39* ± 0.068 % (8)
Means + SEM are presented. Numbers in () represent number of animals. * = EX values
significantly different from SED value (p < 0.05).

JAP-00061-2003-R1
34
TABLE 3
E2 measured by RIA (pg E2/ml), by RIA after LH-20 chromatography (pg E2/ml) to separate E2
from estrone, and by bioassay of total biologically active estrogen present in serum (pg equivalents
of E2 /ml) in boars and in mixed diestrous females.
FEMALE, (Mixed Diestrus)
RIA LH-20/RIA Bioassay
Mean 3.93ns 4.11ns 3.80ns
SEM ± 0.54 ± 0.57 ± 0.715
n 22 20 22
MALE
RIA LH-20/RIA Bioassay
Mean 83.2a 57.8b 35.0c
SEM ± 9.9 13.3 ± 4.95
n 28 15 26
nsFemale values not statistically different
a,b,cMale values with different letter superscripts differ statistically from each other, P < 0.05.

JAP-00061-2003-R1
35
TABLE 4
E2 measured by RIA (pg E2 /ml) and by bioassay of total biologically active estrogen present in
serum (pg equivalents of E2/ml) in staged females.
FEMALE (Peak Estrus)
RIA Bioassay
Mean 23.9 23.2ns
SEM ± 1.27 ± 1.42
n 10 10
FEMALE (Day 5 Diestrus)
RIA Bioassay
Mean 1.6 2.3ns
SEM ± 0.67 ± 0.28
n 10 10
nsFemale RIA values not statistically different from bioassay values

JAP-00061-2003-R1
36
FIGURE LEGENDS
Figure 1. Immunoblot analysis of gender effects on eNOS, SOD-1 and SOD-2 protein levels in
brachial and femoral arteries of sedentary pigs. Brachial artery data is illustrated in panels A, C,
and E; femoral artery data in panels B, D, and F; eNOS data in panels A and B; SOD-1 data in
panels C and D; and SOD-2 in panels E and F. At the top of each panel, a representative
immunoblot is included representing 4 different samples (animals) from each indicated group.
At the bottom of each panel are the summary data from n = 6 animals per group, each analyzed
in triplicate immunoblots. Data are expressed as means ± SE, normalized within each artery type
to set mean densitometric data for the male group to unity. *, P < 0.05; †, P < 0.01, male vs
female within an artery type.
Figure 2. Representative panels of immunohistochemistry performed for eNOS in brachial and
femoral arteries. A: male brachial; B: female brachial; C: male femoral; D: female femoral.
Staining for eNOS is restricted to the endothelium. Staining is more intense in females (B & D)
than males (A & C). Bar = 100 micron.
Figure 3. Representative panels of immunohistochemistry performed for SOD-1 in brachial and
femoral arteries. A: male brachial; B: female brachial; C: male femoral; D: female femoral.
Staining for SOD-1 occurs in both endothelium and smooth muscle cells. Staining in the
brachial artery is similar in males and females (A & B) and less intense in the femoral artery of

JAP-00061-2003-R1
37
males than females C & D). Bar = 100 micron.
Figure 4. Immunoblot analysis of anatomical location effects on eNOS, SOD-1 and SOD-2
protein levels in arteries from male and female sedentary pigs. Male data is illustrated in panels
A, C, and E; female data in panels B, D, and F; eNOS data in panels A and B; SOD-1 data in
panels C and D; and SOD-2 in panels E and F. In each panel the summary data from n = 6
animals per group, each analyzed in triplicate immunoblots are illustrated. Data are expressed as
means ± SE, normalized within each gender to set mean densitometric data for the brachial
group to unity. *, P < 0.05; †, P < 0.01, brachial vs femoral artery within a gender.
Figure 5. RT-PCR analysis of gender effect on ERα mRNA levels in aortic endothelial cells of
sedentary pigs. Top, photograph of representative agarose gel of ERα RT-PCR products derived
from male and female aortic endothelial cell RNA. Four individual samples (animals) per group
are illustrated. Bottom, summary data from 5 male and 6 female animals each analyzed in
duplicate. Data are expressed as means ± SE normalized to set mean densitometric data for the
male group to unity.
Figure 6. Representative panels of immunohistochemistry performed on abdominal aorta. A:
estrogen receptor-a (ERα ) in female; B: ERβ in female; C: ERα in male, inset: preimmune
serum control; D: ERβ in male, inset: preimmune serum control. There is similar staining in
endothelial and smooth muscle cells for ERα in aorta from females and males (A&C). There is

JAP-00061-2003-R1
38
similar staining in endothelial cells for ERβ in aorta from females and males (B&D).
Figure 7. Immunoblot analysis of exercise effects on eNOS, SOD-1 and SOD-2 protein levels in
brachial (left column) and femoral (right column) arteries of male and female pigs. Brachial
artery data is illustrated in panels A, C, and E; femoral artery data in panels B, D, and F; eNOS
data in panels A and B; SOD-1 data in panels C and D; and SOD-2 in panels E and F. The
summary data from n = 6 animals per group, each analyzed in triplicate immunoblots is
illustrated in each panel. Data are expressed as means ± SE, normalized within each artery type
to set mean densitometric data for the male SED group to unity. *, P < 0.05; †, P < 0.01, SED vs
EX within a given gender and artery type.
Figure 8. HPLC/Bioassay separation of estrogenic activities in serum from two boars. Serum
proteins in 300 µl serum were precipitated with 600 µl methanol, and the supernatant was
separated by HPLC in methanol:water 65:35. Fractions (0.275 ml) were collected every 30
seconds and each was assayed for estrogenic activity by bioassay as described, at 7.5% serum
equivalent per ml in the growth medium. Peaks of estrogenic activity at 1.5 and 2.5 minutes in
the data for pig # 1284, represent conjugates in and near the solvent front; MCF-7 cells can
deconjugate some steroid sulfates (42) to release E1 or E2 from molecules that in the animal are
inactive. Results for two animals are represented. Pig 1284 (top, E2 by RIA = 100 pg/ml;
chosen to determine if very high levels of E1 were present that could cross react with the E2 RIA.
Importantly, high E1 was NOT present in this pig serum) and Pig 1343 (bottom, E2 by RIA = 5

JAP-00061-2003-R1
39
pg/ml; chosen to determine if estrogenic activities due to estrogens other than E2 were present;
biologically significant levels of estrogens other than E2 were NOT present). Dark bars represent
bioassay of HPLC fractions. Controls: Light bars represent plate controls for proliferation assay:
Ctrl = control (untreated) samples, AE = treated with antiestrogen (raloxifene, 1 x 10-7 M), and
samples labeled E11 and E10 are treated with estradiol at 1 x 10-11 M and 1 x 10-10 M
respectively. In the serum from Pig 1284, a peak of estrogenic activity was detected at 7 minutes
in the fraction where estradiol runs, equivalent to only 42 pg of E2/ml serum. Note that no other
peaks of activity comparable to E2 were detected.
Figure 9. Average serum estradiol levels for 10 female pigs across one hear cycle. Data for 10
pigs were averaged and plotted relative to the day of peak estradiol concentration as day 0.
Values are mean + SE. The average time of estrus relative to the day of peak estradiol
concentrations is also illustrated. E2 was measured by RIA as described.

JAP-00061-2003-R1
40

JAP-00061-2003-R1
41
REFERENCES
1. Bailie, M., S. Wixson, and M. Landi. Vascular-access-port implantation for serial blood
sampling in conscious swine. Lab. Anim. Sci. 36: 431-433, 1986.
2. Barber, D. A., and V. M. Miller. Gender differences in endothelium-dependent relaxations do
not involve NO in porcine coronary arteries. Am J Physiol 273: H2325-32, 1997.
3. Baulieu, E. E., I. Fabre-Jung, and L. G. Huis in't Veld. Dehydroepinadrosterone sulfate: a
secretory product of the boar testis. Endocrinology 81: 34-8., 1967.
4. Bearden, H. J., and J. W. Fuquay. Reproductive Management. In: Applied Animal
Reproduction, edited by J. W. Fuquay. Upper Saddle River: Prentice-Hall, 1997, p. 23-86.
5. Bell, D. R., H. J. Rensberger, D. R. Koritnik, and A. Koshy. Estrogen pretreatment directly
potentiates endothelium-dependent vasorelaxation of porcine coronary arteries. Am J Physiol
268: H377-83, 1995.
6. Bokencamp, D., P. W. Jungblunt, and H. H. Thole. The C-terminal half of the porcine
estradiol receptor contains no post-translational modification: determination of the primary
structure. Mol. Cell. Endocrinol. 104: 163-172, 1994.
7. Booth, W. D. In-vitro metabolism of unconjugated androgens, oestrogens and the sulphate
conjugates of androgens and oestrone by accessory sex organs of the mature domestic boar. J
Endocrinol 96: 457-64., 1983.
8. Claus, R., and B. Hoffmann. Oestrogens, compared to other steroids of testicular origin, in
blood plasma of boars. Acta Endocrinol (Copenh) 94: 404-11., 1980.

JAP-00061-2003-R1
42
9. Claus, R., D. Schopper, and H. G. Wagner. Seasonal effect on steroids in blood plasma and
seminal plasma of boars. J Steroid Biochem 19: 725-9., 1983.
10. Colenbrander, B., F. H. de Jong, and C. J. Wensing. Changes in serum testosterone
concentrations in the male pig during development. J Reprod Fertil 53: 377-80., 1978.
11. Conley, A. J., C. J. Corbin, M. M. Hinshelwood, Z. Liu, E. R. Simpson, J. J. Ford, and N.
Harada. Functional aromatase expression in porcine adrenal gland and testis. Biol Reprod 54:
497-505., 1996.
12. Delp, M. D., and M. H. Laughlin. Time course of enhanced endothelium-mediated dilation in
aorta of trained rats. Med Sci Sports Exerc 29: 1454-61, 1997.
13. Delp, M. D., R. M. McAllister, and M. H. Laughlin. Exercise training alters endothelium-
dependent vasoreactivity of rat abdominal aorta. J Appl Physiol 75: 1354-63, 1993.
14. Ford, J. J. Serum estrogen concentrations during postnatal development in male pigs. Proc
Soc Exp Biol Med 174: 160-4., 1983.
15. Ford, J. J. Sustained influence of previous estradiol or testosterone treatments on sexual
behaviors of female pigs. Hormones & Behavior 24: 484-496, 1990.
16. Grady, L. H., D. J. Nonneman, G. E. Rottinghaus, and W. V. Welshons. pH-Dependent
cytotoxicity of contaminants of phenol red for MCF-7 breast cancer cells. Endocrinology 129:
3321-3330, 1991.
17. Green, D. J., N. T. Cable, C. Fox, J. M. Rankin, and R. R. Taylor. Modification of forearm
resistance vessels by exercise training in young men. J Appl Physiol 77: 1829-33, 1994.
18. Hayashi, T., J. M. Fukuto, L. J. Ignarro, and G. Chaudhuri. Basal release of nitric oxide from

JAP-00061-2003-R1
43
aortic rings is greater in female rabbits than in male rabbits: implications for atherosclerosis.
Proc Natl Acad Sci U S A 89: 11259-63, 1992.
19. Hayashi, T., K. Yamada, T. Esaki, E. Mutoh, and A. Iguchi. Effect of estrogen on isoforms
of nitric oxide synthase: possible mechanism of anti-atherosclerotic effect of estrogen.
Gerontology 43 Suppl 1: 24-34, 1997.
20. Henricks, D. M., H. D. Guthrie, and D. L. Handlin. Plasma estrogen, progesterone and
luteinizing hormone levels during the estrous cycle in pigs. Biol Reprod 6: 210-8., 1972.
21. Johnson, L. R., J. W. Rush, J. R. Turk, E. M. Price, and M. H. Laughlin. Short-term exercise
training increases ACh-induced relaxation and eNOS protein in porcine pulmonary arteries. J
Appl Physiol 90: 1102-10., 2001.
22. Johnson, M. H., and B. J. Everitt. Essential Reproduction. Oxford: Blackwell Scientific
Publications, 1988.
23. Kirby, C. J., M. F. Smith, D. H. Keisler, and M. C. Lucy. Follicular function in lactating
dairy cows treated with sustained- release bovine somatotropin. J Dairy Sci 80: 273-85, 1997.
24. Kleinert, H., T. Wallerath, C. Euchenhofer, I. Ihrig-Biedert, H. Li, and U. Forstermann.
Estrogens increase transcription of the human endothelial NO synthase gene: analysis of the
transcription factors involved. Hypertension 31: 582-8., 1998.
25. Koller, A., A. Huang, D. Sun, and G. Kaley. Exercise training augments flow-dependent
dilation in rat skeletal muscle arterioles. Role of endothelial nitric oxide and prostaglandins. Circ
Res 76: 544-50, 1995.
26. Laughlin, M. H., C. C. Hale, L. Novela, D. Gute, N. Hamilton, and C. D. Ianuzzo.

JAP-00061-2003-R1
44
Biochemical characterization of exercise-trained porcine myocardium. J Appl Physiol 71: 229-
35., 1991.
27. Laughlin, M. H., K. A. Overholser, and M. J. Bhatte. Exercise training increases coronary
transport reserve in miniature swine. Journal of Applied Physiology 67: 1140-1149, 1989.
28. Laughlin, M. H., J. S. Pollock, J. F. Amann, M. L. Hollis, C. R. Woodman, and E. M. Price.
Training induces nonuniform increases in eNOS content along the coronary arterial tree. J Appl
Physiol 90: 501-10., 2001.
29. Laughlin, M. H., W. G. Schrage, R. M. McAllister, H. A. Garverick, and A. W. Jones.
Interaction of gender and exercise training: vasomotor reactivity of porcine skeletal muscle
arteries. J Appl Physiol 90: 216-27., 2001.
30. Lerner, D. J., and W. B. Kannel. Patterns of coronary heart disease morbidity and mortality
in the sexes: a 26-year follow-up of the Framingham population. Am Heart J 111: 383-90, 1986.
31. Levis, D. G., and J. J. Ford. The influence of androgenic and estrogenic hormones on sexual
behavior in castrated adult male pigs. Hormones & Behavior 23: 393-411, 1989.
32. MacRitchie, A. N., S. S. Jun, Z. Chen, Z. German, I. S. Yuhanna, T. S. Sherman, and P. W.
Shaul. Estrogen upregulates endothelial nitric oxide synthase gene expression in fetal pulmonary
artery endothelium. Circ Res 81: 355-62, 1997.
33. McAllister, R. M., and M. H. Laughlin. Short-term exercise training alters responses of
porcine femoral and brachial arteries. J Appl Physiol 82: 1438-44, 1997.
34. Miller, V. M. Gender, estrogen, and NOS: cautions about generalizations [editorial;
comment]. Circ Res 85: 979-81, 1999.

JAP-00061-2003-R1
45
35. Montano, M. M., W. V. Welshons, and F. S. vom Saal. Free estradiol in serum and brain
uptake of estradiol during fetal and neonatal sexual differentiation in female rats. Biol. Reprod.
53: 1198-1207, 1995.
36. Nagel, S. C., F. S. vom Saal, and W. V. Welshons. The effective free fraction of estradiol and
xeonestrogens in human serum measured by whole cell uptake assays: physiology of delivery
modifies estrogenic activity. Proc. Soc. Exp. Biol. Med. 217: 300-309, 1998.
37. Nataragan, N., G. E. Shambaugh, K. M. Elseth, G. K. Haines, and J. A. Radosevich.
Adaptation of the diphenylamine (DPA) assay to a 96-well plate tissue culture format and
comparison with the MTT assay. Biotechniques 17: 166-171, 1994.
38. New, G., S. J. Duffy, R. W. Harper, and I. T. Meredith. Estrogen improves acetylcholine-
induced but not metabolic vasodilation in biological males. Am J Physiol 277: H2341-7, 1999.
39. New, G., K. L. Timmins, S. J. Duffy, B. T. Tran, R. C. O'Brien, R. W. Harper, and I. T.
Meredith. Long-term estrogen therapy improves vascular function in male to female
transsexuals. J Am Coll Cardiol 29: 1437-44, 1997.
40. Oparil, S. Arthur C. Corcoran Memorial Lecture. Hormones and vasoprotection.
Hypertension 33: 170-6, 1999.
41. Otis, C., B. R. Wamhoff, and M. Sturek. Hyperglycemia-induced insulin resistance in
diabetic dyslipidemic Yucatan Swine. Comp. Med. , 2003.
42. Pasqualini, J. R., C. Gelly, B. L. Nguyen, and C. Vella. Importance of estrogen sulfates in
breast cancer. J. Steroid Biochem 34: 155-63, 1989.
43. Powell, K. E., P. D. Thompson, C. J. Caspersen, and J. S. Kendrick. Physical activity and the

JAP-00061-2003-R1
46
incidence of coronary heart disease. Annual Review of Public Health 8: 253-287, 1987.
44. Raeside, J. I., H. L. Christie, and R. L. Renaud. Androgen and estrogen metabolism in the
reproductive tract and accessory sex glands of the domestic boar (Sus scrofa). Biol Reprod 61:
1242-8., 1999.
45. Raeside, J. I., H. L. Christie, and R. L. Renaud. Metabolism of oestrone and oestradiol-
17beta to conjugated steroids by the accessory sex glands of the male pig. J Endocrinol 163: 49-
53., 1999.
46. Rush, J. W. E., M. H. Laughlin, C. R. Woodman, and E. M. Price. SOD-1 expression in pig
coronary arterioles is increased by exercise training. Am. J. Physiol. 279: H1890-H1897, 2000.
47. Rush, J. W. E., J. R. Turk, and M. H. Laughlin. Exercise training regulates SOD-1 and
oxidative stress in porcine aortic endothelium. Am. J. Physiol. Heart Circ. Physiol. 284: H1378-
H1387, 2003.
48. Sasano, H., J. I. Mason, and N. Sasano. Immunohistochemical analysis of cytochrome P-450
17 alpha-hydroxylase in pig adrenal cortex, testis and ovary. Mol Cell Endocrinol 62: 197-202.,
1989.
49. Schwarzenberger, F., G. S. Toole, H. L. Christie, and J. I. Raeside. Plasma levels of several
androgens and estrogens from birth to puberty in male domestic pigs. Acta Endocrinol (Copenh)
128: 173-7., 1993.
50. Sessa, W. C., K. Pritchard, N. Seyedi, J. Wang, and T. H. Hintze. Chronic exercise in dogs
increases coronary vascular nitric oxide production and endothelial cell nitric oxide synthase
gene expression. Circ Res 74: 349-53., 1994.

JAP-00061-2003-R1
47
51. Setchell, B. P., M. S. Laurie, A. P. Flint, and R. B. Heap. Transport of free and conjugated
steroids from the boar testis in lymph, venous blood and rete testis fluid. J Endocrinol 96: 127-
36., 1983.
52. Srere, P. A. Citrate synthase. Methods in Enzymology 13: 3-5, 1969.
53. Taddei, S., A. Virdis, L. Ghiadoni, P. Mattei, I. Sudano, G. Bernini, S. Pinto, and A. Salvetti.
Menopause is associated with endothelial dysfunction in women. Hypertension 28: 576-82,
1996.
54. Tietz, N. W. Clinical Guide to Laboratory Tests. Philadelphia: W. B. Saunders, 1995.
55. Vagnoni, K. E., C. E. Shaw, T. M. Phernetton, B. M. Meglin, I. M. Bird, and R. R. Magness.
Endothelial vasodilator production by uterine and systemic arteries. III. Ovarian and estrogen
effects on NO synthase. Am J Physiol 275: H1845-56., 1998.
56. Van Straaten, H. W., and C. J. Wensing. Leydig cell development in the testis of the pig. Biol
Reprod 18: 86-93., 1978.
57. Velle, W. Urinary oestrogens in the male. J. Reproduct. Fert. 12: 65-73, 1966.
58. vom Saal, F. S., B. G. Timms, M. M. Montano, P. Palanza, K. A. Thayer, S. C. Nagel, M. D.
Dhar, V. K. Ganjam, S. Parmigiani, and W. V. Welshons. Prostate enlargement in mice due to
fetal exposure to low doses of estradiol or diethylstilbestrol and opposite effects at high doses.
Proc Natl Acad Sci U S A 94: 2056-61, 1997.
59. Wang, J., M. S. Wolin, and T. H. Hintze. Chronic exercise enhances endothelium-mediated
dilation of epicardial coronary artery in conscious dogs. Circ Res 73: 829-38., 1993.
60. Wang, X., D. A. Barber, D. A. Lewis, C. G. A. McGregor, G. C. Sieck, L. A. Fitzpatrick, and

JAP-00061-2003-R1
48
V. M. Miller. Gender and transcriptional regulation of NO synthase and ET-1 in procine aortic
endothelial cells. American Journal of Physiology 273: 1962-1967, 1997.

brachial femoral
B
male female0
1
2
3
male femaleSOD-1
Nor
mal
ized
SO
D-1
prot
ein
sign
al
C
male female0
1
2
3
male femaleeNOS
Nor
mal
ized
eN
OS
prot
ein
sign
al
A
†
male female0
1
2
3
male femaleSOD-1D
†
male female0
1
2
3
male femaleeNOS
†
male female0
1
2
male femaleSOD-2F
†
male female0
1
2
male femaleSOD-2
Nor
mal
ized
SO
D-2
prot
ein
sign
al
E
*
Figure 1

A B
C D
Fig 2

A B
C D
Fig 3
brachial femoral0
1
2
brachial femoral0
1
2
brachial femoral0
1
2
brachial femoral0
1
2
brachial femoral0
1
2
brachial femoral0
1
2
Nor
mal
ized
eN
OS
prot
ein
sign
al
A
Nor
mal
ized
SO
D-1
prot
ein
sign
al
C
Nor
mal
ized
SO
D-2
prot
ein
sign
al
E
B
D
F
male female
†
*
Figure 4
male female0.0
0.2
0.4
0.6
0.8
1.0
1.2
Nor
mal
ized
ER
m
RN
A s
igna
l
ERα
primer
dye
male female
Figure 5
A B
DC
Fig 6

SED EX SED EXmale female
Nor
mal
ized
eN
OS
prot
ein
sign
al5
4
3
2
1
0SED EX SED EX
male female
5
4
3
2
1
0
SED EX SED EXmale female
Nor
mal
ized
SO
D-2
prot
ein
sign
al
3
2
1
0SED EX SED EX
male female
3
2
1
0
Nor
mal
ized
SO
D-1
prot
ein
sign
al
3
2
1
0SED EX SED EX
male female
3
2
1
0SED EX SED EX
male female
A B
C D
E F
brachial femoral
†
**
*
†
Figure 7

Figure 8
Pig 1343
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.61.1 pg E2 eq./ml serum
Minutes
Pig 1284
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.642 pg E2 eq./ml serum
Minutes
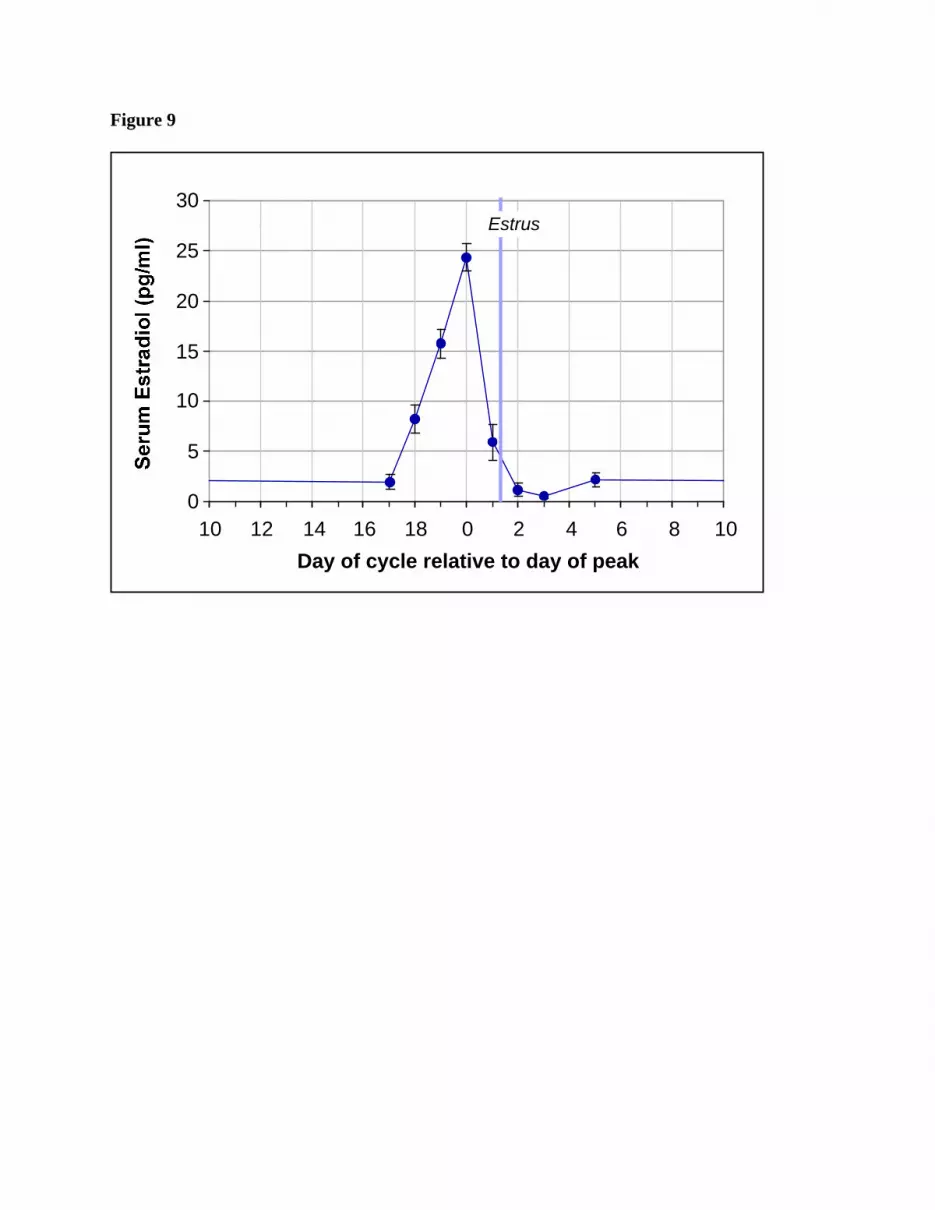
Figure 9
0
5
10
15
20
25
30
10 12 14 16 18 0 2 4 6 8 10
Day of cycle relative to day of peak
Estrus
Copyright © 2022 FDOKUMEN




















