
Galiellalactone is a Direct Inhibitor of STAT3 in Prostate Cancer Cells
11
Galiellalactone Is a Direct Inhibitor of the Transcription Factor STAT3 in Prostate Cancer Cells * Received for publication, March 10, 2014, and in revised form, April 21, 2014 Published, JBC Papers in Press, April 22, 2014, DOI 10.1074/jbc.M114.564252 Nicholas Don-Doncow ‡ , Zilma Escobar § , Martin Johansson ‡ , Sven Kjellstro ¨m ¶ , Victor Garcia , Eduardo Munoz , Olov Sterner § , Anders Bjartell ‡ , and Rebecka Hellsten ‡1 From the ‡ Division of Urological Cancers, Lund University, SE-205 02 Malmo ¨, Sweden, the § Division of Organic Chemistry, Lund University, SE-221 00 Lund, Sweden, the ¶ Department of Biochemistry and Structural Biology, Lund University, SE-221 00 Lund, Sweden, and the Maimonides Institute for Research in Biomedicine of Cordoba, Reina Sofia University Hospital, University of Co ´rdoba, 14004 Co ´rdoba, Spain Background: STAT3 is constitutively active in castration-resistant prostate cancer and the fungal metabolite galiellalactone inhibits STAT3 signaling. Results: Galiellalactone binds covalently to one or more cysteines in STAT3 and prevents STAT3 binding to DNA. Conclusion: Galiellalactone inhibits STAT3 signaling by binding directly to STAT3. Significance: Galiellalactone is a promising direct STAT3 inhibitor for treatment of castration-resistant prostate cancer. The transcription factor STAT3 is constitutively active in several malignancies including castration-resistant prostate cancer and has been identified as a promising therapeutic target. The fungal metabolite galiellalactone, a STAT3 signaling inhibitor, inhibits the growth, both in vitro and in vivo, of prostate cancer cells expressing active STAT3 and induces apoptosis of prostate cancer stem cell-like cells expressing phosphorylated STAT3 (pSTAT3). However, the molecular mechanism of this STAT3-inhibiting effect by galiellalactone has not been clarified. A biotinylated ana- logue of galiellalactone (GL-biot) was synthesized to be used for identification of galiellalactone target proteins. By adding strepta- vidin-Sepharose beads to GL-biot-treated DU145 cell lysates, STAT3 was isolated and identified as a target protein. Confocal microscopy revealed GL-biot in both the cytoplasm and the nucleus of DU145 cells treated with GL-biot, appearing to co-local- ize with STAT3 in the nucleus. Galiellalactone inhibited STAT3 binding to DNA in DU145 cell lysates without affecting phosphor- ylation status of STAT3. Mass spectrometry analysis of recombi- nant STAT3 protein pretreated with galiellalactone revealed three modified cysteines (Cys-367, Cys-468, and Cys-542). Here we dem- onstrate with chemical and molecular pharmacological methods that galiellalactone is a cysteine reactive inhibitor that covalently binds to one or more cysteines in STAT3 and that this leads to inhibition of STAT3 binding to DNA and thus blocks STAT3 sig- naling without affecting phosphorylation. This further validates galiellalactone as a promising direct STAT3 inhibitor for treatment of castration-resistant prostate cancer. The transcription factor signal transducer and activator of transcription 3 (STAT3) 2 is involved in many cellular processes including proliferation, survival, and immune response, and the transient activation of STAT3 is tightly regulated under normal conditions (1). Furthermore, the STAT3 signaling pathway is constitutively activated in several malignancies, including mul- tiple forms of solid and hematological cancers, and is shown to be involved in anti-apoptotic effects and drug resistance (1–3). The mode of constitutive activation of STAT3 through post- translational modifications, e.g. tyrosine and serine phosphor- ylation and lysine acetylation, may vary between different types of cancer (4). Aberrant signaling of upstream tyrosine kinases may be due to mutations or gene amplifications as well as increased expression of growth factors, cytokines, and ligand receptors, which all may lead to constitutive activation of STAT3 and malignant transformation (5, 6). Somatic muta- tions of the STAT3 gene are uncommon, although such are described in leukemia and hepatocellular adenomas (7, 8). Prostate cancer is the second most common cancer in men world wide and the fifth most common cancer overall (2008) (9). Initially, prostate cancer cells respond to androgen deprivation therapy, but within 12–18 months many patients develop castra- tion-resistant prostate cancer with a need for second-line therapy (10). Although cytotoxic drugs and recently approved drugs for more efficient blockade of androgen signaling, including abi- raterone and enzalutamide, are available, there is an obvious need for new and efficient treatment strategies in metastatic castration-resistant prostate cancer (11). Activated STAT3 has been correlated to the malignant potential of prostate cancer cells, disease progression, and increased Gleason score (12–14) and shown to promote meta- static progression of prostate cancer (15). Furthermore, the JAK/STAT signaling pathway is associated with a cancer stem * This work was supported by grants from the Swedish Cancer Society, the Swedish Research Council Medicine, government funding of clinical research within the National Health Service, Lund University (Sweden), governmental strategic funding for cancer research at Lund and Go ¨ teborg Universities (BioCare), the Percy Falk Foundation, and the Gunnar Nilsson Cancer Foundation. This article was selected as a Paper of the Week. 1 To whom correspondence should be addressed: Division of Urological Cancers, Lund University, CMP, Malmo ¨ University Hospital, Jan Waldenstro ¨ ms gata 59, SE-205 02 Malmo ¨ , Sweden. Tel.: 46-40-336069; E-mail: rebecka.hellsten@ med.lu.se. 2 The abbreviations used are: STAT3, signal transducer and activator of tran- scription 3; pSTAT3, phosphorylated STAT3; GL, galiellalactone; GL-biot, biotinylated galiellalactone analogue ZE139; SUMO, small ubiquitin-like modifier; Bis-Tris, 2-(bis(2-hydroxyethyl)amino)-2-(hydroxymethyl)pro- pane-1,3-diol; DMSO, dimethyl sulfoxide. THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 289, NO. 23, pp. 15969 –15978, June 6, 2014 © 2014 by The American Society for Biochemistry and Molecular Biology, Inc. Published in the U.S.A. JUNE 6, 2014 • VOLUME 289 • NUMBER 23 JOURNAL OF BIOLOGICAL CHEMISTRY 15969 by guest on June 15, 2016 http://www.jbc.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Galiellalactone is a Direct Inhibitor of STAT3 in Prostate Cancer Cells

Galiellalactone Is a Direct Inhibitor of the Transcription FactorSTAT3 in Prostate Cancer Cells*�
Received for publication, March 10, 2014, and in revised form, April 21, 2014 Published, JBC Papers in Press, April 22, 2014, DOI 10.1074/jbc.M114.564252
Nicholas Don-Doncow‡, Zilma Escobar§, Martin Johansson‡, Sven Kjellstrom¶, Victor Garcia�, Eduardo Munoz�,Olov Sterner§, Anders Bjartell‡, and Rebecka Hellsten‡1
From the ‡Division of Urological Cancers, Lund University, SE-205 02 Malmo, Sweden, the §Division of Organic Chemistry, LundUniversity, SE-221 00 Lund, Sweden, the ¶Department of Biochemistry and Structural Biology, Lund University, SE-221 00 Lund,Sweden, and the �Maimonides Institute for Research in Biomedicine of Cordoba, Reina Sofia University Hospital, University ofCordoba, 14004 Cordoba, Spain
Background: STAT3 is constitutively active in castration-resistant prostate cancer and the fungal metabolite galiellalactoneinhibits STAT3 signaling.Results: Galiellalactone binds covalently to one or more cysteines in STAT3 and prevents STAT3 binding to DNA.Conclusion: Galiellalactone inhibits STAT3 signaling by binding directly to STAT3.Significance: Galiellalactone is a promising direct STAT3 inhibitor for treatment of castration-resistant prostate cancer.
The transcription factor STAT3 is constitutively active in severalmalignancies including castration-resistant prostate cancer andhas been identified as a promising therapeutic target. The fungalmetabolite galiellalactone, a STAT3 signaling inhibitor, inhibitsthe growth, both in vitro and in vivo, of prostate cancer cellsexpressing active STAT3 and induces apoptosis of prostate cancerstem cell-like cells expressing phosphorylated STAT3 (pSTAT3).However, the molecular mechanism of this STAT3-inhibitingeffect by galiellalactone has not been clarified. A biotinylated ana-logue of galiellalactone (GL-biot) was synthesized to be used foridentification of galiellalactone target proteins. By adding strepta-vidin-Sepharose beads to GL-biot-treated DU145 cell lysates,STAT3 was isolated and identified as a target protein. Confocalmicroscopy revealed GL-biot in both the cytoplasm and thenucleus of DU145 cells treated with GL-biot, appearing to co-local-ize with STAT3 in the nucleus. Galiellalactone inhibited STAT3binding to DNA in DU145 cell lysates without affecting phosphor-ylation status of STAT3. Mass spectrometry analysis of recombi-nant STAT3 protein pretreated with galiellalactone revealed threemodified cysteines (Cys-367, Cys-468, and Cys-542). Here we dem-onstrate with chemical and molecular pharmacological methodsthat galiellalactone is a cysteine reactive inhibitor that covalentlybinds to one or more cysteines in STAT3 and that this leads toinhibition of STAT3 binding to DNA and thus blocks STAT3 sig-naling without affecting phosphorylation. This further validatesgaliellalactone as a promising direct STAT3 inhibitor for treatmentof castration-resistant prostate cancer.
The transcription factor signal transducer and activator oftranscription 3 (STAT3)2 is involved in many cellular processesincluding proliferation, survival, and immune response, and thetransient activation of STAT3 is tightly regulated under normalconditions (1). Furthermore, the STAT3 signaling pathway isconstitutively activated in several malignancies, including mul-tiple forms of solid and hematological cancers, and is shown tobe involved in anti-apoptotic effects and drug resistance (1–3).
The mode of constitutive activation of STAT3 through post-translational modifications, e.g. tyrosine and serine phosphor-ylation and lysine acetylation, may vary between different typesof cancer (4). Aberrant signaling of upstream tyrosine kinasesmay be due to mutations or gene amplifications as well asincreased expression of growth factors, cytokines, and ligandreceptors, which all may lead to constitutive activation ofSTAT3 and malignant transformation (5, 6). Somatic muta-tions of the STAT3 gene are uncommon, although such aredescribed in leukemia and hepatocellular adenomas (7, 8).
Prostate cancer is the second most common cancer in menworld wide and the fifth most common cancer overall (2008)(9). Initially, prostate cancer cells respond to androgen deprivationtherapy, but within 12–18 months many patients develop castra-tion-resistant prostate cancer with a need for second-line therapy(10). Although cytotoxic drugs and recently approved drugs formore efficient blockade of androgen signaling, including abi-raterone and enzalutamide, are available, there is an obviousneed for new and efficient treatment strategies in metastaticcastration-resistant prostate cancer (11).
Activated STAT3 has been correlated to the malignantpotential of prostate cancer cells, disease progression, andincreased Gleason score (12–14) and shown to promote meta-static progression of prostate cancer (15). Furthermore, theJAK/STAT signaling pathway is associated with a cancer stem
* This work was supported by grants from the Swedish Cancer Society, theSwedish Research Council Medicine, government funding of clinicalresearch within the National Health Service, Lund University (Sweden),governmental strategic funding for cancer research at Lund and GoteborgUniversities (BioCare), the Percy Falk Foundation, and the Gunnar NilssonCancer Foundation.
� This article was selected as a Paper of the Week.1 To whom correspondence should be addressed: Division of Urological Cancers,
Lund University, CMP, Malmo University Hospital, Jan Waldenstroms gata 59,SE-205 02 Malmo, Sweden. Tel.: 46-40-336069; E-mail: [email protected].
2 The abbreviations used are: STAT3, signal transducer and activator of tran-scription 3; pSTAT3, phosphorylated STAT3; GL, galiellalactone; GL-biot,biotinylated galiellalactone analogue ZE139; SUMO, small ubiquitin-likemodifier; Bis-Tris, 2-(bis(2-hydroxyethyl)amino)-2-(hydroxymethyl)pro-pane-1,3-diol; DMSO, dimethyl sulfoxide.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 289, NO. 23, pp. 15969 –15978, June 6, 2014© 2014 by The American Society for Biochemistry and Molecular Biology, Inc. Published in the U.S.A.
JUNE 6, 2014 • VOLUME 289 • NUMBER 23 JOURNAL OF BIOLOGICAL CHEMISTRY 15969
by guest on June 15, 2016http://w
ww
.jbc.org/D
ownloaded from

cell-like phenotype in prostate cancer, and blocking of thispathway may inhibit the initiation of tumors (16). Targeting thetranscription factor STAT3 appears to be a promising treat-ment strategy for patients with advanced prostate cancer, andSTAT3 has been identified as a relevant target protein for thedevelopment of new therapies in this group of patients (17, 18).
The fungal metabolite galiellalactone is a small non-toxic andnon-mutagenic molecule that has been shown to preventSTAT3 signaling by blocking the binding of STAT3 to STAT3-specific transcriptional DNA elements (19). We have previ-ously demonstrated that galiellalactone inhibits the growth,both in vitro and in vivo, of prostate cancer cells expressingactivated STAT3 and inhibits the expression of STAT3-regu-lated genes and proteins (20). Furthermore, galiellalactoneinhibits growth and induces apoptosis of prostate cancer stemcell-like cells expressing phosphorylated STAT3 (pSTAT3)(21).
Galiellalactone contains a reactive �,�-unsaturated lactonefunctionality, and galiellalactone has been demonstrated toreact with nitrogen- and sulfur-nucleophiles to produce inac-tive adducts (22). With the reactive potential of galiellalactonetoward biological nucleophiles in consideration, we were inter-ested in investigating whether galiellalactone can alkylateSTAT3 and thereby inhibit the DNA binding as there is prece-dence that direct covalent modification of STAT3 with smallmolecules (23, 24) or through cysteine oxidation (25) can blockthe transcriptional activity of STAT3. The aim of the presentstudy is to elucidate in more detail the mechanism of action ofgaliellalactone using human prostate cancer cells as a model.
EXPERIMENTAL PROCEDURES
Biotinylated Galiellalactone Analogues—Biotinylated galiel-lalactone analogues were synthesized to be used as a tool foridentification of target proteins bound to galiellalactone. Thecomplete synthetic procedure will be published elsewhere.3Galiellalactone was prepared by synthesis as described previ-ously (26).
Synthesis of Galiellalactone-Cysteine Adduct—3.5 mg (0.02mmol) of galiellalactone was dissolved in 1.0 ml of MeOH with2.4 mg (0.018 mmol) of L-cysteine. The solution was stirred atroom temperature for 1 h. The solvent was removed underreduced pressure, and the residue was washed with chloroformto afford 5.6 mg (89%) of the pure adduct. 1H NMR (DMSO-d6)d 0.50 (1H, ddd, J1 � 25.4 Hz, J2 � 12.2 Hz, J3 � 12.2 Hz), 1.05(3H, d, J � 6.55 Hz), 1.84 (1H, m), 1.86 (1H, m), 2.01 (1H, m),2.06 (1H, m), 2.70 (1H, dd, J1 � 25.4 Hz, J2 � 12.2 Hz), 2.97 (1H,dd, J1 � 13.8 Hz, J2 � 8.0 Hz), 3.14 (1H, dd, J1 � 13.7 Hz, J2 � 3.5Hz), 3.42 (1H, dd, J1 � 8.0 Hz, J2 � 3.7 Hz), 3.53 (1H, s), 4.47(1H, s); 13C NMR (DMSO-d6) d 20.5, 28.8, 31.3, 31.9, 32.8, 37.0,45.2, 47.3, 52., 54.0, 83.1, 88.7, 168.7, 175.6; HRMS (FAB�)calculated for C14H21NO5S (M�H) 316.1219, found 316.1231.
Cell Culture and Preparation of Cell Lysates—The humanprostate cancer cell lines DU145 and LNCaP (American TypeCulture Collection) were used. The cells were cultured in RPMI1640 medium supplemented with 10% FBS and 1% penicillin-
streptomycin and incubated at 37 °C in a humidified atmo-sphere of 95% O2 and 5% CO2. Cells were used at low passagesand not cultured for more than 2–3 months. Cells were rou-tinely tested for and found free of mycoplasma. Cell identitywas not authenticated by us. For preparation of lysates, cellswere washed and scraped in ice-cold PBS and centrifuged at1400 rpm at 4 °C. The cells were lysed in mild cell lysis buffercontaining 100 mM Tris HCl, 150 mM NaCl, 0.1% Nonidet P-40,1 mM EDTA, Complete Mini EDTA-free protease inhibitor(Roche Applied Science), incubated for 30 min on ice, and cen-trifuged at 10,000 rpm for 30 min at 4 °C. The supernatant wascollected, and protein content was determined using Bradfordprotein assay.
WST-1 Cell Proliferation Assay—The functional activity ofbiotinylated galiellalactone analogues was evaluated usingWST-1 proliferation assay on DU145 cells, which express con-stitutively activated STAT3. DU145 cells were cultured in 96-wellplates (2000 cells/well in 200 �l of medium) and allowed to set for24 h. The cells were treated with 0 – 60 �M galiellalactone orbiotinylated galiellalactone analogue for 72 h. Samples weremade in triplicate. 20 �l of WST-1 solution (Roche AppliedScience) was added per well and incubated at 37 °C for 4 h. Theabsorbance of each well was measured using a scanning multi-well spectrophotometer, ELISA reader at a wavelength of 450nm, and reference wavelength of 690 nm. The results are pre-sented as the percentage of untreated control cells.
Luciferase Reporter Assay—The inhibitory effect of galiella-lactone and biotinylated galiellalactone analogue ZE139 (GL-biot) on STAT3 signaling was assessed using a luciferasereporter gene assay. LNCaP cells were transiently transfectedwith the pTATA-TK-Luc plasmid that contains four copies ofthe STAT binding site. Transient transfections were performedwith Roti-Fect (Carl Roth GmbH) according to the manufactu-rer’s instructions, and 24 h after transfection, the cells werepreincubated with galiellalactone or GL-biot for 1 h and thentreated with IL-6 (50 ng/ml) for another 8 h. For wash-outexperiments, LNCaP cells were transfected with the pTATA-TK-Luc plasmid for 24 h and incubated for 1 h with galiellalac-tone, washed twice with PBS, and stimulated with IL-6 (50ng/ml) for another 4 h. Cells were collected and lysed; luciferaseassay was performed using luciferase assay reagent (Promega)according to the manufacturer’s instructions; and luciferaseactivity was measured in an Autolumat LB9510 luminometer(Berthold Technologies GmbH). Luciferase activity in differentsamples was normalized with protein concentration. Theresults are represented as the percentage of inhibition consid-ering 100% activation, the results that were obtained with IL-6in the absence of galiellalactone.
Western Blot Analysis—Samples were separated on 7.5% pre-cast gel (Mini-PROTEAN TGX; Bio-Rad) or 8% Bis-Tris self-cast gels. The gels were blotted onto PVDF membranes andblocked with 5% milk or 5% BSA. Membranes were incubatedwith primary antibody diluted in 5% milk or 5% BSA for 1 h atroom temperature or overnight at 4 °C with antibodies raisedagainst STAT3, pSTAT3 Tyr-705, pSTAT3 Ser-727 (Cell Sig-naling Technology), or biotin (Santa Cruz Biotechnology).After incubation with secondary anti-mouse or anti-rabbitantibody conjugated with horseradish peroxidase (GE Health-
3 Z. Escobar, M. Johansson, A. Bjartell, R. Hellsten, and O. Sterner, manuscriptin preparation.
Galiellalactone Is a Direct Inhibitor of STAT3
15970 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 289 • NUMBER 23 • JUNE 6, 2014
by guest on June 15, 2016http://w
ww
.jbc.org/D
ownloaded from

care), the membrane was treated with enhanced chemilumines-cent reagent (Santa Cruz Biotechnology or Millipore) followedby exposure to x-ray film or visualized using a ChemiDoc XRSsystem (Bio-Rad).
Isolation of Galiellalactone-interacting Proteins Using Bioti-nylated Galiellalactone Analogue—To isolate target proteins,intact cells or whole cell lysates of DU145 or LNCaP cells weretreated with GL-biot for 1 h. Cell lysates were incubated withmagnetic streptavidin-Sepharose beads (GE Healthcare), andbound proteins were eluted in 2% SDS and identified usingWestern blot analysis. For competition assays, DU145 cells orDU145 cell lysates were pretreated with galiellalactone (0 –100�M) for 1 h followed by the addition of 25 �M GL-biot for 1 h.The reverse was also investigated where DU145 cell lysateswere pretreated with 25 �M GL-biot before the addition of0 –100 �M galiellalactone.
Confocal Microscopy—Localization of GL-biot and possibleco-localization of GL-biot with STAT3 were assessed usingconfocal microscopy. DU145 cells were grown on chamberslides. After 24 h, the cells were treated with 50 �M GL-biot for2 h before the cells were fixed with paraformaldehyde with sub-sequent methanol fixation and permeabilized and blocked with3% BSA, 0.3% Triton-X in PBS for 1 h. Slides were incubatedwith STAT3 primary antibody (Cell Signaling Technology)overnight, washed in PBS, and subsequently incubated withAlexa Fluor 594-conjugated streptavidin and secondary AlexaFluor 488-conjugated anti-mouse antibody (Invitrogen) for 1 h.Slides were washed in PBS and mounted using DAPI-contain-ing VECTASHIELD mounting medium (Vector Laboratories).The cells on the slides were analyzed by confocal laser scanningmicroscopy using the Zeiss LSM 710 system (Carl Zeiss AG).
Electric Mobility Shift Assay—STAT3 consensus sequence(Santa Cruz Biotechnology) was labeled with digoxigenin-ddUTP (Roche digoxigenin gel shift kit, Roche Applied Sci-ence) according to the manufacturer’s protocol. Lysates fromDU145 cells treated with galiellalactone for 1 h or DU145 celllysates treated with galiellalactone for 15 min on ice were incu-bated with labeled probes for 15 min at room temperature.Supershift assays were performed by pretreating the cell lysatewith STAT3 antibody (Cell Signaling Technology). The com-plexes were resolved on a 6% DNA retardation gel (Invitrogen)in 0.5� Tris-borate-EDTA buffer. The DNA protein complexwas transferred onto a positively charged membrane and cross-linked. Visualization of bands was performed according to themanufacturer’s protocol.
Recombinant pSTAT3 Protein—A synthetic version of theSTAT3 gene, codon-optimized for Escherichia coli (DNA 2.0)encoding amino acids 127–722, was amplified by PCR andligated into pETm11-SUMO3 (European Molecular BiologyLaboratory (EMBL)). The resulting plasmid, pETm11-SUMO3-STAT3, encodes STAT3 with a cleavable N-terminalHis6-SUMO tag. The plasmid was verified by sequencing andtransformed into E. coli BL21(DE3) TKB1 (Stratagene), whichharbors a plasmid-encoded tyrosine kinase gene behind a pro-moter inducible with indoleacrylic acid. Cells were grown in LBwith 50 �g/ml kanamycin and 12.5 �g/ml tetracycline at 37 °C,120 rpm. At A600 � 0.6, isopropyl-�-D-thio-galactopyranosidewas added to a final concentration of 1 mM. Three hours after
induction, cells were harvested and resuspended in tyrosinekinase induction medium containing 53 �M indoleacrylic acidand grown for 2 h. Cells were harvested and resuspended inwash buffer (50 mM NaPO4, 300 mM NaCl, 20 mM imidazole,pH 8.0) supplemented with Complete protease inhibitor,EDTA-free (Roche Applied Science) and lysed with a Frenchpress. The lysate was ultracentrifuged (180,000 � g, 60 min,4 °C), and the supernatant was used for affinity chromatogra-phy on a 1-ml HisTrap HP column (GE Healthcare). The col-umn with sample was washed with wash buffer until a stable UVsignal was obtained. The bound protein was eluted with 50 mM
NaPO4, 300 mM NaCl, 500 mM imidazole, pH 8.0. The elutionpeak fractions were digested with His-tagged SenP2 protease atan enzyme:substrate mass ratio of 1:250 in the presence of 2 mM
DTT. To remove the SenP2 protease and the His-SUMO tag,the sample was dialyzed against wash buffer and loaded againonto a 1-ml HisTrap HP column. The column was washed withwash buffer, and the flow-through and wash fractions werepooled and used for size exclusion chromatography on aHiLoad 26/600 Superdex 200-pg gel filtration column using 50mM Tris, 150 mM NaCl, pH 8.0. Peak fractions from the sizeexclusion chromatography were analyzed with SDS-PAGE. Gelfiltration fractions containing STAT3 were pooled and concen-trated. The purity of the protein was estimated to �95% usingSDS-PAGE analysis.
Mass Spectrometry Analysis—Samples obtained after galiel-lalactone derivatization were partly denaturated with 2 M
UREA prior to in-solution digestion. The samples were dilutedwith 50 mM ammonium bicarbonate to raise the pH, and analiquot of trypsin solution (Promega) was added in a 1:50 tryp-sin:protein ratio (w/w). Enzymatic digest was carried out at37 °C overnight and was stopped by the addition of 10% trifluo-roacetic acid. The experiments were performed with an EasyLCnanoflow HPLC interfaced with a nanoEasy spray ion source(Proxeon Biosystems, Odense, Denmark) connected to anLTQ-Orbitrap Velos Pro mass spectrometer (Thermo FisherScientific). The chromatographic separation was performed at40 °C on a 15-cm (75-�m inner diameter) EASY-Spray columnpacked with 3 �m of resin (Proxeon Biosystems). The nano-HPLC was operating at 300 nl min�1 with a gradient of 5–20%solvent B (0.1% (v/v) formic acid, 100% (v/v) acetonitrile inwater) in solvent A (0.1% (v/v) formic acid in water) during 60min and then 20 – 40% during 30 min followed with an increaseto 90% during 5 min. An MS scan (400 –1400 m/z) was recordedin the Orbitrap mass analyzer set at a resolution of 60,000 at 400m/z, 1 � 106 automatic gain control target, and 500-ms maxi-mum ion injection time. The MS was followed by data-depen-dent collision-induced dissociation MS/MS scans on the eightmost intense multiply charged ions. The general mass spectro-metric conditions were as follows: spray voltage, 2.0 kV; nosheath or auxiliary gas flow; S-lens 60%; ion transfer tube tem-perature, 275 °C. Raw data were processed by Mascot distillersearching the Swiss-Prot database with an in-house Mascotdatabase. The search parameters for the Mascot searches were:Taxonomy: Homo sapiens; Enzyme, No enzyme; Variable Mod-ifications, Galiellalactone (Cys); Precursor Tolerance, 20 ppm;MS/MS Fragment Tolerance, 0.1 Da.
Galiellalactone Is a Direct Inhibitor of STAT3
JUNE 6, 2014 • VOLUME 289 • NUMBER 23 JOURNAL OF BIOLOGICAL CHEMISTRY 15971
by guest on June 15, 2016http://w
ww
.jbc.org/D
ownloaded from

RESULTS
Activity of Biotinylated Galiellalactone Analogues and Cys-teine Reactivity of Galiellalactone—Two biotinylated ana-logues of galiellalactone (ZE139 and ZE140) were synthesized(Fig. 1A), and ZE139 was chosen for further experiments basedon the inhibitory effect on cell proliferation and STAT3 activity(Fig. 1, B and C). From previous studies, we knew that an addedphenyl group in position 7 of the tricyclic structure of galiella-lactone gave analogues with increased potency.3 Galiellalac-tone and the biotinylated analogues ZE139 and ZE140 inhibitproliferation of DU145 cells expressing constitutively activeSTAT3 with growth IC50 values of 3.6, 6.6, and 14 �M, respec-tively after 72 h (Fig. 1B). Galiellalactone and ZE139 showedequal inhibitory effect on STAT3 signaling activity as assessed
by luciferase reporter gene assay (Fig. 1C). To demonstrate thecysteine reactivity of galiellalactone, galiellalactone was treatedwith L-cysteine in PBS buffer at 37 °C or in MeOH at roomtemperature. A complete conversion to an isolatable adductwas observed in less than 15 min. L-Cysteine was added to theconjugated double bond in a completely diastereoselectivemanner as determined by NMR (Fig. 1D).
Biotinylated Galiellalactone Analogue Binds Directly toSTAT3—To investigate the suggested covalent protein interac-tions of galiellalactone in an unbiased way, DU145 cell lysatesand intact DU145 cells were treated with GL-biot, and the pro-teins bound were precipitated using streptavidin-Sepharosebeads. STAT3 was successfully isolated and identified as a tar-get protein by immunoblotting (Fig. 2A). GL-biot was able to
FIGURE 1. Chemical structure and activity of biotinylated galiellalactone analogues. A, chemical structures of galiellalactone and the two biotinylatedderivatives utilized in this study. B, galiellalactone and biotinylated galiellalactone analogues ZE139 and ZE140 inhibit proliferation of DU145 cells with growthIC50 values of 3.6, 6.6, and 14 �M, respectively, after 72 h. C, the inhibitory effect of galiellalactone (GL) and ZE139 on STAT3 activity was investigated in aluciferase reporter gene assay using IL-6-stimulated LNCaP cells. The STAT3-dependent luciferase activity is expressed as the percentage of activation ofrelative control and presented as mean � S.E. from two separate experiments. D, galiellalactone undergoes a Michael addition with L-cysteine in PBS or MeOHto form a stable adduct.
Galiellalactone Is a Direct Inhibitor of STAT3
15972 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 289 • NUMBER 23 • JUNE 6, 2014
by guest on June 15, 2016http://w
ww
.jbc.org/D
ownloaded from

bind to STAT3 both in intact tumor cells and in cell lysates. Inaddition, GL-biot was able to bind to recombinant STAT3 pro-tein (Fig. 2A). Furthermore, pretreatment of DU145 cell lysateswith galiellalactone prior to the addition of GL-biot preventedthe binding of the GL-biot to STAT3 in a dose-dependent man-ner (Fig. 2B) demonstrating competitive binding. In whole celllysates of GL-biot-treated DU145 cells subjected to Westernblot analysis, several bands were detected using anti-biotinantibody, clearly demonstrating that GL-biot had bound to sev-eral protein targets (Fig. 2C).
DU145 cells treated with GL-biot were subjected to confocalmicroscopy to demonstrate the localization of the bound com-pound in cellular compartments (Fig. 2D). GL-biot wasobserved in both the cytoplasm and the nucleus of DU145 cells,appearing to co-localize with STAT3 in the nucleus. STAT3was mainly present in the nucleus. The endogenous levels ofbiotin were below the detection limit.
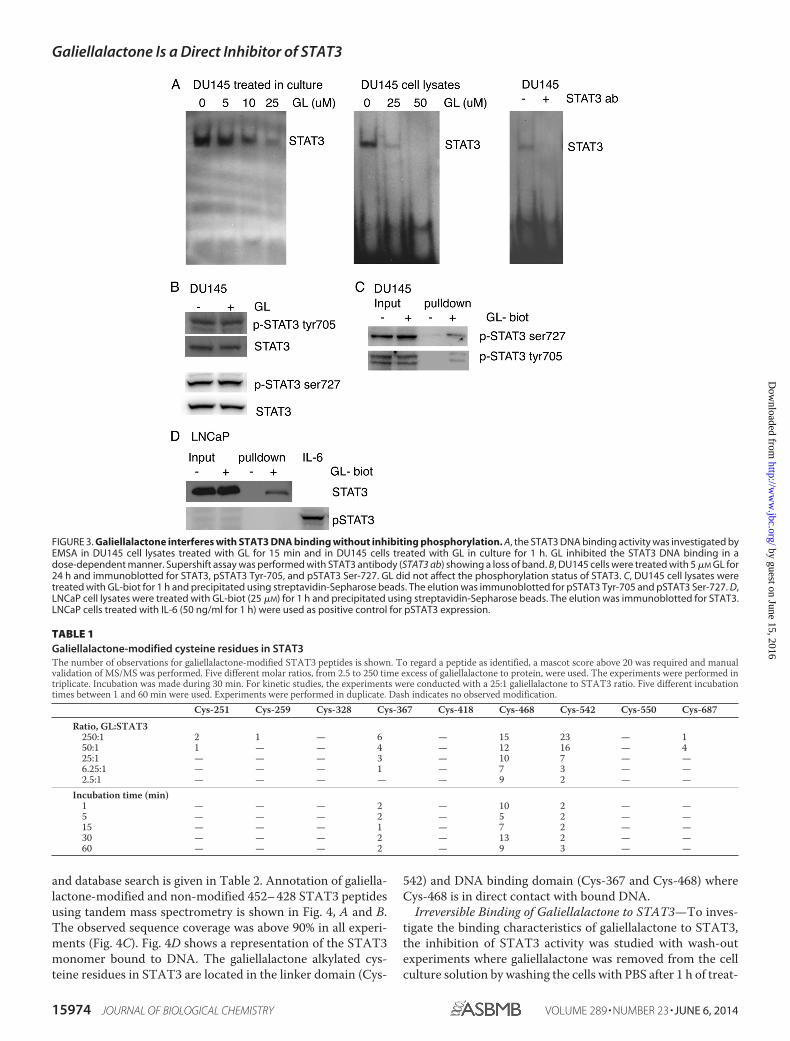
Galiellalactone Inhibits STAT3 Binding to DNA withoutAffecting Phosphorylation Status—When extracts from DU145cells pretreated with galiellalactone and DU145 cell extractsincubated with galiellalactone were subjected to EMSA with
STAT3 probes, we demonstrated a dose-dependent decrease inSTAT3 DNA binding (Fig. 3A). Galiellalactone did not affectphosphorylation of STAT3 Tyr-705 and Ser-727 or the expres-sion of total STAT3 in DU145 cells (Fig. 3B). GL-biot was foundto bind to both pSTAT3 Tyr-705 and pSTAT3 Ser-727 inDU145 cells (Fig. 3C). In LNCaP cells, which do not expressconstitutively active STAT3, GL-biot was able to bind tounphosphorylated STAT3 (Fig. 3D).
Galiellalactone Modifies Cysteine Residues in STAT3—When phosphorylated recombinant STAT3 protein pretreatedwith various concentrations of galiellalactone was digested bytrypsin and subjected to mass spectrometric analysis, we foundthree modified cysteine residues in most of the experiments.The results show that Cys-367, Cys-468, and Cys-542 in recom-binant STAT3 were the predominant cysteines modified bygaliellalactone, and at higher concentrations of galiellalactone,we could also identify modification of Cys-251, Cys-259, andCys-687 (Table 1). Kinetic studies showed that cysteines Cys-367, Cys-468, and Cys-542 all were alkylated after 1 min atmolar ratios 1:25 (protein:galiellalactone) (Table 1). A list ofgaliellalactone-modified STAT3 peptides identified by MS/MS
FIGURE 2. Galiellalactone binds directly to STAT3. A, intact DU145 cells, DU145 cell lysates, and recombinant STAT3 (rec STAT3) protein were incubated withGL-biot for 1 h. The cell lysates and recombinant STAT3 protein were incubated with streptavidin-Sepharose beads, and the elutions were immunoblotted forSTAT3. B, competitive binding was demonstrated by pretreatment of DU145 cell lysates with GL in increasing concentrations (10 –100 �M) for 1 h before theaddition of 25 �M GL-biot for 1 h. The cell lysates were incubated with streptavidin-Sepharose beads, and the elutions were immunoblotted for STAT3. C, DU145cells treated with 25 �M GL-biot for 30 min. Whole cell lysates were run on a gel and subjected to Western blot analysis using anti-biotin antibody. Arrowsindicate bands corresponding to the size of STAT3 (79 and 86 kDa) and NF-�B subunit p65 (65 kDa). D, localization of STAT3 and GL-biot in GL-biot-treatedDU145 cells using confocal microscopy. STAT3 was mainly located in the nucleus, and GL-biot was detected both in the nucleus and in the cytoplasm. GL-biotappeared to co-localize with STAT3. STAT3 was detected using STAT3 primary antibody and secondary Alexa Fluor 488 conjugated anti-mouse antibody. Biotinwas detected using Alexa Fluor 594-conjugated streptavidin. The nucleus was visualized by DAPI.
Galiellalactone Is a Direct Inhibitor of STAT3
JUNE 6, 2014 • VOLUME 289 • NUMBER 23 JOURNAL OF BIOLOGICAL CHEMISTRY 15973
by guest on June 15, 2016http://w
ww
.jbc.org/D
ownloaded from
and database search is given in Table 2. Annotation of galiella-lactone-modified and non-modified 452– 428 STAT3 peptidesusing tandem mass spectrometry is shown in Fig. 4, A and B.The observed sequence coverage was above 90% in all experi-ments (Fig. 4C). Fig. 4D shows a representation of the STAT3monomer bound to DNA. The galiellalactone alkylated cys-teine residues in STAT3 are located in the linker domain (Cys-
542) and DNA binding domain (Cys-367 and Cys-468) whereCys-468 is in direct contact with bound DNA.
Irreversible Binding of Galiellalactone to STAT3—To inves-tigate the binding characteristics of galiellalactone to STAT3,the inhibition of STAT3 activity was studied with wash-outexperiments where galiellalactone was removed from the cellculture solution by washing the cells with PBS after 1 h of treat-
FIGURE 3. Galiellalactone interferes with STAT3 DNA binding without inhibiting phosphorylation. A, the STAT3 DNA binding activity was investigated byEMSA in DU145 cell lysates treated with GL for 15 min and in DU145 cells treated with GL in culture for 1 h. GL inhibited the STAT3 DNA binding in adose-dependent manner. Supershift assay was performed with STAT3 antibody (STAT3 ab) showing a loss of band. B, DU145 cells were treated with 5 �M GL for24 h and immunoblotted for STAT3, pSTAT3 Tyr-705, and pSTAT3 Ser-727. GL did not affect the phosphorylation status of STAT3. C, DU145 cell lysates weretreated with GL-biot for 1 h and precipitated using streptavidin-Sepharose beads. The elution was immunoblotted for pSTAT3 Tyr-705 and pSTAT3 Ser-727. D,LNCaP cell lysates were treated with GL-biot (25 �M) for 1 h and precipitated using streptavidin-Sepharose beads. The elution was immunoblotted for STAT3.LNCaP cells treated with IL-6 (50 ng/ml for 1 h) were used as positive control for pSTAT3 expression.
TABLE 1Galiellalactone-modified cysteine residues in STAT3The number of observations for galiellalactone-modified STAT3 peptides is shown. To regard a peptide as identified, a mascot score above 20 was required and manualvalidation of MS/MS was performed. Five different molar ratios, from 2.5 to 250 time excess of galiellalactone to protein, were used. The experiments were performed intriplicate. Incubation was made during 30 min. For kinetic studies, the experiments were conducted with a 25:1 galiellalactone to STAT3 ratio. Five different incubationtimes between 1 and 60 min were used. Experiments were performed in duplicate. Dash indicates no observed modification.
Cys-251 Cys-259 Cys-328 Cys-367 Cys-418 Cys-468 Cys-542 Cys-550 Cys-687
Ratio, GL:STAT3250:1 2 1 — 6 — 15 23 — 150:1 1 — — 4 — 12 16 — 425:1 — — — 3 — 10 7 — —6.25:1 — — — 1 — 7 3 — —2.5:1 — — — — — 9 2 — —
Incubation time (min)1 — — — 2 — 10 2 — —5 — — — 2 — 5 2 — —15 — — — 1 — 7 2 — —30 — — — 2 — 13 2 — —60 — — — 2 — 9 3 — —
Galiellalactone Is a Direct Inhibitor of STAT3
15974 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 289 • NUMBER 23 • JUNE 6, 2014
by guest on June 15, 2016http://w
ww
.jbc.org/D
ownloaded from
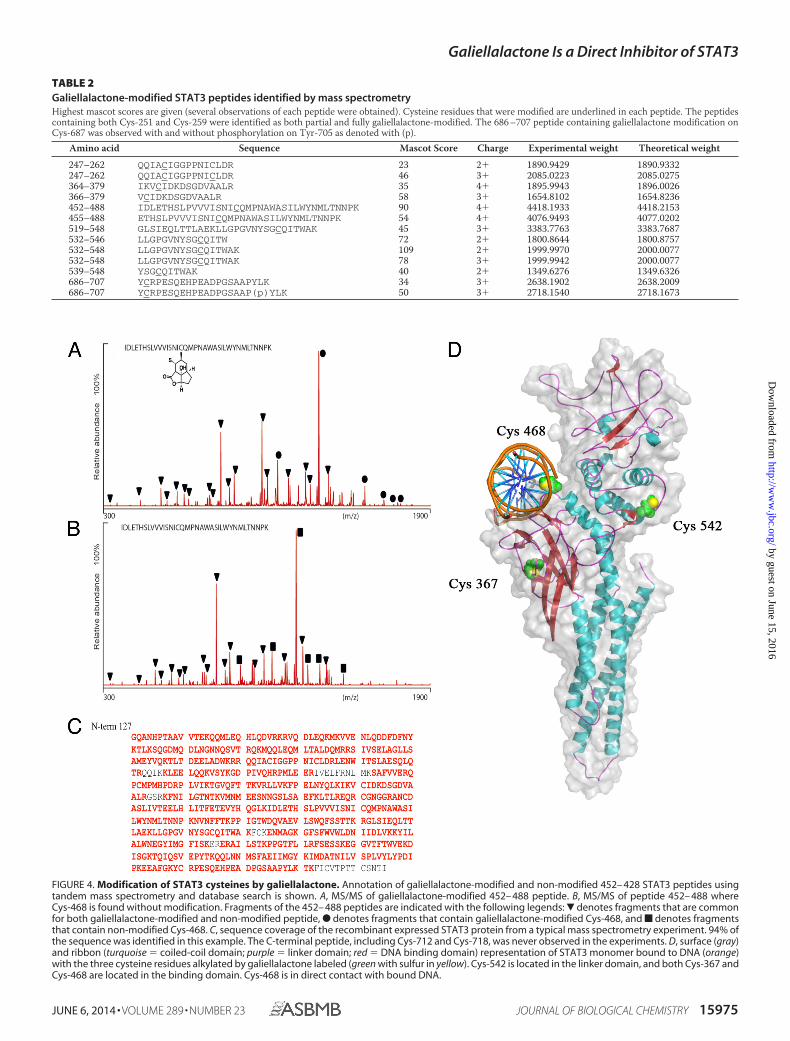
FIGURE 4. Modification of STAT3 cysteines by galiellalactone. Annotation of galiellalactone-modified and non-modified 452– 428 STAT3 peptides usingtandem mass spectrometry and database search is shown. A, MS/MS of galiellalactone-modified 452– 488 peptide. B, MS/MS of peptide 452– 488 whereCys-468 is found without modification. Fragments of the 452– 488 peptides are indicated with the following legends: � denotes fragments that are commonfor both galiellalactone-modified and non-modified peptide, F denotes fragments that contain galiellalactone-modified Cys-468, and f denotes fragmentsthat contain non-modified Cys-468. C, sequence coverage of the recombinant expressed STAT3 protein from a typical mass spectrometry experiment. 94% ofthe sequence was identified in this example. The C-terminal peptide, including Cys-712 and Cys-718, was never observed in the experiments. D, surface (gray)and ribbon (turquoise � coiled-coil domain; purple � linker domain; red � DNA binding domain) representation of STAT3 monomer bound to DNA (orange)with the three cysteine residues alkylated by galiellalactone labeled (green with sulfur in yellow). Cys-542 is located in the linker domain, and both Cys-367 andCys-468 are located in the binding domain. Cys-468 is in direct contact with bound DNA.
TABLE 2Galiellalactone-modified STAT3 peptides identified by mass spectrometryHighest mascot scores are given (several observations of each peptide were obtained). Cysteine residues that were modified are underlined in each peptide. The peptidescontaining both Cys-251 and Cys-259 were identified as both partial and fully galiellalactone-modified. The 686 –707 peptide containing galiellalactone modification onCys-687 was observed with and without phosphorylation on Tyr-705 as denoted with (p).
Amino acid Sequence Mascot Score Charge Experimental weight Theoretical weight
247–262 QQIACIGGPPNICLDR 23 2� 1890.9429 1890.9332247–262 QQIACIGGPPNICLDR 46 3� 2085.0223 2085.0275364–379 IKVCIDKDSGDVAALR 35 4� 1895.9943 1896.0026366–379 VCIDKDSGDVAALR 58 3� 1654.8102 1654.8236452–488 IDLETHSLPVVVISNICQMPNAWASILWYNMLTNNPK 90 4� 4418.1933 4418.2153455–488 ETHSLPVVVISNICQMPNAWASILWYNMLTNNPK 54 4� 4076.9493 4077.0202519–548 GLSIEQLTTLAEKLLGPGVNYSGCQITWAK 45 3� 3383.7763 3383.7687532–546 LLGPGVNYSGCQITW 72 2� 1800.8644 1800.8757532–548 LLGPGVNYSGCQITWAK 109 2� 1999.9970 2000.0077532–548 LLGPGVNYSGCQITWAK 78 3� 1999.9942 2000.0077539–548 YSGCQITWAK 40 2� 1349.6276 1349.6326686–707 YCRPESQEHPEADPGSAAPYLK 34 3� 2638.1902 2638.2009686–707 YCRPESQEHPEADPGSAAP(p)YLK 50 3� 2718.1540 2718.1673
Galiellalactone Is a Direct Inhibitor of STAT3
JUNE 6, 2014 • VOLUME 289 • NUMBER 23 JOURNAL OF BIOLOGICAL CHEMISTRY 15975
by guest on June 15, 2016http://w
ww
.jbc.org/D
ownloaded from

ment. The inhibitory effect on STAT3 activity was then mea-sured after an additional 4 h (Fig. 5A). The results show thatinhibition of STAT3 activity by galiellalactone persisted 4 hafter removal of galiellalactone from the cell medium. Further-more, the binding of GL-biot to STAT3 was not disrupted bythe addition of galiellalactone, indicating a strong and irrevers-ible binding to STAT3 (Fig. 5B).
DISCUSSION
The results indicate that the STAT3 signaling inhibitorgaliellalactone binds directly and covalently to STAT3 and thatthis interaction prevents DNA binding without interfering withupstream activation by phosphorylation. This further validatesgaliellalactone as a promising STAT3 inhibitor for the treat-ment of castration-resistant prostate cancer and other malig-nancies with constitutive activation of STAT3.
Galiellalactone is a reactive Michael acceptor that can reactwith both N-nucleophiles and S-nucleophiles including cys-teine. Nucleophiles add diastereoselectively to the double bondof the �,�-unsaturated lactone moiety giving inactive com-pounds, thus demonstrating the importance of the reactivefunctionality for the biological activity of galiellalactone. Herewe demonstrate that galiellalactone does form a stable adductwith cysteine. Galiellalactone is thought to covalently modifycysteine residues in the DNA binding region of STAT3, thuspreventing DNA binding. Here we saw that galiellalactonemodifies the cysteine residues Cys-367, Cys-468, and Cys-542in recombinant STAT3. Cys-542 is located in the linkerdomain, and Cys-367 and Cys-468 are located within the DNAbinding region of STAT3 (Fig. 4D). The linker domain may be
involved in DNA binding as well as transcriptional activity (27).The binding of galiellalactone to one or more of these cysteines,which are located in domains involved with DNA binding, mayprevent the binding of STAT3 to DNA. The corresponding cys-teine residues to the cysteines in STAT3, which were observedto be modified by galiellalactone, are not present in STAT1 andSTAT5 (28), possibly making this mode of action by galiellalac-tone selective for STAT3. Nevertheless, the effect of galiellalac-tone on STAT1 and STAT5 remains to be investigated.
Buettner et al. (23) demonstrated that the small moleculeSTAT3 inhibitor C48 alkylates Cys-468 in the STAT3 DNAbinding domain, although at a high compound concentration.Stattic, a STAT3 inhibitor that has been shown to preventSTAT3 dimerization and phosphorylation (29), was shown toalkylate four cysteine residues in unphosphorylated STAT3(Cys-251, Cys-259, Cys-367, and Cys-426) (24). Furthermore, ithas been shown that the transcriptional activity of STAT3 issensitive to changes in cellular redox conditions as treatment ofSTAT3 with hydrogen peroxide leads to covalent modifications(oxidation) of specific cysteine residues (25). Taken together,these studies, including this one, demonstrate that STAT3 transcrip-tional activity can be modulated by covalent modifications.
Strategies for the direct targeting of STAT3 include inhibi-tion of the STAT3 dimerization process by binding of smallmolecules or peptide mimetics to the SH2 domain and inhibit-ing STAT3 DNA binding with oligonucleotides (30). Thesestrategies have been thoroughly investigated and have pro-duced compounds with in vivo activity. Recently, it has beenshown that alkylation of reactive cysteine residues in STAT3 isa potential novel strategy for direct inhibiting of STAT3 func-tion. Alkylation of Cys-468 in the DNA binding domain by thecompound C48 is shown to prevent DNA binding, and methyl-2-cyano-3,12-dioxooleana-1,9(11)-dien-28-oate (CDDO-Me) pre-vented STAT3 dimerization by alkylation of Cys-259 (23, 31). Inaddition, the reactive compound Stattic inhibits STAT3 bybinding to multiple cysteine residues in unphosphorylatedSTAT3 as determined by mass spectrometry analysis. Thesefindings are in line with STAT3 signaling being sensitive toredox-dependent cysteine modifications (25, 32).
Galiellalactone shows a strong irreversible binding to STAT3and sustains inhibitory activity after the compound is removed,which could prolong the effect of the compound even after thecompound exposure is below the level of detection thus poten-tially disconnecting pharmacokinetics from pharmacodynam-ics. The long half-life of STAT3 (�8 h) indicates that STAT3activity is not regulated by protein degradation (33).
In addition to binding to STAT3, other binding proteinswere detected using GL-biot, which is not unexpected consid-ering the reactive nature of the inhibitor. The surprisingly smallnumber of additional target proteins, considering the reactivityof galiellalactone toward cysteine, was not identified in thisstudy. However, galiellalactone has been shown to inhibitNF-�B signaling by direct interaction with the NF-�B subunitp65 (34). Furthermore, galiellalactone has been observed toinhibit TGF-� signaling by preventing binding of SMAD2/3 toDNA (35). This demonstrates that galiellalactone is not specificfor STAT3, although the identification of STAT3 as a directbinding target correlates well with the inhibitory effects seen in
FIGURE 5. Irreversible binding of galiellalactone to STAT3. A, STAT3 lucif-erase reporter gene transfected LNCaP cells were treated with GL for 1 h andstimulated with IL-6 (50 ng/ml) for 4 h to induce STAT3 activity. For wash-out(wo) experiments, the cells were treated with GL for 1 h and washed with PBSbefore stimulation with IL-6. The STAT3-dependent luciferase activity isexpressed as the percentage of activation of relative control and presented asmean � S.E. from three separate experiments. B, DU145 cell lysates weretreated with 25 �M GL-biot for 1 h before the addition of increasing concen-trations (10 –100 �M) of GL for 1 h. Cell lysates were incubated with streptavi-din-Sepharose beads, and elutions were immunoblotted for STAT3. GL-biotwas not displaced by GL, indicating a strong binding to STAT3.
Galiellalactone Is a Direct Inhibitor of STAT3
15976 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 289 • NUMBER 23 • JUNE 6, 2014
by guest on June 15, 2016http://w
ww
.jbc.org/D
ownloaded from

galiellalactone-treated DU145 cells expressing constitutivelyactive STAT3. In addition, galiellalactone has been shown to bewell tolerated in animal studies with repeat dosing and displaysminimal cytotoxic effects in non-STAT3-dependent cells,which would not be expected with a non-discriminate andhighly reactive compound (20).
Considering the multitude of STAT3 activation mechanismsin malignancies and the risk of developing therapy resistance tokinase inhibitors, it is in theory desirable to develop inhibitorsacting directly on STAT3. Direct targeting of STAT3 instead ofupstream activators of STAT3 such as tyrosine kinases maylead to a decrease in side effects and increased efficacy. Further-more, direct targeting of STAT3 circumvents the nature ofupstream activation pathway. Here we show that galiellalactoneexerts its inhibitory actions by direct interaction with STAT3.
STAT3 may have different roles in the various stages of can-cer and cancer development with different activation modes ofSTAT3 (phosphorylation of pSTAT3 Tyr-705 or Ser-727) (4).The activation of STAT3 by phosphorylation of Tyr-705 or Ser-727 induces the transcription of different subsets of genes. Thebinding of galiellalactone to STAT3 was independent of phos-phorylation status and bound to both phosphorylated andunphosphorylated STAT3. Unphosphorylated STAT3 hasbeen shown to regulate the expression of genes involved inoncogenesis (36). Binding of galiellalactone to unphosphory-lated STAT3 in prostate cancer cells may be an additionalway to prevent tumor progression; however, the role ofunphosphorylated STAT3 in prostate cancer has not yetbeen evaluated.
Here we demonstrate with chemical and molecular pharma-cological methods that the reactive fungal metabolite galiella-lactone is a cysteine reactive inhibitor that covalently binds toone or more cysteines in STAT3 and that this leads to inhibitionof STAT3 binding to DNA and thus blocks STAT3 signalingwithout affecting tyrosine phosphorylation. Considering thedifficulty in targeting transcription factors with small, drug-likemolecules and the rekindled interest in covalent inhibitors, thisstudy further validates galiellalactone as a promising chemicalstarting point for designing highly specific STAT3 inhibitorsand as a promising STAT3 inhibitor in itself for the treatmentof castration-resistant prostate cancer and other malignancieswith constitutive activation of STAT3.
Acknowledgments—We thank Annika Rogstam and the Lund ProteinProduction Platform at Lund University, Sweden for production ofrecombinant STAT3 protein.
REFERENCES1. Calo, V., Migliavacca, M., Bazan, V., Macaluso, M., Buscemi, M., Gebbia,
N., and Russo, A. (2003) STAT proteins: from normal control of cellularevents to tumorigenesis. J. Cell Physiol. 197, 157–168
2. Barre, B., Vigneron, A., Perkins, N., Roninson, I. B., Gamelin, E., and Co-queret, O. (2007) The STAT3 oncogene as a predictive marker of drugresistance. Trends Mol. Med. 13, 4 –11
3. Peyser, N. D., and Grandis, J. R. (2013) Critical analysis of the potential fortargeting STAT3 in human malignancy. Onco. Targets Ther. 6, 999 –1010
4. Sellier, H., Rebillard, A., Guette, C., Barre, B., and Coqueret, O. (2013)How should we define STAT3 as an oncogene and as a potential target fortherapy? JAKSTAT 2, e24716
5. Quintas-Cardama, A., and Verstovsek, S. (2013) Molecular pathways: Jak/STAT pathway: mutations, inhibitors, and resistance. Clin. Cancer Res.19, 1933–1940
6. Sansone, P., and Bromberg, J. (2012) Targeting the interleukin-6/Jak/statpathway in human malignancies. J. Clin. Oncol. 30, 1005–1014
7. Koskela, H. L., Eldfors, S., Ellonen, P., van Adrichem, A. J., Kuusanmaki,H., Andersson, E. I., Lagstrom, S., Clemente, M. J., Olson, T., Jalkanen,S. E., Majumder, M. M., Almusa, H., Edgren, H., Lepisto, M., Mattila, P.,Guinta, K., Koistinen, P., Kuittinen, T., Penttinen, K., Parsons, A.,Knowles, J., Saarela, J., Wennerberg, K., Kallioniemi, O., Porkka, K.,Loughran, T. P., Jr., Heckman, C. A., Maciejewski, J. P., and Mustjoki, S.(2012) Somatic STAT3 mutations in large granular lymphocytic leukemia.N. Engl. J. Med. 366, 1905–1913
8. Pilati, C., Amessou, M., Bihl, M. P., Balabaud, C., Nhieu, J. T., Paradis, V.,Nault, J. C., Izard, T., Bioulac-Sage, P., Couchy, G., Poussin, K., and Zuc-man-Rossi, J. (2011) Somatic mutations activating STAT3 in human in-flammatory hepatocellular adenomas. J. Exp. Med. 208, 1359 –1366
9. Ferlay, J., Shin, H. R., Bray, F., Forman, D., Mathers, C., and Parkin, D. M.(2010) Estimates of worldwide burden of cancer in 2008: GLOBOCAN2008. Int. J. Cancer 127, 2893–2917
10. Mohler, J. L., Armstrong, A. J., Bahnson, R. R., Boston, B., Busby, J. E.,D’Amico, A. V., Eastham, J. A., Enke, C. A., Farrington, T., Higano, C. S.,Horwitz, E. M., Kantoff, P. W., Kawachi, M. H., Kuettel, M., Lee, R. J.,MacVicar, G. R., Malcolm, A. W., Miller, D., Plimack, E. R., Pow-Sang,J. M., Roach, M., 3rd, Rohren, E., Rosenfeld, S., Srinivas, S., Strope, S. A.,Tward, J., Twardowski, P., Walsh, P. C., Ho, M., and Shead, D. A. (2012)Prostate cancer, Version 3.2012: featured updates to the NCCN guide-lines. J. Natl. Compr. Canc. Netw. 10, 1081–1087
11. Sridhar, S. S., Freedland, S. J., Gleave, M. E., Higano, C., Mulders, P.,Parker, C., Sartor, O., and Saad, F. (2014) Castration-resistant prostatecancer: from new pathophysiology to new treatment. Eur. Urol. 65,289 –299
12. Horinaga, M., Okita, H., Nakashima, J., Kanao, K., Sakamoto, M., andMurai, M. (2005) Clinical and pathologic significance of activation of sig-nal transducer and activator of transcription 3 in prostate cancer. Urology66, 671– 675
13. Mora, L. B., Buettner, R., Seigne, J., Diaz, J., Ahmad, N., Garcia, R., Bow-man, T., Falcone, R., Fairclough, R., Cantor, A., Muro-Cacho, C., Living-ston, S., Karras, J., Pow-Sang, J., and Jove, R. (2002) Constitutive activationof Stat3 in human prostate tumors and cell lines: direct inhibition of Stat3signaling induces apoptosis of prostate cancer cells. Cancer Res. 62,6659 – 6666
14. Tam, L., McGlynn, L. M., Traynor, P., Mukherjee, R., Bartlett, J. M., andEdwards, J. (2007) Expression levels of the JAK/STAT pathway in thetransition from hormone-sensitive to hormone-refractory prostate can-cer. Br. J. Cancer 97, 378 –383
15. Abdulghani, J., Gu, L., Dagvadorj, A., Lutz, J., Leiby, B., Bonuccelli, G.,Lisanti, M. P., Zellweger, T., Alanen, K., Mirtti, T., Visakorpi, T., Buben-dorf, L., and Nevalainen, M. T. (2008) Stat3 promotes metastatic progres-sion of prostate cancer. Am. J. Pathol. 172, 1717–1728
16. Kroon, P., Berry, P. A., Stower, M. J., Rodrigues, G., Mann, V. M., Simms,M., Bhasin, D., Chettiar, S., Li, C., Li, P. K., Maitland, N. J., and Collins,A. T. (2013) JAK-STAT blockade inhibits tumor initiation and clonogenicrecovery of prostate cancer stem-like cells. Cancer Res. 73, 5288 –5298
17. Shodeinde, A. L., and Barton, B. E. (2012) Potential use of STAT3 inhibi-tors in targeted prostate cancer therapy: future prospects. Onco. TargetsTher. 5, 119 –125
18. Turkson, J. (2004) STAT proteins as novel targets for cancer drug discov-ery. Expert Opin. Ther. Targets 8, 409 – 422
19. Weidler, M., Rether, J., Anke, T., and Erkel, G. (2000) Inhibition of inter-leukin-6 signaling by galiellalactone. FEBS Lett. 484, 1– 6
20. Hellsten, R., Johansson, M., Dahlman, A., Dizeyi, N., Sterner, O., and Bjar-tell, A. (2008) Galiellalactone is a novel therapeutic candidate against hor-mone-refractory prostate cancer expressing activated Stat3. Prostate 68,269 –280
21. Hellsten, R., Johansson, M., Dahlman, A., Sterner, O., and Bjartell, A.(2011) Galiellalactone inhibits stem cell-like ALDH-positive prostate can-cer cells. PLoS One 6, e22118
Galiellalactone Is a Direct Inhibitor of STAT3
JUNE 6, 2014 • VOLUME 289 • NUMBER 23 JOURNAL OF BIOLOGICAL CHEMISTRY 15977
by guest on June 15, 2016http://w
ww
.jbc.org/D
ownloaded from

22. von Nussbaum, F., Hanke, R., Fahrig, T., and Benet-Buchholz, J. (2004)The high-intrinsic Diels-Alder reactivity of (�)-galiellalactone: generat-ing four quaternary carbon centers under mild conditions. Eur. J. OrganicChem. 2004, 2783–2790
23. Buettner, R., Corzano, R., Rashid, R., Lin, J., Senthil, M., Hedvat, M., Schr-oeder, A., Mao, A., Herrmann, A., Yim, J., Li, H., Yuan, Y. C., Yakushijin,K., Yakushijin, F., Vaidehi, N., Moore, R., Gugiu, G., Lee, T. D., Yip, R.,Chen, Y., Jove, R., Horne, D., and Williams, J. C. (2011) Alkylation ofcysteine 468 in Stat3 defines a novel site for therapeutic development. ACSChem. Biol. 6, 432– 443
24. Heidelberger, S., Zinzalla, G., Antonow, D., Essex, S., Basu, B. P., Palmer, J.,Husby, J., Jackson, P. J., Rahman, K. M., Wilderspin, A. F., Zloh, M., andThurston, D. E. (2013) Investigation of the protein alkylation sites of theSTAT3:STAT3 inhibitor Stattic by mass spectrometry. Bioorg. Med.Chem. Lett. 23, 4719 – 4722
25. Li, L., Cheung, S. H., Evans, E. L., and Shaw, P. E. (2010) Modulation ofgene expression and tumor cell growth by redox modification of STAT3.Cancer Res. 70, 8222– 8232
26. Johansson, M., and Sterner, O. (2001) Synthesis of (�)-galiellalactone:absolute configuration of galiellalactone. Org. Lett. 3, 2843–2845
27. Lim, C. P., and Cao, X. (2006) Structure, function, and regulation of STATproteins. Mol. Biosyst. 2, 536 –550
28. Becker, S., Groner, B., and Muller, C. W. (1998) Three-dimensional struc-ture of the Stat3� homodimer bound to DNA. Nature 394, 145–151
29. Schust, J., Sperl, B., Hollis, A., Mayer, T. U., and Berg, T. (2006) Stattic: asmall-molecule inhibitor of STAT3 activation and dimerization. Chem.Biol. 13, 1235–1242
30. Yue, P., and Turkson, J. (2009) Targeting STAT3 in cancer: how successfulare we? Expert Opin. Investig. Drugs 18, 45–56
31. Ahmad, R., Raina, D., Meyer, C., and Kufe, D. (2008) Triterpenoid CDDO-methyl ester inhibits the Janus-activated kinase-1 (JAK1)3signal trans-ducer and activator of transcription-3 (STAT3) pathway by direct inhibi-tion of JAK1 and STAT3. Cancer Res. 68, 2920 –2926
32. Zgheib, C., Kurdi, M., Zouein, F. A., Gunter, B. W., Stanley, B. A., Zgheib,J., Romero, D. G., King, S. B., Paolocci, N., and Booz, G. W. (2012) Acyloxynitroso compounds inhibit LIF signaling in endothelial cells and cardiacmyocytes: evidence that STAT3 signaling is redox-sensitive. PLoS One 7,e43313
33. Heinrich, P. C., Behrmann, I., Muller-Newen, G., Schaper, F., and Graeve,L. (1998) Interleukin-6-type cytokine signalling through the gp130/Jak/STAT pathway. Biochem. J. 334, 297–314
34. Perez, M., Soler-Torronteras, R., Collado, J. A., Limones, C. G., Hellsten,R., Johansson, M., Sterner, O., Bjartell, A., Calzado, M. A., and Munoz, E.(2014) The fungal metabolite galiellalactone interferes with the nuclearimport of NF-�B and inhibits HIV-1 replication. Chem. Biol. Interact. 214,69 –76
35. Rudolph, K., Serwe, A., and Erkel, G. (2013) Inhibition of TGF-� signalingby the fungal lactones (S)-curvularin, dehydrocurvularin, oxacyclodo-decindione and galiellalactone. Cytokine 61, 285–296
36. Yang, J., Chatterjee-Kishore, M., Staugaitis, S. M., Nguyen, H., Sch-lessinger, K., Levy, D. E., and Stark, G. R. (2005) Novel roles of unphos-phorylated STAT3 in oncogenesis and transcriptional regulation. CancerRes. 65, 939 –947
Galiellalactone Is a Direct Inhibitor of STAT3
15978 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 289 • NUMBER 23 • JUNE 6, 2014
by guest on June 15, 2016http://w
ww
.jbc.org/D
ownloaded from

Garcia, Eduardo Munoz, Olov Sterner, Anders Bjartell and Rebecka HellstenNicholas Don-Doncow, Zilma Escobar, Martin Johansson, Sven Kjellström, Victor
Cancer CellsGaliellalactone Is a Direct Inhibitor of the Transcription Factor STAT3 in Prostate
doi: 10.1074/jbc.M114.564252 originally published online April 22, 20142014, 289:15969-15978.J. Biol. Chem.
10.1074/jbc.M114.564252Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/289/23/15969.full.html#ref-list-1
This article cites 36 references, 10 of which can be accessed free at
by guest on June 15, 2016http://w
ww
.jbc.org/D
ownloaded from




















