
Rapid activation of the Stat3 transcription complex in liver regeneration
7
Rapid Activation of the Complex in Liver Stat3 Transcription Regeneration DREW E. CRESSMAN, 1 ROBERT H. DIAMOND, 1'2 AND REBECCA TAUB L3 Liver regeneration in response to partial hepatectomy is a physiological growth response observed in the intact ~nimal. Understanding the early signals that trigger liver regeneration is of vital importance to understand the liver's response to injury. It has been observed that several growth factors and cytokines, including epider- mal growth factor (EGF) and interleukin-6 (IL-6), can activate members of the signal transducers and activa- tors of transcription (Stat) family of transcription fac- tors resulting in tyrosine phosphorylation of these fac- tors, nuclear tr~n~location, and an active DNA binding transcriptional complex. Because Stat3 participates in the regulation of primary growth response genes, we wondered if it is induced in the early phase of liver re- generation. We found that Stat3 DNA-binding activity is increased in the remnant liver within 30 minutes of par- tial hepatectomy and peaks at more than 30-fold at 3 hours. This induction is not observed after sham sur- gery. The induction of Stat3 appears to be part of the initial response of the remnant liver to partial hepatec- tomy, because it occurs in the presence of cyclohexi- mide-mediated protein synthesis blockade. Activation of Stat3 is unusual, because it extends beyond the immedio ate-early time period and remains near peak level at 5 hours posthepatectomy. Although insulin-treated H35 cells activate many of the same immediate-early genes as regenerating liver, Stat3 is not induced in these cells. Because Stat factors are known to be inactivated by pro- tein tyrosine phosphatases (PTPase), we showed that a PTPase is able to eliminate the DNA binding of hepatic Stat3. It is likely that Stat3 contributes to the transcrip- tional activation of a subset of immediate-early genes that are induced over a prolonged time in the G1 phase of hepatic cells following partial hepatectomy. The iden- tification of Stat3 as an early factor in liver regeneration provides clues to the activation of signal transduction pathways in the remnant liver within the first minutes Abbreviations: EGF, epidermal growth factor; IL-6, interleukin-6; IKB-a, inhibitor of KB-a; NF-KB, nuclear factor KB; EMSA, electrophoretic gel shil~ assay; PTPase, protein tyrosine phosphatase; YOP, yersinia protein tyrosine phosphatase; PRL-1, phosphatase of regenerating liver. From the 1Department of Genetics, 2Division of Gastroenterology, and the 3Department of Medicine, Howard Hughes Medical Institute, University of Pennsylvania School of Medicine, Philadelphia, PA. Received September 12, 1994; accepted December 7, 1994. Address reprint requests to Rebecca Taub, MD, Department of Genetics and Medicine, Howard Hughes Medical Institute, University of Pennsylvania School of Medicine, Philadelphia, PA 19104-6145. Copyright © 1995 by the American Association for the Study of Liver Diseases. 0270-9139/95/2105-003053.00/0 to several hours after partial hepatectomy. (HEPATOLOGY 1995;21:1443-1449.) Within minutes after a two-thirds partial hepatec- tomy, the majority of cells in the remnant liver that are normally quiescent rapidly reenter the cell cycle. DNA synthesis initially takes place at 12 to 16 hours posthepatectomy and peaks at 24 hours, and restora- tion of the original liver mass occurs in 7 to 10 days. Several different growth factors have been implicated as having a role in this process including hepatocyte growth factor, 1 epidermal growth factor (EGF), trans- forming growth factor, insulin, glucagon, and the cyto- kines interleukin-6 (IL-6) and tumor necrosis factor- a, but the exact signals that trigger liver regeneration have remained elusive. TM The identification of primary or immediate-early growth response genes that are activated in the rem- nant liver within minutes of the partial hepatectomy in a protein synthesis-independent manner can help identify the earliest signals in the regenerating liver. 5~2 We have identified more than 70 immediate-early genes that are activated in the regenerating liver and defined the temporal course of their induction. ~3 We can then ask what transcription factors are responsible for activating the transcription of these immediate- early genes. For instance, identifying inhibitor of KB- (IKB-a) as an immediate-early gene in regenerating liver allowed us to search for the transcription factor complex nuclear factor-KB (NF-KB) that is known to be responsible for its activation. We found that posthepa- tectomy factor/NF-KB, a DNA-binding factor comprised of NF-KB subunits, is activated within minutes of the partial hepatectomy in a protein synthesis-indepen- dent fashion, and is likely to be responsible for trans- activation of a subset of immediate-early genes includ- ing IKB-a and others. 1417 Recently, the Stat (signal transducers and activators of transcription) family of transcription factors have been identified, ls-27 Stat proteins, which contain both the signal transduction domains, SH2 and SH3, and a DNA-binding region, are activated by a number of cytokines and growth factors. The intracellular signal generated by growth factor or cytokine action causes tyrosine phosphorylation of quiescent cytoplasmic Stat factors. This tyrosine phosphorylation may be medi- ated by the Janus kinase family or the tyrosine kinase 1443
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Rapid activation of the Stat3 transcription complex in liver regeneration

Rapid Activation of the Complex in Liver
Stat3 Transcription Regeneration
DREW E. CRESSMAN, 1 ROBERT H. DIAMOND, 1'2 AND REBECCA TAUB L3
Liver regenerat ion in re sponse to partial h e p a t e c t o m y is a phys io log ica l g r o w t h r e s p o n s e observed in the intact ~nimal. U n d e r s t a n d i n g the early s ignals that trigger l iver regenerat ion is o f vital i m p o r t a n c e to unders tand the liver's re sponse to injury. It has b e e n observed that several growth factors and cytokines , inc luding epider- mal g r o w t h factor (EGF) and inter leukin-6 (IL-6), can act ivate m e m b e r s o f the s ignal t ransducers and activa- tors o f t ranscr ipt ion (Stat) fami ly o f transcr ipt ion fac- tors resul t ing in tyros ine phosphory la t i on o f these fac- tors, nuc lear tr~n~locat ion, and an act ive DNA b inding transcr ipt ional complex . B e c a u s e Stat3 part ic ipates in the regu la t ion o f pr imary g r o w t h re sponse genes , w e w o n d e r e d if it is induced in the early phase of l iver re- generat ion . We found that Stat3 DNA-binding act iv i ty is increased in the r e m n a n t l iver wi th in 30 m i n u t e s of par- t ial h e p a t e c t o m y and peaks at m o r e than 30-fold at 3 hours . This induc t ion is no t observed after s h a m sur- gery. The induct ion o f Stat3 appears to be part o f the init ial r e s p o n s e of the r e m n a n t l iver to partial hepatec- tomy, because it occurs in the presence of cyclohexi- mide -media ted prote in synthes i s b lockade. Act ivat ion o f Stat3 is unusual , because it ex tends b e y o n d the immedio ate-early t ime per iod and remains near p e a k level at 5 hours pos thepatec tomy . A l though insul in- treated H35 cells act ivate m a n y o f the s a m e immediate -ear ly genes as regenera t ing liver, Stat3 is no t induced in these cells. B e c a u s e Stat factors are k n o w n to be inact ivated by pro- te in tyros ine p h o s p h a t a s e s (PTPase), w e s h o w e d that a PTPase is able to e l iminate the DNA b inding o f hepat ic Stat3. It is l ikely that Stat3 contr ibutes to the transcrip- t ional ac t ivat ion of a subset o f immediate -ear ly genes that are induced over a pro longed t ime in the G1 phase o f hepat ic cel ls fo l lowing part ial hepatec tomy. The iden- t i f icat ion o f Stat3 as an early factor in l iver regenerat ion provides c lues to the act ivat ion o f s ignal t ransduct ion p a t h w a y s in the r e m n a n t l iver w i th in the first minute s
Abbreviations: EGF, epidermal growth factor; IL-6, interleukin-6; IKB-a, inhibitor of KB-a; NF-KB, nuclear factor KB; EMSA, electrophoretic gel shil~ assay; PTPase, protein tyrosine phosphatase; YOP, yersinia protein tyrosine phosphatase; PRL-1, phosphatase of regenerating liver.
From the 1Department of Genetics, 2Division of Gastroenterology, and the 3Department of Medicine, Howard Hughes Medical Institute, University of Pennsylvania School of Medicine, Philadelphia, PA.
Received September 12, 1994; accepted December 7, 1994. Address reprint requests to Rebecca Taub, MD, Department of Genetics
and Medicine, Howard Hughes Medical Institute, University of Pennsylvania School of Medicine, Philadelphia, PA 19104-6145.
Copyright © 1995 by the American Association for the Study of Liver Diseases.
0270-9139/95/2105-003053.00/0
to several hours after partial hepatec tomy. (HEPATOLOGY 1995;21:1443-1449.)
Within minutes after a two-thirds partial hepatec- tomy, the majority of cells in the remnant liver that are normally quiescent rapidly reenter the cell cycle. DNA synthesis initially takes place at 12 to 16 hours posthepatectomy and peaks at 24 hours, and restora- tion of the original liver mass occurs in 7 to 10 days. Several different growth factors have been implicated as having a role in this process including hepatocyte growth factor, 1 epidermal growth factor (EGF), trans- forming growth factor, insulin, glucagon, and the cyto- kines interleukin-6 (IL-6) and tumor necrosis factor- a, but the exact signals that trigger liver regeneration have remained elusive. TM
The identification of primary or immediate-early growth response genes that are activated in the rem- nant liver within minutes of the partial hepatectomy in a protein synthesis-independent manner can help identify the earliest signals in the regenerating liver. 5~2 We have identified more than 70 immediate-early genes that are activated in the regenerating liver and defined the temporal course of their induction. ~3 We can then ask what transcription factors are responsible for activating the transcription of these immediate- early genes. For instance, identifying inhibitor of KB-
(IKB-a) as an immediate-early gene in regenerating liver allowed us to search for the transcription factor complex nuclear factor-KB (NF-KB) that is known to be responsible for its activation. We found that posthepa- tectomy factor/NF-KB, a DNA-binding factor comprised of NF-KB subunits, is activated within minutes of the partial hepatectomy in a protein synthesis-indepen- dent fashion, and is likely to be responsible for trans- activation of a subset of immediate-early genes includ- ing IKB-a and others. 1417
Recently, the Stat (signal transducers and activators of transcription) family of transcription factors have been identified, ls-27 Stat proteins, which contain both the signal transduction domains, SH2 and SH3, and a DNA-binding region, are activated by a number of cytokines and growth factors. The intracellular signal generated by growth factor or cytokine action causes tyrosine phosphorylation of quiescent cytoplasmic Stat factors. This tyrosine phosphorylation may be medi- ated by the Janus kinase family or the tyrosine kinase
1443

1444 CRESSMAN, DIAMOND, AND TAUB HEPATOLOGY May 1995
receptors themselves. The activated Stat transcription factors then translocate into the nucleus, where they are able to bind to specific DNA-sequence elements and activate the transcription of a number of target genes. DNA binding is dependent on tyrosine phosphoryla- tion. Currently, a number of Stat proteins have been identified including S ta t l (c~ and/~), 91-kd and 84-kd isoforms that are activated by interferon a and inter- feron-T; Stat2, a l l3 -kD distinct protein that is acti- vated by IFN-a; and most recently, Stat3, a 92-kd pro- tein tha t is activated by EGF and IL-6 but not by the interferons. 26'27 Stat3 has highest affinity for an ele- ment (serum inducible element) in the c-fos promoter and may be in par t responsible for activation of the c- fos gene during mitogen stimulation mediated by EGF and other growth factors. 26-3° Trea tment of mice with EGF induces a complex indistinguishable from the Stat3 homodimer in the liver. Because of the hypothe- sis tha t EGF and IL-6 may play a role in liver regenera- tion 1-3 and the finding tha t immediate-early genes tha t may contain Star promoter elements are activated in the regenerat ing liver, we examined regenerat ing liver for the activation of Stat transcription factors.
MATERIALS AND METHODS
Animals and Cells. Adult female Fischer rats (160 to 200 g) were anesthetized with ether and subjected to midventral laparotomy with approximately 70% liver restriction (left lat- eral and median lobes) using protocols approved by the Insti- tutional Animal Care and Use Committee, University of Pennsylvania, Philadelphia, PA. For cycloheximide-treated samples, rats were pretreated 15 minutes before the laparot- omy with 50 mg of cycloheximide per kg of body weight (5% solution in phosphate-buffered saline) intraperitoneally. Nu- clear extracts were prepared from the remaining liver lobes at the indicated times after partial hepatectomy, as pre- viously described. 14'31 Extracts were prepared in the presence of 2 pg/mL of antipain, aprotinin, bestatin, and leupeptin (Boehringer Mannheim, Indianapolis, IN). A-431 cells were grown and treated with EGF before preparation of nuclear extract. 26 H35 cells were grown as described by Taub et a132 and were serum deprived for 3 days before treatment with 10 s mol/L of insulin and before preparation of nuclear ex- tracts.
Electrophoretic Gel Shift Assays. Binding reactions were performed essentially as described by Tewari et al, ~4 using nuclear extracts from posthepatectomy rat liver cells or A- 431 human epidermoid carcinoma cells. All experiments were performed using an excess of probe. Four micrograms of ex- tract were incubated with radiolabeled oligonucleotide (1 ng) for 20 minutes at 30°C in binding buffer (10 mmol/L of N-[2- hydroxyethyl]piperazine-N'-[2-ethanesulfonic acid], pH 7.9/ 50 mmol/L of NaC1/1 mmol/L ethylenediaminetetraacetic acid/10% glycerol) and electrophoresed on non-denaturing 5% polyacrylamide gels in 0.5× TBE buffer (44 mmo]/L Tris/ 44 mmol/L boric acid/1 mmol/L ethylenediaminetetraacetic acid). The gels were dried and exposed to radiograph film. One microgram of poly(dI-dC) was used as a nonspecific DNA competitor in each reaction. The probe used was a prean- nealed high-performance liquid chromatography-purified double-stranded oligonucleotide from the serum-inducible- factor binding element in the c-fos promoter (GATCCTCCA- GCATTTCCCGTAAATCCTCCAG) 3° and end-labeled with
[T-32P]ATP. One-hundredfold excess of unlabeled oligonucleo- tide was incubated with extracts for 20 minutes at 30°C be- fore the addition of the radiolabeled probe in cold competition experiments.
Phosphatase Assays. As described in previous studies, 33'34 p-nitrophenylphosphate assays were performed. Four micro- grams of regenerating liver nuclear extracts were incubated with 1 #g of purified protein tyrosine phosphatase (PTPase) protein in phosphatase buffer (50 mmol/L N-[2-hydroxyethyl]- piperazine-N'-[2-ethanesulfonic acid]; pH 7.9/0.1%//%Mer- captoethanol) at 37°C for 20 minutes. Binding buffer was then added, and extracts were incubated at 30°C for 20 min- utes and electrophoretic gel shift assays (EMSAs) were car- ried out as described above. Purified protein preparations used were wild-type prolactin, mutant prolactin in which a cysteine in the active site (amino acid 104) is converted to a serine, control protein IKB-a, an inhibitor of the DNA-binding activity of the NF-KB transcription factor, and control protein YOP phosphatase (New England BioLabs, Beverly, MA), a 51-kd tyrosine phosphatase from the Escherichia coli strain Yersinia enterocolitica.
RESULTS
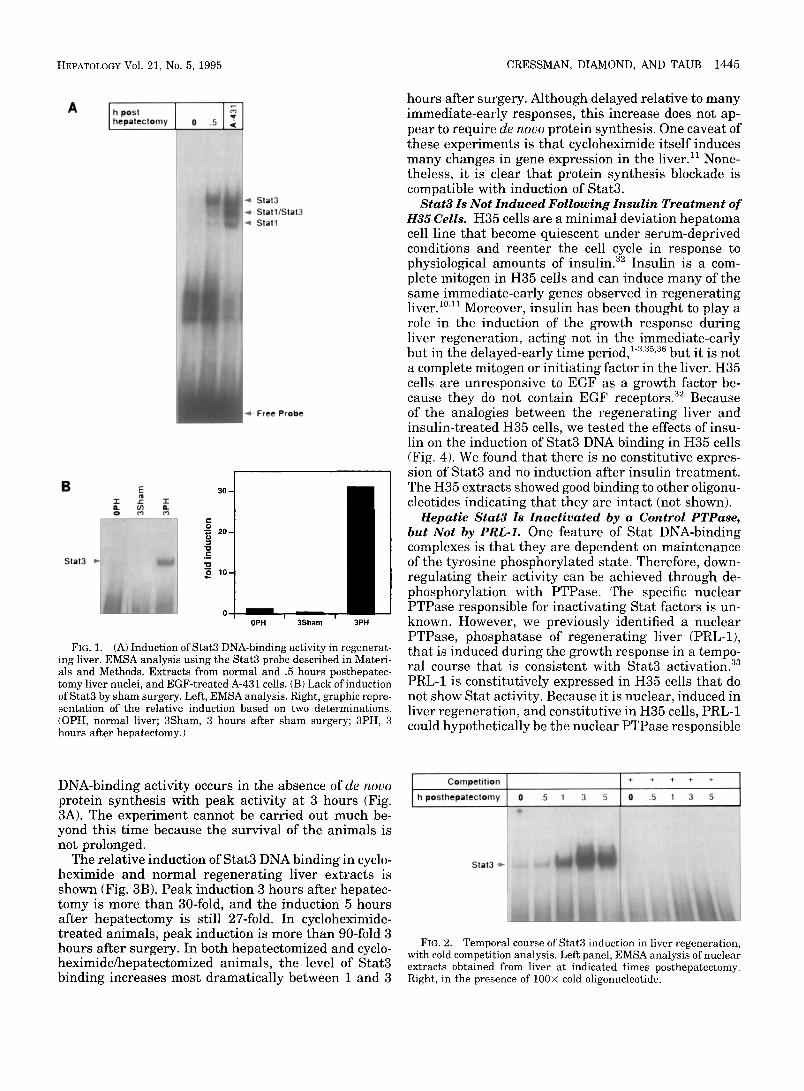
Identification of Induced Stat DNA-Binding Activity in Regenerating Liver. Stat nuclear factors from EGF- st imulated A-431 cells tha t bind to the high-affinity binding site, human serum inducible element, have been well characterized. 26 The upper complex contains a Stat3 homodimer, the middle complex contains a Sta t3 /Sta t l heterodimer, and the lower complex con- tains a S ta t l homodimer. As shown in Fig. 1A, liver nuclear extracts 30 minutes after hepatectomy contain an induced complex that comigrates exactly with the Stat3 homodimer. There is little induction of the lower complexes, and a low level of the complex correspond- ing to the S ta t l homodimer appears to be constitutive. Based on the precise comigration of these complexes with the A-431 complexes, we will refer to the induced complex as Stat3, assuming that it is the Stat3 homodi- mer. In support of this, prel iminary studies with anti- Stat3 antibodies disrupt and supershift this complex. As shown in Fig. 1B, there is no induction of Stat3 by sham surgery 3 hours after surgery. Densitometry shows the relative induction in the 3 hours after hepa- tectomy and the 3 hours after sham compared with normal liver.
We examined the temporal course and specificity of this induction. As shown, peak Stat3 activity occurs 3 hours after hepatectomy, and the level remains ele- vated even 5 hours after hepatectomy (Fig. 2). All com- plexes compete completely with cold oligonucleotide, an indication that they are specific.
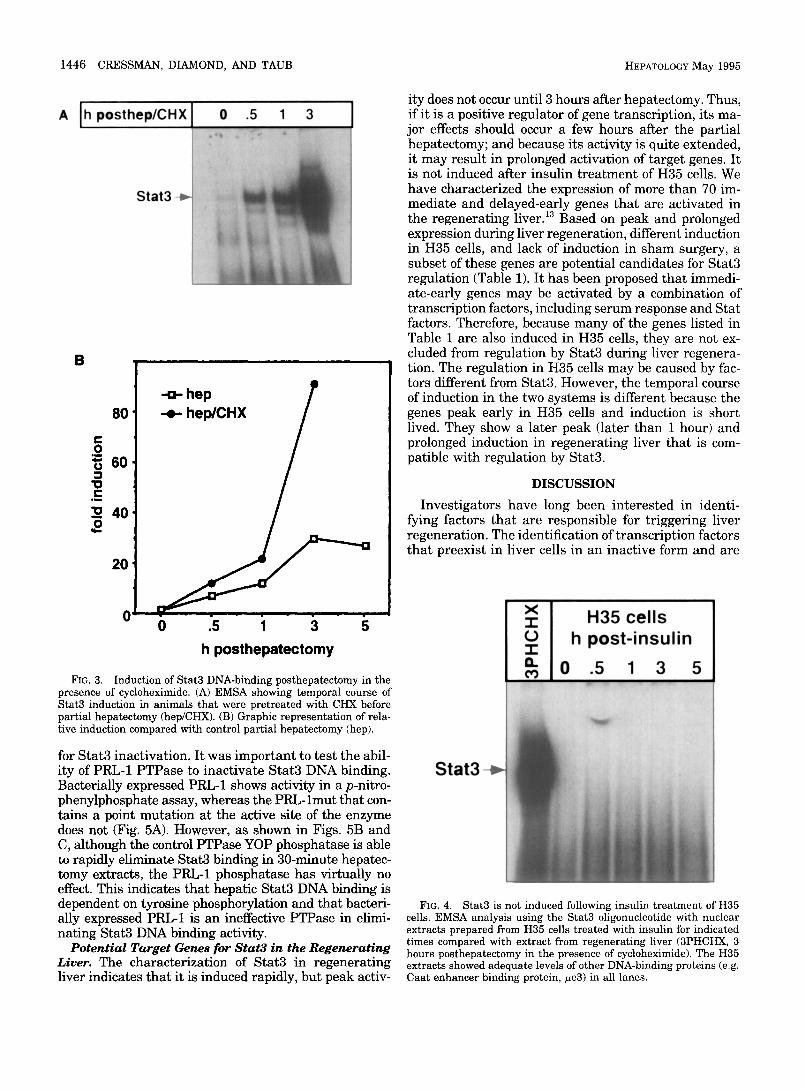
Star3 is Induced in the Absence of de Novo Protein Synthesis. The hal lmark of the pr imary growth re- sponse is its occurrence in the absence of new protein synthesis. Protein synthesis can be blocked by treat ing animals with cycloheximide. Confirmation of protein synthesis blockage by cycloheximide in these extracts was established by determining the level of IKB-a com- pared with control levels. 1~ In these extracts, the level of IKB-c~ decreased rapidly because of the short half- life of this protein. As shown, the induction of Stat3
HEPATOLOGY Vol. 21, No. 5, 1995 CRESSMAN, DIAMOND, AND TAUB 1445
A I h post [ hepatectomy
Stat3
=robe
B E O. ¢/) O
3 0 - - r O.
p,
O 2 0 -
I 0 ._c "o o ~ 0 -
0PH 3Sham 3PH
FIG. 1. (A) Induction of Star3 DNA*binding activity in regenerat- ing liver. EMSA analysis using the Stat3 probe described in Materi- als and Methods. Extracts from normal and .5 hours posthepatec- tomy liver nuclei, and EGF-treated A-431 cells. (B) Lack of induction of Stat3 by sham surgery. Left, EMSA analysis. Right, graphic repre- sentation of the relative induction based on two determinations. (OPH, normal liver; 3Sham, 3 hours after sham surgery; 3PH, 3 hours after hepatectomy.)
hours after surgery. Although delayed relative to many immediate-early responses, this increase does not ap- pear to require de novo protein synthesis. One caveat of these experiments is that cycloheximide itself induces many changes in gene expression in the liver. 11 None- theless, it is clear that protein synthesis blockade is compatible with induction of Stat3.
Star3 Is Not Induced Following Insul in Trea tment of t i35 Cells. H35 cells are a minimal deviation hepatoma cell line that become quiescent under serum-deprived conditions and reenter the cell cycle in response to physiological amounts of insulin. 32 Insulin is a com- plete mitogen in H35 cells and can induce many of the same immediate-early genes observed in regenerating liver. 1°~11 Moreover, insulin has been thought to play a role in the induction of the growth response during liver regeneration, acting not in the immediate-early but in the delayed-early time period, 1-3':~'3~ but it is not a complete mitogen or initiating factor in the liver. H35 cells are unresponsive to EGF as a growth factor be- cause they do not contain EGF receptors. 32 Because of the analogies between the regenerating liver and insulin-treated H35 cells, we tested the effects of insu- lin on the induction of Stat3 DNA binding in H35 cells (Fig. 4). We found that there is no constitutive expres- sion of Stat3 and no induction after insulin treatment. The H35 extracts showed good binding to other oligonu- cleotides indicating that they are intact (not shown).
Hepat ic Stat3 Is Inac t i va ted by a Control PTPase, but Not by PRL-1. One feature of Stat DNA-binding complexes is that they are dependent on maintenance of the tyrosine phosphorylated state. Therefore, down- regulating their activity can be achieved through de- phosphorylation with PTPase. The specific nuclear PTPase responsible for inactivating Stat factors is un- known. However, we previously identified a nuclear PTPase, phosphatase of regenerating liver (PRL-1), that is induced during the growth response in a tempo- ral course that is consistent with Stat3 activation. 33 PRL-1 is constitutively expressed in H35 cells that do not show Stat activity. Because it is nuclear, induced in liver regeneration, and constitutive in H35 cells, PRL-1 could hypothetically be the nuclear PTPase responsible
DNA-binding activity occurs in the absence of de novo protein synthesis with peak activity at 3 hours (Fig. 3A). The experiment cannot be carried out much be- yond this time because the survival of the animals is not prolonged.
The relative induction of Stat3 DNA binding in cyclo- heximide and normal regenerating liver extracts is shown (Fig. 3B). Peak induction 3 hours after hepatec- tomy is more than 30-fold, and the induction 5 hours after hepatectomy is still 27-fold. In cycloheximide- t reated animals, peak induction is more than 90-fold 3 hours after surgery. In both hepatectomized and cyclo- heximide/hepatectomized animals, the level of Stat3 binding increases most dramatically between 1 and 3
Competition
h posthepatectomy
Stat3-~
FIG. 2. Temporal course of Stat3 induction in liver regeneration, with cold competition analysis. Left panel, EMSA analysis of nuclear extracts obtained from liver at indicated times posthepatectomy. Right, in the presence of 100× cold oligonucleotide.
1446 CRESSMAN, DIAMOND, AND TAUB HEPATOLOGY May 1995
A Oh posth
B
-o- hep p 80
=u 60
4o
20
i i i i i i i . - . _ . _ . _ . 0 0 .5 1 3 5 h posthepatectomy
FIG. 3. Induction of Stat3 DNA-binding posthepatectomy in the presence of cycloheximide. (A) EMSA showing temporal course of Stat3 induction in animals that were pretreated with CHX before partial hepatectomy (hep/CHX). (B) Graphic representation of rela- tive induction compared with control partial hepatectomy (hep).
for Stat3 inactivation. It was important to test the abil- ity of PRL-1 PTPase to inactivate Stat3 DNA binding. Bacterially expressed PRL-1 shows activity in a p-nitro- phenylphosphate assay, whereas the PRL-lmut that con- tains a point mutation at the active site of the enzyme does not (Fig. 5A). However, as shown in Figs. 5B and C, although the control PTPase YOP phosphatase is able to rapidly eliminate Stat3 binding in 30-minute hepatec- tomy extracts, the PRL-1 phosphatase has virtually no effect. This indicates that hepatic Stat3 DNA binding is dependent on tyrosine phosphorylation and that bacteri- ally expressed PRL-1 is an ineffective PTPase in elimi- nating Stat3 DNA binding activity.
Potential Target Genes for Star3 in the Regenerating Liver. The characterization of Stat3 in regenerating liver indicates that it is induced rapidly, but peak activ-
ity does not occur until 3 hours after hepatectomy. Thus, if it is a positive regulator of gene transcription, its ma- jor effects should occur a few hours after the partial hepatectomy; and because its activity is quite extended, it may result in prolonged activation of target genes. It is not induced after insulin t reatment of H35 cells. We have characterized the expression of more than 70 im- mediate and delayed-early genes that are activated in the regenerating liver. 13 Based on peak and prolonged expression during liver regeneration, different induction in H35 cells, and lack of induction in sham surgery, a subset of these genes are potential candidates for Stat3 regulation (Table 1). It has been proposed that immedi- ate-early genes may be activated by a combination of transcription factors, including serum response and Stat factors. Therefore, because many of the genes listed in Table 1 are also induced in H35 cells, they are not ex- cluded from regulation by Stat3 during liver regenera- tion. The regulation in H35 cells may be caused by fac- tors different from Stat3. However, the temporal course of induction in the two systems is different because the genes peak early in H35 cells and induction is short lived. They show a later peak (later than 1 hour) and prolonged induction in regenerating liver that is com- patible with regulation by Stat3.
DISCUSSION
Investigators have long been interested in identi- fying factors that are responsible for triggering liver regeneration. The identification of transcription factors that preexist in liver cells in an inactive form and are
Stat3
FIG. 4. Stat3 is not induced following insulin treatment of H35 cells. EMSA analysis using the Stat3 oligonucleotide with nuclear extracts prepared from H35 cells treated with insulin for indicated times compared with extract from regenerating liver (3PHCHX, 3 hours posthepatectomy in the presence of cycloheximide). The H35 extracts showed adequate levels of other DNA-binding proteins (e.g. Caat enhancer binding protein, #e3) in all lanes.

rapidly activated within minutes of the partial hepatec- tomy may provide clues to the initiating signals. Spe- cifically, we now know that both NF-KB and Stat3 are rapidly activated in the remnant liver following partial hepatectomy. Whatever growth factor(s) are responsi- ble for initiating the regenerative response must be
A o.5,
0.4,
0
o T"
• 0.3 u c
0.2, w ,a <
0.1
B i
PRL PRL mut
PTPase
h posthep
HEPATOLOGY Vo]. 21, No. 5, 1995 CRESSMAN, DIAMOND, AND TAUB 1447
Stat3-~
C
u
O > o
_u
3o I
PRL PRL mut
Con YOP
capable of directly activating these factors. Stat3 pre- exists in the liver in an inactive form and is rapidly activated in a protein synthesis-independent fashion with peak activity 3 hours after hepatectomy. Dissec- tion of the Stat pathway of signal transduction sug- gests that tyrosine phosphorylation may be mediated by the just another kinase family of protein tyrosine kinases. ~3 DNA binding of Stat3 in regenerating liver is dependent on tyrosine phosphorylation as DNA bind- ing is rapidly eliminated by the YOP phosphatase. Fur- ther studies will determine if the Janus family of ki- nases are part of the signal transduction cascade in liver regeneration.
Growth factors show some specificity for activation of particular Stat factors. This may allow for the identi- fication of the particular growth factors that are im- portant in liver regeneration. For instance, EGF has been shown to activate the DNA-binding complex that comigrates with Star3 even in quiescent liver. 21 EGF acts as a mitogen in isolated hepatocytes, and recently, it has been shown that EGF is rapidly produced in the regenerating liver in a temporal course that correlates with the immediate-early growth responses EGF does not act as a mitogen in H35 cells that have no EGF receptors, and Stat3 is not activated by mitogen treat- ment of H35 cells. Therefore, there is a correlation be- tween EGF and Stat3 that supports the hypothesis that EGF may be a triggering factor in liver regeneration. In addition, IL-6 can also activate Stat3, and IL-6 levels have been shown to increase in the sera ofhepatectom- ized animals, providing some support for IL-6 action in liver regeneration. 4 It is possible that in vitro systems, including isolated hepatocytes, will provide an environ- ment in which Star3 regulation by specific growth fac- tors can be shown. Recently, we have shown that hepa- tocyte growth factor and EGF can induce some immediate-early genes in isolated hepatocytes. 3s By de- termining if this regulation is mediated by Stat3, we will be able to provide additional insight into the impor- tance of hepatocyte growth factor and/or EGF in trig- gering liver regeneration.
It is probable that Stat3 is responsible for transcrip- tional induction of a subset of immediate-early genes in regenerating liver, perhaps in concert with other transcription factors. Interestingly, the activation of Stat3 is prolonged in regenerating liver, whereas the activation of Stat factors in many other cytokine- and
F~G. 5. Dependence of Stat3 binding on tyrosine phosphorylation. (A) Activity of bacterially expressed PRL-1 in p-nitrophenylphos- phate assay. Two hundred fifty micrograms of bacterially expressed PRL-1 per milliliter, of PRL-lmut per milliliter, or of control protein per milliliter were incubated with p-nitrophenylphosphate for 1 hour before measurement of absorbance at 410 nm. (B) EMSA using Stat3 oligonucleotide and indicated liver nuclear extracts incubated with 62.5 t~g/mL bacterially expressed PRL-1, mutant PRL-1 (PRLmut; cysteine to serine change at the active site), control (Con; bacterially expressed IKB-c~), or commercially available yersinia PTPase (yer- sinia PTPase). (C) Graphic representation of results in (A).

1448 CRESSMAN, DIAMOND, AND TAUB HEPATOLOGY May 1995
TABLE 1. C a n d i d a t e I m m e d i a t e - E a r l y G e n e s T h a t May Be Activated by Stat3 in R e g e n e r a t i n g L i v e r
H35 Gene (peak expression, h) RL Peak (RL) (h) Growth-Induced Sham
PRL-1 (C) + 3 + c-jun +(.5) + 4 + /3-actin +(1) + 3 + JunB +(.5) + 2 + c-myc +(2) + 2 + Fibronectin - + 4 + RL-53 + 2 + RL-98(PC3) +(.5) + 2 +
NOTE. Expression characteristics of these genes have been reportedJ °-~3''~3'3~ Growth-induced genes 13 that are expressed for a prolonged time period during liver regeneration corresponding with the major proliferative phase.
Abbreviations: RL, regenerating liver; C, constitutive; +, expressed; - , not expressed.
g r o w t h f a c t o r - a c t i v a t e d cells is of r e l a t i v e l y s ho r t du- r a t i on , 30 m i n u t e s to a n hour . ls-2v T h i s i m p l i e s t h a t the s i g n a l s r e s p o n s i b l e for i t s a c t i v a t i o n a re p r o t r a c t e d a n d / o r t h a t i t s d o w n r e g u l a t i o n is r e l a t i v e l y slow. As a r e su l t , S t a t 3 m a y be ab le to t r a n s c r i p t i o n a l l y a c t i v a t e t a r g e t genes t h a t a re e x p r e s s e d for a l o n g e r pe r iod du r - i n g t he f i r s t G1 p h a s e o f h e p a t o c y t e s d u r i n g l ive r r e g e n - e r a t i on . T h i s pos s ib i l i t y a l lows us to i d e n t i f y p o t e n t i a l t a r g e t genes for S t a t 3 r e g u l a t i o n . I t is a l r e a d y k n o w n t h a t S t a t fac tors m a y c o n t r i b u t e to t he r e g u l a t i o n of c- fos a n d c-myc. 2s-3° Howeve r , d u r i n g l i ve r r e g e n e r a t i o n t he e x p r e s s i o n p a t t e r n of c-myc m e s s e n g e r R N A corre- s p o n d s m o r e p rec i se ly t h a n c-fos w i t h t he t e m p o r a l course of S t a t 3 i n d u c t i o n . L ikewise , a n u m b e r of o t he r genes i den t i f i ed in T a b l e 1 show t e m p o r a l e x p r e s s i o n t h a t is c o m p a t i b l e w i t h a c t i v a t i o n by S ta t3 . More de- t a i l ed a n a l y s e s of t h e i r p r o m o t e r r eg ions wil l a l low us to a s c e r t a i n the c o n t r i b u t i o n of S t a t3 to t h e i r r e g u l a - t i on d u r i n g l ive r r e g e n e r a t i o n .
Acknowledgment: We t h a n k B r e n t C o c h r a n for A- 431 e x t r a c t s a n d L a u r i e Z i m m e r m a n for m a n u s c r i p t p r e p a r a t i o n .
R E F E R E N C E S
1. Fausto N, Mead JE. Regulation of liver growth: protooncogenes and transforming growth factors. Lab Invest 1989;60:4-13.
2. Michalopoulos GK. Liver regeneration: molecular mechanisms of growth control. FASEB J 1990;4:176-187.
3. Fausto N, Webber EM. Liver regeneration. In: Arias IM, Boyer JL, Fausto N, Jacoby WB, Schachter D, Shafritz DA, eds. The liver: biology and pathobiology. New York: Raven, 1988:1059- 1079.
4. Akerman P, Cote P, Yang SQ, McClain C, Nelson S, Bagby G J, Diehl AM. Antibodies to tumor necrosis factor-a inhibit liver regeneration after partial hepatectomy. Am J Physiol 1992; G579-G585.
5. Lau LF, Nathans D. Identification of a set of genes expressed during the G0/G1 transition of cultured mouse cells. EMBO J 1985;4:3145-3151.
6. Lau LF, Nathans D. Expression of a set of growth-related imme- diate early genes in BALB/c3T3 cells: coordinate regulation with c-fos or c-myc. Proc Natl Acad Sci U S A 1987;84:1182-1186.
7. Almendral JM, Sommer D, MacDonald-Bravo H, Burckhardt J, Perera J, Bravo R. Complexity of the early genetic response to growth factors in mouse fibroblasts. Mol Cell Biol 1988;8:2140- 2148.
8. Zipfel PF, Irving SG, Kelly K, Siebenlist U. Complexity of the
primary genetic response to mitogenic activation of human T cells. Mol Cell Biol 1989;9:1041-1048.
9. Herschman HR. Primary response genes induced by growth fac- tors and tumor promoters. Annu Rev Biochem 1991;60:281-319.
10. Mohn KL, Laz TM, Melby AE, Taub R. Immediate-early gene expression differs between regeneration liver, insulin-stimu- lated H-35 cells, and mitogen-stimulated 3T3 cells: liver specific induction patterns of gene 33, PEPCK, and the jun, fos and egr families. J Biol Chem 1990;265:21914-21921.
11. Mohn KL, Lax TM, Hsu J-C, Melby AE, Bravo R, Taub R. The immediate-early growth response in regenerating liver and insu- lin-stimulated H-35 cells: comparison to serum-stimulated 3T3 cells and identification of 41 novel immediate-early genes. Mol Cell Biol 1991; 11:381-390.
12. Goyette M, Petropoulos C, Shanko P, Fausto N. Regulated tran- scription of c-Ki-ras and c-myc during compensatory growth of rat liver. Mol Cell Biol 1984;4:1493-1498.
13. Haber BA, Mohn KL, Diamond RH, Taub R. Induction patterns of 70 genes during nine days after hepatectomy define the tempo- ral course of liver regeneration. J Clin Invest 1993;91:1319-1326.
14. Tewari M, Dobrzanski P, Mohn KL, Hsu J-C, Bravo R, Taub R. Rapid induction in regenerating liver of RL/IF-1, an IkB that inhibits NH-kB, Rel-p50; and PHF, a novel kB site binding com- plex. Mol Cell Biol 1992; 12:2898-2908.
15. Fitzgerald MJ, Webber EM, Donovan JR, Fausto N. Rapid DNA binding by NF-KB in hepatocytes at the start of liver regenera- tion. Cell Growth Differ, in press.
16. Cressman DE, Taub R. Physiologic turnover of NF-KB by nuclear proteolysis. J Biol Chem 1994;269:26594-26597.
17. Cressman DE, Greenbaum LE, Haber BA, Taub R. Rapid activa- tion of PHF/NFKB in hepatocytes, a primary response in the regenerating liver. J Biol Chem 1995;269:30429-30435.
18. Sadowski HB, Shuai K, Darnell JE, Jr, Gilman MZ. A common nuclear signal transduction pathway activated by growth factor and cytokine receptors. Science 1993;261:1739-1744.
19. Shuai K, Stark GR, Kerr IM, Darnell JE Jr. A single phosphoty- rosine residue of Stat91 required for gene activation by inter- feron-y. Science 1993;261:1744-1746.
20. Larner AC, David M, Feldman GM, Igarashi K-I, Hackett RH, Webb DSA, Sweitzer SM, et al. Tyrosine phosphorylation of DNA binding proteins by multiple cytokines. Science 1993;261:1730- 1733.
21. Ruff-Jamison S, Chen K, Cohen S. Induction by EGF and inter- feron-y of tyrosine phosphorylated DNA binding proteins in mouse liver nuclei. Science 1993;261:1733-1736.
22. Fu X-Y, Zhang J-J. Transcription factor p91 interacts with the epidermal growth factor receptor and mediates activation of the c-fos gene promoter. Cell 1993;74:1135-1145.
23. Darnell JE, Jr, Kerr IM, Stark GR. Jak-STAT pathways and transcriptional activation in response to IFNs and other extra- cellular signaling proteins. Science 1994;264:1415-1421.
24. Improta T, Schindler C, Horvath CM, Kerr IM, Stark GR, Dar- nell JE, Jr. Transcription factor ISGF-3 formation requires phos-

HEPATOLOGY Vo1. 21, No. 5, 1995 CRESSMAN, DIAMOND, AND TAUB 1449
phorylated Stat91 protein, but Stat113 protein is phosphorylated independently of Stat91 protein. Proc Natl Acad Sci U S A 1994; 91:4776-4780.
25. Shuai K, Horvath CM, Huang LHT, Qureshi SA, Cowburn D, Darnell JE Jr. Interferon activation of the transcription factor Stat91 involves dimerization through SH2-phosphotyrosyl pep- tide interactions. Cell 1994; 76:821-828.
26. Zhong Z, Wen Z, Darnell JE Jr. Stat3: A STAT family member activated by tyrosine phosphorylation in response to epidermal growth factor and interleukin-6. Science 1994;264:95-98.
27. Zhong Z, Wen Z, Darnell JE Jr. Stat3 and Stat4: members of the family of signal transducers and activators of transcription. Proc Natl Acad Sci U S A 1994;91:4806-4810.
28. Hayes TE, Kitchen AM, Cochran BH. Inducible binding of a factor to the c-fos regulatory region. Proc Natl Acad Sci U S A 1987; 84:1272-1276.
29. Sacca R, Cochran BH. Identification of a PDGF-responsive ele- ment in the murine c-myc gene. Oncogene 1990;5:1499-1505.
30. Wagner BJ, Hayes TE, Hoban CJ, Cochran BH. The SIF binding element confers sis/PDGF inducibility onto the c-fos promoter. EMBO J 1990;9:4477-4484.
31. Gorski K, Carneiro M, Schibler U. Tissue-specific in vitro transcrip- tion from the mouse albumin promoter. Cell 1986;47:767-776.
32. Taub R, Roy A, Dieter R, Koontz J. Insulin as a growth factor
in rat hepatoma cells: stimulation of protooncogene expression. J Biol Chem 1987;262:10893-10897.
33. Diamond RH, Cressman DE, Laz TM, Abrams CS, Taub R. PRL- 1, a unique nuclear protein Tyr-phosphatase, affects cell growth. Mol Cell Biol 1994; 14:3752-3762.
34. Dunphy WG, Kumagai A. The cdc25 protein contains an intrinsic phosphatase activity. Cell 1991;67:189-196.
35. Taub R, Mohn KL, Diamond RH, Du K, Haber BA. Molecular aspects of insulin-regulated hepatic growth. In: Draznin B, Le- Roith D, eds. Molecular biology of diabetes, Part II. Totawa, NJ: Humana, 1994:301-320.
36. Diamond RH, Du K, Lee VM, Mohn KL, Haber BA, Tewari DS, Taub R. Novel delayed-early and highly insulin-induced growth response genes. J Biol Chem 1993;268:15185-15192.
37. Mullhaupt B, Feren A, Fodor E, Jones A. Liver expression of epidermal growth factor RNA. Rapid increases in immediate- early phase of liver regeneration. J Biol Chem 1994;269:19667- 19670.
38. Weir E, Che Q, DeFrances MC, Bell A, Taub R, Zarnegar R. Rapid induction of mRNAs for liver regeneration factor (LRF-1) and insulin-like growth factor binding protein-1 (IGFBP-1) in primary cultures of rat hepatocytes by hepatocyte growth factor and epidermal growth factor. HEPATOLOGY 1994;20:955-960.




















