
Functional Recognition of the Modified Human tRNALys3UUU Anticodon Domain by HIV's Nucleocapsid...
18
Functional Recognition of the Modified Human tRNA Lys3 UUU Anticodon Domain by HIV's Nucleocapsid Protein and a Peptide Mimic William D. Graham 1 †, Lise Barley-Maloney 1 †, Caren J. Stark 2 †, Amarpreet Kaur 1 , Khrystyna Stolyarchuk 1 , Brian Sproat 3 , Grazyna Leszczynska 4 , Andrzej Malkiewicz 4 , Nedal Safwat 1 , Piotr Mucha 5 , Richard Guenther 1 and Paul F. Agris 2 ⁎ 1 Molecular and Structural Biochemistry, North Carolina State University, Raleigh NC 27695, USA 2 The RNA Institute, Biological Sciences, University at Albany, Albany, NY 12222, USA 3 Integrated DNA Technologies BVBA, Interleuvenlaan 12A, B-3001 Leuven, Belgium 4 Institute of Organic Chemistry, Technical University, Żeromskiego 116, 90-924, Łódź, Poland 5 Department of Chemistry, University of Gdansk, Sobieskiego 18, 80-952 Gdansk, Poland Received 28 January 2011; received in revised form 6 April 2011; accepted 11 April 2011 Edited by M. F. Summers Keywords: Modified nucleosides; N 6 -threonylcarbamoyla- denosine; 5-methoxycarbonylmethyl-2- thiouridine; NCp7; anticodon stem and loop The HIV-1 nucleocapsid protein, NCp7, facilitates the use of human tRNA Lys3 UUU as the primer for reverse transcription. NCp7 also remodels the htRNA's amino acid accepting stem and anticodon domains in preparation for their being annealed to the viral genome. To understand the possible influence of the htRNA's unique composition of post- transcriptional modifications on NCp7 recognition of htRNA Lys3 UUU , the protein's binding and functional remodeling of the human anticodon stem and loop domain (hASL Lys3 ) were studied. NCp7 bound the hASL Lys3 UUU modified with 5-methoxycarbonylmethyl-2-thiouridine at position-34 (mcm 5 s 2 U 34 ) and 2-methylthio-N 6 -threonylcarbamoyladenosine at position- 37 (ms 2 t 6 A 37 ) with a considerably higher affinity than the unmodified hASL Lys3 UUU (K d = 0.28 ± 0.03 and 2.30 ± 0.62 μM, respectively). NCp7 denatured the structure of the hASL Lys3 UUU -mcm 5 s 2 U 34 ;ms 2 t 6 A 37 ;Ψ 39 more effectively than that of the unmodified hASL Lys3 UUU . Two 15 amino acid peptides selected from phage display libraries demonstrated a high affinity (average K d = 0.55 ± 0.10 μM) and specificity for the ASL Lys3 UUU - mcm 5 s 2 U 34 ;ms 2 t 6 A 37 comparable to that of NCp7. The peptides recognized a t 6 A 37 -modified ASL with an affinity (K d = 0.60 ± 0.09 μM) comparable to that *Corresponding author. Present address: The RNA Institute, University at Albany-SUNY, 1400 Washington Ave., Albany, NY 12222, USA. E-mail addresses: [email protected]; [email protected]; [email protected]; [email protected]; [email protected]. † These authors contributed equally to the research. For W.D.G., the work is in partial fulfillment of the graduate degree. Present addresses: L. Barley-Maloney, BD Diagnostics – Women's Health and Cancer, 21 Davis Dr., RTP, NC 277709, USA; B. Sproat, Chemconsilium GCV, Jaarmarktstraat 48, 2221, Booischot, Belgium; N. Safwat, Clinical Marketing – Microbiology, bioMerieux Inc., 100 Rodolphe St., Durham, NC 27712, USA; R. Guenther, Department of Plant Pathology, North Carolina State University, Raleigh, NC 27695, USA. Abbreviations used: NCp7, HIV nucleocapsid protein; hASL Lys UUU , anticodon stem and loop of human tRNA Lys3 UUU ; mcm 5 s 2 U 34 , 5-methoxycarbonylmethyl-2-thiouridine at anticodon wobble position 34; ms 2 t 6 A 37 , 2-methylthio-N 6 - threonylcarbamoyladenosine at position 37; Ψ 39 , pseudouridine at anticodon stem position 39. doi:10.1016/j.jmb.2011.04.025 J. Mol. Biol. (2011) 410, 698–715 Contents lists available at www.sciencedirect.com Journal of Molecular Biology journal homepage: http://ees.elsevier.com.jmb 0022-2836/$ - see front matter © 2011 Elsevier Ltd. All rights reserved.
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Functional Recognition of the Modified Human tRNALys3UUU Anticodon Domain by HIV's Nucleocapsid...

doi:10.1016/j.jmb.2011.04.025 J. Mol. Biol. (2011) 410, 698–715
Contents lists available at www.sciencedirect.com
Journal of Molecular Biologyj ourna l homepage: ht tp : / /ees .e lsev ie r.com. jmb
Functional Recognition of the Modified HumantRNALys3
UUU Anticodon Domain by HIV'sNucleocapsid Protein and a Peptide Mimic
William D. Graham1†, Lise Barley-Maloney1†, Caren J. Stark2†,Amarpreet Kaur1, Khrystyna Stolyarchuk1, Brian Sproat3,Grazyna Leszczynska4, Andrzej Malkiewicz4, Nedal Safwat1,Piotr Mucha5, Richard Guenther1 and Paul F. Agris2⁎1Molecular and Structural Biochemistry, North Carolina State University, Raleigh NC 27695, USA2The RNA Institute, Biological Sciences, University at Albany, Albany, NY 12222, USA3Integrated DNA Technologies BVBA, Interleuvenlaan 12A, B-3001 Leuven, Belgium4Institute of Organic Chemistry, Technical University, Żeromskiego 116, 90-924, Łódź, Poland5Department of Chemistry, University of Gdansk, Sobieskiego 18, 80-952 Gdansk, Poland
Received 28 January 2011;received in revised form6 April 2011;accepted 11 April 2011
Edited by M. F. Summers
Keywords:Modified nucleosides;N6-threonylcarbamoyla-denosine;5-methoxycarbonylmethyl-2-thiouridine;NCp7;anticodon stem and loop
*Corresponding author. Present addNY 12222, USA. E-mail addresses: [email protected]; PAgris@a† These authors contributed equallPresent addresses: L. Barley-Malo
USA; B. Sproat, Chemconsilium GCMicrobiology, bioMerieux Inc., 100 RNorth Carolina State University, RaAbbreviations used: NCp7, HIV n
mcm5s2U34, 5-methoxycarbonylmeththreonylcarbamoyladenosine at pos
0022-2836/$ - see front matter © 2011 E
The HIV-1 nucleocapsid protein, NCp7, facilitates the use of humantRNALys3
UUU as the primer for reverse transcription. NCp7 also remodelsthe htRNA's amino acid accepting stem and anticodon domains inpreparation for their being annealed to the viral genome. To understandthe possible influence of the htRNA's unique composition of post-transcriptional modifications on NCp7 recognition of htRNALys3
UUU, theprotein's binding and functional remodeling of the human anticodon stemand loop domain (hASLLys3) were studied. NCp7 bound the hASLLys3
UUUmodified with 5-methoxycarbonylmethyl-2-thiouridine at position-34(mcm5s2U34) and 2-methylthio-N6-threonylcarbamoyladenosine at position-37 (ms2t6A37) with a considerably higher affinity than the unmodifiedhASLLys3
UUU (Kd=0.28±0.03 and 2.30±0.62 μM, respectively). NCp7denatured the structure of the hASLLys3
UUU-mcm5s2U34;ms2t6A37;Ψ39more effectively than that of the unmodified hASLLys3
UUU. Two 15 aminoacid peptides selected from phage display libraries demonstrated a highaffinity (average Kd=0.55±0.10 μM) and specificity for the ASLLys3
UUU-mcm5s2U34;ms2t6A37 comparable to that of NCp7. The peptides recognized at6A37-modified ASL with an affinity (Kd=0.60±0.09 μM) comparable to that
ress: The RNA Institute, University at Albany-SUNY, 1400 Washington Ave., Albany,[email protected]; [email protected]; [email protected];lbany.edu.y to the research. For W.D.G., the work is in partial fulfillment of the graduate degree.ney, BD Diagnostics – Women's Health and Cancer, 21 Davis Dr., RTP, NC 277709,V, Jaarmarktstraat 48, 2221, Booischot, Belgium; N. Safwat, Clinical Marketing –odolphe St., Durham, NC 27712, USA; R. Guenther, Department of Plant Pathology,leigh, NC 27695, USA.ucleocapsid protein; hASLLys
UUU, anticodon stem and loop of human tRNALys3UUU;
yl-2-thiouridine at anticodon wobble position 34; ms2t6A37, 2-methylthio-N6-ition 37; Ψ39, pseudouridine at anticodon stem position 39.
lsevier Ltd. All rights reserved.

Fig. 1. HIV nucleocapsid protein,nucleosides. (a) NCp7 sequence of(b) Sequence and secondary structureviral RNA. The secondary structuresequences, is drawn from that repoEurope and Oceania.17 The A-rich budomain of the htRNALys3
UUU and issequence with significant complemenwith the anticodon domain fragmetRNALys3 (htRNALys3
UUU) with all odihydrouridine at positions 16, 20 anmcm5s2U34 (∗U34 in red); 2-methylthm7G; 5-methylcytidine at positionsN1-methyladenosine-58, m1A. (d) Chloop domain of htRNALys3
UUU: mcm(hASLLys3UUU-mcm5s2U34;ms2t6A37;Ψsynthesized with a G27-C34 terminalmcm5s2U34;ms2t6A37;Ψ39 with a 3′-tems2t6A37 was used for the characterhASLLys3UUU was synthesized with a(D). (g) The anticodon domain of hhASLLys1CUU-t
6A37. The modified anNCp7 were comprised of residues 29
699Modification-dependent Peptide Binding of tRNALys3
for hASLLys3UUU-mcm5s2U34;ms2t6A37, indicating a preference for the t6A37
modification. Significantly, one of the peptides was capable of relaxing thehASLLys3
UUU-mcm5s2U34;ms2t6A37;Ψ39 structure in a manner similar to thatof NCp7, and therefore could be used to further study protein recognition ofRNA modifications. The post-transcriptional modifications of htRNALys3
UUUhave been found to be important determinants of NCp7's recognition priorto the tRNALys3
UUU being annealed to the viral genome as the primer ofreverse transcription.
© 2011 Elsevier Ltd. All rights reserved.
Introduction
The lentivirus, Human Immunodeficiency Virustype 1 (HIV-1), is a highly adaptive virus that uses acomplex system of proteolytic cleavages of itspolyprotein precursor Pr55Gag (Gag), to replicateand integrate its genome into host cell chromosomes.The Gag polyprotein contains two zinc finger motifsthat are conserved among most retroviral proteinsand is the only component of the HIV-1 virion that isrequired for virus particle assembly.1-3 The life cycleof HIV-1 includes an assembly and budding stepthat results in the proteolytic cleavage of the Gagpolyprotein by HIV-1 protease (PR) into six matureGag proteins. Arguably, one of the most vitalproteins produced from this cleavage is the nuclearcapsid protein 7 (NC or NCp7) (recently reviewed inRef. 4).1,2,4,5 NCp7 is a strongly basic, 55 amino acidprotein containing two shortened zinc fingers(knuckles) (Fig. 1a). NCp7 functions as a nucleicacid chaperone during various stages of viralprogression.4,6 Both theoretical and empirical studiesindicate that approximately 1400 copies of NCp7
NCp7, human tRNALys3U
55 amino acids and zincof the Primer Binding Site (of the HIV-1 sero(sub)typrted for the HIV-1 sero(slge of the viral serotype-Gadjacent to the 18-residutarity to the anticodon dont for experiments withf its known modified nucd 48, D; pseudouridine-27io-N6-threonylcarbamoyla48 and 49, m5C; 2′-O-memical structures of the h5s2U34, and ms2t6A37. (e)39). The modificationsbase pair instead of therminal biotin was used foization of the peptides. (f)nd without fluorescein contRNALys1
CUU with the md unmodified dodecame-40 and modifications we
coat each of the two copies of HIV's RNA viralgenome in the capsid.7-9
NCp7's chaperone activity is characterized by itsability to destabilize RNA structure, facilitating aconformational rearrangement, or remodeling, that iscritical to the replication of HIV-1. NCp7 is involvedin the specific use of the human host cell lysine tRNAisoacceptor 3, htRNALys3
UUU, as the primer of HIVreverse transcription.10,11 The htRNALys3
UUU withthe anticodon UUU is one of some 45 tRNAs, and theminor species of three lysine tRNAs in human cells. Aconformational alteration of its canonical L-shapedtRNA structure is required for htRNALys3
UUU tofunction as the primer for reverse transcription.12 Infacilitating the annealing of the primer to the HIVgenomic RNA, NCp7 recognizes, binds and de-stabilizes the htRNALys3
UUU.4 The Watson-Crick
hydrogen bonding of the 3′-terminal nucleotidesand that of the U-rich anticodon stem and loop (ASL)are destabilized and annealed to the complementary18 nucleotides of the viral RNA's primer binding site(PBS)13,14 and approximately 12 nucleotides of theA-rich bulge of Loop 1,15,16 respectively (Fig. 1b).
UU, its anticodon stem and loop domain and its modifiedfinger structures. The single tryptophan is denoted, W37.PBS) and A-rich Stem and Loop 1 of HIV-1, sero(sub)type G,e-G, having the highest complementariness of the Loop Iub)type B, the most prevalent serotype in the Americas,Loop 1 sequence (VGL; green) binds the U-rich anticodone primer binding site (blue). The single stranded Loop 1main was used alone and in a truncated construct (bolded)NCp7. (c) Sequence and secondary structure of humanleosides: N2,N2-dimethylguanosine at position 10, m2
2G10;, 39 and 55, Ψ; 5-methoxycarbonylmethyl-2-thiouridine-34,denosine-37, ms2t6A37 (∗A37 in red); N7-methylguanosine,ethylribothymidine-54 (2′-O-methyl-5-methyluridine), Tm;ypermodified nucleosides within the anticodon stem andModified anticodon stem and loop domain of htRNALys3
mcm5s2U34 and ms2t6A37 are in red. The ASLs wereΨ27-A43 in order to stabilize the stem. The hASLLys3UUU-r the peptide selection, whereas the hASLLys3-mcm5s2U34;The unmodified hASLLys3UUU sequence. The unmodifiedjugated to the 5′-terminus (F) and dabsyl at the 3′-terminusodification N6-threonylcarbamoyladenosine (t6A37, in red),r anticodon domain fragments used in experiments withre introduced at U34, A37 and U39).

700 Modification-dependent Peptide Binding of tRNALys3
The htRNALys3UUU is packaged along with
tRNALys1,2,18,19 the human lysyl-tRNA synthetase(KRS)20 and the viral genome into the newly formedvirions and used as the primer during a subsequentinfection.4 NCp7's ability to bind and manipulateboth the viral genomicRNAandhost cell's htRNALys3
for successful viral propagation make it a primarytarget for the development of antiviral therapeutics.21
NCp7binds toRNAs in an ionic strength dependentmanner that is characterized by a range of affinities.22
The protein has a demonstrated preference for singlestranded polyGs, especially in GU (and GT)stretches,23 and appears to melt G●U wobble basepairs, first.6,24,25 In its recognition and binding ofhtRNALys3
UUU, NCp7 exhibits its highest affinity forthe amino acid acceptor stem.4,26 As many as eight to
Fig. 1 (legend on
ten copies of the protein are thought to bind to thestem and loop regions of the tRNA in a non-specificmanner.27-29 In addition to the two zinc fingers,NCp7 has a number of positively charged aminoacids that are suggestive of howNCp7 could attach toany RNA, including the different regions andconformations of htRNALys3
UUU.4,5,30-32 Sequenceanalysis of RNA-binding proteins has led to theidentification of specific motifs that are responsiblefor protein recognition of RNA structure.33 Many ofthese motifs contain positively charged residues, suchas arginine, that interact in a non-specific mannerwith the negatively charged backbone of the RNA.However, any degree of specificity of a protein'srecognition of a particular RNA, such as NCp7 forhtRNALys3
UUU, would be dependent on the RNA's
previous page)

701Modification-dependent Peptide Binding of tRNALys3
nucleotide sequence and resulting conformation.Post-transcriptional modifications alter the confor-mation and dynamics of the RNA structure,specifically the loop regions of tRNAs where mostmodifications are found, thereby creating confor-mations that are favorable for a specific protein-tRNA interaction.34 The modified nucleosides ofhtRNALys3
UUU may affect NCp7 recognition of andbinding to the tRNA.Mature htRNALys3
UUU contains the naturallyoccurring post-transcriptional modifications 5-methoxycarbonylmethyl-2-thiouridine (mcm5s2U34)at wobble position 34 and 2-methylthio-N6-threonylcarbamoyladenosine (ms2t6A37) at position37 within the tRNA's anticodon stem and loopdomain (hASLLys3
UUU; Fig. 1c,d). Pseudouridine,Ψ39 is located at position 39, 3′ to the anticodonloop. Post-transcriptional modifications within theanticodon domain are identity elements that arerecognized by the cognate aminoacyl-tRNA synthe-tases for some tRNAs and are important for effectiveaminoacylation of the tRNA for many, particularlyfor those tRNAs having position 34 modifieduridines (glutamic acid, glutamine and lysine).35,36
The post-transcriptional modifications of U34 andA37 increase the affinity of the hASLLys3
UUU to bindthe AAA and AAG codons on the ribosomal-boundmRNA,37,38 and their bacterial counterparts increasethe efficiency of translation and maintain thetranslational reading frame.39 Thus, the naturallyoccurring post-transcriptional modifications presenton htRNALys3
UUU may assist in the recognition ofhtRNALys3
UUU by NCp7, and other HIV-1 proteins,though the importance of these modifications to thetRNA's recognition by HIV proteins has not beenexplored. The ability of NCp7 to disrupt thestructure of the htRNALys3
UUU anticodon domainfor subsequent annealing to the A-rich bulge of theviral Loop 1 sequence could also be facilitated by thepresence of post-transcriptional modifications in theASL. Anticodon domain modifications are charac-terized, generally, by their ability to maintain anopen ASL loop and yet enhance base stacking forcodon binding.40,41 As a consequence, the open loopwould be more susceptible to conformationalchange by NCp7.We have investigated the relationship between
post-transcriptional modifications and the recog-nition of htRNALys3
UUU by NCp7. Here, we reportthat the anticodon domain modifications consid-erably enhanced NCp7 binding affinity for thehASLLys3
UUU. Modifications facilitated NCp7 dis-ruption of the ASL's ordered conformation as aprelude to the structural rearrangement that pre-cedes its annealing to the HIV genome. In addition,we describe the selection and characterization of a15 amino acid peptide that both mimics the NCp7recognition of the modified hASLLys3
UUU, and isalso capable of disrupting the RNA structure.
Results
NCp7 recognition of the modified anticodondomain of htRNALys3
UUU
NCp7 binds to different RNAs to varyingdegrees.4,6 Studies of NCp7 binding to RNA suggestthat it binds to single stranded regions of RNA withlittle specificity.4,6 When interacting with the unmo-dified transcript of the htRNALys3
UUU and stableisotope labeled tRNA expressed in Escherichia coli,NCp7 appears to bind initially to the nucleotides atthe position of the tertiary interactions at the centerof the L-shaped structure from which the meltingof acceptor stem/T-stem helices and anticodondomain would occur.4,24,42,43 The induced confor-mational change and dynamics now favor theannealing of the disrupted acceptor and T stems tothe complementary PBS and the tRNA's U-richanticodon domain to the virus' A-rich bulge of theLoop 1 sequence.15,16 In order to examine in somedetail the extent to which the naturally occurringmodifications within the hASLLys3
UUU may influenceNCp7 recognition of the tRNA, the protein's bindingto various constructs of the hASLLys3
UUU wasinvestigated. The ASLLys3
UUU and its fragments(residues 32-43) were chemically synthesized withand without the complete and incomplete humanmodifications mcm5s2U34, ms2t6A37, 2-thiouridine(s2U34) and Ψ39, and the E. coli modification5-methylaminomethyluridine (mnm5U34).
38,44 NCp7(75 nM) was titrated with the unmodified ASLLys3
UUU(0–3μM) and the change in fluorescence, or quenching,of NCp7's single tryptophan residue (W37) wasrecorded (Fig. 2a).7 NCp7's interaction with theunmodified hASLLys3UUU yielded a Kd in the lowμM range (Kd=2.3±0.62 μM; Table 1) and as suchestablished a baseline for recognition of the RNA inthe absence of modifications.NCp7 was also titrated with the doubly modified
hASLLys3UUU having the naturally occurring mod-
ifications, mcm5s2U34 at wobble position 34 andms2t6A37 at position 37, 3′-adjacent to the anticodon,hASLLys3
UUU-mcm5s2U34;ms2t6A37 (Fig. 2a). Condi-tions were identical to those of the titration with theunmodified ASLLys3
UUU, described above. NCp7exhibited a 10-fold increase in affinity for themodified ASL (Kd=0.28±0.03 μM; Table 1). Thetitration of NCp7 with the unmodified hASLLys3
UUUresulted in a maximum fluorescence quenching ofonly 68% as compared to 75% reduction achievedwith the doubly modified hASLLys3
UUU (Fig. 2a).When the htRNALys3
UUU is annealed to the viralgenome for the priming of reverse transcription,the U-rich ASLLys3
UUU anneals to the A-rich bulgeof the viral stem and loop (Fig. 1b) which also mustbe destabilized by NCp7. In order to compareNCp7's binding of the modified and unmodified

Fig. 2. NCp7 association with the anticodon stemand loop domain of htRNALys3
UUU. The binding ofhASLLys3UUU to NCp7 was monitored by observingthe change in fluorescence (quenching) of theprotein's one tryptophan (W37). (a) NCp7 (75 nM)titrated with increasing concentrations (0–3 μM of thehASLLys3UUU-mcm5s2U34;ms2t6A37 (●) and the unmo-dified hASLLys3UUU (■). (b) NCp7 binding of the HIV-1,serotype-G, viral Stem and Loop 1. The quenching of theNCp7 W37 was monitored in a single experiment whentitrated with a truncated construct of the A-rich HIV-1,serotype-G, viral Stem and Loop 1 (VSL-1 with VGL). Table 1. Affinity of NCp7, and peptides P6 and P17 for
hASLLys
NCp7 orPeptide
ASLLys, Fragment,or HIV Stem/Loop I
DissociationConstant Kd (μM)
NCp7 Unmodified ASLLys3UUU 2.30±0.62ASLLys3UUU-mcm5s2U34;ms2t6A37 0.28±0.03
ASLLys1CUU-t6A37 ND
HIV Stem/Loop I (VSL-1) 0.3Unmodified Fragment29-40 0.62±0.12
Fragment29-40-s2U34 0.01±0.01
Fragment29-40-Ψ39 0.48±0.03Fragment29-40-mnm5U34 0.20±0.07Fragment29-40-t
6A37 0.15±0.06Fragment29-40-mnm5U34;Ψ39 0.25±0.05Fragment29-40-mnm5U34;t
6A37 0.26±0.06A-rich Loop 1 dodecamer 0.06±0.01Duplex Anticodon/Loop 1 1.11±0.16
P6 Unmodified ASLLys3UUU IDASLLys3UUU-mcm5s2U34;ms2t6A37 0.50±0.10
ASLLys1CUU-t6A37 0.60±0.09
P17 Unmodified ASLLys3 IDASLLys3UUU-mcm5s2U34;ms2t6A37 0.60±0.10
ASLVal3ACU-m6A37 0.49±0.31
ASLLys1CUU-t6A37 1.80±0.20
P21 Unmodified ASLLys3UUU 0.98±0.03ASLLys3UUU-mcm5s2U34;ms2t6A37 1.30±0.04
ND=experiment not done.ID=Indeterminable, could not be calculated from the data.
702 Modification-dependent Peptide Binding of tRNALys3
ASLLys3UUU to that of the A-rich bulge of the viral
RNA, the NCp7 was titrated with a 27 nucleotide,stem and loop construct of the HIV-1, serotype-GRNA (VSL-1) and the fluorescence of tryptophan 37was monitored (Fig. 2b). NCp7 bound the VSL-1with an affinity comparable to its binding of thehASLLys3
UUU-mcm5s2U34;ms2t6A37, and to otherstem/loop constructs of viral RNA (reviewed inRef. 45), but considerably greater than that of itsbinding to the unmodified ASLLys3
UUU (Fig. 2b;Table 1).We also assessed the binding of the NCp7 to
fragments of the hASLLys3UUU that were composed of
the single stranded, dodecamer oligoribonucleotideresidues 29-40, and its single stranded complemen-tary sequence of the viral A-rich Loop 1 (Fig. 1b).The fragments were chosen to be non-self comple-mentary, and thus could not form a stem and loopstructure. Chemical constraints to the syntheses ofthese fragments in sufficient quantities limited us tothe introduction of the incomplete modifications of
s2U34 and N6-threonylcarbamoyladenosine (t6A37),along with Ψ39. In addition, the bacterial analog ofmcm5U34, mnm5U34, was incorporated into thefragments. Because of its known preference forsingle stranded RNA, it is not surprising that NCp7exhibited a higher affinity for the shorter, unstruc-tured fragments of the anticodon and Loop 1 thanfor the unmodified hASLLys3
UUU hairpin (Table 1).In fact, NCp7 bound with high affinity to thesingle stranded unmodified anticodon fragment(Kd=0.62±0.12 μM) and even more so to the A-richLoop 1 (Kd=0.06±0.01 μM) than to a doublestranded RNA composed of these two fragments(Kd=1.11±0.16 μM) or the unmodified hASLLys3
UUU(Kd=2.30±0.62 μM).Perhaps more interesting is the comparison of
binding affinities between modified and unmodi-fied fragments. Introduction of the modificationsfurther enhanced the affinity of NCp7 for thesefragments. In particular, the individual introduc-tions of s2U34, mnm5U34 and t6A37 significantlyincreased the protein's affinity for the anticodonfragment (Table 1). NCp7 bound the modifieddodecamer fragments of the hASLLys3
UUU withan affinity higher than that reported for DNAdodecamers.46 These results for single strandedmodified anticodon fragments along with those ofthe ASL study suggest that NCp7's binding ofhtRNALys3
UUU is considerably enhanced whenmodifications are present. The modifications maynot be necessary for NCp7's initial non-specificbinding of the tRNA's acceptor stem, but they

703Modification-dependent Peptide Binding of tRNALys3
appear to be of considerable importance for theprotein's recognition of the anticodon stem and loopdomain.
Peptide recognition of the modified anticodondomain of htRNALys3
UUU
Phage Display Library selection of peptides isan effective way of screening a large number ofpeptide-RNA interactions under biologically rele-vant conditions to determine the contribution ofmodifications to protein recognition of tRNAs,and the amino acids required.47,48 Peptides thatbound the hASLLys3
UUU-ms2t6A37;mcm5s2U34;Ψ39were selected using two different phage displaylibraries and two different conditions for elutionfrommicroplates.49 This triply modified hASLLys3
UUUwas chemically synthesized with biotin at the3′-end, and bound to streptavidin-coated, highcapacity microplates. The first round of screeningwas conducted with the unmodified hASLLys3
UUUbound to streptavidin plates, followed by multiplerounds of screening with the triply modifiedhASLLys3
UUU. Phage were eluted from the platesusing both alkaline and acid conditions.49 Phagethat demonstrated an affinity to bind the unmo-dified hASLLys3
UUU after four rounds of selectionwere catalogued and eliminated from furtherscreening. In the subsequent step, phage thatdid not bind hASLLys3
UUU were selected usinghASLLys3
UUU-mcm5s2U34; ms2t6A37;Ψ39. After fouriterations of phage selection against the fullymodified hASLLys3
UUU, 155 distinct colonies werechosen for DNA sequencing. Several sequences werefound to occur repeatedly among the 155 colonies.Twenty-five peptides were chosen for further study,however five sequences either failed chemical
Table 2. Peptides generated from phage display library scree
Clone No. 15 and 16 Amino acid Peptides
1 FSVSFPSLPAPPDRS3 GRVTYYSCGVSLLFQ4 AGPVPLHSLSYYYNQ5 RAVMTVVWPVSFAGF6 RVTHHAFLGAHRTVG8 PAVASTSSLIIDGPF9 PKAFQYGGRAVGGLW10 AAHVSEHYVSGSLRP11 ASVGPAPWAMTPPVS12 APALWYPWRSLLPLY13 ASLHPVPKTWFFLLS14 WSHSRNTADVPVSML15 HRGYCRDRWNCGEYF17 PHRQCSAPAKSCKILP19 TLPACHELPKHCKRRG20 TLPACHELPKHCNEAR21 NGPECNAYMVRCRGYH23 GNSNCPMLNEQCPWQD24 HTETCINIRNTCTTVA25 LKLPCKITINNCQLAG
synthesis or were insoluble due to a high degree ofhydrophobicity. Thus, 20 peptides were synthesizedwith a conjugated fluorescein isothiocyanate (FITC)at the N-terminus for analysis of their interactionwith the ASLs using fluorescence spectroscopy.Peptides 6 (P6), 15 (P15), 17 (P17) and 21 (P21)were chosen for further study as they appearedmultiple times from separate colonies during thephage display selection (Table 2) suggesting thatthese four would bind to the hASLLys3
UUU-ms2t6A37;mcm5s2U34 with high affinity. Unfortunately, P15lacked the solubility required of the assay and waseliminated from the study.Though the biotin conjugated hASLLys3
UUU-mcm5s2U34;ms2t6A37;Ψ39 was used for the selectionof phage, its synthesis and purification in quantitiesrequired for characterizing peptide affinity andspecificity was cost prohibitive. We and othershave found that Ψ39 slightly improves the thermalstability of hASLs, but adds little to their chemistryand conformation, and nothing to their affinity forthe cognate codon on the ribosome.50,51 Therefore,the affinity of the fluorescein-conjugated peptidesfor the doubly modified hASLLys3-mcm5s2U34;ms2t6A37 was determined by monitoring the fluo-rescence quenching upon binding of the ASLs(Fig. 3). Each of the peptides exhibited fluores-cence quenching with increasing concentrations ofdoubly modified hASLLys3
UUU. Both P6 and P17bound the hASLLys3
UUU-mcm5s2U34;ms2t6A37 witha high degree of affinity (Kd=0.5 μM±0.1 and0.6±0.1 μM, respectively; Table 1), and comparableto that of NCp7.The specificities of P6, P17 and P21 for the
unmodified ASLLys3UUU were assessed and com-
pared. In contrast to their binding of the modifiedASLLys3
UUU, P6 and P17 demonstrated little to no
ning
Library/Elution Times Appearing
Fuse5/basic 18Fuse5/basic 4Fuse5/basic 1Fuse5/acidic 5Fuse5/acidic 10Fuse5/acidic 2Fuse5/acidic 1Fuse5/acidic 1Fuse5/acidic 1Fuse5/acidic 1Fuse5/acidic 1Fuse5/acidic 1F88-cys6/basic 8F88-cys6/basic 8F88-cys6/basic 4F88-cys6/basic 1F88-cys6/acidic 4F88-cys6/acidic 1F88-cys6/acidic 1F88-cys6/acidic 1

Fig. 3. The association of phage display selectedpeptides with hASLLys3
UUU-mcm5s2U34;ms2t6A37. (a) Pep-tide P6 binding of hASLLys3
UUU-mcm5s2U34;ms2t6A37. Thefluorescence of the FITC-conjugated peptide 6 (P6)(percent quenching) was monitored with addition of theASL. P6 titrated with hASLLys3
UUU-mcm5s2U34;ms2t6A37( ); P6 titrated with the unmodified hASLLys3
UUU ( ); P6titrated with E. coli ASLVal3
UAC-m6A37 ( ). (b) Peptide P17
binding of hASLLys3UUU-mcm5s2U34;ms2t6A37. P17 titrated
with the hASLLys3UUU-mcm5s2U34;ms2t6A37 ( ); P17
titrated with the unmodified hASLLys3UUU ( ); P17
titrated with ASLVal3UAC-m
6A37 ( ). (c) The anticodonstem and loop of E. coli tRNAVal3
UAC with themodification N6-methyladenosine at position 37 (m6A37)(ASLVal3
UAC-m6A37).
704 Modification-dependent Peptide Binding of tRNALys3
interaction with this ASL, and a dissociation constantcould not be determined (Fig. 3a,b; Table 1).However, P21 bound the hASLLys3
UUU-mcm5s2U34;ms2t6A37 and the unmodified hASLLys3
UUU withconsiderable, and almost equal, affinity (Kd=0.98±0.03 μM and Kd=1.30±0.04 μM, respectively)demonstrating that this peptide's binding of theASLs is relatively non-specific. To determine if
the peptides were non-specifically recognizing thefive base-paired stem and/or the seven-residueloop of the ASL hairpin structure, we tested theirabilities to bind the E. coli tRNAVal3
UAC. The E. colitRNAVal3
UAC has a different stem, and an antico-don loop nucleoside sequence similar to that ofhASLLys3
UUU (Fig. 3c). The tRNAVal3UAC construct
had its A37 modified at N6, but with only a methylgroup, i.e. m6A37 (ASLVal3
UAC-m6A37). P6 did not
bind the E. coli ASLVal3UAC-m
6A37 indicating thatpeptide P6 is specifically recognizing the modifica-tions of the hASLLys3
UUU-mcm5s2U34;ms2t6A37.However, the fluorescence of peptide P17 wasquenched when titrated with the ASLVal3
UAC-m6A37
indicating recognition of an N6-modified A37(Fig. 3a,b).The specificity and affinity of P6 and P17 for
hASLLys3UUU-mcm5s2U34;ms2t6A37 may be due to
only one of the two modifications rather thanboth. Therefore, we determined their affinities forthe singly modified anticodon stem and loop ofthe human tRNALys species 1 (hASLLys1
CUU-t6A37)
(Fig. 1g). Though htRNALys1CUU has an unmodi-
fied C34, A37 is modified to t6A37 and naturallylacks the ms2-moiety. Otherwise the stem and loopsequences of the ASLs are the same. Both peptidesexhibited significant affinities for hASLLys1
CUU-t6A37
(Kd=0.60±0.09 μM and 1.8±0.20 μM, respectively;Table 1). Not only was the affinity of the peptidesfor the hASLLys1
CUU-t6A37 considerable, but the
fluorescence was quenched to over 50% when thepeptide was saturated with hASLLys1
CUU-t6A37, as
compared to just 12% with the unmodifiedhASLLys3
UUU. Apparently, the single modificationt6A37 is an important recognition element.
NCp7 and a peptide mimic alter the hASLLys3
conformation in a modificationdependent manner
NCp7 has the ability to disrupt RNA secondarystructure to enable the annealing of the primerhtRNALys3
UUU to the HIV genomic RNA which isenergetically favored. To test NCp7's ability todisrupt the base stacking and hydrogen bonds ofthe hASLLys3
UUU, we designed an unmodifiedhASLLys3
UUU to exhibit a fluorescence (Förster)resonance energy transfer (FRET) between thedonor, 5′-conjugated fluorescein, and the acceptor,a 3′-conjugated accepting quencher, dabsyl (Fig. 1f).Because fluorescence emission from the 5′-terminalfluorescein is transferred to the neighboring 3′-terminal dabsyl, increases in fluorescence would beexpected should the NCp7 disrupt the structure ofthe hASLLys3
UUU and thereby increase the distancebetween the two termini. In fact, the protein's abilityto denature the unmodified hASLLys3
UUU wassimilar to the thermal denaturation of the sameASL (Fig. 4). At a molar ratio of ∼2:1 (NCp7 to

3.5
3.0
2.5
2.0
1.5
1.0
0.5
100
80
60
40
20
0
20 30 40 50 60 70 80 90 100
0 2 4 6 8 10
Flu
ores
cenc
e (a
rbitr
ary
units
x 1
0-5 )
Abs
orba
nce
(per
cent
)
Temperature (°C)
NCp7/hASLLys3 (molar ratio)
Fig. 4. NCp7 disruption of the htRNALys3UUU antico-
don stem and loop domain compared to thermaldenaturation. An unmodified anticodon stem and loopdomain construct (hASLLys3
UUU) with a 5′-conjugatedfluorescein and a 3′-conjugated dabsyl was titrated withNCp7 and the increase in fluorescence monitored ( ). ThehASLLys3
UUU was half “melted” at an NCp7 to RNA ratioof ∼2. In addition, an unmodified hASLLys3
UUU (withoutfluorescein or dabsyl) was thermally denatured/renaturedfrom 20 to 95 °C and the UV absorbance at 260 nm wasrecorded, averaged and normalized (percent absorbance, ).The temperature at which the sample was half-melted was∼40 °C.
705Modification-dependent Peptide Binding of tRNALys3
hASLLys3UUU) and a temperature of∼40 °C, the ASL
appears to be half melted. Having established thatNCp7 will disrupt the structure of the unmodifiedhASLLys3
UUU, we were interested in knowing if thenatural modifications had an effect on this functionand whether the peptides were capable of mimick-ing NCp7's denaturation of the RNA. However, forthese experiments we were compelled to use adifferent approach because the synthesis of the fullymodified hASLLys3
UUU with fluorescein and dabsylwas chemically difficult and inordinately expensive.Changes in nucleic acid base stacking, and
indirectly hydrogen bonding, caused by an alter-ation in the nucleotide sequence, changes in tem-perature, or by binding of ligands can be monitoredby circular dichroism (CD) spectrapolarimetry.52,53
The titration of an RNA with either a protein orpeptide adds ellipticity to the CD spectrum of thenucleic acid, especially in the wavelength rangewhere backbone interactions are observed, 220-250 nm. Fortunately, both proteins and peptideshave a spectral null in the wavelength range ofinterest for observing base stacking interactions,
250-280 nm,52 enabling the recording of changes inthe RNA conformation even in the presence ofproteins. NCp7 alone exhibited the expected spec-tral null in the wavelength range of 260-280 nm(Fig. 5a). The unmodified hASLLys3
UUU (1.5 μM)was titrated with NCp7 and its CD ellipticitymonitored for a change in RNA conformation(Fig. 5a). The CD spectra of the unmodifiedhASLLys3
UUU when titrated with increasing concen-trations of NCp7 exhibited a decrease in ellipticity,indicating that the protein binds and relaxes theASL structure (Fig. 5a), which is consistent withdecreased stacking interactions, as would beexpected from the results of the prior FRETexperiment.The triply modified hASLLys3
UUU-mcm5s2U34;ms2t6A37;Ψ39 was also titrated with NCp7 and theprotein again appeared to disrupt the base stackinginteractions and to relax the ASLLys3
UUU structure(Fig. 5b). We compared the changes in ellipticitycaused by NCp7 for unmodified ASLLys3
UUU andmodified hASLLys3
UUU at the wavelength maxi-mum and found that hASLLys3
UUU-mcm5s2U34;ms2t6A37;Ψ39 required less protein to cause dena-turation of the ASL structure (Fig. 5c). The amountof NCp7 required to achieve the point at which halfof the ASL was melted was 0.3 μM for the modifiedhASLLys3
UUU, and 0.46 μM for the unmodifiedhASLLys3
UUU (Fig. 5c). Unlike the binding experi-ments monitored by fluorescence, the CD experi-ments are not indicative of the degree to whichbinding occurs. At concentrations of NCp7 abovethe ratio of 1, NCp7 to ASL, there was little changein CD ellipticity with addition of more protein.Since we did not perform a kinetic analysis, wecannot say that at any snapshot in time, therewould be more NCp7 on the modified ASL versusthe unmodified ASL under conditions in whicheach is saturated with protein. However, thesedata confirmed that the hASLLys3
UUU-mcm5s2U34;ms2t6A37;Ψ39 is bound and its secondary structuremore effectively disrupted by NCp7 than is thatof the unmodified hASLLys3
UUU.The hASLLys3
UUU-mcm5s2U34;ms2t6A37;Ψ39 andunmodified hASLLys3
UUU constructs were alsotitrated with the P6 peptide to determine if it wasable to mimic the ability of NCp7 to bind andunfold the ASL structure. The spectra indicatedthat the peptide mimics NCp7 in its ability notonly to bind hASLLys3
UUU-mcm5s2U34;ms2t6A37;Ψ39better than the unmodified hASLLys3
UUU, but alsoin its ability to relax the ASLLys3 structures (Fig. 5d).Substantive differences were evident in theP6-facilitated denaturation of the triply-modifiedhASLLys3
UUU in comparison to that of the unmodi-fied hASLLys3
UUU. When changes in ellipticity werecompared for increasing concentrations of P6 atthe wavelength maximum, it was apparent that thepeptide completely disrupted the structure of the
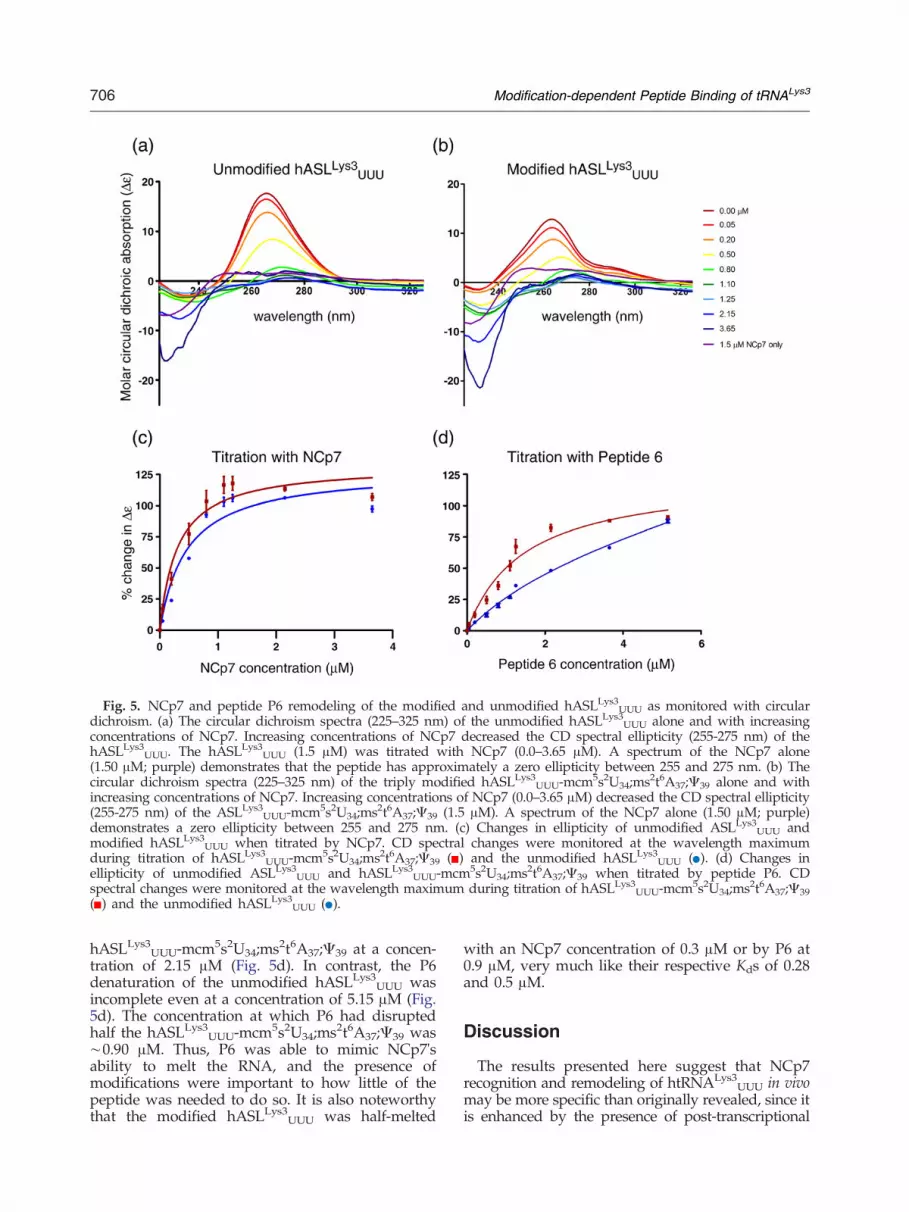
Fig. 5. NCp7 and peptide P6 remodeling of the modified and unmodified hASLLys3UUU as monitored with circulardichroism. (a) The circular dichroism spectra (225–325 nm) of the unmodified hASLLys3UUU alone and with increasingconcentrations of NCp7. Increasing concentrations of NCp7 decreased the CD spectral ellipticity (255-275 nm) of thehASLLys3UUU. The hASLLys3UUU (1.5 μM) was titrated with NCp7 (0.0–3.65 μM). A spectrum of the NCp7 alone(1.50 μM; purple) demonstrates that the peptide has approximately a zero ellipticity between 255 and 275 nm. (b) Thecircular dichroism spectra (225–325 nm) of the triply modified hASLLys3UUU-mcm5s2U34;ms2t6A37;Ψ39 alone and withincreasing concentrations of NCp7. Increasing concentrations of NCp7 (0.0–3.65 μM) decreased the CD spectral ellipticity(255-275 nm) of the ASLLys3
UUU-mcm5s2U34;ms2t6A37;Ψ39 (1.5 μM). A spectrum of the NCp7 alone (1.50 μM; purple)demonstrates a zero ellipticity between 255 and 275 nm. (c) Changes in ellipticity of unmodified ASLLys3
UUU andmodified hASLLys3UUU when titrated by NCp7. CD spectral changes were monitored at the wavelength maximumduring titration of hASLLys3UUU-mcm5s2U34;ms2t6A37;Ψ39 ( ) and the unmodified hASLLys3UUU ( ). (d) Changes inellipticity of unmodified ASLLys3
UUU and hASLLys3UUU-mcm5s2U34;ms2t6A37;Ψ39 when titrated by peptide P6. CDspectral changes were monitored at the wavelength maximum during titration of hASLLys3UUU-mcm5s2U34;ms2t6A37;Ψ39( ) and the unmodified hASLLys3UUU ( ).
706 Modification-dependent Peptide Binding of tRNALys3
hASLLys3UUU-mcm5s2U34;ms2t6A37;Ψ39 at a concen-
tration of 2.15 μM (Fig. 5d). In contrast, the P6denaturation of the unmodified hASLLys3
UUU wasincomplete even at a concentration of 5.15 μM (Fig.5d). The concentration at which P6 had disruptedhalf the hASLLys3
UUU-mcm5s2U34;ms2t6A37;Ψ39 was∼0.90 μM. Thus, P6 was able to mimic NCp7'sability to melt the RNA, and the presence ofmodifications were important to how little of thepeptide was needed to do so. It is also noteworthythat the modified hASLLys3
UUU was half-melted
with an NCp7 concentration of 0.3 μM or by P6 at0.9 μM, very much like their respective Kds of 0.28and 0.5 μM.
Discussion
The results presented here suggest that NCp7recognition and remodeling of htRNALys3
UUU in vivomay be more specific than originally revealed, since itis enhanced by the presence of post-transcriptional

707Modification-dependent Peptide Binding of tRNALys3
modifications. The recruitment of htRNALys3UUU for
the priming of HIV-1 reverse transcription involvesthe Gag and the Gag-Pol polyprotein, and thehuman KRS, or probably a composite of theseproteins.4 The NCp7 domain of Gag is an RNAchaperone of htRNALys3
UUU and facilitates itsannealing to the viral RNA.4 The anticodon domainof tRNALys species is an identity determinant for thesynthetase.54,55 The synthetase is involved in theunacylated tRNA's recruitment to the viral RNA. Itis proposed that the synthetase may be recognizingan htRNALys3
UUU-like element (TLE) of the viralRNA that has sequence homology with the U-richanticodon region and thereby directs the tRNA tothe complementary viral sequences for annealing.4
The complexity of protein components and thedetailed mechanisms by which they recognize,recruit and anneal htRNALys3
UUU to the viralgenome have yet to be described.NCp7 facilitates the use of the htRNALys3
UUUspecies as the primer for reverse transcription. Theprotein remodels the tRNA conformation, thusfacilitating its annealing to the viral genome forthe initiation of reverse transcription during thenext round of infection.4,56 The HIV-1 capsidsare packaged with viral RNA and concentratehtRNALys3
UUU some 60-fold to a ratio of tRNA toviral genome of approximately 6:1 to 8:1.4,57 Thetwo zinc fingers of NCp7 provide the proteinwith a highly structure-specific binding of RNA,whereas a large number of basic residues produce ahigh degree of promiscuity in its binding to a varietyof nucleic acids with varying affinities in vitro.4
NCp7 exhibits some sequence discrimination,6,24,25
however, prior studies did not include the poten-tial influence of the naturally occurring, post-transcriptional modifications that are found in thehost cell tRNALys3
UUU. As would be expected for aprotein involved in the recognition and remodelingof the htRNALys3
UUU conformation, NMR spectros-copy revealed that NCp7 bound to the base pairedregions inside of the canonical L-shape of theacceptor stem of tRNA wherein lies tertiarystructure hydrogen bonding and the G6●U67 andT54●A58 pairs.24 With 15N-labeling, the iminoprotons of the base paired stems of a htRNALys3
UUUproduct of E. coli cloning and thus with bacterialmodifications were observable, but those of theanticodon loop were in too fast an exchange to beobserved. In experiments with unmodified D- andT-domain fragments of the tRNA, NCp7 exhibitedan affinity for the D-domain fragment comparableto the association that we observed with theanticodon domain.58
Modifications had a considerable impact uponNCp7 recognition of ASLLys3
UUU. In fact, theaffinity of NCp7 for the hASLLys3
UUU-mcm5s2U34;ms2t6A37 (Kd=0.28±0.03 μM) was equivalent tothe affinity reported for its binding to the native
bovine tRNALys3UUU (Kd=0.31 μM) using, as we did,
the inherent fluorescence of the protein's W37 andunder conditions of comparable ionic strength.28
NCp7 appears to bind nucleic acids in a salt-effectedmanner as would be expected of an electrostaticinteraction with the phosphodiester backbonecontributing significantly to the binding constant.59
The binding of the stem loop structures of HIV'sΨ-domain contribute significantly to our knowl-edge of its affinities for these structures. NCp7 hasa high affinity for model hairpins (20 mers) of theΨ-domain stem and loops SL2 and SL3 (Kd≈0.1-0.2 μM) and a lower affinity for SL4 (Kd≈14 μM ).60-62
The affinities of NCp7 that we have observed forthe hASLLys3
UUU-mcm5s2U34;ms2t6A37 was compa-rable (Kd=0.28±0.03 μM) and acquired under similarsalt conditions. The equilibrium binding constant ofthe bacterial modified Fragment29-40-mnm5U34;t
6A37(0.26±0.06 μM) was comparable to that of thehASLLys3
UUU-mcm5s2U34;ms2t6A37 (0.28±0.03 μM),and little different than that of the fragment witht6A37 only (0.15±0.06 μM) especially when one takesinto consideration the errors of the binding study.The human tRNALys3
UUU has 18 nucleotides atthe 3′-terminus, most of which are engaged inA-form RNA duplexes of the amino acidacceptor stem and the T-stem. These nucleotidesare complementary to the viral PBS and mustfirst be freed from their internal hydrogenbonding to participate in binding to the PBS.Though this sequence is an important distinctiveproperty of the tRNA and critical to its primingfunction, the combination of modified nucleosidesfound in the anticodon loop of htRNALys3
UUU area chemically rich and exclusive attribute to thattRNA. The modifications s2U34 and t6A37 arefound in other tRNAs.63,64 Therefore, it is interest-ing to note that htRNALys3, htRNALys1,2, htRNAIle,and htRNAAsn are among those tRNAs mostfrequently found within the HIV viral capsid.18,19,65
The htRNALys1,2, htRNAIle, and htRNAAsn have thenucleoside modification t6A37. We have found thatNCp7 binds with high affinity to the hASLLys1
CUUwith t6A37, as well as to the hASLLys3
UUU-mcm5s2U34;ms2t6A37, but not to the unmodifiedhASLLys3
UUU. Thus, the modifications are them-selves recognition determinants for NCp7. Alterna-tively, the anticodon domain conformation resultingfrom the modifications enhances recognition. Wehypothesize that this recognition mechanism pro-vides another level of specificity to protein-nucleicacid interactions in vivo.NCp7 disruption of the hydrogen bonding of the
tRNA's amino acid accepting stem and the stem ofthe T-domain is well documented and is an acceptedfunctional mechanism of facilitating the binding ofthe tRNA to the viral PBS.4 Presumably, NCp7 isalso involved in disrupting the structure of theU-rich anticodon stem and loop domain to

708 Modification-dependent Peptide Binding of tRNALys3
facilitate the subsequent binding of this sequenceto the A-rich bulge of the Loop 1 region of theviral RNA.29,42 Using a FRET analysis between the5′- and 3′-termini of the hASLLys3
UUU, we foundthat when titrated with NCp7, the hASLLys3
UUUstructure is disrupted. This denaturation wasanalogous to the thermal denaturation of the ASL.The interaction of NCp7 with the full length
transcript of tRNALys3 had been monitored previ-ously by circular dichroism and a change in theprotein's spectrum was reported.62 However, littleto no change in the tRNA's spectrum was observedwith the addition of three mole-equivalents of NCp7to one of RNA, ∼1/25 of NCp7 to nucleotides.66
This result contrasts to that of other reports29,42 andalso to our own CD results with the ASLs. Theprotein's denaturation of the hASLLys3
UUU wasobserved by monitoring CD spectra while increas-ing the concentration of NCp7. The presence ofmodifications in the hASLLys3
UUU facilitated NCp7'sdenaturation of the hASLLys3
UUU. The heptadeca-mer ASL was half melted at a stoichiometry ofapproximating 2:1, NCp7 to ASL, consistent withthe reported ratio of one NCp7 per 6-8 nucleotides.At first this stoichiometry appears to contrast withthe one-site, non-linear curve fitting of the bindingcurves to the ASL. The protein does have aconsiderably greater affinity for the single strandedanticodon domain fragments than for the ASLhairpin. Thus, the second site binding of NCp7 tothe ASLs could be weaker than that of the first andnot observed under the conditions of our fluores-cence studies. The 12mer fragments would have oneor possibly two NCp7 bound per RNA, theheptadecamer RNA stem and loops would havetwo molecules of NCp7 bound and the 27mer VSLconstruct (Fig. 2b) being considerably larger couldbind more NC proteins.A stoichiometric binding of NCp7, one per
6-8 nucleotides, would decrease the possibility ofannealing the ASL to the viral RNA until the localamount of the protein is reduced, perhaps throughthe kinetics of on/off rates. In other systems, (e.g.,trans-activating region RNA interaction, TAR-TAR,and the dimer initiation signal interaction, DIS-DIS),it appears that only a small amount of nucleocapsidprotein is sufficient to produce annealing andstructural rearrangement (e.g., from kissing dimerto extended duplex).45,67,68 In fact, our CD experi-ments that monitored disruption of base stackingand not melting indicated that NCp7 at a ratio of0.2:1 (NCp7/ASL) and peptide at 0.5:1 weresufficient to dissolve half the base stacking in-teractions of the modified hASLLys3
UUU (Fig. 5c,d).Though the ratio seems low, CD results average allof the interactions taking place in the experimenteven when the amount of protein is below that of theASL. Thus, we see base stacking being significantlydisrupted with as little as 0.2:1 ratio of NCp7/ASL,
ie. 20% of the ASL is bound by the NCp7. We arenot alone in this observation of NCp7 interactionswith htRNALys3
UUU (personal communication, K.Musier-Forsyth).45,67,68 We found little difference inthe concentrations of NCp7 required for the half-maximal denaturation of the triply modified andunmodified hASLLys3
UUU, “destablization Kd” de-termined by CD (Fig. 5c). This is in contrast to thesignificant difference in the binding Kd (Table 1).NCp7 affinity for the modified hASLLys3
UUU wascomparable to that for NCp7 binding of viral RNAand significantly higher than that for the unmodi-fied hASLLys3
UUU. Although there are some simi-larities between the mechanisms of action of DNAsingle stranded binding protein (SSB) and NCp7,NCp7's faster kinetics and ability to aggregate, aswell as destabilize, RNA and DNA appear to bemost important for its functions.69 Thus, destabili-zation of RNA occurs at sub-saturating NCp7concentrations, whereas annealing/aggregation re-quires saturating levels of NC.70
The effects of modifications on the chemistry,structure and conformational dynamics of theanticodon loop of htRNALys3
UUU increased thebinding affinity of NCp7 and of peptide P6, andfacilitated disruption of the ASL structure. Themajor recurring RNA chemistries available asidentity determinants for protein recognition arethe nucleobase functional groups (NH2 and CO), theribose 2′-OH and the phosphate backbone. But,these per se do not create a unique identity. Thus,with some notable exceptions among the moststudied of RNA-binding proteins,71 there is littleunderstanding of how proteins specifically interactwith their target RNA to form native complexes.Peptide-RNA interactions are more easily studiedin detail, such as the thoroughly investigatedTat-TAR72 and Rev-RRE systems.73 Previously, wehad used Peptide Phage Display libraries toeffectively screen and select peptides that bindspecifically to modified nucleosides with highaffinity and specificity.47-49 In this study, peptideswere selected for their abilities to bind specifically tothe modified ASL region of the human tRNALys3
(hASLLys3UUU-mcm5s2U34;ms2t6A37). The peptides
selected from this screen were analyzed in detail fortheir abilities to bind the hASLLys3
UUU-mcm5s2U34;ms2t6A37. Two peptides were found to haveaffinities and specificities for ASLLys3-mcm5s2U34;ms2t6A37 that were comparable to those of NCp7.In addition, peptide P6 could also mimic NCp7'sability to denature the RNA's hairpin conforma-tion, albeit with somewhat reduced efficiency.Potentially, P6 could facilitate the annealing ofthe hASLLys3
UUU-mcm5s2U34;ms2t6A37 to the A-richbulge of the HIV's stem and Loop 1. In binding tohASLLys1
CUU-t6A37, P6 and P17 appeared to favor
the t6A37 over the mcm5s2U34 as the mostimportant of the modifications in their recognition

709Modification-dependent Peptide Binding of tRNALys3
of the modified hASLLys3UUU. Perhaps in using
mcm5s2U34 and ms2t6A37 and the t6-moiety inparticular as identity elements, the two peptidesbind to the anticodon, the 3′-side of the loop andthe 3′-side of the stem.The short sequence (15 amino acids) of peptide P6
and its lack of structure as determined by CDeffectively mimics the modification-facilitated bind-ing and functionality of the larger and structuredNCp7 (55 amino acids and two zinc fingers). Thebinding of P6 to the anticodon region of themodified hASLLys3
UUU was confirmed with prelim-inary data obtained from mass spectrometry (MS).MS experiments were conducted with the specificaim of observing complex formation between thepeptides and modified nucleosides. Electrosprayionization-MS (ESI-MS), an analytical techniquethat is used to observe non-covalent complexformation between nucleic acids and proteins insolution at nM concentrations,74 has been used toinvestigate the specific interactions between NCp7and the different domains of the HIV-1 packagingsignal.45,75,76 For this reason, we employed ESI-MSas the most cost effective screening technique toobserve the interaction between peptide P6 andhASLLys3
UUU-mcm5s2U34; ms2t6A37. As a control,the modified hASLLys3 was analyzed by ESI-MS inthe absence of the peptide. Spectra indicate a -4charge state for the RNA (Fig. 6). Peptide P6 wasanalyzed alone and under the same conditions.Both the modified and unmodified hASLLys3
UUUwere titrated with P6. A comparison of the resultingspectra led to the observation of a decrease in the
1500 200
m
Rel
ativ
e In
ten
sity
[AS
LL
ys3 U
UU-4
H]4
-
[AS
LL
ys3 U
UU-3
H]3
-
[AS
LL
ys3 U
UU-4
H]4
-
[AS
LL
ys3 U
UU-3
H]3
-
[Pep
•AS
LL
ys3 U
UU
4H]4
-
(a)
(b)
Fig. 6. Mass spectrometry of hASLLys3UUU-mcm5s2U34;ms2t
ms2t6A37 (relative intensity vs. mass/charge ratio) (b) ESI-MS oP6 to ASL is 5/1.
abundance of free peptide in solution, along withan increase in the abundance of its complex withhASLLys3
UUU-mcm5s2U34;ms2t6A37. With equimo-lar amounts of RNA and peptide, ∼50% of thehASLLys3
UUU-mcm5s2U34;ms2t6A37 was in a com-plex with P6, comparable to the results obtainedwhen the fluorescein-conjugated peptide's bindingof the modified hASLLys3
UUU was monitored byfluorescence. At a ratio of 1:5, RNA to peptide,∼100% of the modified hASLLys3
UUU had beenbound by P6 and the free ASL was no longerobserved (Fig. 6b). In these preliminary experiments,small amounts of the unmodified hASLLys3
UUU werebound by P6, and only 50% of E. coli ASLVal3
UACwas in a complex with P6 when the peptide was infive-fold excess (data not shown). Thus, peptide P6bound the modified hASLLys3
UUU preferentially.Anticodon loop modifications create an architec-
ture optimized for the tRNA's binding to the mRNAcodon by reducing thermal stability and conforma-tional dynamics.40,41,77 The modifications at wobbleposition 34, mcm5s2U34 and 3′-adjacent to theanticodon at position 37, ms2t6A37, reduce thermalstability by negating intra-loop hydrogen bonds(Fig. 7). They alter the nucleobase's hydrogenacceptors and donors known to engage in non-canonical, as well as canonical base pairing. Theirprominent physical volume widens the loop. How-ever, modifications at purine-37 also increase thebase stacking of the nucleosides on the 3′-side of theloop and thus increase the order within the loop.41,77
Therefore, the modifications would contribute aphysicochemical advantage, as well as their being
0 2500
/z
[AS
LL
ys3 U
UU-2
H]2
-
[Pep
•AS
LL
ys3 U
UU
3H]3
-
6A37 bound by P6. (a) ESI-MS of hASLLys3UUU-mcm5s2U34;
f hASLLys3-mcm5s2U34;ms2t6A37 bound to P6. The ratio of

Fig. 7. Natural modifications provide identity determinants and a physicochemical advantage to NCp7 remodeling ofhtRNALys3
UUU. (a) NCp7 (blue oval) interaction with the modified anticodon domain of htRNALys3UUU. The
modifications of the anticodon stem and loop domain of htRNALys3UUU, mcm5s2U34 (∗U34) and ms2t6A37 (∗A37), provide
unique identity determinants for NCp7. (b) NCp7 interaction with the unmodified anticodon domain of htRNALys3UUU.
In contrast to the modified anticodon domain of htRNALys3UUU, the unmodified domain lacks the identity determinants
and has intra-loop hydrogen bonds (- - - -) between the invariant U33 and A37.40
710 Modification-dependent Peptide Binding of tRNALys3
chemically distinct identity determinants, for theNCp7 denaturation of the htRNALys3
UUU. In con-trast to an unmodified hASLLys3
UUU, two NCp7molecules could ostensibly bind sequentially, per-haps cooperatively, to the hASLLys3
UUU-mcm5s2U34;ms2t6A37 and thus prepare the ASL for its annealingto the viral RNA (Fig. 7). The introduction of thenatural modifications provides the perfect model forrecognition of htRNALys3
UUU by viral proteins suchas NCp7 when studied in vitro, and may replicatethe higher degree of specificity appreciated in vivo.The abilities of peptides such as P6 to discriminatebetween modified and unmodified htRNALys3, andfor P6 also to be a functional mimic of NCp7 in itsability to disrupt ASL structure, make these peptidesprime candidates for the development of HIV-1antiviral therapeutics. The selection of peptides thatcan effectively function as mimics of RNA bindingproteins, could potentially lead to the developmentof tools in drug discovery and therapeutics thatwould silence viral gene expression in vivo.
Materials and Methods
RNA sample preparation
The unmodified, heptadecamer oligoribonucleotide cor-responding to the anticodon stem and loop of the humantRNALys3
UUU (hASLLys3UUU) and that of the Escherichia coli
tRNAValUAC isoacceptor 3 (ASLVal3UAC) were chemically
synthesized by Dharmacon (Thermo Fisher, Lafayette, CO)
using “ACE” chemistry.78 The hASLLys3UUU-ms2t6A37;
mcm5s2U34;Ψ39 with biotin conjugated at the 3′-end, thehASLLys3
UUU-mcm5s2U34;ms2t6A37, and the ASLVal3UAC
with the natural modification N6-methyladenosine,ASLVal3
UAC-m6A37, were synthesized by Integrated DNA
Technologies BVBA using phosphoramidite chemistries.79
The t6A37-modified ASL of the human tRNALysCUU
isoacceptor 1 (htRNALys1CUU-t
6A37) was synthesized bythe North Carolina State University Nucleic Acid Facilitywith the phosphoramidite of t6A37 having the ribose 2′and 5′ hydroxyls and the threonine carboxyl and hydroxylsuitably protected, and with standard ribonucleosidephosphoramidites and little change in standard synthesisprotocols. The unmodified ASLLys3
UUU with a 5′-conju-gated fluorescein and a 3′-dabsyl, a 27 nucleotideconstruct of the HIV A-rich bulge stem and loop, and 12nucleotide anticodon domain fragments were synthe-sized by the NCSU Nucleic Acid Facility using standard2′-O-TBDMS protected ribonucleoside phosphoramidites(ChemGenes, Wilmington, MA).80 The newly synthesizedASLs were purified by anion exchange HPLC (Nucleo-gen 60-7 DEAE; 250 mm×10 mm column), and desalted(Waters Corporation Sep-pak columns, Milford, MA).81
The purified ASLs were analyzed for their nucleosidecomposition by HPLC,82 and mass spectrometry, and themodifications observed by NMR,40 and X-ray crystallog-raphy bound to codon on the E. coli 30S ribosomalsubunit (Vendeix, A.F.P., Murphy, F.V. 4th and Agris, P.F.,unpublished data).
Isolation and purification of NCp7
The 55 amino acid NCp7 expression clone, pRD2, andNCp7 protein were the gifts of Dr. Michael Summers

711Modification-dependent Peptide Binding of tRNALys3
(University ofMaryland, Baltimore County). Additionally,NCp7 protein was expressed using BL21 (DE3) E. coli cellsand purified as described previously.61,83
Phage display selection of peptides and peptidesynthesis
Peptides that bound to the fullymodified anticodon stemand loop domain of htRNALys3 (hASLLys3
UUU-ms2t6A37;mcm5s2U34;Ψ39) were selected using two different phagedisplay libraries, fuse5 and f88-cys6, gifts from Dr. GeorgeSmith (University of Missouri-Columbia). The fuse5 libraryis that of a completely random 15 amino acid sequencecontaining more than 107 different phage.84 The f88-cys6 isa 16 amino acid sequence randomized for every residueexcept for the locations of two cysteines and containingmore than 2.7×108 phage.85 The 3′-biotin conjugatedhASLLys3
UUU-ms2t6A37;mcm5s2U34;Ψ39 was bound to96-well, streptavidin-coated, high capacity microplates(Pierce) in TTDBA buffer (1 mg/ml BSA and 0.02%NaN3 in 200/1 TBS/Tween, vol/vol; TBS buffer, 50 mMTris HCl, pH 7.5, 150 mM NaCl). The plates wereincubated with phage for 4 hr at 4 °C, and were thenwashed 5 times with a TBS-tween solution in order toremove any unbound phage. To elute the bound phagefrom the plates with either acidic or alkaline conditions,the plate wells were rinsed with acid elution buffer (Acidicconditions) or alkaline elution buffer (Basic conditions) ona shaker with gentle agitation for 10 minutes.49 The elutionmixture containing eluted phage was transferred to amicrotube containing the respective pH-neutralizingbuffer.49 The selected and isolated phage were amplifiedand the selection was reiterated for a total of five rounds. Ascreen was conducted with the unmodified hASLLys3UUUbound to strepavidin plates. Phage that demonstratedan affinity to bind hASLLys3UUU were catalogued andeliminated. The final round of selection was concludedwith a serial dilution of the isolated phage andquantified to determine yield. Phage clones were thenconstructed on starved E. coli K91BluKan cells. Of thephage that demonstrated an affinity for the fully modifiedhASLLys3UUU and not the unmodified hASLLys3UUU, 155distinct colonies were chosen, DNA was isolated andsequenced.49 The resulting sequences were used to selectthe peptides to be synthesized. Some sequences werefound to occur repeatedly among the 155 colonies (Table 1).Twenty-five sequences were chosen, but five sequenceseither failed chemical synthesis or were insoluble due to ahigh degree of hydrophobicity. Thus, 20 peptides weresynthesized with a conjugated fluorescein isothiocyanate(FITC) at the N-terminus (Table 1).The 20 peptides were chemically synthesized (Sigma-
Aldrich) with fluorescein isothiocyanate (FITC) conjugatedto the N-terminus for assaying peptide interaction withthe ASLs by fluorescence spectroscopy. The final concen-tration was determined by the Bradford assay using astandard curve generated from bovine serum albumin,and a peptide of known concentration. Peptides of lowsolubility were eliminated from further consideration.
Fluorescence spectroscopy
The importance of anticodon domain modified nucle-osides to NCp7's recognition of htRNALys3
UUU was
characterized by monitoring the changes in the intrinsicfluorescence properties of the protein's one tryptophanresidue (position 37, Fig. 1) upon the addition of RNA. Forbinding of the ASL's oligonucleotide fragments, trypto-phan fluorescence was monitored with a QuantaMaster™Model C 61 spectrofluorometer (Photon TechnologyInternational Inc.). The titrations were performed at25 °C with increasing amounts of RNA added to a fixedconcentration of 75 nM NCp7 in 400 μL buffer (50 mMHEPES, 50 mM NaCl, 0.04% PEG 8000 buffer, pH 7.5; orwith equivalent results 20 mM phosphate buffer, 20 mMNa2HPO4 and 20 mM KH2PO4 of equal volume, pH 6.8).NCp7 fluorescence was measured at 340 nm emission(290 nm excitation) in a 1 cm path length cuvette. Inmonitoring the fluorescence of fluorescein 3′-conjugatedhASLLys3
UUU with a 5′-dabsyl, fluorescence was measuredat 518 nm emission (490 nm excitation) in a 1 cm pathlength cuvette. Experiments were repeated three timeswith fluorescence detection collected over 10 seconds afteran initial 0.5 min period of equilibration. In assessing thebinding of NCp7 to the heptadecamer ASLs, a microplatespectrofluorometer was used (Molecular Devices Spectra-max Gemini XS with dual scanning monochrometers,Sunnyvale, CA). Under the conditions of 20 mM phos-phate buffer, the tryptophan fluorescence was observed at360 nm (285 nm excitation). Experiments were performedin triplicate and repeated at least twice. The resultingspectra were normalized and the percentage change influorescence was plotted against respective RNA concen-trations. An equilibrium binding constant (Kd) wasderived from the analysis of the curve using a one sitebinding, non-linear regression model (Prism, GraphPadSoftware, San Diego, CA). To assess and correct for inner-filter effects and background, fluorescence intensities wereadjusted for dilution, buffer fluorescence, and screeningeffects due to the presence of RNA. The RNAs inner-filtereffect was minimal, and the buffer chosen had insignifi-cant effects on NCp7 tryptophan fluorescence. Photo-bleaching was determined to be only 3% of totalfluorescence during a period of 10 min. Peptide interactionwith the ASLs was monitored through changes in thefluorescence of the N-terminal FITC that was observed at524 nm (486 nm excitation). The peptides (0.5 μM) weretitrated with varying concentrations of ASLs in 20 mMphosphate buffer. The Kds for each fluorescence experi-ment were assessed using the one site binding equationY=[Bmax][X]/[Kd+X]. As a control a non-RNA bindingpeptide (P31; AGPVPLHSLSYYYNQ) was used as abaseline (data not shown). The data from the P31 wasnot represented on the graph because of the high rate oferror that occurred with this non-specific interaction.
UV-monitored, thermal denaturations
Human ASLLys3UUU was dissolved to a concentration of
2 μM (20 mM phosphate buffer). Thermal denaturationsand renaturations, performed in triplicate in cells of 1 cmpath length, were monitored by UV absorbance (260 nm)using a Cary 3 spectrophotometer as previouslydescribed.50 Data points were averaged over 20 secondsand collected three times/minute with a temperatureramp of 1 °C/min from 5-90 °C. Data from denaturationsand renaturations were treated similarly. No hysteresiswas observed.

712 Modification-dependent Peptide Binding of tRNALys3
Circular dichroism (CD)
CD experiments, performed in triplicate, were con-ducted with a Jasco J815 spectropolarimeter and aninterfaced computer.86,87 RNA samples (1.5 μM) wereprepared in 10 mM phosphate buffer, pH 6.8, and spectracollected from 225 nm to 325 nm. To maintain RNA andbuffer concentrations and to minimize volume changes,the RNA samples were added to lyophilized protein orpeptide samples of known quantity resulting in a range ofprotein concentrations from 0.05 μM to 3.65 μM andpeptide concentration from 0.05 μM to 5.15 μM. The CDspectra were collected at 4 °C, at a rate of 10 nm perminute, a resolution of 1 nm, and the spectra wereaveraged over 6 runs. Normalization of spectra wasperformed by calculating molar circular dichroism Δɛ(cm2/mmol) using the formula Δɛ=(θ/32980)(C)(L)(N),where θ=raw CD amplitude (mdeg), C=concentration(mol/L), L=path length (cm) and N=number of nucleo-tides in RNA. Data were plotted and a one-site (NCp7) ortwo-site (P6), non-linear regression analysis was performedusing Prism 5 (GraphPad Software, San Diego, CA).
Acknowledgements
The authors wish to acknowledge the help ofWinnell Newman in the synthesis of some of theRNA at the North Carolina State University NucleicAcid Facility, Michael Summers for NCp7 and aclone of the protein, George Smith for the specificphage display libraries, Karin Musier-Forsyth for hersuggestions for the manuscript, Glenn Björk for themodified nucleoside composition analysis by HPLC,and Daniele Fabris for help with mass spectrometryand for manuscript suggestions. This work wassupported by grants from the National Institutes ofHealth, the National Science Foundation, and theNorth Carolina Biotechnology Center to P.F.A.(2RO1-GM23037, MCB0548602 and MRG 1102/Agris, respectively).
References
1. Freed, E. O. (1998). HIV-1 Gag proteins: Diversefunctions in the virus life cycle. Virology, 251, 1–15.
2. Coffin, J., Hughes, S. & Varmus, H. (1997). Retroviruses.Cold Spring Harbor Press, Plainview, NY.
3. Coren, L. V., Thomas, J. A., Chertova, E., Sowder, R. C.,Gagliardi, T. D., Gorelick, R. J. & Ott, D. E. (2007).Mutational analysis of the C-terminal gag cleavage sitesin human immunodeficiency virus type 1. J. Virol. 81,10047–10054.
4. Levin, J. G., Mitra, M., Mascarenhas, A. & Musier-Forsyth, K. (2010). Role of HIV-1 nucleocapsid proteinin HIV-1 reverse transcription. RNA Biol. 7, 754–774.
5. Thomas, J. A. & Gorelick, R. J. (2008). Nucleocapsidprotein function in early infection processes. Virus Res.134, 39–63.
6. Levin, J. G., Guo, J., Rouzina, I. & Musier-Forsyth, K.(2005). Nucleic acid chaperone activity of HIV-1nucleocapsid protein: critical role in reverse transcrip-tion and molecular mechanism. Prog. Nucleic Acid Res.Mol. Biol. 80, 217–286.
7. You, J. C. & McHenry, C. S. (1993). HIV nucleocapsidprotein. Expression in Escherichia coli, purification, andcharacterization. J. Biol. Chem. 268, 16519–16527.
8. Lapadat-Tapolsky, M., de Rocquigny, H., Van Gent,D., Roques, B., Plasterk, R. & Darlix, J. L. (1993).Interactions between HIV-1 nucleocapsid protein andviral DNA may have important functions in the virallife cycle. Nucleic Acids Res. 21, 831–839.
9. de Baar, M. P., van der Horn, K. H. M., Goudsmit, J.,de Ronde, A. & deWolf, F. (1999). Detection of humanImmunodeficiency virus type 1 nucleocapsid proteinp7 in vitro and in vivo. J. Clin. Microbiol. 37, 63–67.
10. Wain-Hobson, S., Sonigo, P., Danos, O., Cole, S. &Alizon, M. (1985). Nucleotide sequence of the AIDSvirus, LAV. Cell, 40, 9–17.
11. Ratner, L., Haseltine, W., Patarca, R., Livak, K. J.,Starcich, B., Josephs, S. F. et al. (1985). Completenucleotide sequence of the AIDS virus, HTLV-III.Nature, 313, 277–284.
12. Marquet, R., Isel, C., Ehresmann, C. & Ehresmann, B.(1995). tRNAs as primer of reverse transcriptases.Biochimie, 77, 113–124.
13. Mak, J. & Kleiman, L. (1997). Primer tRNAs forreverse transcription. J. Virol. 71, 8087–8095.
14. Isel, C., Ehresmann, C. &Marquet, R. (2010). Initiationof HIV reverse transcription. Viruses, 2, 213–243.
15. Isel, C., Marquet, R., Keith, G., Ehresmann, C. &Ehresman, B. (1993). Modified nucleosides of tRNALys3
modulate primer/template loop-loop interaction inthe initiation complex of HIV-1 reverse transcription.J. Biol. Chem. 268, 25269–25272.
16. Bilbille, Y., Vendeix, F. A. P., Guenther, R., Malkiewicz,A., Ariza, X., Vilarrasa, J. & Agris, P. F. (2009). Thestructure of the human tRNALys3 anticodon bound tothe HIV genome is stabilized by modified nucleosidesand adjacentmismatch base pairs.Nucleic Acids Res. 37,3342–3353.
17. Wilkinson, K. A., Gorelick, R. J., Vasa, S. M., Guex, N.,Rein, A., Mathews, D. H. et al. (2008). High-throughputSHAPE analysis reveals structures in HIV-1 genomicRNA strongly conserved across distinct biologicalstates. PLoS Biol. 6(e96), 0883–0899.
18. Jiang, M., Mak, J., Ladha, A., Cohen, E., Klein, M.,Rovinski, B. & Kleiman, L. (1993). Identification oftRNAs incorporated into wild-type and mutanthuman immunodeficiency virus type 1. J. Virol. 67,3246–3253.
19. Pavon-Eternod, M., Wei, M., Pan, T. & Kleiman, L.(2010). Profiling non-lysyl tRNAs in HIV-1. RNA, 16,267–273.
20. Cen, S., Khorchid, A., Javanbakht, H., Gabor, J., Stello,T., Shiba, K. et al. (2001). Incorporation of lysyl-tRNAsynthetase into human immunodeficiency virustype 1. J. Virol. 75, 5043–5048.
21. Druillennec, S. & Roques, B. P. (2000). HIV-1 NCp7 asa target for the design of novel antiviral agents. DrugNews Perspect. 13, 337–349.
22. Athavale, S. S., Ouyang, W., McPike, M. P., Hudson,B. S. & Borer, P. N. (2010). Effects of the nature and

713Modification-dependent Peptide Binding of tRNALys3
concentration of salt on the interaction of the HIV-1nucleocapsid protein with SL3 RNA. Biochemistry, 49,3525–3533.
23. Fisher, R. J., Rein, A., Fivash, M., Urbaneja, M. A.,Casas-Finet, J. R., Medaglia, M. & Henderson, L. E.(1998). Sequence-specific binding of human immuno-deficiency virus type 1 nucleocapsid protein to shortoligonucleotides. J. Virol. 72, 1902–1909.
24. Tisné, C., Roques, B. P. & Dardel, F. (2001). Hetero-nuclear NMR studies of the interaction of tRNA3Lys
with HIV-1 nucleocapsid protein. J. Mol. Biol. 306,443–454.
25. Tisné, C., Roques, B. P. & Dardel, F. (2003). Specificrecognition of primer tRNALys3 by HIV-1 nucleocap-sid protein: involvement of the zinc fingers and theN-terminal basic extension. Biochimie, 85, 557–561.
26. Berkowitz, R., Fisher, J. & Goff, S. P. (1996). RNApackaging.Curr. TopicsMicrobiol. Immunol. 214, 177–218.
27. Khan, R. & Giedroc, D. P. (1992). Recombinant humanimmunodeficiency virus type 1 nucleocapsid (NCp7)protein unwinds tRNA. J. Biol. Chem. 267, 6689–6695.
28. Mély, Y., de Rocquigny, H., Sorinas-Jimeno, M., Keith,G., Roques, B. P., Marquet, R. & Gérard, D. (1995).Binding of the HIV-1 nucleocapsid protein to theprimer tRNA3Lys, in vitro, is essentially not specific.J. Biol. Chem. 270, 1650–1656.
29. Hargittai, M. R. S., Mangla, A. T., Gorelick, R. J. &Musier-Forsyth, K. (2001). HIV-1 nucleocapsid proteinzinc finger structures induce tRNALys3 structuralchanges but are not critical for primer/templateannealing. J. Mol. Biol. 312, 985–997.
30. Guo, J., Wu, T., Anderson, J., Kane, B. F., Johnson,D. G., Gorelick, R. J. et al. (2000). Zinc fingerstructures in the human immunodeficiency virustype 1 nucleocapsid protein facilitate efficient minus-and plus-strand transfer. J. Virol. 74, 8980-–8988.
31. Morellet, N., de Rocquigny, H., Mély, Y., Jullian, N.,Déméné, H., Ottmann, M. et al. (1994). Conformationalbehaviour of the active and inactive forms of thenucleocapsid NCp7 of HIV-1 studied by 1H NMR.J. Mol. Biol. 235, 287–301.
32. Poon, D. T., Wu, J. & Aldovini, A. (1996). Chargedamino acid residues of human immunodeficiencyvirus type 1 nucleocapsid p7 protein involved in RNApackaging and infectivity. J. Virol. 70, 6607–6616.
33. Cusack, S. (1999). RNA-protein complexes. Curr.Opin. Struct. Biol. 9, 66–73.
34. Agris, P. F. (1996). The Importance of being modified:roles of modified nucleosides and Mg2+ in RNAstructure and function. Progr. Nucleic Acid Res. Mol.Biol. 53, 79–129.
35. Seno, T., Agris, P. F. & Söll, D. (1974). Involvement ofthe anticodon region of Escherichia coli tRNAGln andtRNAGlu in the specific interaction with cognateaminoacyl tRNA synthetase. Alteration of the2-thiouridine derivatives located in the anticodon ofthe tRNAs by BrCN or sulfur deprivation. Biochim.Biophys. Acta, 349, 328 338.
36. Madore, E., Florentz, C., Giegé, R., Sekine, S.,Yokoyama, S. & Lapointe, J. (1999). Effect of modifiednucleotides on Escherichia coli tRNAGlu structure andon its aminoacylation by glutamyl-tRNA synthetase.Predominant and distinct roles of the mnm5 and s2
modifications of U34. Eur. J. Biochem. 266, 1128–1135.
37. Ashraf, S. S., Sochacka, E., Cain, R., Guenther, R.,Malkiewicz, A. & Agris, P. F. (1999). Single atommodification (O→S) of tRNA confers ribosomebinding. RNA, 5, 188–194.
38. Yarian, C., Marszalek, M., Sochacka, E., Malkiewicz,A., Guenther, R., Miskiewicz, A. & Agris, P. F.(2000). Modified nucleoside dependent Watson-Crickand wobble codon binding by tRNALys
UUU species.Biochemistry, 39, 13390–13395.
39. Björk, G. R., Durand, J. M., Hagervall, T. G.,Leipuviene, R., Lundgren, H. K., Nilsson, K. et al.(1999). Transfer RNA modification: influence ontranslational frameshifting and metabolism. FEBSLett. 452, 47–51.
40. Stuart, J. W., Gdaniec, Z., Guenther, R., Marszalek, M.,Sochacka, E., Malkiewicz, A. & Agris, P. F. (2000).Functional anticodon architecture of human tRNALys3
includes disruption of intraloop hydrogen bonding bythe naturally occurring amino acid modification, t6A.Biochemistry, 39, 13396–13404.
41. Gustilo, E. M., Vendeix, F. A. & Agris, P. F. (2008).tRNA's modifications bring order to gene expression.Curr. Opin. Microbiol. 11, 134–140.
42. Chan, B., Wiedermaier, K., Yip, W. T., Barbara, P. F.& Musier-Forsyth, K. (1999). Intra-tRNA distancemeasurements for nucleocapsid protein-dependenttRNA unwinding during priming of HIV reversetranscription. Proc. Natl Acad. Sci. USA, 96, 459–464.
43. Barat, C., Schatz, O., Le Grice, S. & Darlix, J. L. (1993).Analysis of the interactions of HIV1 replication primertRNALys,3 with nucleocapsid protein and reversetranscriptase. J. Mol. Biol. 231, 185–190.
44. Yarian, C., Townsend, H., Czestkowski, W., Sochacka,E., Malkiewicz, A. J., Guenther, R. et al. (2002).Accurate translation of the genetic code depends ontRNA modified nucleosides. J. Biol. Chem. 277,16391–16395.
45. Hagan, N. & Fabris, D. (2003). Direct mass spectro-metric determination of the stoichiometry and bind-ing affinity of the complexes between nucleocapsidprotein and RNA stem-loop hairpins of the HIV-1 Ψ-recognition element. Biochemistry, 42, 10736–10745.
46. Avilov, S. V., Godet, J., Piémont, E. & Mély, Y. (2009).Site-specific characterization of HIV-1 nucleocapsidprotein binding to oligonucleotides with two bindingsites. Biochemistry, 48, 2422–2430.
47. Agris, P. F., Marchbank, M. T., Newman, W.,Guenther, R., Ingram, P., Swallow, J. et al. (1999).Experimental models of protein-RNA interaction:isolation and analyses of tRNA(Phe) and U1snRNA-binding peptides from bacteriophage displaylibraries. J. Protein Chem. 18, 425–435.
48. Mucha, P., Szyk, A., Rekowski, P., Weiss, P. A. &Agris, P. F. (2001). Anticodon domain methylatednucleosides of yeast tRNAPhe are significant recogni-tion determinants in the binding of a phage displayselected peptide. Biochemistry, 40, 14191–14199.
49. Eshete, M., Marchbank, M. T., Deutscher, S. L., Sproat,B., Leszczynska, G., Malkiewicz, A. & Agris, P. F.(2007). Specificity of phage display selected peptidesfor modified anticodon stem and loop domains oftRNA. Protein J. 26, 61–73.
50. Yarian, C. S., Basti, M. M., Cain, R., Ansari, G.,Guenther, R. H., Sochacka, E. et al. (1999). Structural

714 Modification-dependent Peptide Binding of tRNALys3
and functional roles of the N1- and N3-protons ofΨ attRNA's position 39. Nucleic Acids Res. 27, 3543–3549.
51. Durant, P. C. & Davis, D. R. (1999). Stabilization of theanticodon stem-loop of tRNALys,3 by an A+-C base-pair and by pseudouridine. J. Mol. Biol. 285, 115–131.
52. Fasman, G. D. (1996). Circular dichroism and theconformational analysis of biomolecules. Springer-Verlag,NY.
53. Martin, S. R. & Schilstra, M. J. (2008). Circulardichroism and its application to the study ofbiomolecules. Methods Cell Biol. 84, 263–293.
54. Cusack, S., Yaremchuk, A. & Tukalo, M. (1996).The crystal structures of T. thermophilus lysyl-tRNA synthetase complexed with E. coli tRNALys
and a T. thermophilus tRNALys transcript: anticodonrecognition and conformational changes uponbinding of a lysyl-adenylate analogue. EMBO J.15, 6321–6334.
55. Francklyn, C., Musier-Forsyth, K. & Martinis, S. A.(1997). Aminoacyl-tRNA synthetases in biology anddisease: new evidence for structural and functionaldiversity in an ancient family of enzymes. RNA, 3,954–960.
56. Bampi, C., Jacquenet, S., Lener, D., Décimo, D. &Darlix, J. L. (2004). The chaperoning and assistanceroles of the HIV-1 nucleocapsid protein in proviralDNA synthesis and maintenance. Curr. HIV Res. 2,79–92.
57. Huang, Y., Mak, J., Cao, Q., Li, Z., Wainberg, M. A. &Kleiman, L. (1994). Incorporation of excess wild typeand mutant tRNALys3 into human immunodeficiencyvirus type 1. J. Virol. 68, 7676–7683.
58. Barraud, P., Gaudin, C., Dardel, F. & Tisné, C.(2007). New insights into the formation of HIV-1reverse transcription initiation complex. Biochimie,89, 1204–1210.
59. Urbaneja, M. A., Kane, B. P., Johnson, D. J., Gorelick,R. J., Henderson, L. E. & Casas-Finet, J. R. (1999).Binding properties of the human immunodeficiencyvirus type 1 nucleocapside protein p7 to a modelRNA: Elucidation of the structural determinants forfunction. J. Mol. Biol. 287, 59–75.
60. De Guzman, R. N., Wu, Z. R., Stalling, C. C.,Pappalardo, L., Borer, P. N. & Summers, M. F.(1998). Structure of the HIV-1 nucleocapsid proteinbound to the SL3Ψ-RNA recognition element. Science,279, 384–388.
61. Amarasinghe, G. K., De Guzman, R. N., Turner, R. B.,Chancellor, K. J., Wu, Z. R. & Summers, M. F. (2000).NMR structure of the HIV-1 nucleocapsid proteinbound to stem-loop SL2 of the psi-RNA packagingsignal. Implications for genome recognition. J. Mol.Biol. 301, 491–511.
62. Amarasinghe, G. K., Zhou, J., Miskimon, M.,Chancellor, K. J., McDonald, J. A., Matthew, A. G.et al. (2001). Stem-loop SL4 of the HIV-1 Ψ RNApackaging signal exhibits weak affinity for thenucleocapsid protein. Structural studies and impli-cations for genome recognition. J. Mol. Biol. 314,961–970.
63. Cantara, W. A., Crain, P. F., Rozenski, J., McCloskey,J. A., Harris, K. A., Zhang, X. et al. (2011). The RNAmodification database, RNAMDB: 2011 update.Nucleic Acids Res. 39, D195–D201.
64. Jühling, F., Mörl, M., Hartmann, R. K., Sprinzl, M.,Stadler, P. F. & Pütz, J. (2009). tRNAdb 2009:compilation of tRNA sequences and tRNA genes.Nucleic Acids Res. 37, D159–D162.
65. Wakefield, J. K., Wolf, A. G. & Morrow, C. D. (1995).Human immunodeficiency virus type 1 can usedifferent tRNAs as primers for reverse transcriptionbut selectively maintains a primer binding sitecomplementary to tRNA3Lys. J. Virol. 69, 6021–6029.
66. Gregoire, C. J., Gautheret, D. & Loret, E. P. (1997). NotRNA3Lys unwinding in a complex with HIV NCp7.J. Biol. Chem. 272, 25143–25148.
67. Huthoff, H. & Berkhout, B. (2001). Two alter-nating structures of the HIV-1 leader RNA. RNA, 7,143–157.
68. Zeng, Y., Liu, H. W., Landes, C. F., Kim, Y. J., Ma, X.,Zhu, Y. et al. (2007). Probing nucleation, reverseannealing, and chaperone function along the reactionpath of HIV-1 single-strand transfer. Proc. Natl Acad.Sci. USA, 104, 12651–12656.
69. Wu, H., Rouzina, I. & Williams, M. C. (2010). Single-molecule stretching studies of RNA chaperones. RNABiol. 7, 73–84.
70. My-Nuong Vo, M. N., Barany, G., Rouzina, I. &Musier-Forsyth, K. (2006). Mechanistic studies ofmini-TAR RNA/DNA annealing in the absence andpresence of HIV-1 nucleocapsid protein. J. Mol. Biol.363, 244–261.
71. Cléry, A., Blatter, M. & Allain, F. H. (2008). RNArecognition motifs: boring? Not quite. Curr. Opin.Struct. Biol. 18, 290–298.
72. Davidson, A., Leeper, T. C., Athanassiou, Z.,Patora-Komisarska, K., Karn, J., Robinson, J. A. &Varani, G. (2009). Simultaneous recognition of HIV-1TAR RNA bulge and loop sequences by cyclicpeptide mimics of Tat protein. Proc. Natl Acad. Sci.USA, 106, 11931–11936.
73. Michael, L. A., Chenault, J. A., Miller, B. R., 3rd,Knolhoff, A. M. & Nagan, M. C. (2009). Water, shaperecognition, salt bridges, and cation-pi interactionsdifferentiate peptide recognition of the HIV rev-responsive element. J. Mol. Biol. 392, 774–786.
74. Hofstadler, S. A., Sannes-Lowery, K. A. & Hannis,J. C. (2005). Analysis of nucleic acids by FTICR MS.Mass Spectrom. Rev. 24, 265–285.
75. Turner, K. B., Brinson, R. G., Yi-Brunozzi, H. Y.,Rausch, J. W., Miller, J. T., Le Grice, S. F. et al. (2008).Structural probing of the HIV-1 polypurine tractRNA:DNA hybrid using classic nucleic acid ligands.Nucleic Acids Res. 36, 2799–2810.
76. Hagan, N. A. & Fabris, D. (2007). Dissecting theprotein-RNA and RNA-RNA interactions in thenucleocapsid-mediated dimerization and isomeri-zation of HIV-1 stem loop 1. J. Mol. Biol. 365,396–410.
77. Agris, P. F., Vendeix, F. A. P. & Graham, W. D. (2007).tRNA's wobble decoding of the genome: 40 years ofmodification. J. Mol. Biol. 366, 1–13.
78. Hartsel, S. A., Kitchen, D. E., Scaringe, S. A. &Marshall, W. S. (2005). RNA oligonucleotide synthesisvia 5′-silyl-2′-orthoester chemistry. Methods Mol. Biol.288, 33–50.
79. Agris, P. F., Malkiewicz, A., Kraszewski, A., Everett,K., Nawrot, B., Sochacka, E. et al. (1995). Site-selected

715Modification-dependent Peptide Binding of tRNALys3
introduction of modified purine and pyrimidineribonucleosides into RNA by automated phosphor-amidite chemistry. Biochimie, 77, 125–134.
80. Ogilvie, K. K., Usman, N., Nicoghosian, K. &Cedergren, R. J. (1988). Total chemical synthesis of a77-nucleotide-long RNA sequence having methio-nine-acceptance activity. Proc. Natl Acad. Sci. USA,85, 5764–5768.
81. Guenther, R. H., Gopal, D. H. & Agris, P. F.(1988). Purification of transfer RNA species bysingle-step ion-exchange high performance liquidchromatography. J. Chromatogr. 444, 79–87.
82. Gehrke, C. W., Kuo, K. C., McCune, R. A.,Gerhardt, K. O. & Agris, P. F. (1982). Quantitativeenzymatic hydrolysis of tRNAs: reversed-phasehigh-performance liquid chromatography of tRNAnucleosides. J. Chromatogr. 230, 297–308.
83. Lee, B. M., De Guzman, R. N., Turner, B. G., Tjandra,N. & Summers, M. F. (1998). Dynamical behavior of
the HIV-1 nucleocapsid protein. J. Mol. Biol. 279,633–649.
84. Cheadle, C., Ivashchenko, Y., South, V., Searfoss, G. H.,French, S., Howk, R. et al. (1994). Identification of a SrcSH3 domain binding motif by screening a randomphage display library. J. Biol. Chem. 269, 24034–24039.
85. Chen, L., Wang, Y., Liu, X., Dou, S., Liu, G.,Hnatowich, D. J. & Rusckowski, M. (2008). A newTAG-72 cancer marker peptide identified by phagedisplay. Cancer Lett. 272, 122–132.
86. Scheunemann, A. E., Graham, W. D., Vendeix, F. A. P.& Agris, P. F. (2010). Binding of aminoglycosideantibiotics to helix 69 of 23S rRNA. Nucleic Acids Res.38, 3094–3105.
87. Gagnon, K. T., Zhang, X., Agris, P. F. & Maxwell,E. S. (2006). Assembly of the archaeal box C/D sRNPcan occur via alternative pathways and requirestemperature-facilitated sRNA remodeling. J. Mol. Biol.362, 1025–1042.




















