
Arginase I in myeloid suppressor cells is induced by COX2 in lung carcinoma

FD
SB
*M
Bp((AtilbCciadmwCti�aCatnsHotafmowCC
Ctsft
GASTROENTEROLOGY 2007;132:1055–1065
unctional Antagonism Between RNA Binding Proteins HuR and CUGBP2etermines the Fate of COX-2 mRNA Translation
RIPATHI M. SUREBAN,* NABENDU MURMU,* PAVEL RODRIGUEZ,‡ RANDAL MAY,* REETI MAHESHWARI,‡
RIAN K. DIECKGRAEFE,‡ COURTNEY W. HOUCHEN,* and SHRIKANT ANANT*,§
Department of Medicine and §Department of Cell Biology, University of Oklahoma Health Science Center, Oklahoma City, Oklahoma, and the ‡Department of
edicine, Washington University School of Medicine, St. Louis, Missourintttsmictl
mMh(ti2eHi
ibtABcsgcsiapi
ct
BA
SIC–
ALI
MEN
TARY
TRA
CT
ackground & Aims: Cyclooxygenase-2 (COX-2) ex-ression is regulated at the levels of messenger RNAmRNA) stability and translation by AU-rich elementsARE) located in its 3= untranslated region (3=UTR).lthough structurally homologous RNA binding pro-
eins HuR and CUGBP2 stabilize COX-2 mRNA, HuRnduces whereas CUGBP2 inhibits COX-2 mRNA trans-ation. This study aimed to determine the antagonismetween these proteins on COX-2 expression. Methods:OX-2 ARE binding activity was determined by nitro-
ellulose filter binding and UV cross-linking assays us-ng recombinant glutathione S-transferase (GST)/HuRnd GST/CUGBP2. Protein:protein interactions wereetermined by GST pull-down, yeast 2-hybrid, and im-unocytochemistry assays. Nucleocytoplasmic shuttingas determined by heterokaryon analyses. The effect ofUGBP2 and HuR on COX-2 ARE-dependent transla-
ion was shown by a chimeric luciferase mRNA contain-ng COX-2 3=UTR. HT-29 cells were subjected to 12 Gy-irradiation in a cesium irradiator. Results: CUGBP2nd HuR bind with similar affinities to COX-2 ARE, butUGBP2 competes with HuR for binding. In vitro, HuRnd CUGBP2 heterodimerize. Furthermore, FLAG-agged HuR and myc-tagged CUGBP2 colocalize in theucleus of HCT-116 cells. Moreover, both proteinshuttled between the nucleus and cytoplasm. In vitro,uR enhanced whereas CUGBP2 inhibited translation
f the chimeric luciferase COX-2 3=UTR mRNA. Fur-hermore, CUGBP2 competitively inhibited HuR-medi-ted translation of the transcript. In HT-29 cells trans-ected with HuR and CUGBP2, a switch in COX-2
RNA binding from predominantly HuR to CUGBP2ccurred after radiation treatment, which was coupledith increased silencing of the COX-2 mRNA.onclusions: CUGBP2 overrides HuR and suppressesOX-2 mRNA translation.
yclooxygenase-2 (COX-2), an immediate early re-sponse gene, is a rate-limiting enzyme in the pros-
aglandin synthesis pathway.1 Increased COX-2 expres-ion occurs in response to cytokines and mitogenicactors such as interleukin-1� and epithelial growth fac-
or.2 COX-2 is overexpressed early in epithelial malig-ancy and colon cancer, indicating its importance inumorigenesis.3 In addition to being regulated at theranscriptional level, COX-2 expression is controlled athe posttranscriptional levels of messenger RNA (mRNA)tability and translation.4,5 In normal cells, COX-2
RNA not only is transcribed at a very low rate, but alsos degraded rapidly.6 On the other hand, in transformedells, COX-2 mRNA levels are up-regulated significantlyhrough both increased transcription rate and higherevels of mRNA stability.7
The 3= untranslated region (3=UTR) region of COX-2RNA contains several AUUUA destabilizing motifs.ore importantly, the first 60 nucleotides of the 3=UTR
as a cluster of the nanomer AU-rich elements (ARE)[U/A][A/U]UUU[A/U][U/A]), which is present in bothranscripts.8,9 The half-life of COX-2 mRNA is short, buts increased by various RNA binding proteins.10 There are
groups of RNA binding proteins, one that increases thexpression of COX-2 such as hnRNP A1, apobec-1, anduR, and a second group that inhibits COX-2 expression
ncluding CUGBP2, TIA1, TTP, TIAR, and AUF1.11–13
HuR, identified as specific tumor antigens in cancers ofndividuals with paraneoplastic neurologic disorder, sta-ilizes rapidly degraded transcripts through its bindingo the ARE in the 3=-UTR.14,15 HuR can stabilize manyRE-encoding mRNAs including COX-2, cyclin A, cyclin1, and proliferation-associated genes such as c-fos, vas-ular endothelial growth factor (VEGF), and c-myc, andubsequently increase the protein levels of theseenes.14,16,17 Originally, HuR was only thought to in-rease translation. However, more recently it has beenhown that HuR overexpression in myeloid cells resultedn translational silencing of specific inflammatory medi-tors through interactions with a translation inhibitorrotein TIA-1, suggesting HuR function may depend on
ts binding partners.18
Abbreviations used in this paper: ARE, AU-rich element; COX-2,yclooxygenase-2; GST, glutathione S-transferase; RRM, RNA recogni-ion motif; SDS, sodium dodecyl sulfate; 3=UTR, 3= untranslated region.
© 2007 by the AGA Institute0016-5085/07/$32.00
doi:10.1053/j.gastro.2006.12.031

Ei3iHnacbRnib
Ctpiw3bpsaivsuH
ncldUbmwt
apmabPmEp
8eNgp(XcE(cAmt
dt3
(tnBb1GpbcCC
aCbmwtctmrdl(
gcFH
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1056 SUREBAN ET AL GASTROENTEROLOGY Vol. 132, No. 3
We also previously showed that CUGBP2 (also calledTR-3, Brunol, and Napor) increases COX-2 mRNA stabil-
ty after binding to the ARE sequences in the COX-2=UTR.12 However, in contrast to HuR, CUGBP2 inhib-
ted translation of COX-2 mRNA. Both CUGBP2 anduR encode 3 RNA binding domains of the RNA recog-ition motif (RRM) type, 2 located at the amino terminalnd another one located at the carboxy terminal.14,19 Aritical difference between CUGBP2 and HuR is in theridge region located between the second and thirdRMs. This region is extended in CUGBP2 and is ala-ine- and glutamine-rich. The role of this bridge region
n CUGBP2 currently is not well understood, but iselieved to encode a nuclear localization signal.20
Given that HuR enhances, whereas CUGBP2 inhibits,OX-2 mRNA translation, it suggests opposing func-
ions in regulating COX-2 mRNA translation for the 2roteins. To identify this, we first performed RNA bind-
ng studies. The results revealed that the 2 proteins bindith similar affinities to the ARE sequences in COX-2=UTR. However, CUGBP2 competed with HuR for theinding. More importantly, our results showed a directhysical interaction between the 2 proteins, which re-ulted in colocalization of the proteins in the nucleus. Welso showed that ectopic expression of CUGBP2 proteins sufficient to block cellular mRNA translation by pre-enting HuR binding to the COX-2 mRNA. These resultsupport a novel mechanism of translation inhibitionsed by CUGBP2, which complements the blockage ofuR binding.
Materials and MethodsCell CultureHCT-116, HT-29 (both human colon adenocarci-
omas), and NIH-3T3 (mouse immortalized fibroblast)ells were obtained from the American type culture col-ection and grown in Dulbecco’s modified Eagle’s me-ium supplemented with 10% fetal bovine serum and 100/mL of penicillin-streptomycin in a humidified cham-er at 37°C with 5% CO2. Cells were incubated with 100g/mL of cycloheximide (Sigma, St. Louis, MO) alongith the media, where indicated, to inhibit protein syn-
hesis.
Recombinant Protein Expression andPlasmid ConstructsRecombinant CUGBP2 and HuR were expressed
s N-terminal glutathione S-transferase (GST) fusionroteins from the plasmid pGEX-4T3 (Amersham-Phar-acia, Piscataway, NJ), and purified using glutathione-
garose beads (Sigma). The proteins were purified furthery Sephadex G-75 column chromatography (Amersham-harmacia), and dialyzed against a buffer containing 20mol/L HEPES (pH 7.9), 0.1 mol/L KCl, 0.2 mmol/L
DTA, 0.5 mmol/L dithiothreitol, and 20% glycerol. The
urity of the product was determined to be greater than w5% by sodium dodecyl sulfate (SDS)–polyacrylamide gellectrophoresis analyses.21 For mammalian expression,-terminal FLAG and Myc epitope-tagged CUGBP2 were
enerated by cloning the full-length coding region intolasmid pCMV-Tag 2B (for FLAG) and pCMV-Tag 3B
for Myc) (Invitrogen, Carlsbad, CA) at the Hind III andho1 restriction sites. Myc-tagged HuR was generated byloning the HuR complementary DNA (cDNA) betweencoR I and Sal I restriction sites in plasmid pCMV-Tag 3B
Invitrogen). The full-length (FL) 2.2 kb COX-2 3=UTRloned in the pGL3 basic vector was a generous gift fromubrey Morrison.9 Renilla luciferase reporter gene plas-id pRL-TK (Promega, Madison, WI) was used as con-
rol.
Nitrocellulose Filter-Binding AssaysRecombinant GST/CUGBP2 and GST/HuR were
iluted to 500 ng in 10 �L of binding buffer and addedo 20 �L of binding buffer containing 1.5 nmol/L2P-labeled COX-2 3=UTR complementary RNA (cRNA)15,000 cpm), and incubated for 20 minutes at roomemperature. The mixture then was passed through aitrocellulose filter (HAWP, 0.45-�m pore size; Millipore,illerica, MA) that was pre-equilibrated with bindinguffer, and analyzed by scintillation spectroscopy (model500; Packard, PerkinElmer, Boston, MA) using Ultimaold scintillation cocktail (Packard, PerkinElmer). Eachoint in the binding curve represents the average ofinding reactions performed in triplicate. Dissociationonstants were determined as previously described.22
ontrol experiments revealed that GST alone showed noOX-2 RNA binding activity (data not shown).
UV Cross-LinkingRecombinant GST/CUGBP2 and GST/HuR were
dded at the indicated concentrations to a 32P-labeledOX-2 3=UTR cRNA template (50,000 cpm) in a bindinguffer containing 20 mmol/L HEPES (pH 7.9), 100mol/L KCl, and 1 mmol/L dithiothreitol. The mixtureas incubated for 20 minutes at room temperature and
hen treated sequentially with RNase T1 (1 U/mL finaloncentration) and heparin (5 mg/mL final concentra-ion). The mixture was subjected to UV cross-linking (250
J/cm2) in a Stratalinker (Stratagene, La Jolla, CA) andesolved by SDS–polyacrylamide gel electrophoresis un-er reducing conditions. The intensity of the bound
abeled RNA was determined by Phosphoimager analysismodel PSI; Amersham-Pharmacia).23
Heterokaryon AssayInterspecies human-mouse heterokaryons were
enerated as described previously24 with minor modifi-ations. CUGBP2 and HuR were expressed as N-terminalLAG-tagged proteins from the plasmid pCMVTag-2B.CT-116 cells grown on 35-mm dishes were transfected
ith 2 �g of each plasmid using Fugene 6 (Roche, Alam-
ewdNmsfay3dw
agDggGcdntnMecgudAJ
wmtPopptewbtmwbm8S
Pc
bNrtita2vbap
3dhfIlRtttunmrve
ltCgspSotsvrCIiW
BA
SIC–
ALI
MEN
TARY
TRA
CT
March 2007 CUGBP2 INHIBITS HUR ACTIVITY 1057
da, CA). Twenty-four hours after transfection, the cellsere trypsinized and seeded onto coverslips in 35-mmishes at 3 � 105 cells/coverslip and incubated overnight.IH-3T3 cells, preincubated for 30 minutes with 100g/mL cycloheximide were seeded onto the same cover-
lips at 3 � 105 cells/coverslip The cells were coculturedor 3 hours in the presence of 100 mg/mL cycloheximide,nd then allowed to fuse in the presence of 50% polyeth-lene glycol 3350/phosphate-buffered saline (Sigma) forminutes. After 3 hours of recovery in the growth me-
ium containing 100 mg/mL cycloheximide, the cellsere fixed and processed for immunofluorescence.
ImmunofluorescenceCells were fixed with 10% formalin solution (Sigma),
nd subsequently probed with rabbit anti-FLAG immuno-lobulin (Ig)G and/or mouse monoclonal anti-myc IgG.etection was performed with secondary antibody conju-
ated to Cy3-, Cy5-, or fluorescein isothiocyanate–conju-ated secondary IgG (Jackson ImmunoResearch, Westrove, PA). For standard immunofluorescence micros-
opy, nuclei were identified by 4,6-diamidino-2-phenylin-ole. For the HCT-116-NIH-3T3 heterokaryon assay, theuclei were stained with Hoechst dye 33342 (Invitrogen)o permit discrimination between human and murineuclei. Slides were examined using a Zeiss Axiaskop 2OT plus microscope (Carl Zeiss, Inc., Thornwood, NY)
quipped with a 40� plan neofluar objective and aharged coupled device camera (DAGE-MTI Inc., Michi-an City, IN). A Zeiss Attoarc variable intensity lamp wassed with filter sets designed for Cy3, Cy5, and 4,6-iamidino-2-phenylindole. Images were processed usingdobe Photoshop 8.0 software (Adobe Systems Inc., San
ose, CA).
In Vitro Protein–Protein Interaction AssayFLAG epitope-tagged CUGBP2 and HuR proteins
ere synthesized and metabolically labeled with 35S-ethionine in a coupled transcription/translation sys-
em (TNT T7/T3 Coupled Reticulocyte Lysate System;romega). T7 RNA polymerase was used for productionf CUGBP2 from the pCMV Tag 2B vector, and T3 RNAolymerase was used for generating HuR from theCDNA vector. The radiolabeled proteins were allowedo interact at 4°C for 2 hours with equimolar amounts ofither recombinant GST/CUGBP2 or GST/HuR thatere previously immobilized on glutathione–sepharoseeads (Amersham-Pharmacia) in a binding buffer con-aining 15 mmol/L HEPES, pH 7.9, 50 mmol/L KCl, 0.1
mol/L EDTA, and 0.2% Triton X-100 supplementedith 200 mmol/L NaCl. After washing the beads, theound proteins were eluted using a buffer containing 10mol/L reduced glutathione in 50 mmol/L Tris-HCl (pH
.0). The eluent and the washings were separated in an
DS–polyacrylamide gel electrophoresis and subjected to ahosphoimager analysis (model PSI, Amersham-Pharma-ia).25
Yeast 2-Hybrid InteractionsHuR and CUGBP2 cDNAs were inserted into the
ait plasmid pGBKT7 or prey plasmid pGADT7 betweende1 and Sal1, and EcoR1 and Xho1 restriction sites,
espectively (both plasmids from BD Bioscience Clon-ech, Palo Alto, CA). The plasmids were cotransformednto Saccharomyces cerevisiae AH 109, and synthetic dex-rose minimal media plates lacking -His, -Leu, and -Trp,nd containing X-�-Gal. Positive interaction between theproteins results in production of �-galactosidase, con-
erting X-�-gal to a blue product, thereby resulting inlue-colored colonies. To show positive control for inter-ction, p53 in pGBKT7 and SV40 T large antigen inGADT7 interaction was used.
Luciferase Reporter Gene AssayThe 2.2-kb full-length region of the COX-2
=UTR, cloned into plasmid pGL3-Basic at the Xba1 siteownstream of the firefly (Photinus pyralis) luciferase gene,as been described previously.26 HT-29 cells were trans-
ected transiently using FuGENE 6 (Roche, Indianapolis,N) with the luciferase construct along with the Renillauciferase expressing plasmid pRL-TK (Promega). TotalNA from the cells was isolated and translated in vitro in
he presence of the recombinant proteins. Luciferase ac-ivity was determined as per the manufacturer’s instruc-ions (Dual-Luciferase Reporter Assay System; Promega)sing a Monolight 2010 luminometer (Analytical Lumi-escence Laboratory, San Diego, CA). The activity, nor-alized to Renilla Luciferase activity, is presented as
elative luciferase units relative to control with � SEMalues. Assays were performed in triplicate wells andxperiments were repeated 2 times.
ImmunoprecipitationHT-29 cells were transfected transiently with the
uciferase construct and plasmid constructs expressinghe HA-tagged CUGBP2 and FLAG-tagged HuR.UGBP2 was cloned in the plasmid pHOOK-2 (Invitro-en) at the Hind III and Xho1 restriction sites for expres-ion as a carboxy terminal HA epitope-tagged fusionrotein. The cells were processed as described earlier.27
onicated lysates were immunoprecipitated with anti-HAr anti-FLAG IgG using the Seize X protein A purifica-ion kit (Pierce, Rockford, IL). The pellet and supernatantubsequently were incubated at 70°C for 1 hour to re-erse the cross-links. RNA was isolated and subjected toeverse-transcription polymerase chain reaction forOX-2 and glyceraldehyde-3-phosphate dehydrogenase.
n addition, the lysates were subjected to luciferase activ-ty measurements using the dual luciferase assay kit.
estern blot analyses for COX-2 protein were performed
s previously described.12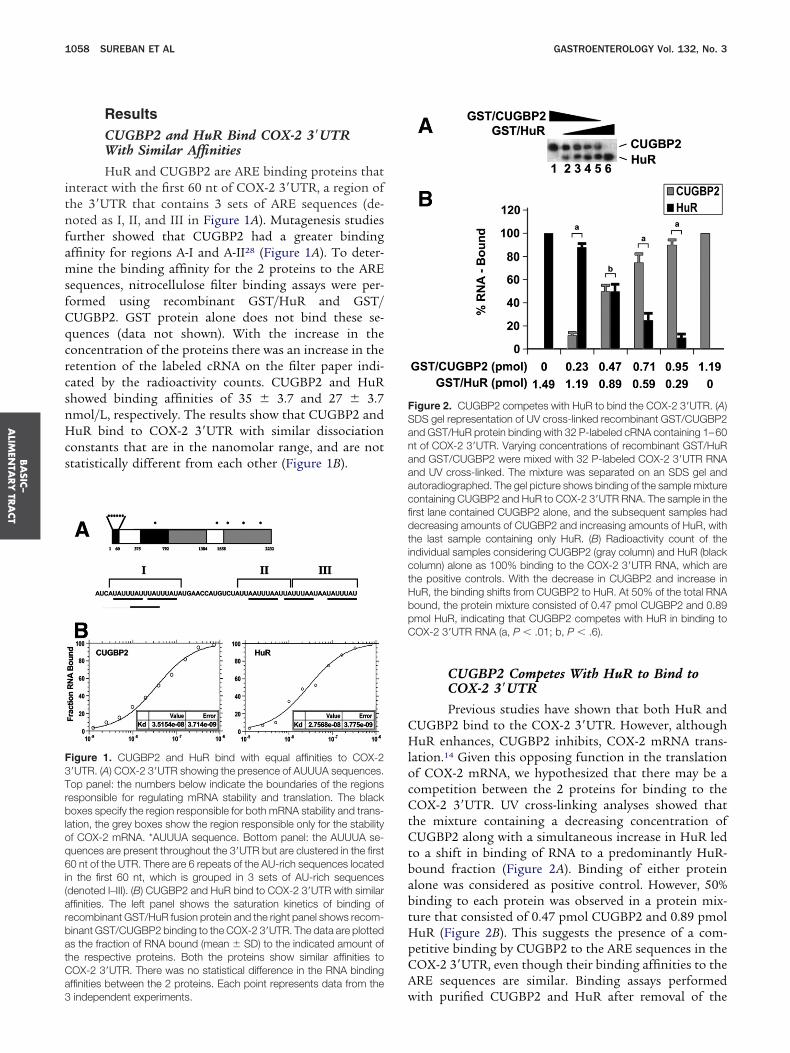
itnfamsfCqcrcsnHcs
CHlocCtCtbabtHpCA
F3Trbloq6i(arbatCa3
FSanaaacfidtictHbpC
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1058 SUREBAN ET AL GASTROENTEROLOGY Vol. 132, No. 3
ResultsCUGBP2 and HuR Bind COX-2 3=UTRWith Similar AffinitiesHuR and CUGBP2 are ARE binding proteins that
nteract with the first 60 nt of COX-2 3=UTR, a region ofhe 3=UTR that contains 3 sets of ARE sequences (de-oted as I, II, and III in Figure 1A). Mutagenesis studies
urther showed that CUGBP2 had a greater bindingffinity for regions A-I and A-II28 (Figure 1A). To deter-ine the binding affinity for the 2 proteins to the ARE
equences, nitrocellulose filter binding assays were per-ormed using recombinant GST/HuR and GST/UGBP2. GST protein alone does not bind these se-uences (data not shown). With the increase in theoncentration of the proteins there was an increase in theetention of the labeled cRNA on the filter paper indi-ated by the radioactivity counts. CUGBP2 and HuRhowed binding affinities of 35 � 3.7 and 27 � 3.7mol/L, respectively. The results show that CUGBP2 anduR bind to COX-2 3=UTR with similar dissociation
onstants that are in the nanomolar range, and are nottatistically different from each other (Figure 1B).
igure 1. CUGBP2 and HuR bind with equal affinities to COX-2=UTR. (A) COX-2 3=UTR showing the presence of AUUUA sequences.op panel: the numbers below indicate the boundaries of the regionsesponsible for regulating mRNA stability and translation. The blackoxes specify the region responsible for both mRNA stability and trans-
ation, the grey boxes show the region responsible only for the stabilityf COX-2 mRNA. *AUUUA sequence. Bottom panel: the AUUUA se-uences are present throughout the 3=UTR but are clustered in the first0 nt of the UTR. There are 6 repeats of the AU-rich sequences located
n the first 60 nt, which is grouped in 3 sets of AU-rich sequencesdenoted I–III). (B) CUGBP2 and HuR bind to COX-2 3=UTR with similarffinities. The left panel shows the saturation kinetics of binding ofecombinant GST/HuR fusion protein and the right panel shows recom-inant GST/CUGBP2 binding to the COX-2 3=UTR. The data are plotteds the fraction of RNA bound (mean � SD) to the indicated amount ofhe respective proteins. Both the proteins show similar affinities toOX-2 3=UTR. There was no statistical difference in the RNA bindingffinities between the 2 proteins. Each point represents data from the
windependent experiments.
CUGBP2 Competes With HuR to Bind toCOX-2 3=UTRPrevious studies have shown that both HuR and
UGBP2 bind to the COX-2 3=UTR. However, althoughuR enhances, CUGBP2 inhibits, COX-2 mRNA trans-
ation.14 Given this opposing function in the translationf COX-2 mRNA, we hypothesized that there may be aompetition between the 2 proteins for binding to theOX-2 3=UTR. UV cross-linking analyses showed that
he mixture containing a decreasing concentration ofUGBP2 along with a simultaneous increase in HuR led
o a shift in binding of RNA to a predominantly HuR-ound fraction (Figure 2A). Binding of either proteinlone was considered as positive control. However, 50%inding to each protein was observed in a protein mix-ure that consisted of 0.47 pmol CUGBP2 and 0.89 pmoluR (Figure 2B). This suggests the presence of a com-etitive binding by CUGBP2 to the ARE sequences in theOX-2 3=UTR, even though their binding affinities to theRE sequences are similar. Binding assays performed
igure 2. CUGBP2 competes with HuR to bind the COX-2 3=UTR. (A)DS gel representation of UV cross-linked recombinant GST/CUGBP2nd GST/HuR protein binding with 32 P-labeled cRNA containing 1–60t of COX-2 3=UTR. Varying concentrations of recombinant GST/HuRnd GST/CUGBP2 were mixed with 32 P-labeled COX-2 3=UTR RNAnd UV cross-linked. The mixture was separated on an SDS gel andutoradiographed. The gel picture shows binding of the sample mixtureontaining CUGBP2 and HuR to COX-2 3=UTR RNA. The sample in therst lane contained CUGBP2 alone, and the subsequent samples hadecreasing amounts of CUGBP2 and increasing amounts of HuR, withhe last sample containing only HuR. (B) Radioactivity count of thendividual samples considering CUGBP2 (gray column) and HuR (blackolumn) alone as 100% binding to the COX-2 3=UTR RNA, which arehe positive controls. With the decrease in CUGBP2 and increase inuR, the binding shifts from CUGBP2 to HuR. At 50% of the total RNAound, the protein mixture consisted of 0.47 pmol CUGBP2 and 0.89mol HuR, indicating that CUGBP2 competes with HuR in binding toOX-2 3=UTR RNA (a, P � .01; b, P � .6).
ith purified CUGBP2 and HuR after removal of the

Gsa
cfctHpwatit1c(tta
tHips
mCdoGwHCoHfbsCyeotLadrgtpftfoitC
FmHftFcwcewssrt3atbit
BA
SIC–
ALI
MEN
TARY
TRA
CT
March 2007 CUGBP2 INHIBITS HUR ACTIVITY 1059
ST moiety by thrombin cleavage showed similar results,uggesting that the presence of the GST moiety did notffect the binding activity (data not shown).
Both HuR and CUGBP2 Shuttle BetweenNucleus and CytoplasmHuR has been shown to shuttle between the nu-
leus and cytoplasm.29 Furthermore, CUGBP2 relocalizesrom the nucleus to the cytoplasm after exposure of theells to ionizing radiation.12 To evaluate CUGBP2 shut-ling, we performed the heterokaryon assay in whichCT-116 intestinal epithelial cells, transfected with thelasmid-expressing FLAG-tagged CUGBP2, was fusedith mouse NIH-3T3 cells. As a positive control, studieslso were performed with FLAG-tagged HuR. Presence ofhe CUGBP2 in the mouse nuclei after fusion wouldndicate that the protein relocalized from one nucleus tohe other by going through the cytoplasm. When HCT-16 cells were fused with NIH-3T3 cells in the presence ofyclohexamide to prevent protein synthesis, both HuRFigure 3A) and CUGBP2 (Figure 3D) were detected inhe murine nuclei in the heterokaryons. This suggestshat both HuR and CUGBP2 shuttle between the nucleusnd cytoplasm in intestinal epithelial cells.
HuR and CUGBP2 Bind to Each Other andColocalize In VivoBecause many RNA binding proteins are known
o heterodimerize with other proteins, we predicted thatuR and CUGBP2 might interact. Furthermore, the sim-
larity in the shuttling activity between the 2 proteins, cou-led with the competition for binding to the COX-2 3=UTR,uggests a functional antagonism through heterodimer for-
igure 3. Nuclear-cytoplas-ic shuttling of CUGBP2 anduR. HCT-116 cells, trans-
ected with N-terminal FLAG-agged CUGBP2 or N-terminalLAG-tagged HuR (as positiveontrol for shuttling) was fusedith untransfected NIH-3T3ells in the presence of poly-thylene glycol. Cycloheximideas added to the cells before fu-ion to prevent de novo proteinynthesis. (A, D) Presence of theespective proteins. (B, E) Dis-inct punctuated staining of NIH-T3 mouse nuclei (indicated byn arrow) with Hoechst 33342;he HCT-116 cells stain uniformlue. (C, F) Merged images. Sim-
lar to HuR, CUGBP2 in the both
he human and mouse nuclei.ation between the 2 proteins. To determine whether theUGBP2 interacts with HuR, we performed a GST pull-own assay. HuR was generated in vitro in the presencef 35S-methionine, and incubated with recombinantST/HuR or GST/CUGBP2, followed by purificationith a glutathione-sepharose column. The 36-kilodaltonuR protein interacted with both GST/HuR and GST/UGBP2 (Figure 4A). However, no such interaction wasbserved with GST alone (data not shown) or when theuR was added directly to the column without the GST
usion proteins (Figure 4A). This implies that HuR canoth homodimerize and heterodimerize with CUGBP2 inolution, in the absence of any RNA. To test whetherUGBP2 and HuR also interact in vivo, we performed aeast 2-hybrid analysis. Both HuR and CUGBP2 werexpressed as fusion proteins with either the DNA bindingr activation domains of Gal4. Positive interaction be-ween the 2 proteins resulted in the expression of HIS3,eu, and Trp, thereby allowing growth of the yeast cell intriple drop-out media (Figure 4B). In addition, it in-
uced expression of MEL1 gene encoding �-galactosidase,esulting in blue colonies after hydrolysis of the chromo-enic substrate X-�-Gal. Interaction between SV40 T an-igen and tumor-suppressor protein p53 was used asositive control for the studies. However, neither proteinormed colonies with either HuR or CUGBP2, suggestinghe specificity of interaction (Figure 4B). To confirmurther that the interaction between HuR and CUGBP2ccurs in mammalian cells, immunocytochemistry exper-
ments were performed. HCT-116 cells were transfectedransiently with N-terminal myc epitope–taggedUGBP2 and N-terminal FLAG epitope–tagged HuR. Co-

lippwidd
itte3Cw
tclddhttcui3ttteH
FGfr3Gtp2gapCl nylind
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1060 SUREBAN ET AL GASTROENTEROLOGY Vol. 132, No. 3
ocalization was observed by immunofluorescence stain-ng of the cells with anti-myc monoclonal and anti-FLAGolyclonal antibodies. The protein complex was localizedredominantly in the nucleus, but some localization alsoas observed in the cytoplasm (Figure 4C). These find-
ngs show that the HuR and CUGBP2 proteins formimeric complexes in vitro, and have the potential toimerize in vivo.
CUGBP2 Overrides HuR and Inhibits COX-2mRNA TranslationGiven that CUGBP2 and HuR compete for bind-
ng to the COX-2 3=UTR, we next determined whetherhis competition affected mRNA translation. In vitroranslation assays were performed with a chimeric lucif-rase mRNA that contained the full-length COX-2=UTR (Figure 5A). As controls, the effect of HuR andUGBP2 on luciferase mRNA without the COX-2 3=UTR
igure 4. CUGBP2 and HuR heterodimerize. (A) GST pull-down assST/HuR and GST/CUGBP2 proteins, and purified over a column con
rom column incubated with the 35 S-labeled HuR alone used as negaecombinant proteins were separated on an SDS–polyacrylamide gel el5 S-HuR in the samples bound with recombinant GST/HuR, indicatinST/CUGBP2, indicating that HuR heterodimerizes with CUGBP2. The
ative of 3 experiments. (B) Yeast 2-hybrid assay. HuR and CUGBP2 cDrotein with the Gal4 DNA binding domain and Gal4 DNA activation domproteins known to interact, were expressed with the Gal4 DNA bindinrowth of the cell, and also in blue colonies in the presence of X-�-Gal ond CUGBP2 homodimerized and heterodimerized to form blue coloniroteins, showing specificity of the interaction in the analyses (negative cUGBP2 were co-expressed in HCT-116 cells and stained with Cy3 (
abeling are in yellow. The nucleus is stained with 4,6-diamidino-2-phe
as tested. The addition of HuR to the in vitro transla- i
ion reaction containing the luciferase–COX-2 3=UTRhimeric mRNA resulted in a dose-dependent increase inuciferase levels (Figure 5B). CUGBP2, on the other hand,ecreased luciferase mRNA translation in a dose-depen-ent manner (Figure 5C). This suggests that HuR en-ances, whereas CUGBP2 suppresses, COX-2 mRNAranslation. In contrast, there was no change in the ac-ivity by either CUGBP2 or HuR treatment when theontrol luciferase mRNA lacking the COX-2 3=UTR wassed in the in vitro translation reaction, implying spec-
ficity of the 2 RNA binding proteins to the COX-2=UTR-mediated effects. We next determined whetherhere is competition between HuR and CUGBP2 in theranslation of the luciferase–COX-2 3=UTR mRNA. Forhis, we performed 2 types of experiments. In the firstxperiment, the combined amount of CUGBP2 anduR was kept constant. When CUGBP2 levels were
vitro translated 35 S-labeled HuR was incubated with recombinantg glutathione sepharose beads. Total input protein (T), eluted sampleontrol (�), and eluted sample (P) and the unbound sample (S) from thehoresis and autoradiographed. The gel picture shows the presence of
t HuR homodimerizes, and also in the sample containing recombinanternatant samples did not show the presence of 35 S-HuR. Represen-were cloned in both the bait and test plasmids for expression as fusionespectively. As a positive control, p53 and SV40 T large antigen (RECT),activation domains, respectively. Presence of an interaction results in
o transcription of the MEL1 gene encoding �-galactosidase. Both HuRn the other hand, these proteins did not interact with the RecT or p53l). (C) Immunofluorescence analysis. Flag-tagged HuR and myc-taggedr myc tag) and Cy5 (green for FLAG tag). Overlapping red and greenole. The colocalization of CUGBP2 and HuR in the nucleus is shown.
ay. Intainintive cectropg tha
supNAsain, rg andwing tes. Oontrored fo
ncreased, while at the same time HuR levels were

deHicaiCtCm
tm
pCpcCp(fttmCv
FCtHiooctC(ie
BA
SIC–
ALI
MEN
TARY
TRA
CT
March 2007 CUGBP2 INHIBITS HUR ACTIVITY 1061
ecreased, there was a simultaneous decrease in lucif-rase levels (Figure 5D). In a second experiment, whenuR levels were kept constant and CUGBP2 levels were
ncreased gradually, 50% translation inhibition oc-urred when the reaction contained 0.3 pmol CUGBP2nd 0.75 pmol HuR (Figure 5D). Furthermore, a 75%nhibition of the luciferase activity was observed whenUGBP2 levels were increased to 0.6 pmol in the reac-
ion. These data, taken together, indicate thatUGBP2 overrides HuR and inhibits the COX-2RNA translation.To determine whether CUGBP2 competitively inhibits
ranslation in vivo, we performed transfection experi-
igure 5. CUGBP2 inhibits, whereas HuR enhances, in vitro COX-2 mOX-2 3=UTR. (B) The Luc/COX-2 3=UTR mRNA (black columns) and t
he presence of increasing concentrations of recombinant GST/HuR, auR there was a corresponding increase in the translation of the Luc
ncreasing concentrations of CUGBP2 as indicated, there was greater subserved with the control Luc mRNA (gray columns; *P � .01). (D) Luc/f increasing concentrations of CUGBP2 and a corresponding decreaonsidered as 100%. There was a consistent decrease in the translaranslation was observed in a protein mixture that contained 0.47 pmUGBP2 over HuR in regulating the luciferase/COX-2 mRNA translation
black columns) translation was performed in the presence of 0.75 pmonhibition (*P � .01) was observed beginning with 25 ng CUGBP2. Gxperiments.
ents in HT-29 cells. The cells were transfected with c
lasmids encoding either Flag-tagged HuR or HA-taggedUGBP2. In addition, the cells were transfected with alasmid encoding the firefly luciferase gene under theontrol of the cytomegalovirus (CMV) promoter and theOX-2 3=UTR. After transfection, cell lysates were pre-ared and subjected to immunoprecipitation with anti-HA
for CUGBP2) or anti-FLAG (for HuR) antibodies. RNArom the immunoprecipitates was isolated and subjectedo reverse-transcriptase– coupled polymerase chain reac-ion for COX-2 mRNA. Higher levels of the COX-2
RNA were found to be bound to HuR compared withUGBP2 in these basal conditions (Figure 6A). We pre-iously showed that endogenous CUGBP2 levels in these
translation. (A) Luciferase transcript containing the full-length murinentrol Luc mRNA (gray columns) were subjected to in vitro translation ine luciferase activity was measured. With increasing concentrations of-2 3=UTR mRNA, but not the control Luc mRNA (*P � .01). (C) Withssion of the Luc/COX-2 3=UTR mRNA (black columns). No effects were2 3=UTR mRNA (black columns) was in vitro translated in the presenceconcentration of HuR. Translation in the presence of HuR alone wasith the increasing concentration of CUGBP2. The 50% reduction inGBP2 and 0.89 pmol HuR, suggesting a competitive advantage for.01). Gray columns, control Luc mRNA. (E) Luc/COX-2 3=UTR mRNA
, and increasing concentrations of CUGBP2 (0–0.6 pmol). Significantcolumns, control Luc mRNA. These results are from 3 independent
RNAhe cond th
/COXppre
COX-singtion wol CU(*P �l HuRray
ells are low, but that they are induced after 12 Gy

�bCrt�bnw(
m3tseCwla
uHTpwelsHweelojlrTCh
4Fmee3ctCIwpwNMmctfiptsefwsctpnahCcic.
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1062 SUREBAN ET AL GASTROENTEROLOGY Vol. 132, No. 3
-irradiation.12 This increased expression led to CUGBP2inding to the ARE in the COX-2 3=UTR and inhibitingOX-2 mRNA translation. To determine whether the
adiation affected the RNA binding capacity of the ec-opically expressed proteins, we also administered 12 Gy-irradiation to the cells after transfection. In contrast toaseline, the majority of the COX-2 mRNA was immu-oprecipitated with anti-HA antibody (for CUGBP2)hen the cells also were exposed to the ionizing radiation
Figure 6A).Because CUGBP2 inhibits translation of COX-2RNA after binding to the ARE sequences in COX-2
=UTR, we next determined whether COX-2 mRNAranslation is affected in the cells. Western blot analysishowed increased COX-2 protein expression in the pres-nce of HuR (Figure 6B). However, in the presence ofUGBP2, the expression was decreased. Furthermore,hen the 2 proteins were co-expressed, COX-2 protein
evels were suppressed. This was even more pronouncedfter exposure of the cells to 12 Gy �-radiation (Fig-
�
re 6B), suggesting that CUGBP2 competitively inhibiteduR-mediated induction of COX-2 mRNA translation.o confirm these results, expression of the luciferaserotein from the luciferase-COX-2 3=UTR constructs alsoas determined. In the absence of radiation treatment,
xpression of HuR alone significantly induced luciferaseevels in the cells, whereas CUGBP2 inhibited the expres-ion (Figure 6C). Furthermore, when both FLAG-taggeduR and HA-tagged CUGBP2 were co-expressed, thereas neither an induction nor suppression of luciferase
xpression (Figure 6B). In contrast, in cells that also werexposed to 12 Gy �-irradiation, there was no induction ofuciferase levels even in the HuR-expressing cells. More-ver, cells transfected with HA-tagged CUGBP2 and sub-
ected to the radiation treatment showed suppression ofuciferase expression, to below baseline levels. Similaresults were obtained in HCT-116 cells (data not shown).hese data, taken together, suggest that after radiationUGBP2 competitively binds to COX-2 3=UTR and in-ibits translation.
™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™igure 6. CUGBP2 competes with HuR in regulating ARE-drivenRNA translation in vivo. (A) HT-29 cells were cotransfected with HA
pitope–tagged CUGBP2, FLAG epitope–tagged HuR, and firefly lucif-rase gene containing the full-length 2.2-kb COX-2 3=UTR (Luc/COX-2=UTR). Cells also were subjected to 12 Gy �-irradiation where indi-ated (lanes 6–9). Total cell lysates were prepared and immunoprecipi-ated with anti-HA and anti-FLAG antibodies to immunoprecipitateUGBP2 and HuR, respectively. As a negative control, a normal rabbit
gG (NRS) preparation was used. RNA bound to the immunoprecipitatesere isolated and subjected to semiquantitative reverse-transcriptionolymerase chain reaction for luciferase gene. The luciferase mRNAas found in anti-HA and anti-FLAG immunoprecipitates, but not withRS, showing in vivo binding of the 2 proteins to the COX-2 3=UTR.oreover, in cells that were not subjected to the radiation treatment,ore of the Luc/COX-2 3=UTR mRNA was bound to HuR, whereas in
ells also subjected to 12 Gy �-radiation (12 Gy), the majority of theranscript was bound to CUGBP2. (B) Total cell lysates from the trans-ections were subjected to Western blot analyses for COX-2. HuR alonencreased COX-2 protein levels (lane 3), but this was inhibited in theresence of cotransfected CUGBP2 (lane 4). Furthermore, after radia-ion, COX-2 protein levels did not increase even with HuR overexpres-ion (lane 7), and there was a further decrease in COX-2 levels in cellsxpressing both HuR and CUGBP2 (lane 8). Actin was used as controlor protein loading. (C) The total cell lysates from the transfections alsoere subjected to an in vitro luciferase activity assay. In the unirradiatedamples, cells expressing FLAG epitope–tagged HuR showed in-reased luciferase activity, whereas those that expressed HA epitope–agged CUGBP2 showed a decreased luciferase activity when com-ared with the controls. When cells expressed both proteins, there waso effect. After radiation there was an overall decrease in the luciferasectivity. In contrast, in cells subjected to 12 Gy �-irradiation, HuR did notave any effect on translation. Furthermore, expression of HA-taggedUGBP2 enhanced the translation inhibition effect (a, P � .01 whenomparing irradiated with unirradiated cells; b, P � .01 when compar-
ng FLAG-HuR–transfected with untransfected cells; c, P � .01 whenomparing HA-CUGBP2–transfected with untransfected cells; d, P �5 when comparing cells transfected with both with untransfected cells).
, No IR; s, 12 Gy.
CswsdtAmienbwcfCpcCmprptpcptpHtw
bcttRtrlsmtdi
bpiaibT
tdtttattorlhhttiost
ctmCstCcbHCtll
aathhalgspbps
eiiast
BA
SIC–
ALI
MEN
TARY
TRA
CT
March 2007 CUGBP2 INHIBITS HUR ACTIVITY 1063
DiscussionThe present study presents evidence that
UGBP2 is a functional antagonist of HuR. There is atriking structural similarity between the 2 proteins,ith each encoding 2 RRMs in the N-terminus and a
ingle RRM in the C-terminus, but the hinge region isifferent.14,19 Evidence of the similarity comes fromhe binding affinities of the 2 proteins to the COX-2RE sequences, with binding constants in the nano-olar range. Although HuR had a slightly higher bind-
ng constant, the difference was not significant. Nev-rtheless, for RNA binding proteins binding in theanomolar range is considered to be high-affinityinding. The first evidence of functional antagonismas seen in the results from the in vitro binding
ompetition assays. Indeed, the switch in ARE bindingrom HuR to CUGBP2 occurred at even lower levels ofUGBP2 when compared with the amount of HuRresent in the binding reaction. The second evidenceame from the in vitro translation studies in whichUGBP2 not only suppressed translation of the chi-eric luciferase–COX-2 3=UTR mRNA in a dose-de-
endent manner, but also abolished its stimulation inesponse to HuR. Further evidence that CUGBP2 com-etitively suppressed HuR-mediated COX-2 mRNAranslation was shown by the lower levels of COX-2rotein and that of the luciferase protein in HT-29ells when the 2 proteins were co-expressed. The sup-ression was especially pronounced after radiationreatment. This suggests the presence of additionalosttranslational modifications that occurred either inuR or CUGBP2, or both, after radiation treatment
hat resulted in enhanced CUGBP2 binding coupledith decreased HuR binding.This study showed homodimerization activity for
oth HuR and CUGBP2. The fact that the 2 proteinsolocalize in the nucleus could suggest potential func-ional significance in relation to their nuclear activi-ies. Although HuR is thought to bind ARE-containingNAs and transport them to the cytoplasm,29 the func-
ional significance of CUGBP2 in epithelial cells cur-ently is unknown. CUGBP2 has been shown to regu-ate alternative splicing of muscle- or neuron-specificplicing regulation events via CUG- or UG-rich ele-
ents, implying that CUGBP2 is a splicing regula-or.30,31 Although HuR has not been shown to have airect effect on alternative splicing, the possibility ex-
sts that HuR may do so through CUGBP2.Many multifunction RNA binding proteins have
een shown to shuttle between the nucleus and cyto-lasm.32 HuR plays an important role in ARE-contain-
ng cellular transcripts by regulating their stabilitynd/or nuclear export. More importantly, HuR bind-ng to the ARE protects the RNA from degradation,ut appears to have little effect on deadenylation.33
his is probably because HuR binds to ARE-containing a
ranscripts in the nucleus and protects them fromegradation during the transit to the cytoplasm.34 Ini-ial studies predicted the hinge region to have bidirec-ional transport signal, but other studies have shownhat this region encodes only a nuclear import signalnd that it interacts with transport factors of theransportin family.29 Additional studies have predictedhat RRM3 interacts with leucine zipper proteins pp32r APRIL, which in turn bind to the CRM1 exporteceptor.35 We previously showed that CUGBP2 trans-ocates to the cytoplasm after radiation exposure, andere we have shown that the protein shuttles in theeterokaryon assay. Recent studies have predicted thathe export signal lies in the hinge region.20 Moreover,he nuclear localization signal is believed to be locatedn the C-terminal end of the protein.20 This is the exactpposite of what has been proposed with HuR, whichuggests the presence of alternative mechanisms ofransport for the proteins.
CUGBP2 is a novel translational regulator thatounteracts HuR in binding to AU-rich sequences,hereby preventing COX-2 protein expression. This is
ost likely a mechanism for in vivo inhibition ofOX-2 expression, in conditions in which overexpres-
ion of COX-2 could be deleterious. One such condi-ion is in cancer, in which COX-2 is overexpressed.ytoplasmic localization of HuR also is increased in
ancer cells.36 Furthermore, increased tumorigenesis iselieved to occur in part due to increased binding ofuR to the COX-2 AREs, thereby stabilizing theOX-2 mRNA, and increasing COX-2 mRNA transla-
ion.14 Inhibiting COX-2 mRNA translation is thus aikely additional mechanism by which the cell regu-ates the amount of COX-2 protein.
The possibility exists that the relative levels of HuRnd CUGBP2 may determine the fate of transcripts suchs COX-2 that are critical for cell growth and/or protec-ion from apoptosis-inducing mechanisms. On the otherand, recent studies have shown that HuR also can in-ibit mRNA translation. In a recent study, Katsanou etl18 showed that HuR overexpression induced the trans-ational silencing of specific cytokine mRNAs by syner-izing with another translational silencer, TIA-1. Thisuggests that HuR function may be modulated by itsrotein partners. Although our studies show differentialinding of CUGBP2 and HuR after radiation, it is equallyossible that CUGBP2 and HuR function together toilence COX-2 mRNA.
Finally, not all RNA binding proteins have negativeffects on gene expression in response to DNA-damag-ng agents. hnRNP K, another RNA binding proteinmplicated in RNA splicing, export, and translationccumulates in the cells in response to UV expo-ure.37,38 Subsequently, hnRNP K cooperates with p53o elicit the activation of p53 target genes such as p21
nd thereby trigger cell-cycle– checkpoint events.38
Tpedat
1
1
1
1
1
1
1
1
1
1
2
2
2
2
2
2
2
2
2
2
3
3
3
3
3
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1064 SUREBAN ET AL GASTROENTEROLOGY Vol. 132, No. 3
aken together, these data suggest that RNA bindingroteins play a major role in regulating various cellularvents in response to DNA-damaging agents, and un-erstanding the mechanism(s) by which these proteinsre regulated would give us insights into their use asargets for anticancer therapies.
References
1. Cao Y, Prescott SM. Many actions of cyclooxygenase-2 in cellulardynamics and in cancer. J Cell Physiol 2002;190:279–286.
2. Yucel-Lindberg T, Ahola H, Carlstedt-Duke J, Modeer T. Involve-ment of tyrosine kinases on cyclooxygenase expression and pros-taglandin E2 production in human gingival fibroblasts stimulatedwith interleukin-1beta and epidermal growth factor. Biochem Bio-phys Res Commun 1999;257:528–532.
3. Khan KN, Masferrer JL, Woerner BM, Soslow R, Koki AT. En-hanced cyclooxygenase-2 expression in sporadic and familialadenomatous polyposis of the human colon. Scand J Gastroen-terol 2001;36:865–869.
4. Hla T, Bishop-Bailey D, Liu CH, Schaefers HJ, Trifan OC. Cycloox-ygenase-1 and -2 isoenzymes. Int J Biochem Cell Biol 1999;31:551–557.
5. Dixon DA. Dysregulated post-transcriptional control of COX-2gene expression in cancer. Curr Pharm Des 2004;10:635–646.
6. Sully G, Dean JL, Wait R, Rawlinson L, Santalucia T, Saklatvala J,Clark AR. Structural and functional dissection of a conserveddestabilizing element of cyclo-oxygenase-2 mRNA: evidenceagainst the involvement of AUF-1 [AU-rich element/poly(U)-bind-ing/degradation factor-1], AUF-2, tristetraprolin, HuR (Hu antigenR) or FBP1 (far-upstream-sequence-element-binding protein 1).Biochem J 2004;377:629–639.
7. Shao J, Sheng H, Inoue H, Morrow JD, DuBois RN. Regulation ofconstitutive cyclooxygenase-2 expression in colon carcinomacells. J Biol Chem 2000;275:33951–33956.
8. Sengupta S, Jang BC, Wu MT, Paik JH, Furneaux H, Hla T. TheRNA-binding protein HuR regulates the expression of cyclooxygen-ase-2. J Biol Chem 2003;278:25227–25233.
9. Cok SJ, Morrison AR. The 3=-untranslated region of murine cyclo-oxygenase-2 contains multiple regulatory elements that altermessage stability and translational efficiency. J Biol Chem 2001;276:23179–23185.
0. Cok SJ, Acton SJ, Sexton AE, Morrison AR. Identification ofRNA-binding proteins in RAW 264.7 cells that recognize a lipopo-lysaccharide-responsive element in the 3-untranslated region ofthe murine cyclooxygenase-2 mRNA. J Biol Chem 2004;279:8196–8205.
1. Cok SJ, Acton SJ, Morrison AR. The proximal region of the 3=-untranslated region of cyclooxygenase-2 is recognized by a mul-timeric protein complex containing HuR, TIA-1, TIAR, and theheterogeneous nuclear ribonucleoprotein U. J Biol Chem 2003;278:36157–36162.
2. Mukhopadhyay D, Houchen CW, Kennedy S, Dieckgraefe BK,Anant S. Coupled mRNA stabilization and translational silencingof cyclooxygenase-2 by a novel RNA binding protein, CUGBP2.Mol Cell 2003;11:113–126.
3. Murmu N, Jung J, Mukhopadhyay D, Houchen CW, Riehl TE,Stenson WF, Morrison AR, Arumugam T, Dieckgraefe BK, AnantS. Dynamic antagonism between RNA-binding protein CUGBP2and cyclooxygenase-2-mediated prostaglandin E2 in radiationdamage. Proc Natl Acad Sci U S A 2004;101:13873–13878.
4. Dixon DA, Tolley ND, King PH, Nabors LB, McIntyre TM, Zimmer-man GA, Prescott SM. Altered expression of the mRNA stabilityfactor HuR promotes cyclooxygenase-2 expression in colon can-
cer cells. J Clin Invest 2001;108:1657–1665.5. King PH, Redden D, Palmgren JS, Nabors LB, Lennon VA. Huantigen specificities of ANNA-I autoantibodies in paraneoplasticneurological disease. J Autoimmun 1999;13:435–443.
6. Brennan CM, Steitz JA. HuR and mRNA stability. Cell Mol Life Sci2001;58:266–277.
7. Peng SY, Chou SP, Hsu HC. Association of downregulation ofcyclin D1 and of overexpression of cyclin E with p53 mutation,high tumor grade and poor prognosis in hepatocellular carci-noma. J Hepatol 1998;29:281–289.
8. Katsanou V, Papadaki O, Milatos S, Blackshear PJ, Anderson P,Kollias G, Kontoyiannis DL. HuR as a negative posttranscriptionalmodulator in inflammation. Mol Cell 2005;19:777–789.
9. Chen CY, Xu N, Shyu AB. Highly selective actions of HuR inantagonizing AU-rich element-mediated mRNA destabilization.Mol Cell Biol 2002;22:7268–7278.
0. Ladd AN, Cooper TA. Multiple domains control the subcellular local-ization and activity of ETR-3, a regulator of nuclear and cytoplasmicRNA processing events. J Cell Sci 2004;117:3519–3529.
1. Anant S, MacGinnitie AJ, Davidson NO. Apobec-1, the catalyticsubunit of the mammalian apolipoprotein B mRNA editing en-zyme, is a novel RNA-binding protein. J Biol Chem 1995;270:14762–14767.
2. Carey J, Lowary PT, Uhlenbeck OC. Interaction of R17 coat pro-tein with synthetic variants of its ribonucleic acid binding site.Biochemistry 1983;22:4723–4730.
3. Anant S, Henderson JO, Mukhopadhyay D, Navaratnam N,Kennedy S, Min J, Davidson NO. Novel role for RNA-bindingprotein CUGBP2 in mammalian RNA editing. CUGBP2 modulatesC to U editing of apolipoprotein B mRNA by interacting withapobec-1 and ACF, the apobec-1 complementation factor. J BiolChem 2001;276:47338–47351.
4. Akakura S, Yoshida M, Yoneda Y, Horinouchi S. A role for Hsc70in regulating nucleocytoplasmic transport of a temperature-sen-sitive p53 (p53Val-135). J Biol Chem 2001;276:14649–14657.
5. Blanc V, Navaratnam N, Henderson JO, Anant S, Kennedy S,Jarmuz A, Scott J, Davidson NO. Identification of GRY-RBP as anapolipoprotein B RNA-binding protein that interacts with bothapobec-1 and apobec-1 complementation factor to modulate C toU editing. J Biol Chem 2001;276:10272–10283.
6. Zhao D, Keates AC, Kuhnt-Moore S, Moyer MP, Kelly CP, Pothou-lakis C. Signal transduction pathways mediating neurotensin-stimulated interleukin-8 expression in human colonocytes. J BiolChem 2001;276:44464–44471.
7. Niranjanakumari S, Lasda E, Brazas R, Garcia-Blanco MA. Revers-ible cross-linking combined with immunoprecipitation to studyRNA-protein interactions in vivo. Methods 2002;26:182–190.
8. Singh G, Charlet BN, Han J, Cooper TA. ETR-3 and CELF4 proteindomains required for RNA binding and splicing activity in vivo.Nucleic Acids Res 2004;32:1232–1241.
9. Fan XC, Steitz JA. HNS, a nuclear-cytoplasmic shuttling sequencein HuR. Proc Natl Acad Sci U S A 1998;95:15293–15298.
0. Ladd AN, Charlet N, Cooper TA. The CELF family of RNA bindingproteins is implicated in cell-specific and developmentally regu-lated alternative splicing. Mol Cell Biol 2001;21:1285–1296.
1. Gromak N, Matlin AJ, Cooper TA, Smith CW. Antagonistic regula-tion of alpha-actinin alternative splicing by CELF proteins andpolypyrimidine tract binding protein. RNA 2003;9:443–456.
2. Wilkinson MF, Shyu AB. Multifunctional regulatory proteins thatcontrol gene expression in both the nucleus and the cytoplasm.Bioessays 2001;23:775–787.
3. Gantt K, Cherry J, Tenney R, Karschner V, Pekala PH. An early eventin adipogenesis, the nuclear selection of the CCAAT enhancer-binding protein {beta} (C/EBP{beta}) mRNA by HuR and its translo-cation to the cytosol. J Biol Chem 2005;280:24768–24774.
4. Wang JG, Collinge M, Ramgolam V, Ayalon O, Fan XC, Pardi R,Bender JR. LFA-1-dependent HuR nuclear export and cytokine mRNA
stabilization in T cell activation. J Immunol 2006;176:2105–2113.
3
3
3
3
ote
Co
c
March 2007 CUGBP2 INHIBITS HUR ACTIVITY 1065
5. Gallouzi IE, Brennan CM, Steitz JA. Protein ligands mediate the CRM1-dependent export of HuR in response to heat shock. RNA 2003;9:1410.
6. Lopez de Silanes I, Fan J, Yang X, Zonderman AB, Potapova O,Pizer ES, Gorospe M. Role of the RNA-binding protein HuR incolon carcinogenesis. Oncogene 2003;22:7146–7154.
7. Bomsztyk K, Denisenko O, Ostrowski J. hnRNP K: one proteinmultiple processes. Bioessays 2004;26:629–638.
8. Moumen A, Materson P, O’Conner MJ, Jackson SP. hnRNP K: anHDM2 target and transcriptional coactivator of p53 in response
to DNA damage. Cell 2005;123:1065–1078. aReceived August 16, 2006. Accepted November 16, 2006.Address requests for reprints to: Shrikant Anant, PhD, Department
f Medicine, University of Oklahoma Health Science Center, 920 Stan-on L. Young Boulevard, WP 1360, Oklahoma City, Oklahoma 73104.-mail: [email protected] by National Institutes of Health grants DK-62265 and
A109269 (S.A.), DK-060106 (B.K.D.), and an American Gastroenter-logy Association Research Scholars Award (S.A.).The authors thank Aubrey Morrison for the pGL3-COX-2 3=UTR luciferase
onstruct, and Dr Joan Steitz for the pCDNA3-HuR plasmid construct. The
uthors also thank Dr. William F. Stenson for helpful discussions.BA
SIC–
ALI
MEN
TARY
TRA
CT
Copyright © 2022 FDOKUMEN




















