
Functional and kinetic characterization of granulocyte colony-stimulating factor-primed CD34 −...
10
Functional and kinetic characterization of granulocyte colony- stimulating factor-primed CD34 ) human stem cells Roberto M. Lemoli, 1 Francesco Bertolini, 2 Maria T. Petrucci, 3 Chiara Gregorj, 3 Maria R. Ricciardi, 3 Miram Fogli, 1 Antonio Curti, 1 Cristina Rabascio, 2 Simona Pandolfi, 1 Sergio Ferrari, 4 Robin Fo, 3 Michele Baccarani 1 and Agostino Tafuri 3 1 Istituto di Ematologia e Oncologia Medica ‘L. & A. Sera `gnoli’, University of Bologna, Bologna, 2 Divisione di Ematologia e Oncologia, Istituto Europeo di Oncologia, Milan, 3 Dipartimento di Biotecnologie Cellulari ed Ematologia, Sezione di Ematologia, University ‘La Sapienza’ of Rome, Rome, and 4 Dipartimento di Scienze Biomediche, University of Modena, Modena, Italy Received 27 June 2003; accepted for publication 14 August 2003 Summary. We assessed the functional properties and the kinetic status in vitro, and the engraftment potential in vivo of human haematopoietic stem cells according to the expression of CD34 antigen. Lin ) CD34 ) and Lin ) CD34 + cells were isolated from granulocyte colony-stimulating factor-primed peripheral blood (PB) cells of healthy donors. The CD34 ) cell fraction did not contain either clonogenic cells in semisolid culture or long-term culture initiating cells (LTC-IC). However, stroma-dependent liquid cultures and cytokines induced CD34 expression on a minority of stem cells, acquisition of clonogenic capacity and generation of LTC-IC. Significantly higher percentages of quiescent G 0 cells and lower percentages of cycling G 1 cells were found in Lin ) CD34 ) cells when compared with Lin ) CD34 + cells. Kinetic quiescence of Lin ) CD34 ) cells was associated with a significantly higher expression of the negative regulators of the cell cycle, p27 Kip1 and p21 cip1/waf1 . Cytokine-mediated induction of CD34, in vitro, resulted in cycling of stem cells and downregulation of p27. There was a higher rate of human long-term engraftment in immunocompromised non-obese diabetic (NOD)/recombination activating gene 1 null and NOD/severe combined immunodeficient-b 2 micro- globulin null mice injected with CD34 + cells. Thus, our study indicated that CD34 expression on human PB stem cells was associated with haematopoietic activity, cell-cycle recruit- ment and downregulation of p27 Kip1 in vitro and higher engraftment capacity in vivo. Keywords: haematopoietic stem cells, cell cycle, CD34. The CD34 antigen (Ag) is a phosphoglycoprotein that is expressed on the cell membrane of human haematopoietic progenitor cells in bone marrow (BM), peripheral blood (PB) and cord blood (CB) (reviewed in Krause et al, 1996). Animal models (Berenson et al, 1988) and clinical trans- plantation studies using highly enriched cell populations have also demonstrated the capacity of CD34 + cells to behave as true stem cells, as they engraft in lethally irradiated allogeneic hosts (Bensinger et al, 1996). How- ever, recent studies have challenged the dogma that all pluripotent stem cell express the CD34 Ag. In particular, the generation of CD34 knockout mice demonstrated that the presence of the CD34 molecule is not functionally necessary to develop multilineage haematopoiesis (Cheng et al, 1996). Furthermore, transplantation of single Lin ) CD34 ) cells induced the sustained long-term engraftment of lethally irradiated mice (Osawa et al, 1996). Following these initial observations, several studies have addressed the issue of the stem cell capacity of CD34 ) cells in different animal systems and in humans using xenograft repopulation assays (Goo- dell et al, 1997; Bhatia et al, 1998; Zanjani et al, 1998; Fujisaki et al, 1999; Sato et al, 1999; Andrews et al, 2000; Tajima et al, 2000; Verfaillie et al, 2000; Gao et al, 2001). However, experiments using human cells gave widely varying results. Whereas Zanjani et al (1998) and Bhatia et al (1998) reported significant numbers of long-term xenogenic repopulating cells within the CD34 ) cell popu- lation, Gao et al (2001) and Fujisaki et al (1998) found that CD34 ) cells make hardly any contribution to human haematopoietic engraftment in non-obese diabetic/severe combined immunodeficient (NOD/SCID) mice. The relationship between CD34 + and CD34 ) stem cells has been studied in vitro (Nakamura et al, 1999) and in vivo Correspondence: Roberto M. Lemoli MD, Istituto di Ematologia e Oncologia Medica ‘L. & A. Sera `gnoli’, Via Massarenti, 9, 40100 Bologna, Italy. E-mail: [email protected] British Journal of Haematology, 2003, 123, 720–729 720 Ó 2003 Blackwell Publishing Ltd
Transcript of Functional and kinetic characterization of granulocyte colony-stimulating factor-primed CD34 −...

Functional and kinetic characterization of granulocyte colony-
stimulating factor-primed CD34) human stem cells
Roberto M. Lemoli,1
Francesco Bertolini,2
Maria T. Petrucci,3
Chiara Gregorj,3
Maria R. Ricciardi,3
Miram Fogli,1
Antonio Curti,1
Cristina Rabascio,2
Simona Pandolfi,1
Sergio Ferrari,4
Robin Fo,3
Michele Baccarani1
and Agostino Tafuri3 1Istituto di Ematologia e
Oncologia Medica ‘L. & A. Seragnoli’, University of Bologna, Bologna, 2Divisione di Ematologia e Oncologia, Istituto
Europeo di Oncologia, Milan, 3Dipartimento di Biotecnologie Cellulari ed Ematologia, Sezione di Ematologia, University
‘La Sapienza’ of Rome, Rome, and 4Dipartimento di Scienze Biomediche, University of Modena, Modena, Italy
Received 27 June 2003; accepted for publication 14 August 2003
Summary. We assessed the functional properties and thekinetic status in vitro, and the engraftment potential in vivoof human haematopoietic stem cells according to theexpression of CD34 antigen. Lin)CD34) and Lin)CD34+
cells were isolated from granulocyte colony-stimulatingfactor-primed peripheral blood (PB) cells of healthy donors.The CD34) cell fraction did not contain either clonogeniccells in semisolid culture or long-term culture initiating cells(LTC-IC). However, stroma-dependent liquid cultures andcytokines induced CD34 expression on a minority of stemcells, acquisition of clonogenic capacity and generation ofLTC-IC. Significantly higher percentages of quiescent G0
cells and lower percentages of cycling G1 cells were found inLin)CD34) cells when compared with Lin)CD34+ cells.Kinetic quiescence of Lin)CD34) cells was associated with a
significantly higher expression of the negative regulators ofthe cell cycle, p27Kip1 and p21cip1/waf1. Cytokine-mediatedinduction of CD34, in vitro, resulted in cycling of stem cellsand downregulation of p27. There was a higher rate ofhuman long-term engraftment in immunocompromisednon-obese diabetic (NOD)/recombination activating gene1null and NOD/severe combined immunodeficient-b2micro-globulinnull mice injected with CD34+ cells. Thus, our studyindicated that CD34 expression on human PB stem cells wasassociated with haematopoietic activity, cell-cycle recruit-ment and downregulation of p27Kip1 in vitro and higherengraftment capacity in vivo.
Keywords: haematopoietic stem cells, cell cycle, CD34.
The CD34 antigen (Ag) is a phosphoglycoprotein that isexpressed on the cell membrane of human haematopoieticprogenitor cells in bone marrow (BM), peripheral blood (PB)and cord blood (CB) (reviewed in Krause et al, 1996).Animal models (Berenson et al, 1988) and clinical trans-plantation studies using highly enriched cell populationshave also demonstrated the capacity of CD34+ cells tobehave as true stem cells, as they engraft in lethallyirradiated allogeneic hosts (Bensinger et al, 1996). How-ever, recent studies have challenged the dogma that allpluripotent stem cell express the CD34 Ag. In particular, thegeneration of CD34 knockout mice demonstrated that thepresence of the CD34 molecule is not functionally necessaryto develop multilineage haematopoiesis (Cheng et al, 1996).
Furthermore, transplantation of single Lin)CD34) cellsinduced the sustained long-term engraftment of lethallyirradiated mice (Osawa et al, 1996). Following these initialobservations, several studies have addressed the issue of thestem cell capacity of CD34) cells in different animal systemsand in humans using xenograft repopulation assays (Goo-dell et al, 1997; Bhatia et al, 1998; Zanjani et al, 1998;Fujisaki et al, 1999; Sato et al, 1999; Andrews et al, 2000;Tajima et al, 2000; Verfaillie et al, 2000; Gao et al, 2001).However, experiments using human cells gave widelyvarying results. Whereas Zanjani et al (1998) and Bhatiaet al (1998) reported significant numbers of long-termxenogenic repopulating cells within the CD34) cell popu-lation, Gao et al (2001) and Fujisaki et al (1998) found thatCD34) cells make hardly any contribution to humanhaematopoietic engraftment in non-obese diabetic/severecombined immunodeficient (NOD/SCID) mice.
The relationship between CD34+ and CD34) stem cellshas been studied in vitro (Nakamura et al, 1999) and in vivo
Correspondence: Roberto M. Lemoli MD, Istituto di Ematologia e
Oncologia Medica ‘L. & A. Seragnoli’, Via Massarenti, 9, 40100
Bologna, Italy. E-mail: [email protected]
British Journal of Haematology, 2003, 123, 720–729
720 � 2003 Blackwell Publishing Ltd

(Sato et al, 1999; Tajima et al, 2000). Taken together, thesestudies indicated that CD34) cells can be forced to expressCD34 either following cytokine stimulation in vitro or in vivoor following exposure to cytotoxic drugs. Thus, Sato et al(1999) proposed that only one class of murine stem cellsexists, albeit in two functional states distinguishable byCD34 expression. According to this model, CD34) stem cellsare extremely quiescent and can be activated by differentstimuli to generate a CD34+ cell population with highengraftment potential. Murine CD34+ cells can then revertto a CD34) phenotype after transplantation and retainsecondary long-term repopulating activity. In humans, thereversibility of CD34 expression on BM haematopoietic stemcells has been recently demonstrated by xenogenic trans-plant studies where CD34) stem cells were generated fromCD34+ progenitors and these CD34) cells regained CD34expression upon secondary transplantation (Dao et al,2003; Zanjani et al, 2003). However, it is still unclearwhether different subsets of human stem cells mobilized intoPB have different functional properties, and also whetherthe modulation of stem cell phenotype depends on cellactivation or induction of cell cycling.
In this study we addressed the issue of whether changesin CD34 expression correlate with the kinetic state of stemcells. Moreover, we assessed the functional characteristics,in vitro, and the engraftment potential of granulocytecolony-stimulating factor (G-CSF)-primed humanLin)CD34+ and CD34) cells transplanted in immunocom-promised NOD/recombination activating gene (RAG) 1null
and NOD/SCID-b2Microgobulin (b2M)null mice. Our resultsshowed that expression of CD34 on human stem cells wasassociated in vitro with haematopoietic activity, cell cyclerecruitment and downregulation of the cyclin-dependentkinase inhibitors (CKI) p27Kip1 (p27) and p21cip1/waf1 (p21),which are negative cell-cycle regulators maximallyexpressed during quiescence (Hengst & Reed, 1996; Chenget al, 2000), and higher engraftment capacity in vivo.
MATERIALS AND METHODS
Cells. Normal adult BM cells were obtained from fourallogeneic BM transplant donors. BM specimens wereobtained in steady state conditions, prior to G-CSF mobiliza-tion, from three of the normal volunteers and in one caseduring G-CSF treatment on the same day as the PBSCharvest. To minimize PB cell contamination, no more than2–3 ml of BM were collected from each aspiration. To obtainPB progenitor cells, six healthy donors received glycosylatedrecombinant human (rh) G-CSF (Lenograstim, Rhone-Pou-lenc Rorer, Milan, Italy) administered subcutaneously at10 lg/kg/d for 5–6 d. Leukaphereses were performed ondays 5 and 6 as previously described (Lemoli et al, 1997;Lemoli et al, 2000). Haematopoietic stem cell purificationand subsequent studies were always performed using day-5collections at the peak time of PB CD34+ cells. The protocolwas approved by the ethical committee of the UniversityHospital and each donor gave written informed consent.
Mononuclear light density (<1Æ077 g/ml) cells (MNC)were enriched using Ficoll-Paque (Pharmacia, Uppsala,
Sweden) and resuspended in 1% bovine serum albumin(BSA; Sigma Chemical Co, St Louis, MO, USA).
Stem cell selection and phenotypic analysis. The MNC fromBM or PB of healthy donors were processed by MiniMacshigh-gradient magnetic separation column (Miltenyi Biotec,Bergisch Gladbach, Germany) to obtain highly purifiedCD34+ cells as already reported (Lemoli et al, 1997). Thepercentages of CD34+ cells in BM and in PB samples were1Æ6 ± 1%, and 0Æ9 ± 0Æ3% of the MNC fraction respectively.After magnetic separation, the percentages of CD34+ in BMand PB samples were 92 ± 4% and 95 ± 5% respectively.Propidium iodide (2 lg/ml) was added for the detection ofnon-viable cells, which were excluded from the analyses. Inseparate experiments, MNC were stained for 30 min at 4�Cwith a mixture of lineage-specific antibodies (CD2, CD3,CD14, CD16, CD19, CD24, CD56, CD66b, CD41 andGlycophorin A) provided by the manufacturer (Stem CellTechnologies, Vancouver, Canada), followed by addition ofsecondary antibody conjugated to magnetic dextran-ironparticles for a further 30 min at 4�C. Cells were then elutedthrough a magnetized column (StemSep; Stem Cell Tech-nologies) to remove Lin+ cells. Lin) cells were then collectedaccording to the manufacturers instructions, incubatedwith anti-human CD34-fluorescein isothiocyanate (FITC)monoclonal antibody (MoAb) (HPCA-2; Becton Dickinson,San Jose, CA, USA) and sorted on a FACStar Plus equipment(Becton Dickinson). The sorting gates were set on isotypecontrol antibody-stained samples. Sort windows wereadjusted to minimize the chance of weakly positive cellscontaminating the CD34) cell population. Aliquots of sortedLin)CD34+ and Lin)CD34) cells, were reanalysed byFacScan to assess their purities, which were 98Æ5 ± 0Æ7%and 99Æ8 ± 0Æ1% respectively. A representative example ofsorting gates and flow cytometric reanalysis of sorted cells isshown in Fig 1. In some experiments, contamination ofCD34+ cells in the CD34) cell population was ruled out byreverse transcription-polymerase chain reaction (RT-PCR)for CD34 mRNA (data not shown).
Freshly separated Lin)CD34+ and Lin)CD34) cells werealso analysed by flow cytometry for the expression ofimportant molecules affecting homing to BM and engraft-ment. Briefly, 1 · 106 haematopoietic cells were incubatedwith 10 ll of anti-human CXCR4-phycoerythrin (PE) (R &D Systems Europe LDS, Abingdon, UK), anti-human CD49e-PE (VLA-5; Cymbus Biotechnology LTD, Chandlers Ford,Hampshire, UK) and anti-human CD49d-FITC (VLA-4;Cymbus Biotechnology) MoAbs at room temperature for20 min. After two washes, cells were analysed by FacScan.
Progenitor assays. Human colony-forming cells (CFU)were cultured in methylcellulose as previously described(Lemoli et al, 1997). Briefly, 2–5 · 103 CD34+ or 1 · 104
CD34) cells were plated in duplicate in culture mediumconsisting of 1 ml of Iscove’s modified Dulbecco’s medium(IMDM) supplemented with 24% fetal calf serum (FCS) (SeraLab, Crawley Down, Sussex, UK), 10)4 mol/l 2-mercapto-ethanol (Sigma), 0Æ2 mmol/l bovin haemin (Sigma), 0Æ8%BSA (Sigma), 2 U/ml of rh erythropoietin (Dompe Biotec,Milan, Italy), 50 ng/ml of stem cell factor (SCF) (Amgen,Thousand Oaks, CA, USA), 10 ng/ml of granulocyte-
Kinetic Characterization of Human Stem Cells 721
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 123: 720–729
macrophage CSF (GM-CSF; Immunex, Seattle, WA, USA)and 10 ng/ml of interleukin-3 (IL-3; Immunex). The finalconcentration of methylcellulose was 1Æ32%. CFU-granulo-cyte-macrophage (CFU-GM), burst-forming units erythroid(BFU-E) and multilineage colonies (CFU-Mix) (togetherreferred to as CFU-C), were scored after 14 d of incubationat 37�C in a fully humidified 5% CO2 atmosphere.
For long-term culture initiating cell (LTC-IC) assays, cellswere plated onto irradiated murine stromal cells (M2-10B4)genetically engineered to produce G-CSF and IL-3 with aweekly half-medium change. After 5 weeks in culture, thecells were then evaluated for their secondary CFU-C activity,and the number of LTC-IC was calculated as earlier reported(Lemoli et al, 1997).
Long-term cultures. Haematopoietic Lin)CD34+ andLin)CD34) stem cells were cultured onto irradiatedM2-10B4 cells (see above) at an initial density of4–10 · 104 cells/ml in IMDM supplemented with 10%FCS (Sera Lab), l-glutamine and antibiotics. All cultureswere maintained at 37�C in humidified 5% CO2 atmospherefor 3 weeks in presence of optimized concentrations of thefollowing rh cytokines: SCF (50 ng/ml; Amgen), IL-11(50 U/ml; Endogen, Woburn, MA, USA), fms-like tyrosinekinase-3 (FLT3)-ligand (50 ng/ml; Immunex) and megak-aryocyte growth and development factor (MGDF)(100 ng/ml, Amgen). Twice a week, the culture mediumwas replaced by fresh medium and cytokines and haema-topoietic cells were adjusted to the initial cell concentration.
Aliquots of cell suspension were cultured at weekly intervalsin methylcellulose and in liquid culture to evaluate thepresence of secondary CFU-C and of LTC-IC respectively (seeabove).
Flow cytometric evaluation of p27 and p21 expression. Aspreviously reported (Ricciardi et al, 2001; Curti et al, 2002),5–10 · 105 haematopoietic cells/ml were fixed and perme-abilized using the Fix & Perm Cell Permeabilization kit(Caltag, Burlingame, CA, USA) and then incubated for30 min in the dark, at room temperature, with the mousemonoclonal antibodies anti-human p27 (clone 57) (SantaCruz Biotechnology, Santa Cruz, CA, USA) and anti-humanp21 (Oncogene Research Products, San Diego, CA, USA)both used at the concentration of 1 lg/ml. Irrelevant mouseantibody of the appropriate subtype was used as negativecontrol to determine background fluorescence. After twowashes, the cells were incubated with FITC-conjugated goatF(ab’)2 fragment mouse IgG (H-L) for 30 min in the dark atroom temperature. Cells were then ready for flow cytometricanalysis. The human T-lymphoblastoid CEM cell line andnormal PB lymphocytes were used as positive control forp27 evaluation. Results were indicated as the meanfluorescence index (MFI) obtained by the ratio of themedian fluorescence value of the anti-human p27 dividedthat obtained by the control antibody. Expression of p21was analysed using the Kolmogorov–Smirnov (KS) statistictest (D value), which enables the objective and accurateidentification of small differences in fluorescence intensity
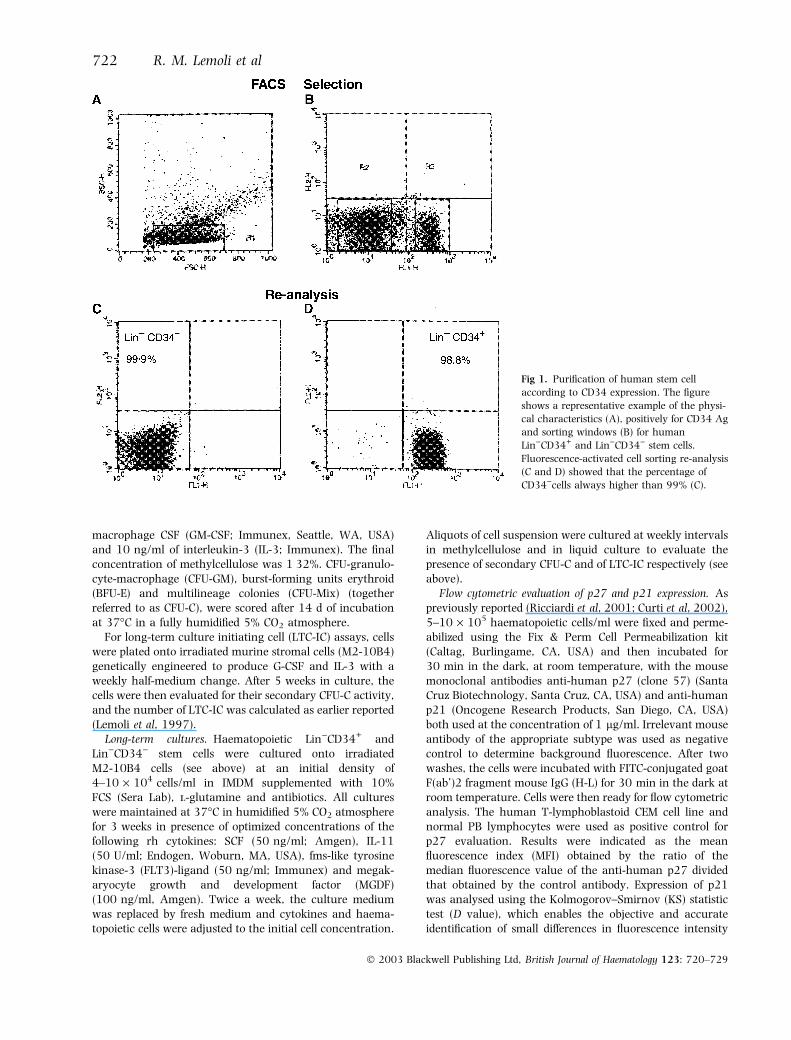
Fig 1. Purification of human stem cell
according to CD34 expression. The figure
shows a representative example of the physi-
cal characteristics (A), positively for CD34 Ag
and sorting windows (B) for human
Lin)CD34+ and Lin)CD34) stem cells.
Fluorescence-activated cell sorting re-analysis
(C and D) showed that the percentage of
CD34)cells always higher than 99% (C).
722 R. M. Lemoli et al
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 123: 720–729

(Tafuri et al, 2002). U937 cell line (KS ¼ 0Æ48) and normalPB lymphocytes (PBL) (KS ¼ 0) were used as positive andnegative controls respectively, for the expression of p21.
Flow cytometric evaluation of cell cycle distribution. Cell-cycle distribution was evaluated using the Acridine Orange(AO) technique. As previously described (Lemoli et al, 1997;Tafuri et al, 1999) the percentage of cells in G0, G1, S, G2M,and the mean RNA content of G1 cells were determined bysimultaneously measuring the DNA and RNA total cellularcontent. RNA content of G1 cells was expressed as RNA-Index (RNA-I G1), determined as the ratio of the mean RNAcontent of G1 cells of the samples multiplied by 10, dividedby the median RNA content of control lymphocytes. G0 cellswere defined as cells with an RNA content equal or lowerthan that of normal PBL.
Flow cytometry. Flow cytometry analysis was performedusing a FACScan flow cytometer (Becton Dickinson),operating at 488 nm. Five to ten thousand cells weremeasured for each analysis at separate wavelength bands:F530-DNA (AO) and –FITC (anti-p27MoAb) and F>620-RNA(AO). Samples were analysed using a Hewlett Packardmicrocomputer and Becton Dickinson software includingCellfit, Lysis II, CellQuest and ModFit.
Animal models. Six 8-week-old NOD-Rag1null (abbreviatedas NOD/RAG) (Shultz et al, 2000) or b2 microglobulinknockout NOD/-prkdcscid (abbreviated as NOD/SCID/B2M)(Kollet et al, 2000) mice were sublethally irradiated with500 or 200 cGy respectively, and injected intravenouslywith 5000–300 000 highly purified Lin)CD34+ orLin)CD34) cells. Sixty to 90 d after transplant, mice werekilled and their blood and marrow collected for theevaluation of human haematopoietic engraftment. Allprocedures involving animals were performed in accordancewith national and international laws and policies.
Measurement of human haematopoietic engraftment by flowcytometry and RT-PCR. Blood and marrow of NOD/RAG andNOD/SCID/B2M mice were evaluated by four-colour flowcytometry using a panel of MoAbs reacting with humanCD19, CD34 and CD45. After red cell lysis, cell suspensionswere evaluated by a FACSCalibur (Becton Dickinson) usinganalysis gates designed to exclude dead cells, platelets anddebris. After acquisition of at least 100 000 cells persample, analyses were considered as informative whenadequate numbers of events (i.e. >50; typically 100–200)were collected in the human cell enumeration gates.Percentages of stained cells were determined and comparedwith the appropriate negative controls. Positive stainingwas defined as being greater than non-specific backgroundstaining. Annexin V and 7-amino-actinomycin D (7AAD)were used to exclude apoptotic and dead cells (Dell’Agnolaet al, 2002).
The expression of human CD45 in blood and marrow oftransplanted mice was confirmed by RT-PCR and Southernblotting (Dell’Agnola et al, 2002). The sensitivity (onepositive cell/105 background cells) and specificity of humanCD45 RT-PCR was evaluated by serial dilutions of CD45positive NB4 cells in CD45 negative Hela cells andconfirmed by Southern blotting with probes that wereobtained by 32P labelled reverse primers.
Statistical analysis. The results are expressed as themean ± standard deviation (SD) of at least three differentexperiments. For transplant experiments, statistical com-parisons were performed using the t-test, analysis ofvariance (anova) and linear regression when data werenormally distributed and the non-parametric analyses ofSpearman and Mann–Whitney when data were skewed.Results of in vitro studies were analysed with the paired non-parametric Wilcoxon rank sum test. P-values lower than0Æ05 were considered as statistically significant.
RESULTS
Frequency and clonogenic growth of Lin)CD34+ and CD34)
cellsLin)CD34) stem cells were purified from the BM (n ¼ 4) orfrom G-CSF-primed PB (n ¼ 6) of healthy donors. Thefrequencies of PB Lin) cells and Lin)CD34) were1Æ09 ± 0Æ32% and 0Æ11 ± 0Æ04% of MNC respectively. Insteady-state BM, the frequencies of Lin) and Lin)CD34)
were 1Æ3 ± 0Æ6% and 0Æ09 ± 0Æ03% of MNC respectively.The slight differences between the two sources of stem cellswere not statistically significant. G-CSF-mobilizedLin)CD34+ and Lin)CD34) cells were plated in semisolidassay to determine the frequency of CFU-C, and in liquidculture onto M2-10B4 feeder layers to assess the content ofLTC-IC. In five of six cases of Lin)CD34), we did not observeany growth of either early or clonogenic progenitor cells:the frequency of both CFU-C and LTC-IC was <0Æ01% in onecase. Conversely, the mean clonogenic efficiency of PBLin)CD34+ cells was 1Æ94 ± 0Æ54% and the mean numberof LTC-IC was 49Æ5 ± 12 · 104 cells. Similar results wereobserved when BM-derived progenitor cells were assayed insemisolid and liquid culture (data not shown).
Production of CFU-C and LTC-IC from Lin)CD34+ and CD34)
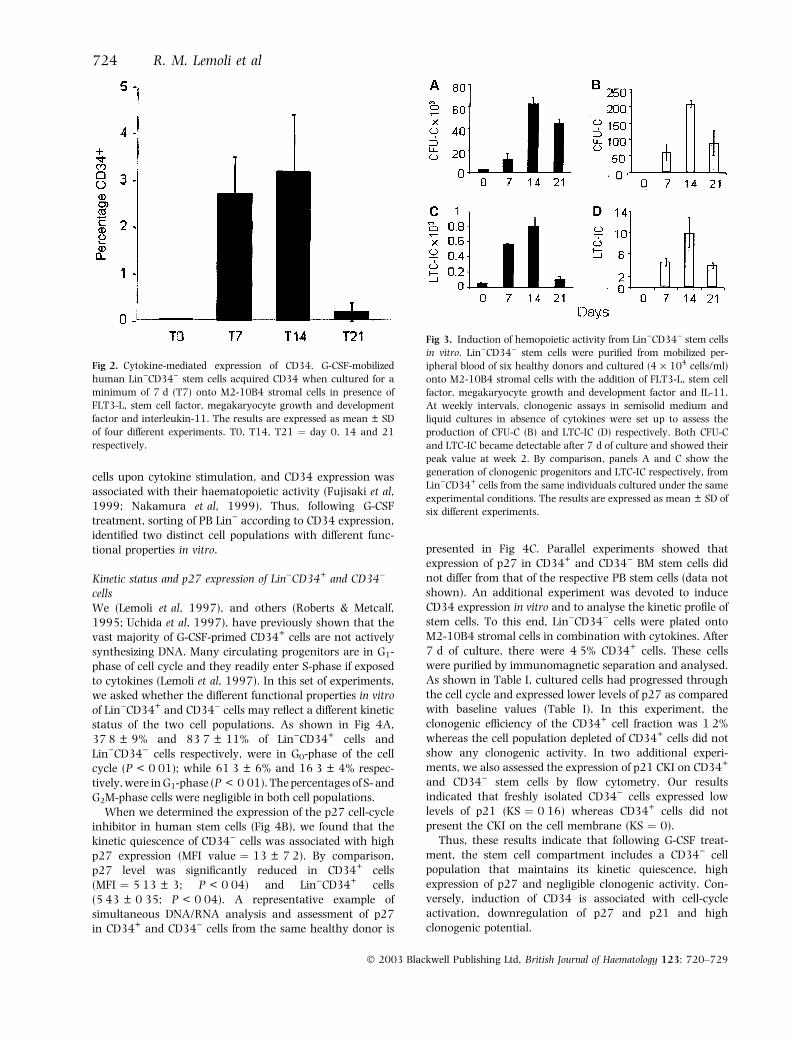
cells in liquid cultureTo further determine the functional characteristics ofG-CSF-primed stem cells, we plated Lin)CD34) ontoM2-10B4 in combination with rh FLT3-L, SCF, MGDF andIL-11. The surface expression of CD34 in these cultured cellsand the output of CFU-C and LTC-IC were analysed atweekly intervals. The mean frequency of CD34+ cells was2Æ7 ± 0Æ8% after 1 week, 3Æ2 ± 1Æ2% at week 2 and0Æ2 ± 0Æ2% at week 3 (Fig 2). Moreover, both CFU-C andLTC-IC (Fig 3) became readily detectable after 7 d of cultureand showed their peak value at week 2. Figure 3 shows thegeneration of clonogenic progenitors and LTC-IC fromLin)CD34+ cells from the same individuals cultured underthe same experimental conditions. The percentage of CD34+
cells declined to 65 ± 5%, 27 ± 8% and 3 ± 2% at week 1,2 and 3 respectively. However, because of the majorexpansion of the total number of cells during long-termculture, at week 2 we found mean fold-increases of 321Æ6and 16Æ6 in CFU-C and LTC-IC respectively.
Taken together, these results confirm that, similarly tounprimed BM and CB cells, G-CSF-stimulated Lin)CD34)
cells did not retain any colony-forming or LTC-IC ability.However, they could be induced to differentiate into CD34+
Kinetic Characterization of Human Stem Cells 723
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 123: 720–729
cells upon cytokine stimulation, and CD34 expression wasassociated with their haematopoietic activity (Fujisaki et al,1999; Nakamura et al, 1999). Thus, following G-CSFtreatment, sorting of PB Lin) according to CD34 expression,identified two distinct cell populations with different func-tional properties in vitro.
Kinetic status and p27 expression of Lin)CD34+ and CD34)
cellsWe (Lemoli et al, 1997), and others (Roberts & Metcalf,1995; Uchida et al, 1997), have previously shown that thevast majority of G-CSF-primed CD34+ cells are not activelysynthesizing DNA. Many circulating progenitors are in G1-phase of cell cycle and they readily enter S-phase if exposedto cytokines (Lemoli et al, 1997). In this set of experiments,we asked whether the different functional properties in vitroof Lin)CD34+ and CD34) cells may reflect a different kineticstatus of the two cell populations. As shown in Fig 4A,37Æ8 ± 9% and 83Æ7 ± 11% of Lin)CD34+ cells andLin)CD34) cells respectively, were in G0-phase of the cellcycle (P < 0Æ01); while 61Æ3 ± 6% and 16Æ3 ± 4% respec-tively, were in G1-phase (P < 0Æ01). The percentages of S- andG2M-phase cells were negligible in both cell populations.
When we determined the expression of the p27 cell-cycleinhibitor in human stem cells (Fig 4B), we found that thekinetic quiescence of CD34) cells was associated with highp27 expression (MFI value ¼ 13 ± 7Æ2). By comparison,p27 level was significantly reduced in CD34+ cells(MFI ¼ 5Æ13 ± 3; P < 0Æ04) and Lin)CD34+ cells(5Æ43 ± 0Æ35; P < 0Æ04). A representative example ofsimultaneous DNA/RNA analysis and assessment of p27in CD34+ and CD34) cells from the same healthy donor is
presented in Fig 4C. Parallel experiments showed thatexpression of p27 in CD34+ and CD34) BM stem cells didnot differ from that of the respective PB stem cells (data notshown). An additional experiment was devoted to induceCD34 expression in vitro and to analyse the kinetic profile ofstem cells. To this end, Lin)CD34) cells were plated ontoM2-10B4 stromal cells in combination with cytokines. After7 d of culture, there were 4Æ5% CD34+ cells. These cellswere purified by immunomagnetic separation and analysed.As shown in Table I, cultured cells had progressed throughthe cell cycle and expressed lower levels of p27 as comparedwith baseline values (Table I). In this experiment, theclonogenic efficiency of the CD34+ cell fraction was 1Æ2%whereas the cell population depleted of CD34+ cells did notshow any clonogenic activity. In two additional experi-ments, we also assessed the expression of p21 CKI on CD34+
and CD34) stem cells by flow cytometry. Our resultsindicated that freshly isolated CD34) cells expressed lowlevels of p21 (KS ¼ 0Æ16) whereas CD34+ cells did notpresent the CKI on the cell membrane (KS ¼ 0).
Thus, these results indicate that following G-CSF treat-ment, the stem cell compartment includes a CD34) cellpopulation that maintains its kinetic quiescence, highexpression of p27 and negligible clonogenic activity. Con-versely, induction of CD34 is associated with cell-cycleactivation, downregulation of p27 and p21 and highclonogenic potential.
Fig 2. Cytokine-mediated expression of CD34. G-CSF-mobilized
human Lin)CD34) stem cells acquired CD34 when cultured for a
minimum of 7 d (T7) onto M2-10B4 stromal cells in presence of
FLT3-L, stem cell factor, megakaryocyte growth and development
factor and interleukin-11. The results are expressed as mean ± SD
of four different experiments. T0, T14, T21 ¼ day 0, 14 and 21
respectively.
Fig 3. Induction of hemopoietic activity from Lin)CD34) stem cells
in vitro. Lin)CD34) stem cells were purified from mobilized per-
ipheral blood of six healthy donors and cultured (4 · 104 cells/ml)
onto M2-10B4 stromal cells with the addition of FLT3-L, stem cell
factor, megakaryocyte growth and development factor and IL-11.
At weekly intervals, clonogenic assays in semisolid medium and
liquid cultures in absence of cytokines were set up to assess the
production of CFU-C (B) and LTC-IC (D) respectively. Both CFU-C
and LTC-IC became detectable after 7 d of culture and showed their
peak value at week 2. By comparison, panels A and C show the
generation of clonogenic progenitors and LTC-IC respectively, from
Lin)CD34+ cells from the same individuals cultured under the same
experimental conditions. The results are expressed as mean ± SD of
six different experiments.
724 R. M. Lemoli et al
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 123: 720–729
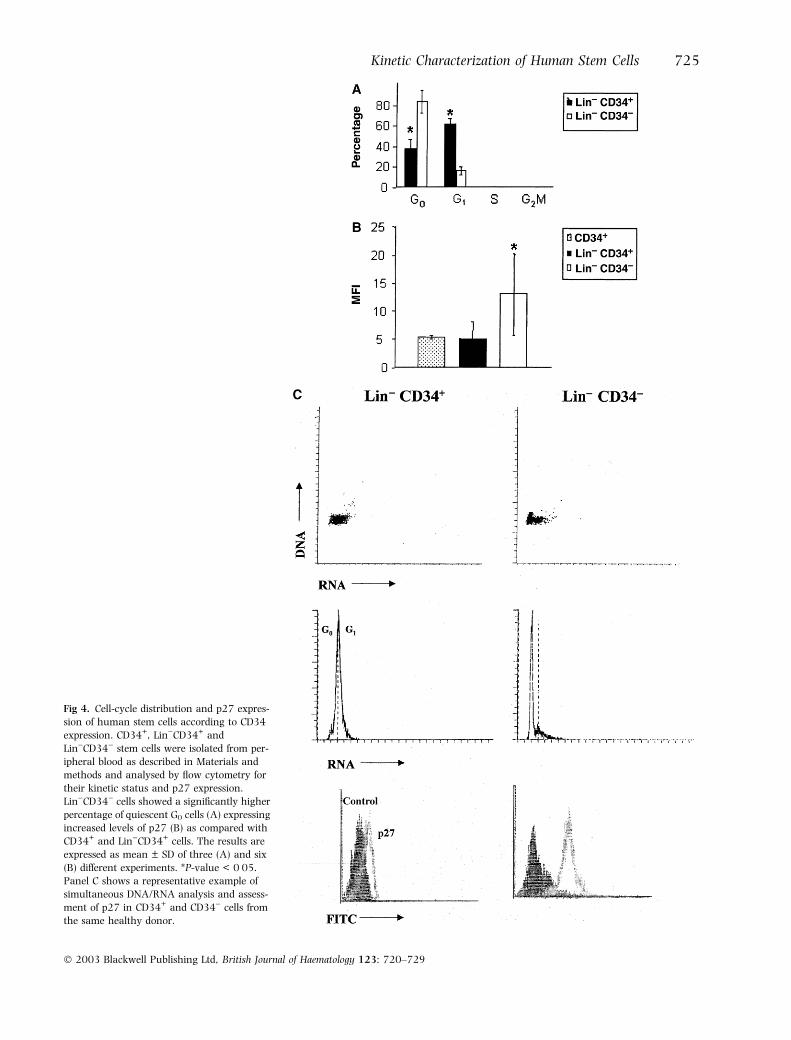
Fig 4. Cell-cycle distribution and p27 expres-
sion of human stem cells according to CD34
expression. CD34+, Lin)CD34+ and
Lin)CD34) stem cells were isolated from per-
ipheral blood as described in Materials and
methods and analysed by flow cytometry for
their kinetic status and p27 expression.
Lin)CD34) cells showed a significantly higher
percentage of quiescent G0 cells (A) expressing
increased levels of p27 (B) as compared with
CD34+ and Lin)CD34+ cells. The results are
expressed as mean ± SD of three (A) and six
(B) different experiments. *P-value < 0Æ05.
Panel C shows a representative example of
simultaneous DNA/RNA analysis and assess-
ment of p27 in CD34+ and CD34) cells from
the same healthy donor.
Kinetic Characterization of Human Stem Cells 725
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 123: 720–729

Lin)CD34+ and CD34) cells have different engraftment capacityin xenograft transplant modelsOn injection of purified G-CSF-primed Lin)CD34+ orLin)CD34) cells to sublethally irradiated NOD/RAG orNOD/SCID/B2M mice, we found that both progenitor cellpopulations showed haematopoietic potential. Figure 5reports the flow cytometric assessment of human myeloid(CD45+CD19)), B-lymphoid (CD45+CD19+) and progenitor(CD34+CD19)) cell engraftment in the BM of a representa-tive NOD/SCID/B2M mouse receiving 300 000 Lin)CD34)
cells, evaluated 90 d after transplant. Interestingly,Lin)CD34) cells were able to generate CD34+ cells in theBM of transplanted mice.
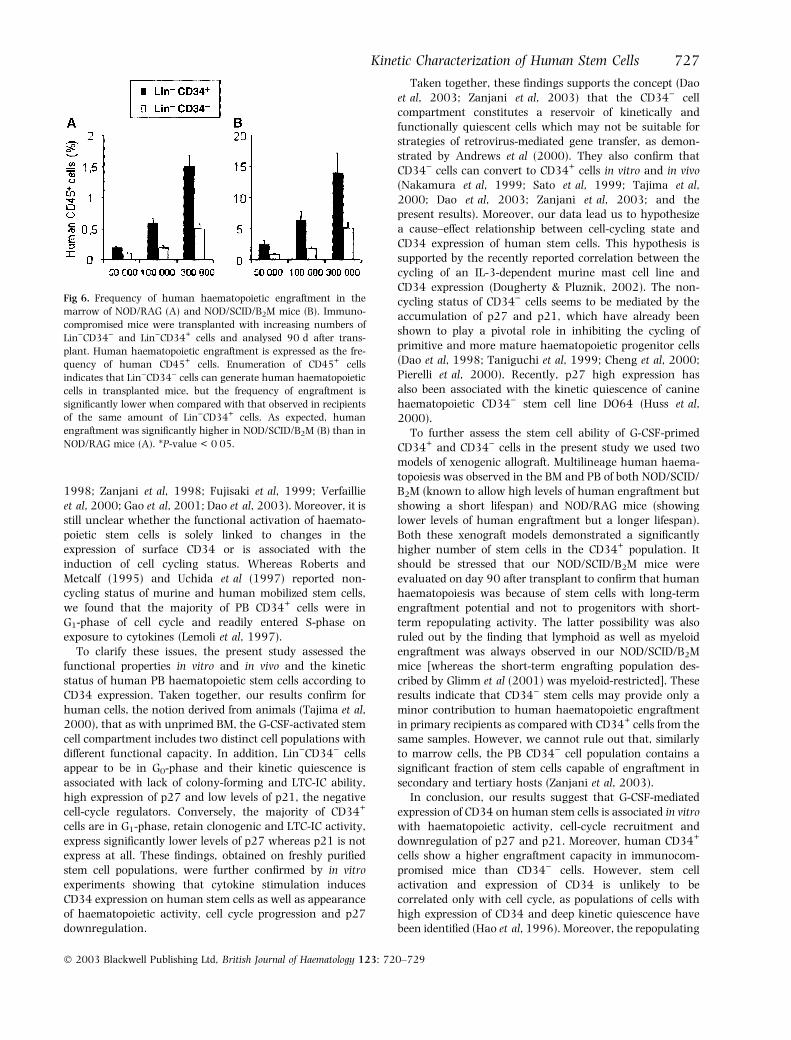
The RT-PCR evaluation of the expression of human CD45in PB of transplanted mice, showed human haematopoieticengraftment in 20 of 20 NOD/SCID/B2M mice and in five ofsix NOD/RAG mice (data not shown). The NOD/RAG mouselacking human engraftment was transplanted with 50 000Lin)CD34) cells. Quantification of human haematopoietic
engraftment by enumeration of human CD45+ cells indica-ted that Lin)CD34) cells can generate human haematopoi-etic cells in transplanted mice, but the frequency ofengraftment was significantly lower when compared withthat observed in recipients of the same amount of Lin)CD34+
cells (P < 0Æ05) (Fig 6). As expected, human engraftmentwas significantly higher in NOD/SCID/B2M than in NOD/RAG mice (Fig 6).
To explain the different engraftment results, we assessedthe expression of important molecules affecting homing toBM and engraftment of human haematopoietic stem cells inimmunodeficient mice. Flow cytometric analysis performedin two experiments demonstrated that a mean of 46, 69 and78% of Lin)CD34+ and 43, 59 and 76% of Lin)CD34)
expressed the chemokine SDF-1 (CXCL12) receptor CXCR4and the integrins CD49e (VLA-5) and CD49d (VLA-4)respectively, with no significant difference between the twocell populations as for the percentage of positive cells andthe MFI (data not shown).
DISCUSSION
Earlier observations have prompted the hypothesis thatmurine (Sato et al, 1999; Tajima et al, 2000) and human(Bhatia et al, 1998; Zanjani et al, 1998; Dao et al, 2003)haematopoietic stem cells exist in two functional states thatcan be distinguished by CD34 expression. As CD34) stemcells are extremely quiescent, they need to be activated bythe induction of CD34 in order to reach a functional statesuitable for efficient engraftment of recipient mice. In thisregard, G-CSF induces CD34 expression on murine stemcells, as well as their mobilization into PB (Tajima et al,2000). However, it remains to be determined whether thesame model of CD34 activation applies to human stem cellsduring G-CSF-mediated mobilization.
At present, conflicting published data exist on the stemcell capacity of human CD34+ and CD34) cells (Bhatia et al,
Fig 5. Flow cytometric assessment of human
stem cells engraftment in the bone marrow of
immunocompromised mice. A representative
NOD/SCID/B2M mouse was transplanted with
300 000 human Lin)CD34) cells and evalu-
ated 90 d after transplant. The top left panel
shows the gate used to exclude platelets,
debris and dead cells. The top right panel
shows the negative controls and the lower
panels show the enumeration gates for
human B cells (CD19+ CD45+), myeloid cells
(CD19) CD45+) and progenitor cells (CD34+
CD19)).
Table I. Induction of CD34 results in progression through the cell
cycle and downregulation of p27.
CD34+
cells
Cell-cycle distribution (%)p27
Expression (MFI)G0 G1 S G2M
T0 0 87Æ8 11Æ2 0Æ6 0Æ4 20Æ7T7 4Æ5 87Æ4 3 7Æ8 1Æ8 5Æ4
Peripheral blood Lin)CD34) cells were plated onto M2-10B4 cells
in combination with FLT3-L, stem cell factor, megakaryocyte
growth and development factor and interleukin-11 for 7 d. After
liquid culture (T7), cells expressing the CD34 antigen were purified
and analysed for their cell-cycle distribution and p27 expression.
The results were compared with those observed before culture (T0).
MFI, mean fluorescence intensity.
726 R. M. Lemoli et al
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 123: 720–729
1998; Zanjani et al, 1998; Fujisaki et al, 1999; Verfaillieet al, 2000; Gao et al, 2001; Dao et al, 2003). Moreover, it isstill unclear whether the functional activation of haemato-poietic stem cells is solely linked to changes in theexpression of surface CD34 or is associated with theinduction of cell cycling status. Whereas Roberts andMetcalf (1995) and Uchida et al (1997) reported non-cycling status of murine and human mobilized stem cells,we found that the majority of PB CD34+ cells were inG1-phase of cell cycle and readily entered S-phase onexposure to cytokines (Lemoli et al, 1997).
To clarify these issues, the present study assessed thefunctional properties in vitro and in vivo and the kineticstatus of human PB haematopoietic stem cells according toCD34 expression. Taken together, our results confirm forhuman cells, the notion derived from animals (Tajima et al,2000), that as with unprimed BM, the G-CSF-activated stemcell compartment includes two distinct cell populations withdifferent functional capacity. In addition, Lin)CD34) cellsappear to be in G0-phase and their kinetic quiescence isassociated with lack of colony-forming and LTC-IC ability,high expression of p27 and low levels of p21, the negativecell-cycle regulators. Conversely, the majority of CD34+
cells are in G1-phase, retain clonogenic and LTC-IC activity,express significantly lower levels of p27 whereas p21 is notexpress at all. These findings, obtained on freshly purifiedstem cell populations, were further confirmed by in vitroexperiments showing that cytokine stimulation inducesCD34 expression on human stem cells as well as appearanceof haematopoietic activity, cell cycle progression and p27downregulation.
Taken together, these findings supports the concept (Daoet al, 2003; Zanjani et al, 2003) that the CD34) cellcompartment constitutes a reservoir of kinetically andfunctionally quiescent cells which may not be suitable forstrategies of retrovirus-mediated gene transfer, as demon-strated by Andrews et al (2000). They also confirm thatCD34) cells can convert to CD34+ cells in vitro and in vivo(Nakamura et al, 1999; Sato et al, 1999; Tajima et al,2000; Dao et al, 2003; Zanjani et al, 2003; and thepresent results). Moreover, our data lead us to hypothesizea cause–effect relationship between cell-cycling state andCD34 expression of human stem cells. This hypothesis issupported by the recently reported correlation between thecycling of an IL-3-dependent murine mast cell line andCD34 expression (Dougherty & Pluznik, 2002). The non-cycling status of CD34) cells seems to be mediated by theaccumulation of p27 and p21, which have already beenshown to play a pivotal role in inhibiting the cycling ofprimitive and more mature haematopoietic progenitor cells(Dao et al, 1998; Taniguchi et al, 1999; Cheng et al, 2000;Pierelli et al, 2000). Recently, p27 high expression hasalso been associated with the kinetic quiescence of caninehaematopoietic CD34) stem cell line DO64 (Huss et al,2000).
To further assess the stem cell ability of G-CSF-primedCD34+ and CD34) cells in the present study we used twomodels of xenogenic allograft. Multilineage human haema-topoiesis was observed in the BM and PB of both NOD/SCID/B2M (known to allow high levels of human engraftment butshowing a short lifespan) and NOD/RAG mice (showinglower levels of human engraftment but a longer lifespan).Both these xenograft models demonstrated a significantlyhigher number of stem cells in the CD34+ population. Itshould be stressed that our NOD/SCID/B2M mice wereevaluated on day 90 after transplant to confirm that humanhaematopoiesis was because of stem cells with long-termengraftment potential and not to progenitors with short-term repopulating activity. The latter possibility was alsoruled out by the finding that lymphoid as well as myeloidengraftment was always observed in our NOD/SCID/B2Mmice [whereas the short-term engrafting population des-cribed by Glimm et al (2001) was myeloid-restricted]. Theseresults indicate that CD34) stem cells may provide only aminor contribution to human haematopoietic engraftmentin primary recipients as compared with CD34+ cells from thesame samples. However, we cannot rule out that, similarlyto marrow cells, the PB CD34) cell population contains asignificant fraction of stem cells capable of engraftment insecondary and tertiary hosts (Zanjani et al, 2003).
In conclusion, our results suggest that G-CSF-mediatedexpression of CD34 on human stem cells is associated in vitrowith haematopoietic activity, cell-cycle recruitment anddownregulation of p27 and p21. Moreover, human CD34+
cells show a higher engraftment capacity in immunocom-promised mice than CD34) cells. However, stem cellactivation and expression of CD34 is unlikely to becorrelated only with cell cycle, as populations of cells withhigh expression of CD34 and deep kinetic quiescence havebeen identified (Hao et al, 1996). Moreover, the repopulating
Fig 6. Frequency of human haematopoietic engraftment in the
marrow of NOD/RAG (A) and NOD/SCID/B2M mice (B). Immuno-
compromised mice were transplanted with increasing numbers of
Lin)CD34) and Lin)CD34+ cells and analysed 90 d after trans-
plant. Human haematopoietic engraftment is expressed as the fre-
quency of human CD45+ cells. Enumeration of CD45+ cells
indicates that Lin)CD34) cells can generate human haematopoietic
cells in transplanted mice, but the frequency of engraftment is
significantly lower when compared with that observed in recipients
of the same amount of Lin)CD34+ cells. As expected, human
engraftment was significantly higher in NOD/SCID/B2M (B) than in
NOD/RAG mice (A). *P-value < 0Æ05.
Kinetic Characterization of Human Stem Cells 727
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 123: 720–729

capacity of human mobilized PB CD34+ cells in NOD/SCIDmice was shown to be mainly expressed in G0 cells asopposed to G1 cells (Gothot et al, 1998). This is in contrastwith the kinetic status and the repopulating capacity ofCD34+ and CD34) cells shown in this study. Thus, themechanism(s) determining the ‘activation’ status and theincreased engraftment potential of human haematopoieticstem cells remain partially unknown. In this regard, we didnot find significant differences between CD34+ and CD34)
stem cells for the expression of major b1 integrins very lateantigen (VLA)-4 and VLA-5 and CXC chemokine receptor 4(CXCR4), which have been demonstrated to be crucial forthe engraftment of human haematopoietic cells in NOD/SCID mice (Peled et al, 1999; Peled et al, 2000). It may wellbe that cell cycle transit is associated with a continuallychanging stem cell phenotype and different functionalproperties in vitro and in vivo (Quesenberry et al, 2002).Further studies are in progress to correlate the functionalproperties of different subsets of stem cells with theirgenomic profile.
ACKNOWLEDGMENTS
The research was supported by AIL (Associazione ItalianaLeucemie), Rome, AIRC (Associazione Italiana per laRicerca contro il Cancro), Milan, University of Bologna(funds for selected topics), Italy.
REFERENCES
Andrews, R.G., Peterson, L.J., Morris, J., Potter, J., Heyward, S.,
Gough, M., Bryant, E. & Kiem, H.P. (2000) Differential engraft-
ment of genetically modified CD34+ and CD34) hematopoietic
cell subsets in lethally irradiated baboons. Experimental Hema-
tology, 28, 508–518.
Bensinger, W.I., Buckner, C.D., Shannon-Dorcy, K., Rowley, S.,
Appelbaum, F.R., Benyunes, M., Clift, R., Martin, P., Demirer, T.,
Storb, R., Lee, M. & Schiller G. (1996) Transplantation of allo-
geneic CD34+ peripheral blood stem cells in patients with
advanced hematologic malignancy. Blood, 88, 4132–4138.
Berenson, R.J., Andrews, R.G., Bensinger, W.I., Kalamasz, D.,
Knitter, G., Buckner, C.D. & Bernsrein, I.D. (1988) Antigen
CD34+ marrow cells engraft lethally irradiated baboons. Journal
of Clinical Investigation, 81, 951–955.
Bhatia, M., Bonnet, D., Murdoch, B., Gan, O.I. & Dick, J.E. (1998) A
newly discovered class of human hematopoietic cells with SCID-
repopulating activity. Nature Medicine, 9, 1038–1045.
Cheng, J., Baumhueter, S., Cacalano, G., Carver-Moore, K., Thibo-
deaux, H., Thomas, R., Broxmeyer, H.E., Cooper, S., Hague, N.,
Moore, M. & Lasky L.A. (1996) Hematopoietic defects in mice
lacking the sialomucin CD34. Blood, 87, 479–484.
Cheng, T., Rodrigues, N., Dombkowsky, D., Stier, S. & Scadden, D.T.
(2000) Stem cell repopulation efficiency but not pool size is
governed by p27kip1. Nature Medicine, 6, 1235–1240.
Curti, A., Tafuri, A., Ricciardi, M.R., Tazzari, P., Petrucci, M.T.,
Fogli, M., Ratta, M., Lapalombella, R., Ferri, E., Tura, S., Bacca-
rani, M. & Lemoli, R.M. (2002) Interleukin-11 induces pro-
liferation of human T-cells and its activity is associated with
downregulation of p27kip1. Haematologica, 87, 373–380.
Dao, M.A., Taylor, N. & Nolta, J.A. (1998) Reduction in levels of
the cyclin-dependent kinase inhibitor p27kip-1 coupled with
transforming growth factor b neutralization induces cell-cycle
entry and increases retroviral transduction of primitive human
hematopoietic cells. Proceedings of the National Academy of Sciences
of the United States of America, 95, 13 006–13 011.
Dao, M.A., Arevalo, J. & Nolta, J.A. (2003) Reversibility of CD34
expression on human hematopoietic stem cells that retain the
capacity for secondary reconstitution. Blood, 101, 112–118.
Dell’Agnola, C., Rabascio, C., Mancuso, P., Capillo, M., Pruneri, G.,
Gobbi, A., Minucci, S., Ronzoni, S., Volorio, S., Calabrese, L.,
Tradati, N., Martinelli, G., Shultz, L. & Bertolini, F. (2002) In vitro
and in vivo hematopoietic potential of human stem cells residing
in muscle tissue. Experimental Hematology, 30, 905–914.
Dougherty, S. & Pluznik, D. (2002) Expression of the hematopoietic
stem cell marker CD34 is related to the cell cycle status
(Abstract). Experimental Hematology, 30(Suppl. 1), 57a.
Fujisaki, T., Berger, M.G., Rose-John, S. & Eaves, C.J. (1999) Rapid
differentiation of a rare subset of adult human Lin-CD34-CD38-
cells stimulated by multiple growth factors in vitro. Blood, 94,
1926–1932.
Gao, Z., Fackler, M.J., Leung, W., Lumkul, R., Ramirez, M., Theo-
bald, N., Malech, H.L. & Civin, C.I. (2001) Human CD34+ cell
preparations contain over 100-fold greater NOD/SCID mouse
engrafting capacity than do CD34-cell preparations. Experimental
Hematology, 29, 910–921.
Glimm, H., Eisterer, W., Lee, K., Cashman, J., Holyoake, T.L.,
Nicolini, F., Shultz, L.D., von Kalle, C. & Eaves, C.J. (2001) Pre-
viously undetected human hematopoietic cell populations with
short-term repopulating activity selectively engraft NOD/SCID-
beta2 microglobulin-null mice. Journal of Clinical Investigation,
107, 199–206.
Goodell, M.A., Rosenzweig, M., Kim, H., Marks, D.F., DeMaria, M.,
Paradis, G., Grupp, S.A., Sieff, C.A., Mulligan, R.C. & Johnson,
R.P. (1997) Dye efflux studies suggest that hematopoietic stem
cells expressing low or undetectable levels of CD34 antigen exist
in multiple species. Nature Medicine, 3, 1337–1344.
Gothot, A., van der Loo, J.C. & Srour, E.F. (1998) Cell cycle-related
changes in repopulating capacity of human mobilized peripheral
blood CD34+ cells in non-obese diabetic/severe combined
immune-deficient mice. Blood, 92, 2641–2649.
Hao, Q.L., Thiemann, F.T., Petersen, D., Smogorzewska, E.M. &
Crooks, G.M. (1996) Extended long-term culture reveals a highly
quiescent and primitive human hematopoietic progenitor popu-
lation. Blood, 88, 3306–3313.
Hengst, L. & Reed, S. (1996) Translation control of p27 accumu-
lation during the cell cycle. Science, 271, 1861–1864.
Huss, R., Gatsios, P., Graeve, L., Lange, C., Eissner, G., Kolb, H.J.,
Thalmeier, K. & Heinrich, P.C. (2000) Quiescence of CD34-
negative hematopoietic stem cells is mediated by downregulation
of cyclin B and no Stat activation. Cytokine, 12, 1195–1204.
Kollet, O., Peled, A., Byk, T., Ben-Hur, H., Greiner, D., Shultz, L. &
Lapidot T. (2000) Beta2 microglobulin-deficient (B2m(null))
NOD/SCID mice are excellent recipients for studying human stem
cell function. Blood, 95, 3102–3105.
Krause, D.S., Fackler, M.J., Civin, C.I. & May, W.S. (1996) CD34:
structure, biology and clinical utility. Blood, 87, 1–13.
Lemoli, R.M., Tafuri, A., Fortuna, A., Petrucci, M.T., Ricciardi,
M.R., Catani, L., Rondelli, D., Fogli, M., Leopardi, G., Ariola, C. &
Tura S. (1997) Cycling status of CD 34+ cells mobilized into
peripheral blood of healthy donors by recombinant human gra-
nulocyte colony-stimulating factor. Blood, 89, 1189–1196.
Lemoli, R.M., Martinelli, G., Zamagni, E., Motta, M.R., Rizzi, S.,
Terragna, C., Rondelli, R., Ronconi, S., Curti, A., Bonifazi, F.,
Tura, S. & Cavo, M. (2000) Engraftment, clinical, and molecular
follow-up of patients with multiple myeloma who were reinfused
728 R. M. Lemoli et al
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 123: 720–729

with highly purified CD34+ cells to support single or tandem
high-dose chemotherapy. Blood, 95, 2234–2239.
Nakamura, Y., Ando, K., Chargui, J., Sato, T., Tsuji, T., Hotta, T. &
Kato, S. (1999) Ex vivo generation of CD34+ cells from CD34-
hematopoietic cells. Blood, 94, 4053–4059.
Osawa, M., Hanada, K., Hamada, H. & Nakauchi, H. (1996) Long-
term lymphohematopoietic reconstitution by a single Cd34-low/
negative hematopoietic stem cell. Science, 273, 242–245.
Peled, A., Petit, I., Kollet, O., Magid, M., Ponomaryov, T., Byk, T.,
Nagler, A., Ben-Hur, H., Many, A., Shultz, L., Lider, O., Alon, R.,
Zipori, D. & Lapidot, T. (1999) Dependence of human stem cell
engraftment and repopulation of NOD/SCID mice on CXCR4.
Science, 283, 845–848.
Peled, A., Kollet, O., Ponomaryov, T., Petit, I., Franitza, S., Grab-
ovsky, V., Slav, M.M., Nagler, A., Lider, O., Alon, R., Zipori, D. &
Lapidot, T. (2000) The chemokine SDF-1 activates the integrins
LFA-1, VLA-4 and VLA-5 on immature human CD34+ cells: role
in transendhotelial/stromal migration and engraftment of NOD/
SCID mice. Blood, 95, 3289–3296.
Pierelli, L., Marone, M., Bonanno, G., Mozzetti, S., Rutella, S.,
Morosetti, R., Rumi, C., Mancuso, S., Leone, G. & Scambia, G.
(2000) Modulation of bcl-2 and p27 in human primitive pro-
liferating hematopoietic progenitors by autocrine TGF-b1 is a cell
cycle-independent effect and influences their hematopoietic
potential. Blood, 95, 3001–3010.
Quesenberry, P.J., Colvin, G.A. & Lambert, J.-F. (2002) The
chiaroscuro stem cell: a unified stem cell theory. Blood, 100,
4266–4271.
Ricciardi, M.R., Petrucci, M.T., Gregorj, C., Ariola, C., Lemoli, R.M.,
Fogli, M., Mauro, F.R., Cerretti, R., Foa, R., Mandelli, F. & Tafuri
A. (2001) Reduced susceptibility to apoptosis correlates with
kinetic quiescence in disease progression of chronic lymphocytic
leukemia. British Journal of Haematology, 113, 1–10.
Roberts, A.W. & Metcalf, D. (1995) Noncycling state of peripheral
blood progenitor cells mobilized by granulocyte colony-stimulat-
ing factor and other cytokines. Blood, 86, 1600–1605.
Sato, T., Laver, J.H. & Ogawa, M. (1999) Reversible expression of
CD34 by murine hematopoietic stem cells. Blood, 94, 2548–2554.
Shultz, L.D., Lang, P.A., Christianson, S.W., Gott, B., Lyons, B.,
Umeda, S., Leiter, E., Hesselton, R., Wagar, E.J., Leif, J.H., Kollet,
O., Lapidot, T. & Greiner, D.L. (2000) NOD/LtSz-Rag1null mice:
an immunodeficient and radioresistant model for engraftment of
human hematolymphoid cells, HIV infection, and adoptive
transfer of NOD mouse diabetogenic T cells. Journal of
Immunology, 164, 2496–2507.
Tafuri, A., Lemoli, R.M., Petrucci, M.T., Ricciardi, M.R., Fogli, M.,
Bonsi, L., Ariola, C., Strippoli, P., Gregorj, C., Petti, M.C., Tura, S.,
Mandelli, F. & Bagnara, G.P. (1999) Thrombopoietin and inter-
leukin-11 have different modulatory effects on cell cycle and
programmed cell death in primary acute myeloid leukemia cells.
Experimental Hematology, 27, 1255–1263.
Tafuri, A., Gregorj, C., Petrucci, M.T., Ricciardi, M.R., Mancini,
M., Cimino, G., Mecucci, C., Tedeschi, A., Fioritoni, G., Ferrara,
F., Di Raimondo, F., Gallo, E., Liso, V., Fabbiano, F., Cascavilla,
N., Pizzolo, G., Camera, A., Pane, F., Lanza, F., Cilloni, D.,
Annino, L., Vitale, A., Vegna, M.L, Vignetti, M., Foa, R. &
Mandelli, F for the GIMEMA Group. (2002) MDR1 protein
expression is an independent predictor of complete remission in
newly diagnosed adult acute lymphoblastic leukemia. Blood,
100, 974–981.
Tajima, F., Sato, T., Laver, J.H. & Ogawa, M. (2000) CD34
expression by murine hematopoietic stem cells mobilized by
granulocyte colony-stimulating factor. Blood, 96, 1989–1993.
Taniguchi, T., Endo, H., Chikatsu, N., Uchimaru, K., Asano, S.,
Fujita, T., Nakahata, T. & Motukura, T. (1999) Expression of
p21Cip1/waf1/Sdi1 and p27kip-1 Cyclin-dependent kinase inhibitors
during human hematopoiesis. Blood, 93, 4167–4178.
Uchida, N., He, D., Friera, A.M., Reitsma, M., Sasaki, D., Chen, B. &
Tsukamoto, A. (1997) The unexpected G0/G1 cell cycle status of
mobilized hematopoietic stem cells from peripheral blood. Blood,
89, 465–472.
Verfaillie, C.M., Almeida-Porada, G., Wissink, S. & Zanjani, E.D.
(2000) Kinetics of engraftment of CD34) and CD34+ cells from
mobilized blood differs from that of CD34) and CD34+ cells from
bone marrow. Experimental Hematology, 28, 1071–1079.
Zanjani, E.D., Almeida-Porada, G., Livingston, A.G., Flake, A.W. &
Ogawa, M. (1998) Human bone marrow CD34) cells engraft
in vivo and undergo multilineage expression that includes
giving rise to CD34+ cells. Experimental Hematology, 26, 353–
360.
Zanjani, E.D., Almeida-Porada, G., Livingston, A.G., Zeng, H.Q. &
Ogawa, M. (2003) Reversible expression of CD34 by adult
human bone marrow long-term engrafting hematopoietic stem
cells. Experimental Hematology, 31, 406–412.
Kinetic Characterization of Human Stem Cells 729
� 2003 Blackwell Publishing Ltd, British Journal of Haematology 123: 720–729




















