
Hybrid HIV/MSCV LTR Enhances Transgene Expression of Lentiviral Vectors in Human CD34 +...
11
Hybrid HIV/MSCV LTR Enhances Transgene Expression of Lentiviral Vectors in Human CD34 + Hematopoietic Cells JOHN KIM CHOI, a NGHIA HOANG, a ANTONINA M. VILARDI, a PATRICIA CONRAD, c STEPHEN G. EMERSON, b ALAN M. GEWIRTZ b a Department of Pathology and Laboratory Medicine and b Department of Medicine, University of Pennsylvania, Philadelphia, USA; c Department of Pediatrics, Children’s Hospital of Philadelphia, Philadelphia, Pennsylvania, USA Key Words. Lentiviral vector · MSCV-based vector · Human CD34 + cells · Hybrid LTR ABSTRACT HIV-based lentiviral vectors can transduce nondivid- ing cells, an important advantage over murine leukemia virus (MLV)-based vectors when transducing slowly divid- ing hematopoietic stem cells. However, we find that in human CD34 + hematopoietic cells, the HIV-based vectors with an internal cytomegalovirus (CMV) promoter express transgenes 100- to 1,000-fold less than the MLV-based retroviral vector murine stem cell virus (MSCV). To increase the expression of the integrated lentivirus, we replaced CMV promoter with that of the Rous sarcoma virus or MSCV and obtained a modest augmentation in expression. A more dramatic effect was seen when the CMV enhancer/promoter was removed and the HIV long- terminal repeat (LTR) was replaced by a novel HIV/MSCV hybrid LTR. This vector retains the ability to transduce nondividing cells but now expresses its transgene (enhanced green fluorescent protein) 10- to 100-fold greater than the original HIV-based vector. When compared under identical conditions, the HIV vector with the hybrid LTR transduced a higher percentage of CD34 + cells than the MSCV-based retroviral vector (19.4% versus 2.4%). The number of transduced cells and level of transgene expres- sion remain constant over 5-8 weeks as determined by long- term culture-initiating cells, fluoresence-activated cell sorting, and nonobese diabetic/severe combined immuno- deficiency repopulation assay. Stem Cells 2001;19:236-246 STEM CELLS 2001;19:236-246 www.StemCells.com Correspondence: John Kim Choi, M.D., Ph.D., University of Pennsylvania, 413A SCL, 422 Curie Boulevard, Philadelphia, Pennsylvania 19104, USA. Telephone: 215-573-6527; Fax: 215-573-6523; e-mail: [email protected]; Alan M. Gewirtz, M.D., University of Pennsylvania School of Medicine, Department of Pathology and Internal Medicine, Rm 713 BRB II/III, 421 Curie Boulevard, Philadelphia, Pennsylvania 19104, USA. Telephone: 215-898-4499; Fax: 215-573-2078; e-mail: [email protected] Received January 18, 2001; accepted for publication February 19, 2001. ©AlphaMed Press 1066-5099/2001/$5.00/0 INTRODUCTION HIV-based vectors are attractive vehicles for delivering transgenes into slow to nondividing cells such as human hematopoietic stem cells [1-5], neurons [6], skeletal myocytes [7], and hepatocytes [7]. However, one significant disadvan- tage of the HIV-based vectors for hematopoietic applications is the weak activity of the HIV long-terminal repeat (LTR) in cells of this lineage. To increase the activity, HIV-based vec- tors that express the HIV accessory protein tat have been engi- neered [1, 3, 5]. A concern with this strategy is that the tat protein enhances transcription of other cellular genes that may be detrimental to the cells [8-13]. Furthermore, tat pro- tein may also enter into neighboring cells [14] with similar effects. Other first and second generation HIV-based vectors do not express tat but use an internal cytomegalovirus (CMV) promoter to drive the expression of the transgene [2, 4]. While exploring various experimental systems to express ectopic oncogenes in primary human CD34 + cells, we found that the HIV-based vectors with an internal CMV poorly express its transgene when compared to the murine leukemia virus (MLV)-based vector murine stem cell virus (MSCV). This poor expression could hamper studies that require a threshold quantity of transgene for a biologic effect. The poor expression also has clinical implications in gene therapy. To explore approaches for increasing transgene expression, we engineered various modifications of a first generation
Transcript of Hybrid HIV/MSCV LTR Enhances Transgene Expression of Lentiviral Vectors in Human CD34 +...

Hybrid HIV/MSCV LTR Enhances Transgene Expression of LentiviralVectors in Human CD34+ Hematopoietic Cells
JOHN KIM CHOI,a NGHIA HOANG,a ANTONINA M. VILARDI,a PATRICIA CONRAD,c
STEPHEN G. EMERSON,b ALAN M. GEWIRTZb
aDepartment of Pathology and Laboratory Medicine and bDepartment of Medicine,
University of Pennsylvania, Philadelphia, USA; cDepartment of Pediatrics, Children’s Hospital
of Philadelphia, Philadelphia, Pennsylvania, USA
Key Words. Lentiviral vector · MSCV-based vector · Human CD34+ cells · Hybrid LTR
ABSTRACT
HIV-based lentiviral vectors can transduce nondivid-ing cells, an important advantage over murine leukemiavirus (MLV)-based vectors when transducing slowly divid-ing hematopoietic stem cells. However, we find that inhuman CD34+ hematopoietic cells, the HIV-based vectorswith an internal cytomegalovirus (CMV) promoter expresstransgenes 100- to 1,000-fold less than the MLV-basedretroviral vector murine stem cell virus (MSCV). Toincrease the expression of the integrated lentivirus, wereplaced CMV promoter with that of the Rous sarcomavirus or MSCV and obtained a modest augmentation inexpression. A more dramatic effect was seen when theCMV enhancer/promoter was removed and the HIV long-
terminal repeat (LTR) was replaced by a novel HIV/MSCVhybrid LTR. This vector retains the ability to transducenondividing cells but now expresses its transgene(enhanced green fluorescent protein) 10- to 100-fold greaterthan the original HIV-based vector. When compared underidentical conditions, the HIV vector with the hybrid LTRtransduced a higher percentage of CD34+ cells than theMSCV-based retroviral vector (19.4% versus 2.4%). Thenumber of transduced cells and level of transgene expres-sion remain constant over 5-8 weeks as determined by long-term culture-initiating cells, fluoresence-activated cellsorting, and nonobese diabetic/severe combined immuno-deficiency repopulation assay. Stem Cells 2001;19:236-246
STEM CELLS 2001;19:236-246 www.StemCells.com
Correspondence: John Kim Choi, M.D., Ph.D., University of Pennsylvania, 413A SCL, 422 Curie Boulevard, Philadelphia,Pennsylvania 19104, USA. Telephone: 215-573-6527; Fax: 215-573-6523; e-mail: [email protected]; Alan M.Gewirtz, M.D., University of Pennsylvania School of Medicine, Department of Pathology and Internal Medicine, Rm 713 BRBII/III, 421 Curie Boulevard, Philadelphia, Pennsylvania 19104, USA. Telephone: 215-898-4499; Fax: 215-573-2078; e-mail:[email protected] Received January 18, 2001; accepted for publication February 19, 2001. ©AlphaMed Press1066-5099/2001/$5.00/0
INTRODUCTION
HIV-based vectors are attractive vehicles for deliveringtransgenes into slow to nondividing cells such as humanhematopoietic stem cells [1-5], neurons [6], skeletal myocytes[7], and hepatocytes [7]. However, one significant disadvan-tage of the HIV-based vectors for hematopoietic applicationsis the weak activity of the HIV long-terminal repeat (LTR) incells of this lineage. To increase the activity, HIV-based vec-tors that express the HIV accessory protein tat have been engi-neered [1, 3, 5]. A concern with this strategy is that the tatprotein enhances transcription of other cellular genes thatmay be detrimental to the cells [8-13]. Furthermore, tat pro-tein may also enter into neighboring cells [14] with similar
effects. Other first and second generation HIV-based vectorsdo not express tat but use an internal cytomegalovirus (CMV)promoter to drive the expression of the transgene [2, 4].
While exploring various experimental systems to expressectopic oncogenes in primary human CD34+ cells, we foundthat the HIV-based vectors with an internal CMV poorlyexpress its transgene when compared to the murine leukemiavirus (MLV)-based vector murine stem cell virus (MSCV).This poor expression could hamper studies that require athreshold quantity of transgene for a biologic effect. The poorexpression also has clinical implications in gene therapy.To explore approaches for increasing transgene expression,we engineered various modifications of a first generation

Choi, Hoang, Vilardi et al. 237
HIV-based vector driven by an internal CMV enhancer/pro-moter [15]. Of these, transgene expression was highest with avector in which the internal CMV enhancer/promoter wasdeleted and a portion of the U3 region of the 3′ HIV LTR wasreplaced with the U3 region of MSCV LTR. The 3′ hybrid LTRduplicates and replaces the 5′ LTR during viral reverse tran-scription leading to expression that is driven by the HIV/MSCVhybrid enhancer/promoter. This modified vector retains theproperty to transduce nondividing cells and expresses its trans-gene 10-100-fold greater than the original HIV-based vectorcontaining the internal CMV enhancer/promoter.
MATERIALS AND METHODS
VectorsThe original HIV vector pHR’CMVLacZ, the packaging
plasmid CMV∆R8.2, and the VSV-G envelope plasmid pMD.G[6] were generously provided by Inder M. Verma, Salk Institute;La Jolla, CA. The MSCV-based vector MIGR1 [16] wasgenerously provided by Warren Pear, University ofPennsylvania, Philadelphia, PA. The RSV and MSCVenhancer/promoters were isolated by polymerase chain reac-tion (PCR) amplification of the pBK-RSV (Stratagene; LaJolla, CA; http://www.Stratagene.com) and MIGR1 plasmids,respectively. The hybrid MSCV/HIV LTR was generated byPCR amplification and extension using the primers listed inTable 1. The PCR products were verified by sequencing andintroduced into the original HIV vector using standard methods.
Generation of Virus
MSCV-Based RetrovirusRetroviruses were generated using previously described
methods [16]. Briefly, MIGR1 and a vector that encodes theviral envelope protein VSV-G were transiently cotransfected
into the packaging cell line glycoprotein (GP) (generously pro-vided by Garry P. Nolan, Stanford University Medical Center;Stanford, CA). During the next 3 days, the transfected GP cellsrelease retroviruses into the culture medium; each ml of mediacontains 105 to 107 infectious viral particles using K562 as tar-get cells. The culture media were collected 48 and 72 hourspost-transfection and stored in small aliquots at -70ºC.
HIV-Based LentivirusLentiviral particles were generated in 293T cells using
transient transfection with plasmids encoding the RNAgenome, viral protein, and VSV-G envelope protein fol-lowing described methods [6]. Viral particles were col-lected and stored as described for the MSCV-basedretrovirus. Viral collection was tested for replication-com-petent viruses by detecting for rescue of integrated provirusthat expresses enhanced green fluorescent protein (EGFP).
Viral ConcentrationVSV-G pseudotyped retroviruses were concentrated by
filtration using Biomax low-speed centrifuge filtration con-centration (Millipore; Medford, MA).
The titer of the virus was determined by transducingK562 as target cells, and the number of EGFP+ cells wasdetermined by fluorescence-activated cell sorter analysis(FACS) 5-7 days later.
TransductionFour-hundred thousand primary CD34+ cells, 50 µl of
unconcentrated or concentrated virus, and 8 µg/ml of polybrene(Sigma; St. Louis, MO; http://www.sigma-aldrich.com) werecentrifuged in a 96-well plate at 1,500 × g for 90 minutes atroom temperature following published protocols [17]. For allexperiments, cells were transduced using a single exposure tothe viral stock. Afterwards, the cells were washed once with10 volumes of serum-free media and then cultured.
Tissue Culture of Cells
Cell LinesK562 cells were cultured in RPMI supplemented with
10% fetal bovine serum (FBS). 293T, GP, and HeLa cellswere cultured in Dulbecco’s modified Eagle’s medium sup-plemented with 10% FBS. Nondividing HeLa cells wereestablished using published protocols [18]. Briefly, HeLacells were seeded to generate plates that were 25% confluentand then treated with aphidicolin (15 µg/ml) 24 hours priorto transduction and daily thereafter.
Primary Human Hematopoietic CellsCells from bone marrow (BM) aspirates or umbilical cord
blood (UCB) were purified by Ficoll gradient and plated in
Table 1. Oligonucleotides to generate enhancer promoter
RSV
5′ GAA GAT CTG CAT CTG CTC CCT GCT TGT5′ CCG CTC GAG GGA TCC GCT AGC CAG GTG CAC ACC AA
MSCV
5′ CCA TCG ATA GAT CTG AAA GAC CCC ACC TGT AGG5′ CCG CTC GAG GGA TCC ACG GGT ACC CGG GCG ACG CA
MSCV/HIV hybrid
HIV-U3 5′A: 5′ CCG CTC GAG ACC TGG AAA AAC ATGHIV-U3 3′C: 5′ AGG TGG GGT CTT TCA ATC CAC AGA TCA AGG ATA TC
MSCV-LTR 3′C: 5′ TGA GGG CTC GCC ACT ACG GGT ACC CGG GCG ACG CAMSCV-LTR 5′C: 5′ TGA AAG ACC CCA CCT GTA GG
HIV-RU5 5′B: 5′ AGT GGC GAG CCC TCA GAT GCHIV-RU5 3′A: 5′ GGA CTA GTG AAG CAC TCA AGG CAA GCT

238 Expression of Novel Lentiviral Vectors in CD34 Cells
Iscove’s modified Dulbecco’s medium (IMDM) supple-mented with 10% FBS at 5-10 million cells/ml. After over-night incubation, the nonadherent mononuclear cells werefurther processed for CD34+ cells by positive immunomag-netic selection using the MACS system (Miltenyi Biotec;Auburn, CA; http://www.miltenyibiotec.com). CD34+ cellswere cultured in IMDM supplemented with 10.0% FBSsupplemented with the growth factors stem cell factor(SCF, 100 ng/ml), thrombopoietin (TPO, 50 ng/ml), andflk-3 (100 ng/ml) and the cells cultured for 2-3 days prior totransduction. For transduction without growth factors,freshly isolated CD34+ cells were transduced.
Long-Term Bone Marrow CulturePrimary CD34+ BM or UCB cells were transduced and
cultured on a monolayer of irradiated primary BM stromalcells for 5-8 weeks with IMDM supplemented with 12.5%horse serum and 12.5% FBS following published protocols[19]. At weekly intervals, half of the media was replaced withfresh media and cells were demi-depleted. After 5-8 weeks,unattached cells and trypsinized attached cells were cultured inmethylcellulose (Stem Cell Technology; Vancouver, BC,Canada; http://www.stemcell.com) supplemented with SCF(100 ng/ml), interleukin 3 (IL-3, 20 ng/ml), IL-6 (20 ng/ml),IL-11 (50 ng/ml), GM-CSF (20 ng/ml), and erythropoietin (1 µ/ml). Colonies were scored 10-15 days later under phaseand ultraviolet (UV) microscopy.
AnimalsA colony of NOD/LtSZ-scid/scid mice (NOD/SCID)
was established in the animal facility of the University ofPennsylvania. They were housed in microisolator cagesand provided with autoclaved food and water. For therepopulation assay, mice were irradiated with two 150 cGyfractions 2 hours prior to injection. 2 × 106 transducedUCB cells were injected via tail vein. After 5 weeks, themice were sacrificed, and BM cells were isolated from thefemurs and tibias.
DNA IsolationGenomic DNA was isolated using the QIAamp DNA
purification columns (Qiagen; Valencia, CA; http://www.qiagen.com) using manufacturer’s protocol. DNA was quan-tified by UV absorption spectrophotometry and ethidiumbromide binding.
PCRPCR was performed using standard procedure. Briefly, 5 ng
of genomic DNA was amplified in 50 µl reaction for 30 cyclesusing 0.5 units of Amplitaq Gold (Perkin Elmer; Norwalk, CT;http://www.instruments.perkinelmer.com/index.asp), 0.5 uM ofeach primer, and PCR parameters that consisted of denaturing at95°C for 30 seconds, annealing at 60°C for 30 seconds, andelongating at 72°C for 30 seconds. EGFP primer sequenceswere 5′ ATC CTG GTC GAG CTG GAC GGC and 5′ CGTGCT GCT TCA TGT GGT CGG G. Insulin receptor primersequences were 5′ GGA ACA ACC TCA CTA GGT TGC Aand 5′ CTG GCA GTG ACT ATG AGT CCA A.
FACS AnalysisFACS analysis was performed on FACSTAR and ana-
lyzed using Cell-Quest software (Becton Dickinson; San Jose,CA; http://www.bd.com). Phycoerythrin-conjugated antibod-ies to CD34 and CD45 were purchased from PharMingen (San Diego, CA; http://www.pharmingen.com).
RESULTS
Integration and Expression of HIV Lentiviral VectorCompared to MSCV Vector
The original HIV-based lentiviral vector encodes forbeta-galactosidase while the MSCV-based MIGR1 retrovi-ral vector encodes for EGFP. The different transgenes inthe two vectors require different detection systems and thusprevent direct comparison of the transduction efficiencies.To better compare the two vectors, the beta-galactosidasesequence of the original HIV vector was replaced with the
Figure 1. Diagram of constructedretroviral vectors. The two parentalvectors MIGR1 and pHR′CMVLacZwere obtained and modified usingstandard and PCR cloning tech-niques. To directly comparebetween MSCV and HIV-based sys-tems, the multicloning site (MCS)and LacZ of the original lentiviralvector were replaced with the MCS,internal ribosomal entry site, andenhanced green fluorescent protein (EGFP) from a bicistronic MSCV-based vector. LTR = long terminal repeats; ψ = viral packagingsignal; CMV = CMV enhancer/promoter; RSV = RSV enhancer/promoter; MSCV = MSCV enhancer/promoter; MCS = multicloning site;IRES = internal ribosomal entry site; GFP = enhanced green fluorescent protein.
MSCV
MSCV
MSCVGFP
MSCV
LacZ
GFP
GFP
GFP
GFP
MCS
MCS
MCS
MCS
MCS
MCS IRES
IRES
IRES
IRES
IRES
RSV
CMV
CMV
LTR
LTR
LTR
LTR
LTR
LTR
LTR
LTR
LTR
ψ
ψ
ψ
ψ
ψ
ψ
MIGR1
pHR'CMVLacZ
pHR'CMVGFP
pHR'RSVGFP
pHR'MSCVGFP
HIV/MSCVHybrid
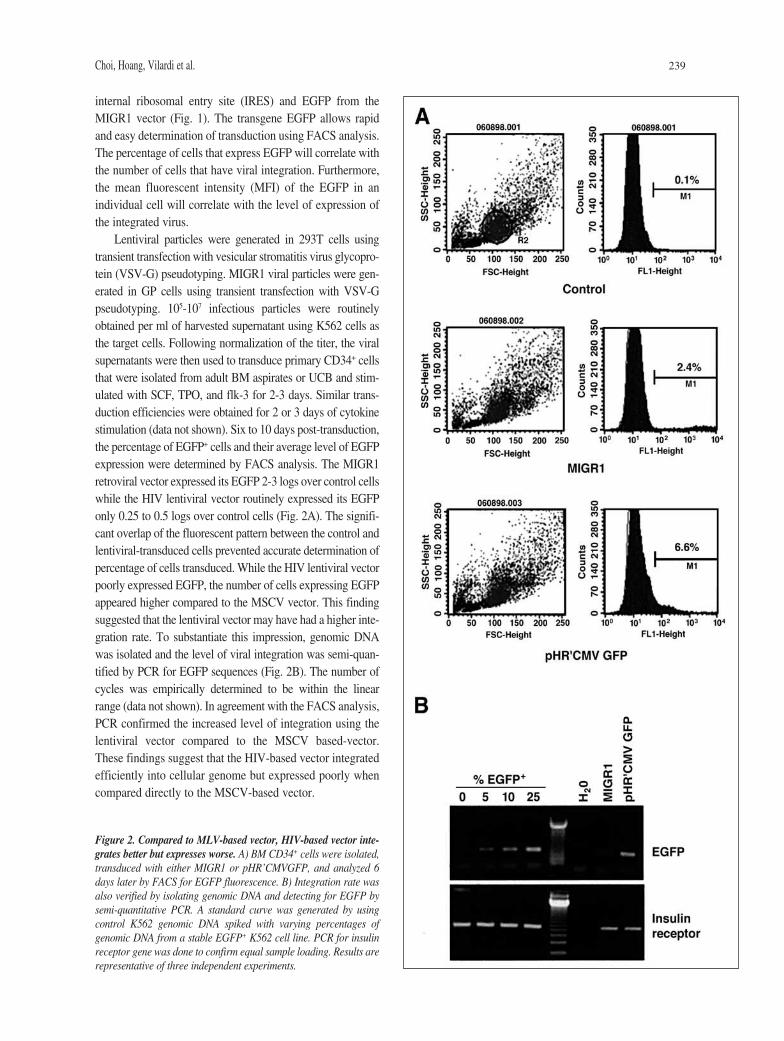
Choi, Hoang, Vilardi et al. 239
internal ribosomal entry site (IRES) and EGFP from theMIGR1 vector (Fig. 1). The transgene EGFP allows rapidand easy determination of transduction using FACS analysis.The percentage of cells that express EGFP will correlate withthe number of cells that have viral integration. Furthermore,the mean fluorescent intensity (MFI) of the EGFP in anindividual cell will correlate with the level of expression ofthe integrated virus.
Lentiviral particles were generated in 293T cells usingtransient transfection with vesicular stromatitis virus glycopro-tein (VSV-G) pseudotyping. MIGR1 viral particles were gen-erated in GP cells using transient transfection with VSV-Gpseudotyping. 105-107 infectious particles were routinelyobtained per ml of harvested supernatant using K562 cells asthe target cells. Following normalization of the titer, the viralsupernatants were then used to transduce primary CD34+ cellsthat were isolated from adult BM aspirates or UCB and stim-ulated with SCF, TPO, and flk-3 for 2-3 days. Similar trans-duction efficiencies were obtained for 2 or 3 days of cytokinestimulation (data not shown). Six to 10 days post-transduction,the percentage of EGFP+ cells and their average level of EGFPexpression were determined by FACS analysis. The MIGR1retroviral vector expressed its EGFP 2-3 logs over control cellswhile the HIV lentiviral vector routinely expressed its EGFPonly 0.25 to 0.5 logs over control cells (Fig. 2A). The signifi-cant overlap of the fluorescent pattern between the control andlentiviral-transduced cells prevented accurate determination ofpercentage of cells transduced. While the HIV lentiviral vectorpoorly expressed EGFP, the number of cells expressing EGFPappeared higher compared to the MSCV vector. This findingsuggested that the lentiviral vector may have had a higher inte-gration rate. To substantiate this impression, genomic DNAwas isolated and the level of viral integration was semi-quan-tified by PCR for EGFP sequences (Fig. 2B). The number ofcycles was empirically determined to be within the linearrange (data not shown). In agreement with the FACS analysis,PCR confirmed the increased level of integration using thelentiviral vector compared to the MSCV based-vector.These findings suggest that the HIV-based vector integratedefficiently into cellular genome but expressed poorly whencompared directly to the MSCV-based vector.
Figure 2. Compared to MLV-based vector, HIV-based vector inte-grates better but expresses worse. A) BM CD34+ cells were isolated,transduced with either MIGR1 or pHR’CMVGFP, and analyzed 6days later by FACS for EGFP fluorescence. B) Integration rate wasalso verified by isolating genomic DNA and detecting for EGFP bysemi-quantitative PCR. A standard curve was generated by usingcontrol K562 genomic DNA spiked with varying percentages ofgenomic DNA from a stable EGFP+ K562 cell line. PCR for insulinreceptor gene was done to confirm equal sample loading. Results arerepresentative of three independent experiments.
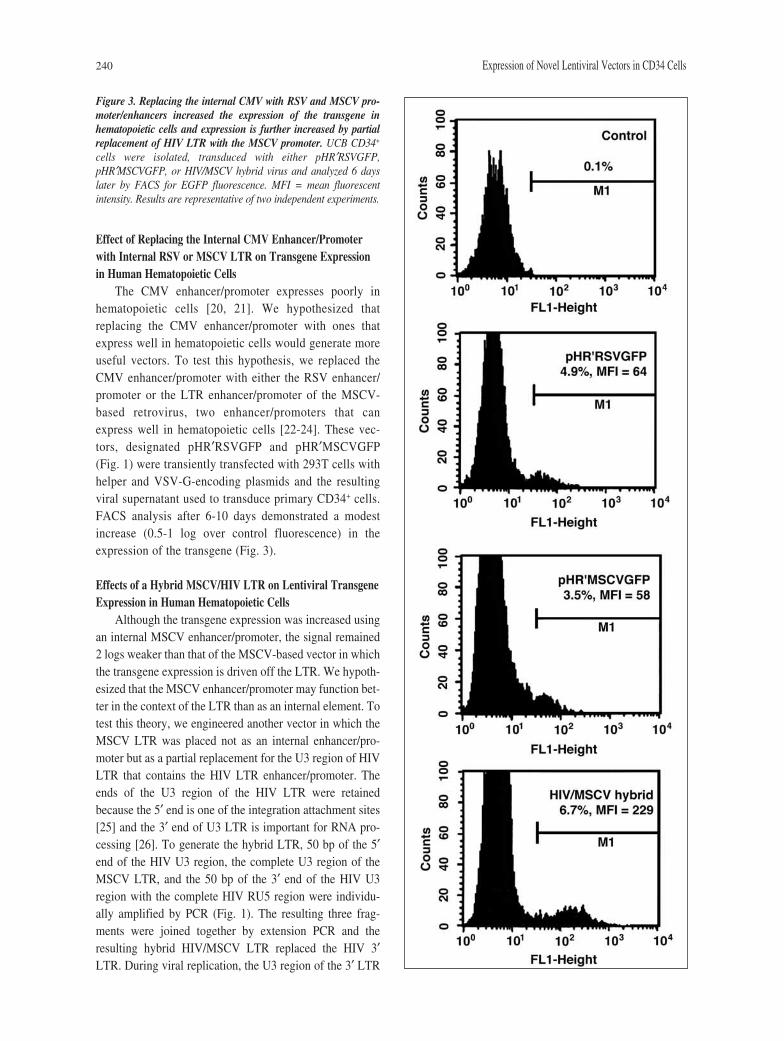
240 Expression of Novel Lentiviral Vectors in CD34 Cells
Effect of Replacing the Internal CMV Enhancer/Promoterwith Internal RSV or MSCV LTR on Transgene Expressionin Human Hematopoietic Cells
The CMV enhancer/promoter expresses poorly inhematopoietic cells [20, 21]. We hypothesized thatreplacing the CMV enhancer/promoter with ones thatexpress well in hematopoietic cells would generate moreuseful vectors. To test this hypothesis, we replaced theCMV enhancer/promoter with either the RSV enhancer/promoter or the LTR enhancer/promoter of the MSCV-based retrovirus, two enhancer/promoters that canexpress well in hematopoietic cells [22-24]. These vec-tors, designated pHR′RSVGFP and pHR′MSCVGFP(Fig. 1) were transiently transfected with 293T cells withhelper and VSV-G-encoding plasmids and the resultingviral supernatant used to transduce primary CD34+ cells.FACS analysis after 6-10 days demonstrated a modestincrease (0.5-1 log over control fluorescence) in theexpression of the transgene (Fig. 3).
Effects of a Hybrid MSCV/HIV LTR on Lentiviral TransgeneExpression in Human Hematopoietic Cells
Although the transgene expression was increased usingan internal MSCV enhancer/promoter, the signal remained2 logs weaker than that of the MSCV-based vector in whichthe transgene expression is driven off the LTR. We hypoth-esized that the MSCV enhancer/promoter may function bet-ter in the context of the LTR than as an internal element. Totest this theory, we engineered another vector in which theMSCV LTR was placed not as an internal enhancer/pro-moter but as a partial replacement for the U3 region of HIVLTR that contains the HIV LTR enhancer/promoter. Theends of the U3 region of the HIV LTR were retainedbecause the 5′ end is one of the integration attachment sites[25] and the 3′ end of U3 LTR is important for RNA pro-cessing [26]. To generate the hybrid LTR, 50 bp of the 5′end of the HIV U3 region, the complete U3 region of theMSCV LTR, and the 50 bp of the 3′ end of the HIV U3region with the complete HIV RU5 region were individu-ally amplified by PCR (Fig. 1). The resulting three frag-ments were joined together by extension PCR and theresulting hybrid HIV/MSCV LTR replaced the HIV 3′LTR. During viral replication, the U3 region of the 3′ LTR
Figure 3. Replacing the internal CMV with RSV and MSCV pro-moter/enhancers increased the expression of the transgene inhematopoietic cells and expression is further increased by partialreplacement of HIV LTR with the MSCV promoter. UCB CD34+
cells were isolated, transduced with either pHR′RSVGFP,pHR′MSCVGFP, or HIV/MSCV hybrid virus and analyzed 6 dayslater by FACS for EGFP fluorescence. MFI = mean fluorescentintensity. Results are representative of two independent experiments.
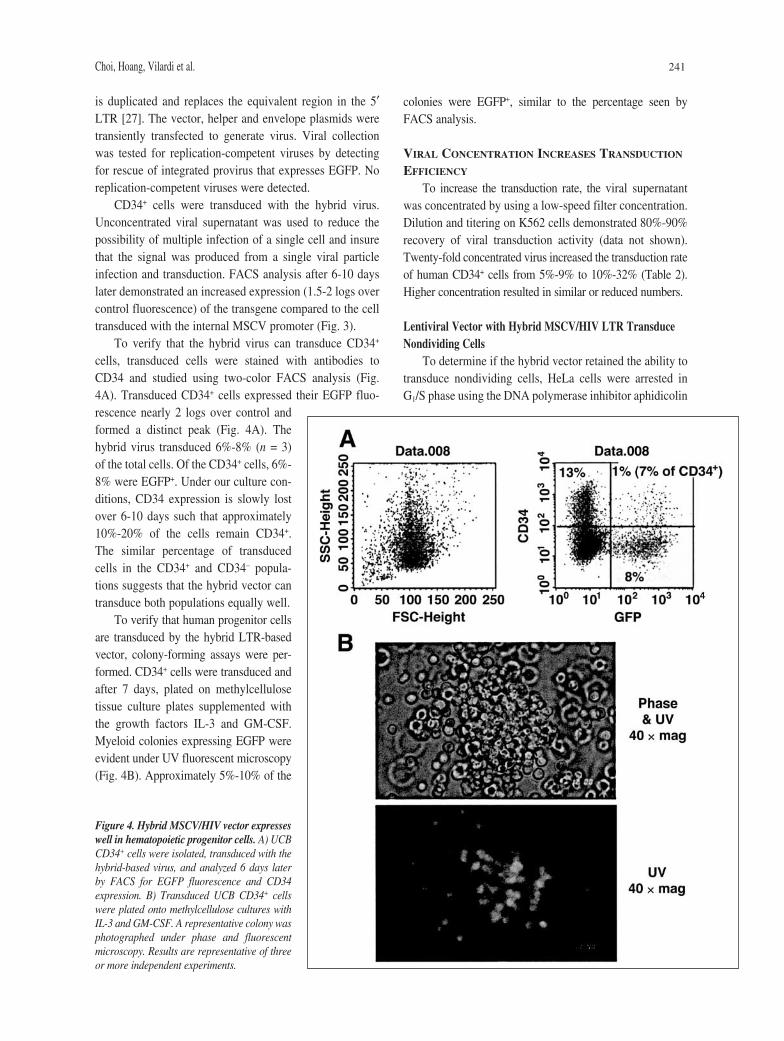
Choi, Hoang, Vilardi et al. 241
is duplicated and replaces the equivalent region in the 5′LTR [27]. The vector, helper and envelope plasmids weretransiently transfected to generate virus. Viral collectionwas tested for replication-competent viruses by detectingfor rescue of integrated provirus that expresses EGFP. Noreplication-competent viruses were detected.
CD34+ cells were transduced with the hybrid virus.Unconcentrated viral supernatant was used to reduce thepossibility of multiple infection of a single cell and insurethat the signal was produced from a single viral particleinfection and transduction. FACS analysis after 6-10 dayslater demonstrated an increased expression (1.5-2 logs overcontrol fluorescence) of the transgene compared to the celltransduced with the internal MSCV promoter (Fig. 3).
To verify that the hybrid virus can transduce CD34+
cells, transduced cells were stained with antibodies toCD34 and studied using two-color FACS analysis (Fig.4A). Transduced CD34+ cells expressed their EGFP fluo-rescence nearly 2 logs over control andformed a distinct peak (Fig. 4A). Thehybrid virus transduced 6%-8% (n = 3)of the total cells. Of the CD34+ cells, 6%-8% were EGFP+. Under our culture con-ditions, CD34 expression is slowly lostover 6-10 days such that approximately10%-20% of the cells remain CD34+.The similar percentage of transducedcells in the CD34+ and CD34– popula-tions suggests that the hybrid vector cantransduce both populations equally well.
To verify that human progenitor cellsare transduced by the hybrid LTR-basedvector, colony-forming assays were per-formed. CD34+ cells were transduced andafter 7 days, plated on methylcellulosetissue culture plates supplemented withthe growth factors IL-3 and GM-CSF.Myeloid colonies expressing EGFP wereevident under UV fluorescent microscopy(Fig. 4B). Approximately 5%-10% of the
colonies were EGFP+, similar to the percentage seen byFACS analysis.
VIRAL CONCENTRATION INCREASES TRANSDUCTION
EFFICIENCY
To increase the transduction rate, the viral supernatantwas concentrated by using a low-speed filter concentration.Dilution and titering on K562 cells demonstrated 80%-90%recovery of viral transduction activity (data not shown).Twenty-fold concentrated virus increased the transduction rateof human CD34+ cells from 5%-9% to 10%-32% (Table 2).Higher concentration resulted in similar or reduced numbers.
Lentiviral Vector with Hybrid MSCV/HIV LTR TransduceNondividing Cells
To determine if the hybrid vector retained the ability totransduce nondividing cells, HeLa cells were arrested inG1/S phase using the DNA polymerase inhibitor aphidicolin
Figure 4. Hybrid MSCV/HIV vector expresseswell in hematopoietic progenitor cells. A) UCBCD34+ cells were isolated, transduced with thehybrid-based virus, and analyzed 6 days laterby FACS for EGFP fluorescence and CD34expression. B) Transduced UCB CD34+ cellswere plated onto methylcellulose cultures withIL-3 and GM-CSF. A representative colony wasphotographed under phase and fluorescentmicroscopy. Results are representative of threeor more independent experiments.
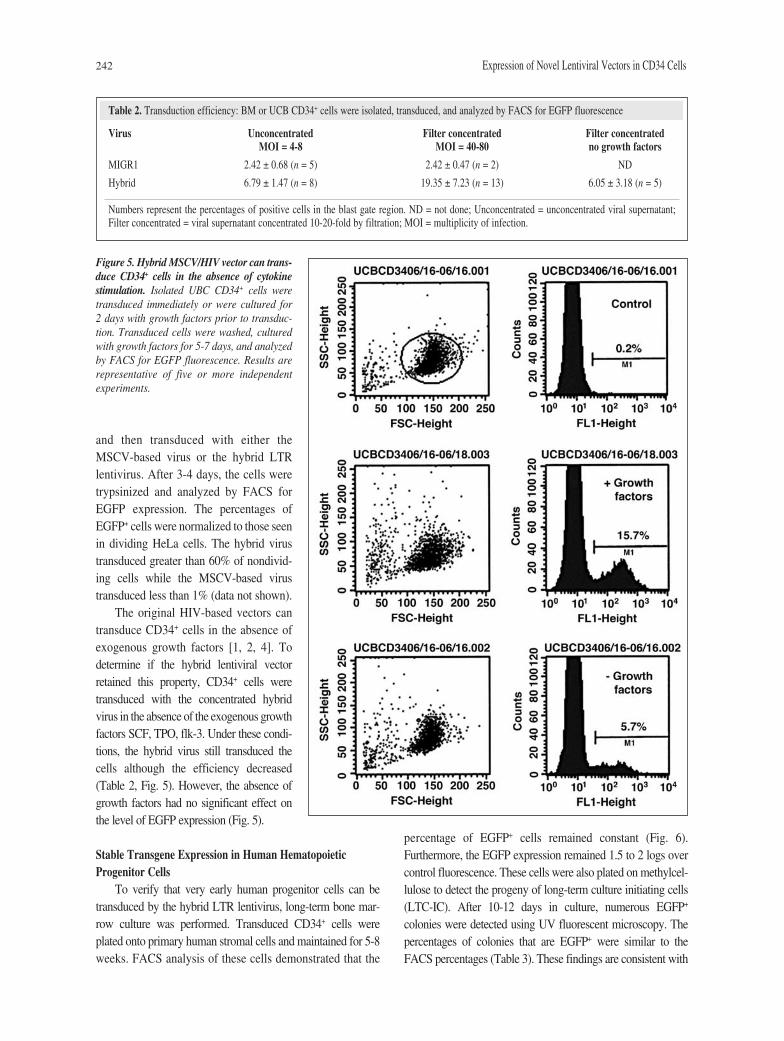
242 Expression of Novel Lentiviral Vectors in CD34 Cells
and then transduced with either theMSCV-based virus or the hybrid LTRlentivirus. After 3-4 days, the cells weretrypsinized and analyzed by FACS forEGFP expression. The percentages ofEGFP+ cells were normalized to those seenin dividing HeLa cells. The hybrid virustransduced greater than 60% of nondivid-ing cells while the MSCV-based virustransduced less than 1% (data not shown).
The original HIV-based vectors cantransduce CD34+ cells in the absence ofexogenous growth factors [1, 2, 4]. Todetermine if the hybrid lentiviral vectorretained this property, CD34+ cells weretransduced with the concentrated hybridvirus in the absence of the exogenous growthfactors SCF, TPO, flk-3. Under these condi-tions, the hybrid virus still transduced thecells although the efficiency decreased(Table 2, Fig. 5). However, the absence ofgrowth factors had no significant effect onthe level of EGFP expression (Fig. 5).
Stable Transgene Expression in Human HematopoieticProgenitor Cells
To verify that very early human progenitor cells can betransduced by the hybrid LTR lentivirus, long-term bone mar-row culture was performed. Transduced CD34+ cells wereplated onto primary human stromal cells and maintained for 5-8weeks. FACS analysis of these cells demonstrated that the
percentage of EGFP+ cells remained constant (Fig. 6).Furthermore, the EGFP expression remained 1.5 to 2 logs overcontrol fluorescence. These cells were also plated on methylcel-lulose to detect the progeny of long-term culture initiating cells(LTC-IC). After 10-12 days in culture, numerous EGFP+
colonies were detected using UV fluorescent microscopy. Thepercentages of colonies that are EGFP+ were similar to theFACS percentages (Table 3). These findings are consistent with
Table 2. Transduction efficiency: BM or UCB CD34+ cells were isolated, transduced, and analyzed by FACS for EGFP fluorescence
Virus Unconcentrated Filter concentrated Filter concentratedMOI = 4-8 MOI = 40-80 no growth factors
MIGR1 2.42 ± 0.68 (n = 5) 2.42 ± 0.47 (n = 2) ND
Hybrid 6.79 ± 1.47 (n = 8) 19.35 ± 7.23 (n = 13) 6.05 ± 3.18 (n = 5)
Numbers represent the percentages of positive cells in the blast gate region. ND = not done; Unconcentrated = unconcentrated viral supernatant;Filter concentrated = viral supernatant concentrated 10-20-fold by filtration; MOI = multiplicity of infection.
Figure 5. Hybrid MSCV/HIV vector can trans-duce CD34+ cells in the absence of cytokinestimulation. Isolated UBC CD34+ cells weretransduced immediately or were cultured for 2 days with growth factors prior to transduc-tion. Transduced cells were washed, culturedwith growth factors for 5-7 days, and analyzedby FACS for EGFP fluorescence. Results arerepresentative of five or more independentexperiments.

Choi, Hoang, Vilardi et al. 243
transduction of very early hematopoietic precursor cells withno evidence of promoter shutdown of the hybrid LTR.
Stable Transgene Expression in NOD/SCID RepopulationAssay
To confirm transduction of very early progenitor cells,the hybrid vector was tested in a repopulation assay. CD34+
UCB cells that were stimulated with SCF, TPO, flt-3 for 2days, and then transduced by spinoculation. Analysis of thepre-injected CD34+ cells revealed that 20% were EGFP+
(data not shown). After infection, 2.0 × 106 UCB cells wereimmediately injected via tail vein into sublethally irradiated
male NOD/SCID mice. At week 5 post-injection, mice weresacrificed and the BM cells were isolated and analyzed byFACS for the presence of human CD45+ and EGFP+ cells. Ofthe engrafted human CD45+ cells, 20% were EGFP+ (Fig. 7)similar to the pre-injection levels. Furthermore, the EGFPexpression was 1.5 logs over control fluorescence. These find-ings are consistent with transduction and stable expression ofthe transgene in repopulating cells.
DISCUSSION
We have engineered a novel lentiviral vector with a hybridMSCV/HIV LTR that successfully weds the high transgeneexpression of the MLV-based vectors with the high trans-duction efficiency of lentiviral vectors. To our best knowl-edge, this is the first report that demonstrates that a
Figure 6. Hybrid-based vector maintains the number of cells and the level ofexpression in transduced CD34+ cells. UCB CD34+ cells were isolated and stim-ulated with SCF, TPO, and flt-3 for 2 days. The cells were transduced with thehybrid virus, cultured on irradiated human stromal cells for 5 weeks, and thenanalyzed by FACS for EGFP fluorescence. Results are representative of threeindependent experiments.
Table 3. Long-term transduction efficiency
EGFP+ EGFP+ EGFP+
after 1 week after 5 weeks colonies
10.16 ± 2.99% 10.63 ± 1.82% 8.60 ± 3.05%(n = 241 colonies)
CD34+ UCB cells were transduced with the hybrid virus, cultured onstromal cell layer, and analyzed by FACS for EGFP fluorescence at1 week and 5 weeks post-transduction. After 5 weeks, cells were alsoplated in methylcellulose and the colonies scored 10-15 days later forEGFP fluorescence. Results are summary of three independentexperiments.
Figure 7. Hybrid-based vector transduces NOD/SCID repopulatingcells. UCB CD34+ cells were isolated and stimulated with SCF, TPO,and flt-3 for 2 days. The cells were transduced with the hybrid virusand immediately injected into sublethally irradiated NOD/SCIDmice. At week 5, mice were sacrificed and the BM cells analyzed byFACS for human CD45+ and EGFP+ cells. % represents the percent-age of human CD45+ cells that are also EGFP+.

244 Expression of Novel Lentiviral Vectors in CD34 Cells
promoter can be functional when embedded in the HIVLTR as has been demonstrated for the oncoviral retrovirusvectors. In human CD34+ cells, the hybrid vector expressesits transgene 10- to 100-fold over the original HIV-basedvector. This high level of expression and the percentage ofEGFP+ cells were maintained over 8 weeks in LTC-ICassays and 5 weeks in NOD/SCID repopulation assay, indi-cating that the hybrid vector transduces very earlyhematopoietic precursors and that promoter shutdown doesnot occur. Thus, this vector is a valuable research tool for highand stable expression of transgenes in human hematopoieticprogenitor cells.
Similar to the original HIV-based vector, the hybrid vec-tor was able to transduce nondividing HeLa cells, an expectedfinding since all known HIV viral proteins that promote trans-duction of nondividing cells [7, 28-31] are present. Thehybrid vector also transduced human CD34+ hematopoieticcells with greater efficiency than the MSCV-based vectorwhen assayed under identical transduction conditions withnormalized viral titers. The transduction efficiencies were1%-4% and 10%-30% for the MLV-based and hybrid-basedviruses, respectively. The efficiency for the hybrid-basedvirus decreased to 2%-10% in the absence of exogenousgrowth factors. While these numbers are less than thosereported by others, the differences probably represent differ-ent experimental conditions [1, 2, 4]. For example, we did notuse fibronectin or multiple exposures to viral supernatant. Wetransduced the CD34+ cells only once 18-24 hours after isola-tion. In contrast, Evans et al. transduced CD34+ cells twicewithin 24 hours of isolation [1]. These experimental differ-ences and not the inherent differences in the viral vectors arethe probable explanation for the differences in the reportedtransduction efficiencies. Further supporting this premise arethe markedly different transduction efficiencies (10% versus54%) that are seen in unstimulated CD34+ cells transducedwith the same HIV-based vector [2, 4].
Although lentiviral vectors represent promising toolsfor gene therapy, numerous issues need to be addressedprior to clinical trial. Many of the current advances in vec-tor design deal with the biosafety issue and have led to thesecond [31] and third generations [32] of the HIV-basedvectors. The third generation vector has a deleted 3′ LTR(self-inactivating-vector) which in theory will decrease therisk of expression of oncogenes downstream of the viralinsertion site, although such an event has yet to be reportedin human studies using retroviral vectors with intact LTRs.
More recent advances in lentiviral vector design dealwith the poor expression of the transgene in human hemato-poietic cells. This has important implications in using lentivi-ral vectors as therapeutic and research tools. Both the first[18] and second generation vectors contain an internal CMV
promoter that does not express well in hematopoietic cells.The initial third generation vector [32] replaced the internalCMV promoter with the phosphoglycerate kinase gene(PGK) promoter but a recent study indicates that this vectordoes not express well in hematopoietic cells [33]. The addi-tion of a post-transcriptional regulatory element of wood-chuck hepatitis virus (Wpre) [34] increases the expression ofthe transgene to the same level as our hybrid vector [33, 35].Interestingly, Wpre decreases the expression of the transgeneoff the human elongation factor EF1α promoter, suggestingthat the effects of Wpre can depend on the promoter. Studiesare ongoing to determine if the Wpre can further increase theexpression of our hybrid vector.
The relatively weak activity of the PGK promoter in theabsence of the Wpre is similar to the modest increase inexpression seen in our vectors in which the internal CMVpromoter was replaced with the RSV or MSCV promoter.A modest increase in expression has been reported in Tcells using an HIV-based vector in which the CMV internalenhancer/promoter was replaced with an internal MLVLTR enhancer/promoter [21]. The increased expressionprobably indicates that the network of transcription factorsand coactivators in hematopoietic cells favors the expres-sion of the leukemia viral LTR over the immediate pro-moter of CMV, an expected finding considering thatleukemia virus is specialized to infect hematopoietic cellswhile CMV infects a wider variety of cell types.Understanding which of the many potential DNA elementsof the LTR contributes to the increased expression will leadto further improvement in the design of the enhancer/promoterfor higher expression in hematopoietic cells.
The expression of the transgene is further increased whenthe internal MSCV enhancer/promoter is moved to the LTR.Three possible mechanisms for the increased expression of thehybrid vector include: A) loss of promoter interference fromthe HIV LTR [21]; B) duplication of the MSCV enhancerleading to synergistic increases in the activity of the 5′ LTR,and C) utilization of the mRNA splicing sites that are 5′ to theinternal enhancer/promoter site—since splicing is linked tonuclear export, transcripts driven of the 5′ LTR will be splicedand more efficiently exported out of the nucleus. While thesemechanisms may contribute to the increase in expression,they do not completely explain the greater than 10-foldincrease. Promoter interference appears to decrease expres-sion by only two- to threefold [21]. Increasing the number ofcopies of the MSCV enhancers (2, 3, and 4 copies) at theinternal site did not increase the expression of the transgene(data not shown). Splicing of the HIV viral transcript is con-sidered inefficient, contributing to the nuclear entrapment ofthe transcript in the absence of the Rev protein [36]. Althoughthe actual mechanisms for the increased expression remain

Choi, Hoang, Vilardi et al. 245
REFERENCES
1 Evans JT, Kelly PF, O’Neill E et al. Human cord bloodCD34+CD38– cell transduction via lentivirus-based gene transfervectors. Hum Gene Ther 1999;10:1479-1489.
2 Case SS, Price MA, Jordan CT et al. Stable transduction ofquiescent CD34(+)CD38(-) human hematopoietic cells byHIV-1-based lentiviral vectors. Proc Natl Acad Sci USA1999;96:2988-2993.
3 Uchida N, Sutton RE, Friera AM et al. HIV, but not murineleukemia virus, vectors mediate high efficiency gene transferinto freshly isolated G0/G1 human hematopoietic stem cells.Proc Natl Acad Sci USA 1998;95:11939-11944.
4 Miyoshi H, Smith KA, Mosier DE et al. Transduction ofhuman CD34+ cells that mediate long-term engraftment ofNOD/SCID mice by HIV vectors. Science 1999;283:682-686.
5 Sutton RE, Wu HT, Rigg R et al. Human immunodeficiencyvirus type 1 vectors efficiently transduce human hematopoieticstem cells. J Virol 1998;72:5781-5788.
6 Naldini L, Blomer U, Gage FH et al. Efficient transfer, inte-gration, and sustained long-term expression of the transgenein adult rat brains injected with a lentiviral vector. Proc NatlAcad Sci USA 1996;93:11382-11388.
7 Kafri T, Blomer U, Peterson DA et al. Sustained expressionof genes delivered directly into liver and muscle by lentiviralvectors. Nat Genet 1997;17:314-317.
8 Chirivi RG, Taraboletti G, Bani MR et al. Human immuno-deficiency virus-1 (HIV-1)-Tat protein promotes migration ofacquired immunodeficiency syndrome-related lymphomacells and enhances their adhesion to endothelial cells. Blood1999;94:1747-1754.
9 Weiss JM, Nath A, Major EO et al. HIV-1 Tat induces monocytechemoattractant protein-1-mediated monocyte transmigrationacross a model of the human blood-brain barrier and up-regu-lates CCR5 expression on human monocytes. J Immunol1999;163:2953-2959.
10 Barillari G, Sgadari C, Palladino C et al. Inflammatory cytokinessynergize with the HIV-1 Tat protein to promote angiogenesisand Kaposi’s sarcoma via induction of basic fibroblast growthfactor and the alpha v beta 3 integrin. J Immunol 1999;163:1929-1935.
11 Demarchi F, Gutierrez MI, Giacca M. Human immunodefi-ciency virus type 1 tat protein activates transcription factorNF-kappaB through the cellular interferon-inducible, dou-ble-stranded RNA-dependent protein kinase, PKR. J Virol1999;73:7080-7086.
12 Maggirwar SB, Tong N, Ramirez S et al. HIV-1 Tat-mediatedactivation of glycogen synthase kinase-3beta contributes toTat-mediated neurotoxicity. J Neurochem 1999;73:578-586.
13 Lefevre EA, Krzysiek R, Loret EP et al. Cutting edge: HIV-1Tat protein differentially modulates the B cell response ofnaive, memory, and germinal center B cells. J Immunol1999;163:1119-1122.
14 Schwarze SR, Ho A, Vocero-Akbani A et al. In vivo proteintransduction: delivery of a biologically active protein into themouse. Science 1999;285:1569-1572.
15 Blomer U, Naldini L, Kafri T et al. Highly efficient and sus-tained gene transfer in adult neurons with a lentivirus vector.J Virol 1997;71:6641-6649.
16 Pear WS, Miller JP, Xu L et al. Efficient and rapid induction ofa chronic myelogenous leukemia-like myeloproliferative dis-ease in mice receiving P210 bcr/abl-transduced bone marrow.Blood 1998;92:3780-3792.
17 Kotani H, Newton PB 3rd, Zhang S et al. Improved methods ofretroviral vector transduction and production for gene therapy.Hum Gene Ther 1994;5:19-28.
18 Naldini L, Blomer U, Gallay P et al. In vivo gene delivery andstable transduction of nondividing cells by a lentiviral vector[see comments]. Science 1996;272:263-267.
19 Coulombel L, Eaves AC, Eaves CJ. Enzymatic treatment oflong-term human marrow cultures reveals the preferentiallocation of primitive hemopoietic progenitors in the adherentlayer. Blood 1983;62:291-297.
20 Baskar JF, Smith PP, Ciment GS et al. Developmental analy-sis of the cytomegalovirus enhancer in transgenic animals. J Virol 1996;70:3215-3226.
21 Kung SK, An DS, Chen IS. A murine leukemia virus (MuLV)long terminal repeat derived from rhesus macaques in the con-text of a lentivirus vector and MuLV gag sequence results inhigh-level gene expression in human T lymphocytes. J Virol2000;74:3668-3681.
22 Karlsson S, Humphries RK, Gluzman Y et al. Transfer of genesinto hematopoietic cells using recombinant DNA viruses. ProcNatl Acad Sci USA 1985;82:158-162.
23 Zarrin AA, Malkin L, Fong I et al. Comparison of CMV, RSV,SV40 viral and Vlambda1 cellular promoters in B and T lym-phoid and non-lymphoid cell lines. Biochim Biophys Acta1999;1446:135-139.
24 Conneally E, Bardy P, Eaves CJ et al. Rapid and efficientselection of human hematopoietic cells expressing murine
unidentified, our results predict that some promoters will bemore active when placed in the LTR versus at an internal site.This approach would be useful in designing lentiviral vectorsfor restricted expression in specific cell lineages, especially incases in which Wpre decreases the expression of a particularpromoter.
ACKNOWLEDGMENTS
This work was supported by grants from the NationalInstitutes of Health (CA75330) and the McCabe Fund,Philadelphia, PA.
A.M.G. is a Distinguished Clinical Scientist of the DorisDuke Charitable Foundation.

246 Expression of Novel Lentiviral Vectors in CD34 Cells
heat-stable antigen as an indicator of retroviral-mediated genetransfer. Blood 1996;87:456-464.
25 Masuda T, Kuroda MJ, Harada S. Specific and independentrecognition of U3 and U5 att sites by human immunodeficiencyvirus type 1 integrase in vivo. J Virol 1998;72:8396-8402.
26 Dougherty JP, Temin HM. A promoterless retroviral vectorindicates that there are sequences in U3 required for 3′ RNAprocessing. Proc Natl Acad Sci USA 1987;84:1197-1201.
27 Yu SF, von Ruden T, Kantoff PW et al. Self-inactivating retro-viral vectors designed for transfer of whole genes into mam-malian cells. Proc Natl Acad Sci USA 1986;83:3194-3198.
28 Bukrinsky MI, Haggerty S, Dempsey MP et al. A nuclearlocalization signal within HIV-1 matrix protein that governsinfection of non-dividing cells. Nature 1993;365:666-669.
29 Gallay P, Swingler S, Song J et al. HIV nuclear import is gov-erned by the phosphotyrosine-mediated binding of matrix tothe core domain of integrase. Cell 1995;83:569-576.
30 Connor RI, Chen BK, Choe S et al. Vpr is required for effi-cient replication of human immunodeficiency virus type-1 inmononuclear phagocytes. Virology 1995;206:935-944.
31 Zufferey R, Nagy D, Mandel RJ et al. Multiply attenuatedlentiviral vector achieves efficient gene delivery in vivo. NatBiotechnol 1997;15:871-875.
32 Dull T, Zufferey R, Kelly M et al. A third-generation lenti-virus vector with a conditional packaging system. J Virol1998;72:8463-8471.
33 Salmon P, Kindler V, Ducrey O et al. High-level transgeneexpression in human hematopoietic progenitors and differenti-ated blood lineages after transduction with improved lentiviralvectors. Blood 2000;96:3392-3398.
34 Zufferey R, Donello JE, Trono D et al. Woodchuck hepatitisvirus posttranscriptional regulatory element enhances expres-sion of transgenes delivered by retroviral vectors. J Virol1999;73:2886-2892.
35 Follenzi A, Ailles LE, Bakovic S et al. Gene transfer by lentivi-ral vectors is limited by nuclear translocation and rescued byHIV-1 pol sequences. Nat Genet 2000;25:217-222.
36 Olsen HS, Cochrane AW, Rosen C. Interaction of cellularfactors with intragenic cis-acting repressive sequences withinthe HIV genome. Virology 1992;191:709-715.




















