
Tmt1: the first LTR-retrotransposon from a Tuber spp
12
Curr Genet (2008) 53:23–34 DOI 10.1007/s00294-007-0155-9 123 RESEARCH ARTICLE Tmt1: the Wrst LTR-retrotransposon from a Tuber spp. Claudia Riccioni · Andrea Rubini · Beatrice BelWori · Valentina Passeri · Francesco Paolocci · Sergio Arcioni Received: 16 July 2007 / Revised: 3 September 2007 / Accepted: 13 September 2007 / Published online: 31 October 2007 © Springer-Verlag 2007 Abstract Retrotransposons are suitable targets for devel- oping molecular markers for population genetics studies. Transposable elements have not yet been isolated from ectomycorrhizal fungi of the genus Tuber. In this paper, we report on the isolation and characterization of Tmt1, an LTR-retrotransposon from Tuber melanosporum. The Tmt1 sequence shows relatedness to Ty3/gypsy retrotransposons from which it diVerentiates for the presence of a dUTPase extra-domain between protease and reverse transcriptase. Phylogenetic analyses suggest a horizontal transfer of the dUTPase gene (dut) from a fungal host genome. The pres- ence of non-identical LTRs and degenerate ORFs substanti- ate an ancient integration of Tmt1 in T. melanosporum genome. Furthermore, transcripts analyses proved an inac- tive status of Tmt1, whereas Southern analysis showed that Tmt1 is a repetitive T. melanosporum species-speciWc ele- ment. Tmt1-based markers will help us to gain more insights into population biology in this fungal species. Keywords Tuber melanosporum · Ascomycete · Ectomycorrhizas · LTR-retrotransposon · dUTPase Introduction Transposable elements (TEs) are discrete segments of DNA that are able to move and replicate within their host genomes. TEs have been found to be ubiquitous in most living organisms, and to represent a major contribution of non-coding, repetitive DNA to eukaryotic genome sizes (Kidwell and Lisch 1997; Kidwell 2002). TEs act as agents of genome restructuring and mutagenesis by promoting a wide range of mutations, from modiWcation in the size and arrangement of whole genomes, to substitutions, deletions and insertions of even single nucleotides. TEs are valuable genetic tools: for example they are widely used for gene tagging in forward genetics (Daboussi and Capy 2003) and, by analyzing their chromosomal distribution and/or by developing diVerent types of transposon-based molecular markers, for population genetics studies (Kumar and Hirochika 2001; Daboussi and Capy 2003; Schulman et al. 2004). Eukaryotic transposons are grouped into two main clas- ses, according to the mechanism of transposition (Kempken and Kück 1998). Class I elements transpose via the reverse transcription of RNA intermediates into full-length DNA copies, followed by the integration of the DNA into new sites within the host genomes. By contrast, most Class II elements transpose via DNA excision and reintegration (Flavell 1999; Goodwin et al. 2003). Class I elements, are divided into two subclasses, the LTR (long terminal repeat)-retrotransposons and the non-LTR retrotranspo- sons (LINEs and SINEs—long and short interspersed nuclear elements, respectively). Based on the comparison of their reverse transcriptase (RT) sequences, LTR-retro- transposons are classiWed in Wve distinct groups: Ty3/ gypsy, Ty1/copia, BEL, DIRS1 and vertebrate retroviruses (Goodwin and Poulter 2001). Electronic supplementary material The online version of this article (doi:10.1007/s00294-007-0155-9) contains supplementary material, which is available to authorized users. Communicated by U. Kück. C. Riccioni · A. Rubini · B. BelWori · V. Passeri · F. Paolocci · S. Arcioni (&) National Research Council, Plant Genetics Institute-Perugia, Perugia Division, Via Madonna Alta 130, 06128 Perugia, Italy e-mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Tmt1: the first LTR-retrotransposon from a Tuber spp

Curr Genet (2008) 53:23–34
DOI 10.1007/s00294-007-0155-9RESEARCH ARTICLE
Tmt1: the Wrst LTR-retrotransposon from a Tuber spp.
Claudia Riccioni · Andrea Rubini · Beatrice BelWori · Valentina Passeri · Francesco Paolocci · Sergio Arcioni
Received: 16 July 2007 / Revised: 3 September 2007 / Accepted: 13 September 2007 / Published online: 31 October 2007© Springer-Verlag 2007
Abstract Retrotransposons are suitable targets for devel-oping molecular markers for population genetics studies.Transposable elements have not yet been isolated fromectomycorrhizal fungi of the genus Tuber. In this paper, wereport on the isolation and characterization of Tmt1, anLTR-retrotransposon from Tuber melanosporum. The Tmt1sequence shows relatedness to Ty3/gypsy retrotransposonsfrom which it diVerentiates for the presence of a dUTPaseextra-domain between protease and reverse transcriptase.Phylogenetic analyses suggest a horizontal transfer of thedUTPase gene (dut) from a fungal host genome. The pres-ence of non-identical LTRs and degenerate ORFs substanti-ate an ancient integration of Tmt1 in T. melanosporumgenome. Furthermore, transcripts analyses proved an inac-tive status of Tmt1, whereas Southern analysis showed thatTmt1 is a repetitive T. melanosporum species-speciWc ele-ment. Tmt1-based markers will help us to gain moreinsights into population biology in this fungal species.
Keywords Tuber melanosporum · Ascomycete · Ectomycorrhizas · LTR-retrotransposon · dUTPase
Introduction
Transposable elements (TEs) are discrete segments of DNAthat are able to move and replicate within their hostgenomes. TEs have been found to be ubiquitous in mostliving organisms, and to represent a major contribution ofnon-coding, repetitive DNA to eukaryotic genome sizes(Kidwell and Lisch 1997; Kidwell 2002). TEs act as agentsof genome restructuring and mutagenesis by promoting awide range of mutations, from modiWcation in the size andarrangement of whole genomes, to substitutions, deletionsand insertions of even single nucleotides. TEs are valuablegenetic tools: for example they are widely used for genetagging in forward genetics (Daboussi and Capy 2003) and,by analyzing their chromosomal distribution and/or bydeveloping diVerent types of transposon-based molecularmarkers, for population genetics studies (Kumar andHirochika 2001; Daboussi and Capy 2003; Schulman et al.2004).
Eukaryotic transposons are grouped into two main clas-ses, according to the mechanism of transposition (Kempkenand Kück 1998). Class I elements transpose via the reversetranscription of RNA intermediates into full-length DNAcopies, followed by the integration of the DNA into newsites within the host genomes. By contrast, most Class IIelements transpose via DNA excision and reintegration(Flavell 1999; Goodwin et al. 2003). Class I elements, aredivided into two subclasses, the LTR (long terminalrepeat)-retrotransposons and the non-LTR retrotranspo-sons (LINEs and SINEs—long and short interspersednuclear elements, respectively). Based on the comparisonof their reverse transcriptase (RT) sequences, LTR-retro-transposons are classiWed in Wve distinct groups: Ty3/gypsy, Ty1/copia, BEL, DIRS1 and vertebrate retroviruses(Goodwin and Poulter 2001).
Electronic supplementary material The online version of this article (doi:10.1007/s00294-007-0155-9) contains supplementary material, which is available to authorized users.
Communicated by U. Kück.
C. Riccioni · A. Rubini · B. BelWori · V. Passeri · F. Paolocci · S. Arcioni (&)National Research Council, Plant Genetics Institute-Perugia, Perugia Division, Via Madonna Alta 130, 06128 Perugia, Italye-mail: [email protected]
123

24 Curr Genet (2008) 53:23–34
Both autonomous and non-autonomous members arefound in many diVerent families of both Class I and IItransposons. Whereas an autonomous element encodes acomplete set of enzymes characteristic of its family, a non-autonomous element encodes none, or only some of them,and depends on enzymes encoded by its autonomous rela-tive for transposition.
In fungi, the horizontal gene transfer by non-infectiveselWsh genetic elements such as transposons, plasmids andintrons has been suggested as a possible mechanismaccounting for the multiple emergence of and adaptationsto parasitic and symbiotic growth (Tunlid and Talbot2002). In ectomycorrhizal basidiomycetes the occurrenceof retroelements has been reported (Murata and Yamada2000), whereas no TEs were so far isolated from ectomy-corrhizal ascomycetes.
Tuber melanosporum Vittad. is an ectomycorrhizal asco-mycete fungus producing hypogeous, black fruit bodiesappreciated worldwide for their unique taste and aroma. T.melanosporum, also known as “tartufo nero pregiato di Nor-cia e Spoleto” or “Perigorde truZe” is patchily distributed inItaly, France and Spain. Since the worldwide demand forthis species greatly exceeds its availability, artiWcial truZeplantations have been set, by outplanting nursery producedmycorrhizal plants, not only in Southern Europe where thisspecies is endemic. However, many aspects concerning theecological-agronomic requirements of any given truZe spe-cies have still to be fully clariWed. The diYculties of grow-ing Tuber spp. under experimental conditions along with thelack of phylogenetically closed model species have been infact the main obstacles in assessing crucial issues concern-ing their biology. Despite this, molecular markers haverecently provided helpful hints to gain insights into the lifecycle and reproductive system of Tuber spp. (Rubini et al.2005, 2007; Paolocci et al. 2006). In T. melanosporummolecular markers as microsatellites, random ampliWedpolymorphic DNA (RAPD) and internal transcribedregion—single nucleotide polymorphisms (ITS-SNP), havedetected low levels of genetic diversity (Bertault et al. 1998,2001; Paolocci et al. 2000; Murat et al. 2004). Thus, it hasbeen claimed that this species experienced a population bot-tleneck during the last glaciation which should havedepressed its genetic variability, population size and distri-bution (Bertault et al. 1998, 2001).
With the aim of characterizing as many informative mul-tilocus markers as possible we have undertaken researchwork to isolate transposable elements from Tuber spp. Inthis paper, we report on the isolation and characterizationof Tmt1 (T. melanosporum transposon 1), the Wrst LTR-ret-rotransposon-like element from a Tuber spp. Tmt1 is a T.melanosporum species-speciWc sequence showing peculiarfeatures and it may provide mycologists with an additionaltool to gain insights into truZe biodiversity and phylogeny.
Materials and methods
Sample source and DNA isolation
Ascocarps of T. melanosporum, T. brumale Vittad., T. indi-cum Cooke et Massee, T. aestivum Vittad., T. magnatumPico, T. borchii Vittad., T. macrosporum Vittad. and T.rufum Pico, were used in this study. Each ascocarp was Wrstexamined for macroscopic and microscopic features forspecies determination (Ceruti 1960; Montecchi and Lazzari1993). Small pieces of the gleba of each sample were storedat ¡70°C into sterile bags for molecular analyses. GenomicDNA was isolated from each ascocarp and puriWed by aCsCl gradient step as previously described (Paolocci et al.1995). Molecular analyses with ITS species-speciWc anduniversal ITS1/ITS4 primer pairs (Paolocci et al. 1997;Amicucci et al. 1998; Rubini et al. 1998, 2001) were alsoperformed to conWrm the specimen identity.
Isolation and characterization of T. melanosporum TEs
Genomic DNA isolated from a T. melanosporum ascocarp(me312 collected in Umbria, Italy) was used for the isola-tion of retrotransposon fragments, basically following thebiotin–streptavidin enrichment procedure reported byPearce et al. (1999). In brief, genomic DNA was restrictedwith EcoRI for 2 h and ligated with the relative EcoRIadapter (Vos et al. 1995). The ligated template was puriWedwith JETquick PCR Product PuriWcation Spin Kit (Gen-omed) and PCR-ampliWed with biotinylated RNaseHprimer 1 (Pearce et al. 1999) and EcoRI adapter primer.The PCR product was puriWed as above. Biotinylated prod-ucts were recovered using Streptavidin MagneSphere®
Paramagnetic Particles (Promega) and ampliWed withRnaseH primer 2 (Pearce et al. 1999) and EcoRI adapterprimer. The resulting amplicons were size fractionated byagarose gel electrophoresis. Single DNA bands in the 200–900 bp size range were recovered from gel using the JET-quick Gel Extraction Spin kit (Genomed), submitted to asecond round of PCR, cloned into pGEM®-T Easy Vector(Promega) and sequenced using the Big Dye Terminator kitV 3.1 and an ABI Prism 310 Genetic Analyzer (AppliedBiosystems).
The extension of a selected PCR amplicon (see Results)towards the 5� and 3� ends was obtained by Genome Walk-ing (GW; Siebert et al. 1995) using the Universal Genome-Walker™ Kit (Clontech) according to the supplier’sinstructions. At this purpose, DNA from the T. melanospo-rum ascocarp me312 was used to create the “libraries” ofadaptor-ligated fragments. Multiple steps of GW procedurewere performed using either AP1 or AP2 adapter primersprovided with the kit, coupled with the speciWc primersreported in Fig. 1a and Table 1. The resulting amplicons of
123
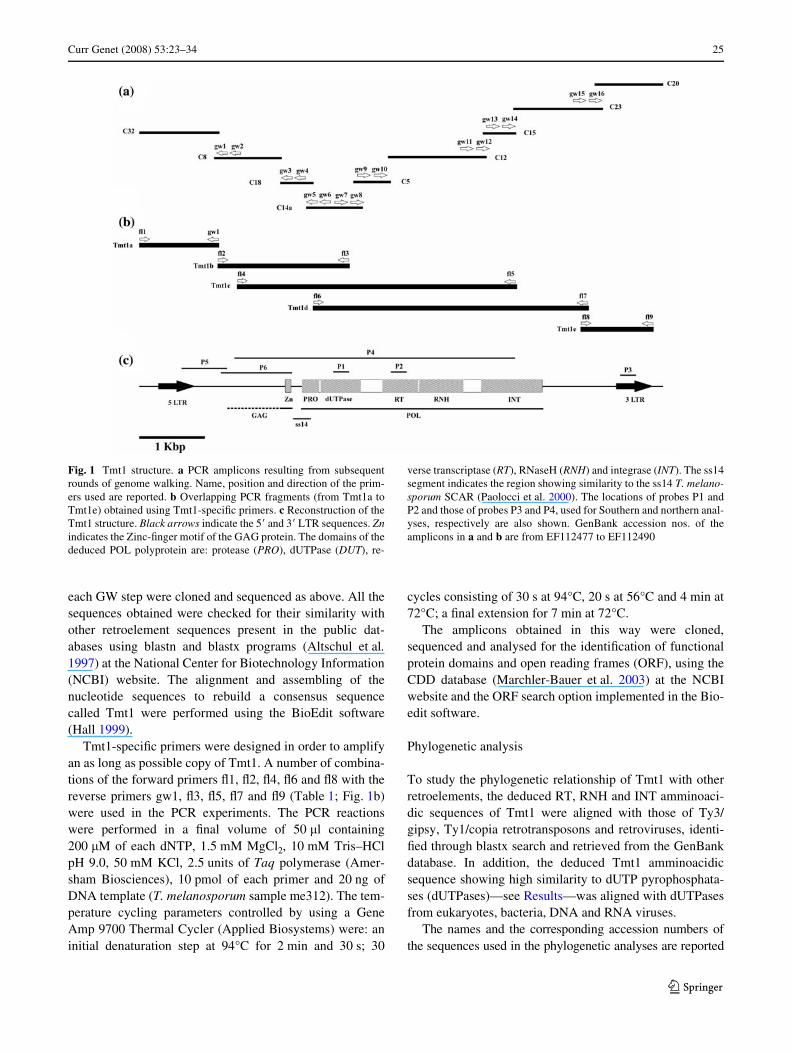
Curr Genet (2008) 53:23–34 25
each GW step were cloned and sequenced as above. All thesequences obtained were checked for their similarity withother retroelement sequences present in the public dat-abases using blastn and blastx programs (Altschul et al.1997) at the National Center for Biotechnology Information(NCBI) website. The alignment and assembling of thenucleotide sequences to rebuild a consensus sequencecalled Tmt1 were performed using the BioEdit software(Hall 1999).
Tmt1-speciWc primers were designed in order to amplifyan as long as possible copy of Tmt1. A number of combina-tions of the forward primers X1, X2, X4, X6 and X8 with thereverse primers gw1, X3, X5, X7 and X9 (Table 1; Fig. 1b)were used in the PCR experiments. The PCR reactionswere performed in a Wnal volume of 50 �l containing200 �M of each dNTP, 1.5 mM MgCl2, 10 mM Tris–HClpH 9.0, 50 mM KCl, 2.5 units of Taq polymerase (Amer-sham Biosciences), 10 pmol of each primer and 20 ng ofDNA template (T. melanosporum sample me312). The tem-perature cycling parameters controlled by using a GeneAmp 9700 Thermal Cycler (Applied Biosystems) were: aninitial denaturation step at 94°C for 2 min and 30 s; 30
cycles consisting of 30 s at 94°C, 20 s at 56°C and 4 min at72°C; a Wnal extension for 7 min at 72°C.
The amplicons obtained in this way were cloned,sequenced and analysed for the identiWcation of functionalprotein domains and open reading frames (ORF), using theCDD database (Marchler-Bauer et al. 2003) at the NCBIwebsite and the ORF search option implemented in the Bio-edit software.
Phylogenetic analysis
To study the phylogenetic relationship of Tmt1 with otherretroelements, the deduced RT, RNH and INT amminoaci-dic sequences of Tmt1 were aligned with those of Ty3/gipsy, Ty1/copia retrotransposons and retroviruses, identi-Wed through blastx search and retrieved from the GenBankdatabase. In addition, the deduced Tmt1 amminoacidicsequence showing high similarity to dUTP pyrophosphata-ses (dUTPases)—see Results—was aligned with dUTPasesfrom eukaryotes, bacteria, DNA and RNA viruses.
The names and the corresponding accession numbers ofthe sequences used in the phylogenetic analyses are reported
Fig. 1 Tmt1 structure. a PCR amplicons resulting from subsequentrounds of genome walking. Name, position and direction of the prim-ers used are reported. b Overlapping PCR fragments (from Tmt1a toTmt1e) obtained using Tmt1-speciWc primers. c Reconstruction of theTmt1 structure. Black arrows indicate the 5� and 3� LTR sequences. Znindicates the Zinc-Wnger motif of the GAG protein. The domains of thededuced POL polyprotein are: protease (PRO), dUTPase (DUT), re-
verse transcriptase (RT), RNaseH (RNH) and integrase (INT). The ss14segment indicates the region showing similarity to the ss14 T. melano-sporum SCAR (Paolocci et al. 2000). The locations of probes P1 andP2 and those of probes P3 and P4, used for Southern and northern anal-yses, respectively are also shown. GenBank accession nos. of theamplicons in a and b are from EF112477 to EF112490
123

26 Curr Genet (2008) 53:23–34
in Fig. 4a and b. Multiple sequence alignments were per-formed with ClustalX (Thompson et al. 1997) following aminor manual adjustment of the gaps using Bioedit software.Only the best conserved residues were retained and used forthe phylogenetic analyses. The alignments are available inFASTA format as supplementary Wles (electronic supplem-entry materials of SF1 and SF2). Phylogenetic analyses wereperformed with the Neighbor-joining method using the soft-ware Mega version 3.1 (Kumar et al. 2004).
Southern analyses
About 2 �g of genomic DNA from 8 Tuber species: T.melanosporum (6 samples), T. brumale (3), T. indicum(2), T. aestivum (2), T. magnatum (1), T. borchii (2), T.macrosporum (1) and T. rufum (1) were restricted over-night with HindIII (New England Biolabs), electrophore-sed in 1% agarose gel in the presence of a size-standardGeneRuler DNA Ladder-mix (Fermentas) and blotted
Table 1 Primers used in this study
N. Name Direction Sequence(5�–3�)
1 gw1 r GAGGGTCCTGATGTGACAGGATCTAGG
2 gw2 r TGGGGTTACTGTGCTTGCTGCTCTCGA
3 gw3 r TATCCGATGCCCTGCTTTTCCACAC
4 gw4 r CCATTATTTCCAGTAGCTCTGTTTCCC
5 gw5 r GCTGTCAATGTCCATAGGTACTGTTG
6 gw6 r CTATCTCATGGTTCTTTAGATCTGCC
7 gw7 f GGACTACCAGGTCCAGAAAGGCGATC
8 gw8 f GGAAACAAGGGATTTGGAAGTATGGAC
9 gw9 f GAGACTCTTTCGCGTCAGGAGGATC
10 gw10 f AGAAAGATGGAAAAATGAGACTATGCG
11 gw11 f CCGATATCTGAACGGCAGGACACGGAA
12 gw12 f GACACGGAAGAACGGCTAAAACCTCAG
13 gw13 f GAACACTATTCACATCGGGACTATGGAAC
14 gw14 f GTTCCACCCACAAACAGATGGACACAG
15 gw15 f AAAACTCCTTGGCAGAGACGGAACC
16 gw16 f ATGTCAGGAGCCTACACAGGCAGAT
17 X1 f CTGTAAGGAGAATATTCTGGT
18 X2 r GCCCTAGATCCTGTCACAT
19 X3 r GCCTTTCTGGACCTGGTAGTCC
20 X4 f CTTAGCGCAATGCGAGGG
21 X5 r AAGCCCTTAAATATTGTTCGAG
22 X6 f AGGGAAAAGGCAACATTTCAAGTGGC
23 X7 r TTCCATCTTTCTGCGGGTAG
24 X8 f CAAAGAAACTGGACTACAAGAAAATCG
25 X9 r GAATCCAAGGCATCAAGGCCCATCATG
26 dutpasefw f CACCCCCACAGTGGTCTAGC
27 dutpasebk r GCCTTTCTGGACCTGGTAGTCC
28 rtfw f CGCCTCAAGATACGGAATG
29 rtbk r CGAGTATATGAGTACATCGTCC
30 probeltrf f CAAGCCTTGCGMATATGATTTATG
31 probeltrr r CTTAGAACAAAGCGTGAAGAACC
32 nancltr3 f TAYAMAAYACCTTTACGATC
33 nancltr4 r AAGCYAYRTTCDTGACA
34 18f f GTGGAAAAGCAGGGCATC
35 ltr3 f TATAMAAYACCTTTACGATCTTTATCYTTA
36 esp1f f CAGTCGATAGCTCTCCTGCTG
37 esp1b r TCTGAGTAGAGATTCCACATATGG
38 esp3b r GCTTGACTCATRTCGTATTGTAAGGf Forward, r reverse
123

Curr Genet (2008) 53:23–34 27
(Sambrook et al. 1989) onto Hybond N+ membrane(Amersham). The membrane was hybridised with two dis-tinct DNA probes: a 130 bp long probe (P1, Fig. 1c) span-ning the dUTPase conserved motives 2, 3 and 4 (Harriset al. 1999) and a 150 bp long probe (P2, Fig. 1c) span-ning the RT conserved motives 4 and 5 (Xiong and Eick-bush 1990). The probes were PCR ampliWed using theprimer pairs dutpasefw/dutpasebk and rtfw/rtbk, respec-tively (Table 1). The PCR reactions were performed in aWnal volume of 50 �l containing 200 �M of each dNTP,1.5 mM MgCl2, 10 mM Tris–HCl pH 9.0, 50 mM KCl,2.5 units of Taq polymerase (Amersham Biosciences),10 pmol of each primer and 20 ng of T. melanosporumgenomic DNA. The temperature cycling parameters con-trolled by using a Gene Amp 9700 Thermal Cycler(Applied Biosystems) were: an initial denaturation step at94°C for 2 min and 30 s; 30 cycles consisting of 30 s at94°C, 20 s at 58°C and 30 s at 72°C; a Wnal extension for7 min at 72°C. Probe labelling was performed using theReady To Go DNA labelling Kit (Amersham Biosciences)with �-P32 labelled dCTP.
Pre-hybridizations and hybridizations were done at 65°Caccording to the Hybond N+ supplier’s instructions. Post-hybridization washes and autoradiography were performedas previously reported (Paolocci et al. 2000).
Northern and RT-PCR analyses
Total RNA was isolated from T. melanosporum, T. indi-cum, T. brumale, T. aestivum and T. magnatum ascocarps(1 g), from ectomycorrhizae of T. melanosporum on Quer-cus pubescens and from Q. pubescens uninoculated rootsaccording to Chang et al. (1993). The RNA preparationswere treated with DNase using the DNA-freeTM kit(Ambion) according to the enzyme supplier’s instructions.The absence of residual DNA was veriWed by the null PCRampliWcation in the presence of the ITS1/ITS4 primer pair.Quality and quantity of RNA was veriWed by agarose gelelectrophoresis and spectrophotometric analysis. For north-ern analysis 20 �g of RNA were run, blotted and hybridizedas reported by Damiani et al. (1998). Five probes, namelyP1, P3, P4, P5 and P6 (Fig. 1c) were PCR ampliWed usingthe primer pairs dUTPasefw/dUTPasebk, probeltrf/prob-eltrr, X4/X5, ltr3/esp1b and esp1f/gw4 (Fig. 1; Table 1),respectively. The PCR conditions and probe labelling pro-cedures were as described for Southern analysis.
For RT-PCR analysis 5 �g of total RNA from T. melano-sporum ascocarps and ectomycorrhizae and from Q. pubes-cens uninoculated roots were reverse transcribed in thepresence of 200 units of SuperScriptIII Reverse Transcrip-tase (Invitrogen) using either random hexamers (AmershamBiosciences) or the Tmt1-speciWc reverse primer esp3b(Fig. 1b; Table 1) in combination with the reverse primer
EFtBw designed on the 1-� elongation factor gene (Paol-occi et al. 2004). The resulting cDNAs were PCR ampliWedusing six Tmt1-speciWc primer-pairs (X4/X5, rtfw/X5, 18f/rtbk, nancltr3/X5, nancltr3/nancltr4 and 18f/nancltr4,Fig. 1b; Table 1). As positive control, the same cDNAswere ampliWed with the primer pair EFtFw/EFtBw speciWcto the housekeeping gene 1-� elongation factor (Paolocciet al. 2004). PCR conditions were as follows: an initialdenaturation step at 94°C for 1 min and 30 s; 30 cycles con-sisting of 30 s at 94°C, 20 s at 45°C and 6 min and 30 s at72°C; a Wnal extension for 7 min at 72°C. Negative controls(no cDNA template and cDNA from uninoculated roots)were always performed.
Results
Isolation of retrotransposon-like sequences
Following the enrichment method for TEs proposed byPearce et al. (1999), seven T. melanosporum DNA frag-ments, with diVerent levels of similarity to retrotransposonsequences from public databases, were cloned. The highestlevels were shown by the 860 bp long clone C14a (Fig. 1a)presenting intriguing features with respect to the otherclones: (1) an approximately 250 bp long sequence at the 5�
end that showed similarity to pol genes of transposable ele-ments while retaining a high nucleotidic similarity to ss14,a previously isolated T. melanosporum species-speciWcSCAR (Sequence Characterized AmpliWed Region), Gen-Bank accession no. AF210077 (Paolocci et al. 2000); (2) anadditional 230 bp long trait showing amminoacidic similar-ity to dUTPases. Thus, the entire retroelement containingthe C14a genomic fragment was searched in subsequentrounds of GW (Fig.1a). Single GW experiments oftenyielded multiple clones, always showing point-mutationsrespect to each other (data not shown). Among the multipleclones, those showing the highest similarity to retroele-ments and harbouring the longest fragment were selected todesign primers for the next GW round. The overlappingregions of contiguous GW clones also showed point-muta-tions, suggesting to us that the retroelement under consider-ation represents a repetitive sequence in the T.melanosporum genome. The assembling of the eight puta-tively contiguous GW clones (C32, C8, C18, C14a, C5,C12, C15, C23, C20; Fig. 1a) led us to reconstruct the com-plete structure of the Tmt1 LTR-retroelement, (Fig. 1c).PCR primers were then designed to amplify a single full-length copy of Tmt1. Although no full-length Tmt1sequences were obtained, this strategy allowed us to reducethe number of contiguous clones from nine to Wve (Tmt1a,b, c, d and e, Fig. 1b) to reconstruct the entire structure ofTmt1 (Fig. 1).
123

28 Curr Genet (2008) 53:23–34
Structure of Tmt1
The Tmt1 element is 7499 bp long and it is Xanked by twoputative LTRs (Figs. 1c, 2), of about 560 bp, with 86%sequence similarity. They are not bordered by the invertedrepeats typical of retrotransposon and retroviral genomes.Searches were carried out for transcription regulatory ele-ments in the LTR sequences. As a result, three putativeTATA boxes and two putative CT boxes were identiWed(Fig. 2). A polyadenilation signal was not clearly detected.A polypurine tract (AGGAGGAGGGGAG), likely corre-sponding to the priming site for the (+)-strand DNA synthe-sis, is located immediately upstream of the 3� LTR. Theprimer binding site (PBS) at which minus strand reversetranscription initiates, could not be identiWed at the 3� endof the 5� LTR.
The coding region of Tmt1 is spanned by the largelyoverlapping clones Tmt1b, Tmt1c and Tmt1d (Fig. 1b, c).Given the high similarity at nucleotide level betweenTmt1b and Tmt1c and between Tmt1c and Tmt1d (95 and93.5%, respectively) we conventionally used Tmt1c as ref-erence sequence, except for the INT which was derivedfrom Tmt1d. The ORF search revealed that the putativecoding regions were disrupted by single nucleotide muta-tions generating frameshifts or stop codons. As a conse-quence the deduced amminoacidic sequences weremanually edited to recreate the complete structure of Tmt1.This analysis showed that Tmt1 contains the gag and polgenes typical of retrotransposons. The pol gene of Tmt1contains the aspartic protease (PRO), reverse transcriptase(RT), RNaseH (RNH) and integrase (INT) domains which
are arranged in the same order that typiWes the Ty3/gypsygroup of LTR-retrotransposons. However, the organizationof the pol gene in Tmt1 diVers signiWcantly from that of allother LTR retrotransposons for the presence of a dUTPasedomain located between the PRO and the RT domains(Fig. 1c).
The alignments of the putative GAG, PRO, RT, RNH andINT amminoacidic sequences of Tmt1 with those of otherfungal retroelements as well as of the putative Tmt1-dUTPasewith dUTPases from viruses and fungi are shown in Fig. 3.
The GAG protein is generally poorly conserved amongdiverse retroelements and retroviruses, with exception for thezinc-Wnger motif (CX2–C–X4–H–X4–C) (Covey 1986). Thismotif is responsible for the binding of GAG protein to geno-mic RNA of LTR retrotransposons. The putative zinc-Wngermotif of Tmt1 is CLKCGKAGHRIREG (Fig. 3a), endingwith a glycine residue instead of the conserved cysteine.
A region of about 86 amminoacids (aa) with similarity toPRO, containing the characteristic protease motifMLDSGA was found in the initial region of pol (Fig. 3b).The consensus amminoacidic sequence of this motif in themajority of retroelements is (hydrophobic residue)2-D-S/T-G-A/S (Dunn et al. 2002).
Downstream the PRO domain, an approximately 203 aalong region with strong similarity to dUTPases was found.The dUTPase of Tmt1 contains all Wve conserved motifsoccurring in viral and non-viral dUTPases (Harris et al.1999) (Fig. 3c).
The RT domain of Tmt1 (Fig. 3d) is about 180 aa long.It contains the seven characteristic motifs of RT (Xiong andEickbush 1990), with motif 5 including the YXDD box (X
Fig. 2 Alignment of Tmt1 5� and 3� LTR regions. Black lines indicate putative TATA-boxes a and CT-boxes b
09 08 07 06 05 04 03 02 01 |....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....
ATCCAATATACGCACTTTGTTATTGTATGGACACTGAGTATTGGTACACTGTGCGCTTTTGGGAAAGGAGTGATTCGGTGCAAGAACTGT '5 RTLACTCAGTATACACACTTTGTTATTGTATGGACACTGAGTATTGATACGCAGTGCACTTTTAGCAAAGGAGTGATTCGATGTAAGCACTGT '3 RTL
081 071 061 051 041 031 021 011 001 |....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....
TATGAGCTATCCACGTACAGGGTACTGAGTATGAGTTCGGTACACACATTGGTAGTGTTTAGGGTGTGTATTGATTAGTATGTTGGGCTT '5 RTLTATGAGCAGTCCACGTGCAGGGTACTGAGTATGA-CTCAGTACACGCTTTCGTAGTG-TTAGGGTGTGTATTGTTTAGTATACTGGGATT '3 RTL
072 062 052 042 032 022 012 002 091 |....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....
ATAAGTGCTTTGGGTACTGTAGTATTACTCCAGTACACGCTTTGTATTTAGTATACGCGTTCCGAACGGGCATTCCGTATGAGCATTCCG '5 RTLATACGCACTT--GGTACTG--AGATTACTCCAGTACACGGTTTGTATTTAGTATAAGCGTTCCGAACGGGCATTCCGCATGAGCCTTCCG '3 RTL
063 053 043 033 023 013 003 092 082 |....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....
GTTTTGGGTACTGGAATATCGCTCCAGTACACGTTTTGAGTATTTGGTGTAGTATTGCGTTATTTGTTCAGTGTACTGAGCTGACTCATT '5 RTLG-TTTGGGTACTGGAATATCGCTCCAGTACATG--TTGAGTATT---GATAGGATTGTGTTA-TTGTTCAGTATACTGAGCTGACTAG-- '3 RTL
054 044 034 024 014 004 093 083 073 |....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....
AAGGGCGGAAATTAGTCCACTGGATTTCATCAGTGTACACGTTTTGGTTTAGTTTTGTTTGATCAGTACACTGGTTTCACTAGTGTACGA '5 RTLAAGGGTGGGTATCAGTACACTGGATTTCACCAGTGTACACGCTTTGGTTTAGTATTGTTTGATTAGTTCACTGGTTTCACTAGT-TACGA '3 RTL
045 035 025 015 005 094 084 074 064 |....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....
AAGCAATTGCATAATACATCCTTTTCAAGAATCTTGTTTCGCACTTCTTGGATAGAAGGAGTGCGCGTAGCCGGAGGAATATATCACAAG '5 RTLAAGCAATTTCATTATTCATCCTTTTCATGAATCTTGTTTCGCACTTCTTGGATAGAAGGAGCCGGCGTAGCCGGAGGAAAATATCATAAG '3 RTL
065 055 .|....|....|....|....|....
CTATTTCTAGCATTTCCACAAAATAT '5 RTLCTATAGCTAGCATTTCCACAAAACAT '3 RTL
a
a b
ba
123

Curr Genet (2008) 53:23–34 29
is a leucine residue in Tmt1) speciWc of the active site (Var-mus and Brown 1989).
An approximately 224 aa long region of Tmt1 showssimilarity to RNH sequences of fungal retrotransposons.They usually contain the conserved motif FLG-X5-RXFI(Neuveglise et al. 1996). The corresponding Tmt1-RNHmotif is TLRFCNFYRKFI (Fig. 3e).
The INT region of Tmt1 (Fig. 3f) is about 310 aa longand it retains the three typical conserved domains describedby Khan et al. (1991) and Malik and Eickbush (1999): theN-terminal domain containing a zinc-binding site H–X6–H–X29–C–X2C, the core domain containing the catalytic motifD,D35-E and the C-terminal domain with the GPY/F mod-ule, which is constituted by the sequence G–(D/E)–X10–20–K–L–X2–(R/K)–(F/Y/W)–X–G–P–(F/Y)–X–(I/V). The GPY/F module is not universal in the Ty3/Gypsy group and it israrely found in retroviruses. The INT domain ends with thismodule in Tmt1, whereas in some members of the Ty3 line-age it continues with a sequence similar to a chromodomain(chromatin organization modiWer domain), that often occursin transcription factors playing a role in chromatin modiWca-tions (Malik and Eickbush 1999).
Finally, a 401 bp-long region of Tmt1 placed betweenthe gag–pol transition sequence and the PRO 3� end(Fig. 1c) shows a high similarity (76% of sequence iden-tity) with ss14, a T. melanosporum species-speciWc SCAR(Paolocci et al. 2000).
Tmt1 distribution within and among Tuber spp.
Southern analysis of Tuber spp. genomic DNAs was carriedout to evaluate the repetitiveness and species-speciWcity ofTmt1. The dUTPase (P1) and the RT (P2) probes produceda similar pattern with eight main bands on T. melanospo-rum DNA restricted with HindIII (Fig. 4 and data notshown), which did not cut between and inside the twoprobes. Both probes resulted to be T. melanosporum-spe-ciWc since no hybridization signal was detected in any otherTuber spp. (data not shown). As far as the technique usedpermits, no polymorphic signals among the 6 T. melano-sporum samples were revealed (Fig. 4).
Tmt is an inactive retrotransposon
With the aim to test for the presence of Tmt1 transcriptstotal RNAs from ascocarps of T. melanosporum, T. indicum,T. brumale, T. aestivum and T. magnatum were hybridizedwith two Tmt1-speciWc probes: the probe P3 designed onthe LTR and the probe P4 spanning most of the putativeTmt1 coding sequence. Neither RNAs from T. melanospo-rum fruit bodies nor from other Tuber spp. showed hybrid-izing bands with the two probes (data not shown). To checkfor the presence of Tmt1 transcripts in another truZe life
stage, northern analysis was then performed on RNAisolated from T. melanosporum ectomycorrhizae. To thispurpose replicated Wlters containing RNAs from T. melano-sporum ectomycorrhizae and ascocarps as well as fromuninoculated Q. pubescens roots were hybridized with theprobes spanning the dut gene (P1), the LTR (P3), a regionbetween the 5� LTR and the gag gene (P5) and the gag gene(P6). None of the probes produced hybridization signals onthe target RNAs (data not shown). The lack of Tmt1 tran-scripts was also conWrmed by RT-PCR analysis performedon cDNAs from T. melanosporum ascocarp and ectomy-corrhizae as well as from uninoculated host roots using sixTmt-1 speciWc primer-pairs. Positive and negative controlswere as expected (data not shown).
Phylogenetic analysis
The phylogenetic comparison of the RT, INT and RNHproteins with those of other retroelements showed thatTmt1 belongs to the highly supported cluster of Ty3/gypsyretrotransposons that is clearly separated from both retrovi-ruses and Ty1/copia retrotransposons. More speciWcally,the phylogram clearly indicates that Tmt1 is highly relatedto the other fungal and plant Ty3 retrotransposons withwhich it forms a cluster with more than 70% of bootstrapsupport (Fig. 5a).
Although supported by low bootstrap values, the phylo-genetic tree based on the alignment of the conserved motifsof Tmt1-dUTPases with those of other organisms indicatesthat the Tmt1-dUTPase is likely more related to dUTPasesof eukaryotes, mostly fungi and plants, than to those of ret-roviruses, the elements with the highest relatedness to LTRretrotransposons (Fig. 5b). Furthermore, from the sameanalysis it emerges that Tmt1-dUTPase does not show ahigh relatedness with the dUTPase of PcGR1, a Ty3/gypsyretrotransposon, isolated from the basidiomycete Phanero-chaete chrysosporium (accession no. AAR04354), that is,to the best of our knowledge, the only LTR retrotransposonharbouring a dut gene so far reported.
Discussion
In this paper, we report on the identiWcation of Tmt1, theWrst LTR-retrotransposon from a Tuber species. Tmt1 is aT. melanosporum-speciWc consensus sequence generatedfrom Wve overlapping PCR amplicons obtained by extend-ing to the 5� and 3� termini, through a genome walkingapproach, a core DNA fragment isolated by the enrichmentmethod proposed by Pearce et al. (1999). Tmt1 is 7,499-bplong and it contains 560-bp LTRs. The structural featuresand the phylogenetic analyses based on the deduced aminoacidic sequences of RT, RNH and INT indicate that Tmt1
123

30 Curr Genet (2008) 53:23–34
GA
G Z
INC
-FIN
GE
R
PR
OT
EA
SE
dU
TP
ase
cft-1
TDAMIDSGASG-NFASESFVTRNRIATCKKKEGYELIAVDG----S orfvirus -----------MAARHTSLAMEFCC--------------TETLR-----VVRLSQNATIPTRGSPGAAGLDLCSAYDCVVPSHCSCVVFTDLLIKLPPGCYG
cft-1
PSSRETRTCYGCGKPGHIARDCRGKNMVRR real
AQALVDSGCLCYSLVNKKFAYRHRLERFQIP-VRIIEGVNG----- fowl
-----------MDP-FGSSSVPPCS--------------TSDLPEPKLYFVRLSPHAVPPVRATHGAAGYDLFSAYDIKVPARGRALVPTDLVFQFPPGCYG
real
KTKRAPLTCYSCGKPGHIARDCQSTTRVRR grh
VNAQVDSGCECYAAMSDKCATRLRIERIPLPQARHVGTAVG----- S.cer
---------------------------------------MTATSDKVLKIQLRSASATVPTKGSATAAGYDIYASQDITIPAMGQGMVSTDISFTVPVGTYG
grh
QRRRETGACLRCGNSGHQVADCTYAAALRP maggy TYALTDCGAEGKCFLDQGWAEERQLQMYPLRNPFDIEVFDG---RT PcGR1
MAKVKQFHKKYSDTPKGISLLQCHHNEEDEFFVRELMLLQSWGADDQFEVELMNKDAKLPSRGSPESAGLDLFIMEDLTLMPHDRKLVPTGLRMCTPPGTYA
maggy
HNRKENMLCYRCGSQEHFVAKCPEPDTRRT skippy LSALVDSGADM-NFISPTTVNELRLPWKDKNDPYTVHDGQGETYLY
Tmt1c
ENKMTFIGKLCATQCLNSSPVVYSTTIREAEEETWHAKFAEMRQDKRVRIQKLDQEAKVPTKGSKGAAGHDLYANEGIKIPANGQATIATGIAIGLPEGTYG
skippy KRDKSKVTCYNCGKKGHYERECKNPVKTNQ Tmt1c
IYAMLDSGATE-DFIDKEICNKYGLHTKQAKKARQVYLADGK--PS
Tmt1c
ARRLNGKLCLKCGKAGHRIREGRSRLDTGy
cft-1
SLPSVERETIPLPLAIQRHHEEITLDVTD-MASHDIVLGMPWLRKH orfvirus RIAPRSGLAVKHFIDVGAGVIDEDYRGNVGVVLFNFGNSDFEVKKGDRIAQLICERISH-PAVQEVNRLDDTDRGDSGFGSTGSGACGGSGDTV--WYIS---
real
KLS-EINEVARFSFKLHGHEETAYAYVMDLSFGEDIYLGRGWMNHN fowl
RIAPRSGLAAKFFIDVGAGVIDPDYRGNVSVVLFNFSESSFNIRRGDRVAQLILERIMV-PELSELTQLGETDRGASGFGSTGMGAVDRNQRSVLEWLTPGSR
grh
RAQPMIRELAKCEMDVDGWVTPMLFYIVP-GLARDVILGLPWMTHR S.cer
RIAPRSGLAVKNGIQTGAGVVDRDYTGEVKVVLFNHSQRDFAIKKGDRVAQLILEKIVDDAQIVVVDSLEESARGAGGFGSTGN-------------------
maggy
AESGKCTHYVRGQLRIKDHIQKNALFFVTQLAHYPIVLGMPWLKQH
PcGR1
QVALQSGLSLR-GIDIGAGIINRDYQEELKILVINNSPNTFTFHKGDRVAQLMLERIFY-AEPTPVSRLDSTIRSSDGFGSTGI-------------------
skippy ENGNITREIDHLKVFVNGKNQGIDFDIIP-VWRYDLVLGYPWLLRY
Tmt1c
RIVPHSGLAVKHRLMTMAGVIDVDYTGEIKVVLANLAQEDYQVQKGDRIAQLIIEKINKED-LHEVDQPEETMRGNKGFGSTDQNQLDSTKRIEIKEITAQAF
Tmt1c AMGPIT-HTATVPMDIDSHRETATFQVAD-LKNHKIVLEMPWLRKH
RE
VE
RSE
TR
AN
SCR
IPT
ASE
cft-1 EKLAKGWIRRSTSSAGTPCMFVPKANG-KLRLVQDYRKLNEITIKNRYPLPNIEEAQDRLTGSDWYTKIDLRDAFYAIRMAEGEEWKTAF RTRYGLYEFLVMPMGLTNAPASCQDLVNETLRDLLDVCVVAYMDDILVYTKGSLQEHTKQVQDVFERLTKSGFKTAPEKCEFHKKEVKFLGFII
real SLLEKGFIRVSSSPAAAPVLFAKKPGG-GLRLCIDYRALNAITKKDRYPLPLIRETLNNLSKAKWFTKLDVIAAFHKIRVAEGDEWKTAF RTRFGLFEWLVTPFGMANSPSTFQRYINWTLREFLDDFCSAYLDDVLIYTDGSLKQHQEHVRKVLRKLQDAGLQVDIKKCEFEVKSTKYLGFII
grh DMMDKGWIRASSSSAAAPVLMVRKASG-GWRLCVDYRALNSITMQDRYPLPLIKETIRSLTGARWFTKVDVRAAFHKLRIAEGDEHLTAF RTRFGLFEWLVCPFGLAGAPATFQRYVNGVLGDTLGDYASAYLDDILIYSSGSKSDHWSKVTRVLDKLAAAGLNLDLDKSAFAVKEVKYLGFIV
maggy AELKKGFIRPSSSSVASPVLFVKKQGG-GLRFCVDYRALNNITVKDRYPLPLVRETLNNLAGMKFFSKIDIVSAFNNIRIKKGEEYLTAF RTRFGLYESLVMPFGLTGAPATFQRYINDSLREYLDVFCTAYLDDILIYSR-TRTEHEEHLKLVLEALRKAGLYANAAKCEFFVTETKFLGLLV
skippy DMIRKGYIRPSKSSAGFPVMFVPKPNSNKLRLVVDYRQLNEITEKDRTSLPLITELKDRLFGKKWFTALDLKSAYNLIRIKEADEWKTAF RTKYGLFEYLVMPFGLTNAPAVFQRMITNVLREYLDIFVVCYLDDILIFSD-TEEEHTEHVHKVLKALQDANMLVEPTKSHFHQSQVTYLGHEI
boty DNLAKGWIRESASQVASPTMWVPKKDG-PDRLVVDYRKLNTLTKKDRYPLPLATELRDRLGGRTIFTKMDLRNGYHLIRMKEGEEWKTAF KTRYGLYEYQVMPFGLTNAPATFMRLMNNVLSQYLDTCCICYLDDILVYSN-NKVQHIKDVSNILESLSKADLLCKPSKCEFHVTETEFLGFTV
afut1 KILKKRWIRESSSPAAALVLFIKKLDR-GLCLCINY*GLNAIIVKN*YPLP*INKLIDRI*GVKFFTKLDLWDAYYYIYI*QGNK*KIAF RTCYS*FKYLVMPFRLTNTPVTF*AYINKIMQGILNKFCIVYLDNILIYSQ-TEEEYE*YIKEVL*HLNSMNLYTKLLKYKFYKTEVKFLSFLI
Tmt1c
QNLRRGWIRDSFASRGPLILFVKRKDG-KMRLCVDYRALNSVTKKDRYPLPLIGEALDRL*SAKYFTKLDIKDAYHNVRIMAGDEWKTTF ASRYGMFEYLVMPFGLTNAPASFQRW-NRVLNKYLDICCIVYLDDVLIYSE-NIQQHRKDISNIMAAIQESGMKIKPTKCKFHKEETEYLGFII
RN
aseH
cft-1 STTGITIDPAKTQSIREWPEPKTVKDVQSFLGLANYNRKFIKDYSKTAAPMTMLTRKDV------NWKWGKEQTEAFKRLKEQCASAPTLRLFDGSKEVHIETDASDMAIGACLT------
real -------------------------------------------FSQLAAPLTRLT-GDV------SFRWGPEEQSAFDKLKEIFVSEPTLASFDPERETVLETDSSGFAVGGVLS------
grh AGEGVQADPEKIKAIRDWEAPTRLRGLRGFLGFANFYRDFIDGYSTLTAPLLALTKKGT------PFRWTEELEGAFEALKHAFLQAPILAQWDDAKDTRMETDCSGAALGGCLS------
maggy GVEGVKMDPEKITAVLDWQTPKKLTDVQAFLGFGNFYRRFIRDFSKIVAPLTRLTKKDV------AFEWNSACETAFRLLKRKFTEAPVLAHFDWEKDVILETDASDYVSAGILS------
skippy SHNEIRMDRRKIAAVAEWKVPTSVKETQSFLGFANYYRRFIKDFSKTAIPLTEITKKDK------QFQWNDKAQEAFEKLKSAITSEPVLVMFDPDRQVELETDASDFALGGQIG------
boty -SQGLKMSKDKVKAVLEWKQPTTIKEVQSFLGFVNFYRRFIKGYSGITTPLTTLTRKDQG-----SFEWTAKAQESFDTLKQAVAEEPILLTFDPEKEIIVETDSSDFAIGAVLS------
afut1 G*EGVWVDPI*LSTVSKWLVPYLFYNI*IFLGFIGFF*YFIKAYLQITIPLTNLLKGVKNRYNPRSFT*TEEAQKAF*DLKDAFIKTLILAHFNPKKLILLITNILGFIIIGILL*PDSNL
Tmt1c IEVGSRLTQLKQRPFGTGPNQTTKQMF-STLRFCNFYRKFIEGFSRLAKPLYNLTKKDK------KWE*GENEQKAYDAIRTHLTSALILVHFDPXXKTMIKPDASKYVCSGILS------
cft-1 -QTHD-GKRHPVAYYSRKMTTAEQNYDIHDKELLAIVAAMQHWRVYVEG-PPKLTILSDHKNLTYFTTTKELTRRQARWSELLGQYKFEIKYTPGTENGPADALSRRSDYMEGKEPV
real -QYGDDGVLRPCAYFSRKNNAHECNYEIHDKELLAVVRCLEEWDSELRSVER-FKVITDHKNLEYFMKPRMLNERQIRWSLLLGRYNMELLYRPGKQNVRADALSRREQDLPVG---
grh -QKGTDGLWRPVAFHSAKLTDAQRNYTIHDKELLAVIACLKAWDAELRSVRRPFLILTDHKALEYFSKPREVSERQMRWAETLSKFNYNLRFRPGRLAGVPDALSRREQDECTTPRL
maggy -QYGDDGILRPVAFFSKKHTATECNYEIYDKELLAIIRCFEEWRPELEGTSSPVQIITDHRNLEYFTTTKMLNRRQARWAEFLSRFNFRITYRPGKQGAKPDALTRRSEDMPEEGDE
skippy -QRDDNGVLHPIAFYSHKMHGAELNYPIYDKEFLAIVNCFKEFRHYLRGSKHPVKVFTDHKNIAYFATTQELNRRQLRYAEYLCEFDFTIAHCKGTDNGRADAISRRPDFDTGTVKT
boty -QPGQNGKYQPIAFYSRKLSPAELNYEIYDKELLAIVDAFREWRVYLEG-RYTVQVYTD---LV--TSPQRAYRR------------------------------------------
afut1 IETYSSHN*RLITFYLRKLYNTK**YEVYNQELLIIIKCFKY*RHYLEGSCYLIRV*TDYINLTYFFTTKTLNIRQA*WAELLAAYNFIIKYKLRQLN-PIDILLRY*DYKLTRDEE
Tmt1c -QQCDDGKWRPVSY*SKTITPAECNYDVANEELLAIVQALKEWRRYVSNNDHPVRVLTDHKNLVPFMTTKELNDCQIRWMEHLSKYNFKIEYRPGKESGKPDALTRIASDKPQMKED
INT
EG
RA
SE
cft-1 ECIRQHHDEPTYGHPGTSKTVDLIQRSFSFPQMRLKVLRYIKKCVHCQQNKAARHAKYGHLQFRTPPTKPWDEVTMDFITKLPRSKDRVTGQAYDMILVMVDRLTKYAHFIPASEIYTAEQLGYLVLDRLIRYHGFPEVFITDRDKLFTSNYWKTLMGTIGIKHKLSTAYHPETDGQTER
real QIISGAHDSLATGHPGREVTYKILARDYFWPGMTQTIRRYVRNCSTCGRSKSWREGKQGLLKPLPIPAQIWKEISMDFVEGLPESEG------MTNLMVITDRLSKGTIFVPLP-NIKTDTVVQKFIERVVAYHWLPDAITSDRGRQFVSVLWTKLCELLKINRRLSTAYHPQTDGATER
grh AVLQRTHESPMVGHSGRDGTFAILARDYHWDGMAEHVRRFVRNCDICRRTKPSRRARQGLLQPLPIPDRFWKQISIDFMTDLP-GNGEVT---PRYLMVITDRLSKYVQLEAMH-SMKAEDCAARFLSSWWRFRGFPSQIISDRGSDWVGGFWTELCRQTGVEQLLSTSYHPETDGGTER
maggy EILRRCHDSPVAGHPGKAKTYDLLSREYYWPGMLHYVSLWVKKCQTCRRINPSREGHQGLLRPLPTPERSWQHLSMDFITHLPQSNG------HDAILVVVDRLTKMRHFVPCKGTCNAEDTANLYLHHVWKLHGLPLTIVSDRGTQFVSKFWKHLTTRLKIDSLLSTAHHPETDGQTER
skippy EFIYEIHAHPLHGHQGVTKTMKRLQELGYRHFKKGQVEKVIKQCDLCAKTKAQRHKPYGQLQPLPVAQRPWDSITMDFITKLPLSEEPSTGIFYDSIMVIVDRLTKFSYYLPYREATDAEELSYVFYRHIVSIHGLPTEILSDRGPTFAATFWQSLMARLGLNHRLTTAFRPQVDGQTER
afut1 ELLKIHYNNPYTGHLGVEKIVALL*YKFY*EGM*HNIKEYIYIYAAY*KTKTPQHLTTGKLALLPILKEP*SNLSIDFITSLPPLA*--GQEVFNAILVIIN*YTKIAKYIPIRKIITTEELANVFLEKVIL*YSMLRSLVTNRDTLFTSYY*TNFYRSLRIKK*ISIVFYSQTNSQTER
Tmt1d DII*QLHDIRXAGHG-TAKTTEFITRSFYWPGLREDVK*YVKNCDICQRTKMAKHVPYGLLKSNEISEQPWETITMNFIMDLPTSEN------YDSILVVVDCLTKMSHFILVRKDIKLRDFIQTFFKEVFRLHGLPKNIITDRGTLFTSGLWKKVTGEMGIQNRMSTAFHPQTDGQTEW
cft-1 TNQTLEQYLRHYINYAQDNWVSLLPMAQIALNNHKSETTSTTPFMRTLARTLTYPE---------MAPLLKE-------GDKVYLLTKN-LKTRRQTKKLDHVKVGPFFID
real MNSVWETYIRSFTNWAQNDWALLCPMAQIAINGRTATSTSMSPFFLQHGYEVNPL---SQQEQERQANKTRREAQNFRVGDKVWLKLDQQYSTGRSSKKLDWKN-AKYTVI
grh ANQEVQQYLRAYIAFDQGDWPDHLGAAQLALNNRNSSVTGTSPNKLLLGFDIEAVP-------------------------------------------------------
maggy FNASLEQYLRAYVAYLQDDWESWLPLAEFTANSHKSETTGTSPFYATYGFHPRMGF--AQARYEEQANRHRTPARRLTVGQYVWLDARN-IQTARPQKKLDWKNLGPFRIS
skippy MNQVLEQYLRCYINYEQNDWVEKLPIAQLAYNTAYNESTKLTPAYANFGFTPNAYH--VRKRMKNYYDPKRLKGPTFSEGDMVYLATKN-IKTDRPSHKLDYKFIGPYKVL
afut1 QNQILEVYLRIFINY**DN*VI*LQIAEFAYNYTPYTLTKITPFKAAYGYVLVLPQ-------------------------------------------------------
Tmt1d
VNVILEQYIRAYINYQQDDWYKLLSMAECAYNNGKQETIGTTPFFANYGQHPQSQT-QAQQRQKEQYDKQRRPDPNIQPGDMVWLIPRN-IHTTQPSKKLDYKKIGPFRIL
mot
if 2
mot
if 1
mot
if 1
m
otif
2
mot
if 3
m
otif
7
zinc
bin
ding
sit
e co
re d
omai
n
mot
if 3
m
otif
5
mot
if 4
zn
mot
if 4
m
otif
5
mot
if 6
C-t
erm
inal
dom
ain
GP
Y/F
(a)
(d)
(e)
(f)
(b)
(c)
123

Curr Genet (2008) 53:23–34 31
�
belongs to the Ty3/gypsy group of LTR-retrotransposons.However, it also possesses an additional domain betweenthe PRO and the RT showing a high similarity with dUTP-ases.
The dUTPase catalyses the hydrolysis of deoxyuridinetriphosphate (dUTP) to deoxyuridine monophosphate(dUMP) and inorganic pyrophosphate (PPi). Thus, dUT-Pase, minimising the intracellular pool of dUTP, preventsuracil misincorporation into DNA by DNA polymerases.The dUTPase has proved to be an essential enzyme inE. coli and S. cerevisiae (el-Hajj et al. 1988; Gadsden et al.1993) and it is ubiquitous in archaea, eubacteria andeukaryotes. This enzyme is also present in some virusesand in particular in Retroviridae, Poxviridae and Herpesvir-
idae (Baldo and McClure 1999). The dUTPase motif wasalso identiWed in a few human endogenous retroviruses, e.g.the HERV-L family where dUTPase is located downstreamof the endonuclease/integrase domain (Cordonnier et al.1995). Indeed, to the best of our knowledge, informationabout LTR-retrotransposons carrying dut genes are limitedto the sole presence in public databases of a nucleotidesequence relative to a dut-containing Ty3/gypsy retrotrans-poson isolated from Phanerochaete chrysosporium (seeResults). Thus, in order to evaluate the probable origin ofthe dut gene present in Tmt1, a phylogenetic analysis ofdUTPase proteins from a large number of phylogeneticallydistant organisms was performed (Fig. 5b). The resultingphylogenetic tree suggests that TmT1-dUTPase is likelymore related to dUTPases from eukaryotes and, more spe-ciWcally from fungi, than to those from retroviruses. Inter-estingly, horizontal transfers of the dut genes have beenshown to occur between viruses and their hosts, such asbetween eukaryotes and Poxviruses and between eubacteriaand bacteriophages (Baldo et al. 1999). Thus, it cannot beruled out the hypothesis that a horizontal transfer of dutgene between a fungal host genome and Tmt1 occurred,most likely before the introgression of this retroelementinto the T. melanosporum genome.
Tmt1 is a degenerate retrotransposon with not identicalLTRs. This suggests an ancient integration of this retroele-ment into the T. melanosporum genome, after which theLTRs have undergone mutations. Further support to thisthesis is lent by the fact that we could not obtain any full-length copies of Tmt1, even using degenerated PCR prim-ers and low stringency conditions. Still better, no longORFs could be detected in the Tmt1 coding region, whichconversely is littered with many point mutations generatingstop codons, frameshifts and disruptions of functionaldomains. Intact and active retroelements are in fact gener-ally transcribed into a long polycistronic mRNA, that istranslated into a polyprotein, subsequently cleaved into themature proteins (Kingsman and Kingsman 1988).
Along this line, only a few of the approximately 30 LTRand non-LTR retrotransposons recognised in fungi (e.g.Skippy, Maggy and Cft-1) have been demonstrated to beactive elements (Anaya and Roncero 1995; Nakayashikiet al. 1999), the others most likely being “fossils” of activeelements, presenting numerous stop codons and frameshiftmutations in conserved domains (Daboussi and Capy2003). In both Class I and Class II TEs, fully functional
Fig. 3 Amminoacidic alignments of the putative GAG zinc-Wnger (a),PRO (b), RT (d), RNH (e) and INT (f) domains from Tmt1 and otherretrotransposons from Wlamentous fungi. c Shows the alignment of theTmt1-dUTPase with dUTPase of retroviruses and fungi. Accessionnos. are: cft-1, Cladosporium fulvum GenBank AF051915; real, Alter-naria alternata EMBL AB025309; grh, Magnaporthe grisea EMBLM77661; maggy, Magnaporthe grisea GenBank L35053; skippy,
Fusarium oxysporum GenBank L34658; boty, Botrytis cinerea EMBLX81791; afut1, Aspergillus fumigatus GenBank L76086; orf virus,GenBank NC005336; fowl adenovirus, GenBank AC000014; pcGR1,Phanerochaete chrysosporium GenBank AY425728; dUTPase fromSaccharomyces cerevisiae, GenBank CAA52322. Triangles under thealignment indicate the location of nucledotidic mutations generatingframeshifts. Asterisks indicate stop codons
Fig. 4 Southern analysis of HindIII-digested genomic DNA from sixT. melanosporum samples, hybridized with the P1 probe (dUTPase).The P2 probe (RT) yielded the same pattern (not shown). Numbersindicate fragment-sizes (kbp) according to GeneRuler DNA LadderMix (Fermentas)
123

32 Curr Genet (2008) 53:23–34
elements tend to be in the minority in most genomes. The vastmajority of copies are usually non-autonomous elements, oreven small fragments of full-size copies (Kidwell 2002).However, non-autonomous elements may still be activatedby transacting factors encoded by functional elements ofthe same family. Northern and RT-PCR analyses prompt usto exclude this possibility for Tmt1. As a matter of factTmt1-speciWc transcripts were not detected when RNAsfrom T. melanosporum ascocarps and ectomycorrhizaewere analysed. Nevertheless, we could not rule out the
hypotheses that Tmt1 is actively transcribed in diVerent lifestages and that it might be mobilized under the correct stim-uli. However, the possibilities of activating transposition asa response to controlled experimental stresses as well as ofstudying the Tmt1 expression pattern on diVerent fungallife stages are hindered by the diYculty of reproducing theTuber melanosporum life cycle in the lab.
Since transposons can trigger a wide range of eVects inthe host genomes, as changes in gene expression, genesequence and/or even chromosomal organization, mecha-
Fig. 5 Phylogenetic analysis of Tmt1. a Phylogram based on thealignment of conserved RT, RNH and INT domains from retrotranspo-sons and retroviruses. MarY1, Tricholoma matsutake (5002510); Mag-gy, Magnaporthe grisea (AAA33420); Grasshopper, M. grisea(AAA21442); Real, Alternaria alternata (BAA89272); CfT1, Clado-sporium fulvum (AAF21678); Cgret, Glomerella cingulata(AAG24792); Skippy, Fusarium oxysporum (AAA88791); Tf2, Sac-charomyces pombe (CAB58169); Sushi, Takifugu rubripes(AAC33526); IFG, Pinus radiata (AJ004945); Gm, Glycine max poly-protein (AAO23078); At1, Arabidopsis thaliana polyprotein(AAC69377); Os, Oryza sativa putative retrotransposon protein(ABA97430); Skypper, Dictyostelium discoideum (AAC48336); Ty3,Saccharomyces cerevisiae (AAA98435); At2, A. thaliana poliprotein(AAF24531); Ulysses, Drosophila virilis (CAA39967); Dr, Danio re-rio (XP_698865); Osvaldo, D. buzzatii (CAB39733); Mdg1, D. mela-nogaster (S70430); 412, D. melanogaster (CAA27750); Gypsy, D.melanogaster (P10401); 297, D. melanogaster (CAB57796); Tom, D.ananassae (CAA80824); TED, Autographa californica (AAA92249);ZAM, D. melanogaster (CAA04050); BLV, Bovine leukemia virus(BAA00544); RSV, Rous Sarcoma Virus (NP_056886); HIV1, Hu-man immunodeWciency virus 1 (AAP76556); Copia, Drosophila sp.(1107279B); Ty4, S. cerevisiae (AAA91746). b Phylogram based onthe alignment of dUTPase domain. Sp, Schizosaccharomyces pombe(Q9P6Q5); Cn, Cryptococcus neoformans (AAW46925); Af, Asper-gillus fumigatus (XP_750039); Nc, Neurospora crassa (CAE76286);Os, O. sativa (XP_469212); At, A. thaliana (NP_190278); Le, Lycop-
ersicon esculentum (AAB22611); Um, Ustilago maydis (EAK83957);Pb-cv, Paramecium bursaria Chlorella virus 1 (O41033); Ca, Candidaalbicans (CAA54897); Sc1, S. cerevisiae (NP_009811); Sc2, S. cere-visiae (CAA52322); Fa1, Fowl adenovirus 1 (AP_000401); Fa8, Fowladenovirus 8 (AAC71662); Rt-rv, Rana tigrina ranavirus(NP_572002). Ov, Orf virus (41057070); Bpsv, Bovine papular stoma-titis virus (AAR98364); Hs, Homo sapiens (AAB71393); Mm, Musmusculus (AAH19979); Rn, Rattus norvegicus (NP_446044); Pc,Phanerochaete chrysosporium retrotransposon polyprotein(AAR04354); Vv, Vaccinia virus (AAB96413); Swv, Swinepox virus(AAL69752); Shv, Sheeppox virus (NP_659591); Lsdv, lumpy skindisease virus (AAN02742); Asg, Agrotis segetum granulovirus(YP_006327); Mcn, Mamestra conWgurata nucleopolyhedrovirus B(AAM95060); Ws, Wolinella succinogenes (Q7M9N1); Hp, Helico-bacter pylori (Q9ZKY3); Rc, Rickettsia colorii (Q92I74); Atu, Agro-bacterium tumefaciens (Q8UII1); Mm, Magnetospirillum magneticum(BAE49005); r1t, Lactococcus bacteriophage r1t (NP_695048);TP901-1, Lactococcus lactis bacteriophage TP901-1 (NP_112684);T5, Enterobacteria phage T5 (AAS77177); SPBc2, BacteriophageSPBc2 (AAC13138); Jsrt, Jaagsiekte sheep retrovirus (P31625); Mtv1,Endogenous mouse mammary tumor virus Mtv1 (AAF31464); Eiav,Equine infectious anemia virus (1DUN); Srl, Small ruminant lentivirus(AAR22709); Fiv, Feline immunodeWciency virus (AAM13444); Pw,Pyrococcus woesei (AAR15897); St, Sulfolobus tokodaii(NP_377604)
123

Curr Genet (2008) 53:23–34 33
nisms of silencing of the foreign genetic material throughprocesses such as RIP (repeat-induced point mutation) andMIP (methylation induced premeiotically) can be activatedby the recipient genome (Daboussi 1997). These processeshave been envisioned to ensure genome stability. Prelimi-nar analyses based on the comparison of dinucleotide ratiosbetween Tmt1 and T. melanosporum structural genes (Mar-golin et al. 1998; Bibbins et al. 1998) reveal that likely RIPis not the inactivation mechanism for Tmt1 (data notshown).
As suggested by the high level of single nucleotide poly-morphism (SNP) among overlapping sequences obtainedby GW and, above all, as shown by Southern analysis,Tmt1 is a repeated element in the genome of T. melanospo-rum. Furthermore, the dut gene is very likely present inmany if not all Tmt1 copies, as the dUTPase and RT probesproduced two overlapping hybridization patterns. This evi-dence suggests an ancient introgression of dut gene into theTmt1 structure.
Moreover, since all the other Tuber spp. tested, do notcarry any Tmt1 homologous copy it is conceivable thatTmt1 have been integrated into the T. melanosporumgenome after separation from the most closely related blacktruZe species T. brumale and T. indicum (Rubini et al.1998; Roux et al. 1999). A similar hypothesis was alsoenvisaged for the ss14, a SCAR of T. melanosporum (Paol-occi et al. 2000).
To the best of our knowledge Tmt1 is the Wrst retroele-ment isolated from an ectomycorrhizal ascomycete. Theusefulness of retrotransposon-based strain identiWcation hasbeen recently reported for the cultivated ectomycorrhizalbasidiomycete Tricholoma matsutake (Murata et al. 2005).Along this line, a preliminary Tmt1-based REMAP(Retotransposon-Microsatellite AmpliWed Polymorphism)survey allowed us to detect polymorphic bands amongT. melanosporum samples of diVerent geographical origin(C. Riccioni, unpublished results). Thus, Tmt1-basedmolecular markers may represent a tool to respond to theneed for polymorphic markers to trace T. melanosporumstrains and populations. Research is also in progress toidentify putative intact and active retroelements in Tuberspp. that might be used in the near future for transposon-mediated mutagenesis.
Acknowledgments This research was partially funded by RegioneUmbria and Fondazione Cassa di Risparmio—Perugia. Contributionno. 103 from the Plant Genetics Institute, Perugia Division.
References
Altschul SF, Madden TL, SchaVer AA, Zhang J, Zhang Z, Miller W,Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a new gen-eration of protein database search programs. Nucleic Acids Res25:3389–3402
Amicucci A, Zambonelli A, Giomaro G, Potenza L, Stocchi V (1998)IdentiWcation of ectomycorrhizal fungi of the genus Tuber by spe-cies-speciWc ITS primers. Mol Ecol 7:273–277
Anaya N, Roncero MI (1995) Skippy, a retrotransposon from the fun-gal pathogen Fusarium oxysporum. Mol Gen Genet 249:637–647
Baldo AM, McClure MA (1999) Evolution and horizontal transfer ofdUTPase-encoding genes in viruses and their hosts. J Virol73:7710–7721
Bertault G, Raymond M, Berthomieu A, Callot G, Fernandez D (1998)TriXing variation in truZes. Nature 394:734
Bertault G, Rousset F, Fernandez D, Berthomieu A, Hochberg ME,Callot G, Raymond M (2001) Population genetics and dynamicsof the black truZe in a man-made truZe Weld. Heredity 86:451–458
Bibbins M, Cummings NJ, Connerton IF (1998) DAB1: a degenerateretrotransposon-like element from Neurospora crassa. Mol GenGenet 258(4):431–436
Ceruti A (1960) Elaphomycetales et Tuberales. In: Bresadola J (eds)Iconographia Mycologica vol. 28 suppl. II. Tridenti. Trento, Italy
Chang S, Puryear J, Cairney J (1993) A simple and eYcient method forisolating RNA from pine trees. Plant Mol Biol Rep 11:113–116
Cordonnier A, Casella JF, Heidmann T (1995) Isolation of novel hu-man endogenous retrovirus-like elements with foamy virus-relat-ed pol sequence. J Virol 69:5890–5897
Covey SN (1986) Amino acid sequence homology in gag region of re-verse transcribing elements and the coat protein gene of cauli-Xower mosaic virus. Nucleic Acids Res 14:623–633
Daboussi MJ (1997) Fungal transposable elements and genome evolu-tion. Genetica 100:253–260
Daboussi MJ, Capy P (2003) Transposable elements in Wlamentousfungi. Annu Rev Microbiol 57:275–99
Damiani F, Paolocci F, Consonni G, Crea F, Tonelli C, Arcioni S(1998) A maize anthocyanin transactivator induces pigmentationin hairy roots of dicotyledonous species. Plant Cell Rep 17:339–344
Dunn BM, Goodenow MM, Gustchina A, Wlodawer A (2002) Retro-viral proteases. Genome Biol 3:1–7
el-Hajj HH, Zhang H, Weiss B (1988) Lethality of a dut (deoxyuridinetriphosphatase) mutation in Escherichia coli. J Bacteriol170:1069–1075
Flavell AJ (1999) Long terminal repeat retrotransposons jump betweenspecies. Proc Natl Acad Sci USA 96:12211–12212
Gadsden MH, McIntosh EM, Game JC, Wilson PJ, Haynes RH (1993)dUTP pyrophosphatase is an essential enzyme in Saccharomycescerevisiae. EMBO J 12:4425–4431
Goodwin TJD, Poulter RTM (2001) The DIRS1 group of retrotranspo-sons. Mol Biol Evol 18:2067–2082
Goodwin TJD, Dalle Nogare DE, Butler MI, Poulter RTM (2003) Ty3/gypsy-like retrotransposons in Candida albicans and Candidadubliniensis: Tca3 and Tcd3. Yeast 20:493–508
Hall TA (1999) BioEdit: a user-friendly biological sequence alignmenteditor and analysis program for Windows 95/98/NT. Nucleic Ac-ids Symp Ser 41:95–98
Harris JM, McIntosh EM, Muscat GEO (1999) Structure/functionanalysis of a dUTPase: catalytic mechanism of a potential chemo-therapeutic target. J Mol Biol 288:275–287
Kempken F, Kück U (1998) Transposons in Wlamentous fungi—factsand perspectives. Bioessays 20:652–659
Khan E, Mack JP, Katz RA, Kulkosky J, Skalka AM (1991) Retroviralintegrase domains: DNA binding and the recognition of LTR se-quences. Nucleic Acids Res 19:851–860
Kidwell MG, Lisch D (1997) Transposable elements as sources of var-iation in animals and plants. Proc Natl Acad Sci USA 94:7704–7711
Kidwell MG (2002) Transposable elements and the evolution of ge-nome sizes in eukaryotes. Genetica 115:49-63
123

34 Curr Genet (2008) 53:23–34
Kingsman AJ, Kingsman SM (1988) Ty: a retroelement moving for-ward. Cell 53:333–334
Kumar A, Hirochika H (2001) Applications of retrotransposons as ge-netic tools in plant biology. Trends Plant Sci 6(3):127–34
Kumar S, Tamura K, Nei M (2004) MEGA3: integrated software formolecular evolutionary genetics analysis and sequence align-ment. Brief Bioinform 5:150–163
Malik HS, Eickbush TH (1999) Modular evolution of the integrase do-main in the Ty3/Gypsy class of LTR retrotransposons. J Virol73:5186–5190
Marchler-Bauer A, Anderson JB, DeWeese-Scott C, Fedorova ND,Geer LY, He S, Hurwitz DI, Jackson JD, Jacobs AR, LanczyckiCJ, Liebert CA, Liu C, Madej T, Marchler GH, Mazumder R,Nikolskaya AN, Panchenko AR, Rao BS, Shoemaker BA, Simo-nyan V, Song JS, Thiessen PA, Vasudevan S, Wang Y, YamashitaRA, Yin JJ, Bryant SH (2003) CDD: a curated Entrez database ofconserved domain alignments. Nucleic Acids Res 31:383–387
Margolin BS, Garrett-Engele PW, Stevens JN, Fritz DY, Garrett-Eng-ele C, Metzenberg RL, Selker EU (1998) A methylated Neuros-pora 5S rRNA pseudogene contains a transposable elementinactivated by repeat-induced point mutation. Genetics149:1787–1797
Montecchi A, Lazzari G (1993) Atlante FotograWco dei funghi ipogei.Associazione Micologica Bresadola, Trento
Murat C, Díez J, Luis P, Delaruelle C, Dupré C, Chevalier G, BonfanteP, Martin F (2004) Polymorphism at the ribosomal DNA ITS andits relation to postglacial re-colonization routes of the PerigordtruZe Tuber melanosporum. New Phytol 164:401–411
Murata H, Yamada A (2000) marY1, a member of the gypsy group oflong terminal repeat retroelements from the ectomycorrhizalbasidiomycete Tricholoma matsutake. Appl Environ Microbiol66:3642–3645
Murata H, Babasaki K, Yamada A (2005) Highly polymorphic DNAmarkers to specify strains of the ectomycorrhizal basidiomyceteTricholoma matsutake based on sigmamarY1, the long terminalrepeat of gypsy-type retroelement marY1. Mycorrhiza 15:179–86
Nakayashiki H, Kiyotomi K, Tosa Y, Mayama S (1999) Transpositionof the retrotransposon MAGGY in heterologous species of Wla-mentous fungi. Genetics 153:693–703
Neuveglise C, Sarfati J, Latge JP, Paris S (1996) Afut1, a retrotranspo-son-like element from Aspergillus fumigatus. Nucleic Acids Res24:1428–1434
Paolocci F, Cristofari E, Angelini P, Granetti B, Arcioni S (1995) Thepolymorphism of the rDNA region in typing ascocarps and ecto-mycorrhizae of truZe species. In: Stocchi V et al. (eds) Biotech-nology of Ectomycorrhizae. Plenum, New York, pp 171–184
Paolocci F, Rubini A, Granetti B, Arcioni S (1997) Typing Tuber mel-anosporum and Chinese black truZe species by molecular mark-ers. FEMS Microbiol Lett 153:255–260
Paolocci F, Rubini A, Riccioni C, Granetti B, Arcioni S (2000) Cloningand characterization of two repeated sequences in the symbiotic fun-gus Tuber melanosporum Vitt. FEMS Microbiol Ecol 34:139–146
Paolocci F, Rubini A, Riccioni C, Topini F, Arcioni S (2004) Tuberaestivum and Tuber uncinatum: two morphotypes or two species?FEMS Microbiol Lett 235:109–115
Paolocci F, Rubini A, Riccioni C, Arcioni S (2006) Re-evaluation ofthe life cycle of Tuber magnatum. Appl Environ Microbiol72:2390–2393
Pearce SR, Stuart-Rogers C, Knox MR, Kumar A, Ellis THN, FlavellAJ (1999) Rapid isolation of plant Ty1-copia group retrotranspo-son LTR sequences for molecular markers studies. Plant J19:711–717
Roux C, Séjalon-Delmas N, Martins M, Parguey-Leduc A, Dargent R,Bécard G (1999) Phylogenetic relationships between Europeanand Chinese truZes based on parsimony and distance analysis ofITS sequences. FEMS Microbiol Lett 180:147–155
Rubini A, Paolocci F, Granetti B, Arcioni S (1998) Single step molec-ular characterization of morphologically similar black truZe spe-cies. FEMS Microbiol Lett 264:7–12
Rubini A, Paolocci F, Granetti B, Arcioni S (2001) Morphologicalcharacterization of molecular-typed Tuber magnatum ectomy-corrhizae. Mycorrhiza 11:179–185
Rubini A, Paolocci F, Riccioni C, Vendramin GG, Arcioni S (2005)Genetic and phylogeographic structures of the symbiotic fungusTuber magnatum. Appl Envir Microbiol 71:2701–2705
Rubini A, Riccioni C, Arcioni S, Paolocci F (2007) Troubles withtruZes: unveiling more of their biology. New Phytol (OnlineEar-ly Articles) doi:10.1111/j.1469–8137.2007.01976.x
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular cloning—a lab-oratory manual, 2nd edn, CSH Press, Cold Spring Harbor
Schulman AH, Flavell AJ, Noel Ellis TH (2004) The application ofLTR retrotransposons as molecular markers in plants. In: MillerWJ, Capy P (eds) Mobile genetic elements. Humana Press, Tot-owa, pp 145–174
Siebert PD, Chenchik A, Kellogg DE, Lukyanov KA, Lukyanov SA(1995) An improved method for walking in uncloned genomicDNA. Nucleic Acids Res 23:1087–1088
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG(1997) The CLUSTAL_X windows interface: Xexible strategiesfor multiple sequence alignment aided by quality analysis tools.Nucleic Acids Res 25:4876–4882
Tunlid A, Talbot NJ (2002) Genomics of parasitic and symbiotic fungi.Curr Opin Microbiol 5:513–519
Varmus H, Brown P (1989) Retroviruses. In: Berg DE, Howe MM(eds) Mobile DNA. American Society for Microbiology, Wash-ington DC, pp 53–108
Vos P, Hogers R, Blecker M, Reijans M, Van de Lee T, Hornes M, Frij-ters A, Pot J, Pelman J, Kuiper M, Zabeau M (1995) AFLP: a newtechnique for DNA Wngerprinting. Nucleic Acids Res 23:4407–4414
Xiong Y, Eickbush TH (1990) Origin and evolution of retroelementsbased upon their reverse transcriptase sequences. EMBO J9:3353–3362
123




![Samsung SCX-4521F LTR [USA33839] Rep.pdf](https://static.fdokumen.com/doc/165x107/63192fdf1a1adcf65a0e8beb/samsung-scx-4521f-ltr-usa33839-reppdf.jpg)















