
FRAGMENTASI HUTAN : Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
142
FRAGMENTASI HUTAN FRAGMENTASI HUTAN Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan Hendra Gunawan Lilik Budi Prasetyo Pusat Penelitian dan Pengembangan Konservasi dan Rehabilitasi Badan Penelitian dan Pengembangan Kehutanan KEMENTERIAN KEHUTANAN
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of FRAGMENTASI HUTAN : Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan

FRAGMENTASI HUTAN FRAGMENTASI HUTAN Teori yang mendasari penataan ruang hutan
menuju pembangunan berkelanjutan
Hendra Gunawan
Lilik Budi Prasetyo
Pusat Penelitian dan Pengembangan Konservasi dan Rehabilitasi
Badan Penelitian dan Pengembangan Kehutanan
KEMENTERIAN KEHUTANAN

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
i
ISBN :978–602–1681–03-9

PRAGMENTASI HUTAN
ii
Judul : FRAGMENTASI HUTAN :
Teori yang mendasari penataan ruang hutan
menuju pembangunan berkelanjutan
ISBN : 978–602–1681–03-9
Penulis : Hendra Gunawan
Lilik Budi Prasetyo
Penelaah Ilmiah : Prof. Dr. Tukirin Partomiharjo, M.Sc.
Disain dan Tata Letak : Tatang Rohana
Foto Sampul Depan : Hendra Gunawan
Diterbitkan oleh : Pusat Penelitian dan Pengembangan Konservasi dan Rehabilitasi
Badan Penelitian dan Pengembangan Kehutanan Kementerian Kehutanan
Jl. Gunung Batu No.5, Bogor 16610
© Copyright 2013
Hak cipta dilindungi oleh undang-Undang
Dilarang memperbanyak buku ini tanpa izin tertulis dari penerbit.
Boleh dikutip dengan menyebutkan sumbernya.
Saran pengutipan :
Gunawan, H. dan L.B. Prasetyo. 2013. Fragmentasi Hutan :
Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan. Pusat Penelitian dan Pengembangan Konservasi dan
Rehabilitasi. Bogor.
Saran tentang buku mohon disampaikan ke :

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
iii
KATA PENGANTAR KEPALA PUSAT PENELITIAN DAN PENGEMBANGAN
KONSERVASI DAN REHABILITASI, BADAN PENELITIAN DAN PENGEMBANGAN KEHUTANAN
Salah satu tugas pokok lembaga penelitian adalah menghasilkan dan
mempublikasikan buku buku, baik yang bersifat teoritis maupun praktis,
baik hasil penelitian maupun hasil pemikiran dan pengkajian literatur.
Buku berjudul ”Fragmentasi Hutan : Teori yang mendasari penataan ruang
hutan menuju pembangunan berkelanjutan” merupakan salah satu produk
Pusat Penelitian dan Pengembangan Konservasi dan Rehabilitasi yang
diharapkan dapat membantu para prakitsi kehutanan, khususnya dalam
pengaturan tata ruang kawasan hutan.
Kepada penulis disampaikan penghargaan dan terima kasih atas
upayanya membukukan berbagai teori dalam sebuah buku yang mudah
dimengerti, baik oleh para praktisi maupun akademisi sehingga dapat
menjadi referensi, baik dalam penelitian maupun pengambilan keputusan.
Kepada Penelaah Ilmiah juga disampaikan terima kasih atas telaah dan
saran perbaikan untuk buku tersebut.
Akhirnya, semoga buku ini dapat bermanfaat untuk meningkatkan
kualitas pengelolaan hutan dalam rangka pembangunan berkelanjutan dan
berwawasan lingkungan.
Bogor, November 2013
Kepala Pusat,
Ir. Adi Susmianto, M.Sc.
NIP.195712211982031002

PRAGMENTASI HUTAN
iv
KATA PENGANTAR
Isu kepunahan keanekaragaman hayati tropika menjadi isu krusial
yang menjadi perhatian internasional pada dua dekade terakhir.
Fragmentasi hutan merupakan salah satu penyebab utama punahnya
keanekaragaman hayati di beberapa lokasi. Fragmentasi hutan merupakan
proses dan hasil dari peri laku manusia dalam memanfaatkan sumbedaya
hutan, seperti konversi, penebangan liar, pembakaran hutan, perladangan
dan perambahan kawasan hutan.
Keberhasilan pengelolaan keanekaragaman hayati di kantong-
kantong hutan yang telah terfragmentasi memerlukan pemahaman konsep-
konsep yang lahir dari teori biogeografi pulau, seperti fragmentasi, koridor
dan single large or several small (SLOSS). Buku ini membantu para
mahasiswa, praktisi konservasi, manajer satwaliar serta pengambil
kebijakan penataan ruang pembangunan dalam memahami fragmentasi
hutan dan kaitannya dengan pengelolaan dan konservasi keanekaragaman
hayati.
Buku ini merupakan kompilasi dari hasil terjemahan, penelaahan
dan analisis dari berbagai buku teks dan website yang relevan. Untuk
memahami dan mengetahui lebih detail, para pembaca dipesilakan
membaca buku-buku teks yang menjadi sumber penulisan buku ini.
Penulis menyadari, buku ini masih jauh dari kesempurnaan, oleh
karena itu saran-saran dari peminat akan dipertimbangkan untuk perbaikan.
Akhirnya, semoga buku ini bermanfaat bagi para peminat dan pemerhati
masalah konservasi keanekaragaman hayati.
Bogor, November 2013
Penulis

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
v
DAFTAR ISI
Halaman
BAB 1. FRAGMENTASI: SUATU PENDAHULUAN ..................... 1
BAB 2. MASALAH DAN PROSES FRAGMENTASI ...................... 10
BAB 3. TEORI BIOGEOGRAFI PULAU vs FRAGMENTASI HUTAN ... 16
BAB 4. FRAGMENTASI HABITAT DIPANDANG DALAM SKALA LANSKAP ........................................................
30
BAB 5. KONSEP KORIDOR DAN SINGLE LARGE OR SEVERAL SMALL (SLOSS) .................................................
42
BAB 6. DINAMIKA POPULASI PADA SKALA LANSKAP ................ 52
BAB 7. PENGARUH FRAGMENTASI TERHADAP SATWALIAR ....... 71
BAB 8. PERPINDAHAN SATWA DALAM LANSKAP .................... 95
BAB 9. FRAGMENTASI DAN DINAMIKA METAPOPULASI ............. 103
BAB 10. IMPLIKASI PENGELOLAAN KONSERVASI ..................... 109
BAB 11. STUDI KASUS: Evaluasi Lanskap Kawasan Merapi –
Merbabu Dan Telaah Kemungkinan Sebaran Macan Tutul Di Katong-Kantong Hutan Yang Terfragmentasi ...
112
RIWAYAT PENULIS ..................... ..................... .............. 130

PRAGMENTASI HUTAN
vi
DAFTAR GAMBAR Nomor Teks Halaman
Gambar 1. Model konseptual pengaruh fragmentasi
(Kupfer et al. 2004) ...............................................
13
Gambar 2. Jumlah jenis relatif pada (a) pulau-pulau kecil, dekat,
(b) pulau-pulau besar, jauh atau kecil, dekat, dan (c) pulau-pulau besar, dekat (Mac Arthur and
Wilson 1967) ..................................................
21
Gambar 3. Mekanisme bekerjanya pengaruh isolasi ................. 27
Gambar 4. Klasifikasi rejim gangguan berdasarkan pada luas yang
terganggu dan luas lanskap (diadaptasi oleh Shugart 1984; dari Shugart and West 1981)……………..............
32
Gambar 5. Empat tipe gangguan yang ditunjukkan oleh derajat
gangguan dan luas geografis yang dipengaruhinya (Morrison et al. 1992) .......................................
32
Gambar 6. Fungsi insiden dari pelatuk totol besar (Picoides
major) (Dari Moore dan Hooper dalam Wilcove et al. 1986) ...................................................
35
Gambar 7. Contoh koridor perpindahan yang dibuat di
pegunungan Costa Rica (Dari Stiles and Clark 1989) …
45
Gambar 8. Perbandingan SLOSS (Dari berbagai sumber) ............ 50
Gambar 9. Gradien pertukaran di dalam metapoulasi dari
berbagai struktur internal (Morrison et al. 1992) ......
56
Gambar 11a. Batas Das dan sistem riparian (riverine) (Morrison et al. 1992) .......................................
58
Gambar 11b. Kejadian sepuluh patches dari hutan tua
(Morrison et al. 1992) .......................................
58
Gambar 11c. Kejadian vertebrata obligat dewasa di tujuh dari 10
patches (Morrison et al. 1992) .............................
59

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
vii
Gambar 11d. Seleksi tiga patches untuk pemanenan tebang habis
(Morrison et al. 1992) ....................................... 59
Gambar 11e. Akibat langsung dari ganguan pemanenan : hilangnya
spesies pada tiga patches (Morrison et al. 1992) ......
60
Gambar 11f. Kehilangan spesies kemudian pada patch hutan yang
jauh, terisolasi dan kecil (faunal relaxation) (Morrison et al. 1992) ...................................................
60
Gambar 11g. Masih kehilangan spesies kemudian pada suatu patch
hutan lebih besar yang kini terisolasi (Morrison et al. 1992) .......................................
61
Gambar 12. Contoh habitat yang terfragmentasi oleh: (1) jaringan
listrik, (2) jalan raya, (3) tebang habis dan (4)
pertanian (Foto: Hendra Gunawan, 2008) ...............
73
Gambar 13. Sekumpulan patch hutan yang ditempati oleh
populasi-populasi penghuni hutan. Tebal tipisnya
panah menunjukkan laju pertukaran individu antar populasi (panah tebal menunjukkan lebih banyak
pertukaran). Catatan: Laju pertukaran dikoreksi oleh seberapa dekat patch-patch dalam ruang (Sumber:
http://chesapeake.towson. edulandscape/forestfrag/effects.asp ....................
76
Gambar 14. Penghunian (occupancy) sekumpulan patch hutan yang
mendukung suatu metapopulasi spesies penghuni hutan pada dua waktu yang berbeda. Patch hitam
ditempati dan patch putih tidak ditempati. Catatan: penghunian pada patch individual berubah sepanjang
waktu (seperti patch-patch dikolonisasi dan populasi-populasi di dalam patch punah), tetapi jumlah patch-
patch yang ditempati tetap sama sepanjang waktu (Sumber: http://chesapeake. towson.edu/landscape/
forestfrag/effects.asp ......................................
76
Gambar 15. Grafik menunjukkan hubungan antara kelembaban
tanah dan edge dan core habitat dala patch hutan.
Banyak spesies burung tidak dapat mentolerir kondisi tepi habitat (edge habitat) dan hanya ditemukan di
core habitat (Sumber:http://chesapeake.towson. edulandscape/forestfrag/effects.asp) ...................
78
Gambar 16. Berbagai bentuk fragment dan gambaran luas
interior-nya ...................................................
80

PRAGMENTASI HUTAN
viii
Gambar 17. Pengaruh kehilangan dan fragmentasi habitat pada
ukuran patch dan isolasi (Fahrig 1997) …………………….. 82
Gambar 18. Ilustrasi zona pengaruh tepi (edge effect) suatu pulau
habitat………………………………………………………….............
86
Gambar 19. Koridor harus cukup lebar untuk memberikan manfaat
positif lebih banyak bagi satwaliar. Koridor riparian yang sempit seperti yang tampak dalam gambar
berpeluang menghasilkan damak negatif bagi satwaliar ………………………………………………………………………
101
Gambar 20. Metapopulasi adalah suatu populasi sumber (source)
dan populasi penerima (sink) (Barnes 2000) ……….………
106
Gambar 21. Ukuran, bentuk, konfigurasi dan jumlah patch semua
mempengaruhi jumlah habitat interior dalam patch kecil, tunggal, patch-patch persegi memberikan
jumlah habitat interior yang kecil dan patch-patch lingkaran besar memberikan habitat interior terbesar
(Barnes 2000) ................................................
110
Gambar 22. Tahapan prosedur kajian evaluasi lanskap dan ana lisis
spasial .........................................................
115
Gambar 23. Komposisi luas setiap kelas penutupan lahan di
kawasan lanskap Merapi Merbabu .........................
118
Gambar 24. Jumlah patch setiap kelas penutupan lahan ……………… 119
Gambar 25. Luas patch rata-rata setiap kelas penutupan lahan .... 120
Gambar 26. Total edge setiap kelas penutupan lahan ................ 120
Gambar 27. Edge density setiap kelas penutupan lahan ............. 121
Gambar 28. Mean patch edge setiap kelas penutupan lahan ………… 121
Gambar 29. Mean shape index setiap kelas penutupan lahan …..... 121
Gambar 30. Mean patch fractal dimension untuk setiap kelas
penutupan lahan …………………...............................
123
Gambar 31. Lokasi dugaan habitat macan tutul terpilih (warna
kuning) hasil query ..........................................
125

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
ix
DAFTAR TABEL
Nomor Teks Halaman
Tabel 1. Implikasi dispersi habitat pada skala berbeda untuk berbagai atribut lanskap terfragmentasi ....................
33
Tabel 2. Klasifikasi faktor-faktor untuk dievaluasi dalam suatu studi gangguan lanskap ……………...............................
54
Tabel 3. Komposisi pentutupan lahan AOI Kawasan Merapi-Merbabu 116
Tabel 4. Hasil patch analyses pada skala lanskap kawasan Merapi-
Merbabu ............................................................
117
Tabel 5. Hasil patch analyses skala kelas kawasan Merapi–Merbabu 117

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
1
1
FRAGMENTASI : SUATU PENDAHULUAN
PENDAHULUAN
DEFINISI DAN PENGERTIAN
MENGAPA PERLU MEMPELAJARI FRAGMENTASI?
DAFTAR PUSTAKA
I. PENDAHULUAN
Mempelajari fragmentasi merupakan bagian dari i lmu ekologi
lanskap. Ekologi lanskap merupakan i lmu baru yang diturunkan dari disiplin
terdahulu sebagai suatu gabungan arsitektur lanskap, zoogeografi, geografi
tumbuhan dan sinekologi. Ekologi lanskap merupakan i lmu yang
mempelajari respon spesies atau komunitas terhadap pola-pola (patterns)
patch dalam suatu lanskap (Morrison et al. 1992). Banyak prinsip-prinsip
dasar ekologi lanskap diturunkan dari biogeografi pulau, zoogeografi dan
fitogeografi, yaitu i lmu yang mempelajari penyebaran dan pergerakan
satwa dan tumbuhan melintasi pulau-pulau dan wilayah geografi yang lebih
luas (Morrison et al. 1992).
Ekologi lanskap banyak memfokuskan pada dinamika tumbuhan dan
satwa di dalam patches (kantong habitat), khususnya di pulau-pulau dan
lingkungan terisolasi lainnya. Teori pulau klasik menyatakan bahwa di
pulau, dinamika populasi dan struktur komunitas ditentukan oleh beberapa
faktor : ukuran (pulau kecil memiliki resiko kepunahan lokal lebih besar);
jarak dari sumber spesies yang mengkolonisasi (semakin jauh, semakin
tinggi laju kepunahannya); dan atribut spesies meliputi kemampuan

PRAGMENTASI HUTAN
2
dispersal, demografik (survivorship, rekrutmen); dan spesialisasi
penggunaan habitat (Morrison 1992).
Pemukiman manusia dan kegiatan terkait lainnya, seperti hutan
tanaman dan pertanian telah mengubah lanskap alami menghasilkan suatu
mosaik dari habitat yang terfragmentasi. Fragmentasi habitat dapat
memberikan pengaruh merugikan pada flora dan fauna dari habitat alami
yang sebelumnya masih utuh dan berkesinambungan. Ada dua pengaruh
utama dari fragmentasi yaitu : mengurangi total luas dari habitat asal dan
menciptakan wilayah tepi (edge area) di antara habitat asal dengan lanskap
yang terganggu oleh manusia, yang dikenal sebagai efek tepi (edge effects).
Dalam fragmentasi habitat ada enam proses terpisah yang dapat
dipertimbangkan yaitu:
Berkurangnya luas total dari habitat
Meningkatnya jumlah wilayah tepi (edge)
Berkurangnya luasan habitat interior
Terisolasinya suatu fragment (potongan) habitat dari wilayah habitat
lainnya
Terpecahnya satu patch (kantong) habitat menjadi beberapa patch
(kantong) habitat yang lebih kecil
Berkurangnya ukuran rata-rata setiap patch (kantong) habitat
II. DEFINISI DAN PENGERTIAN
A. Ekologi Lanskap
Ekologi lanskap merupakan suatu bagian dari i lmu ekologi yang
mempelajari bagaimana struktur lanskap mempengaruh kelimpahan dan
distribusi organisme. Ekologi lanskap juga didefinisikan sebagai ilmu yang
mempelajari pengaruh pola (pattern) dan proses, dimana pola di sini
khususnya mengacu pada struktur lanskap. Dengan demikian secara
lengkap ekologi lanskap dapat didefinisikan sebagai ilmu yang mempelajari
bagaimana struktur lanskap mempengaruhi (memproses dan membentuk)
kelimpahan dan distribusi organisme.
Definisi lain menyebutkan, ekologi lanskap merupakan sub disiplin
ekologi dan geografi yang khusus mempelajari variasi spasial dalam lanskap

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
3
yang mempengaruhi proses-proses ekologi seperti distribusi, aliran energi,
materi dan individu dalam lingkungannya (yang pada gilirannya mungkin
mempengaruhi ditribusi elemen-elemen lanskap itu sendiri). Teori
biogeografi pulau dari MacArtur dan Wilson merupakan fokus dari ekologi
lanskap yang oleh Levin digunakan untuk menjelaskan model metapopulasi.
Dalam ekologi lanskap dapat dipelajari bagaimana fragmentasi habitat
mempengaruhi daya hidup suatu populasi (population viability). Dalam
perkembangannya ekologi lanskap banyak menggunakan teknologi Sistem
Informasi Geografis (SIG) dan banyak data habitat yang tersedia (seperti
citra satelit dan foto udara.
Sebagai bagian dari ekologi, ekologi lanskap ditujukan mempelajari
sebab dan akibat dari heterogenitas spasial (Forman 1995). Heterogenitas
merupakan ukuran bagaimana bagian-bagian suatu lanskap berbeda satu
sama lain. Ekologi lanskap melihat pada bagaimana struktur spasial
mempengaruhi kelimpahan organisme pada skala lanskap, serta perilaku
dan fungsi lanskap secara keseluruhan. Hal ini berarti juga mempelajari
pola, atau keteraturan internal lanskap, proses atau operasi kontinu dari
fungsi organisme (Turner 1989). Ekologi lanskap juga mencakup
geomorfologi dalam penerapannya untuk disain dan arsitektur lanskap
(Allaby 1998). Geomorfologi merupakan i lmu yang mempelajari bagaimana
formasi geologi mempengaruhi struktur lanskap.
B. Fragmentasi
Fragmentasi hutan terjadi karena hutan yang luas dan menyambung
terpecah menjadi blok-blok lebih kecil akibat pembangunan jalan,
pertanian, urbanisasi atau pembangunan lain. Akibatnya mengurangi fungsi
hutan sebagai habitat berbagai spesies tumbuhan dan satwaliar.
Fragmentasi juga mempengaruhi struktur, temperatur, kelembaban dan
pencahayaan yang akan mengganggu satwa hutan yang adpatasinya telah
terbentuk selama ribuan tahun. Fragmentasi didefinisikan sebagai
pemecahan habitat organisme menjadi kantong-kantong (patches) habitat
yang membuat organisme kesulitan melakukan pergerakan dari kantong
habitat yang satu ke yang lainnya. Fragmentasi dapat disebabkan oleh
penghilangan vegetasi pada areal yang luas atau oleh jalan yang

PRAGMENTASI HUTAN
4
memisahkan habitat bahkan oleh jaringan kabel listrik (Rusak & Dobson
2007).
Fragmentasi adalah proses pemecahan suatu habitat, ekosistem
atau tipe landuse menjadi bidang-bidang lahan yang lebih kecil dan
fragmentasi juga merupakan sebuah hasil dimana proses fragmentasi
mengubah atribut-atribut habitat dan karakteristik suatu lanskap yang ada.
Fragmentasi habitat mengubah konfigurasi spasial suatu kantong habitat
(habitat patches) besar dan menciptakan isolasi atau perenggangan
hubungan antara kantong-kantong (patches) habitat asli karena terselingi
oleh mosaik yang luas atau tipe habitat lain yang tidak sesuai bagi spesies
yang ada (Wiens 1990).
Franklin et al. (2002) mengembangkan definisi baru tantang
fragmentasi sebagai hasil (outcome) dan proses. Hasil (outcome) dari
fragmentasi habitat adalah diskontinuitas yang diperoleh dari serangkaian
mekanisme, di dalam distribusi spasial suatu sumberdaya dan kondisi yang
ada dalam suatu areal pada suatu skala tertentu yang mempengaruhi
okupansi, reproduksi atau survival suatu spesies. Fragmentasi habitat
didefinisikan sebagai serangkaian mekanisme yang mengakibatkan
diskontinuitas distribusi spasial suatu habitat. Ada empat komponen kunci
dari dua definisi tersebut yaitu : (1) diskontinuitas, (2) mekanisme, (3)
distribusi spasial dari suatu sumberdaya dalam suatu area, dan (4) atribut
demografik (Franklin et al. 2002).
Konsep fragmentasi habitat diturunkan dari teori biogeografi pulau
(MacArthur & Wilson 1967), yakni jumlah spesies meningkat dengan
meningkatnya ukuran pulau (Haila 2002). Fragmentasi penting mendapat
perhatian karena berpengaruh pada kekayaan spesies dari komunitas, trend
populasi beberapa spesies dan keanekaragaman hayati ekosistem secara
keseluruhan (Morrison et al. 1992).
Menurut Wilcove (1987) dalam Morrison et al. (1992) ada empat
cara fragmentasi dapat menyebabkan kepunahan lokal : (1) spesies dapat
mulai keluar dari kantong habitat yang terlindungi; (2) kantong habitat
gagal menyediakan habitat karena pengurangan luas atau hilangnya
heterogenitas internal; (3) fragmentasi menciptakan populasi yang lebih
kecil dan terisolasi yang memiliki resiko lebih besar terhadap bencana,

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
5
variabilitas demografik, kemunduran genetik atau disfungsi sosial; (4)
fragmentasi dapat mengganggu hubungan ekologis yang penting sehingga
dapat menimbulkan sebab sekunder kepunahan dari hilangnya spesies kunci
dan pengaruh merugikan dari lingkungan luar dan efek tepi (edge effect).
C. Patch
Patch, merupakan suatu terminologi dasar dalam ekologi lanskap
didefinisikan sebagai sebuah area yang relatif homogen yang berbeda
dengan sekelilingnya (Forman 1995). Patch merupakan unit dasar dari
lanskap yang berubah dan berfluktuasi. Proses perubahan dan fluktuasi ini
disebut sebagai dinamika patch. Patch memiliki bentuk tertentu dan
konfigurasi spasial, dan dapat digambarkan dalam komposisi variabel-
variabel internalnya seperti jumlah pohon, jumlah jenis pohon, tinggi
pohon, atau variabel lainnya (Forman 1995).
D. Matrix
Matrix merupakan latar belakang (background) sistem ekologi dari
suatu lanskap dengan derajat konektifitas yang tinggi. Konektivitas adalah
ukuran bagaimana suatu koridor, jaringan (network) atau matrix terhubung
atau berkesinambungan (Forman, 1995). Sebagai contoh, suatu lanskap
berhutan (matrix) yang memiliki sedikit celah (gap) dalam tutupan
hutannya berarti memiliki konektifitas lebih tinggi. Koridor memiliki fungsi
penting sebagai jalur penghubung antara suatu tipe tutupan lahan yang
berbeda dengan tetangga di kedua sisinya (Forman, 1995). Suatu jaringan
(network) merupakan suatu sistem hubungan antar koridor, sementara
mosaik menggambarkan pola (pattern) dari patch, koridor dan matrix yang
membentuk suatu lanskap dalam suatu kesatuan (Forman 1995).
E. Boundary dan Edge
Patches bisa memiliki batas (boundary) yang jelas atau tidak jelas
(kabur) (Sanderson & Harris 2000). Suatu zona yang tersusun atas ekosistem
edge di perbatasan disebut boundary (Forman 1995). Edge (tepi) berarti
bagian dari suatu ekosistem yang berdekatan dengan garis kelilingnya
(perimeter), dimana pengaruh-pengaruh dari patch yang berdekatan dapat

PRAGMENTASI HUTAN
6
menyebabkan perbedaan lingkungan antara interior suatu patch dengan
tepiannya (edge). Efek tepi (edge effect) ini meliputi perbedaan komposisi
spesies atau kelimpahan di bagian luar patch (Forman 1995). Sebagai
contoh, ketika suatu lanskap merupakan sebuah mosaik dari tipe-tipe
seperti hutan berdekatan dengan padang rumput, maka edge-nya adalah
lokasi di mana kedua tipe tersebut bergabung. Dalam lanskap yang
kontinu, seperti dari hutan ke kebun kayu, maka lokasi edge-nya menjadi
kabur dan kadang-kadang dibedakan oleh gradien lokal ketika melampaui
suatu ambang batas, misalnya penutupan pohonnya di bawah 35 persen
(Turner and Gardner 1991).
F. Habitat
Habitat adalah suatu tipe komunitas biotik atau kesatuan komunitas
biotik dimana spesies atau populasi hidup (Bailey, 1984). Habitat adalah
suatu unit lingkungan, alami maupun tidak (meliputi iklim, makanan, cover
dan air) dimana seekor satwa, tumbuhan atau populasi secara alami dan
normal hidup dan berkembang (Helms, 1998). Definisi habitat terbaru yang
relevan untuk pengelola satwa liar datang dari Hall et al. (1997) yaitu
sumberdaya dan kondisi yang ada pada suatu tempat yang memberikan
tempat hidup (occupancy), termasuk survival dan reproduksi suatu
organisme. Definisi ini berimplikasi bahwa habitat adalah sejumlah
sumberdaya spesifik yang dibutuhkan oleh suatu spesies (Hall et al. 1997).
III. MENGAPA PERLU MEMPELAJARI FRAGMENTASI?
Sampai saat ini para ahli satwa memiliki pandangan tradisioanal
tentang edges dan ekoton sebagai sesuatu yang positif. Dalam buku-buku
teks klasik (Leopold 1933; Thomas et al. 1979; Yoakum & Dasmann 1971
dalam Morrison et al. 1992) merekomendasikan bahwa pengelola satwa
seharusnya meningkatkan jumlah edges, karena satwa melimpah pada
pertemuan habitat (mereka menganggap habitat sebagai vegetasi atau
lingkungan secara umum, bukan habitat spesifik suatu spesies yang
memerlukan syarat khusus). Anggapan bahwa pembukaan dan pembersihan
hutan menguntungkan satwa ternyata tidak selalu benar. Dalam beberapa
dekade terakhir terbukti bahwa banyak hutan yang dulunya

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
7
berkesinambungan sekarang terancam karena dibuka untuk membuat
ekoton sehingga berdampak negatif bagi satwa seperti meningkatnya
pemangsaan dan overbrowsing. Anggapan bahwa menciptakan edges dan
membuka hutan itu bermanfaat perlu diteliti lagi dengan seksama (Morrison
et al. 1992).
Jumlah spesies satwa dan tumbuhan berhubungan dengan kondisi
yang ada dalam interior yang relatif luas dari vegetasi matang atau
dipengaruhi oleh vegetasi terganggu dan pengaruh tepi. Komposisi dan
kelimpahan relatif dari asosiasi spesies dalam komunitas biotik bervariasi
antar lanskap menurut jumlah dan sebaran habitat secara spasial. Dengan
demikian fragmentasi berpengaruh terhadap kekayaan spesies dari suatu
komunitas, kecenderungan populasi spesies tertentu dan keanekaragaman
hayati secara keseluruhan dari suatu ekosistem. Apakah pengaruh tersebut
disukai atau tidak bergantung pada jumlah dan komposisi spesies yang ada
pada suatu area. Hal ini memiliki implikasi kuat untuk pengelolaan lanskap
(Morrison et al. 1992).
Fragmentasi hutan juga menjadi perhatian di dalam hutan tropis.
Klein (1989) dalam Morrison et al. (1992) yang melakukan penelitian
terhadap kumbang bangkai dan organisme pada kotoran (feces) di hutan
bersinambung dan hutan terfragmentasi, menyimpulkan bahwa fragmentasi
menyebabkan berkurangnya spesies pengurai kotoran dan secara tidak
langsung fragmentasi hutan berpengaruh pada siklus hara dan proses -
proses ekosistem terkait lainnya, dan mungkin mempengaruhi produktivitas
tapak dalam jangka panjang.
Fragmentasi hutan temperate di Amerika Serikat bagian barat
dapat mengurangi atau menghilangkan populasi pengerat kecil pemakan
jamur (mycophagus) yang menjadi agen kunci penyebaran sejumlah spesies
jamur hypogeous (di bawah tanah), khususnya mychorrizae yang penting
bagi hutan konifer (Maser et al. 1978 dalam Morrison et al. 1992). Jamur-
jamur michorrizae tumbuh di rambut-rambut akar pohon konifer dan
membantu menyerap hara. Pengerat (rodent) kecil yang bertindak sebagai
agen kunci penyebaran membutuhkan tegakan hutan yang tua atau pohon
tumbang yang besar. Fragmentasi hutan dapat mengurangi ketersediaan
tegakan hutan tua tersebut dan elemen-elemen vegetasi lainnya, yang

PRAGMENTASI HUTAN
8
mungkin secara tidak langsung mengurangi produktivitas hutan dalam
jangka panjang (Morrison et al. 1992).
Fragmentasi lingkungan merupakan suatu isu perencanaan dan
konservasi habitat, yakni peningkatan fragmentasi dapat mempengaruhi
ketahanan populasi dan keragaman spesies dan komunitas. Fragmentasi
dapat mengisolasi individu, unit berkembangbiakan, dan sub populasi dari
spesies-spesies dalam patch interior. Isolasi ini dapat meningkatkan resiko
kepunahan lokal karena peningkatan variasi ukuran populasi dalam peluang
untuk hidup dan kesempatan berkembangbiak; fluktuasi di lingkungan dan
kualitas dan kuantitas sumberdaya; peningkatan kerentanan untuk punah
dari patch yang lebih kecil, lebih terisolasi jika menghadapi bencana seperti
angin badai dan kebakaran (Morrison et al. 1992).
Saat ini di banyak negara, perhatian publik terhadap perencanaan
habitat telah meningkat karena adanya peningkatan isu terkait seperti
penurunan keanekaragaman hayati, peningkatan fragmentasi lingkungan
yang dianggap berdampak buruk bagi keanekaragaman hayati dan hilangnya
hutan-hutan tua serta semakin langka dan berkurangnya habitat. Oleh
karena itu, baik dari perspektif sosial dan perhatian publik maupun
perspektif ilmiah, fragmentasi lingkungan dan pengelolaan habitat dalam
skala lanskap perlu mendapat perhatian dan dipelajari secara benar.
DAFTAR PUSTAKA
Allaby, M. 1998. Oxford Dictionary of Ecology. Oxford University Press, New
York, NY.
Bailey, J.A. 1984. Princples of Wildlife Management. John Wiley and Sons. New York.
Forman R.T.T. 1995. Land mosaics: the ecology of landscapes and regions.
Cambridge University Press, Cambridge, 632 pp.
Franklin, A.B., B.R. Noon, And T. L.George. 2002. What Is Habitat
Fragmentation? Studies in Avian Biology No. 25:20-29. http://www. humboldt. edu/-tlg2/publications/whatis
habitat20fragmentation.Pdf. Diakses Tanggal 11 Mei 2007.

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
9
Haila, Y. 2002. A conceptual genealogy of fragmentation research: from island biogeography to landscape ecology. Ecological Applications
12:321–334.
Hall, L. S., P. R. Krausman, And M. L.Morrison. 1997. The habitat concept and a plea for standard terminology. Wildlife Society Bulletin
25:173–182.
Helms, J.a. (ed). 1998. The Dictionary of Forestry. The Society of
American Forestry and CABI Publishing. Bethesda, MD and Oxon, UK.
http://www.everythingbio.com/glos/definition.php?word=fragmentation.
Diakses Tanggal 17 Oktober 2006.
http://en.wikipedia.org/wiki/Habitat_fragmentation. Diakses Tanggal 17
Oktober 2006.
http://en.wikipedia.org/wiki/Landscape ecology. Diakses Tanggal 24 Februari 2007.
http://www.carleton.ca/lands-ecol/whatisle.html. Diakses Tanggal 24
Februari 2007.
MacArthur, R. H., and E. O. Wilson. 1967. The theory of island
biogeography. Princeton University Press, Princeton, New Jersey, USA.
Morrison, M.L., B.G. Marcot and R.W. Mannan. 1992. Wildlife-Habitat
Relationships. The University of Wisconsisn. Madison, Wisconsin.
Rusak, H. and C. Dobson. 2007. Forest Fragmentation. www.ontarionature
.org. Diakses tanggal 26 Februari 2007.
Sanderson, J. and L. D. Harris (eds.). 2000. Landscape Eco logy: A Top-Down Approach. Lewis Publishers, Boca Raton, Florida, USA.
Turner, M.G. and R. H. Gardner (eds.). 1991. Quantitative Methods in Landscape Ecology. Springer-Verlag, New York, NY, USA.
Wiens, J. A. 1989. Spatial scaling in ecology. Functional Ecology 3:385–397.

PRAGMENTASI HUTAN
10
2
MASALAH DAN PROSES FRAGMENTASI
MASALAH FRAGMENTASI
PROSES FRAGMENTASI
PENGARUH FRAGMENTASI
MENGUKUR FRAGMENTASI
DAFTAR PUSTAKA
I. MASALAH FRAGMENTASI
Fragmentasi habitat merupakan masalah penting di seluruh dunia.
Penyempitan habitat secara luas memang cukup serius, tetapi ketika
dikombinasikan dengan fragmentasi maka dapat meruntuhkan kesatuan
ekosistem secara keseluruhan. Jalan, urbanisasi dan pertanian merupakan
kegiatan utama manusia yang memecah-mecah wilayah alami yang
seringkali diikuti oleh malapetaka bagi satwa. Wilayah yang sebelumnya
merupakan hutan yang kompak dan utuh menjadi terpecah-pecah,
berukuran kecil dan terisolasi, sehingga beberapa jenis satwa tidak dapat
melakukan perpindahan atau pergerakan untuk mencari makan atau untuk
berkembang biak.
Di sisi lain, ada beberapa jenis satwa dan tumbuhan yang lebih
menyukai habitat interior berupa hutan yang rapat dan gelap, maka jika
50% hutan tersebut dibabat, misal untuk membuat jalan atau lapangan
parkir dan sisanya terbelah oleh jalan, maka hutan yang lebat, sejuk dan
gelap berubah menjadi terang benderang, kelembaban dan temperaturnya
berubah dan tidak sesuai lagi sebagai habitat satwa dan tumbuhan tersebut.
Dengan demikian, fragmentasi juga meningkatkan efek tepi (edge effect).
Sejalan dengan itu area habitat interior dipengaruhi oleh kondisi berbeda

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
11
dari habitat lainnya di sekitarnya. Semakin kecil suatu habitat, semakin
besar proporsi yang terkena efek tepi dan hal ini dapat menyebabkan
perubahan yang dramatis bagi komunitas satwa dan tumbuhan.
Bila suatu populasi satwa menjadi kesulitan menyeberangi jalan
raya yang menghalanginya untuk mencapai kantong habitat lainnya yang
berjarak cukup jauh, maka satwa tersebut hanya dapat berkembang biak
terbatas di kantong habitat tempatnya tinggal yang dapat dikatakan sebagai
pulau dan populasinya akan menghadapi resiko inbreeding. Lebih jauh, jika
populasi diserang wabah penyakit atau bencana alam lainnya, maka satwa
tersebut akan mengalami kepunahan lokal dan sulit untuk rekolonisasi dari
populasi lainnya. Sejalan dengan itu, maka jelas bahwa kesinambungan
atau konektivitas dari hutan sangat penting.
Suatu hutan yang sehat dan cukup besar akan mampu mendukung
organisme dengan jelajah luas, seperti jenis-jenis satwa besar atau satwa
pemangsa. Pengurangan luas dapat memiliki dampak langsung pada spesies
ini dan karena predator sering memainkan peran penting sebagai pengatur
populasi spesies lain, maka keseimbangan ekos istem dapat sangat
terganggu. Dalam skala luas, perubahan ik lim juga mungkin memaksa suatu
spesies untuk bermigrasi, jika habitat alaminya sangat terfragmentasi,
banyak di antara mereka tidak dapat bermigrasi dan akan menghadapi
resiko kepunahan. Ini juga membuktikan betapa pentingnya suatu lanskap
yang berkesinambungan.
II. PROSES FRAGMENTASI
Fragmentasi umumnya terjadi melalui hilangnya habitat (habitat
loss), sebaliknya hilangnya habitat dapat dipandang sebagai akibat
fragmentasi. Tetapi fragmentasi dapat disertai hi langnya habitat
(berkurangnya jumlah) seiring dengan pemecahan atau pembagian kantong
habitat besar menjadi kantong-kantong habitat berukuran kecil dan lebih
terisolasi (Hunter 1997; Haila 1999; Franklin et al. 2002; Fahrig 2003). Jika
hilangnya habitat dan fragmentasi dipandang secara terpisah, maka
hilangnya habitat memiliki dampak lebih signifikan bagi kelangsungan hidup
(viability) spesies daripada fragmentasi (Haila 2002; Fahrig 2003). Namun,

PRAGMENTASI HUTAN
12
karena fragmentasi dan hilangnya habitat terjadi bersamaan maka sangat
sulit untuk menentukan mana yang lebih penting bagi perubahan habitat
(Haila 1999).
Fragmentasi bekerja dalam empat cara ketika hilangnya habitat dan
fragmentasi digabung untuk menggambarkan dan mengkategorikan
prosesnya (Franklin et al. 2002; Fahrig 2003) : (1) habitat hilang tanpa
fragmentasi; (2) pengaruh kombinasi hilangnya habitat dan pemecahan
habitat menjadi patches lebih kecil; (3) pemecahan habitat menjadi patch-
patch lebih kecil tanpa kehilangan habitat; dan (4) hilangnya habitat dan
pemecahan habitat menjadi patch-patch lebih kecil serta penurunan
kualitas habitat. Contoh ini berlaku untuk lanskap yang terdiri lebih dari
satu habitat dan dikeli lingi oleh matriks di dalam suatu kesatuan lanskap.
Kasus pertama dan kedua berlaku ketika lanskap keseluruhan berisi satu
habitat dan tidak ada matriks di sekeli lingnya. Dalam kenyataan, kasus dua
dan empat merupakan cara yang paling umum dalam fragmentasi habitat.
Fragmentasi habitat merupakan satu aspek dari tahapan proses
yang secara spasial dan temporal mengubah habitat dan lanskap yang
diakibatkan oleh sebab-sebab alami maupun antropogenik (Forman 1995).
Tetapi, perubahan habitat tidak dapat dihindari karena tidak ada habitat
atau lanskap yang tetap (Forman 1995). Lanskap berubah melalui lima
proses spasial dengan berbagai derajat overlap sepanjang periode
perubahan lahan (Forman 1995), dan fragmentasi hanyalah satu outcome.
Proses ini dapat diakibatkan oleh penyebab alami dan antropogenik.
Perforasi (Perforation) merupakan proses membuat lubang di dalam
habitat. Pemotongan (Dissection) adalah pemotongan atau pembagian area
menjadi habitat berbeda dengan lebar yang relatif sama. Fragmentasi
(Fragmentation) adalah pemecahan habitat menjadi potongan-potongan
yang lebih kecil. Penyusutan (Shrinkage) terjadi seiring potongan habitat
berlanjut dengan penurunan luas. Erosi habitat (Attrition) adalah proses
dimana kantong habitat yang tersisa berangsur hilang karena degradasi
habitat atau suksesi.
Fragmentasi dimulai dengan dissection ketika jalan, jaringan
transmisi, sungai dan fitur linear lainnya menjadi penghalang pergerakan.
Kemudian diikuti perforation ketika muncul kantong-kantong habitat

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
13
(habitat patches) kecil yang dibuat oleh manusia atau sebab alami dan efek
tepi menjadi nyata. Fragmentasi merupakan tahap ketiga yang terjadi
ketika kantong habitat yang lebih kecil meningkat frekuensinya dan
berkurang luasnya sampai pada tingkat di mana habitat yang terfragmentasi
mulai mendominasi lanskap. Attrition merupakan tahap akhir dimana lahan
alami atau habitat asli tersisa sebagai kantong yang kecil dan terisolasi di
tengah-tengah lanskap yang sekarang didominasi oleh suatu mosaik habitat
yang telah berubah dan terfragmentasi. Proses fragmentasi membuat
habitat menjadi tidak sesuai atau memiliki kesesuaian rendah bersamaan
dengan berkurangnya kualitas habitat satwaliar. Sebaliknya, jika proses
gangguan mengubah mosaik habitat tetapi tidak ada perubahan kualitas
habitat berarti tidak terjadi fragmentasi, atau habitatnya berubah tetapi
tidak terfragmentasi (Hunter 1997)
III. PENGARUH FRAGMENTASI
Ada empat cara primer fragmentasi hutan dapat mempengaruhi
keanekaragaman hayati, yaitu: (1) keterwakilan (sample effect); (2) luas
area (area effect); (3) isolasi (isolation effect) dan (4) pengaruh tepi (edge
effect) (Gambar 1). Masing-masing pada gilirannya akan berpengaruh pada
sebaran populasi, komunitas dan proses ekosistem (Kupfer et al. 2004).
Gambar 1. Model konseptual pengaruh fragmentasi (Kupfer et al. 2004).

PRAGMENTASI HUTAN
14
Mekanisme dan proses fragmentasi menghasilkan tiga tipe
pengaruh: (1) ukuran patch; (2) pengaruh tepi (edge effect); dan (3)
pengaruh isolasi (Fahrig, 2003). Ahli satwa harus memperhatikan semua
karena ketiganya terjadi pada fragmentasi habitat dan masing-masing
memerlukan penanganan yang berbeda (Franklin et al. 2002; Fahrig 2003).
Fragmentasi habitat dapat dipandang dari segi positif dan negatif.
Pengaruh positifnya adalah meningkatkan keragaman habitat, menciptakan
penjajaran habitat yang bermanfaat, dan meningkatkan edge yang disukai
spesies satwaliar generalis. Fragmentasi memberikan pengaruh negatif
ketika: (1) ada habitat yang hilang; (2) terbentuk kantong habitat lebih
kecil yang mendorong pada kepunahan lokal dan isolasi; (3) habitat-habitat
tidak lagi bersambungan, khususnya jika fragmentasi disebabkan oleh
aktifitas non kehutanan; dan (4) jumlah edge meningkat sehingga
fragmentasi habitat merugikan spesies interior (Barnes 2000).
IV. MENGUKUR FRAGMENTASI
Mengukur fragmentasi sangat menarik, ada banyak ukuran yang
dapat digunakan untuk mengkuantifikasikan fragmentasi habitat. Sebagai
contoh dalam program FRAGSTATS tersedia 100 matrix fragmentasi habitat
(McGarigal and Marks 1995), tetapi hanya sedikit kesepakatan matrik mana
yang paling sesuai dan bisa diterjemahkan ke dalam tindakan manajemen
konservasi (Garrison 2005).
Indeks-indeks struktur lanskap seringkali digunakan untuk
mengkuantifikasikan fragmentasi habitat. Indeks-indeks telah
dikembangkan untuk mengukur tiga aspek struktur lanskap: (1) komposisi
lanskap; (2) konfigurasi lanskap; dan (3) bentuk patch di dalam lanskap.
Komposisi menunjukkan jumlah dari tipe penutupan (cover) yang berbeda
yang ditemukan dalam lanskap. Konfigurasi menunjukkan bagaimana patch
dari tipe cover yang sama atau berbeda tersusun di dalam lanskap dan
hubungannya satu sama lain. Lanskap dengan komposisi sama dapat
memiliki konfigurasi yang berbeda, sehingga diperlukan beberapa indeks
aspek untuk menggambarkan suatu lanskap. Harus dicatat bahwa beberapa
indeks tidak sesuai benar untuk ketiga kategori tersebut.

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
15
Program Patch Anlayst yang kompatibel dengan ArcView 3.x cukup
handal untuk menghitung statistik fragmentasi, karena merupakan
modifikasi dari program Fragstats dan dapat digunakan untuk menghitung
statistik spasial, baik file poligon (seperti shape files) dan file raster
(seperti Arc grids) (Elkie et al., 1999). Penjelasan dan contoh aplikasi
program Patch Analyst dibahas secara terpisah pada Bab 11.
DAFTAR PUSTAKA
Barnes, T.G. 2000. Landscape Ecology and Ecosystems Management. Cooperative Extension Services, University of Kentucky, College of Agriculture. UK. http://www.ca.uky.edu. Diakses Tangal 24 Februari 2007.
Elkie, P.C., R.S. Rempel and A.P. Carr. 1999. Patch Analyst User’s Manual. Ontario Ministry of Natureal Resources, Northwest Science & Technology. Thunder Bay. Ontario.
Fahrig, L. 2003. Effects of habitat fragmentation on biodiversity. Annual Reviews of Ecology and Systematics 34:487–515.
Forman, R. T. T. 1995. Some general principles of landscape and regional ecology. Landscape Ecology 10:133–142.
Franklin, A.B., B.R. Noon, and T. L.George. 2002. What Is Habitat Fragmentation? Studies in Avian Biology No. 25:20-29. http://www. humboldt. edu/-tlg2/publications/whatis habitat20fragmentation.Pdf. Diakses Tanggal 11 Mei 2007.
Garrison, B.A. 2005. Fragmentation of Terrestrial Habitat : An Overview for Wildlife Biologists. Trans.W.Sect.Wildl. Soc. 41:2005.
Haila, Y. 2002. A conceptual genealogy of fragmentation research: from island biogeography to landscape ecology. Ecological Applications 12:321–334.
http://chesapeake.towson.edu/landscape/forestfrag/what_habitat.asp. Diakses Tanggal 1 Mei 2007.
Hunter, M. L., JR. 1997. The biological landscape. Pages 57–67 in K. A. Kohm and J. Franklin (eds). Creating a Forestry for the 21st Century. Island Press. Washington, D.C.
Kupfer, J.A., G.P. Malanson and S.B. Franklin. 2004. Identifying the Biodiversity Research Needs Related to Forest Fragmentation. A report prepared for the National Commission on Science for Sustainable Forestry (NCSSF) and funded by the National Council for Science and the Environment (NCSE).
McGarigal, K., and B. J. Marks. 1995. Fragstats: spatial pattern analysis program for quantifying landscape structure. USDA For. Serv. Gen. Tech. Rep. PNW-351. http://www. innovativegis.com/basis/Supplements/BM_Aug_99/FRAG_expt.htm. Diakses Tanggal 12 April 2006.

PRAGMENTASI HUTAN
16
3
TEORI BIOGEOGRAFI PULAU vs FRAGMENTASI HUTAN
PENDAHULUAN
TIPE-TIPE PULAU
HUBUNGAN SPESIES-AREA
KEPUNAHAN LOKAL DAN PERUBAHAN KOMUNITAS
EFEK ISOLASI
PERBEDAAN ANTARA BIOGEOGRAFI PULAU SEJATI DENGAN PULAU HABITAT
DAFTAR PUSTAKA
I. PENDAHULUAN
Teori Biogeografi pulau pertama dikemukakan oleh Mac Arthur dan
Wilson (1967). Dalam teorinya mereka memprediksi jumlah spesies yang
mungkin akan bertahan pada suatu pulau yang baru ter bentuk. Dalam
biogeografi pulau dipelajari dan dijelaskan faktor-faktor yang
mempengaruhi keragaman spesies dari sauatu komunitas tertentu. Dalam
konteks ini, “pulau” dapat berupa areal habitat yang dikeli lingi oleh areal
lain yang tidak sesuai untuk spesies dalam “pulau” tersebut; bukan hanya
pulau sesungguhnya yang dikeli lingi lautan, tetapi juga gunung yang
dikeli lingi oleh gurun pasir, danau yang dikelilingi daratan, dan fragment
hutan yang dikeli lingi oleh lanskap yang terganggu oleh manusia.
Model biogeografi pulau adalah suatu model kaidah umum mengenai
penyebaran keanekaragaman hayati yang menjelaskan hubungan antara
luas areal dan jumlah spesies (species-area relationship). Pulau-pulau yang
luas memiliki jumlah spesies yang lebih banyak dari pada pulau-pulau kecil.
Hal ini karena pulau-pulau yang lebih luas biasanya memiliki tipe habitat

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
17
dan komunitas yang lebih banyak, menyediakan kemungkinan isolasi
geografis dan jumlah populasi yang lebih banyak untuk setiap spesies
sehingga memperbesar kemungkinan spesiasi dan memperkecil
kemungkinan kepunahan dari spesies yang baru terbentuk atau dari spesies
yang baru datang (Primack et al. 1998). Model biogeografi pulau
memperkirakan jika 50% dari wilayah pu lau rusak, sekitar 10% spesies yang
hidup di pulau tersebut akan punah. Apabila spesies ini endemik di wilayah
tersebut, maka spesies akan punah. Apabila 90% dari habitat rusak, pulau
akan kehilangan 50% spesiesnya, dan jika 99% habitatnya hilang, maka 75%
spesies alami akan hilang (Primack et al. 1998).
Biogeografi pulau dapat diaplikasikan pada fragmentasi lanskap
sebagai model bagaimana “pulau-pulau” habitat kecil dapat berpengaruh
buruk pada keragaman hayati habitat as linya (Harris 1984). Teori ini sangat
penting dalam mendisain kawasan konservasi karena memberikan panduan
kuantitatif tentang luas kawasan dan kesinambungan antara kawasan
konservasi yang bertetangga, berdasarkan karakteristik ekologi di wilayah
tersebut (Diamond 1975).
Menurut teori biogeografi pulau (Mac Arthur & Wilson 1967),
kekayaan spesies suatu pulau bergantung pada:
Isolasi pulau, karena isolasi mempengaruhi laju kolonisasi. Pulau yang
terisolasi atau jauh, memiliki spesies yang lebih sedikit dari pada pulau
yang dekat dengan sumber spesies yang mengkolonisasi. Pulau yang
lebih jauh, lebih sedikit didatangi pengkoloni dibandingkan pulau yang
lebih dekat. Jika ada pulau-pulau di antara sumber kolonisasi (daratan
utama) dengan pulau, maka dapat berperan sebagai batu loncatan
(stepping stones) dan dapat meningkatkan laju kolonisasi pulau yang
jauh. Jika ada dua pulau dengan jarak yang sama dari sumber
kolonisasi, maka pulau yang lebih besar akan memiliki laju kolonisasi
yang lebih tinggi karena adanya Target Effect (The bigger targets are
easier to hit).
Luas pulau, karena luas pulau mempengaruhi laju kepunahan. Pulau
yang besar memiliki jumlah spesies yang lebih banyak dibandingkan
pulau yang kecil. Pulau kecil memiliki ukuran populasi yang lebih kecil,

PRAGMENTASI HUTAN
18
lebih sedikit refugia (area untuk mengungsi) dan memiliki laju
kepunahan lebih tinggi.
Dinamika kolonisasi dan kepunahan, kolonisasi menggantikan spesies
yang punah (species turnover).
Kekayaan spesies mencerminkan suatu keseimbangan (equilibrium)
antara kolonisasi dan kepunahan.
II. TIPE-TIPE PULAU
Pulau adalah daratan yang dikeli lingi air. Kondisi fisik pulau
biasanya sempit, tipe habitatnya sedikit, sedangkan pengaruh laut sangat
besar. Pulau digunakan oleh para ahli ilmu pengetahuan alam sebagai
laboratorium lapangan untuk menelaah masalah biogeografi. Darwin dan
Wallace membedakan antara pulau benua (continental island) dan pulau
laut (oceanic island) (Haris 1984). Pulau laut adalah pulau yang belum
pernah berhubungan dengan daratan lainnya (misalnya : P. Nias, P.
Bawean, P. Natuna, P. Belitung, P. Sumba dan P. Aru). Pulau Benua adalah
pulau yang pada masa lampau mempunyai hubungan dengan darata n atau
benua lainnnya karena turunnya permukaan laut (misalnya : Pulau Sumatra,
P. Jawa, P. Kalimantan, P. Papua, P. Simeulue, P. Enggano, P. Buru, P. Kai
dan P. Tanimbar) (Primack et al. 1998).
Ahli lain membagi pulau menjadi tiga tipe yaitu (Ripley 1985):
1. Pulau benua (continental island) yang dulunya merupakan bagian
daratan benua tetapi sekarang terpisah dari daratan induknya, ciri -ciri
kehidupan serupa dengan benua/daratan utamanya dan dekat dengan
benua.
2. Pulau gunung berapi (volcanic island) yang berasal dari gunung berapi
dan tidak terbentuk sebagai bagian benua. Pulau-pulau gunung berapi
berada di dekat pulau-pulau besar membentuk busur pulau gunung
berapi di laut. Pulau-pulau gunung berapi di bagian sisi yang
menghadap benua biasanya dikeli lingi perairan dangkal sedangkan di
tepi luarnya selalu mempunyai tebing yang terjal yang turun ke bagian
laut paling dalam.

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
19
3. Pulau samudera (oceanic island) sejati yang terbentuk dari proses
geologi. Biasanya berukuran kecil dan seringkali letaknya jauh dari
daratan utama. Pulau ini dapat berbentuk pulau tunggal atau
membentuk kepulauan kecil. Suksesi yang terjadi di pulau ini adalah
suksesi primer
Disamping ketiga tipe pulau tersebut juga ada tipe pulau lain yaitu
(Whitten et al., 1988):
1. Pulau atol. Pulau atol terbentuk dari binatang karang yang telah mati,
berukuran kecil, biasanya berbentuk cincin dan banyak terdapat di
Lautan Pasifik (Contoh : Pulau Takabonerate dan Tukang Besi).
2. Pulau purba (ancient island). Pulau purba telah terpisah dari daratan
utama sejak lama sekali (lebih dari 100 juta tahun) oleh karenanya
memiliki tingkat endemisme tinggi (misalnya P. Madagaskar dan
Kepulauan Selandia Baru).
Pulau-pulau benua memperoleh keanekaragaman hayatinya ketika
pulau-pulau tersebut berhubungan dengan benua. Selanjutnya pulau-pulau
ini memperoleh tambahan dari hasil migrasi hewan/tumbuhan (Primack et
al. 1998). Pulau laut asal mulanya merupakan substrat kosong, komunitas
hewan berkembang dari kolonisasi awal yang kemudian berkembang
semakin kaya dan kompleks (Haris 1984). Dengan perkataan lain, pulau-
pulau laut hanya dapat memperoleh keanekaragaman hayatinya dari
kemampuan migrasi hewan dan tumbuhan yang berasal dari tempat-tempat
lain (Primack et al. 1998). Laju imigasi dan kolonisasi serta faktor-faktor
lingkungan laut sangat mempengaruhi karakteristik biota pulau-pulau laut,
setidakya pada tingkat hunian awalnya (Haris 1984).
III. HUBUNGAN SPESIES-AREA
Menurut teori biogeografi, jumlah spesies (jumlah equilibrium)
yang terdapat di suatu pulau ditentukan oleh dua faktor, yaitu jarak dari
daratan utama dan ukuran pulau. Keduanya akan mempengaruhi laju
kepunahan di pulau dan tingkat imigrasi. Pulau-pulau yang dekat dengan
daratan utama kemungkinan menerima imigran dari daratan utama lebih
besar dari pada pulau-pulau yang jauh dari daratan utama. Pada pulau-

PRAGMENTASI HUTAN
20
pulau yang lebih kecil peluang kepunahan lebih besar dari pada pulau-pulau
besar. Pulau-pulau besar memiliki jumlah spesies lebih banyak dari pada
pulau-pulau kecil (Mac Arhtur & Wilson 1967).
Kekayaan spesies suatu pulau juga tergantung pada: (1) isolasi,
karena isolasi mempengaruhi laju kolonisasi dan (2) ukuran pulau karena
ukuran pulau mempengaruhi laju kepunahan. Kolonisasi dan kepunahan
adalah dinamis, kolonisasi menggantikan spesies yang punah, atau disebut
turn over. Kekayaan spesies suatu pulau menunjukkan keseimbangan
(equilibrium) antara kecepatan kolonisasi dan kecepatan kepunahan (Mac
Arhtur & Wilson 1967).
Hubungan antara jumlah spesies dan luas pulau digambarkan
dengan rumus sebagai berikut (Mac Arhtur & Wilson 1967):
Dimana S = jumlah spesies, A = luas pulau, z dan c adalah konstanta yang
diperlukan untuk menyesuaikan data luas (dalam m2, km2, dll) dengan
jumlah spesies. Hubungan ini dapat dibuat linier dengan menggunakan log
sehingga rumusnya menjadi (Mac Arhtur & Wilson 1967):
Log S = Log c + z log A
Laju kolonisasi lebih tinggi pada pulau yang dekat dengan daratan
utama karena lebih banyak spesies yang dapat menyeberang laut yang
relatif dekat. Laju kepunahan lebih besar pada pulau yang lebih kecil
karena populasi-populasi berukuran lebih kecil dan kemungkinan terkena
penyakit dan kejadian merugikan lainnya yang dapat menghabiskan populasi
atau menurunkannya sampai tingkat yang tidak viable. Hubungan ini
merupakan prinsip dasar teori biogeografi pulau seperti dtunjukkan pada
Gambar 2.
Hubungan antara ukuran pulau dan jumlah jenis relatif konstan
untuk kelompok-kelompok hewan atau tumbuhan. Secara umum
berkurangnya sepuluh kali ukuran pulau mengurangi setengah jumlah jenis.
Jika suatu pulau mendukung lebih sedikit jenis dari yang diharapkan sampai
di bawah garis (Gambar 2), mungkin disebabkan oleh (Whitten et al. 1988):
1. Kelompok tersebut belum diketahui dengan baik.

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
21
2. Keseimbangan jumlah jenis belum tercapai.
3. Pulau terdiri atas jumlah habitat yang relatif terbatas atau habitat
tidak mendukung jumlah jenis yang banyak.
4. Pulau sangat jauh dan sulit dikolonisasi.
Jika suatu pulau mendukung lebih banyak jenis dari pada yang
diharapkan sampai di atas garis (Gambar 2), mungkin disebabkan oleh
(Whitten et al. 1988):
1. Jumlah melebihi keseimbangan, namun beberapa jenis pada waktunya
nanti akan hilang.
2. Pulau sangat kaya akan tipe-tipe habitat.
3. Pulau merupakan pusat penyebaran jenis kelompok tertentu.
Gambar 2. Jumlah jenis relatif pada (a) pulau-pulau kecil, dekat, (b)
pulau-pulau besar, jauh atau kecil, dekat, dan (c) pulau-pulau besar, dekat (Mac Arthur and Wilson 1967).
Laju Imigrasi Laju Kepunahan
Jumlah spesies pada sebuah pulau
Jauh
Dekat
Kecil
Besar
a b c

PRAGMENTASI HUTAN
22
Sebagai contoh, untuk total jenis, Sulawesi berada di bawah garis
dari jumlah yang seharusnya untuk hewan dan tumbuhan, tetapi berada di
atas garis untuk mamalia, hal ini mungkin disebabkan oleh sebaran jenis-
jenis tikus yang luar biasa dan juga ular. Sulawesi juga berada di atas garis
untuk jenis endemik, hal ini disebabkan oleh sejarah geologinya (Whitten et
al. 1988).
Pengaruh ukuran pulau pada tingkat endemisitas juga sangat tinggi.
Pulau-pulau besar mempunyai jumlah spesies dengan tingkat endemisitas
yang jauh lebih tinggi, tetapi korelasinya tidak begitu tampak dengan
kekayaan spesies. Spesies endemik berkorelasi negatif dengan kekayaan
spesies dan berkorelasi positif dengan tingkat isolasi. Disamping itu juga
ada perbedaan antara tumbuhan dan hewan dalam tingkat endemisitas.
Tingginya jumlah hewan endemik tidak selalu diikuti oleh tingginya
tumbuhan endemik, tetapi seringkali tingginya tumbuhan endemik diikuti
oleh tingginya hewan endemik (Primack et al. 1998).
Tumbuhan endemik lebih terpengaruh oleh ukuran pulau dari pada
isolasi geografik. Sebaliknya endemisitas burung sangat bergantung pada
isolasi geografik. Pulau-pulau yang jauh dan kecil dapat mempunyai tingkat
endemisitas burung yang tinggi, tetapi tingkat endemisitas tumbuhannya
dapat rendah (Primack et al. 1998).
IV. KEPUNAHAN LOKAL DAN PERUBAHAN KOMUNITAS
Istilah pulau bisa diartikan mulai dari sebuah pulau kecil, pulau
karang (atol) terpencil yang dikeli lingi laut sampai pada kawasan hutan
yang ditetapkan secara hukum. Suatu komunitas alami yang ada sebagai
sebuah pusat dari suatu habitat regional yang lebih besar akan berisi banyak
spesies langka yang tergantung pada sistem yang lebih besar untuk
eksistensinya. Seiring dengan cepatnya perubahan penggunaan lahan di
sekitarnya, maka akan tercipta suatu kantong pulau habitat (habitat patch)
yang terisolasi. Seiring dengan pulau habitat yang semakin terisolasi
dengan cepat dari vegetasi di sekitarnya yang serupa, spesies langka dengan
cepat akan hilang.

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
23
Kepunahan mengurangi satwa di setiap pulau habitat sehingga
memainkan peran penting dalam menentukan komposisi spesies. Tidak
adanya kompensasi kolonisasi, proses tersebut menghasilkan kemiringan
(slope) yang lebih tajam pada kurva species-area. Dengan demikian
perbedaan nilai kemiringan antara pulau-pulau samudera dan pulau-pulau
benua dapat dijelaskan oleh ketidakseimbangan relatif dari proses
kepunahan dan kolonisasi (Harris 1984).
Picton (1979) dalam Harris (1984) menganalisis perubahan populasi
10 spesies mamalia besar pada 24 wilayah pegunungan kontinental yang
semi terisolasi di Rocky Mountains bagian utara. Luas arealnya bervariasi
dari 11 – 4.480 mil persegi (29 – 11.600 km2). Persentasi spesies asli yang
telah hilang selama periode pembagunan pemukiman, pertanian dan
peternakan ranch, perburuan subsisten, pembalakan dan penambangan
berbanding terbalik dengan ukuran luas areal. Sementara areal yang lebih
kecil kehilangan lebih dari 50% spesies aslinya, areal yang lebih besar
kehilangan sekitar 4% dari spesies yang pernah ada (Picton 1979 dalam
Harris 1984). Dengan perkataan lain, faktor-faktor yang meningkatkan
perbedaan antara kepunahan dan kolonisasi menyebabkan kemiringan
hubungan spesies-area meningkat seperti ni lai yang ditunjukkan oleh pulau-
pulau sebenarnya.
Kepunahan lokal atau hilangnya spesies dari taman nasional, cagar
alam dan pulau-pulau habitat mempengaruhi kelompok spesies tertentu
lebih dari lainnya. Secara umum, spesies yang paling rawan untuk hilang
ditunjukkan oleh populasi yang kecil. Populasi kecil dapat dihasilkan oleh
keterbatasan sumberdaya spasial dan temporal (seperti habitat yang sangat
spesialis atau sumberdaya pakan), eksploitasi berlebih atau areal yang
relatif lebih kecil dari pada daerah jelajah spesies tersebut. Spesies yang
sangat spesialis, yang menunjukkan jumlah terbesar dari kelas makanannya
(seperti raptor, karnivora darat, pemakan serangga di batang pohon) dan
yang berada pada tropic level yang lebih tinggi biasanya yang pertama
mengalami kepunahan. Walaupun spesies dengan jelajah luas memiliki
potensi kolonisasi yang lebih tinggi, laju kepunahan lokalnya lebih tinggi
dari rata-rata untuk semua spesies dalam komunitas (Harris 1984).

PRAGMENTASI HUTAN
24
Untuk berbagai alasan, fenomena yang digambarkan di atas sangat
penting untuk mamalia. Beruang grizzly, srigala abu-abu, dan fisher telah
hilang dari Oregon Barat dan wolverine (sejenis anjing hutan) dan Lynx
menjadi sangat jarang. Semua jenis tersebut adalah karnivora puncak.
Olterman & Verts (1982) dalam Harris (1984) meninjau status 41 spesies
mamalia di Oregon. Tujuh dari 8 spesies (88%) yang pernah ada di fragment
bagian barat dan dinyatakan ”punah”, ”jarang” atau ”terancam punah
(endangered)” adalah karnivora. Sebaliknya, lima dari delapan spesies
(62%) dalam kategori ”tidak jarang atau tidak endagered” adalah herbivora.
Picton (1979) dalam Harris (1984) membatasi analisisnya untuk wialyah
Rocky Mountains pada herbivora besar karena karnivora sudah sangat serius
menurun sehingga analisisnya tidak dapat dijamin. Di Great Basin, spesies
herbivora dengan kebutuhan habitat yang umum (generalis) dan berukuran
tubuh kecil sampai sedang masih bertahan pada sebagian besar pulau
habitat, herbivora dengan ukuran tubuh besar dan/atau memerlukan
habitat khusus (spesialis) serta karnivora memiliki laju kepunahan yang
lebih tinggi dan hanya bertahan di sedikit areal dari 19 pulau habitat
(Brown 1978 dalam Harris 1984).
V. EFEK ISOLASI
Baik komunitas tropika maupun temperate sama-sama menghadapi
masalah inbreeding dan hilangnya keragaman genetik yang disebabkan oleh
sub populasi tumbuhan dan satwa yang terisolasi satu sama lain akibat
fragmentasi. Jika jarak yang memisahkan antara dua fragment terlalu
besar dan suatu spesies tidak dapat menyeberanginya, populasi di kedua
fragment tersebut benar-benar terpisahkan. Inbreeding biasa terjadi jika
sub populasi di fragment tersebut kecil. Walaupun hal ini belum ada
catatannya tetapi potensial terjadi (Harvey and Lyles 1986).
Hilangnya keragaman genetik dapat terjadi bahkan tanpa
inbreeding, dan homozigositas yang dihasilkan dalam gen-gen tertentu
dapat membawa suatu spesies pada kepunahan secara perlahan
(evolusioner) (Soule 1986). Untuk meningkatkan pertukaran antar
fragment, banyak pengelola satwa memanfaatkan koridor yang
menghubungkan dua atau lebih pulau habitat. Koridor meningkatkan

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
25
kekayaan spesies burung-burung yang sedang berkembangbiak (MacClintock
et al. 1977) dan meningkatkan mobilitas satwa penyebar biji (Haris 1984).
Koridor mungkin bisa menghambat hilangnya keragaman genetik dan
memungkinkan penyebaran spesies antar fragment, tetapi pengurangan
ekosistem alami ke dalam fragment-fragment terus saja terjadi. Penurunan
luas ekosistem alami ini hanya dapat merugikan spesies yang bergantung
padanya untuk bertahan hidup.
Salah satu prinsip biogeografi yang telah lama dianut adalah
pentingnya isolasi dalam menentukan karakteristik komunitas biotik.
Walaupun jarak dianggap sebagai ukuran utama dari isolasi, tetapi
konsepnya jauh lebih kompleks. Mungkin ukuran jarak absolut lebih mudah
dipahami oleh pengelola, tetapi kenyataannya tidak selalu demikian karena
yang dihadapi oleh spesies-spesies tumbuhan dan satwa berbeda. Ada
ribuan perbedaan antara jarak tempuh normal rutin bulanan antara
mamalia kecil seperti mole (sejenis tikus mondok) dan beruang grizzly atau
srigala. Dengan demikian jarak 0,6 mil bisa 75 kali radius jelajah seekor
tikus mondok, tetapi hanya 1% dari radius jelajah seekor coughar (Harris
1984).
Alasan kedua mengapa ukuran jarak absolut tidak begitu menjadi
perhatian, karena perilaku menetap (sedentary) versus berpindah-pindah
(migratory) setiap spesies berbeda. Suatu spesies migratory dengan
anggota yang menjelajahi ribuan kilometer secara musiman memiliki
peluang lebih tinggi untuk mengkolonisasi pulau terisolasi dalam jarak
tempuhnya dari pada penghuni yang memiliki sifat menetap. Untuk spesies
seperti pika, jarak 1.000 feet (300 m) antara singkapan talus merupakan
rintangan yang sulit dan sangat tidak mungkin menyebar dengan jarak lebih
dari beberapa kilometer dari lingkungannya (Smith 1974 dalam Harris
1984).
Pertimbangan ketiga adalah spesialisasi habitat dan toleransi
terhadap variasi klimatik, edafik dan perubahan (gradient) vegetasi.
Spesies tertentu (misalnya beruang hitam) menjelajah seluruh wilayah
ketinggian, seluruh gradient kelembaban dan seluruh tahapan suksesi
vegetasi. Spesies lain seperti red tree vole (sejenis tikus kecil) mungkin
menghadapi rintangan besar yang tak terlihat (Grinnell 1941b dalam Harris

PRAGMENTASI HUTAN
26
1984), hanya ada di tajuk beberapa spesies konifer (kecuali dalam situasi
tidak umum), dan hanya terbatas di Oregon bagian barat dan California
Barat Laut. Dengan demikian tebang habis yang luas dan hutan sekunder,
bagi spesies tertentu bisa tidak berpengaruh tetapi bisa mengisolasi secara
total bagi spesies lainnya (Harris 1984).
Derajat isolasi harus dilihat sebagai suatu kontinum bagi spesies
secara spesifik dan harus dilihat dari aspek biologi spesies tersebut dan
kondisi lingkungannya. Findley & Anderson (1956) dalam Harris (1984)
menyatakan bahwa spesies arboreal seperti tupai dan spesies lain seperti
marten dan fisher yang sangat tergantung pada habitat hutan, akan sangat
terbatasi oleh sungai besar dan ngarai. Mereka juga menyatakan bahwa
penyebaran spesies mamalia di Colorado Rockies merupakan kebalikan dari
ketergantungannya pada hutan. Keberadaan hutan riparian yang dapat
digunakan sebagai koridor penyebaran sangat penting bagi penyebaran
banyak spesies (Findley & Anderson 1956 dalam Harris 1984).
Pengaruh tidak langsung tambahan dari isolasi adalah sangat
penting bagi satwa, karena sebaran tumbuhan di habitat terisolasi juga
masalah bagi sebaran satwa. Isolasi mungkin memiliki pengaruh
mengurangi keragaman sumberdaya tumbuhan dan kualitas habitat
(Johnson 1975 dalam Harris 1984). Dengan demikian meskipun suatu
spesies satwa dapat mencapai suatu pulau habitat, habitat tersebut
mungkin tidak cukup beragam dan kompleks untuk mendukung spesies
tersebut. Johnson (1975) dalam Harris (1984) menyimpulkan bahwa
pemiskinan spesies burung dan kerapatan yang lebih rendah spesies di
Pegunungan Great Basin diakibatkan oleh pemiskinan tumbuhan dan
serangga yang merupakan makanan dan habitatnya.
Pengaruh gabungan dari semua faktor tampaknya bekerja secara
signifikan pada amfibia dan reptilia, diikuti oleh mamalia, burung penghuni
tetap dan burung migran. Dengan kata lain, burung secara proporsional
lebih melimpah di komunitas terisolasi seperti Mary’s Peak dan Steens
Mountains dari pada rata-rata di habitat terfragmentasi lainnya. Isolasi
juga kurang penting untuk menentukan di tipe habitat apa burung berada di
pulau-pulau habitat pegunungan tetapi penting untuk menentukan

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
27
keberadaan mamalia (Brown 1978 dalam Harris 1984). Secara grafis, efek
isolasi diperlihatkan pada Gambar 3.
VI. PERBEDAAN ANTARA BIOGEOGRAFI PULAU SEJATI DENGAN PULAU HABITAT
Disamping kemiripan antara biogeografi pulau samudera sejati
dengan pulau-pulau habitat (habitat islands) dan pegunungan, terdapat
beberapa perbedaan yang harus dijelaskan (Wilcox 1980 dalam Harris
1984). Pulau-pulau sejati dikeli lingi oleh media yang bisa berfungsi sebagai
agen penyebaran tetapi tidak akan cukup sebagai habitat spesies daratan.
Puncak pegunungan Mesic dikelilingi oleh lingkungan yang sangat ekstrim
perbedaannya seperti gurun pasir bisa menjadi analogi baik sebagai
perbandingan. Tebang habis secara total di sekeli ling suatu pulau tua juga
berbeda dari habitat tua yang tidak sesuai bagi penghuni yang lama tinggal
disana. Hal ini digunakan untuk membenarkan analogi, tetapi analogi ini
tidak memiliki konsistensi kuat (Harris 1984).
Gambar 3. Mekanisme bekerjanya pengaruh isolasi.
Perbedaan kedua antara pulau-pulau sejati dan pulau-pulau tua
masa mendatang bisa memiliki konsekuensi yang lebih berat. Pulau-pulau

PRAGMENTASI HUTAN
28
sehati selalu dianggap dekat dengan benua yang berperan sebagai sumber
imigran. Benua secara implisit berisi sejumlah besar dan pemasok jenis-
jenis potensial yang tak pernah habis menjadi pengkoloni. Imigrasi dan
kolonisasi dari sumber spesies selalu mungkin. Tetapi, beberapa dekade
mendatang ketika hutan terdiri dari tanaman monokultur rotasi pendek dan
ketika banyak spesies terbatasi hanya di cagar alam yang terpisah jauh,
maka tidak ada “benua” sumber spesies yang dapat mengkoloni. Sama
halnya, tidak akan ada benua sumber spesies untuk memperkaya pulau-
pulau tua. Seperti halnya Burgess & Sharpe (1981) dalam Harris (1984) yang
menganggap pulau-pulau tua akan terjadi di sebuah lautan tanpa “benua”
di Amerika Serikat Bagian Timur. Pulau-pulau tersebut menggambarkan
kantong-kantong habitat yang harus berperan sebagai satu-satunya sumber
spesies. Tidak akan ada lagi cadangan spesies yang dapat menjadi sumber
imigrasi (Harris 1984).
DAFTAR PUSTAKA
Amos, W.H. 1980. Wildlife of Islands. H.arry N. Abrams, Incorporated. New York.
Brown, J.H. and M.V. Lomolino. 1998. Biogeography. Sinauer Associates,
Inc. Sunderland, Massachusetts.
Diamond, J.M. 1975. The Island Dillema : Lessons of Modern Biogeograpgic
Studies for the Design of Natural Reserves. Biological Conservation 7:129-146.
Haris, L.D. 1984. The Fragmented Forest : Island Biogeography Theory and the Preservation of Biotic Diversity. The University of Chicago
Press. Chicago.
Harvey, P.H. and A.M. Lyles. 1986. Inbreeding in natural populatio n of
birds and mammals. In M.E. Soule (ed). Conservation Biology : The Science of Scarcity and Diversity. Sinauer Assoc. Inc., Sunderland,
M.A.
http://learning.turner.com/efts/rforest/habfrag.htm. Eco-Link Habitat
Fragmnentation. Diakses Tanggal 17-10-2006.
http://www.abdn.ac.uk/zoohons/lecture4/sld017.htm. Diakses Tanggal 6-
11-2007.

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
29
Mac Arthur, R.H. and E.O. Wilson. 1967. The Theory of Island Biogeography. Princeton University Press. Princeton, New Jersey.
MacClintock, L., R.F. Whitcomb and B.L. Whitcomb. 1977. Island Biogeography and Habitat Island of Eastern Forests. II. Evidence
for the Value of Corridors and Minimization of Isolation in Preservation of Biotic Diversity. American Birds 31: 6-16.
Primack, R.B., J. Supriatna, M. Indrawan dan P. Kramadirata. 1998. Biologi Konservasi. Yayasan Obor Indonesia. Jakarta.
Ripley, S.D. 1985. Tropical Asia. Time-Life Books Inc. U.S.A.
Whitten, A.J., M. Mustafa dan G.S. Henderson. 1988. The Ecology of
Sulawesi. Gadjahmada University Press. Yogyakarta.
Wikipedia. 2007. Island Biogeography. http://en.wikipedia.org/
wiki/Island_ biogeography. Diakses Tanggal 30-01-2007.
WWF. 1973. The Atlas of World Wildlife. Portland House. Barcelona.

PRAGMENTASI HUTAN
30
4
FRAGMENTASI HABITAT DIPANDANG DALAM SKALA LANSKAP
DINAMIKA PATCH
DESKRIPSI POLA PATCH
PENGARUH UKURAN PATCH
PENGARUH FAKTOR-FAKTOR LAIN
DAFTAR PUSTAKA
I. DINAMIKA PATCH
Kejadian (occurance) dan distribusi patch-patch dalam suatu
lanskap adalah tidak statis. Mereka terus berubah sepanjang waktu
dibawah kekuatan sistematis seperti suksesi dan erosi serta di bawah
kekuatan bencana alam seperti badai dan gangguan jangka pendek oleh
manusia. Gangguan bencana alam umumnya menciptakan patch-patch
dengan suksesi awal di dalam suatu lanskap dan mengurangi patch-patch
dengan suksesi lebih tua yang sudah ada. Hal ini merupakan ciri khas
proses dimana hutan yang sudah matang terfragmentasi menjadi blok-blok.
Gangguan bencana alam juga mempengaruhi ukuran patch, biasanya
meningkatkan patch-patch dengan suksesi baru dan mengurangi ukuran
patch-patch dengan suksesi lama. Tetapi beberapa gangguan bisa lebih
berpengaruh di dalam patch itu sendiri dari pada antar patch.
Heterogenitas dalam patch vegetasi dapat meningkat dengan membuka
kanopi, menciptakan pohon tumbang, dan meningkatkan pancaran cahaya
dan panas ke strata kanopi yang lebih rendah dan permukaan tanah.
Gangguan mengubah rejim energi dan aliran hara di dalam lanskap,

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
31
walaupun juga dapat meningkatkan atau menurunkan aliran-aliran tertentu
tergantung pada tipe gangguannya (Morrison et al. 1992).
Shugart (1984) mengusulkan suatu cara untuk mengklasifikasi
gangguan berdasarkan pada hubungan antara luas lanskap dan luas yang
terganggu (Gambar 4). Sistemnya mengkarakterisasi gangguan sebagai
rentang dari pohon kecil tumbang yang mempengaruhi kurang dari 1.000 m2
dalam hutan individual 100 ha sampai kebakaran hutan yang mempengaruhi
lebih dari 100 ha di hutan wisata lebih dari 10.000 ha atau badai yang
mempengaruhi 1.000.000 ha atau lebih dari suatu pulau. Sughart
mengusulkan bahwa kondisi kesetimbangan dalam suatu lanskap yang
sedang terganggu memerlukan sesuatu kurang lebih luas lanskap mendekati
50 kali dari ukuran gangguan. Dengan demikian, suatu kumpulan pohon
kecil dapat menyerap gangguan pohon tumbang, tetapi luas hutan nasional
atau taman nasional diperlukan untuk menyerap kebakaran acak untuk
menjaga kesetimbangan lanskap. Pada lanskap yang tidak dalam
kesetimbangan, dimana gangguan bervariasi ukurannya, mungkin diperlukan
oleh manajer untuk mengubah skala gangguan atau untuk meningkatkan
luas di bawah manajemen dalam rangka memberikan kondisi gangguan
kesetimbangan (Morrison et al. 1992).
Cara lain untuk mengkarakterisasi gangguan adalah dengan derajat
perubahan lingkungan. Empat tipe gangguan umum dapat digambarkan
dalam cara ini (Gambar 5). Gangguan Tipe I adalah bencana lingkungan
seperti gunung meletus, kebakaran luas yang berdampak pada skala
geografis luas. Gangguan Tipe II adalah bersifat lebih lokal dan meliputi
pengaruh topan, serangga dan penyakit. Gangguan Tipe III tersebar luas
tetapi relatif berdampak rendah per unit waktu dan meliputi pengaruh
kronik dan sistematik dari predator dan kompetitor dan perubahan gradual
dalam penggunaan lahan dalam suatu wilayah yang luas. Gangguan Tipe IV
adalah bersifat setempat dan umumnya berdampak rendah dan meliputi
perubahan kecil pada lingkungan yang disebabkan oleh kebakaran lokal dan
pembangunan oleh manusia (Morrison et al. 1992).
Kebanyakan perhatian tentang dampak dari fragmentasi hutan
diturunkan dari pengurangan dan isolasi habitat lokal dari gangguan Tipe I
dan Tipe IV. Tetapi dampak sekunder dari fragmentasi seperti

PRAGMENTASI HUTAN
32
meningkatnya pemangsaan atau spesies kompetitor, terjadi pada skala
sebaran yang lebih luas dan sulit untuk diramalkan dan dimonitor secara
lokal. Sementara, para perencana ingin melacak dampak kumulatif dari
gangguan Tipe IV. Dampak kumulatif ini mungkin bisa menjadi gangguan
Tipe III pada wilayah yang lebih luas. Dampak yang menyebar lebih luas ini
harus dipelajari sebagai bagian dari program monitoring fragmentasi atau
penelitian (Morrison et al. 1992).
Gambar 4. Klasifikasi rejim gangguan berdasarkan pada luas yang
terganggu dan luas lanskap (diadaptasi oleh Shugart, 1984; dari Shugart & West 1981).
Gambar 5. Empat tipe gangguan yang ditunjukkan oleh derajat gangguan dan luas geografis yang dipengaruhinya (Morrison et al. 1992).
Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
33
Kondisi lingkungan mencirikan edges dari patch-patch yang tersisa –
sinar matahari langsung dan sering serta fluktuasi temperatur dan
kelembaban yang besar – sering masuk ke suatu patch. Dengan demikian,
ukuran patch yang secara ekologis efektif, dimana sejumlah lingkungan
interior yang sebenarnya adalah lebih kecil dari pada ukuran fisik patch.
Demikian juga bentuk patch dipengaruhi oleh fragmentasi. Gangguan dan
fragmentasi bisa menciptakan edges, islands, rings, peninsulas dan bentuk
patch-patch lainnya yang tidak kondusif bagi spesies interior tetapi
menguntungkan spesies yang menyukai suksesi edges dan spesies suksesi
awal. Pada umumnya, jumlah dan konfigurasi patch-patch dan
perubahannya dari waktu ke waktu bervariasi menurut kondisi gangguan
yang berbeda (Morrison et al. 1992).
Kita juga dapat mengkarakterisasi fragmentasi lingkungan menurut
skala spasial dimana terjadinya serta menurut ukuran dan pola penggunaan
areal oleh spesies satwaliar. Lord dan Norton (1990) memandang
fragmentasi sebagai suatu diskontinuitas gradient lingkungan dan skalanya
bebas. Mereka menyarankan agar sebaran patch-patch dipandang dalam
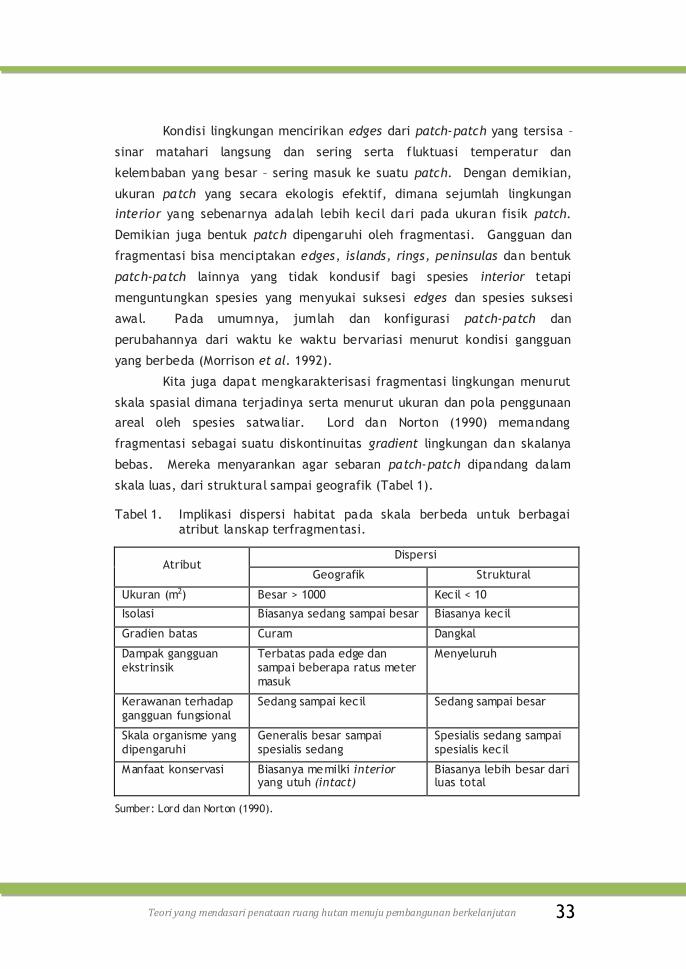
skala luas, dari struktural sampai geografik (Tabel 1).
Tabel 1. Implikasi dispersi habitat pada skala berbeda untuk berbagai atribut lanskap terfragmentasi.
Atribut Dispersi
Geografik Struktural
Ukuran (m2) Besar > 1000 Kecil < 10
Isolasi Biasanya sedang sampai besar Biasanya kecil
Gradien batas Curam Dangkal
Dampak gangguan ekstrinsik
Terbatas pada edge dan sampai beberapa ratus meter masuk
Menyeluruh
Kerawanan terhadap gangguan fungsional
Sedang sampai kecil Sedang sampai besar
Skala organisme yang dipengaruhi
Generalis besar sampai spesialis sedang
Spesialis sedang sampai spesialis kecil
Manfaat konservasi Biasanya memilki interior yang utuh (intact)
Biasanya lebih besar dari luas total
Sumber: Lord dan Norton (1990).

PRAGMENTASI HUTAN
34
II. DESKRIPSI POLA PATCH
Memahami dampak gangguan dan proses-proses dalam fragmentasi
lingkungan merupakan kunci untuk menggambarkan dan meramalkan pola-
pola patch. Pada skala lanskap dari beberapa sampai lusinan patch,
penggambaran matematis konfigurasi patch, travel corridor ( lintasan
koridor) dan pathway serta struktur edge merupakan alat yang berguna
untuk mengkarakterisasi pola patch pada suatu lanskap (Morrison et al.
1992).
Forman & Gordon (1986) me-review sejumlah cara untuk
menggambarkan pola patch. Hal ini meliputi menggambarkan patch-patch
sebagai matrix dan jaringan (network). Dengan penggambaran tersebut,
matematika network dapat digunakan untuk menganalisis bagaimana pola
patch tertentu memberikan konektivitas (connectivity) – potensi pergerakan
satwa – antar patch-patch. Forman dan Gordon juga menyarikan dalam
sebuah matrix ada enam ukuran karakteristik patch meliputi: patch shape,
isolation, accessibility, interaction dan dispersion (Morrison et al. 1992).
Patch-patch juga telah dideskripsikan dari perspektif kehadiran
edges dalam suatu lanskap. Kemampuan satwa untuk bergerak melalui
berbagai tipe edge yang disebut porosity atau permeability dari suatu edge.
Dari perspektif ini, beberapa model pergerakan satwa dalam suatu lanskap
berhubungan langsung dengan pola patch-patch sebagaimana
mempengaruhi jalan-jalan lebar (highways) untuk pergerakan, filters dan
barriers. Model demikian berguna untuk menggambarkan kondisi spesifik
spesies dari patch terisolasi dan terkoneksi (Buechner, 1987a,1987b; Stamps
et al. 1987). Buechner (1987b) memodelkan pergerakan satwa melintasi
taman nasional semenanjung dan menyimpulkan bahwa arah dan magnitude
pergerakan dapat dipengaruhi oleh ratio perimeter-area patch, edge
permeability, tingkah laku dan preferensi habitat suatu spesies dan ukuran
relatif menurunnya dispersal dan sumber asal satwa yang berpindah.
Konektivitas patch dan permebilitas edge bervariasi menurut
ukuran tubuh spesies, kekhususan habitat dan luas daerah jelajah (home
range). Patch yang berperan sebagai kesesuaian habitat secara keseluruhan
dalam tipe dan jumlah untuk melestarikan suatu satwa bertubuh kecil
berhabitat spesifik seperti red tree vole (Arborimus longicaudus), suatu

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
35
jenis spesialis pada Douglas fir, juga mungkin berperan, pada banyak skala
lebih luas, hampir sebagai batu loncatan (stepping stone) dalam dispersal
spesies bertubuh lebih besar dan tidak memerlukan habitat spesifik seperti
singa gunung (Felis concolor) (Morrison et al. 1992).
Spesies berbeda merespon lanskap dengan berbeda pula yang
mencerminkan apa yang dimaksud environmental grain size (ukuran
hambatan lingkungan). Satwa dengan perbedaan sumberdaya orientasi,
ukuran tubuh dan luas home range menerima dan menggunakan patch dan
sumberdaya pada skala yang berbeda. Serangkaian patch-patch spesifik
dapat muncul sebagai coarse grained (hambatan berat) bagi spesies dengan
jelajah sempit dan fine grained (hambatan ringan) untuk spesies dengan
jelajah luas. Dengan demikian asesmen apakah suatu konfigurasi patch-
patch tertentu dan derajat fragmentasi menguntungkan atau merugikan
sangat banyak tergantung pada karakteristik spesies dalam menggunakan
lanskap (Morrison et al. 1992).
III. PENGARUH UKURAN PATCH
Ukuran patch atau jumlah tipe lingkungan yang ada dalam suatu
lanskap secara langsung mempengaruhi kolonisasi oleh individu, ketahanan
(persistence) individu dan unit perkembangbiakan (breeding unit) serta
jumlah spesies dalam areal tersebut. Ukuran patch hutan di hutan gugur
sebelah timur telah dihubungkan dengan peluang (propbability) kehadiran
burung penyanyi dan
jenis jenis burung
lainnya (Whitcomb et
al., 1981; lihat
Gambar 6).
Gambar 6. Fungsi insiden dari pelatuk totol besar (Picoides major)
(Dari Moore & Hooper dalam Wilcove et al., 1986).

PRAGMENTASI HUTAN
36
Kurva peluang demikian disebut sebagai fungsi insiden (incidence
function). Fungsi insiden menggambarkan kemiripan spesies yang
ditemukan di dalam suatu patch habitat dari suatu areal tertentu. Fungsi
insiden dapat juga diinterpretasikan sebagai proporsi dari patch dengan
ukuran tertentu mengandung spesies yang ada.
Fungsi insiden juga digunakan untuk menggambarkan jumlah spesies
berbeda yang ada pada suatu areal (island atau patch) dengan ukuran
tertentu. Hubungan demikian disebut “hubungan kekayaan spesies - area”
(species richness - area relationship) dan secara matematis digambarkan
sebagai :
zCAS
dimana S adalah jumlah spesies yang ada, A adalah luas pulau atau patch, C
adalah konstanta skala yang nilainya bervariasi menurut takson dan lokasi,
dan z adalah laju dimana jumlah spesies meningkat dengan meningkatnya
luas. Dari sejumlah penelitian kekayaan spesies pada kepulauan samudera,
z bervariasi dari 0,24 untuk lahan perkembangbiakan dan burung-burung air
di West Indies dan vertebrata daratan di Kepulauan Danau Michigan, sampai
0,49 untuk lahan perkembangbiakan dan burung-burung air di teluk Guinea
(MacArthur & Wilson 1967). Preston (1962a, 1962b) menghitung sebuah
nilai teoritis 0,263. Ketika diplotkan pada sebuah hubungan log-log, dimana
S = log C + z log A, fungsinya tampak sebagai sebuah garis lurus dan z
menjadi slope dari garis tersebut. Sebagai konsekuensi dari faktor z,
sebagai hukum umum, dua kali lipat jumlah spesies tampaknya
membutuhkan 10 kali luas (Darlington 1957; Haris 1984). Hubungan
species-area melemah (nilai z menurun) dengan semakin terisolasinya pulau
dan dengan spesies yang kurang vagil.
Parameter atau ukuran-ukuran karakteristik patch dalam sebuah
matrix antara lain (Morrison et al. 1992):
Bentuk Patch (Shape of Patch)

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
37
dimana Di adalah indeks bentuk dari patch i, P adalah perimeter patch dan
A adalah luas patch.
Isolasi sebuah Patch (Isolation of a Patch)
dimana ri adalah indeks isolasi patch i, n adalah jumlah bertetangga yang
dipertimbangkan dan di j adalah jarak antara patch i dan patch
tetangganya j.
Isolasi Beberapa Patch (Isolation of Patches)
dimana D adalah indeks isolasi seluruh patch yang ada. Patch-patch
diletakkan pada sebuah grid dengan koordinat x dan y. Rata-rata lokasi
dan ragam (variance) untuk semua patch dihitung untuk kordinat y,
22
yx vv masing-masing adalah ragam pada koordinat x dan y.
Aksesibilitas sebuah patch (Accessibility of a Patch)
n
i
iji da1
dimana ai adalah indeks aksesibilitas patch i; dij adalah jarak yang
menghubungkan antara patch i dan patch manapun dari n patch tetangga j.
Interaksi antara patch-patch (Interction among patches)
n
i j
j
id
AI
12

PRAGMENTASI HUTAN
38
dimana Ii adalah derajat interaksi dari patch i dengan n patch-patch
tetangganya; A adalah luas patch tetangga j; dan d j adalah jarak antara
edges dari patch i dan patch-patch j.
Dispersi Patches (Dispersion of Pacthes)
cc dR 2
dimana Rc adalah indeks dispersi; dc adalah jarak rata-rata dari suatu patch
(pusatnya atau centroid) ke patch tetangga terdekatnya; dan adalah
kepadatan rata-rata dari patch-patch. Rc =1 untuk patch-patch tersebar
acak; Rc <1 untuk patch-patch menggerombol (aggregat); dan 1 < Rc ≤
2,149 untuk patch-patch tersebar teratur (regularly). Dengan demikian Rc
adalah ukuran penggerombolan (aggregation).
IV. PENGARUH FAKTOR-FAKTOR LAIN
Peluang kehadiran spesies tertentu atau tingkat kekayaan spesies
total tidak hanya dipengaruhi oleh luas (area) (Usher, 1985). Faktor kunci
lainnya meliputi keberadaan sumberdaya esensial seperti pakan, air dan
bahan pembuat sarang; heterogenitas lingkungan; dan keberadaan pesaing,
pemangsa dan penyakit (Morrison et al. 1992).
Freemark & Merriam (1986) melaporkan bahwa keragaman avi-
fauna hutan di antara 21 fragment hutan pada suatu lanskap pertanian
dekat Ottawa, Canada tergantung pada ukuran dan heterogenitas hutan
tetapi bahwa fragment berukuran besar penting untuk spesies interior
hutan dan penghuni tetap (resident). Askin (1984) dan Askin et al. 1987)
melaporkan bahwa keragaman dan kelimpahan burung-burung interior
hutan di tenggara Connecticut berkaitan dengan luas fragment hutan dan
luas hutan regional dan bahwa sebaran hutan regional merupakan faktor
yang lebih signifikan. Tempat yang lebih terisolasi dari hutan-hutan lainnya
memiliki lebih sedikit spesies interior hutan.

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
39
Lynch dan Whigham (1984) menghitung kembali kelimpahan lokal
spesies burung yang berkembang biak di hutan interior upland di pantai
Maryland ternyata secara signifikan dipengaruhi oleh luas hutan, isolasi,
struktur dan floristik. Mereka menemukan bahwa pengaruh fragmentasi
hutan adalah kompleks dan bersifat spesifik terhadp spesies. Walaupun
fragmentasi secara umum memiliki pengaruh negatif pada spesies interior
hutan, struktur hutan dan floristik ternyata lebih penting dari pada ukuran
patch dan isolasi bagi banyak spesies.
Keberadaan spesies lain dapat sangat mempengaruhi efek prinsip
species-area. Sebagai contoh, laba-laba orb (Aranae) mencapai kepadatan
ekstrim sangat tinggi di pulau-pulau sub tropikal karena tidak ada predator
(Schoener dan Toft, 1983).
Soule et al. (1979) menyimpulkan bahwa laju kepunahan mamalia
besar di 19 cagar alam di Afrika Timur akibat ukuran cagar yang kecil tetapi
kenyataannya cagar terbesarpun gagal memberikan habitat yang cukup
untuk memelihara viabilitas populasi selama beberapa abad. Kushlan
(1979) mengingatkan bahwa ukuran saja tidak cukup sebagai kriteria untuk
menetapkan cagar alam. Ia mencontohkan kasus Taman Nasional
Everglades di Florida yang kehilangan spesies walaupun ukurannya relatif
besar. Taman nasional tersebut bukan ekosistem mandiri tetapi sangat
tergantung pada nutrien dan air yang diterima dari luar batas taman
nasional.
Soule et al. (1988) juga melaporkan bahwa empat variabel biotik
dan biogeografik di habitat chaparral di California Selatan,
bertanggungjawab atas 90% dari variasi dalam kekayaan spesies burung.
Kekayaan spesies burung yang lebih besar berkorelasi dengan umur patch-
patch yang lebih muda, chaparral-nya lebih luas, luas total lembah yang
diteliti lebih besar dan keberadaan coyote (Canis latrans) dan ketiadaan
rubah abu-abu (Urocyon cinereoargenteus). Peneliti berhipotesis bahwa
keberadaan coyote menekan pemangsaan oleh rubah abu-abu, sementara
coyote sendiri tidak memakan burung.

PRAGMENTASI HUTAN
40
DAFTAR PUSTAKA
Askins, R.A. 1984. Effect of regional Forest Configuration on the Species Richness and Density of Forest Birds. Abstract of Paper Presented
at the 1984 Meeting of the American Ornitologists’ Union, August, in Lawrence, Kans.
Askins, R.A., M.J.Philbrick dan D.S. Sugeno. 1987. Relationship Between the Regional Abudance of Forest and the Compposition of Forest
Bird Comunities. Biological Conservation 39 : 129-152.
Buechner, M. 1987a. Conservation in Insular Parks : Simulation Models of
Factors Affecting the Movement of Animals Across Park Boundaries. Biological Conservation 41 : 57 – 76.
Buechner, M. 1987b. A Geometric Model of Vertebrate Dispersal : Tests and Implications. Ecology 68 : 310-318.
Forman, R.T.T. dan M. Gordon. 1986. Landscape Ecology. John Wiley and Sons. New York.
Freemark, K.E. dan H.G. Merriam. 1986. Important of Area and Habitat Heterogenity to Bird Assemblages in Temperate Forest Fragments.
Biological Conservation 36 : 115 -141.
Kushlan, J.A. 1979. Design and Management of Continental Nature
Reserves. Lessons from the Everglades. Biological Conservation 15 : 281-290.
Lord, J.M. dan D.A. Norton. 1990. Scale and the Spatial Concept of Fragmentation. Conservation Biology 4 : 197-2002.
Lynch, J.F. dan D.F.Whigham. 1984. Effects of Forest Fragmentation on Breeding Bird Communities in Maryland, USA. Biological
Conservation 28 : 287-324.
Morrison, M.L., B.G. Marcot and R.W. Mannan. 1992. Wildlife-Habitat Relationship: Consepts and Applications. The University of
Wisconsisn Press. Madison, Wisconsin.
Schoener, T.W. dan C.A. Toft. 1983. Spider Populations : Extraordinary
high Densities on Islands Without Top Predator. Science 219 : 1353-1355.
Shugart, H.H., Jr. 1984. A theory of Forest Dynamics. Springer-Verlag. New York.
Shugart, H.H., Jr. dan D.C. West. 1981. Long-term Dynamics of Forest Ecosystems. American scientist 69 : 672-652.

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
41
Soule, M.E., B.A.Wilcox dan C. Holtby. 1979. Benign Neglect : A Model of faunal Colapse in the Game Reserves of east Africa. Biological
Conservation 15 : 259-272.
Soule, M.E., D.T. Bolger, A.C. Alberts, J.Wright, M.Sorice dan S.Hill. 1988.
Reconstructed Dynamics of Rapid Extinctions of Chaparral-Requiring Birds in Urban Habitat Islands. Conservation Biology 2 : 75-92.
Stamps, J.A., M. Buechner dan V.V. Krishnan. 1987. The Effects of Edge Permeaability and Habitat Geometry on Emigration from Patches of
Habitat. American Naturalist 129 : 533 – 552.
Usher, M.B. 1985. Implication of Species-Area Relationship for Wildlife
Conservation. Journal of Environmental Management 21 : 181-191.
Whitcomb, R.F., C.S. Robbins, J.F. Lynch, B.L. Whitcomb,
M.K.Klimkiewecz, and D.Bystrak. 1981. Effects of Forest Fragmentation on Avifauna of the Eastern Deciduous Forest. In Ed.
R.L. Burgess and D.M. Sharpe (eds). Forest Island Dynamics in Man-Dominate Landscape. Pp.125-206. Springer-Verlag. Bew York.
Wilcove, D.S., C.H. McLellan dan A.P. Dobson. 1986. Habitat Fragmentation in the Temperate Zone. In M.E.Soule (ed). Conservation Biology. Pp. 237-256. Sinauer Associates.
Sunderland, Mass.

PRAGMENTASI HUTAN
42
5
KONSEP KORIDOR DAN SINGLE LARGE OR SEVERAL SMALL
(SLOSS)
KORIDOR
PERANAN KORIDOR
SINGLE LARGE OR SEVERAL SMALL (SLOSS)
DAFTAR PUSTAKA
I. KORIDOR
Koridor merupakan komponen lanskap berbentuk strip atau jalur
lahan yang berbeda dengan matrix di sekitarnya. Koridor merupakan areal
yang menghubungkan antar patch-patch sehingga berperan sebagai lintasan
atau saluran bagi organisme untuk bertukar atau berpindah dari suatu patch
ke patch lain. Koridor dapat menjadi saluran untuk perpindahan atau
penghalang (barrier) atau penyaring (filter), misalnya untuk aliran gen.
Bentuk lain konektivitas habitat adalah batu loncatan (stepping stone) yaitu
satu atau lebih kantong habitat (habitat patches) yang secara ekologis
terisolasi yang memberikan sumberdaya dan tempat pengungsian bagi satwa
dalam perpindahan menjelajahi suatu lanskap.
Struktur dan fungsi koridor tergantung pada beragam faktor
meliputi derajat liku-liku (semakin berliku-liku semakin banyak edges),
perpotongan dengan matrix (bagi tumbuhan dapat menghambat aliran
spesies, gen dan energi), penyempitan (dapat menghentikan perpindahan
beberapa spesies), perpotongan antar koridor (kadang-kadang ditemukan
banyak jenis interior) dan konektivitas yang harus dijaga agar tidak
terputus. Hal paling penting dalam koridor adalah pola lanskap yang mampu

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
43
mendukung konektivitas bagi spesies, komunitas dan proses ekologi
sehingga konservasi populasi dan komunitas dapat efektif dan proses
ekologi dalam lanskap terpelihara.
Walaupun koridor dapat berupa jalur terisolasi, tetapi sering
terhubungkan pada patch-patch dengan karakteristik vegetasi yang mirip
(Forman & Godron 1986). Beberapa studi telah menunjukkan bahwa koridor
dapat membantu organisme berpindah dari satu patch ke patch lainnya,
karena menghubungkan habitat dengan kondisi yang sesuai. Koridor dapat
dibagi ke dalam tiga kategori yaitu: koridor habitat, koridor perpindahan
(movement) dan koridor penghalang (barrier) yang dideskripsikan sebagai
berikut (Forman 1985) :
1. Koridor habitat – elemen lanskap linear yang memberikan daya
survival, natalitas dan pergerakan, juga dapat memberikan habitat,
baik sementara maupun permanen. Koridor ini secara pasif
meningkatkan konektivitas patch-patch. Contoh koridor habitat
adalah sabuk lahan basah bervegetasi yang dapat melestarikan
populasi salamander.
2. Koridor yang memfasilitasi perpindahan–elemen lanskap linear yang
memberikan daya survival dan perpindahan antara patch-patch
habitat tetapi tidak sampai diperlukan untuk melahirkan di dalam
koridor. Koridor yang memfasilitasi perpindahan secara aktif
meningkatkan konektivitas. Suatu lahan bervegetasi jalur yang
memiliki karakteristik edge dapat memberikan suatu lintasan untuk
suatu jenis satwa hutan interior tetapi tdak memberikan habitat.
Sebagai contoh, jalur vegetasi sepanjang jalan dapat memberikan
sumber pakan untuk seekor vole tetapi tidak cukup memberikan
lindungan untuk anak-anaknya.
3. Koridor penghalang (barrier) atau penyaring (filter) – elemen
lanskap linear yang menghalangi (barrier) atau menghambat (filter)
aliran energi, mineral, nutrien dan/atau spesies untuk melintasinya
(seperti : aliran tegak lurus terhadap panjang koridor). Sebagai
contoh koridor barrier atau filter adalah suatu jalan tol yang
menghalangi satwa melintasinya.

PRAGMENTASI HUTAN
44
Koridor satwaliar menjadi pertimbangan dalam konservasi di
populasi-populasi kecil yang terfragmentasi karena merupakan sarana untuk
menghubungkan populasi-populasi tersebut. Dengan memberikan lintasan
untuk perpindahan antar populasi tersebut melalui koridor maka dapat
meningkatkan peluangnya untuk survival. Koridor satwaliar dapat
berbentuk seperti tanaman pagar, kanal, selokan, pinggiran jalan dan rel,
serta sungai (Meret 2007).
II. PERANAN KORIDOR
Manfaat atau keuntungan potensial dari koridor satwaliar adalah
(Meret, 2007):
1. Meningkatkan laju imigrasi antara populasi sehingga dapat memelihara
keragaman, meningkatkan ukuran populasi, menurunkan kemungkinan
kepunahan dan menghindarkan inbreeding.
2. Meningkatkan areal untuk mencari makan bagi spesies dengan jelajah
yang luas.
3. Memberikan tempat melarikan diri dan bersembunyi dari predator,
kebakaran dan gangguan lainnya.
Koridor juga memiliki kerugian, seperti meningkatnya predasi
terhadap spesies asli terutama jenis-jens satwa kecil serta meningkatnya
perburuan atau perusakan o leh manusia karena lebih mudah dilakukan
terutama di koridor yang sempit. Koridor juga dapat menjadi saluran bagi
alien species yang bersifat invasif, penyakit dan patogen. Prinsipnya
semakin lebar koridor adalah semakin baik karena menurunkan peluang
predator menemukan mangsanya, memberikan habitat bagi jenis -jenis
interior dan memberikan fungsi yang lebih baik bagi pergerakan satwa.
Potensi merugikan lainnya dari koridor bagi satwaliar adalah
(Meret 2007):
1. meningkatkan imigrasi dapat menyebarkan penyakit, hama, spesies
asing, menurunkan tingkat keragaman genetik dan tekanan
outbreeding.

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
45
2. Memfasilitasi penyebaran kebakaran dan meningkatkan keterbukaan
terhadap predator, pemburu dan pencuri.
3. Biaya
Koridor satwaliar telah diusulkan untuk Gunung Kilimanjaro di
Tanzania. Meningkatnya pertanian di sekeli ling kaki gunung telah banyak
mengisolasi padang rumput dataran tinggi atau habitat moorland di gunung
dari padang rumput dataran rendah atau habitat savana. Isolasi ini telah
menjadi faktor dalam kepunahan dua spesies mamalia pegunungan. Ada
satu koridor yang menghubungkan dua areal dan banyak mamalia
menggunakannya. Gajah menggunakan koridor ini secara musiman untuk
berpindah antara gunung dan taman nasional yang jauhnya sekitar 20 km.
Kerbau, eland (antelop tanduk spiral) dan anjing liar telah diteliti
menggunakan koridor ini. Pergerakan satwa dari habitat dataran tinggi ke
dataran rendah melalui koridor
ini, barangkali penting dalam
menjaga populasi yang dapat
bertahan (viable population) dari
spesies yang ada (Meret 2007).
Sutcliffe & Thomas
(1996) dalam (Meret 2007) telah
mempelajari kupu-kupu ikal kecil
(ringlet butterfly) di Inggris
menggunakan metode mark-
recapture, menunjukkan bahwa
koridor dapat mengurangi resiko
kepunahan dan dapat membantu
rekolonisasi areal populasi kupu-
kupunya yang telah punah
(Gambar 7)
Gambar 7.
Contoh koridor perpindahan yang dibuat di pegunungan Costa Rica
(Dari Stiles & Clark, 1989).

PRAGMENTASI HUTAN
46
Haas (1995) dalam (Meret 2007) mempelajari persebaran burung-
burung dalam patch-patch berhutan. Tidak seperti yang diduga, ternyata
koridor tidak penting bagi burung-burung migran (karena mereka dapat
terbang melewati areal-areal yang tidak cocok dengan rute tahunan ke
suatu areal perkembangbiakan), Haas menemukan bahwa tempat-tempat
bervegetasi adalah penting sebagai batu loncatan (stepping stone) dan
dapat berfungsi sebagai koridor untuk burung robin dan trasher. Dengan
demikian koridor ini menjadi penting bagi burung-burung di areal yang
terisolasi.
III. SINGLE LARGE OR SEVERAL SMALL (SLOSS)
Perdebatan SLOSS (Single Large Or Several Small) adalah
perdebatan dalam ekologi dan biologi konservasi selama tahun 1970-an dan
1980-an yang memperdebatkan apakah cagar tunggal besar (single large)
atau kecil banyak (several small) yang terbaik untuk mengkonservasi
keanekaragaman hayati pada suatu habitat terfragmentasi. Pada Tahun
1975 Jared Diamond menyarankan beberapa “aturan” untuk mendisain
kawasan konservasi berdasarkan teori Bogeografi pulau dari MacArthur dan
Wilson (1967). Salah satu dari sarannya adalah bahwa suatu cagar tunggal
besar lebih baik dari pada cagar banyak tetapi lebih kecil walaupun luas
totalnya sama. Karena kekayaan spesies meningkat seiring luas habitat,
suatu blok habitat yang lebih besar akan mendukung lebih banyak spesies
dari pada beberapa blok habitat kecil-kecil. Ide ini dipopulerkan oleh
banyak pakar ekologi lainnya, dan telah dimasukkan ke dalam sebagian
besar buku teks standar dalam biologi konservasi, juga telah digunakan
dalam praktek perencanaan konservasi di dunia. Ide ini ditentang oleh
murid pendahulu Wilson, yaitu Daniel Simberloff yang menunjukkan bahwa
ide ini didasarkan pada asumsi cagar lebih kecil memiliki suatu komposisi
spesies yang terkumpul (nested) - dengan asumsi bahwa setiap cagar besar
memiliki semua spesies yang ada di cagar-cagar yang lebih kecil. Jika
cagar-cagar lebih kecil memiliki spesies yang tidak dimiliki oleh cagar
lainnya, maka ada kemungkinan bahwa dua cagar kecil dapat memiliki lebih
banyak spesies dari pada sebuah cagar besar tunggal.

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
47
Debat terjadi karena cagar-cagar kecil berbagi spesies satu dengan
lainnya (memiliki spesies yang sama), sehingga membawa pada
pengembangan teori nested subset oleh Bruce Patterson dan Wirt Atmar
pada tahun 1980-an dan pada pembentukan Biological Dynamics of Forest
Fragment Project (BDFFP) dekat Manaus, Brazil pada tahun 1980 oleh
Thomas Lovejoy.
Bila masih ada peluang atau potensi untuk memilih single large,
maka lebih baik memilih single large dari pada several small, tetapi bila
dihadapkan pada keterbatasan kawasan hutan dan fragmentasi yang tidak
dapat dihindarkan, mungkin several small menjadi pilihan. Masing-masing
memiliki kelebihan dan kelemahan, dengan campur tangan manajemen
kelemahan bisa dikurangi.
Single large memang memiliki banyak kelebihan maka lebih
banyak dianut para ahli dan praktisi konservasi. Satu kelemahan fatal
adalah jika terjadi bencana hebat seperti tsunami, kebakaran hutan atau
pandemi suatu penyakit atau hama, maka bisa memusnahkan seluruh
spesies yang ada tanpa ada yang tersisa. Hal tersebut dapat dihindari jika
habitat-habitat tersimpan dalam beberapa lokasi (several small), sehingga
jika satu habis terkena bencana, masih ada habitat lain yang tersisa di
tempat lain.
Kelebihan lain several small adalah mewakili tipe habitat yang
lebih beragam, lebih banyak menampung populasi spesies langka.
Kelemahan pada several small antara lain meningkatnya resiko kepunahan
lokal karena masalah genetik (menurun, inbreeding, hanyutan genetik),
perubahan demografik (laju kelahiran dan kematian terkait variasi atau
peluang acak) dan perubahan lingkungan (pemangsaan, kompetisi,
penyakit, bencana). Habitat yang terfragmentasi juga menurunkan peluang
rekolonisasi dan meningkatnya efek tepi (edge effect).
Untuk mengurangi efek tepi dan efek fragmentasi maka perlu
dibuat koridor yang menghubungkan kantong-kantong habitat yang terpisah
agar dapat saling berhubungan. Namun koridor juga memiliki kelemahan
antara lain dapat menjadi jalan penularan penyakit, meningkatnya predasi
dan perburuan di sepanjang koridor

PRAGMENTASI HUTAN
48
Bagaimanapun juga single large merupakan pilihan prioritas
karena dengan ukuran yang besar dapat mengurangi pengaruh edge, dapat
mencakup lebih banyak spesies dan mempunyai keanekaragaman habitat
yang lebih besar. Kelebihan ini dijelaskan dalam teori island biogeography.
Dukungan awal penggunaan teori biogeografi pulau dengan argumen bahwa
cagar besar tunggal umumnya lebih baik dalam melestarikan populasi lebih
banyak dan lebih besar dari pada luasan yang sama tetapi terbagi ke dalam
sekumpulan cagar kecil. Ada dua alasan untuk argumen tersebut yaitu:
Areal-areal berdekatan lebih bisa melestarikan komunitas utuh dari
spesies-spesies yang saling bebas (interdependent).
Areal-areal lebih bisa untuk memelihara viable population dari spesies
yang terjadi pada kepadatan populasi rendah, khususnya vertebrata
besar.
Meskipun demikian, sedikitnya ada dua masalah dengan dua
argumen tersebut yaitu:
Ukuran besar dipandang dari sudut mana? Untuk tanaman tahunan,
untuk vegetasi perennial dan untuk satwa kecil penghuni tetap,
beberapa hektar mungkin memenuhi semua kebutuhan habitat,
misalnya suatu lumpur gambut atau singkapan tebing. Oleh karena itu,
ahli konservasi yang ingin melindungi keanekaragaman tumbuhan,
mungkin lebih memilih membeli beberapa cagar berukuran 10 – 100
hektar dengan beragam habitat, tipe tanah dan rejim geologi dari pada
membeli satu cagar tunggal dengan luas 2.000 hektar. Tetapi jika kita
tertarik pada burung-burung hutan, cagar-cagar kecil berukuran 100 –
200 hektar mungkin tidak cukup besar untuk mendukung populasi yang
bersarang.
Beberapa kritik cepat menunjuk, bahwa menurut teori biogeografi
pulau, spesies pada pulau kecil tidak perlu menjadi subset dari pulau
besar. Kenyataannya, jika teori kesetimbangan diinterpretasikan
dengan tegas, maka setidaknya mereka memang bukan subset. Sebagai
hasilnya, kita mungkin menyelamatkan lebih banyak spesies dalam
suatu sistem cagar-cagar kecil dari pada dalam cagar besar tunggal,
walaupun setiap cagar bisa saja berisi lebih sedikit spesies.

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
49
Sebagai tambahan, ada kritik mendasar pada pendekatan ini karena ada
sedikit bukti bahwa perbedaan dalam keanekaragaman spesies dalam
pulau adalah kesetimbangan (equilibrium) antara kolonisasi dan
kepunahan. Secara spesifik, terdapat sedikit bukti untuk species
turnover.
Meskipun demikian, debat tersebut secara keseluruhan seperti
kehilangan arah yang menjadi perhatian. Bagaimanapun, kita akan
membuat cagar dimana kita menemukan spes ies atau komunitas yang ingin
kita selamatkan. Kita akan membuat cagar sebesar mungkin yang kita
dapat, atau sebesar yang diperlukan untuk melindungi elemen-elemen yang
menjadi perhatian kita. Kita biasanya tidak berhadapan dengan pilihan
optimasi seperti yang diperdebatkan.
Perdebatan panjang tentang SLOSS membawa implikasi dalam
menetapkan luas cagar, dimana pemilihan single large beralasan karena
cagar besar dapat mengurangi edge effect dan dapat mencakup lebih
banyak spesies dan mempunyai keanekaragaman habitat yang lebih besar
dibandingkan cagar kecil. Hal ini didasarkan pada teori biogeografi pulau
yang berimplikasi praktis: (1) tetapkan kawasan perlindungan seluas
mungkin agar dapat melestarikan spesies sebanyak mungkin, (2) bila
memungkinkan lahan-lahan di sekitar kawasan tersebut diambil alih untuk
meningkatkan luas kawasan yang telah ada (3) bila harus memilih maka
pilih cagar yang lebih besar.
Cagar kecil yang ditempatkan secara baik akan memiliki beberapa
kelebihan antara lain : dapat mencakup tipe-tipe habitat yang lebih
beragam, menampung lebih banyak populasi spesies langka, lebih mudah
terhindar dari bencana seperti spesies asing, penyakit atau kebakaran.
Cagar kecil yang dekat dengan pemukiman juga dapat berfungsi sebagai
pusat pendidikan konservasi yang berguna untuk mendukung usaha
konservasi jangka panjang.
Daerah konservasi yang berbentuk membulat akan meminimalkan
rasio atau perbandingan edge-to-area, sehingga mempunyai pusat yang
berada relatif jauh dari tepi. Kawasan yang berbentuk memajang akan
memiliki tepi atau pinggir yang luas dan seluruh lokasi di kawasan tersebut
akan berada dekat tepi. Kawasan yang berbentuk segi empat bujur sangkar

PRAGMENTASI HUTAN
50
lebih banyak memberikan perlindungan dibandingkan persegi panjang. Ide
tersebut belum diterapkan karena kebanyakan kawasan konservasi
ditetapkan secara kebetulan sehingga bentuknya tidak beraturan.
Untuk mengurangi efek fragmentasi, cagar sebaiknya dikelola
sebagai sistem regional untuk memperlancar aliran genetik serta migrasi
antar populasi dan menjamin perwakilan yang cukup bagi spesies dan
habitat. Bila memungkinkan, kawasan perlindungan perlu mencakup
keseluruhan ekosistem (misalnya DAS, danau maupun gunung) sehingga
memungkinkan pengelola mempertahankannya secara lebih efektif dari
pengaruh luar yang bersifat
merusak.
Untuk mengelola cagar-cagar
kecil dan menyebar perlu dibuat
koridor habitat yang menghubungkan
kawasan-kawasan dilindungi tersebut
sehingga membentuk suatu sistem
yang lebih besar. Koridor dapat
memungkinkan perpindahan
tumbuhan dan satwa untuk
menyebar sehingga memungkinkan
aliran gen serta koloniasasi lokasi
yang sesuai. Koridor dapat berfungsi
melestarikan satwa yang harus
bermigrasi musiman di antara seri
habitat yang berbeda. Kendala
potensial dari koridor adalah dapat
menjembatani pergerakan spesies
hama dan penyakit yang dapat
mengancam kepunahan spesies
langka yang dilindungi dan resiko
predasi yang lebih besar dari satwa
lain atau pemburu yang biasanya
terkonsentrasi di jalur lintasan satwa.
Gambar 8. Perbandingan SLOSS (Dari berbagai sumber)

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
51
DAFTAR PUSTAKA
Forman, R.T.T. and M. Gordon. 1986. Landscape Ecology. John Wiley and Sons. New York.
Forman, R.T.T. 1985. Land Mosaics: The Ecology of Landscapes and Regions. Cambridge University Press, Cambridge, Uk.
Meret, J. 2007. Habitat Fragmentation and Wildlife Corridors. http://www.science. mcmaster.ca.htm. Diakses Tanggal 02-11-
2007.
Stiles, F.G. and D.A. Clark. 1989. Conservation of Tropical rain Forest
Birds : A Case Study from Costa Rica. American Birds 43 (Fall) : 420-428.
Mac Arthur, R.H. and E.O. Wilson. 1967. The Theory of Island Biogeography. Princeton University Press. Princeton, New Jersey.
http://en.wikipedia.org/wiki/SLOSS_Debat. Diakses Tanggal 10 Desember 2007.

PRAGMENTASI HUTAN
52
6
DINAMIKA POPULASI PADA SKALA LANSKAP
PROSES-PROSES LANSKAP
DINAMIKA METAPOPULASI
PULAU SAMUDERA (OCEANIC ISLAND) DAN KANTONG HABITAT (HABITAT PATCH)
KESETIMBANGAN (EQUILIBRIUM ) POPULASI ANTAR KANTONG HABITAT (HABITAT PATCH) DAN SUBPOPULASI
DAFTAR PUSTAKA
I. PROSES-PROSES LANSKAP
Untuk memberikan distribusi habitat yang mencukupi di dalam dan
lintas lanskap, kita harus memahami agen-agen yang mengubah kondisi
lingkungan. Proses-proses lanskap terdiri atas abiotik dan biotik. Proses
abiotik mempengaruhi pola habitat dalam lanskap meliputi pembuangan
massa, pembentukan tanah, kualitas air dan udara serta pengaruh bahan
induk pada perkembangan vegetasi. Secara umum, proses abiotik
melibatkan pertukaran material - udara, gas, partikel, tanah dan air – di
dalam dan di antara lanskap. Mengelola habitat pada skala lanskap harus
menjawab bagaimana pertukaran material dari dan ke tapak mempengaruhi
tujuan perencanaan tapak dan bagaimana kegiatan transport materia l di
tapak tersebut ke areal lainnya. Menilai dampak kumulatif dari kedua
aktivitas di dalam dan di luar tapak adalah penting ketika merencanakan
jumlah dan konfigurasi habitat spesifik.
Gangguan alami juga merupakan bagian dari proses abiotik dalam
lanskap. Gangguan dapat bersifat kronik atau perlahan, contohnya meliputi
suksesi vegetasi, perubahan iklim dan perubahan frekuensi dan intensitas

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
53
kebakaran. Gangguan juga dapat bersifat katastropik atau cepat seperti
kebakaran, badai, banjir dan gunung meletus.
Karr and Freemark (1985) menawarkan suatu klasifikasi faktor-
faktor untuk dievaluasi dalam studi gangguan (Tabel 2). Faktor-faktor
untuk dipertimbangkan meliputi tipe dan rejim gangguan, tipe sistem
biologis, dan konteks regional (context). Karr dan Freemark juga
menyebutkan bahwa respon biotik terhadap gangguan, bervariasi mulai dari
kepunahan populasi sampai perubahan dalam laju pertumbuhan yang
membawa perubahan perilaku dan ekologi dalam seleksi habitat.
Shugart and Seagle (1985) memodelkan Okupansi patch (patch
occupancy) oleh vertebrata dalam kehadiran distribusi di suatu ekosistem
hutan hujan basah di Tanzania dan menyimpulkan bahwa keanekaragaman
(diversity) hutan dan lingkungannya secara konstan sepanjang waktu
mempengaruhi kekayaan spesies di dalam suatu lanskap. Semakin besar
lanskap, semakin besar penyokong variasi lingkungan. Dalam modelnya
juga, kehadiran interaksi kompetitif di antara pengkoloni sangat
menurunkan jumlah spesies potensial di dalam suatu lanskap.
Proses-proses biotik meliputi peran invasi spesies sebagai parasit
sarang dan kompetitor. Karena habitatnya dieksploitasi manusia, burung
cowbird kepala cokelat (Molothrus ater) telah menginvasi Sierra Nevada,
California sejak 1930. Mereka sekarang menjadi parasit sarang dari 22
spesies burung yang menjadi inangnya dan dapat mengancam
keberlangsungan survival beberapa spesies di Sierra, khususnya burung
warbling vireo (Vireo gilvus) (Rothstein et al., 1980).
Proses-proses biotik lainnya seperti epidemi, meningkatnya atau
invasi spesies hama yang menekan ukuran populasi atau trend spesies lain–
juga mempengaruhi distribusi dan kelimpahan suatu populasi pada suatu
lanskap. Spesiasi dan subspesiasi merupakan proses biotik yang
mempengaruhi bagaimana spesies merespon kondisi lanskap di berbagai
zona geografis yang luas selama ribuan tahun generasi. Gangguan oleh
manusia, seperti eksploitasi, perburuan dan introduksi spesies eksotik dapat
dipandang sebagai suatu proses biotik.
Ekologi lanskap kebanyakan ditujukan pada bagaimana satwa
menggunakan lingkungan yang heterogen. Mempelajari pola seleksi

PRAGMENTASI HUTAN
54
sumberdaya, perilaku mencari makan (foraging behaviour) serta pola dan
derajat spesialisasi sumberdaya, semua memberikan informasi tentang
penggunaan habitat. Dinamika fauna berhubungan dengan bagaimana
satwa berpindah melalui lanskap. Dengan demikian, kita juga mempelajari
tipe-tipe perpindahan, seperti dispersal satwa muda (juvenile), migrasi
musiman dan tahunan satwa dewasa serta pola perpindahan kurang teratur
(reguler). Yang menjadi perhatian dalam disain lanskap adalah pola, laju
dan jarak pergerakan spesies, serta perilaku perkembangbiakan dan sosial
spesies yang terkait.
Tabel 2. Klasifikasi faktor-faktor untuk dievaluasi dalam suatu studi gangguan lanskap.
Faktor Contoh Hasil Respon Biotik
Tipe Gangguan
Faktor fisik Dampak kekeringan terhadap
distribusi burung dan katak di hutan tropis
Perilaku dan ekologi seleksi
habitat
Faktor biologi Demam kuning menurunkan
populasi monyet howler
Evolusi resistensi penyakit,
kepunahan indang, atau perubahan siklus kelimpahan
monyet
Interaksi faktor fisik dan biologi
Hujan musim semi dan distribusi burung mempengaruhi
ikan di Everglades
Laju pertumbuhan, survivorship dan reproduksi ikan
Rejim Gangguan
Dimensi spasial Distribusi pohon tumbang vs badai di areal hutan yang luas
Waktu untuk rekolonisasi dan kumpulan vertebrata akan bervariasi
Dimensi temporal
Frekuensi Serangan musim kering tahunan vs tidak menentu
Adaptasi fisiologi vs respon perilaku, ekologi dan evolusi
Waktu kejadian Pakan dan sedimentasi selama periode ikan bertelur
(spawning)
Kepunahan kohort (populasi awal)
Tipe Sistem Biologi
Individu –
populasi spesies
Burung dan kadal bervariasi
dalam mobilitas, lamanya dan kemampuan untuk
menghentikan reproduksi pada pulau jembatan (land bridge island)
Laju kepunahan burung tinggi,
tetapi laju kepunahan kadal rendah di Pulau Barro Colorado,
Panama.
Tipe Kumpulan/ lingkungan ekosistem – biotik
dan abiotik
Ikan lebih terbatasi oleh kimiawi lingkungan perairannya dari pada burung oleh kimiawi
udara yang beracun
Rangkaian adaptasi fisiologi dan perilaku yang berbeda, tetapi polusi udara dari manusia
mengubah kesetimbangan ini.

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
55
Faktor Contoh Hasil Respon Biotik
Konteks Regional
Dalam areal Ukuran luas dan sifat internal
mosaik habitat
Survivorship, kolonisasi, pola
kepunahan bervariasi antar pulau.
Antara Areal dan
wilayah didekatnya
Habitat pulau hutan pada
samudera dan pulau hutan pada lahan pertanian
Variasi kepunahan dan dinamika
kolonisasi antar kelompok vertebrata; membedakan
pengaruh kolonisasi dari patch-patch tetangganya dari rumput vs air
Sumber: Karr & Freemark (1985).
II. DINAMIKA METAPOPULASI
Memahami dan memprediksi respon satwaliar terhadap konfigurasi
patch dan fragmentasi pada skala lanskap memerlukan pemahaman
dinamika populasi. Pada bab ini akan dibahas konsep dinamika
metapopulasi. Kita diskusikan implikasi dari konsep metapopulasi untuk
mendisain pola patch pada skala lanskap dalam rangka melestarikan
viabilitas populasi. Kemudian kita membandingkan konsep ekologi pulau
samudera dengan bagaimana populasi menggunakan patch-patch di suatu
setting benua dan menggarisbawahi dinamika kedua setting tersebut saling
memiliki persamaan. Dengan cara demikian kita dapat memahami lebih
baik bagaimana populasi mungkin merespon pola patch-patch habitat dan
kondisi pada tingkat lanskap. Kemudian kita dapat mendiskusikan pengaruh
pola juxtaposition, yang meliputi fragmentasi lingkungan. Akhirnya kita
mendiskusikan bagaimana ukuran populasi dan trends yang mungkin
mencapai kesetimbangan (equilibrium) di antara patch-patch (Morrison et
al. 1992).
Suatu metapopulasi adalah suatu spesies yang jelajahnya terdiri
atas patch-patch yang berbeda secara geografis yang dihubungkan melalui
pola aliran genetik, kepunahan dan rekolonisasi (Lande and Barrowclough
1987). Metapopulasi umumnya terjadi ketika kondisi lingkungan dan
karakteristik spesies memberikan pertukaran yang kurang sempurna dari
sumber individu dan genetik di antara sub populasi. Ini terjadi khususnya
ketika habitat berada pada kondisi heterogen pada suatu wilayah, provinsi
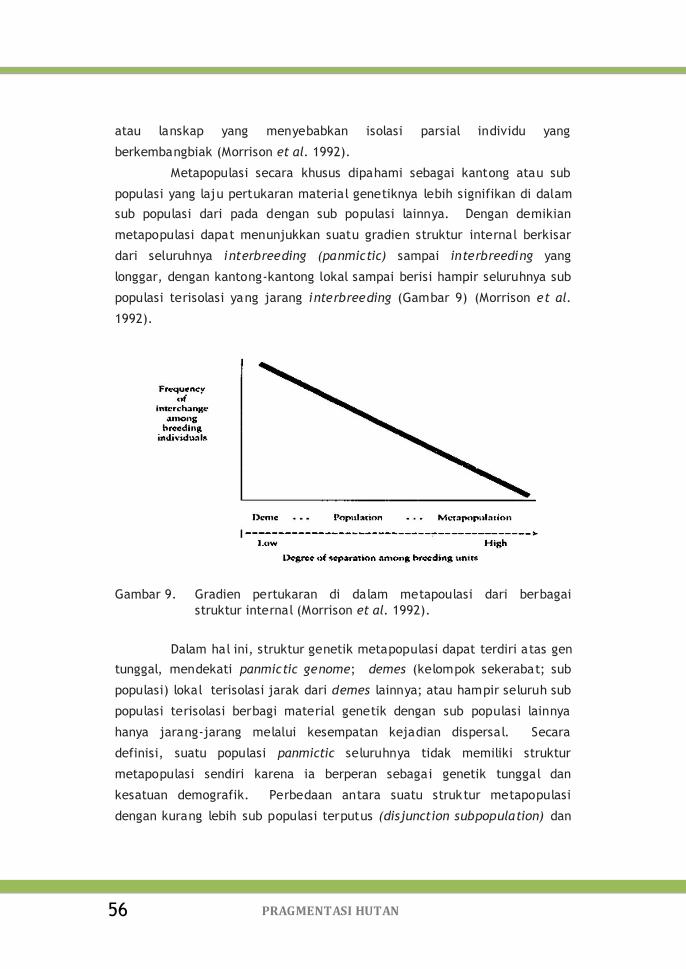
PRAGMENTASI HUTAN
56
atau lanskap yang menyebabkan isolasi parsial individu yang
berkembangbiak (Morrison et al. 1992).
Metapopulasi secara khusus dipahami sebagai kantong atau sub
populasi yang laju pertukaran material genetiknya lebih signifikan di dalam
sub populasi dari pada dengan sub populasi lainnya. Dengan demikian
metapopulasi dapat menunjukkan suatu gradien struktur internal berkisar
dari seluruhnya interbreeding (panmictic) sampai interbreeding yang
longgar, dengan kantong-kantong lokal sampai berisi hampir seluruhnya sub
populasi terisolasi yang jarang interbreeding (Gambar 9) (Morrison et al.
1992).
Gambar 9. Gradien pertukaran di dalam metapoulasi dari berbagai
struktur internal (Morrison et al. 1992).
Dalam hal ini, struktur genetik metapopulasi dapat terdiri atas gen
tunggal, mendekati panmictic genome; demes (kelompok sekerabat; sub
populasi) lokal terisolasi jarak dari demes lainnya; atau hampir seluruh sub
populasi terisolasi berbagi material genetik dengan sub populasi lainnya
hanya jarang-jarang melalui kesempatan kejadian dispersal. Secara
definisi, suatu populasi panmictic seluruhnya tidak memiliki struktur
metapopulasi sendiri karena ia berperan sebagai genetik tunggal dan
kesatuan demografik. Perbedaan antara suatu struktur metapopulasi
dengan kurang lebih sub populasi terputus (disjunction subpopulation) dan

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
57
suatu struktur populasi yang kontinyu (clinal population) dengan
intergrading sub populasi, bagaimanapun merupakan suatu masalah tingkat
pertukaran di antara unit-unit sub populasi (Morrison et al. 1992).
Suatu contoh distribusi metapopulasi terdiri atas hampir
seluruhnya sub populasi terisolasi terjadi pada spesies burung pegunungan
di sepanjang pegunungan Great Basin (Johnson 1975; 1978). Johnson
mengeksplorasi sumber keanekaragaman komunitas burung boreal (Gambar
10). Ia melaporkan bahwa agen berikut diurutkan menurut kore lasi
penurunannya, mempengaruhi jumlah spesies burung boreal; luas hutan
woodland, latitude puncak tertinggi, elevasi puncak tertinggi, dan lebar
barrier (derajat isolasi). Dengan demikian, distrubusi metapopulasi secara
keseluruhan dari suatu spesies dipengaruhi oleh karakteristik lokal yang
mempengaruhi kepunahan lokal seperti halnya isolasi mempengaruhi
kolonisasi (Morrison et al. 1992).
Gambar 10. Sumber keragaman komunitas burung boreal di bagian barat daya Amerika Serikat (Sumber: Johnson 1975).
Secara khusus, ilmuwan mempersepsikan metapopulasi sebagai
kelompok-kelompok individu yang menempati luasan habitat terbatas
secara lokal. Dari waktu ke waktu, individu dalam patch-patch atau
kelompok patch mungkin menjadi punah dari peluang keragaman dalam
survivorship dan rekrutmen atau dari penurunan katastropik atau sistematik
berdasarkan sumbernya (Gambar 10) Tapaknya mungkin bahkan menjadi

PRAGMENTASI HUTAN
58
dikolonisasai oleh individu yang menyebar atau hanyut dari patch-patch di
sekitarnya. Dengan cara ini, pola okupansi patch dalam luasan geografik
yang besar mungkin berlaku sebagai suatu bank dari cahaya berkedip-kedip.
Beberapa cahaya masih berkedip dalam periode waktu yang lebih lama
karena mereka mewakili kondisi optimal, areal-areal dengan jumlah habitat
yang banyak, atau patch-patch yang menjadi pusat dari sebaran geografik.
Cahaya lainnya mungkin menjadi gelap dan berkedip secara jarang dan
hanya untuk periode pendek karena mereka mewakili kondisi sub optimal,
areal-areal dengan jumlah habitat lebih sedikit, atau patch-patch pinggiran
untuk sebaran geografis secara umum. Gambar 11a – 11g berikut ini
menunjukan gambaran skematik dinamika lanskap kolonisasi dan okupansi
patch (Morrison et al. 1992).
11a. Batas Das dan sistem riparian (riverine) (Morrison et al. 1992).
11b. Kejadian sepuluh patches dari hutan tua (Morrison et al. 1992).
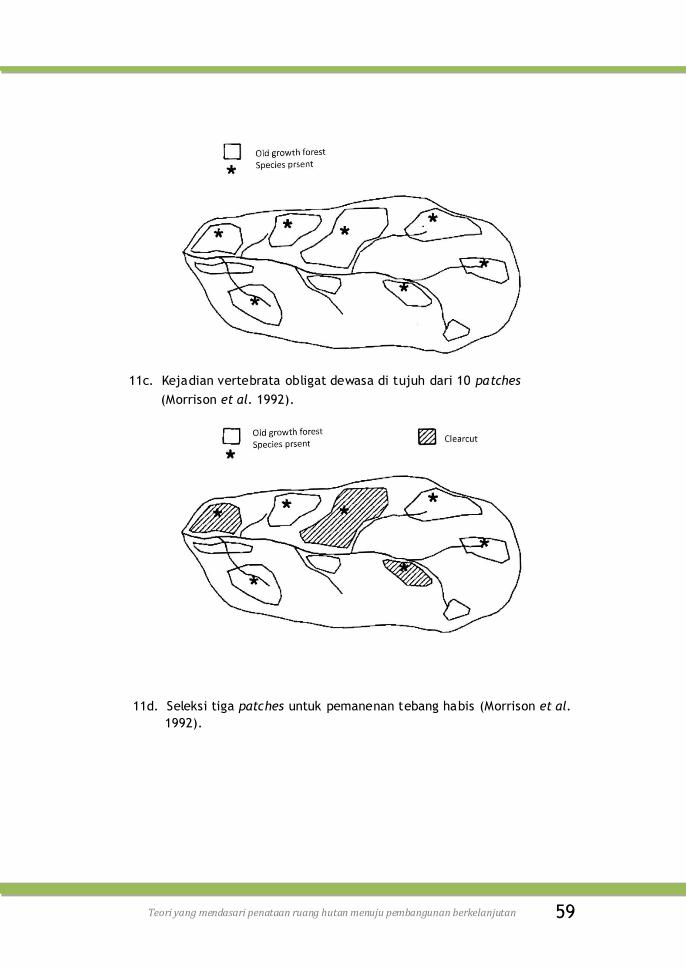
Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
59
11c. Kejadian vertebrata obligat dewasa di tujuh dari 10 patches
(Morrison et al. 1992).
11d. Seleksi tiga patches untuk pemanenan tebang habis (Morrison et al.
1992).

PRAGMENTASI HUTAN
60
11e. Akibat langsung dari ganguan pemanenan : hilangnya spesies.pada
tiga patches (Morrison et al. 1992).
11f. Kehilangan spesies kemudian pada patch hutan yang jauh,
terisolasi dan kecil (faunal relaxation) (Morrison et al. 1992).

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
61
Struktur metapopulasi dapat muncul dari beragam faktor,
termasuk pengurangan populasi yang dulu berdekatan akibat fragmentasi
atau kehilangan habitat; invasi tapak-tapak yang sampai kini tidak dihuni,
karena ekspansi jelajah spesies karena habitatnya menjadi available atau
tidak adanya kompetitor kunci, predator, atau penyakit; kejadian dan
kolonisasi penggunaan sink habitat (habitat yang selama ini tidak
digunakan), untuk reproduktif pada tahun-tahun yang baik; heterogenitas
disribusi sumberdaya, seperti buah-buahan tropika, dari waktu ke waktu di
berbagai ruang; heterogenitas atau distribusi habitat yang berkantong -
kantong (patchy), seperti substrat lithic atau lingkungan akuatik; dan
introduksi populasi oleh manusia (Morrison et al. 1992).
Dinamika metapopulasi adalah kompleks, mencakup dinamika
spasial, temporal dan numerik, seiring kondisi biologis dan sumbedaya
berubah dari waku ke waktu dan sejauh jelajah spesies. Dari perspektif
manajemen, resiko potensial terhadap kepunahan dari struktur
metapopulasi adalah sulit untuk dianalisis. Pertanyaan penting untuk
merencanakan habitat bagi konservasi viabilitas suatu metapopulasi
memperhatikan perkiraan peluang bahwa populasi yang menyebar dengan
11g. Masih kehilangan spesies kemudian pada suatu patch hutan lebih
besar yang kini terisolasi (Morrison et al. 1992).

PRAGMENTASI HUTAN
62
baik akan terpelihara sepanjang waktu. Distribusi patch-patch dalam
lanskap, khususnya karena pengaruhnya terhadap aksesibiltas di antara
individu dan sub populasi, adalah penting untuk membantu menjamin
populasi terdistribusi dengan baik (Morrison et al. 1992).
Apakah implikasi dari dinamika tersebut bagi perencanaan dan
pengelolaan patch atau habitat? Satu panduan yang mungkin adalah untuk
mengatur patch habitat-habitat sub optimal (sink) di sekelilingnya tetapi
dekat ke habitat-habitat (sumberdaya) optimal. Hal ini akan
memungkinkan rekolonisasi sink habitat selama periode dimana
sumberdayanya baik. Namun seringkali kita tidak tahu apa penyusun
kondisi sub optimal dan optimal. Dalam kasus ini, ada dua kemungkinan
penyebab kejadian. Kita dapat melihat pada professional judgement dan
melakukan survei tipe Delphi mencari pendapat ahli untuk memulai
membuat penduan perencanaan patch-patch. Panduan harus
diimplementasikan dan respon individu dan populasi dimonitor. Atau kita
dapat mendasarkan perencanaan awal panduan pada parameter-parameter
habitat untuk memprediksi model seperti model indeks kesesuaian habitat
(habitat suitability index model). Sekali lagi, respon individu dan populasi
harus dimonitor (Morrison et al. 1992).
Pickett & Thompson (1978) juga menyimpulkan bahwa
memelihara sumber kolonisasi adalah vital untuk melestarikan spesies di
dalam cagar alam. Mereka merekomendasikan bahwa disain cagar alam
harus didasarkan pada ”minimum dynamic area” yaitu luas minimal dengan
rejim gangguan alami yang menyeimbangkan laju emigrasi dari sumber
eksternal dengan laju kepunahan internal.
Pada skala home range individu atau unit reproduksi, pedoman
lain adalah untuk memberikan kantong-kantong habitat (habitat patches)
untuk makan dan istirahat di dalam jarak jelajah harian. Tetapi hal ini juga
menyebabkan meningkatnya kebutuhan sumberdaya ketika merencanakan
foraging habitat di dekat struktur atau habitat berkembangbiak; hal ini
akan membantu perencanaan untuk ukuran patch-patch yang cukup dan
pendekatan tipe habitat berbeda yang diperlukan untuk kebutuhan yang
berbeda pada berbagai tahap kehidupan (Morrison et al. 1992).

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
63
Dinamika kolonisasi patch-patch tampaknya tidak semua patch
yang cocok (suitable) akan ditempati oleh satwa dalam satu waktu. Oleh
karena itu, dari perspektif manajemen, tidak perlu mengkonservasi patch-
patch yang potensial suitable jika tidak ditempati. Lebih baik memonitor
patch-patch dari waktu ke waktu untuk mengetahui pola penghunian
(occupancy) sebelum mengubah arah manajemen yang akan sangat
menurunkan kualitas patch-patch. Tampaknya patch-patch yang kosong
mungkin masih vital untuk dispersal dan distribusi. Monitoring harus
digunakan untuk mengetahui frekuensi dan tipe penggunaan patch-patch
untuk keperluan tersebut (Morrison et al. 1992).
Konsep dinamika metaopulasi dapat bermanfaat untuk
menetapkan kriteria dalam mencantumkan (listing) spesies yang terancam
(threatened), dalam bahaya (endangered), jarang (rare) atau rawan
(sensitive) sebagaimana dinamika kepunahan lokal dan kolonisasi
meningkatkan resiko menjadi viability. Sebaliknya, konsep tersebut juga
bisa memberikan pedoman untuk mengeluarkan suatu spesies dari daftar
(delisting). Membuat kriteria delisting akan memerlukan penetapan target
tujuan recovery yang realistik untuk distribusi suatu populasi yang cukup,
memungkinkan tingkat isolasi yang dikehendaki dan interaksi sub populasi
dan derajat realistik penghunian patch (Morrison et al. 1992).
Variasi kondisi dan kualitas patch mungkin diperlukan untuk
menjaga populasi terdistrubusi dengan baik. Kita harus mempe rtimbangkan
areal-areal yang digunakan secara musiman untuk migrasi, dispersal dan
beristirahat dalam rangka membantu pertukaran individu di dalam dan
antar sub populasi. Secara umum, hal tersebut mungkin yang terbaik untuk
memberikan variasi kondisi habitat untuk menjaga keragaman genetik di
seluruh ekotipe, sub populasi dan ras-ras metapopulasi. Memberikan hanya
kondisi optimal – lingkungan dengan okupansi tertinggi atau konsisten –
mungkin tidak memberikan untuk spesiasi jangka panjang dan diversifikasi
stok genetik (Morrison et al. 1992).
Secara umum, memahami dinamika metapopulasi merupakan
kunci untuk memberikan koreksi jumlah, kualitas dan distribusi habitat
pada berbagai skala. Hal tersebut harus dapat memberikan panduan dalam
rehabilitasi habitat dan penambahan lahan, misalnya untuk penggunaan

PRAGMENTASI HUTAN
64
musiman atau migrasi (Gambar 4). Patch lebih besar, berdekatan lebih
disukai dari pada patch kecil dan terisolasi. Tetapi dalam memberikan
sambungan antar patch-patch, hanya ada sedikit pilihan untuk disain
(Morrison et al. 1992).
Suatu struktur metapopulasi tidak selalu lebih buruk (inferior)
dibandingkan suatu populasi panmictic secara keseluruhan. Tidak jelas
apakah menghubungkan sub populasi dari suatu metapopulasi selalu
bermanfaat. Keuntungan yang diberikan oleh suatu struktur metapoulasi
meliputi pertahanan (buffering) yang lebih besar terhadap penyebaran
penyakit dan kejadian-kejadian katastropik. Disamping itu, semakin besar
keragaman lokasi tidak tersambung yang dihuni memungkinkan terjadinya
manfaat evolusi yang lebih besar. Ekotipe lokal mungkin dapat berkembang
menjadi lebih cocok dengan kondisi lingkungan lokal dari pada jika genome
terbanjiri dari panmixia dengan semua sub populasi lainnya (Morrison et al.
1992).
III. PULAU SAMUDERA (OCEANIC ISLAND) DAN KANTONG HABITAT (HABITAT PATCH)
Dalam beberapa hal, dinamika populasi dan metapopulasi pada
kantong (patch) habitat benua (continental) menunjukkan karakteristik
yang berbeda dengan pulau samudera (oceanic island). Misalnya, pada
pulau samudera, laut di sekililingnya tidak dapat dihuni dan dispersal
melaluinya adalah kejadian yang jarang. Sebaliknya, pada lanskap benua,
lahan terganggu mungkin sub optimal tetapi dapat digunakan untuk
dispersal, istirahat atau migrasi musiman atau tahunan (Whitcomb 1977).
Oleh karena itu, pola perpindahan di antara kantong (patch) habitat pada
lanskap benua dapat lebih kompleks dari pada di antara pulau.
Disamping itu, ada perbedaan dalam pengaruh dari pola dan
juxtaposition dari patch habitat. Pada lanskap benua, pola patch secara
langsung mempengaruhi laju penghunian dan dinamika kolonisasi dan
dengan demikian populasi bertahan (persistent) di dalam areal. Mengingat
hal ini juga benar pada pulau samudera, maka umumnya tidak
dipertimbangkan dalam teori kesetimbangan pulau. Dengan demikian

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
65
penggambaran proses kolonisasi dan kepunahan bisa lebih kompleks dalam
lanskap hutan dari pada dalam pulau (Morrison et al. 1992).
Juga terdapat perbedaan dalam pengaruh ukuran patch.
Walaupun sebuah patch kecil dalam lanskap benua dapat berhenti berfungsi
sebagai habitat, sama halnya dengan pulau samudera yang kecil dan
terisolasi, ini dapat menyumbang pada kejadian (occurance) dan
keberadaan (persistence) dari suatu spesies dalam sebuah lanskap jika
kondisi di dekatnya atau sekitarnya suitable untuk spesies yang
dipertanyakan. Patch-patch di dalam lanskap benua memiliki ciri mendapat
tekanan invasi pengaruh tepi (edge effect) ke dalam interior patch.
Semakin kecil patch, semakin besar fraksi patch dipengaruhi oleh edge.
Tidak ada pesamaan yang jelas antara pengaruh tersebut dengan pulau
samudera. Demikian juga, bahkan jika patch lebih kecil dari yang
dibutuhkan untuk memberikan kebutuhan pokok bagi berbagai tahapan
kehidupan suatu asosiasi spesies, patch-patch mungkin masih memberikan
sumbangan berarti bagi dispersal, mencari makan, pelindung atau tempat
istirahat, tergantung pada lanskap dimana patch itu ada. Di sisi lain, suatu
pulau samudera yang kecil tidak memberikan sumbangan apa-apa (Morrison
et al. 1992).
Mungkin ada perbedaan dalam pengaruh kompetisi antara spesies
dalam pulau dan spesies dalam lanskap benua. Martin (1981) meneliti
burung-burung di 69 pulau hutan (shelterbelt) di Dakota selatan bagian
timur menemukan bahwa ketidakhadiran spesies tidak sesederhana karena
preferensi habitat atau isolasi. Interaksi kompetitif antar spesies
mempengaruhi jumlah individu dan spesies dalam komunitas dan struKtur
ekologi komunitas, sementara interaksi kondisi lingkungan, kesempatan dan
kompetisi menentukan pola distribusi individu spesies di antara komunitas.
Sebaliknya, spesies mamalia kecil di pulau barrier Virginia menunjukkan
tidak ada pengaruh dari kompetisi pada sumberdaya yang digunakan oleh
setiap spesies, walaupun tingkat segregasi habitat yang tinggi teramati
(Dueser & Porter 1986).
Laju kepunahan spesies, baik di pulau samudera maupun kantong
habitat dalam suatu lanskap merupakan fungsi dari meningkatnya isolasi,
yang diakibatkan oleh fragmentasi dan menurunnya luasan. Pada pulau-

PRAGMENTASI HUTAN
66
pulau dan dalam lanskap benua, laju kepunahan umumnya lebih tinggi pada
spesies dengan spesialisasi pada sumberdaya atau lingkungan tertentu dari
pada spesies yang lebih generalis (Morrison et al. 1992).
Schoener & Schoener (1983) yang meneliti kadal pada pulau-pulau
kecil Bahama melaporkan bahwa waktu untuk kepunahan secara langsung
meningkat dengan luas pulau. Di atas suatu luasan tertentu, kolonisasi
cepat, tetapi di bawahnya, pengkoloni dengan cepat menjadi punah.
Menurut Whitcomb et al. (1981) dalam kantong habitat benua, fungsi-fungsi
insiden (incidence functions) menunjukkan hubungan yang serupa antara
ukuran patch dan kehadiran spesies ketika patch cukup terisolasi dari
sumber spesies pengkoloni (Gambar 3). Pengaruh-pengaruh tersebut kurang
jelas ketika kantong habitat tenggelam dalam suatu matrix lanskap patch-
patch dengan berbagai kualitas lingkungan dan jarak antar patch.
Suatu konsep yang berhubungan dengan oceanic maupun
continental adalah laju penghunian (occupancy rate). Laju penghunian
suatu spesies dan kekayaan spesies dalam pulau-pulau, seperti juga dalam
patch-patch pada lanskap umumnya dipengaruhi tidak hanya oleh luas.
Sebagai contoh, Howe et al. (1981) melaporkan bahwa sekitar 75% spesies
burung di hutan hujan yang luas yang diteliti di New South Wales juga
teramati di hutan-hutan sisa (remnant forest). Spesies yang tidak tercatat
di remnant forest juga jarang di hutan hujan atau memiliki home range
yang luas sehingga tidak didukung o leh fragment-fragment yang terisolasi.
Ketidak hadiran spesies langka pada fragment-fragment tersebut mungkin
disebabkan oleh pengaruh kombinasi ukuran fragment yang kecil dan
isolasi, juga mungkin pengaruh sampling. Karena ukuran populasi yang
rendah, peluang kepunahan spesies langka di dalam fragment-fragment
tersebut mungkin tinggi, dan laju kolonisasi ke dalam fragment-fragment
tersebut dari sumber spesies di hutan yang lebih luas adalah rendah.
Dilaporkan juga beberapa spesies burung yang biasa di desa terbuka
ditemukan di remnant forest dan tidak di hutan yang luas, hal ini
membuktikan bahwa remnant forest berisi spesies edge dan suksesi awal
(Morrison et al. 1992).
Konsep relevan lainnya adalah prinsip founder (founder principle).
Prinsip ini menyatakan bahwa suatu kumpulan individu yang bermigrasi

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
67
dapat memulai sebuah sub populasi lokal pada suatu areal yang sebelumnya
tidak berpenghuni. Berkaitan dengan prinsip founder adalah rescue effect
yang terjadi ketika seekor imigran mengisi suatu patch-patch tidak
berpenghuni tetapi sebelumnya berpenghuni (Brown & Brown 1977).
Memahami bagaimana segment suatu populasi, seperti nonbreeding floaters
dapat berperan sebagai founder dan penyelamat adalah penting dalam
mendisain lanskap.
Lomolino (1984) melaporkan bahwa rescue effect mungkin
mempengaruhi distribusi dalam pulau (insular distribution) dua spesies vole
(Microtus pennsylbanicus dan Blarina brevicauda) di wilayah pulau seribu
sungai St. Lawrence, New York. Distribusi spesies di pulau-pulau tersebut
dihasilkan dari seleksi imigrasi – seleksi untuk untuk fenotip lebih vagile
dari populasi di dalam daratan sumbernya – dan tidak adanya predator di
pulau-pulau tersebut.
Jika lingkungan terganggu pada laju yang lebih cepat dari pada
yang dapat direspon oleh populasi secara demografik, maka faunal
relaxation penurunan kekayaan spesies atau penghunian pulau atau patch-
patch bisa terjadi. Faunal relaxation terjadi di dalam pulau atau lanskap
yang mengalami isolasi. Penurunan terjadi karena laju kolonisasi menurun
tetapi laju kepunahan lokal tidak. Relaxation tejadi dengan isolasi
mendadak dari lingkungan yang kaya (Morrison et al. 1992).
Faunal relaxation mungkin merupakan penyebab penurunan atau
hilangnya tujuh spesies burung yang menjadi ciri hutan tua di fragment
hutan oak-hemlock, di arboretum Connecticut antara tahun 1853 dan 1976.
Meskipun demikian, pada beberapa tahun belakangan ini, fragment-
fragment telah menjadi kurang terisolasi seiring suksesi vegetasi yang
meluas di areal-areal sekitarnya yang menghasilkan kondisi yang kondusif
untuk mendukung kembali spesies burung-burung hutan tua (Askin dan
Philbrick, 1983). Faunal relaxation mungkin sekarang tejadi di beberapa
taman nasional (Newmark 1986), walaupun bukti-buktinya diperdebatkan.
Faunal relaxation juga dapat terjadi di alam liar setelah 50 tahun
mendatang, seiring hutan tua menjadi berkurang pada lahan hutan produksi
kayu komersial, jika kesinambungan patch tidak dibuat (Morrison et al.
1992).

PRAGMENTASI HUTAN
68
Hasil dari faunal relaxation adalah kepunahan populasi lokal atau
terjadinya populasi relict (populasi bertahan hanya di sedikit lokasi).
Populasi relict tampaknya akhir-akhir ini terjadi di lokasi-lokasi yang
terasing dan terputus dan mungkin merupakan hasil dari kondisi lingkungan
sebelumnya. Kejadian dan distribusi populasi relict mungkin tidak
mencerminkan kondisi lingkungan saat ini maupun kemampuan disperasl
dari spesies. Dengan demikian, akan menjadi salah bila mendefinisikan
kebutuhan manajemen habitat dan lanskap berdasarkan studi pola kejadian
sekarang dan asosiasi lingkungan dari populasi relict tersebut (Morrison et
al., 1992).
Secara umum, pola kehadiran spesies pada patch-patch benua dan
lanskap, seperti halnya pada pulau-pulau samudera, merupakan hasil
turunan dari proses kolonisasi dan kepunahan. Hal ini pada gilirannya
adalah fungsi dari faktor-faktor seperti kemampuan dispersal spesies,
sejarah hidup dan karakteristik demografik; variasi dari kondisi iklim dan
sumberdaya lokal; ukuran patch dan spacing; perubahan dalam koridor
antar pacth dan tipe lingkungan yang digunakan untuk dispersal; dan
kehadiran kompetitor, predator, parasit dan penyakit (Morrison et al.,
1992).
IV. KESETIMBANGAN (EQUILIBRIUM) POPULASI ANTAR KANTONG HABITAT
(HABITAT PATCH) DAN SUB POPULASI
Titik dimana suatu spesies menempati kantong-kantong habitat di
dalam dan di antara lanskap bergantung dinamika pada dua skala;
kesetimbangan antara kolonisasi dengan kepunahan antar patch-patch lokal
dalam lanskap tertentu, dan kesetimbangan antar imigrasi (spesies yang
masuk) dengan emigrasi (spesies yang keluar) dan kematian antar sub
populasi dari suatu metapopulasi. Dua skala ini overlap antar spesies
dengan berbagai ukuran tubuh, areal yang digunakan, dan struktur
populasi. Tetapi prinsip utamanya adalah keseimbangan (balance) antara
laju kolonisasi dan kepunahan. Dengan demikian, kesetimbangan
(equilibrium) populasi merupakan suatu fungsi dari dinamika dispersal,
migrasi musiman, dan pergerakan (movement) lainnya dan menggambarkan

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
69
suatu keseimbangan (balance) antara rekruitmen dan kehilangan
demografik (Morrison et al. 1992).
Temple & Cary (1988) mengembangkan suatu model simulasi
komputer dari suatu lanskap yang berisi populasi hipotetik burung-burung
hutan interior yang fekunditasnya berhubungan negatif dengan
kedekatannya pada suatu edge dalam lanskap. Model ini
mendemonstrasikan jika beberapa spesies yang menunjukkan fekunditas
yang lebih rendah di dekat edge, kemudian persistensi mereka dalam suatu
lanskap dapat dikendalikan dengan mengatur jumlah keberadaan edge.
Dengan suatu populasi yang sederhana, bila emigrasi dari wilayah lain yang
reproduksinya lebih baik sama dengan nol atau sedikit, maka populasi yang
bergantung pada interior dalam lanskap yang sangat terfragmentasi dapat
menjadi punah secara lokal.
Dalam suatu model dinamik patch yang serupa, Fahrig dan Merriam
(1985) mengukur survival dari suatu populasi tikus kaki putih (Peromyscus
leucopus) di antara kantong-kantong hutan (forest patches) dalam suatu
lanskap pertanian. Model tersebut memprediksi bahwa tikus dalam areal-
areal berhutan yang terisolasi akan memiliki laju pertumbuhan lebih rendah
dan akan lebih rawan terhadap kepunahan lokal dari pada di dalam areal-
areal berhutan yang berkesinambungan. Prediksi ini telah diverifikasi
dengan observasi empiris terhadap populasi di alam liar (Morrison et al.
1992).
DAFTAR PUSTAKA
Askin, R.A. and M. Philbrick. 1983. Changes in Bird Community as a Forest
Fragment Becomes Less Isolated. Abstract of a paper presented at the 1983 meeting of the American Ornitologist’ Union, August, in
New York.
Brown, C.W. and A.K. Brown. 1977. Turnover Rates in Insular
Biogeography : Effects of Immigration on Extinction. Ecology 58 : 445-449.
Dueser, R.D. and J.H. Porter. 1986. Habitat Use by Insular Small Mammals. Relative Effects of Competition and Habitat Structure. Ecology 67 :
195-201.
Fahrig, L. and G. Merriam. 1985. Habitat Patch Connectivity and
Population Survival. Ecology 66 : 1762-1768.

PRAGMENTASI HUTAN
70
Howe, R.W., T.D. Howe and H.A. Ford. 1981. Bird Distribution on Small Rainforest Remnants in New South Wales. Australian Wildlife
Research 8 : 637-651.
Johnson, N.K. 1975. Controls of Number of Bird Species on Montane Islands
in Great Basin. Evolution 29:545-567.
Johnson, N.K. 1978. Patterns of Avian Geography and Speciation in the
intermountain. In Intermountain Biogeography : A Symposium. Pp.137-158. Great Basin naturalist Memoirs No. 2.
Karr, J.R. and K.E. Freemark. 1985. Disturbance and vertebrates : An Integrative Perpective. In S.T.A.Picket and P.S.White (eds). The
Ecology of Natural Disturbance and Patch Dynamics. Pp. 153-168. Academic Press. Orlando, Fla.
Lande, R. and G.F.Barrowclough. 1987. Effective Population Size, Genetic Variation, and Their Use in Population Management. In M.E.Soule
(ed). Viable Population for Conservation. Pp.189. Cambridge Univerity Press. Cambridge.
Lomolino, M.V. 1984. Immigrant Selection, Predation, and the Distributions of Microtus pennsylvanicus and Blarina brevicauda on Islands. American Naturalist 123 : 468-483.
Martin, T.E. 1981. Limitation in Small Habitat Islands : Chance or Competition? Auk 98 : 715-734.
Morrison, M.L., B.G. Marcot and R.W. Mannan. 1992. Wildlife-Habitat Relationship: Consepts and Applications. The University of
Wisconsisn Press. Madison, Wisconsin.
Newmark, W. 1986. Mammalian Richness, Colonization, and Extinction in
western North American National Parks. Ph.D. thesis University of Michigan, Ann Arbor.
Pickett, S.T.A. dan J.N. Thomson. 1978. Patch Dynamics and The Design of Nature Reserves. Biological Conservation 13 : 27-37.
Rothstein, S.I., J. Verner and E. Stevens. 1980. Range Expansion and Diurnal Changes in Dispersion of the Brown-Headed Cowbird in the
Sierra Nevada. Auk 97 : 253-267.
Schoener, T.W. and A. Schoener. 1983. The Time to Extinction of a
Colonizing Propagule of Lizards Increases with Siland Area. Natural Resources Conference 45 : 245-251.
Temple, S.A. dan J.R. Cary. 1988. Modeling Dynamcs of Habitat-Interior Bird Populations in Fragmented Landscapes. Conservation Biology 2
: 340-347.
Wilcove, D.S., C.H. McLellan and A.P. Dobson. 1986. Habitat
Fragmentation in the Temperate Zone. In M.E.Soule (ed). Conservation Biology. Pp. 237-256. Sinauer Associates.
Sunderland, Mass.

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
71
7
PENGARUH FRAGMENTASI TERHADAP SATWALIAR
PENDAHULUAN
KERUSAKAN HABITAT
MENGURANGI DAYA HIDUP (VIABILITY)
PENGARUH PADA POPULASI
PENGURANGAN LUAS HABITAT
PERUBAHAN KEANEKARAGAMAN HAYATI
PENGARUH ISOLASI PATCH
EFEK TEPI (EDGE EFFECT)
SPESIES, RELUNG MAKAN DAN FRAGMENTASI
FRAGMENTASI HABITAT DAN KEPUNAHAN
PEMANGSAAN DAN FRAGMENTASI
FRAGMENTASI HABITAT DAN PERILAKU SATWA
DAFTAR PUSTAKA
I. PENDAHULUAN
Fragmentasi habitat merupakan proses perubahan lingkungan yang
penting dalam evolusi dan biologi konservasi. Fragmentasi menggambarkan
terjadinya ketidaktersambungan/ discontinuity (fragmentasi) dalam
lingkungan (habitat) suatu organisme. Fragmentasi habitat dapat
disebabkan oleh proses-proses geologi yang secara perlahan mengubah tata
letak lingkungan fisik atau oleh kegiatan manusia seperti konversi lahan
yang dapat mengubah lingkungan dengan lebih cepat. Proses geologis

PRAGMENTASI HUTAN
72
dianggap sebagai penyebab utama terjadinya spesiasi, sedangkan kegiatan
manusia dianggap sebagai penyebab kepunahan banyak spesies
(http://en.wikipedia.org/wiki/Habitat_fragmentation).
Fragmentasi habitat seringkali disebabkan oleh manusia ketika
vegetasi alami ditebang untuk kegiatan manusia seperti pertanian,
pengembangan pedesaan atau perkotaan. Habitat-habitat yang sebelumnya
kontinyu menjadi terpecah ke dalam fragment-fragment yang terpisah.
Setelah penebangan yang intensif, fragment yang terpisah cenderung
menjadi ”pulau-pulau” yang sangat kecil dan terisolasi satu sama lain oleh
lahan budidaya, peternakan, jalan pengerasan atau bahkan lahan terbuka.
Lahan terbuka seringkali merupakan hasil dari sistem pertanian tebas-bakar
(slash and burn) di hutan-hutan tropika. Di jalur lahan gandum di New
South Wales Tengah bagian barat, Australia, 90% dari vegetasi alami telah
ditebang dan mengakibatkan fragmentasi habitat yang ekstrim
(http://en.wikipedia.org/wiki/Habitat_fragmentation).
Fragmentasi habitat dapat mencakup enam proses diskrit yaitu
(http://en.wikipedia.org/wiki/Habitat_fragmentation):
Pengurangan luas total habitat
Meningkatnya jumlah habitat pinggiran (edge)
Menurunnya jumlah habitat bagian dalam (interior)
Isolasi fragment habitat yang satu dari fragment habitat lainnya
Memecah satu kantong habitat (habitat patch) menjadi beberapa
kantong (patch) yang lebih kecil.
Menurunnya luas rata-rata dari setiap kantong habitat (habitat patch).
II. KERUSAKAN HABITAT
Salah satu cara utama bagaimana fragmentasi habitat
mempengaruhi keanekaragaman hayati adalah mengurangi jumlah habitat
yang tersedia (available) bagi tumbuhan dan satwa seperti hutan hujan,
hutan lainnya, samudera dan lain-lain. Fragmentasi habitat selalu
menyebabkan sejumlah kerusakan habitat. Tumbuhan dan organisme lain
yang tak dapat pindah (sessile) dalam areal ini biasanya langsung punah.
Satwa yang dapat bergerak (khususnya burung dan mamalia)

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
73
menyelamatkan diri ke kantong habitat yang tersisa. Hal ini dapat
menyebabkan terjadinya pengaruh kerumuman (crowded effect) dan
meningkatkan kompetisi. (http://en.wikipedia.org/wiki/Habitat_fragmentation).
Gambar 12. Contoh habitat yang terfragmentasi oleh: (1) jaringan listrik,
(2) jalan raya, (3) tebang habis dan (4) pertanian (Foto: Hendra Gunawan, 2008).
Fragment habitat yang tersisa selalu lebih kecil dari habitat
aslinya. Spesies yang tidak dapat berpindah antar fragment harus bertahan
dengan sumberdaya yang ada pada fragment tunggal dimana mereka
menghabiskan sisa hidupnya. Karena satu dari penyebab utama kerusakan
habitat adalah pembangunan pertanian, fragment habitat jarang mewakili
lanskap semula (http://en.wikipedia.org/wiki/Habitat_fragmentation).
1
1
2
4
3
2
3

PRAGMENTASI HUTAN
74
III. MENGURANGI DAYA HIDUP (VIABILITY)
Luas area merupakan petunjuk utama jumlah spesies di dalam
sebuah fragment. Ukuran fragment akan mempengaruhi jumlah spesies
ketika fragment terjadi, dan akan memepengaruhi kemampuan spesies
tersebut untuk bertahan di dalam fragment itu. Fragment-fragment
habitat yang kecil hanya dapat mendukung populasi-populasi kecil
tumbuhan dan satwaliar dan populasi-populasi kecil lebih rentan terhadap
kepunahan. Fluktuasi kecil dalam iklim, sumberdaya atau fakor-faktor lain
yang dianggap tidak penting dan diharapkan bisa cepat terkoreksi dalam
populasi-populasi besar dapat menjadi kastatropik dalam populasi-populasi
kecil dan terisolasi. Dengan demikian fragmentasi habitat merupakan suatu
penyebab penting dari kepunahan spesies. Dinamika dari populasi -populasi
yang telah terbagi cenderung tidak sama satu sama lain
(http://en.wikipedia.org/wiki/Habitat_fragmentation).
Dalam sebuah lanskap yang tidak terfragmentasi, penurunan
populasi dapat diselamatkan oleh imigrasi dari limpahan populasi di
dekatnya. Dalam lanskap terfragmentasi, jarak antar fragment dapat
mencegah hal tersebut terjadi. Sebagai tambahan, fragment-fragment
habitat yang tidak dihuni yang terpisahkan dari suatu sumber pengkoloni
oleh beberapa penghalang memiliki peluang yang kecil untuk dihuni
kembali (repopulated) dari pada fragment-fragment yang tersambung.
Bahkan spesies kecil seperti katak totol Colombia tergantung pada rescue
effect. Penelitian menunjukkan 25% juvenil menempuh jarak lebih 200m
dibandingkan dengan 4% dewasa. Sisanya, 95% di lokasi baru mereka,
menunjukkan bahwa perjalanan ini diperlukan untuk bertahan hidup.
Dispersal yang tinggi pada amfibia lainnya mengindikasikan bahwa spesies
langka ini terancam kepunahan dalam kelasnya
(http://en.wikipedia.org/wiki/ Habitat_fragmentation).
Sebagai tambahan, fragmentasi habitat membawa pada edge
effect. Perubahan iklim mikro dalam penyinaran, temperatur dan angin
dapat mengubah ekologi di sekitar fragment, di bagian interior dan
eksterior fragment. Kebakaran menjadi lebih berpeluang terjadi seiring
merosotnya kelembaban dan meningkatnya temperatur dan kecepatan
angin. Spesies eksotik dan hama dapat berkembang dengan sendirinya lebih

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
75
mudah dalam lingkungan yang terganggu tersebut, dan hewan ternak yang
berdekatan sering mengganggu ekologi alami. Demikian juga, habitat
sepanjang edge dari fragment memiliki iklim berbeda dan menyenangkan
spesies yang berbeda dari habitat interior. Oleh karena itu, fragment-
fragment kecil tidak disukai oleh spesies yang membutuhkan habitat
interior (http://en.wikipedia.org/wiki/ Habitat_fragmentation).
IV. PENGARUH PADA POPULASI Fragmentasi hutan mempengaruhi populasi-populasi tumbuhan dan
satwa pada beberapa skala. Pada skala lanskap yang lebih besar seperti
Daerah Aliran Sungai (DAS) Teluk Chesapeake, populasi-populasi penghuni
hutan yang sebelumnya berkesinambungan te lah mengalami pemecahan
menjadi sub populasi yang lebih kecil menempati fragment-fragment hutan
yang tersisa. Ilmuwan percaya bahwa sub populasi-sub populasi ini dapat
berperan sebagai metapopulasi. Suatu metapopulasi merupakan kumpulan
dari populasi-populasi kecil yang menempati sejumlah kantong habitat
(Gambar 13). (http://chesapeake.towson. dulandscape/forestfrag/effects.asp).
Individu-individu secara tidak teratur berpindah antar patch, dan
populasi dapat menjadi punah di dalam patch individual sebagai akibat dari
peluang kejadian (chance events). Sebagai contoh, dua jantan dalam suatu
populasi dari enam betina dapat dimakan predator. Jika tidak ada jantan
lain yang imigrasi ke dalam patch tersebut dari patch lain, populasi di
dalam patch tersebut akan mengalami kepunahan. Populasi-populasi kecil
dianggap rawan mengalami tipe peluang kepunahan ini. Tetapi, karena
individu-individu secara tidak teratur berpindah antar patch, maka patch
yang kosong akhirnya akan dikolonisasi dan ditempati lagi di masa
mendatang (Gambar 14) (http://chesapeake.towson.edu/landscape/
forestfrag/effects.asp).

PRAGMENTASI HUTAN
76
Gambar 13. Sekumpulan patch hutan yang ditempati oleh populasi-populasi penghuni hutan. Tebal tipisnya panah menunjukkan laju pertukaran individu antar populasi (panah tebal menunjukkan lebh banyak pertukaran). Catatan : Laju pertukaran dikoreksi oleh seberapa dekat patch-patch dalam ruang (Sumber: http://chesapeake.towson.
edulandscape/forestfrag/effects.asp).
Jika laju kolonisasi pada patch-patch kosong lebih tinggi dari laju
kepunahan, metapopulasi akan bertahan. Hal ini karena ketika beberapa
patch mengalami kepunahan, yang lainnya dikolonisasi. Akibatnya,
perpindahan individu-individu antar populasi mengikat semua populasi ke
dalam suatu metapopulasi yang dapat bertahan dalam lanskap yang
terganggu.
Gambar 14. Penghunian (occupancy) sekumpulan patch hutan yang mendukung
suatu metapopulasi spesies penghuni hutan pada dua waktu yang berbeda. Patch hitam ditempati dan patch putih tidak ditempati. Catatan: penghunian pada patch individual berubah sepanjang waktu (seperti patch-patch dikolonisasi dan populasi-populasi di dalam patch punah), tetapi jumlah patch-patch yang ditempati tetap sama sepanjang waktu. Sumber: http://chesapeake.towson.edu/ andscape/forestfrag/effects.asp).

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
77
Masalah bagi perencana lahan adalah bahwa laju perpindahan
individu-individu antar patch ditentukan oleh seberapa dekat atau
bagaimana patch-patch itu terpisahkan. Patch-patch yang berjauhan dari
patch lainnya tidak akan bertukaran individu denga patch-patch lainnya,
dan populasi kecil yang tersisa dalam patch akhirnya akan mengalami
kepunahan. Patch-patch menjadi lebih terisolasi satu sama lain, laju
kolonisasi merosot ke titik dimana laju kepunahan lebih tinggi dari laju
kolonisasi, dan seluruh metapopulasi akan mengalami kepunahan. Hal ini
akan terjadi sebelum semua patch di dalam suatu lanskap hilang. Yang
perlu digaris-bawahi adalah bahwa patch-patch yang berdekatan satu sama
lain memberikan habitat yang lebih baik dari pada patch-patch yang
terisolasi walau memiliki ukuran yang sama
http://chesapeake.towson.edu/landscape/forestfrag/effects.asp).
Pada skala patch hutan individual, beberapa faktor mempengaruhi
nilainya sebagai habitat tumbuhan dan satwaliar. Secara umum, patch-
patch yang lebih besar mendukung lebih banyak spesies. Hal ini karena
patch hutan yang lebih besar memiliki lebih banyak variasi habitat dan
mendukung populasi yang lebih besar sehingga kurang rawan terhadap
peluang kepunahan. Lagipula, hanya patch-patch besar cenderung berisi
habitat yang cukup untuk mendukung spesies seperti mamalia yang lebih
besar yang membutuhkan areal yang lebih luas
(http://chesapeake.towson.edu/landscape/forestfrag/effects.asp).
Patch hutan individual juga dipengaruhi oleh kondisi di sekitarnya.
Pada tepi hutan (forest edge), angin dan sinar matahari mengakibatkan
kondisi yang lebih kering dari pada di dalam interior patch hutan. Tepi
hutan juga lebih mudah dicapai oleh predator dan parasit yang dapat
terjadi di lahan tetangganya atau areal yang terbangun di sekitarnya.
Sebagai contoh, kucing rumah yang membunuh burung kecil seringkali lebih
umum ada di tepi hutan yang dekat dengan pemukiman. Cowbird, yang
merupakan burung parasit sarang juga lebih umum terdapat di hutan yang
dekat dengan lahan terbuka dimana mereka biasa mencari makan. Cowbird
meletakkan telurnya di sarang burung lain (burung inang). Burung-burung
inang akan merawat telur-telur burung cowbird. Ketika telur menetas,
burung cowbird yang tubuhnya lebih besar akan berebut makanan dengan

PRAGMENTASI HUTAN
78
anak burung inangnya dan bahkan dapat mendorong anak inangnya keluar
dari sarang (http://chesapeake.towson.edu/landscape/forestfrag/effects.asp).
Beberapa spesies burung interior hutan tidak toleran terhadap
kondisi yang lebih kering atau terhadap predator dan parasit yang terjadi di
tepi hutan. Spesies ini hanya ada di pusat habitat dari patch hutan
(Gambar 15).http://chesapeake.towson. edulandscape/forestfrag/effects.asp).
Gambar 15.
Grafik menunjukkan hubungan antara kelembaban tanah dengan
edge dan core habitat dalam patch hutan. Banyak spesies burung tidak
dapat mentolerir kondisi tepi habitat (habitat edge) dan hanya ditemukan
di dalam core habitat (Sumber:http://chesapeake.towson.
edulandscape/forestfrag/ effects.asp).
V. PENGURANGAN LUAS HABITAT
Fragmentasi habitat, meskipun terjadi secara alami tetap saja
mengurangi luas total suatu habitat asli. Keller and Anderson (1992)
menyatakan bahwa kehilangan habitat absolut dari suatu habitat asli dan
pengurangan kepadatan sumberdaya sehubungan dengan fragmentasi
berpotensi mempengaruhi biota lebih dari faktor-faktor tunggal lainnya.
Fragmentasi habitat mempengaruhi flora dan fauna dari suatu ekosistem
dengan mengganti ekosistem alami dengan lanskap yang didominasi
manusia yang mungkin tidak cocok bagi sejumlah spesies asli tertentu.
Meskipun demikian, berbeda dengan samudera sebagai penghalang
geografik, matrix lanskap buatan manusia dapat dicapai/dilewati oleh flora
dan fauna, karena mereka dapat dengan mudah menyebar melewatinya,
jika tidak tinggal di dalamnya
http://learning.turner.com/efts/rforest/habfrag.htm).
Di sisi lain, lanskap buatan manusia dapat secara langsung
menyumbang pada kepunahan spesies di dalam “pulau habitat” (habitat

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
79
island) dengan menggangu keseimbangan ekosistem yang disukai oleh
spesies yang sangat beradaptasi dengan kondisi yang berubah. Sebagai
contoh, meningkatnya jumlah lanskap yang didominasi manusia
memungkinkan spesies tertentu untuk tumbuh secara fenomenal, tetapi
dapat membahayakan spesies yang secara ekslusif tergantung pada patch
habitat interior. Contoh yang sering dikutip adalah burung parasit
cowbird kepala cokelat (Molothrus ater) yang populasinya meningkat secara
dramatis sejak manusia mulai mengganggu lanskap pada skala luas di
Amerika Utara. Cowbird kepala cokelat merupakan parasit sarang, yaitu
mengganti telur-telur burung inangnya dan menggantikannya dengan
telurnya sendiri dan membiarkan burung inang yang tidak menyadarinya
mengerami dan membesarkannya. Peningkatan jumlah burung cowbird
tersebut berdampak negatif terhadap keberhasilan perkembangbiakan
banyak burung-burung penyanyi penghuni tetap hutan (Mayfield 1977).
Disamping gangguan keseimbangan ekosistem yang disukai oleh
spesies yang memiliki kemampuan adaptasi tinggi, kehilangan habitat
akibat fragmentasi dengan mudah dapat menyebabkan populasi dan jelajah
spesies interior berkurang. Saunders (1989) mencatat satu contoh yang
jelas, bagaimana perubahan areal yang luas dari habitat yang utuh menjadi
pulau-pulau yang terfragmentasi mempengaruhi avifauna. Ia meneliti
perubahan avifauna di jalur ladang gandum Australia bagian barat sebagai
akibat dari fragmentasi. Ia menunjukkan bahwa 41% burung-burung asli di
wilayah tersebut telah menurun kelimpahan dan jelajahnya sejak 1900-an
dan menunjukkan bahwa hampir semua perubahan ini secara langsung
diakibatkan oleh fragmentasi habitat dan penurunan kelimpahan vegetasi
alami. Walaupun beberapa spesies meningkat kelimpahannya, ia
menekanankan bahwa lebih banyak spesies yang dirugikan dari pada yang
diuntungkan.
Bentuk fragment hutan mempengaruhi luasan interior yang
terkandung di dalamnya, seperti diperllihatkan pada Gambar 16, dimana
luas dari seluruh fragment hutan contoh adalah sekitar 50 ha. Fragment
hutan mungkin cukup besar tetapi memiliki interior yang kecil karena
bentuknya linear. Bentuk fragment seperti lingkaran atau bujur sangkar
memiliki proporsi interior terbesar dibandingkan terhadap luas totalnya dan

PRAGMENTASI HUTAN
80
dapat memiliki hampir dua kali lipat luas interior fragment berbentuk
persegi panjang sempit dengan luas total yang sama
(http://www.lrconline.com).
Sumber: http://www.lrconline.com
Gambar 16. Berbagai bentuk fragment dan gambaran luas interior-nya.
Spesies yang meningkat kelimpahan dan jelajahnya ketika terjadi
fragmentasi habitat adalah spesies yang mampu beradaptasi. Dengan
perkataan lain, kebutuhan sumberdayanya dapat dipenuhi oleh berbagai
kondisi sehingga sering diuntungkan oleh kegiatan manusia yang mengurangi
kompetisi dengan spesies lain. Karena hal ini, spesies yang diuntungkan
oleh kegiatan manusia bukanlah spesies yang harus kita kelola dan lindungi.
Sebaliknya, kita perlu melindungi spesies yang berjuang sendiri untuk
bertahan dalam habitat yang terus menghilang dengan cepat (Harris 1984).
Kita dapat membuat prediksi skala lanskap tentang efek
kehilangan hutan dari efek isolasi skala patch, karena isolasi patch
merupakan fungsi khas dari jumlah hutan dalam suatu lanskap. Prediksi
skala lanskap berdasarkan pada efek ukuran patch tidaklah jelas karena
ukuran patch dapat dikurangi oleh kehilangan atau oleh kombinasi
kehilangan habitat dan fragmentasi (Gambar 17).

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
81
VI. PERUBAHAN KEANEKARAGAMAN HAYATI
Disamping mengubah fisik bagian habitat as li, berkurangnya luas
habitat as li dapat menurunkan keanekaragaman hayati suatu areal melalui
beberapa cara. Menurunnya keanekaragaman hayati suatu areal dapat
terjadi jika fragment habitat lebih kecil dari pada home range satwa yang
memiliki home range terbesar yang ada di dalam ekosistem utuhnya.
Banyak burung memiliki home range besar karena mereka membutuhkan
sumberdaya yang terdistribusi dalam patch-patch. Sebagai contoh,
sepasang burung pelatuk paruh gading (Campephilus principalis) yang
berkembangbiak membutuhkan hutan dataran bawah (bottomland) kontinyu
yang tak terganggu seluas 6,5-7,6 km2, dan Elang goshawk Eropa (Accipiter
gentilis) membutuhkan 30-50 km2 untuk home range-nya (Wilcove et al.
1986).
Jika fragment habitat yang ada lebih kecil dari pada luas minimum
yang dibutuhkan oleh suatu spesies, individu-individu spesies tersebut
berpeluang tidak akan ditemukan di dalam fragment habitat tersebut.
Sebagai contoh, sejenis burung wabler Lousiana waterthrush (Seirus
motacilla) jarang ditemui di tegakan pohon yang sempit karena mereka
membutuhkan perairan terbuka di dalam home range mereka, dan hampir
semua tegakan pohon yang sempit tidak memiliki sungai atau kolam yang
sepanjang tahun berair (Robbins, 1980). Jika suatu spesies membutuhkan
dua atau lebih tipe habitat, mereka sering dianggap dapat mengalami
kepunahan lokal akibat fragmentasi habitat, karena seringkali mereka tidak
dapat dengan bebas berpindah antara tipe habitat yang berbeda. Burung
blue-grey gnatcatcher (Polioptila caerulea) berpindah dari hutan deciduous
ke padang chaparral selama musim berkembangbiak, dan jika satu dari
kedua habitat tersebut tidak dapat dijangkau, mereka diduga kuat akan
mengalami kepunahan lokal (Wilcove et al. 1986).

PRAGMENTASI HUTAN
82
Gambar 17. Pengaruh kehilangan dan fragmentasi habitat pada ukuran
patch dan isolasi (Fahrig, 1997).
Hilangnya beberapa spesies dari suatu komunitas bisa memiliki
pengaruh sekunder yang beruntun di seluruh ekosistem. Sebagai contoh,
hilangnya predator puncak dari suatu areal karena fragment terlalu kecil
dapat menyebabkan meningkatnya omnivora kecil, yang pada gilirannya
nanti dapat menyebabkan tekanan pemangsaan yang berlebihan pada telur
dan anak burung-burung penyanyi, akhirnya menyebabkan penurunan
keberhasilan perkembangbiakan burung-burung tersebut.
Komunitas tropika sering dianggap kehilangan keanekaragaman
hayati dari pada komunitas daerah temperate karena spesies tropika
biasanya ditemukan dalam kepadatan yang lebih rendah dan tidak
terdistribusi secara luas, dan sering memiliki kemampuan dispersal yang
lebih lemah (Wilcove et al. 1986). Banyak spesies tropika terlibat dalam

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
83
hubungan mutualisme yang kompleks seperti hubungan tumbuhan-
penyerbuk, tumbuhan-penyebar biji, dan parasit-inang, sehingga tidak
dapat dihindari kepunahan lokal salah satu spesies menyebabkan kepunahan
spesies lainnya. Burung kasuari (Casuarius casuarius), pemakan buah di
hutan hujan di Australia, diduga kuat mengalami kepunahan lokal akibat
fragmentasi habitat karena kebutuhan habitatnya berupa hutan hujan
kontinyu yang luas berkaitan dengan hubungan mutualisme yang unik antara
keduanya yaitu tumbuhan-penyebar biji. Burung besar yang tak dapat
terbang ini mengembara secara nomaden dalam mencari musim berbuah.
Kasuari berfungsi sebagai satu dari sedikit penyebar biji -biji berukuran
besar, yang banyak diantaranya perlu dikupas dan dipecah (dicerna)
sebelum berkecambah. Kepunahan kasuari dari fragment hutan hujan tak
dapat dihindarkan akan membawa kepunahan pohon-pohon atau tumbuhan
yang tergantung padanya sebagai penyebar/pemecah biji.
Disamping menjadi rumah bagi spesies terancam punah, komunitas
tropika terancam kerusakan dan fragmentasi karena lokasi fisiknya overlap
dengan batas-batas geografis negara-negara dunia ketiga. Dalam negara-
negara ini, masyarakatnya seringkali terantung pada penghasilan yang
didapatkan dari kayu hutan hujan atau ternak yang dibudidayakan dengan
menebang habis hutan hujan. Tekanan yang konstan pada komunitas hutan
hujan ini mengakibatkan fragmentasi habitat yang sangat luas. Fragment-
fragment kecil terisolasi yang dihasilkan menyebabkan ganguan pada
keseimbangan ekosistem. Ahli ekologi Thiollay dan Myberg (1988) meneliti
status dari semua burung pemangsa (raptor) yang ditemukan tersisa di
habitat hutan hujan di pulau tropika Jawa, dimana hampir semua habitat
asli hanya tersisa di cagar-cagar alam. Hampir semua raptor sangat jarang
ditemukan di luar cagar-cagar alam sebagaimana telah diduga sebelumya.
Mereka juga menemukan bahwa semakin besar ukuran cagar, semakin padat
populasi raptor di dalamnya.
Menariknya, Lovejoy et al. (1986) menemukan fenomena yang
serupa dengan burung-burung Amazon dalam proyek dinamika biologi
fragment hutan (Biological Dynamics of Forest Fragments = BDFF;
sebelumnya Minimum Critical Size of Ecosystem) di Brazil. Tujuan utama
proyek BDFF adalah untuk menemukan bagaimana komunitas hutan hujan

PRAGMENTASI HUTAN
84
merespon setelah suatu ekosistem utuh terpecah menjadi fragment-
fragment dengan ukuran berbeda. Mereka menemukan suatu crowding
effect, dimana kelimpahan burung-burung dalam fragment hutan langsung
meningkat secara signifikan setelah terjadi deforestasi di areal
tetangganya. Meningkatnya jumlah burung disebabkan oleh migrasi burung -
burung dari areal yang baru saja ditebang habis ke dalam fragment hutan
tersebut. Crowding effect menurun dengan meningkatnya ukuran suatu
fragment hutan.
VII. PENGARUH ISOLASI PATCH
Isolasi kantong-kantong habitat (habitat patches) terjadi seiring
lanskap yang terus terfragmentasi. Areal-areal dari tipe yang sama menjadi
terisolasi, tidak hanya oleh jarak tetapi juga oleh ketidakcocokan karena
gangguan lingkungan, memposisikan tumbuhan dan satwa tanpa adaptasi
untuk perpindahan jarak panjang pada situasi yang tidak menguntungkan.
Ketidakmampuan spesies untuk berpindah antar kantong habitat
menyebabkan hilangnya variasi genetik dan keanekaragaman yang akhirnya
dapat mengakibatkan hilangnya spesies tersebut di dalam habitat yang
tefragmentasi itu. Seiring dengan lingkungan yang terganggu menjadi
meningkat ketidakcocokannya, bahkan spesies yang lebih mobile pun dapat
terhambat dalam perpindahan antar pulau habitat. Saat ini banyak
komunitas dan fragment-fragment ekosistem yang terpisah jauh dan sangat
berkurang luasnya sehingga banyak satwa gagal mempertahankan
populasinya. Komunitas-komunitas tersebut menjadi sistem tertutup
sehingga rawan menghadapi perubahan katastropik dari kejadian-kejadian
seperti penyakit, kekeringan, angin topan atau banjir.
Beberapa contoh paling penting isolasi yang dihasilkan dari
fragmentasi habitat atau komunitas dapat ditemukan adalah ada pada apa
yang tertingal dari ekosistem prairie. Ekosistem ini sekarang sangat
berkurang menjadi fragment-fragment kecil yang terisolasi dan terpencar
di tengah ”lautan” pertanian dan lahan yang tidak cocok lainnya bagi
banyak spesies prairie, khususnya invertebrata dan tumbuhan. Tanpa
koridor penghubung atau stepping stones, spesies-spesies tersebut

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
85
menghadapi ancaman inbreeding yang berlanjut pada penurunan jumlah
dan bahkan kepunahan dari kantong habitatnya.
Baik komunitas tropika maupun temperate menghadapi masalah
yang sama yaitu inbreeding dan kehilangan keanekaragaman genetik, yang
dihasilkan oleh sub populasi tumbuhan dan satwa yang terisolasi satu sama
lain akibat dari fragmentasi habitat. Jika ada jarak yang terlalu besar
antara dua fragment dan suatu spesies tidak dapat menyebar melintasi
areal di antaranya, populasi tersebut secara prinsip telah terpecah.
Inbreeding bisa terjadi jika sub populasi pada fragment yang ada kecil. Hal
ini tidak langsung didokumentasikan, tetapi potensial terjadi.
Hilangnya keanekaragaman genetik dapat terjadi bahkan tanpa
inbreeding dan hasilnya homosigositas pada gen-gen tertentu dapat
menyebabkan evolusi yang berakhir pada kematian suatu spesies (Soule
1986). Dalam rangka meningkatkan pertukaran antara fragment-fragment,
banyak manajer satwaliar mengaplikasikan koridor-koridor yang
menghubungkan dua atau lebih pulau-pulau habitat. Berdasarkan
penelitian MacClintock et al. (1977) koridor-koridor meningkatkan kekayaan
spesies burung-burung yang berkembangbiak dan meningkatkan perjalanan
satwa penyebar biji (Harris 1984). Koridor-koridor dapat mencegah
hilangnya keanekaragaman genetik dan memungkinkan dispersal spesies
antar fragment, tetapi hilangnya ekosistem asli menjadi fragment-
fragment terus saja terjadi. Berkurangnya luas ekosistem alami hanya
dapat merugikan spesies yang tergantung padanya untuk hidup.
VIII. EFEK TEPI (EDGE EFFECT)
Edge adalah bagian luar dari batas (boundary) suatu kantong
habitat (habitat patch). Efek tepi (Edge effect) adalah suatu kondisi
dimana habitat yang sesuai menjadi kurang sesuai atau sebaliknya bagi
suatu spesies karena bersebelahan dengan lahan bukan habitat (non
habitat). Degradasi habitat ini dapat terjadi karena pemangsaan dari
spesies yang hidup di luar patch, atau meningkatnya kompetisi dengan
spesies yang hidup di luar kantong habitat.

PRAGMENTASI HUTAN
86
Edge effect terjadi di dekat perbatasan dua atau lebih tipe
habitat yang berbeda. Edge effect menguntungkan bagi banyak spesies
tumbuhan dan satwa karena edge memberikan mereka manfaat dari dua
atau lebih tipe habitat untuk survival mereka. Tetapi banyak spesies lain
yang mendapatkan pengaruh negatif akibat terlalu banyaknya edge.
Konsentrasi banyak spesies dekat edge menyebabkan meningkatnya
kompetisi, pemangsaan dan parasitisme. Perambahan spesies eksotis
sangat berkaitan dengan dinamika edge. Di hutan-hutan, banyak spesies
eksotik masuk ke interior, bermula dari zona tergangu yang berkaitan
dengan gangguan yang ditimbulkan oleh manusia.
Satu aspek lain areal yang
harus dipertimbangkan adalah batas
(boundary) antara habitat yang
menjadi perhatian dan habitat-
habitat di sekelilingnya (Gambar 18).
Hal ini melahirkan pertanyaan
tentang edge effect, karena boundary
umumnya gradual, tidak tajam
http://faculty.plattsburgh.edu/thom
as. wolosz/extinction.htm.
Kita tertarik pada edge bagian luar (outer edge) atau zone edge
effect akan memiliki beberapa kondisi yang berbeda dengan kondisi di
dalam habitat. Jika kita membicarakan tentang boundary antara hutan dan
ladang, misalnya, edge effect dapat meliputi derajat yang lebih tinggi dari
penyinaran matahari di edge hutan, kecepatan angin yang lebih besar, dan
kondisi yang lebih kering dan teduh. Zona edge ini dapat berfungsi sebagai
jebakan ekologi dimana ia menggambarkan habitat yang diinginkan untuk
beberapa satwa, tetapi mungkin tidak diinginkan oleh sejumlah besar
Gambar 18.
Ilustrasi zona pengaruh tepi (edge effect) suatu pulau habitat
(Sumber:http://faculty.plattsburgh.edu/thomas.wolosz/ extinction.htm).

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
87
pemangsaan oleh satwa yang tinggal di habitat sekeli lingnya. Di antara
burung-burung, masalah umum sepanjang zona edge meliputi pemangsaan
sarang dan parasit terhadap anakan oleh spesies seperti burung cowbird.
IX. SPESIES, RELUNG MAKAN DAN FRAGMENTASI
Beberapa spesies sensitif terhadap ukuran habitat dan disebut
“area-sensitive” (sensitif terhadap luas). Dengan demikian spesies interior
hutan yang berkembagbiak seperti burung oven dan wabler Kentucky
menurun populasinya jika terjadi fragmentasi (Gibbs & Faarborg 1990).
Pengaruh fragmentasi hutan tropika pada komunitas kumbang
kotoran dan bangkai telah dipelajari oleh Klein (1989) di Amozonia Tengah.
Fragment-fragment memiliki spesies lebih sedikit, lebih jarang dan lebih
tersebar. Perbedaan ini merupakan bukti ketika fragment satu hektar
dibandingkan dengan hutan tak terganggu. Perpindahan kumbang
terinterupsi oleh penebangan habis dan hanya setelah hutan sekunder
tumbuh kembali, beberapa kumbang dapat berpindah di dalam fragment-
fragment, walaupun jaraknya hanya beberapa meter. Mungkin iklim mikro
dan khususnya pengeringan di batas tegakan kayu menciptakan kondisi
penghalang untuk kumbang-kumbang hutan bagian bawah (understorey)
(Farina 2000).
Peranan kumbang kotoran dan bangkai sebagai pemusnah larva
nematoda (cacing) dan parasit gastrointestinal lainnya dari vertebrata
adalah sangat penting dalam pengendalian penyakit. Dengan meningkatnya
fragmentasi hutan tropika dan konsekuensinya pengurangan jumlah
kumbang tersebut, kita dapat menduga akan terjadi peningkatan pe nyakit
vertebrata (Farina 2000).
Ketika pengaruh-pengaruh fragmentasi habitat diteliti pada skala
spesies tunggal atau kelompok spesies yang berkerabat/berhubungan,
adalah mungkin untuk menemukan hasil yang mengejutkan, menarik dan
tak terprediksi. Contohnya adalah kalajengking Cercophonius squama dan
Amphipoda, famili Tallitridae, sebagaimana dilaporkan oleh Margules et al.
(1994). Dalam waktu jeda 8 tahun yaitu 3 tahun sebelum fragmentasi dan 5
tahun setelah fragmentasi dalam plot yang diberi perlakukan dan terkontrol

PRAGMENTASI HUTAN
88
dari hutan kayu keras Australia yang didominasi oleh Eucalyptus,
fragmentasi tidak memberikan pengaruh pada kelimpahan kalajengking.
Berbeda untuk Amphipoda, yang menurun dengan jelas setelah fragmentasi,
penurunan lebih besar terjadi di sisa hutan yang kecil dibandingkan di sisa
hutan yang luas (Farina 2000).
Kemungkinan kalajengking, sebagai hewan purba menggunakan
perilaku membuat lubang selama periode kering, dan memiliki kemampuan
untuk menghindar dari tekanan lingkungan akibat fragmetasi. Hal ini tidak
dapat dilakukan oleh Amphipoda, yang lebih sensitif terhadap ik lim mikro
dan memiliki sejarah evolusi yang lebih muda. Ini merupakan i lustrasi yang
baik bahwa ketika kita mempelajari fragmentasi pada level spesies,
hasilnya bisa sangat berbeda sama sekali (Farina 2000).
Tindakan pengelolaan harus diambil sehubungan dengan tingkah
laku populasi dan skala lanskap dan populasi di dalam hutan-hutan sisa
(remnant), daripada hanya mempertimbangkan jumlah spesies.
Malacosoma disstria merupakan larva kupu-kupu yang berlindung di hutan
yang mengalami outbreak (meledak) selama periode berbeda dan
berhubungan dengan struktur hutan (Roland 1993). Jumlah edge hutan per
kilometer persegi merupakan penduga yang baik bagi periode outbreak.
Parasitoid mungkin kurang efisien dalam mengendalikan Malacosoma
disstria sepanjang fragment. Sebagaimana dengan banyak spesies
Lepidoptera, Malacosoma disstria meletakkan lebih banyak telurnya
sepanjang edge dan pemangsaan telur spesies ini tampaknya pada tingkat
yang lebih rendah daripada di dalam hutan interior (Farina 2000).
Sepanjang edge, kondisi iklim mikro yang lebih disukai dan
temperatur yang lebih tinggi menurunkan periode perkembangan larva,
sehingga mengurangi resiko pemangsaan dibandingkan de ngan larva di
tegakan yang kontinyu (Farina 2000).
Spesies yang sensitif terhadap fragmentasi habitat kurang efisien
dalam berpindah dan mengkolonisasi habitat baru, dan konsekuensinya
memiliki kemampuan dispersal yang lebih rendah (Villard & Taylor 1994;
Villard et al. 1995). Burung-burung understorey hutan tropis Afrika
khususnya sensitif pada fragmentasi (Newmark 1990). Spesies interior
hutan dan langka merupakan satu yang paling terkena pengaruh buruk oleh

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
89
pengurangan habitat, dan pemeliharaan koridor bisa menjadi langkah
penceghan kepunahan lokal (Farina 2000).
Serangga-serangga penyerbuk di hutan kering sub tropika di
Argentina bagian utara berkurang, menurunkan ukuran fragment-fragment,
tetapi sebaliknya lebah madu (Apis melifera) meningkat frekuensi
kunjungannya ke bunga (Aizen & Feinsinger 1994).
Walaupun diduga kuat adanya laju yang tinggi pada alien species
di tegakan hutan yang kecil dan terisolasi, penginvasi dihentikan oleh
konkurensi cahaya (rendah di interior) dan oleh isolasi tanaman budidaya,
digabung dengan kapasitas yang rendah alien species untuk berpindah.
Fragment-fragment tersebut memiliki edge dan vegetasi semak yang lebat
yang mencegah alien species masuk, walaupun pada waktu yang sama edge
yang hangat ini menarik alien species (Brother & Spingarn 1992).
X. FRAGMENTASI HABITAT DAN KEPUNAHAN
Pengaruh fragmentasi pada keanekaragaman jenis burung telah
didokumentasikan di hutan dataran tinggi San Antonio, Colombia oleh
Kattan et al. (1994). Di wilayah ini hasil sensus burung tahun 1911
dibandingkan dengan data tahun 1959, 1963 dan 1989-1990. Hilangnya 24
spesies atau 31% dari spesies burung-burung asli, merupakan informasi yang
relevan tentang pemiskinan (impoverishment) fauna di lanskap ini. Peneliti
yang melakukan sensus dan perbandingan terkini memberikan alasan bahwa
tingginya laju kepunahan terutama disebabkan oleh posisi banyak spesies
pada batas atas dari sebaran menurut ketinggiannya, dan kelompok yang
lebih rawan adalah burung pemakan serangga pada understorey dan burung
pemakan buah di tajuk-tajuk besar.
Kerja Kattan et al (1994) tersebut penting dari berbagai sudut
pandang, pertama karena sejarah penurunan keanekaragaman jenis burung
secara lengkap terdokumentasikan, kedua karena pengaruh fragmentasi
tergantung di bagian besar biogeografi spesies dan pada kompleksitas dari
struktur foraging.

PRAGMENTASI HUTAN
90
XI. PEMANGSAAN DAN FRAGMENTASI
Tegakan-tegakan hutan (woodlots) yang terisolasi umumnya
mengalami lebih banyak pemangsaan. Wilcove (1985) menemukan laju
pemangsaan yang lebih tinggi pada tegakan-tegakan hutan yang kecil dari
pada tegakan-tegakan hutan yang besar, tetapi pemangsaan juga lebih
tinggi di daerah sub-urban tetangganya dari pada di tegakan-tegakan hutan
yang terisolasi oleh lahan pertanian. Ketika sarang cawan terbuka
ditempatkan di tanah dan pada ketinggian 1-2 m di atas tanah, pemangsaan
ditemukan lebih tinggi di kedua kasus tersebut dari pada di sarang-sarang
percobaan berbentuk lobang. Mengingat kebanyakan burung-burung
penyanyi migran neotropikal membangun sarang-sarang cawan terbuka,
maka penurunan spesies ini yang disebabkan oleh pemangsaan bisa jadi
lebih dari yang diduga.
Pada kelompok-kelompok hutan besar di Virginia, pemangsaan pada
sarang buatan bervariasi dari 5 sampai 40%, tergantung pada banyak
variabel vegetasi dan tekanan komunitas pemangsa (Leimgruber et al.
1994).
Fragmentasi prairie dan marshland terus terjadi dengan sangat
cepat di beberapa wilayah di dunia, seperti di Canada, karena tekanan
dampak agroindustri modern. Walaupun banyak burung-burung air memiliki
kemampuan memulihkan diri (resilient cappacity) dari dampak kehilangan
habitat, namun hilangnya sebagian besar habitat alami tempat
berkembangbiak sangat meningkatkan resiko pemangsaan. Pasitschniak dan
Messier (1995) menggunakan sarang-sarang buatan yang ditempatkan pada
jarak yang berbeda, untuk mensimulasikan resiko pemangsaan di edge.
Resiko pemangsaan sarang itik liar berhubungan dengan jarak dari edge di
dalam cover sarang yang rapat, tetapi tidak ada pengaruh edge yang
teramati di dalam peternakan yang terbengka lai atau ladang pakan ternak
yang belum dipanen. Hal ini bisa jadi karena edge yang tiba-tiba antara
lahan budidaya tersebut dan aksesibilitas yang lebih besar untuk pemangsa.
Oleh karena itu, pada lanskap buatan manusia bisa jadi kurang penting
dibandingkan struktur vegetasi.
Argumen ini dipertanyakan oleh peneliti-peneliti lain. Pemangsaan
sarang di tanah pada fragment-fragment prairie di Missouri diteliti oleh

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
91
Burger et al. (1994). Sarang-sarang buatan di prairie < 15 ha lebih banyak
dimangsa daripada di sisa-sisa prairie yang besar (37% vs 13,9%). Sarang-
sarang yang ditempatkan pada jarak < 60 m dari tegakan hutan (woodlots)
kurang berpeluang untuk berhasil dari pada sarang-sarang buatan yang
ditempatkan lebih jauh (28,7% vs 7,9% dari pemangsaan).
Fragmentasi hutan-hutan holarctic (nearctic dan palearctic) telah
mengganggu dinamika banyak herbivora kecil dibandingkan dengan hutan
tak terganggu. Menghilangnya siklus dalam kelimpahan berpindah ke
selatan terutama disebabkan oleh meningkatnya tekanan dari pemangsa.
Andren et al. (1985) meneliti tekanan pemangsa terhadap tetranoidae
menggunakan sarang-sarang model. Tekanan pemangsa tinggi di selatan,
seperti yang telah diduga, dan pemangsa utamanya adalah corvidae yang
lebih melimpah di wilayah selatan. Burung-burung ini berkorelasi positif
dengan sistem pertanian, fragmentasi hutan dan dengan rejim gangguan
manusia yang lebih besar.
XII. FRAGMENTASI HABITAT DAN PERILAKU SATWA
Fragmentasi habitat mengubah beberapa aspek perilaku satwa,
seperti perpindahan dan pencarian makan. Respon mamalia kecil terhadap
fragmentasi telah diuji oleh Diffendorfer et al. (1995) pada tiga plot 0,5
hektar dengan patch yang lebih besar 5.000 m2, patch medium 288 m2 dan
patch kecil 32 m2 dari tahun 1984 sampai 1992; spesies yang diteliti adalah
tikus kapas (Sigmodon hispidus), tikus rusa (Peromyscus maniculatis) dan
vole (sejenis tikus) prairie (Microtus ochrogaster). Seperti yang sudah
diduga, satwa berpindah menempuh jarak yang panjang dan satwa dalam
proporsi yang lebih rendah berpindah, sehingga meningkatkan fragmentasi.
Burung hantu oranye kecokelatan (Strix aluco) merupakan
pemangsa nokturnal di Eropa, tampaknya sensitif terhadap fragmentasi
hutan (woodland). Peri laku foraging 24 ekor burung hantu yang diberi
radio tracking oleh Redpath (1992) telah dianalisis baik di hutan kontinyu
maupun di lahan pertanian dengan tegakan hutan (woodlots) yang
terpencar. Di dalam tegakan hutan yang terfragmentasi burung hantu
memiliki jarak antar perch (tempat istirahat/roosting) lebih jauh dan waktu

PRAGMENTASI HUTAN
92
perch (beristirahat/ roosting) yang lebih lama. Jantan memiliki waktu
perch yang lebih lama dari pada betina, dan jantan di hutan yang
terfragmentasi menggunakan waktu untuk terbang 40% lebih banyak dari
pada betina di hutan kontinyu. Dengan demikian fragmentasi sangat
berpengaruh terhadap aktifitas dan perilaku burung hantu.
Pada beberapa spesies, adaptasi terhadap fragmentasi bisa jadi
merupakan naluri/bawaan sejak lahir, seperti yang terjadi pada kambing
gunung penghuni padang pasir (Ovis canadensis). Spesies ini teradaptasi
untuk hidup di pegunungan yang terjal, dataran terbuka, terfragmentasi
alami, dan telah mengembangkan kemampuan yang tinggi di habitat
terfragmentasi dalam keadaan dimana koridor yang sesuai tersedia (Bleich
et al. 1990).
Persebaran (dispersion) mengurangi efek isolasi tetapi mendukung
penyebaran penyakit dan kematian yang lebih besar pada organisme yang
berpindah (Burkey 1995).
DAFTAR PUSTAKA
Aizen, M.A. dan P. Feinsinger. 1994. Habitat Fragmentation, Naive Insect Polllinators and Feral Honey Bee in Argentine, Chaco Serrano.
Ecological Application 4 : 378-392.
Andren, H., P. Angelstam, E. Lindtstrom, dan P. Widen. 1985. Difference
in Predation Pressure in Relation to Habitat Fragmentation : An Experiment. Oikos 45 : 273-277.
Bleich, V.C., J.D. Wehausen, dan S.A. Holl. 1990. Dessert Dwelling
Mountain Sheep : Conservation Implication of a Naturally Fragmented Distribution. Conservation Biology 4 : 383-390.
Brothers, T.S. dan A. Spingarn. 1992. Forest Fragmentation and Alien Plant Invasion of Central Indiana Old-Growth Forests.
Conservation Biology 6: 91-100.
Burger, L.D., L.W. Burger, J. Faaborg. 1994. Effects of Prairies
Fragentation on Predation on Artificial Nests. Journal of Wildlife Management 58 : 249-254.
Burkey, T.V. 1995. Extinction Rates in Archipelagoes : Implication for Population in Fragmented Habitats. Conservation Biology 9 : 527-
541.

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
93
Farina, A. 2000. Principles and Methods in Landscape Ecology. Kluwer Academic Publisher. Dordrecht, The Netherlands.
Gibbs, J.P. dan J. Faarborg. 1990. Estimating the Viability of Oven Bird and Kentucky Wabler Populastions in Forest Fragment.
Conservation Biology 4 : 193-196.
Harris, L.D. 1984. The Fragmented Forest. University of Chicago Press.
Chicago, IL.
http://chesapeake.towson.edu/landscape/forestfrag/effects.asp. Diakses
Tanggal 01-02-2008.
http://chesapeake.towson.edu/landscape/forestfrag/glossary.asp. Diakses
Tanggal 01-02-2008.
http://en.wikipedia.org/wiki/Habitat_fragmentation. Diakses Tanggal 01-
02-2008.
http://faculty.plattsburgh.edu/thomas. wolosz/extinction.htm. Diakses
Tanggal 01-02-2008.
http://learning.turner.com/efts/rforest/habfrag.htm. Diakses Tanggal 17-
10-2006.
http://www.brocku.ca/epi/lebk/ lebk.html. Diakses Tanggal 01-02-2008.
http://www.dnr.state. wi.us/org/land/er/biodiversity/concepts/
fragmentation.htm. Diakses Tanggal 01-02-2008.
http://www.Lrconline.Com. Conserving The Forest Interior: A Threatened
Wildlife Habitat. Diakses Tanggal 01-02-2008.
Kattan, G.H., H. Alvares-Lopez, M. Giraldo. 1994. Forest Fragmentation
dan Kepunahan Burung : San Antonio Eighty Years Later. Conservation Biology 8 : 138-146.
Keller, M.E. dan S.H. Anderson. 1992. Avian Use of Habitat Configurations Created by Forest cutting in Southestern Wyoming. Condor 94:
55-65.
Klein, B.C. 1989. Effect of Forest Fragmentation on Dung and Carrion
Beetle Communities in Central Amazonia. Ecology 70 : 1715-1725.
Leimgrurber, P., W.J. McShea, dan J.H. Rappole. 1994. Predation on Artificial Nests in Large Forest Blocks. Journal of Wildlife
Management 58 : 254-260.
MacClintock, L., R.F. Whitcomb dan B.L. Whitcomb. 1977. Island
Biogeography and “Habitat Island” of Eastern Forest. II. Evidence for The Value of Corridors and Minimization of

PRAGMENTASI HUTAN
94
Isolationin Preservation of Biotic Diversity. American Birds 31 : 6-16.
Mayfield, H. 1977. Brown-headed Cowbird : Agent of Extermination? American Birds 31 : 107-112.
Newmark, W. 1986. Mammalian Richness, Colonization, and Extinction in western North American National Parks. Ph.D. thesis University
of Michigan, Ann Arbor.
Pasutschniak, M. dan F. Messier. 1995. The Fragility of Ecosystems : A
Review. Journal of Applied Ecology 32 : 677-692.
Redpath, S.M. 1995. Impact of Habitat Fragmentation on Activity and
Hunting Behaviour in The Tawny Owl, Strix aluco. Ecology 6 : 410-415.
Robbins, C.S. 1980. Effects of Forest Fragmentation on Breeding Bird Populations in the Piedmont of the Mid-Atlantic Region. Atlantic
Naturalist 33 : 31-36.
Roland, J. 1993. Large Scale Forest Fragmentation Increases the Duration
of tent Caterpilar Outbreak. Oecologia 93 : 25-30.
Soule, M.E. 1986. The Fitness and Viability of Populations. Pp. 13-18 in M.E. Soule (ed). Conservation Biology : The Science of Scarcity
and Diversity. Sinauer Assoc. Inc., Sunderland, MA.
Thiollay, J.M. dan B.U. Meyburg. 1988. Forest Fragmentation and the
Conservation of Raptors on the Island of Java. Biological Conservation 44 : 229 – 250.
Villard, M.A. dan Taylor, P.D. 1994. Tolerance to Habitat Fragmentation Influences the Colonization of New Habitat by Forest Birds.
Oecologia 98 : 393-401.
Villard, M.A., G. Merriam, B.A. Maurer. 1995. Dynamics in Subdivided
Population of Neotripocal Migratory Birds in a Fragmented Temperate Forest. Ecology 76 : 27-40.
Wilcove, D.S. 1985. Nest Predation in Forest Tracts and the Decline of Migratory Songbirds. Ecology 66 : 1211-1214.
Wilcove, D.S., C.H. McLellan dan A.P. Dobson. 1986. Habitat Fragmentation in the Temperate Zone. Pp 237-285 in M.E. Soule
(ed). Conservation Biology : The Science of Scarcity and Diversity. Sinauer Assoc. Inc., Sunderland, MA.

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
95
8
PERPINDAHAN SATWA DALAM LANSKAP
POLA PERGERAKAN
SALTATORY MOVEMENT
HOME RANGE
DISPERSAL
MIGRASI
DAFTAR PUSTAKA
I. POLA PERGERAKAN
Suatu obyek berpindah di antara dua titik menunjukkan
pergerakan sinambung (continuous movement) jika kecepatanya tidak
pernah turun sampai nol, tetapi mungkin memiliki kecepatan yang tetap,
meningkat signifikan atau melambat. Teta pi umumnya suatu obyek
menunjukkan pergerakan melompat atau bertahap (saltatory movement),
yaitu berhenti sekali atau beberapa kali selama dalam perpindahannya
antara dua titik (Forman dan Gordon, 1986).
Jika kita memandang aspek spasial dari suatu lanskap, kita
temukan bahwa dua pola perpindahan menggambarkan dua tipe utama
aliran spesies, energi dan materi. Partikel, gas dan energi panas berpindah
secara kontinu melintasi suatu lanskap dalam aliran yang tetap (steady).
Banyak spesies menunjukkan perpindahan yang seragam (uniform)
sepanjang koridor atau melalui matrix lanskap. Perpindahan pada
kecepatan yang relatif konstan cenderung terjadi di areal-areal yang
heterogenitasnya rendah. Elemen-elemen lanskap seperti padang rumput,

PRAGMENTASI HUTAN
96
hutan atau jalan setapak yang disukai oleh spesies akan membuat
perpindahan kontinyu jika heterogen, karena tidak ada penghalang
(barrier) atau areal yang tidak cocok yang dapat menurunkan kecepatan
organisme yang berpindah (Forman & Gordon, 1986).
Percepatan atau perlambatan perpindahan bisa terjadi ketika satu
bagian rute relatif homogen dan bagian lainnnya heterogen. Seekor satwa
mulai dari areal heterogen dimana beberapa elemen lanskap tidak cocok
atau sulit ditinggali sehingga rata-rata bergerak dengan lambat (walaupun
mungkin juga bergerak cepat ketika melintasi tempat berbahaya seperti
jalan). Ketika ia memasuki lintasan (tract) yang lebih homogen atau lahan
yang cocok ditinggali, ia dapat berpindah melewati lintasan (tract) rata-
rata dengan lebih cepat (kecuali tract tersebut adalah tujuannya, dimana
satwa memperlambat mungkin untuk makan). Sebaliknya, seekor satwa
memulai di habitat homogen yang disukai seringkali memperlambat ketika
menghadapi lahan heterogen dengan patch-patch yang tidak disukai
(Forman & Gordon 1986).
Pengaruh-pengaruh dibalik pola perpindahan ini dapat
digambarkan atau diukur lebih tepat. Pada level resolusi pertama, kita
dapat mengukur derajat heterogenitas lanskap. Sebagai tambahan
terhadap ukuran heterogenitas, adalah bermanfaat untuk memperkirakan
derajat kontras yang ada di antara elemen-elemen lanskap yaitu bagaimana
perbedaan berbagai tipe yang dilewati satwa. Elemen-elemen lanskap
dapat dikategorikan pada suatu skala misalnya dari 1 sampai 10
berdasarkan kemiripannya dengan matrix (Forman & Gordon 1986).
Sebuah pendekatan lain dalam memahami pengaruh-pengaruh
dibalik pola perpindahan adalah dengan mempertimbangkan peran batas-
batas (boundaries) antara elemen-elemen lanskap. Batas (boundaries)
adalah tempat-tempat dimana satwa harus melewati dari satu elemen ke
elemen lainnya. Berbeda dengan konektivitas (connectivity) yaitu derajat
kemana suatu tipe elemen lanskap tertentu terpisah atau menyambung di
dalam lanskap, di sini kita menggunakan konsep yang agak mirip yaitu
frekuensi melintasi batas (boundary crossing frequency). Pada dasarnya
merupakan ukuran dari jumlah boundary per unit panjang dimana suatu
obyek melewatinya dalam perpindahannya melintasi lanskap. Satwa yang

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
97
menghadapi ruang yang lebih heterogen memiliki boundary crossing
frequency (BCF) yang lebih rendah dan mungkin berpindah lebih cepat
melintasi suatu areal. BCF dapat dikombinasikan dengan ukuran derajat
kontras elemen lanskap untuk memberikan perkiraan yang sangat tepat
kecepatan relatif dari satwa melintasi lanskap (Forman & Gordon 1986).
Kemudahan mengukur BCF menjadikannya digunakan dalam
perencanaan dan manajemen, karena dapat dihitung untuk sejumlah rute
alternatif yang mungkin antara dua titik. Dengan cara ini, sebagai contoh,
kita dapat langsung membandingkan sebuah garis lurus dengan suatu rute
menggunakan koridor dan dengan suatu rute yang menghindarinya. BCF
menjadi perhatian khusus dalam memahami perpindahan spesies interior,
termasuk spesies yang sulit ditemui (secretive) yang melintasi sedikit
boundary dan membutuhkan areal yang jauh dari edge (Forman & Gordon
1986).
II. SALTATORY MOVEMENT
Suatu obyek dapat berpindah untuk sementara, berhenti dan
berpindah lagi. Selagi ia berpindah melintasi suatu lanskap antara dua
titik, tempat-tempat tertentu sepanjang rute perpindahannya berfungsi
sebagai titik pemberhentian bagi obyek tersebut. Tipe perpindahan ini
disebut saltatory movement (perpindahan melompat) atau dalam hal
spesies disebut jump dispersal (pemencaran melompat) (Pielou 1979).
Jika hujan deras mengguyur ladang yang dibajak di atas bukit, air
mengalir melalui permukaan tanah membawa butiran-butiran partikel
(erosi), membawanya menuruni lereng bukit beberapa jauh dan
mengumpulkannya segera setelah hujan berhenti. Butiran partikel tetap
tinggal sampai hujan besar berikutnya turun yang akan mengerosi lagi dan
membawanya menuruni lereng bukit lebih jauh. Proses perpindahan dan
penghentian ini terus berlangsung di lereng bukit sampai butiran partikel
tertumpuk ke dalam sungai di bawah. Dalam contoh saltatory movement
ini, partikel memiliki sedikit interaksi dengan obyek-obyek sepanjang jalur
yang dilewatinya tetapi bisa memiliki interaksi yang signifikan pada titik -
titik pemberhentian, misalnya memberikan nutrisi mineral kepada

PRAGMENTASI HUTAN
98
tumbuhan pada titik-titik tersebut atau bisa menghaluskan tanah untuk
satwa atau biji. Dengan demikian nilai penting saltatory movement
terutama terletak pada seringnya interaksi utama antara obyek yang
berpindah dengan obyek-obyek pada titik pemberhentian. Pada continuous
movement, interaksi seperti ini sedikit, atau setidaknya tersebar sepanjang
rute dan tidak terkumpul pada titik-titik (Forman & Gordon 1986).
Contoh di atas juga menekankan bagaimana struktur lanskap
mempengaruhi aliran. Jika ladang tunggal yang dibajak sangat luas dengan
galur vertikal lurus menutupi bukit turun ke arah sungai, hujan besar
pertama dapat mengangkut partikel dalam jarak yang jauh, bahkan sampai
ke sungai. Di sisi lain, jika petani mengerjakan bajakan sesuai kontur, laju
air aliran permukaan dan erosi menjadi lambat, dan butiran tanah tidak
berpindah sampai jauh menuruni bukit, atau memiliki tanaman pagar
sejajar dengan kontur lereng bukit, butiran partikel berpindah dalam jarak
yang lebih pendek setiap kali hujan. Tiga perubahan yang dihasilkan dari
heterogenitas tambahan dalam struktur lanskap adalah (Forman dan Gordon
1986):
1. Perubahan partikel dari continuous movement ke saltatory movement.
2. Dengan bertambahnya jumlah pembehentian sepanjang rute,
bertambah pula interaksi antara partikel dengan lingkungan di
sepanjang rute,
3. Penurunan laju perpindahan dari bukit ke sungai – sebagai bagian dari
bertambahnya boundary yang dilewati – dari beberapa jam, barangkali
menjadi beberapa tahun atau bahkan abad.
Demikian juga, seekor satwa berpindah secara kontinyu
(continuous movement) melintasi suatu lanskap biasanya memiliki dampak
minimal pada lanskap tersebut, sementara pada satwa dengan saltatory
movement terdapat banyak interaksi sering terjadi pada pemberhentian.
Satwa tersebut mungkin banyak makan dedaunan (browsing) di tempat
tersebut, menginjak/merusaknya, memupuknya, bersarang di dalamnya
atau dimakan predator di tempat tersebut (Forman and Gordon 1986).
Dua kemungkinan interaksi ini memberikan dua tipe
pemberhentian penting yang dibuat oleh spesies yang menyebar. Ketika
individu satwa sampai pada suatu tempat (spot), tinggal untuk periode yang

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
99
singkat, dan berpindah lagi, tempat tersebut dapat disebut pemberhentian
untuk istirahat (rest stop). Sebaliknya, suatu tempat yang dikolonisasi oleh
suatu spesies – dimana spesies tersebut datang kemudian tumbuh dan
berkembangbiak dengan berhasil – disebut batu loncatan (stepping stone)
(Kimura & Weiss 1964; MacArthur & Wilson 1967). Pada beberapa kasus,
seekor individu mungkin tinggal untuk waktu yang lama pada rest stop
tanpa berkembangbiak, misalnya ketika individu mamalia membuat rumah,
atau suatu benih tumbuh menjadi tumbuhan dewasa tanpa menghasilkan
bunga dan buah. Apa yang penting dari dua tipe pemberhentian ini adalah
ketika suatu spesies menggunakan suatu tempat (spot) tertentu sebagai
stepping stone, spesies tersebut memperluas distrbusinya sebagai individu
yang berkembangbiak. Hal ini memberikan sumber baru untuk penyebaran
individu berikutnya. Sebaliknya, rest stop hanya lokasi sementara untuk
spesies. Penyebaran banyak spesies tumbuhan dari Amerika Selatan ke
utara melintasi Laut Karibia bisa terjadi karena adanya serangkaian pulau
sebagai stepping stones. Suatu penyebaran serupa terjadi melintasi
Samudera Pasifik, karena adanya beberapa stepping stones (Forman dan
Gordon, 1986).
III. HOME RANGE
Satwa di dalam suatu lanskap berpindah dengan tiga cara – di
dalam suatu home range, dalam dispersal, dan dalam migrasi (Swingland
and Greenwood, 1983). Home range seekor satwa adalah suatu areal
sekitar rumahnya (seperti sarang, tempat tinggal atau lubang) yang
digunakan untuk mencari makan dan kegiatan harian lainnya. Umumnya
sepasang satwa dan anak-anak mereka menggunakan suatu home range
bersama, walaupun ada beberapa spesies yang berbagi home range dalam
kelompok besar. Territory adalah suatu areal yang dipertahankan dari
masuknya individu lain dari spesies yang sama. Satwa-satwa memiliki
territory yang dipertahankan, home range mereka biasanya lebh besar,
karena mereka biasanya mencari makan di luar batas yang
dipertahankannya (Forman dan Gordon, 1986).

PRAGMENTASI HUTAN
100
IV. DISPERSAL
Dispersal satwa adalah perpindahan satu arah seekor individu dari
home range dimana ia dilahirkan ke home range yang baru. Home range
baru biasanya jauh dari asalnya, normalnya berjarak beberapa kali dari
diameter home range. Satwa-satwa dewasa muda (sub adult) yang
meninggalkan induknya dan membuat home range sendiri merupakan
sebagian besar dari satwa-satwa yang melakukan dispersal, walaupun ada
juga beberapa dewasa yang menyebar dengan cara ini. Dispersal bisa juga
memperluas distribusi suatu spesies secara keseluruhan. Sebagai contoh,
vole (sejenis tikus) padang rumput (Microtus pennsylvanicus) memperluas
jelajahnya secara cepat ke Illinois tengah (Amerika Serikat) ketika super
highway dibangun (Getz et al. 1978). Jelaslah, bahu jalan yang berumput
kontinyu sepanjang super highway berfungsi menjadi saluran dispersal.
Sebelumnya, dispersal dilakukan melewati koridor berumput dari sisi jalan
yang lebih kecil, namun adanya perkampungan menjadi penghalang yang
menghambat dispersal (Forman & Gordon 1986).
V. MIGRASI
Migrasi adalah siklus perpindahan satwa antar areal yang terpisah
yang digunakan selama musim yang berbeda. Spesies yang bermigrasi telah
teradaptasi dengan iklim dan kondisi lainnya yang berhubungan dengan
perubahan musim dan dengan demikian cenderung untuk menghindari
kondisi lingkungan yang tidak disukainya dan menggunakan lingkungan yang
disukainya. Contoh klasik adalah migrasi sejumlah besar burung-burung
antara wilayah lebih dingin dan lebih hangat dan kelompok besar caribou
yang bermigrasi antara tundra dan batas (edge) hutan boreal. Aliran
beberapa kelelawar dan kupu-kupu dari utara ke selatan dan sebaliknya
merupakan kasus yang sama menariknya yang disebut latitudinal migration.
Migrasi seperti ini umumnya melintasi beberapa atau banyak lanskap
(Forman & Gordon 1986).
Migrasi vertikal merupakan perpindahan spesies satwa antara
elevasi lebih tinggi dan elevasi lebih rendah pada suatu gunung yang umum
terjadi untuk menghindari kondisi tidak disukai dan memilih kondisi yang

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
101
disukai. Sebagai contoh di pegunungan Alpin, Swis, ibex Eropa (Capra ibex
L.) mencari makan di vegetasi alpine pada musim panas dan pada musim
dingin di padang rumput dengan elevasi yang lebih rendah. Banyak spesies
burung di Rocky Mountains berkembang biak di elevasi tinggi pada musim
panas dan di elevasi rendah pada musim dingin. Elk (Cervus canadensis),
rusa mule (Odocoileus hemionus) dan herbivora berkuku lainnya pada
musim dingin dalam kawanan yang besar berada di areal terbuka dengan
elevasi rendah. Pada awal musim panas kawanan tersebut pecah.
Kelompok-kelompok kecil mengikuti salju mencair sampai melalui beberapa
zona hutan ke padang-padang rumput di elevasi tinggi. Pada musim gugur
mereka turun lagi ke tempatnya mencari makan waktu musim dingin. Hal
yang sama terjadi pada beberapa lanskap pegunungan, domba domestik
berkelompok melewati sistem lembah dari elevasi rendah pa da musim
dingin ke wilayah elevasi tinggi pada musim panas (Forman & Gordon 1986).
Foto: Doc. Hendra Gunawan
Gambar 19. Koridor harus cukup lebar untuk memberikan manfaat positif
lebih banyak bagi satwaliar. Koridor riparian yang sempit seperti yang tampak dalam gambar berpeluang menghasilkan
dampak negatif bagi satwaliar.

PRAGMENTASI HUTAN
102
Beberapa perpindahan satwa musim dingin-musim panas dapat
dianggap sebagai transisi antara migrasi dan penjelajahan home range.
Home range spesies tertentu menyempit selama musim dingin ketika
penjelajahannya terhalangi oleh salju. Padahal, beberapa spesies
membutuhkan home range musim dingin yang lebih luas karena distribusi
makanan yang jarang. Pada kedua kasus tersebut, tempat tinggal atau
sarang ada di dalam home range musim dingin dan musim panas. Spesies
lainnya berpindah ke habitat berbeda di dalam suatu lanskap pada musim
dingin dan panas dengan mudahnya, seperti migrasi vertikal. Sebagai
contoh di Maine, rusa (Odocoileus virginiana) terkumpul di rawa cedar
(Thuja, sejenis konifer yang selalu hijau) selama musim dingin karena rawa
ini memberikan persediaan makanan serta melindungi dari predator dan
cuaca. Pheasant (sejenis burung buruan) di Texas berpindah dari tekanan
basah (playas yaitu daerah datar yang mengering) pada musim panas ke
daerah tanaman musim sejuk pada musim semi, dan kemudian ke suatu
lajur tanaman campuran, biji kecil, dan tekanan basah di musim panas dan
gugur (Whiteside & Guthery 1983).
DAFTAR PUSTAKA
Forman, R.T.T. dan M. Gordon. 1986. Landscape Ecology. John Wiley and Sons. New York.
Getz, L.L., R. Cole dan D.L. Gates. 1978. Interstates Roadsides as Dispersal Routes for Microtus Pennsylvanicus. J. Mammal 59 : 208-212.
Kimura, M. dan G.H. Weiss. 1964. The Stepping Stone Model of Population Structure and the Decrease of Genetic. Correlation with Distance. Genetics 49 : 561-576.
MacArthur, R. H., and E. O. Wilson. 1967. The theory of island biogeography. Princeton University Press, Princeton, New Jersey, USA.
Pielou, E.C. 1979. Biogeography. Wiley. New York. Swingland, I.R. dan P.J. Greenwood (eds). 1983. The Ecology of Animal
Movement. Clarendon Press. Oxford. Whiteside, R.W. dan F.S. Guthery. 1983. Ring-Necked Pheasant Movement,
Home Range, and Habitat Us in Wst Texas. J. Wildlife Management 47 : 1097-1104.

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
103
9
FRAGMENTASI DAN DINAMIKA METAPOPULASI
Levins (1970) merupakan orang pertama yang menggunakan istilah
metapopulasi dan memperkenalkan konsep metapopulasi sebagai suatu
populasi yang tersusun atas populasi-populasi lokal yang dianggap akan mati
dan dikolonisasi lagi secara lokal. Ia memperkenalkan model matematik
untuk menggambarkan metapopulasi:
dp/dt = m p (1 - p) - e p ,
dimana p adalah proporsi (fraksi) pusat-pusat populasi (seperti pulau
habitat atau patch), m adalah laju migrasi (kolonisasi), dan e adalah laju
dimana populasi lokal menjadi punah.
Pada equilibrium p*=1 – e/m. Metapopulasi akan terjadi (yaitu p* > 0)
hanya jika e < m.
Suatu metapopulasi dapat hidup di suatu wilayah hanya jika laju
rata-rata kepunahan lebih kecil dari laju rata-rata migrasi. Populasi-
populasi lokal terbangun pada patch habitat tertentu yang bisa ditempati
atau kosong pada suatu waktu. Beberapa individu yang menyebar dapat
meninggalkan suatu patch pergi untuk mengkolonisasi suatu elemen kosong
atau mengisi kembali (reinforce) suatu populasi kecil. Populasi-populasi
yang terbentuk di dalam sebuah patch dapat menghilang megikuti kejadian
lingkungan (kebakaran, pohon tumbang) atau kejadian demografik
(epidemik, penuaan). Model Levins sangat sederhana. Ia memberikan
setiap patch ni lai yang sama sebagai sumber individu yang menyebar dan

PRAGMENTASI HUTAN
104
memiliki peluang kepunahan yang sama, dan peluang keberhasilan dispersal
adalah sama di semua patch (Burel dan Baudry, 2003).
Model dan konsep Levins telah menjadi metapopulasi klasik. Yang
lebih baru didefinisikan oleh Hanski dan Gilpin (1991), metapopulasi
didefinisikan sebagai sekumpulan (set) populasi-populasi lokal yang
berinteraksi melalui perpindahan individu antar populasi-populasi tersebut.
Hanski dan Simberloff (1997) mendefinisikan metapopulasi sebagai
sekumpulan (set) populasi-populasi lokal di dalam beberapa area yang lebih
besar, dimana ditandai migrasi dari satu populasi lokal ke beberapa patch
lain yang memungkinkan.
Dengan demikian, suatu metapopulasi minimal merupakan
sekumpulan populasi-populasi lokal yang relatif terisolasi, terdistribusi
secara spasial yang terikat bersama karena peristiwa dispersal antar
populasi. Peristiwa dispersal yang jaraknya relatif panjang mungkin tidak
sering, tetapi harus terjadi cukup sering untuk memberikan rekolonisasi
populasi yang telah mengalami kepunahan lokal. Metapopulasi regional
menghadapi kepunahan lokal dengan pasti karena dispersal yang cukup
antar populasi. Jika dispersal antar populasi begitu sering dimana
kepunahan lokal tidak terjadi, konsep metapopulasi tidak diperlukan
(berlebihan), dan populasi regional lebih baik dipandang secara sederhana
sebagai sebuah populasi tunggal yang terdistribusi secara spasial (walaupun
berkantong-kantong/patchily). Di sisi lain, jika dispersal terlalu jarang dan
peluang kepunahan lokal tidak nol, metapopulasi regional tidak dapat
berlangsung dan akan punah.
Sebuah metapopulasi biasanya dianggap terdiri dari beberapa
populasi berbeda bersama dengan areal-areal habitat yang cocok yang saat
ini tidak dihuni. Setiap populasi bersiklus dengan relatif bebas dari populasi
lainnya dan akhirnya menjadi punah sebagai akibat stokastiksitas demografi
(fluktuasi dalam ukuran populasi akibat kejadian demografik random);
populasi yang lebih kecil lebih rentan menghadapi kepunahan.
Walaupun populasi-populasi secara sendiri-sendiri memiliki masa
hidup terbatas, populasi secara keseluruhan seringkali stabil karena imigrasi
dari satu populasi (misalnya, yang mungkin mengalami ledakan populasi)
berpeluang merekonlonisasi habitat yang telah ditinggalkan populasi lain

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
105
karena punah. Mereka dapat juga emigrasi ke populasi kecil dan
menyelamatkan populasi tersebut dari kepunahan (disebut rescue effect).
Gangguan manusia pada lanskap seringkali menyebabkan
fragmentasi dan isolasi (insularization) dari habitat satwaliar yang pernah
kontinyu. Persamaan antara patcth-patch yang tidak kontinyu dari habitat
terfragmetasi dan populasi lokal yang relatif terisolasi dari suatu
metapopulasi adalah jelas. Paling tidak secara prinsip, dinamika populasi
dapat memungkinkan keberadaan spesies regional yang menempati suatu
jaringan patch atau pulau habitat yang tidak kontinyu. Tidak mengejutkan
bila biologi konservasi telah berpaling ke dinamika metapopulasi sebagai
solusi yang mungkin untuk keberlangsungan spesies di lanskap yang
terfragmentasi. Meskipun demikian, banyak asumsi yang diberikan untuk
kesederhanaan dan kebaikan model k lasik metapopulasi dari Levins (yaitu
ukuran dan ruang populasi yang sama) berpeluang terlalu membatasi
aplikasi pada spesies tertentu dan lanskap atau wilayah dimana ukuran
patch dan jarak antar patch bisa sangat bervariasi. Populasi pulau (insular)
saat kini terbentuk oleh fragmentasi habitat mungkin tidak berfungsi
sebagai metapopulasi menurut definisi klasik, tetapi mungkin lebih
menggambarkan suatu metapopulasi tidak setimbang (non equilibrium).
Sesungguhnya, mereka mungkin tidak mengambarkan metapopulasi sama
sekali, tetapi lebih sebagai kumpulan populasi terisolasi yang tidak saling
berinteraksi.
Dalam dinamika metapopulasi, individu populasi individual bisa
menjadi punah, tetapi mereka juga dapat dikolonisasi lagi dari populasi-
populasi lain (Gambar 20). Jika kita membuat populasi-populasi ini
menurun sampai jumlah yang cukup rendah dan tidak ada perpindahan
antara populasi-populasi tersebut, maka masalah genetik yang serius bisa
terjadi untuk melestarikan spesies. Walaupun sejumlah kecil perpindahan
antara populasi akan memelihara keadan genetik agak stabil (Barnes, 2000).
Pengembangan teori metapopulasi, dalam hubungannya dengan
pengembangan dinamika Source-Sink, menekankan pentingnya konektivitas
antara populasi-populasi yang tampaknya terisolasi. Walaupun tidak ada
populasi tunggal yang dapat menjamin survival jangka panjang dari suatu
spesies, pengaruh kombinasi dari banyak populasi dapat melakukan ini.

PRAGMENTASI HUTAN
106
Jika tidak ada perpindahan, populasi-populasi tersebut berpeluang
untuk punah, tergantung apakah mereka merupakan patch sumber (source)
atau patch penerima (sink) dari suatu metapopulasi. Patch-patch sumber
akan selalu tetap dalam kondisi lokalnya dan menyumbang individu-individu
ke semua patch lain di dalam lanskap. Patch-patch penerima (sink)
membiarkan populasi-populasi atau indvidu-individu menjadi punah karena
mereka tidak memiliki habitat yang kondusif bagi spesies untuk bertahan.
Contohnya seringkali terjadi dengan spesies satwaliar daratan, suatu patch
sumber habitat atau populasi akan penuh atau mencapai daya dukungnya
sehingga beberapa satwa harus berpindah ke patch sink. Walaupun patch
itu mungkin bukan habitat terbaik untuk beberapa spesies, misalnya
burung-burung mungkin dapat bersarang dan bertahan di sana (Barnes
2000).
Gambar 20. Metapopulasi adalah suatu populasi sumber (source) dan populasi penerima (sink) (Barnes 2000).

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
107
Manfaat lain dari populasi sink adalah jika beberapa kejadian
katastropik terjadi di satu patch sink dan memusnahkan anggotanya, patch
lainnya dapat membantu mengisi kembali populasinya (repopulate).
Dengan demikian, patch-patch sink membantu menstabilkan populasi dari
waktu ke waktu. Apa yang terjadi dalam satu patch tidak sama dengan apa
yang terjadi dalam patch-patch lainnnya (Barnes 2000).
Atribut penting lainnya dari metapopulasi adalah bahwa pada
beberapa kasus mereka sebenarnya dapat mencegah spesies dari kepunahan
karena patch-patch sumber dan patch-patch sink yang relatif terlindungi
dan terisolasi. Sebagai contoh, jika suatu penyakit di satu patch
memusnahkan semua individu di sana, karena tidak ada konektivitas antar
patch, maka tidak ada jalan penyebaran penyakit tersebut ke semua
populasi. Oleh karena itu, konsep metapopulasi memainkan peran yang
sangat penting dalam manajemen satwaliar saat ini. Sebuah contoh
memberikan gambaran pentingnya pemahaman metapopulasi ketika
mengelola satwaliar pada level lanskap. Bobwhite qail (sejenis burung
puyuh) di Amerika Utara masih ada dalam serangkaian metapopulasi. Saat
ini, mereka hidup dalam fragment-fragment habitat dengan beberapa
perpindahan satwa antar fragment. Kadang-kadang perpindahannya sangat
sedikit. Pada musim dingin 1976 dan 1977 di Kentucky, terjadi musim
dingin yang sangat dingin dengan banyak salju dan es yang mengakibatkan
beberapa patch quail kehilangan seluruh populasinya. Dari waktu ke
waktu, walaupun dengan perpindahan yang sangat sedikit, patch-patch
tersebut akhirnya berpopulasi lagi, walaupun memerlukan 20 tahun untuk
memulihkan beberapa populasi lokal (Barnes 2000).
Dalam kasus-kasus lain, perubahan habitat yang signifikan terjadi
sehingga menghambat quail untuk me-repopulasi beberapa areal, dan
menyebabkan populasi-populasi tersebut menjadi punah. Dalam kasus ini,
metapopulasi memungkinkan spesies untuk survive dan akhirnya
memulihkan populasi-populasi keci l melalui penyebarannya (Barnes 2000).
Seringkali ahli biologi membuat keputusan berdasarkan pada level
populasi pada saat ini. Tetapi, dengan dinamika metapopulasi, kita harus
mempetimbangkan kepunahan dan indikasi dari total areal yang potensial.
Seperti pada kasus quail yang memerlukan 20 tahun untuk merestorasi

PRAGMENTASI HUTAN
108
beberapa populasi lokal, walaupun masih banyak yang belum terpulihkan
dan mungkin tidak akan pernah terpulihkan karena degradasi habitat yang
terus berlangsung (Barnes 2000).
DAFTAR PUSTAKA
Barnes, T.G. 2000. Landscape Ecology and Ecosystems Management.
Cooperative Extension Services, University of Kentucky, College of Agriculture. UK. http://www.ca.uky.edu. Diakses Tangal 24
Februari 2007.
Burel, F., dan J. Baudry. 2003. Landscape Ecology : Consepts, Methods
and Application. Science Publishers, Inc. Enfield (NH), USA.
Hanski, I., and D. Simberloff. 1997. The metapopulation approach, its history, conceptual domain, and application to conservation. pp.
5–26. In I. A. Hanski and M. E. Gilpin (eds.), Metapopulation Biology. Academic Press, San Diego, Californina.
Hanski, I., and M. Gilpin. 1991. Metapopulation dynamics: brief history and conceptual domain. Biological Journal of the Linnean Society
42:3–16.
http://en.wikipedia.org/wiki/Habitat_fragmentation. Diakses Tanggal 01-
02-2008.
http://www.esd.ornl.gov/programs/SERDP/EcoModels/metapop.html.
Diakses Tanggal 06-02-2008.
http://en.wikipedia.org/wiki/ Metapopulation. Diakses Tanggal 01-02-2008.

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
109
10
IMPLIKASI PENGELOLAAN KONSERVASI
Dengan memahami dinamika metapopulasi dapat membuat
kebijakan pengelolaan konservasi spesies pada level lanskap lebih baik.
Seiring kita mengubah patch-patch habitat yang relatif seragam di dalam
matrix lanskap, perubahan fisik terjadi dan menciptakan efek pulau (island
effect) pada fragment-fragment yang tercipta. Perubahan-perubahan
tersebut meliputi: (1) penurunan ukuran patch; (2) peningkatan proporsi
edge dan (3) perubahan ik lim mikro pada patch, meliputi peningkatan
penyinaran matahari, fluktuasi temperatur yang lebih besar dan terpaan
angin yang lebih besar. Pengaruh-pengaruh tersebut dapat menyebabkan:
(1) kepunahan organisme secara lokal; (2) mengurangi dispersal dan re-
kolonisasi patch-patch habitat; (3) invasi spesies eksotis atau tidak asli; (4)
meningkatnya parasitisme sarang atau pemangasaan pada burung; dan (5)
menurunnya keanekaragaman spesies satwa interior hutan (Barnes 2000).
Walaupun dinamika metapopulasi dan konsep ekologi lanskap
rumit dan sulit dipahami oleh orang awam, kita harus berupaya memahami
konsep ini karena pentingnya konsep tersebut untuk menentukan keputusan
pada level lanskap dan pengelolaan satwaliar (Barnes, 2000).
Fragmentasi habitat seringkali merupakan penyebab spesies
menjadi terancam atau dalam bahaya kepunahan (endangered).
Keberadaan habitat yang viable adalah penting untuk survival spesies dan
dalam banyak kasus fragmentasi habitat yang tersisa dapat menyulitkan
pengambilan keputusan bagi para ahli biologi konservasi. Sumberdaya
tersedia yang terbatas untuk konservasi, lebih baik untuk melindungi patch-
patch habitat terisolasi yang ada atau untuk membeli kembali lahan untuk
mendapatkan lahan yang lebih luas dan kontinyu sebanyak mungkin.
Satu solusi untuk masalah fragmentasi habitat adalah
menghubungkan fragment-fragment dengan cara menanami koridor dengan

PRAGMENTASI HUTAN
110
vegetasi asli. Hal ini potensial untuk mencegah masalah isolasi tetapi tidak
mencegah kehilangan habitat interior. Dalam beberapa kasus suatu spesies
terancam mungkin mendapatkan keuntungan terlindungi dari penyakit
karena tersebar di habitat-habitat yang terisolasi.
Upaya pencegahan lainnnya adalah memperluas sisa habitat kecil
untuk meningkatkan jumlah habitat interior. Hal ini mungkin tidak dapat
dilakukan karena lahan-lahan terbangun seringkali lebih mahal dan
membutuhkan waktu dan usaha yang tidak sedikit untuk merestorasi.
Solusi terbaik umumnya tergantung pada spesies tertentu atau
ekosistem yang sedang menjadi perhatian. Spesies yang lebih mobile,
seperti burung tidak memerlukan habitat yang tersambung, sementara
beberapa satwa yang lebih kecil seperti pengerat mungkin lebih terbuka
terhadap pemangsaan di lahan terbuka.
Dari pemahaman konsep-konsep terkait dengan fragmentasi
habitat dan metapopulasi dapat dirangkum seperti disajikan pada Gambar
21 sebagai pertimbangan dalam pengelolaan populasi pada level lanskap
dan pengelolaan satwaliar (Barnes 2000).
Gambar 21. Ukuran, bentuk, konfigurasi dan jumlah patch, semua mempengaruhi jumlah habitat interior dalam patch kecil,
tunggal, patch-patch persegi memberikan jumlah habitat interior yang kecil dan patch-patch lingkaran besar
memberikan habitat interior terbesar (Barnes 2000).

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
111
DAFTAR PUSTAKA
Barnes, T.G. 2000. Landscape Ecology and Ecosystems Management. Cooperative Extension Services, University of Kentucky, College
of Agriculture. UK. http://www.ca.uky.edu. Diakses Tangal 24 Februari 2007.
http://en.wikipedia.org/wiki/Habitat_fragmentation. Diakses Tanggal 01-02-2008.

PRAGMENTASI HUTAN
112
11
STUDI KASUS: Evaluasi Lanskap Kawasan Merapi – Merbabu Dan Telaah Kemungkinan Sebaran Macan Tutul Di Kantong-Kantong
Hutan Yang Terfragmentasi1)
PENDAHULUAN
METODOLOGI
HASIL DAN PEMBAHASAN
SIMPULAN DAN SARAN
DAFTAR PUSTAKA
LAMPIRAN
I. PENDAHULUAN
A. Latar Belakang
Bentang lanskap Gunung Merapi – Gunung Merbabu merupakan
kawasan yang sedang terancam karena terkepung oleh daerah pemukiman
dan budidaya yang terus berkembang pesat akibat pertumbuhan penduduk
yang tinggi. Pelan tetapi pasti kedua gunung tersebut seperti dikuliti
vegetasinya dari kaki hingga mendekati puncaknya. Dalam skala lanskap
kedua gunung tersebut tampak seperti bertelanjang kaki, atau dengan
perkataan lain vegetasinya hanya mengumpul di sekitar puncak yang sulit
dijangkau sehingga belum dimanfaatkan.
Kawasan Merapi – Merbabu dikelilingi oleh beberapa kabupaten
yang sedang berkembang cepat seperti Sleman, Magelang, Klaten, Boyolali
1)
Tulisan pada Bab ini merupakan Paper Hendra Gunawan untuk Mata Kuliah Ekologi Lanskap
yang diasuh oleh Prof. Dr. Lilik Prasetyo, M.Sc. pada Program Studi Ilmu Pengetahuan Kehutanan, Sekolah Pasca Sarjana, Institut Pertanian bogor.

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
113
dan Semarang. Pertumbuhan penduduk yang tinggi dan perkembangan
pembangunan yang pesat telah menyebabkan ekspansi wilayah sampai ke
kaki Gunung Merapi dan Merbabu. Akibatnya kawasan ini mengalami
kehilangan habitat yang parah dan tingkat fragmentasi yang tinggi. Hal ini
tentu saja tidak menguntungkan bagi konservasi keanekaragaman hayati
dan perlindunguan sistem penyangga kehidupan yang menjadi peran utama
dari kawasan tersebut.
Mengingat pentingnya kawasan Merapi – Merbabu sebagai kawasan
perlindungan sistem penyangga kehidupan dan konservasi keanekaragaman
hayati, maka kedua Gunung tersebut telah ditetapkan sebagai taman
nasional sejak tahun 2004. TN. Merapi ditetapkan berdasarkan Keputusan
Menteri Kehutanan Nomor : 134/Menhut-II/2004, tanggal 4 Mei 2004
dengan luas luas ± 6.410 Ha dan TN. Merbabu berdasarkan Keputusan
Menteri Kehutanan Nomor : 135/Kpts-II/2004, tanggal 4 Mei 2004 dengan
luas ± 5.725 hektar.
Kawasan Merapi – Merbabu, dari tahun ke tahun terus mengalami
perubahan struktur, komposisi dan konfigurasi lanskapnya akibat adanya
gangguan dan perkembangan pembangunan yang tidak dapat dihindarkan.
Untuk mengetahui kondisi lanskap secara statistik, perlu dilakukan evaluasi
atau analisis spasial dari citra landsat.
B. Tujuan
Kajian ini bertujuan untuk mengevaluasi kondisi lanskap kawasan
Merapi-Merbabu pada tahun 2005 dan memprediksi kemungkinan sebaran
populasi macan tutul berdasarkan syarat kecukupan luas dan tipe vegetasi
di kantong-kantong habitat (habitat patches) yang ada.
II. METODOLOGI
A. Alat dan Bahan
Peralatan yang digunakan adalah seperangkat komputer (laptop)
dan printer. Software yang digunakan adalah ERDAS Imagine 8.5 dan
ArcView GIS 3.2. Bahan yang digunakan adalah citra landsat TM (tahun
2005), band 3, 4 dan 5 yaitu:
L7G120065_06520050509_B30.TIF

PRAGMENTASI HUTAN
114
L7G120065_06520050509_B40.TIF
L7G120065_06520050509_B50.TIF
untuk wilayah Jawa Tengah yang di dalamnya mencakup kawasan Gunung
Merapi dan Gunung Merbabu, yang akan dijadikan Area Of Interest (AOI).
Panduan yang digunakan adalah ERDAS IMAGINE Tour Guides dari ERDAS Inc.
Atlanta, Gorgia (1997).
B. Metode
1. Analisis Citra
Untuk kepentingan interpretasi citra, band combination yang
digunakan adalah 5-4-3. Penutupan lahan (land cover) diklasifikasikan
menjadi delapan kelas yaitu : (1) hutan primer; (2) hutan sekunder; (3)
kebun; (4) lahan terbuka; (5) lahar; (6) pemukiman (7) sawah dan (8)
sungai.
Areal yang akan dikaji dipilih menjadi Area Of Interest (AOI) yaitu
kawasan lanskap Gunung Merapi–Gunung Merbabu. AOI ini kemudian
diklasifikasikan tipe penutupan lahannya (land cover). Klasifikasi dilakukan
secara unsupervised dan bertahap dari 90 kelas pada tahap pertama
direklaifikasi lagi sampai akhirnya menjadi 8 kelas. Hasil klasifikasi
diekspor ke shape file untuk kemudian dilakukan opearasi spasial dengan
program ArcView GIS. Secara skematis tahapan prosedur kajian ini
disajikan pada Gambar 22.
2. Evaluasi Lanskap
Untuk mengetahui struktur lanskap dilakukan analisis patch
dengan extension Patch Analyst yang ada dalam program ArcView GIS.
Analisis dilakukan pada skala lanskap dan skala kelas.
3. Analisis Spasial
Dari literatur diketahui bahwa macan tutul dapat hidup di berbagai
ketinggian dan di berbagai tipe hutan. Oleh karena itu, yang akan dijadikan
persyaratan dalam analisis spasial ini hanyalah tipe vegetasi, dimana yang
diangap cocok untuk habitat macan tutul adalah hutan primer dan hutan
sekunder. Sementara ketinggian tidak dijadikan penentu karena macan
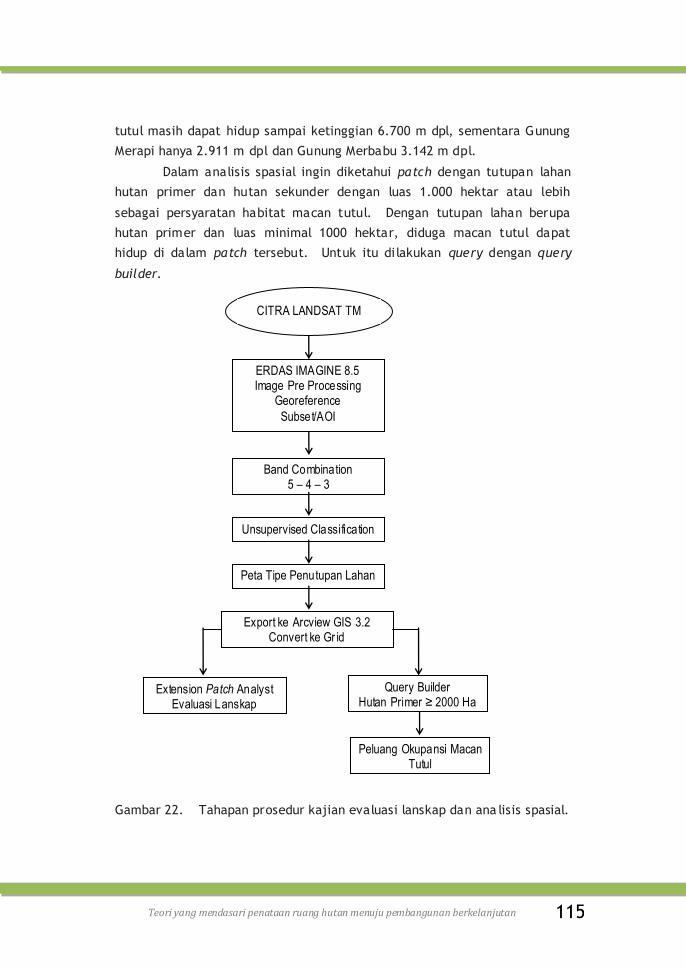
Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
115
tutul masih dapat hidup sampai ketinggian 6.700 m dpl, sementara Gunung
Merapi hanya 2.911 m dpl dan Gunung Merbabu 3.142 m dpl.
Dalam analisis spasial ingin diketahui patch dengan tutupan lahan
hutan primer dan hutan sekunder dengan luas 1.000 hektar atau lebih
sebagai persyaratan habitat macan tutul. Dengan tutupan lahan berupa
hutan primer dan luas minimal 1000 hektar, diduga macan tutul dapat
hidup di dalam patch tersebut. Untuk itu dilakukan query dengan query
builder.
Gambar 22. Tahapan prosedur kajian evaluasi lanskap dan ana lisis spasial.
CITRA LANDSAT TM
ERDAS IMAGINE 8.5 Image Pre Processing
Georeference
Subset/AOI
Unsupervised Classification
Band Combination 5 – 4 – 3
Peta Tipe Penutupan Lahan
Export ke Arcview GIS 3.2 Convert ke Gr id
Extension Patch Analyst Evaluasi Lanskap
Query Builder Hutan Primer ≥ 2000 Ha
Peluang Okupansi Macan Tutul
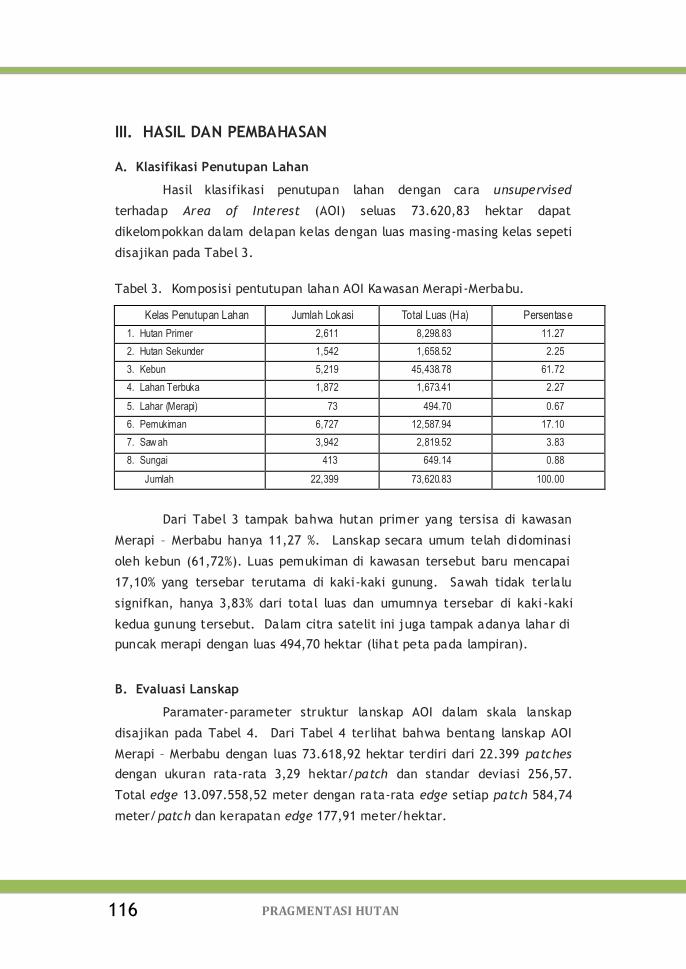
PRAGMENTASI HUTAN
116
III. HASIL DAN PEMBAHASAN
A. Klasifikasi Penutupan Lahan
Hasil klasifikasi penutupan lahan dengan cara unsupervised
terhadap Area of Interest (AOI) seluas 73.620,83 hektar dapat
dikelompokkan dalam delapan kelas dengan luas masing-masing kelas sepeti
disajikan pada Tabel 3.
Tabel 3. Komposisi pentutupan lahan AOI Kawasan Merapi-Merbabu.
Kelas Penutupan Lahan Jumlah Lokasi Total Luas (Ha) Persentase
1. Hutan Primer 2,611 8,298.83 11.27
2. Hutan Sekunder 1,542 1,658.52 2.25
3. Kebun 5,219 45,438.78 61.72
4. Lahan Terbuka 1,872 1,673.41 2.27
5. Lahar (Merapi) 73 494.70 0.67
6. Pemukiman 6,727 12,587.94 17.10
7. Saw ah 3,942 2,819.52 3.83
8. Sungai 413 649.14 0.88
Jumlah 22,399 73,620.83 100.00
Dari Tabel 3 tampak bahwa hutan primer yang tersisa di kawasan
Merapi – Merbabu hanya 11,27 %. Lanskap secara umum telah didominasi
oleh kebun (61,72%). Luas pemukiman di kawasan tersebut baru mencapai
17,10% yang tersebar terutama di kaki-kaki gunung. Sawah tidak terlalu
signifkan, hanya 3,83% dari total luas dan umumnya tersebar di kaki -kaki
kedua gunung tersebut. Dalam citra satelit ini juga tampak adanya lahar di
puncak merapi dengan luas 494,70 hektar (lihat peta pada lampiran).
B. Evaluasi Lanskap
Paramater-parameter struktur lanskap AOI dalam skala lanskap
disajikan pada Tabel 4. Dari Tabel 4 terlihat bahwa bentang lanskap AOI
Merapi – Merbabu dengan luas 73.618,92 hektar terdiri dari 22.399 patches
dengan ukuran rata-rata 3,29 hektar/patch dan standar deviasi 256,57.
Total edge 13.097.558,52 meter dengan rata-rata edge setiap patch 584,74
meter/patch dan kerapatan edge 177,91 meter/hektar.
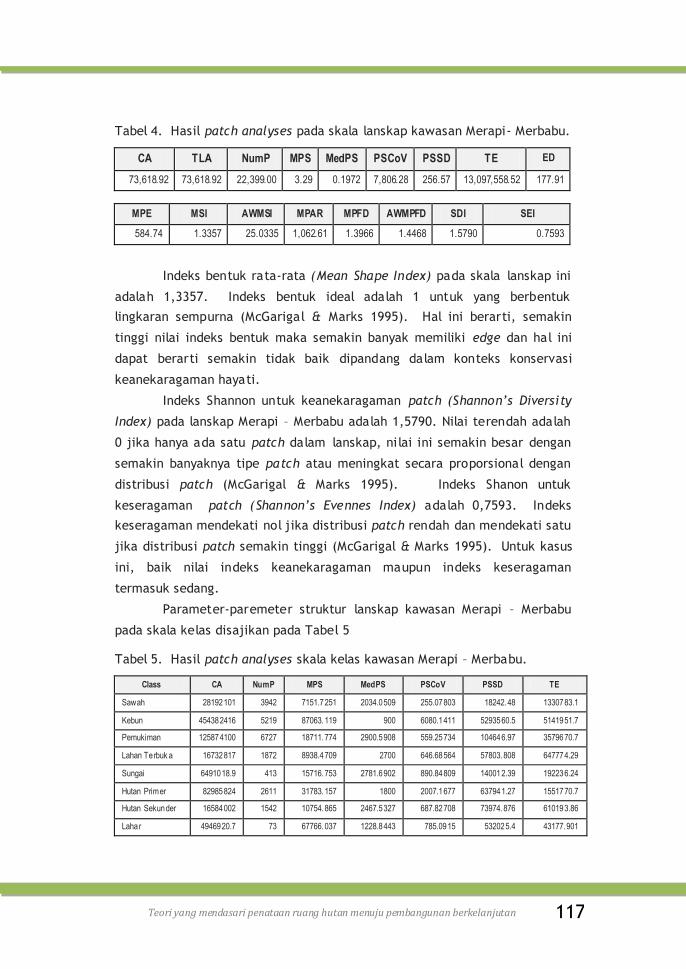
Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
117
Tabel 4. Hasil patch analyses pada skala lanskap kawasan Merapi- Merbabu.
CA TLA NumP MPS MedPS PSCoV PSSD TE ED
73,618.92 73,618.92 22,399.00 3.29 0.1972 7,806.28 256.57 13,097,558.52 177.91
MPE MSI AWMSI MPAR MPFD AWMPFD SDI SEI
584.74 1.3357 25.0335 1,062.61 1.3966 1.4468 1.5790 0.7593
Indeks bentuk rata-rata (Mean Shape Index) pada skala lanskap ini
adalah 1,3357. Indeks bentuk ideal adalah 1 untuk yang berbentuk
lingkaran sempurna (McGarigal & Marks 1995). Hal ini berarti, semakin
tinggi nilai indeks bentuk maka semakin banyak memiliki edge dan hal ini
dapat berarti semakin tidak baik dipandang dalam konteks konservasi
keanekaragaman hayati.
Indeks Shannon untuk keanekaragaman patch (Shannon’s Diversity
Index) pada lanskap Merapi – Merbabu adalah 1,5790. Nilai terendah adalah
0 jika hanya ada satu patch dalam lanskap, nilai ini semakin besar dengan
semakin banyaknya tipe patch atau meningkat secara proporsional dengan
distribusi patch (McGarigal & Marks 1995). Indeks Shanon untuk
keseragaman patch (Shannon’s Evennes Index) adalah 0,7593. Indeks
keseragaman mendekati nol jika distribusi patch rendah dan mendekati satu
jika distribusi patch semakin tinggi (McGarigal & Marks 1995). Untuk kasus
ini, baik nilai indeks keanekaragaman maupun indeks keseragaman
termasuk sedang.
Parameter-paremeter struktur lanskap kawasan Merapi – Merbabu
pada skala kelas disajikan pada Tabel 5
Tabel 5. Hasil patch analyses skala kelas kawasan Merapi – Merbabu.
Class CA NumP MPS MedPS PSCoV PSSD TE
Sawah 28192101 3942 7151.7251 2034.0509 255.07803 18242.48 1330783.1
Kebun 454382416 5219 87063.119 900 6080.1411 5293560.5 5141951.7
Pemukiman 125874100 6727 18711.774 2900.5908 559.25734 104646.97 3579670.7
Lahan Terbuk a 16732817 1872 8938.4709 2700 646.68564 57803.808 647774.29
Sungai 6491018.9 413 15716.753 2781.6902 890.84809 140012.39 192236.24
Hutan Primer 82985824 2611 31783.157 1800 2007.1677 637941.27 1551770.7
Hutan Sekunder 16584002 1542 10754.865 2467.5327 687.82708 73974.876 610193.86
Lahar 4946920.7 73 67766.037 1228.8443 785.0915 532025.4 43177.901

PRAGMENTASI HUTAN
118
Komposisi Luas Setiap Kelas Penutupan Lahan
Di Kawasan Lanskap Merapi - Merbabu
Kebun
62%
Hutan Sekunder
2%Hutan Primer
11%Sungai
1%
Lahan Terbuka
2%
Pemukiman
17%
Lahar
1%Sawah
4%
Class ED MPE MSI AWMSI MPAR MPFD AWMPFD
Sawah 0.0018077 337.59084 1.1753858 1.5932474 0.1064781 1.0358913 1.0753844
Kebun 0.0069846 985.23696 1.1743595 33.639594 0.1188159 1.0333634 1.338805
Pemukiman 0.0048624 532.13479 1.2044895 2.4834018 0.097942 1.0377723 1.109076
Lahan Terbuk a 0.0008799 346.03328 1.15755 2.0174635 0.1016752 1.0329523 1.0920544
Sungai 0.0002611 465.46305 1.1800859 3.5735501 0.0991964 1.0345259 1.1488919
Hutan Primer 0.0021078 594.32046 1.1835092 6.9835049 0.1082887 1.0345488 1.2068266
Hutan Sekunder 0.0008289 395.71586 1.1789416 2.4164773 0.1027523 1.0354675 1.1118753
Lahar 5.865E -05 591.47809 1.1814022 2.6891076 0.1225719 1.0381473 1.1282568
1. Class Area
Class area (CA)
menunjukkan luas masing-
masing kelas penutupan
lahan disajikan pada
Tabel 3, sedangkan
komposisinya dapat di-
lihat pada Gambar 23.
Gambar 23. Komposisi luas setiap kelas penutupan lahan di
kawasan lanskap Merapi Merbabu.
Dari Gambar 23, tampak bahwa kawasan lanskap Merapi – Merbabu
sudah didominasi oleh kebun (62%) dan pemukiman (17%). Sementara hutan
primer yang tersisa tinggal 11%. Dari segi konservasi, tampaknya kawasan
ekosistem Merapi – Merbabu sedang mengalami tekanan yang sangat berat.
Akibatnya kawasan budidaya berkembang pesat dan kawasan lindung
mungkin akan terus menurun. Pada kondisi seperti ini, penataan ruang
perlu menjadi perhatian, mengingat kawasan Merapi–Merbabu merupakan
kawasan yang memiliki fungsi lindung, terutama sebagai daerah tangkapan
air dan dikeli lingi oleh kota-kota penting yang setiap saat dapat terancam
banjir, tanah longsor dan kekeringan.
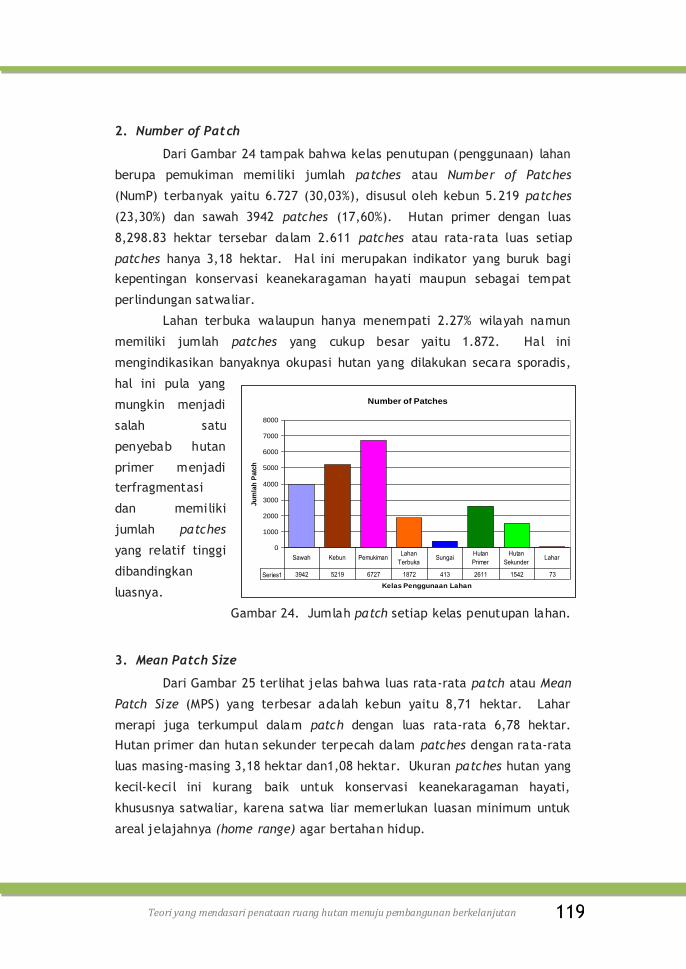
Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
119
Number of Patches
0
1000
2000
3000
4000
5000
6000
7000
8000
Kelas Penggunaan Lahan
Ju
mla
h P
atc
h
Series1 3942 5219 6727 1872 413 2611 1542 73
Sawah Kebun PemukimanLahan
TerbukaSungai
Hutan
Primer
Hutan
SekunderLahar
2. Number of Patch
Dari Gambar 24 tampak bahwa kelas penutupan (penggunaan) lahan
berupa pemukiman memiliki jumlah patches atau Number of Patches
(NumP) terbanyak yaitu 6.727 (30,03%), disusul oleh kebun 5.219 patches
(23,30%) dan sawah 3942 patches (17,60%). Hutan primer dengan luas
8,298.83 hektar tersebar dalam 2.611 patches atau rata-rata luas setiap
patches hanya 3,18 hektar. Hal ini merupakan indikator yang buruk bagi
kepentingan konservasi keanekaragaman hayati maupun sebagai tempat
perlindungan satwaliar.
Lahan terbuka walaupun hanya menempati 2.27% wilayah namun
memiliki jumlah patches yang cukup besar yaitu 1.872. Hal ini
mengindikasikan banyaknya okupasi hutan yang dilakukan secara sporadis,
hal ini pula yang
mungkin menjadi
salah satu
penyebab hutan
primer menjadi
terfragmentasi
dan memiliki
jumlah patches
yang relatif tinggi
dibandingkan
luasnya.
Gambar 24. Jumlah patch setiap kelas penutupan lahan.
3. Mean Patch Size
Dari Gambar 25 terlihat jelas bahwa luas rata-rata patch atau Mean
Patch Size (MPS) yang terbesar adalah kebun yaitu 8,71 hektar. Lahar
merapi juga terkumpul dalam patch dengan luas rata-rata 6,78 hektar.
Hutan primer dan hutan sekunder terpecah dalam patches dengan rata-rata
luas masing-masing 3,18 hektar dan1,08 hektar. Ukuran patches hutan yang
kecil-kecil ini kurang baik untuk konservasi keanekaragaman hayati,
khususnya satwaliar, karena satwa liar memerlukan luasan minimum untuk
areal jelajahnya (home range) agar bertahan hidup.

PRAGMENTASI HUTAN
120
Total Edge
0
1,000
2,000
3,000
4,000
5,000
6,000
Kelas
Km
Series1 1,331 5,142 3,580 648 192 1,552 610 43
Sawah Kebun PemukimanLahan
TerbukaSungai
Hutan
Primer
Hutan
SekunderLahar
Gambar 25. Luas patch rata-rata setiap kelas penutupan lahan.
4. Total Edge
Total Edge (TE) atau keliling dari patch yang terpanjang adalah
pada kelas penutupan lahan kebun yaitu 5.142 km, diikuti oleh pemukiman
3.142 km dan hutan primer 1.552 km. Semakin panjang edge dapat
menjadi indikasi bentuk patch yang semakin tidak beraturan (kompleks) dan
jumlah patch yang semakin banyak (Gambar 26).
Gambar 26. Total edge setiap kelas penutupan lahan.
Mean Patch Size
0.00
2.00
4.00
6.00
8.00
10.00
Kelas
Hekta
r
Series1 0.72 8.71 1.87 0.89 1.57 3.18 1.08 6.78
Sawah Kebun PemukimanLahan
TerbukaSungai
Hutan
Primer
Hutan
SekunderLahar

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
121
Edge Density
0.00
20.00
40.00
60.00
80.00
Kelas
Per H
a
Series1 18.08 69.85 48.62 8.80 2.61 21.08 8.29 0.59
Sawah Kebun PemukimanLahan
TerbukaSungai
Hutan
Primer
Hutan
SekunderLahar
Mean Patch Edge
0.00
200.00
400.00
600.00
800.00
1000.00
1200.00
Kelas
Mete
r/P
atc
h
Series1 337.59 985.24 532.13 346.03 465.46 594.32 395.72 591.48
Sawah Kebun PemukimanLahan
TerbukaSungai
Hutan
Primer
Hutan
SekunderLahar
5. Edge Density
Edge density (ED) adalah jumlah edge relatif terhadap luas lanskap.
Edge density terbesar pada AOI lanskap Merapi – Merbabu adalah kebun
yaitu 69,85 meter/hektar, disusul oleh pemukiman 48,62 meter/hektar dan
hutan primer 21,08 meter/hektar. Semakin tinggi edge density menjadi
indikasi semakin
tidak baik untuk
habitat satwa yang
sensitif terhadap
edge (Gambar 27).
Gambar 27.
Edge density setiap kelas penutupan
lahan.
6. Mean Patch Edge
Mean patch edge (MPE) adalah jumlah rata edge per patch. Pada
AOI lanskap Merapi – Merbabu, nilai MPE tertinggi adalah kebun dengan
985,24 meter/patch. Hutan primer juga memiliki MPE yang relatif tinggi
dibandingkan kelas penutupan lahan lain, yaitu 594,32 mater/patch
dibandingkan dengan sawah, pemukiman, lahan terbuka dan hutan
sekunder (Gambar 28).
Gambar 28. Mean
patch edge setiap kelas penutupan
lahan.

PRAGMENTASI HUTAN
122
7. Mean Shape Index
Mean shape index (MSI) menggambaran kompleksitas bentuk patch.
Tingkat kompleksitas bersifat relatif dibandingkan terhadap bentuk
lingkaran atau bujur sangkar. Nilai MSI lebih dari 1, nilai MSI sama dengan 1
ketika semua patches berbentuk lingkaran atau bujur sangkar (MacGarigal
and Marks, 1995).
Nilai MSI diperoleh dengan membagi jumlah keliling setiap patches
dengan akar kuadrat dari luas patch (hektar) dan disesuaikan untuk standar
lingkaran (poligon) atau bujur sangkar (grid), dibagi dengan jumlah patches
(MacGarigal and Marks, 1995). Hal ini berarti semakin tinggi ni lai MSI suatu
kelas penutupan lahan maka semakin kompleks bentuk-bentuk patches-nya
dan semakin besar juga edge-nya. Hal ini untuk satwa-satwa yang tidak
menyukai edge dapat berakibat buruk karena mengurangi luas habitatnya.
Gambar 29. Mean shape index setiap kelas penutupan lahan.
8. Mean Patch Fractal Dimension
Mean Patch Fractal Dimension (MPFD) juga merupakan ukuran
kompleksitas bentuk patch. Nilai MPFD mendekati satu untuk bentuk
dengan keli ling (perimeter) sederhana dan mendekati dua jika bentuknya
lebih kompleks (MacGarigal and Marks, 1995).
Mean Shape Index
1.1200
1.1400
1.1600
1.1800
1.2000
1.2200
Kelas
Ind
ex
Series1 1.1754 1.1744 1.2045 1.1576 1.1801 1.1835 1.1789 1.1814
Sawah Kebun PemukimanLahan
TerbukaSungai
Hutan
Primer
Hutan
SekunderLahar

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
123
Gambar 30. Mean patch fractal dimension untuk setiap kelas penutupan lahan.
Untuk studi kasus ini, kompleksitas bentuk patches tertinggi dimiliki
oleh patch lahar Gunung Merapi dengan nilai MPFD 1,0381 disusul oleh
pemukiman dengan nilai 1,0378, sawah 1,0359 dan hutan sekunder 1,0355.
Secara umum nilai MPFD pada lanskap Merapi – Merbabu ini lebih mendekati
nilai satu, yang berarti lebih dekat ke bentuk sederhana.
C. Potensi Sebaran Habitat Macan Tutul
Dari hasil query diperoleh hanya ada dua lokasi yang memenuhi
syarat sebagai habitat macan tutul (Gambar 11), masing-masing dengan luas
1.784,039 hektar dan 2.690,736 hektar yang berada di sekitar puncak
Gunung Merapi dan Gunung Merbabu. Keduanya merupakan hutan primer,
sementara hutan sekundernya tidak ada yang memenuhi syarat luas
minimal. Namun karena hutan sekunder yang berada di Gunung Merapi
(warna hijau muda) berada bersambungan dengan hutan primer maka,
secara bersama-sama dengan hutan primer dapat berfungsi sebagai habitat
macan tutul.
Berdasarkan Tabel 3 dan Tabel 5 tampak bahwa ada 2.611 patches
hutan primer dan 1.542 patches hutan sekunder, masing-masing dengan luas
patches rata-rata 3,18 ha untuk hutan primer dan 1,08 untuk hutan
sekunder. Total hutan primer sendiri hanya 8.298,83 hektar, sementara
hutan sekunder 1.658,5 hektar, sehingga areal berhutan di lanskap Merapi
Mean Patch Fractal Dimension
1.0300
1.0320
1.0340
1.0360
1.0380
1.0400
Kelas
Series1 1.0359 1.0334 1.0378 1.0330 1.0345 1.0345 1.0355 1.0381
Sawah Kebun PemukimanLahan
TerbukaSungai Hutan Primer
Hutan
SekunderLahar

PRAGMENTASI HUTAN
124
Merbabu pada tahun 2005 (saat pengambi lan citra) hanyalah 9.957,35
hektar.
Ukuran home range macan tutul sangat bervariasi dan sangat
tergantung pada ketersediaan jumlah dan penyebaran satwa mangsa (IUCN -
The World Conservation Union 1996). Ukuran home range macan tutul
rata-rata berkisar antara 30 – 78km2 (jantan) dan 23 – 33 km2 (betina) di
kawasan yang dilindungi (Bailey 1993). Tetapi home range mungkin jauh
lebih besar ketika ketersediaan makanan berkurang dan kepadatan macan
tutul rendah.
Di Taman Nasional Royal Chitwan, Nepal, jumlah ungulata per
kilometer persegi sangat tinggi, macan tutul betina menjelajahai wilayah
antara 6 – 13 km2. Di Taman Nasional Serengeti dan Tsavo, Afrika Timur,
teritori mereka berkisar antara 11 – 121 km2. Tetapi di Pegunungan
Stellenbosch, Afrika Selatan, di Kalahari dan di Pegunungan Sikhote Alin,
Rusia Timur, macan tutul jantan berburu di teritori yang kadang-kadang
lebih dari 400 km2 (IUCN - The World Conservation Union, 1996).
Berdasarkan informasi home range dan bukti bahwa di Cagar Alam
Pulau Sempu (Kabupaten Malang) dengan luas 877 ha ditemukan macan
kumbang (Surabaya Post Hot News, Selasa, 17/09/1996), maka pada studi
ini, diasumsikan seekor macan tutul memiliki daerah jelajah antara 600 –
1.000 hektar. Dengan demikian kawasan ekosistem Gunung Merapi –
Gunung Mebabu tersebut hanya mampu mendukung 10 - 17 ekor macan
tutul.
Jika dilihat dari Gambar 31, tampak bahwa kecil kemungkinan
masih adanya pertukaran genetik antara populasi macan tutul di Gunung
Merapi dan populasi macan tutul di Gunung Merbabu. Hal ini disebabkan
tidak adanya konektivitas berupa koridor yang menghubungkan kedua
populasi tersebut. Bila diperhatikan dengan seksama, hutan primer yang
tampaknya sebelumnya pernah menyatu antara Gunung Merapi dan Gunung
Merbabu, kini sudah terpecah-pecah dan terpotong oleh pemukiman dan
kebun. Pemukiman dan kebun ini merupakan barrier bagi pergerakan
macan tutul antar kedua populasi.

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
125
Gambar 31. Lokasi dugaan habitat macan tutul terpilih (warna kuning) hasil query.
Meskipun demikian, masih ada harapan untuk membuat koridor
antara Gunung Merapi dan Merbabu. Kemungkinan koridor dapat dibuat
dengan merekonstruksi tutupan hutan melalui tepi sungai yang saling
bertemu dari Gunung Merapi dan dari Gunung Merbabu atau melalui jarak
terdekat melalui kawasan Selo (sela antara G. Merapi dan G. Merbabu)
(Gambar 31).
Tanpa adanya koridor, bila macan tutul dari Gunung Merapi akan
menjelajah ke Gunung Merbabu, maka harus melewati kebun dan
pemukiman. Hal ini dapat membahayakan macan tutul itu sendiri dan
masyarakat. Dengan kondisi habitat macan tutul (hutan) yang dikepung
oleh pemukiman dan kebun, dikhawatirkan akan sering terjadi konflik
G. Merapi
G. Merbabu

PRAGMENTASI HUTAN
126
antara macan tutul dan masyarakat. Hal ini bisa terjadi jika di habitat
alaminya (hutan) kekurangan persediaan satwa mangsa, maka macan tutul
dikhawatirkan akan masuk ke kampung dan memangsa hewan ternak dan
bila bertemu dengan manusia bisa melukai manusia atau manusia yang
membantai macan tutul.
Kondisi kawasan lanskap Merapi-Merbabu dengan struktur,
komposisi dan konfigurasi lanskap seperti saat ini, tampaknya kurang
menguntungkan bagi kepentingan konservasi keanekaragaman hayati,
khususnya satwa langka dengan home range luas seperti macan tutul.
Fragmentasi habitat dan habitat loss telah membatasi pergerakan macan
tutul dan menurunkan kualitas habitatnya sehingga dapat mengancam
kelestariannya.
Upaya-upaya yang perlu dilakukan untuk mengindari kehancuran
ekosistem Merapi – Merbabu dan kepunahan berbagai jenis spesies langka
antara lain dapat di lakukan dengan :
1. Segera menetapkan kawasan-kawasan hutan negara yang memiliki
fungsi lindung hidrologi dan konservasi keanekaragaman hayati sebagai
kawasan konservasi.
2. Melakukan restorasi ekosistem hutan, terutama di sepanjang kawasan
penghubung antara Merapi dan Merbabu.
3. Menghentikan okupasi hutan yang dilakukan secara ilegal dan
merehabilitasi kawasan hutan yang rusak serta melakukan revegetasi
lahan-lahan terbuka di kawasan hutan negara.
4. Meninjau kembali rencana tata ruang wilayah regional, khususnya yang
menyangkut wilayah penyangga kehidupan di bentang lanskap Merapi –
Merbabu.
5. Melakukan perluasan habitat dengan membangun daerah-daerah
penyangga di sekitar kawasan konservasi Taman Nasional Merapi dan
Taman Nasional Merbabu.

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
127
IV. SIMPULAN DAN SARAN
1. Kawasan Merapi – Merbabu sedang mengalami tekanan perubahan
struktur, komposisi dan konfigurasi lanskap.
2. Parameter-parameter lanskap hasil patch analyses menunjukkan kondisi
yang kurang menguntungkan bagi kepentingan konservasi
keanekaragaman hayati dan ekosistemnya.
3. Kawasan berhutan yang tersisa tinggal 13,52% (9.957,35 hektar) dari
sekitar total lanskap seluas 73.620,83 hektar. Kebun dan pemukiman
sudah mendominasi lanskap ini, masing-masing dengan penutupan lahan
61,72% dan 17,10%.
4. Tingkat fragmentasi dan isolasi hutan juga tinggi yang ditunjukkan oleh
banyaknya patches hutan yaitu 2.611 untuk hutan primer dari luasnya
8.298,83 hektar dan 1.542 patches untuk hutan sekunder dari luasnya
1.658,52 hektar.
5. Peluang keberadaan macan tutul (Panthera pardus melas) di lanskap
Merapi – Merbabu hanya berada di dua lokasi yaitu di sekitar puncak
kedua gunung tersebut dengan luas total 8.298,83 hektar hutan primer.
Bila ditambah dengan hutan sekunder 1.658,5 hektar maka luasnya
menjadi 9.957,35 hektar.
6. Dengan asumsi home range seekor macan tutul berkisar 600 – 1.000
hektar maka kawasan tersebut diperkirakan hanya mampu mendukung
populasi 10 – 17 ekor.
7. Untuk mencegah kehancuran ekosistem Merapi – Merbabu lebih parah,
perlu segera dilakukan peninjauan dan pembenahan rencana tata ruang
regional dan penetapan kawasan-kawasan hutan negara menjadi
kawasan konservasi sebagai perluasan TN Merapi dan TN Merbabu.
8. Ekspansi wilayah pemukiman dan budidaya dengan mengokupasi hutan
perlu segera dihentikan dan kawasan hutan yang sudah terlanjur rusak
perlu segera direstorasi.

PRAGMENTASI HUTAN
128
DAFTAR PUSTAKA
http://www.bio.davidson.edu/people/vecase/Behavior/Spring2002/Friedman/socialsystem.html. Diakses Tanggal 4 Mei 2007.
IUCN - The World Conservation Union. 1996. Leopard Panthera pardus Linnaeus 1758. IUCN - The World Conservation Union.
McGarigal, K., and B. J. Marks. 1995. Fragstats: spatial pattern analysis program for quantifying landscape structure. U.S. Forest Service
General Technical Report PNW-GTR-351.
Surabaya Post Hot News, Selasa, 17/09/1996. Perburuan Liar Ancam
Kelestarian Pulau Sempu dan Satwa Langka. http://www.wp.com/64257/170996/05sempu.htm. Diakses 01-02-
2007.

Lampiran : Penutupan Lahan Kawasan Gunung Merapi-Gunung Merbabu.
Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan PRAGMENTASI HUTAN 129

PRAGMENTASI HUTAN
130
BIODATA PENULIS
Dr. Ir. Hendra Gunawan, M.Si. dilahirkan pada 3
April 1964 di Kabupaten Banjarnegara. Putera
keempat dari pasangan Alimah (Ibu) dan
Aswowikarto (ayah) menyelesaikan pendidikan SD
(1976) dan SMP (1980) di Kabupaten Banjarnegara
dan SMA di SMAN 1 Kota Cirebon (1980). Meraih
gelar sarjana kehutanan (1980), Magister Sains
Pengelolaan Sumberdaya Alam dan Lingkungan
(2000) dan Doktor Ilmu Kehutanan (2010) di Institut
Pertanian Bogor.
Dalam karirnya penulis pernah menjadi manajer pembinaan hutan di sebuah
HPH (1989-1991), kemudian menjadi Pegawai Negeri Sipil di Kementerian
Kehutanan sejak 1992 dengan profesi sebagai peneliti di bidang konservasi
sumberdaya alam. Saat ini jabatannya adalah Peneliti Utama di Bidang
Konservasi Sumberdaya Alam di Puslitbang Konservasi dan Rehabilitasi,
Badan Litbang Kementerian Kehutanan.
Penulis telah menghasilkan lebih dari 110 karya tulis ilmiah dan populer.
Selain meneliti, penulis juga sering diminta sebagai konsultan, narasumber,
pembimbing mahasiswa, pengajar diklat dan tenaga ahli di berbagai tim
dan kelompok kerja. Menjadi anggota Dewan Riset Badan Litbang
Kehutanan, Dewan Redaksi di beberapa jurnal dan majalah, anggota Pokja
Konservasi Badak Indonesia dan Pokja Restorasi Ekosistem Kawasan
Konservasi merupakan kesibukannya akhir-akhir ini.
Penulis menaruh perhatian pada dampak lingkungan, baik akibat
pembangunan maupun bencana alam. Di sela kesibukannya sebagai peneliti,
masih menyempatkan diri menjadi penyusun AMDAL dan telah mendapatkan
sertifikasi kompetensi Ketua Tim Penyusun AMDAL (KTPA) dengan bidang
keahlian dampak ekologis.
Kegiatan yang sedang disibukinya saat ini antara lain melanjutkan
penelitian Restorasi Ekosistem Gunung Merapi, penelitian Sebaran Macan
Tutul Di Jawa Barat, penyusunan buku Bioekologi dan Konservasi Badak
Indonesia, buku Restorasi Ekosistem Pegunungan Pasca Perambahan Di
Gunung Ciremai dan buku Penanganan Satwaliar Pasca Erupsi Gunung
Merapi.

Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
131
BIODATA PENULIS
Prof. Dr. Ir. Lilik Budi Prasetyo, MSc, lahir di
Salatiga, 16 Maret 1962. Menyelesaikan pendidikan
dasarnya hingga Seklah menengah Atas di Salatiga.
Gelar sarjana S-1 diperolehnya tahun 1986 dari
Departemen Agronomi, Fakultas Pertanian, Institut
Pertanian Bogor. Penulis mendapatkan kesempatan
melanjutkan pendidikan pascasarjana di bidang
Perencanaan Lanskap, Departemen Lingkungan
University of Tsukuba Jepang dan tamat tahun
1993.
Pendidikan Doktor diambil di universitas yang sama di bidang Manajemen
Hutan dan tamat pada tahun 1996 dengan disertasi berjudul : Application of
Geographical Information System (GIS) for Land-Use Changes Study Toward
Better Understanding of Deforestration Process: case Studies in JABOTABEK
and South Sumatra, Indonesia. Lanskap, Pemodelan Spasial
Penulis saat ini adalah guru besar di bidang ekologi lanskap dan dipercaya
sebagai Kepala Laboratorium Analisis Lingkungan dan Permodelan Spasial,
Departemen Konservasi Sumberdaya Hutan-Fakultas Kehutanan – IPB.
Penulis mengasuh mata kuliah Analisis Spasial Lingkungan, Sistem Informasi
Geografi dan Ekologi Lanskap. Selain itu, penulis juga menjadi Kepala
Program Perubahan Lingkungan Global, dan Kepala Lab. Analisis Lingkungan
Spasial, Pusat Penelitian Lingkungan Hidup-IPB.
Penulis pernah menjadi peneliti tamu di Institut of Agroenvironmental
Sciences, Tsukuba dan Universita Tokyo. Penelitian yang dilakukan saat ini
banyak berkaitan dengan permasalahan deforestasi, perubahan penggunaan
lahan, fragmentasi habitat dan peranan elemen lanskap. Email:
[email protected] dan hompage: http://lbprastdp.staff.ipb.ac.id

FRAGMENTASI HUTAN Teori yang mendasari penataan ruang hutan
menuju pembangunan berkelanjutan
Pusat Penelitian dan Pengembangan Konservasi dan Rehabilitasi
Badan Penelitian dan Pengembangan Kehutanan
KEMENTERIAN KEHUTANAN




















