
Field Performance and Genetic Fidelity of Micropropagated ...
11
Open Life Sci. 2017; 12: 1–11 Skoog; CTAB − Cetyl trimethylammonium bromide; IAA − Indole Acetic Acid; SDS − Sodium Dodecyl Sulphate 1 Introduction Coffee is one of the most important plantation crops and agricultural export commodities in the world. It is grown in about 80 countries of the tropical and subtropical regions of the world, especially in Africa, Asia and Latin America. The total global production of green coffee is about 143.25 million bags (60 kg capacity) with an export earning of over US$31.8 billion during 2014. Although more than 100 coffee species are known, only two species, i.e. Coffea arabica (known as arabica coffee) and Coffea canephora (known as robusta coffee), are commercially cultivated and consumed. Arabica coffee is favored over robusta coffee because of its quality and aroma and constitutes about 60% of total global coffee production. However, in recent years, there has been a shift towards robusta coffee cultivation because of the rising production costs, labor scarcity and other difficulties in providing timely inputs especially for disease and pest management associated with arabica cultivation. This challenging scenario has impelled focused research on robusta coffee in India. Traditionally coffee is propagated through seeds and new plantations are established by planting the seedlings. However, in C. canephora, propagation through seeds is not desirable and leads to the appearance of off type plants in the progeny due to cross pollination and segregation. Thus, in vitro propagation is the method of choice to palliate this problem and propagate elite mother plants in large numbers. There are several reports on micropropagation and in vitro multiplication of plants via somatic embryogenesis in coffee. However, successful reports on field establishment and performance of micropropagated plants of coffee are very much limited. The progress on micropropagation in coffee has been reviewed [1-3]. In Brazil, an experimental field was established to DOI 10.1515/biol-2017-0001 Received July 13, 2016; accepted October 31, 2016 Abstract: This study was conducted to compare the growth and yield of one of the commercial hybrid coffee cultivars (Coffea congensis x Coffea canephora) of robusta coffee established from somatic embryogenesis as well as conventional seedlings. Results indicated no statistically significant differences in the growth pattern or the cumulative yield between the somatic embryogenesis derived plants and the seedlings. The genetic fidelity of somatic embryogenesis derived plants and the mother plant was tested using sequence related amplified polymorphism (SRAP) markers. A total of 24 SRAP primers were employed for DNA analysis which produced a total of 153 clear, distinct and reproducible amplicons of variable size. Out of 24 SRAP primers, 9 primers produced amplification patterns which are identical between the mother plants and plants derived from somatic embryogenesis. Cluster analysis revealed more than 95% genetic similarity between the somatic embryogenesis derived plants and the mother plants indicating a high degree of genetic fidelity. The present study clearly demonstrates the usefulness of SRAP markers in genetic fidelity analysis of coffee. Keywords: Somatic embryogenesis, Micropropagated plants, Field performance, Genetic fidelity, Coffea canephora, SRAP markers Abbreviations: CV − coefficient of variation; 2,4-D − 2,4-dichlorophenoxyaceticacid; MS −Murashige and © 2017 Bychappa Muniswamy et al., published by De Gruyter Open. This work is licensed under the Creative Commons Attribution-NonCommercial-NoDerivs 3.0 License. *Corresponding author: Manoj Kumar Mishra, Plant Biotechnology Division, Unit of Central Coffee Research Institute, Coffee Board, Manasagangothri, Mysore-570 006, India, E-mail: [email protected] Bychappa Muniswamy, Bharathi Kosaraju, Raghuramulu Yenugula, Plant Biotechnology Division, Unit of Central Coffee Research Institute, Coffee Board, Manasagangothri, Mysore-570 006, India Bychappa Muniswamy, Bharathi Kosaraju, Manoj Kumar Mishra*, Raghuramulu Yenugula Field Performance and Genetic Fidelity of Micropropagated Plants of Coffea canephora (Pierre ex A. Froehner) Research Article Open Access
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Field Performance and Genetic Fidelity of Micropropagated ...

Open Life Sci. 2017; 12: 1–11
Skoog; CTAB − Cetyl trimethylammonium bromide; IAA − Indole Acetic Acid; SDS − Sodium Dodecyl Sulphate
1 IntroductionCoffee is one of the most important plantation crops and agricultural export commodities in the world. It is grown in about 80 countries of the tropical and subtropical regions of the world, especially in Africa, Asia and Latin America. The total global production of green coffee is about 143.25 million bags (60 kg capacity) with an export earning of over US$31.8 billion during 2014. Although more than 100 coffee species are known, only two species, i.e. Coffea arabica (known as arabica coffee) and Coffea canephora (known as robusta coffee), are commercially cultivated and consumed. Arabica coffee is favored over robusta coffee because of its quality and aroma and constitutes about 60% of total global coffee production. However, in recent years, there has been a shift towards robusta coffee cultivation because of the rising production costs, labor scarcity and other difficulties in providing timely inputs especially for disease and pest management associated with arabica cultivation. This challenging scenario has impelled focused research on robusta coffee in India.
Traditionally coffee is propagated through seeds and new plantations are established by planting the seedlings. However, in C. canephora, propagation through seeds is not desirable and leads to the appearance of off type plants in the progeny due to cross pollination and segregation. Thus, in vitro propagation is the method of choice to palliate this problem and propagate elite mother plants in large numbers. There are several reports on micropropagation and in vitro multiplication of plants via somatic embryogenesis in coffee. However, successful reports on field establishment and performance of micropropagated plants of coffee are very much limited. The progress on micropropagation in coffee has been reviewed [1-3]. In Brazil, an experimental field was established to
DOI 10.1515/biol-2017-0001Received July 13, 2016; accepted October 31, 2016
Abstract: This study was conducted to compare the growth and yield of one of the commercial hybrid coffee cultivars (Coffea congensis x Coffea canephora) of robusta coffee established from somatic embryogenesis as well as conventional seedlings. Results indicated no statistically significant differences in the growth pattern or the cumulative yield between the somatic embryogenesis derived plants and the seedlings. The genetic fidelity of somatic embryogenesis derived plants and the mother plant was tested using sequence related amplified polymorphism (SRAP) markers. A total of 24 SRAP primers were employed for DNA analysis which produced a total of 153 clear, distinct and reproducible amplicons of variable size. Out of 24 SRAP primers, 9 primers produced amplification patterns which are identical between the mother plants and plants derived from somatic embryogenesis. Cluster analysis revealed more than 95% genetic similarity between the somatic embryogenesis derived plants and the mother plants indicating a high degree of genetic fidelity. The present study clearly demonstrates the usefulness of SRAP markers in genetic fidelity analysis of coffee.
Keywords: Somatic embryogenesis, Micropropagated plants, Field performance, Genetic fidelity, Coffea canephora, SRAP markers
Abbreviations: CV − coefficient of variation; 2,4-D − 2,4-dichlorophenoxyaceticacid; MS −Murashige and
© 2017 Bychappa Muniswamy et al., published by De Gruyter Open. This work is licensed under the Creative Commons Attribution-NonCommercial-NoDerivs 3.0 License.
*Corresponding author: Manoj Kumar Mishra, Plant Biotechnology Division, Unit of Central Coffee Research Institute, Coffee Board, Manasagangothri, Mysore-570 006, India, E-mail: [email protected] Muniswamy, Bharathi Kosaraju, Raghuramulu Yenugula, Plant Biotechnology Division, Unit of Central Coffee Research Institute, Coffee Board, Manasagangothri, Mysore-570 006, India
Bychappa Muniswamy, Bharathi Kosaraju, Manoj Kumar Mishra*, Raghuramulu Yenugula
Field Performance and Genetic Fidelity of Micropropagated Plants of Coffea canephora (Pierre ex A. Froehner)
Research Article Open Access

2 B. Muniswamy, et al.
evaluate the tissue culture plants of Bourbon obtained from somatic embryos produced in both solid and liquid media and compare their performance with the seedlings of the same line [4]. In another study, five robusta clones are being field tested in five coffee producing countries (4000 plants/location): Philippines, Thailand, Mexico, Nigeria and Brazil. The visual inspection of 8000 plants under field conditions in Philippines revealed that all the micropropagated robusta plants have normal vegetative and reproductive growth with normal flowering and fruit set after 2 years of field planting [5]. Similarly, Etienne et al. [6] and Etienne & Bertrand [7] evaluated 20,000 field grown arabica tissue culture plants in four Central American countries and reported the trueness-to- type and agronomic characteristics of tissue cultured plants produced by embryogenic cell suspensions. Muniswamy et al. [8] reported the field performance of tissue cultured plants of Coffea arabica. However, micropropagation through somatic embryogenesis involving a prolonged callus phase is considered to be unreliable due to the genomic changes of in vitro raised plantlets [9,10]. Thus the genetic fidelity of the invitro derived plantlets should be tested as early as possible especially in woody perennials which have a long life cycle.
Several strategies have been employed to assess the genetic fidelity of regenerated plants of which DNA based molecular markers have gained paramount importance during recent years. Various molecular markers such as RAPD [11-14], AFLP [15,16], SSR [17] and ISSR [18] are used for determining the genetic fidelity of micropropagated plants. In coffee (C. arabica), Landey et al. [19] analyzed genetic fidelity of in vitro propagated plants of C. arabica using AFLP and SSAP markers and confirmed that genetic variation observed in micropropagated plants are caused by abnormal chromosome numbers. Interestingly, these authors could not detect genetic variability among micropropagated plants using AFLP markers. Recently, a new class of molecular marker known as sequence related amplified polymorphism (SRAP) has been increasingly used for genetic diversity and gene mapping [20]. Recently we have reported the use of SRAP markers in coffee [21-23]. The SRAP is a PCR based marker system that preferentially amplifies coding sequences randomly distributed throughout the genome. Moreover SRAP markers possessed multi loci and multi allelic features which made them more suitable than other marker systems in genetic analysis compared to other marker systems, the multi allelic features of SRAP markers make them more suitable for genetic analysis [24]. More recently, SRAP markers were used in
assessing the genetic fidelity of some plants obtained through tissue culture [25-27]. Here we describe the field performance and genetic stability of in vitro regenerated plants of Coffea canephora (CxR) hybrids using SRAP marker system. To our knowledge, SRAP has been used for the first time in assessing the genetic fidelity of coffee obtained through somatic embryogenesis.
2 Methods
2.1 Selection of elite plants and somatic embryogenesis
Superior mother plants of C. canephora (cv. CxR) were identified in some of the estates in Karnataka and used as the source material for in vitro multiplication. CxR is an interspecific hybrid between C. congensis and C. canephora which was developed at Central Coffee Research Institute (CCRI) and widely cultivated in India. Leaf explants were incubated in Murashige and Skoog (1962) medium [28] supplemented with 2, 4-D (1 mg L-1) and kinetin (5 mg L-1) for callus induction. After four weeks of incubation in callus induction medium, the developed calli were transferred to MS medium supplemented with 2, 4-D (0.1 mg L-1), IAA (1 mg L-1) and kinetin (4 mg L-1) along with proline (20 mg L-1) for somatic embryo induction. The somatic embryos were transferred to MS medium with kinetin (0.1 mg L-1) for germination and development. The well grown in vitro plantlets were planted in net pots/polybags containing soilrite mixture and hardened in a partially shaded area under polytunnels. After hardening, these somatic embryo derived plants and the seedlings of the corresponding mother plants were field planted and established in different locations. In each location, an average of 600 tissue culture derived plants and seedlings raised from the identified mother plant were planted in the field for agronomic evaluation. One hundred randomly selected tissue culture plants and equal number of seedlings were selected and growth and agronomic features like stem diameter, bush spread, number of primaries, length of primaries, number of nodes per primary, leaf area and total fruit yield were recorded for five consecutive years and data was subjected to statistical analysis. The projected mean yield of somatic embryo derived plants and seedlings were compared statistically The seed samples collected from the somatic embryo derived plants and seedlings were graded and analyzed for both bean parameters as well as organoleptic evaluation at the Coffee Board’s Quality Evaluation Centre.

Genetic fidelity of coffee tissue culture plants 3
2.2 DNA isolation
Genomic DNA was isolated from fresh young leaves using a modified CTAB method as described earlier by Mishra et al. [29]. About 200 mg of freshly collected leaf tissue was ground to fine powder in liquid nitrogen, transferred to a 30 ml tube containing 5 ml preheated extraction buffer (2% CTAB (w/v), 100 mM Tris-HCL (pH 8.0), 25 mM EDTA, 2M NaCl, and 0.1% beta-mercaptoethanol). The tubes were incubated at 60°C for one hour with occasional shaking for thorough mixing. After incubation, the tubes were cooled to room temperature and centrifuged at 6000 rpm for 20 min following which the supernatant was transferred into a fresh tube and extracted twice with chloroform-isoamyl alcohol (24:1). The supernatant was transferred to 2 ml tubes, precipitated with 0.7 volume of ice cold isopropanol at room temperature for 30 min, and then centrifuged at 8000 rpm for 20 min at 4°C. The resultant pellet after centrifugation was washed with 75% (v/v) ethanol for 10 min and dissolved in 60 µl of Tris-EDTA (1-10mM). The concentration of DNA was measured using a 0.8% agarose gel stained with ethidium bromide as well as via a UV spectrophotometer at 260 nm. The ratio of the absorbance at 260 and 280 nm (A 260/280) was used to assess the purity of DNA. The re-suspended DNA was then diluted to 10 mg/l concentration in sterile distilled water and kept at -20°C for further use in amplification reactions.
A total of 40 SRAP primer pairs involving ten forward and thirteen reverse primers were initially screened of which 24 primer pairs (Table 1) have given consistent banding pattern and therefore were used in the genetic fidelity analysis. SRAP analysis was performed by
adapting the procedure described by Li & Quiros [20] with minor modifications as described earlier by Mishra et al. [22]. Each 20 µl PCR reaction mixture consisted of 30 ng template DNA, 2 µL of 10x reaction buffer (75 mM Tris-HCl, pH 8.8, 20 mM (NH4)2 SO4, 0.01% Tween 20), 200 µM dNTP mixture, 2.5 mM MgCl2, 3 µM each of forward and reverse primer, and 1.0 U Taq DNA polymerase. The PCR amplification program followed was 4 min initial denaturation at 96°C; 5 cycles consisting of 1 min denaturation at 94°C, 1.15 min primer annealing at 35°C; and 2 min extension at 72°C followed by 30 cycles consisting of 1 min denaturation at 94°C, 1.15 min primer annealing at 50°C; and 2 min extension at 72°C; and a final extension of 15 min at 72°C. PCR products of SRAP were run on 2.0% (w/w) agarose gels containing 0.5 µg ethidium bromide/ml in 1X TAE buffer and then visualized and photographed using the UV-transilluminator (Biorad).
2.3 Data analysis
SRAP amplified bands were scored for presence (1) or absence (0). The total number of bands, distribution of bands across all the collections, polymorphic bands, and average number bands per primer were calculated. A pairwise similarity matrix was constructed using the Dice similarity coefficient [30]. The relationship between various germplasm collections was displayed as a dendrogram constructed using NTSYS –PC 2.1 software [31] based on the unweighted pair group method using arithmetic averages (UPGMA). Statistical support of the clusters was assessed by means of 1000-bootstrap replicates.
Table 1. Primer sequences used for SRAP analysis.
Name Forward primer (5’ – 3’) Name Reverse primer (5’ – 3’) Primer combinations used
Forward Reverse
Me1 TGAGTCCAAACCGGATA Em1 GACTGCGTACGAATTAAT Me1 Em4/Em7/Em12Me2 TGAGTCCAAACCGGAGC Em2 GACTGCGTACGAATTTGC Me2 Em3/Em4/Em6/Em7/Em12Me3 TGAGTCCAAACCGGAAT Em3 GACTGCGTACGAATTGAC Me3 Em3/Em4/Em11/Em12Me4 TGAGTCCAAACCGGACC Em4 GACTGCGTACGAATTTGA Me4 Em1/Em2/Em11/Em16Me5 TGAGTCCAAACCGGAAG Em5 GACTGCGTACGAATTAAC Me5 Em5Me6 TGAGTCCAAACCGGACA Em6 GACTGCGTACGAATTGCA Me6 Em5/Em15Me10 TGAGTCCAAACCGGAAA Em7 GACTGCGTACGAATTCAA Me10 Em13/Em16Me11 TGAGTCCAAACCGGAAC Em8 GACTGCGTACGAATTCAC Me11 Em16Me13 TGAGTCCAAACCGGAAG Em11 GACTGCGTACGAATTCTA Me13 Em11Me14 GTAGCACAAGCCGGAGC Em12 GACTGCGTACGAATTCTC Me14 Em8
Em13 GACTGCGTACGAATTCTGEm15 GACTGCGTACGAATTGATEm16 GACTGCGTACGAATTGTC

4 B. Muniswamy, et al.
3 Results3.1 Somatic embryogenesis and plant establishment
More than 90% of the leaf explants produced calli following incubation in the callus induction medium. Initially the calli were thin, greenish and developed all along the cut surface of the explant. After one month, the calli developed profusely and enlarged and subsequently turned slightly brownish but still attached to the leaf explants. Forty five days after incubation, globular somatic embryos started developing as tiny structures in the callus mass (Figure 1A). The number of these globular somatic embryos increased and after 90 days of incubation these
somatic embryos were prominent. When the somatic embryos were transferred to auxin free MS medium with kinetin (0.1 mg L-1,) germination and development of somatic embryo took place with the rapid development of cotyledonary leaves (Figure 1B & 1C). However, all the somatic embryos did not give rise to plantlets but continued to produce secondary somatic embryos. The well grown in vitro plantlets with 3-4 pairs of leaves and well grown root system (Figure 1D) were planted in netpots/polybags containing soilrite mixture and hardened in a partially shaded area under polytunnels (Figure 1E). About 30% of the plants survived the hardening process. After hardening, these somatic embryo derived plants and their seedlings were field planted and established in different locations (Figure 1F & 1G).
Figure 1. Somatic embryogenesis, plant regeneration and field performance of somatic embryo derived plants of C. canephora (cv. CxR). (A) Somatic embryogenesis induced from the cultured leaf explants. (B & C) Embryo maturation and germination of somatic embryos and converting into plantlets on half strength MS medium with 0.1 mgL-1 of KN. (D) regenerated plantlet ready for hardening. (E) Hardening of tissue culture plants in polybags containing soil mixture. (F) Cropping pattern of somatic embryo derived plant having bold fruits and close fruit clusters. (G) View of field establishment of somatic embryo derived plants and seedlings.

Genetic fidelity of coffee tissue culture plants 5
3.2 Comparison of growth and yield
The vegetative growth and fruit yield data were recorded for 5 consecutive years and data was analyzed statistically. After the initial plant establishment, height of both the somatic embryo derived plants and their seedlings were restricted to 3 feet by topping. Morphologically, somatic embryo derived plants of C. canephora cv. CxR were more uniform and vigorous than seedlings in all the locations established. The average stem girth obtained in seedlings (6.24 cm) is slightly higher than somatic embryo derived plants (5.92 cm) (Table 2). Similarly, the average bush spread of seedlings (226 cm) was slightly higher than that obtained for somatic embryo derived plants (232 cm). The average number of nodes per primary was found to be similar in both somatic embryo derived plants (7.2) and the seedlings (7.06). The leaf area ranged between 240-266 cm2 in somatic embryo derived plants as compared to seedlings with 240-250 cm2.
In somatic embryo derived plants 20-56 fruits were produced per node whereas only 12-30 fruits were produced per node in seedlings. Fruit yield per plant was higher in somatic embryo derived plants (4.54 kg/plant) as compared to seedlings (3.80 kg/plant) (Table 2). The weight of 100 fruits was similar in both the plant types with 195.8 g in somatic embryo derived plants and 190 g in seedlings. The percentage of floats is more or less same in somatic embryo derived plants (5.1%) and seedlings (4.98%). Values of growth index was computed based on four morphological characters, viz., number of primaries, length of the primary, nodes on primary and stem girth, which are presented in Table 2. Somatic embryo derived plants of C. canephora cv. CxR registered highest growth index (40.21) compared to seedlings (31.15) (Table 2)
Yield data was recorded for five consequent years after four years of field planting in both somatic embryo derived plants and seedlings of C. canephora cv. CxR plants. Highest fruit yield were recorded in the fifth year in somatic embryo derived plants (8091 kg/ha) followed
by seedlings (7096 kg/ha) in one of the trial plot in Coorg, Karnataka (Figure 2). Highest average clean coffee of 1348 kg/ha was recorded in somatic embryo derived plants followed by 1182.7 kg/ha in seedlings.
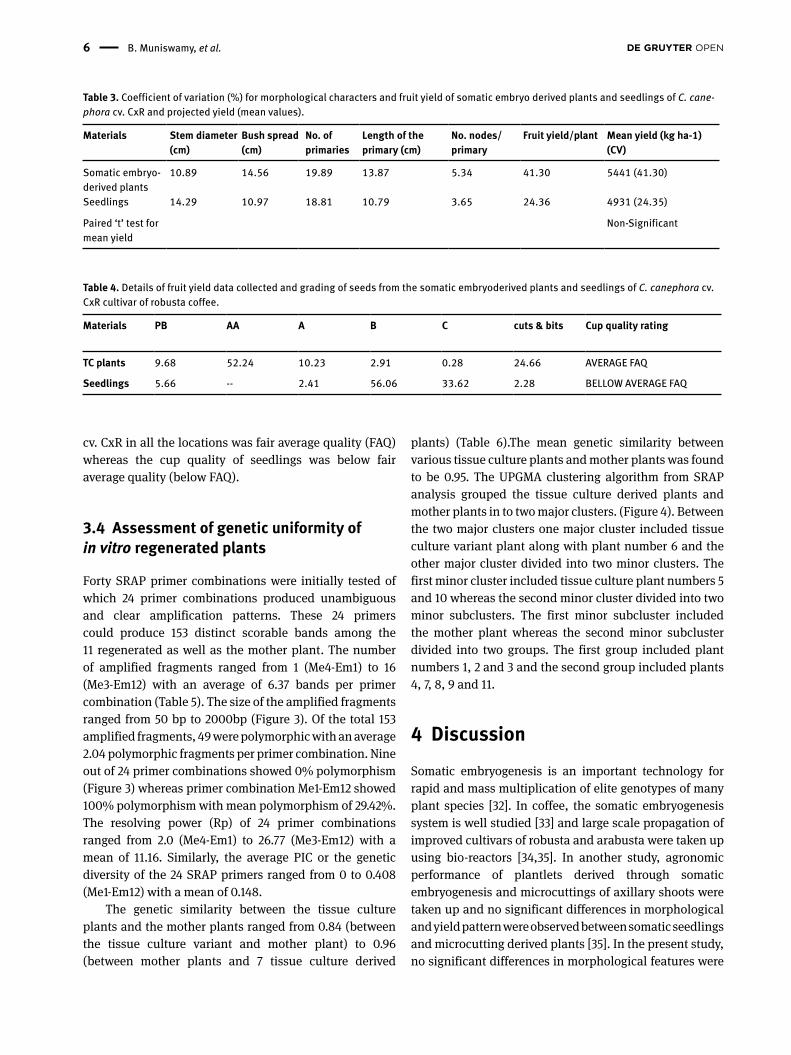
The coefficient of variation (CV) was slightly lower for stem diameter in somatic embryo derived plants, bush spread and length of primaries and highest for number of primaries in somatic embryo derived plants of C. canephora cv. CxR plants (Table 3). Though somatic embryo derived plants showed higher CV for fruit yield and number of nodes per primary in different locations, no significant variation in yield was obtained in a paired t-test (Table 3).
3.3 Bean grade percentage and cup quality
Bean parameters and organoleptic quality analysis was carried out and presented in Table 4. In somatic embryo derived plants percentage of pea berries, AA and A grade seeds recorded was higher than that obtained in the seedlings. Higher percent of B and C grade beans was obtained in seedlings. The average to good cup quality obtained in somatic embryo derived plants of C. canephora
Table 2. Details of morphological data collected from the somatic embryoderived plants and seedlings of C. canephora cv. CxR (mean values) and growth index.
Materials Stem diameter (cm)
Bush spread (cm)
No. of prima-ries
Length of the primary (cm)
No. nodes/primary
Range of fruits /node
Fruit yield/plant
Growth Index Score*
Somatic embryoderived plants
5.92 232 4.56 107.16 7.18 20-56 4.54 40.21
Seedlings 6.24 226 4.4 105 7.06 12-30 3.80 31.15 * Based on the equation, G. I= x1 + 0.46 x2 - 1.62 x3 - 0.34 x4, where: x1= no. of primaries, x2 = length of the primary, x3 = nodes on primary, x4 = stem girth.
Figure 2. Year-wise yield data of somatic embryo derived plants and their seedlings of C. canephora cv. CxR cultivar of robusta coffee.
6 B. Muniswamy, et al.
cv. CxR in all the locations was fair average quality (FAQ) whereas the cup quality of seedlings was below fair average quality (below FAQ).
3.4 Assessment of genetic uniformity of in vitro regenerated plants
Forty SRAP primer combinations were initially tested of which 24 primer combinations produced unambiguous and clear amplification patterns. These 24 primers could produce 153 distinct scorable bands among the 11 regenerated as well as the mother plant. The number of amplified fragments ranged from 1 (Me4-Em1) to 16 (Me3-Em12) with an average of 6.37 bands per primer combination (Table 5). The size of the amplified fragments ranged from 50 bp to 2000bp (Figure 3). Of the total 153 amplified fragments, 49 were polymorphic with an average 2.04 polymorphic fragments per primer combination. Nine out of 24 primer combinations showed 0% polymorphism (Figure 3) whereas primer combination Me1-Em12 showed 100% polymorphism with mean polymorphism of 29.42%. The resolving power (Rp) of 24 primer combinations ranged from 2.0 (Me4-Em1) to 26.77 (Me3-Em12) with a mean of 11.16. Similarly, the average PIC or the genetic diversity of the 24 SRAP primers ranged from 0 to 0.408 (Me1-Em12) with a mean of 0.148.
The genetic similarity between the tissue culture plants and the mother plants ranged from 0.84 (between the tissue culture variant and mother plant) to 0.96 (between mother plants and 7 tissue culture derived
plants) (Table 6).The mean genetic similarity between various tissue culture plants and mother plants was found to be 0.95. The UPGMA clustering algorithm from SRAP analysis grouped the tissue culture derived plants and mother plants in to two major clusters. (Figure 4). Between the two major clusters one major cluster included tissue culture variant plant along with plant number 6 and the other major cluster divided into two minor clusters. The first minor cluster included tissue culture plant numbers 5 and 10 whereas the second minor cluster divided into two minor subclusters. The first minor subcluster included the mother plant whereas the second minor subcluster divided into two groups. The first group included plant numbers 1, 2 and 3 and the second group included plants 4, 7, 8, 9 and 11.
4 DiscussionSomatic embryogenesis is an important technology for rapid and mass multiplication of elite genotypes of many plant species [32]. In coffee, the somatic embryogenesis system is well studied [33] and large scale propagation of improved cultivars of robusta and arabusta were taken up using bio-reactors [34,35]. In another study, agronomic performance of plantlets derived through somatic embryogenesis and microcuttings of axillary shoots were taken up and no significant differences in morphological and yield pattern were observed between somatic seedlings and microcutting derived plants [35]. In the present study, no significant differences in morphological features were
Table 3. Coefficient of variation (%) for morphological characters and fruit yield of somatic embryo derived plants and seedlings of C. cane-phora cv. CxR and projected yield (mean values).
Materials Stem diameter (cm)
Bush spread (cm)
No. of primaries
Length of the primary (cm)
No. nodes/primary
Fruit yield/plant Mean yield (kg ha-1) (CV)
Somatic embryo-derived plants
10.89 14.56 19.89 13.87 5.34 41.30 5441 (41.30)
Seedlings 14.29 10.97 18.81 10.79 3.65 24.36 4931 (24.35)
Paired ‘t’ test for mean yield
Non-Significant
Table 4. Details of fruit yield data collected and grading of seeds from the somatic embryoderived plants and seedlings of C. canephora cv. CxR cultivar of robusta coffee.
Materials PB AA A B C cuts & bits Cup quality rating
TC plants 9.68 52.24 10.23 2.91 0.28 24.66 AVERAGE FAQ
Seedlings 5.66 -- 2.41 56.06 33.62 2.28 BELLOW AVERAGE FAQ

Genetic fidelity of coffee tissue culture plants 7
Table 5. Polymorphism obtained by SRAP analysis of mother plant and somatic embryogenesis derived plants of Coffea canephora (CxR).
Primer combinations Total bands Size range Range and average number of bands
No. of polymorphic bands
Percentage of polymorphism
RP PIC
Me-1-Em4 7 150-2000 3-6 (5.53) 4 57.14 11.08 0.305Me-1-Em-7 3 100-1400 2-3 (2.15) 1 33.33 4.308 0.325Me-1- Em-12 5 100-1200 1-4 (3.53) 5 100 7.077 0.408Me-2 – Em-3 6 50-1300 6-6 (6.0) 0 0.00 12.00 0.000Me-2 – Em-4 12 50-2000 10-12 (11.53) 2 16.66 23.08 0.067Me-2 – Em-6 6 150-1900 6-6 (6.0) 0 0.00 12.00 0.000Me-2 – Em-7 5 250-850 3-5 (4.0) 2 40.0 8.00 0.228Me-2 – Em-12 8 100-850 8-8 (8) 0 0.00 16.00 0.000Me-3 – Em-3 5 150-1500 4-5 (4.15) 2 40.0 8.31 0.252Me-3 – Em-4 2 100-1600 2-2 (2.0) 0 0.00 4.00 0.000Me-3 – Em-11 11 150-1400 7-10 (7.23) 5 45.45 14.46 0.375Me-3 – Em-12 16 50-1400 11-15 (13.38) 5 31.25 26.77 0.222Me-4 – Em-1 1 800 1-1 (1) 0 0.00 2.00 0.000Me-4 – Em-2 3 475-550 3-3 (3) 0 0.00 6.00 0.000Me-4 – Em-11 4 250-600 1-4 (3.38) 3 75.00 6.77 0.263Me-4 – Em-16 8 150-1300 6-8 (7.61) 2 25.00 15.23 0.087Me-5 – Em-5 4 100-850 4-4 (4.0) 0 0.00 8.00 0.000Me-6 – Em-5 6 150-1200 4-6 (5.0) 2 33.33 10.00 0.226Me-6 – Em-15 5 100-1600 5-5 (5.0) 0 0.00 10.00 0.000Me-10 – Em-13 8 250-1400 4-7 (6.76) 5 62.50 13.54 0.198Me-10 – Em-16 7 100-900 2-7 (5.38) 5 71.42 10.77 0.319Me-11– Em-16 8 150-900 5-8 (6.69) 3 37.50 13.38 0.211Me-13 – Em-11 8 75-1900 6-8 (7.61) 3 37.50 15.23 0.088Me-14 – Em-8 5 175-1250 5-5 (5) 0 0.00 10.00 0.000Total 153 49 706.08 268.05 3.574Average 6.37 2.04 29.42 11.16 0.148
Figure 3. PCR amplification of somatic embryogenesis derived plants using various SRAP markers. Lane M − 1kb ladder; lane MP − mother plant; lanes 1-12 − somatic embryo derived plants. (A) SRAP amplification using primers Me14 + Em8.(B) SRAP amplification using primers Me11 + Em16(C) SRAP amplification using primers Me6 + Em15.(D) SRAP amplification using primers Me2 + Em6.

8 B. Muniswamy, et al.
Table 6. Similarity coefficient of Coffea canephora (CxR) mother plant and somatic embryogenesis derived plants using SRAP markers.
Mother plant
TC Plant 1
TC Plant 2
TC Plant 3
TC Plant 4
TC Plant 5
TC Plant 6
TC Plant 7
TC Plant 8
TC Plant 9
TC variant
TC Plant10
TC Plant11
Mother plant 1
TC Plant 1 0.967 1
TC Plant 2 0.960 0.992 1
TC Plant 3 0.963 0.996 0.996 1
TC Plant 4 0.956 0.989 0.989 0.992 1
TC Plant 5 0.963 0.953 0.952 0.956 0.963 1
TC Plant 6 0.914 0.904 0.903 0.907 0.915 0.930 1
TC Plant 7 0.960 0.992 0.992 0.996 0.996 0.960 0.910 1
TC Plant 8 0.960 0.992 0.992 0.996 0.996 0.960 0.910 1 1
TC Plant 9 0.960 0.992 0.992 0.996 0.996 0.960 0.911 1 1 1
TC variant 0.843 0.833 0.840 0.836 0.844 0.851 0.918 0.840 0.840 0.840 1
TC Plant 10 0.950 0.932 0.932 0.936 0.943 0.966 0.963 0.939 0.939 0.939 0.883 1
TC Plant 11 0.956 0.989 0.989 0.992 0.992 0.956 0.906 0.996 0.996 0.996 0.835 0.935 1
Figure 4. Dendrogram generated using the unweighted pair group method with arithmetic average (UPGMA) analysis showing relationships among mother plant and somatic embryogenesis derived plants of Coffea canephora (CxR) using SRAP data.

Genetic fidelity of coffee tissue culture plants 9
obtained between the seedlings and tissue culture plants. In Carcia papaya, no morphogenic differences were observed between the invitro raised plants and seedlings [36]. Jain & Datta [37] compared various morphological characters such as leaf shape, petiole length, area of leaf lamina and internodal distance in both in vtro raised plants of Morus bombycis cultivar, Shimanochi and the vegetative propagated saplings and observed no significant quantitative variation in these characters between them and the results obtained in the present study supports their contention. In the present study, though minor differences in average yield was obtained between tissue culture plants and seedlings they were not statistically significant. In a previous study, Ducos et al. [35] did not observe any significant differences in yield pattern of somatic derived plants and plants derived from microcuttings of coffee and the present observation did not deviate from their findings. In Musa, micropropagated plants did not confer a yield advantage over conventionally propagated plants [38]. Yield is controlled by many factors in the same environment and not always corresponds with the genotypic homogeneity.
Genetic variation has frequently been observed in plants regenerated from tissue cultures [39]. The degree and type of variation is affected by several factors, including genotype and culture conditions [40]. Jain [10] proposed that cultures which are exposed to 2,4-D for prolonged periods can accumulate mutations. According to Phillips et al. [41], the auxin 2,4-D,commonly used for the induction of callus in somatic embryogenesis systems, has been shown to cause genetic and/or epigenetic DNA variation. According to Kaeppler et al. [42], DNA methylation levels increased with increasing amounts of the auxin 2,4-D and may be an important factor affecting tissue culture-induced variation. In coffee, 10% somaclonal variation is highly genotypic dependent and varied from 3 to 30% in somatic embryogenesis derived plants [5].
Etienne & Bertrand [7] observed 3 to 10% off type plants in a population of 30,000 plants belonging to 20 clones of C. arabica F1 hybrids, derived through somatic embryogenesis. In our study, less than 1% of off type plants were observed in the somatic embryogenesis derived plants of Coffea canephora var. CxR which could be mainly attributed to the genotype since 2,4-D was also used by us for callusing and somatic embryogenesis.
In recent years, molecular markers have been increasingly used for capturing the morphologically indistinguishable somaclonal variants regenerated
through various in vitro methods in different crops such as Solanum aculeatissimum [43], Olea europaea [44], Dendrocalamus hamiltonii [45], Capparis deciduas [46] and Camellia spp. [47]. In the presen tstudy, we have used sequence related amplified polymorphism (SRAP) marker for unravelling the genetic variation between the mother plants and the plants regenerated through somatic embryogenesis. Molecular marker analysis demonstrated 95% genetic similarity between mother plants and the regenerated plants, indicating true-to-type progeny. In earlier studies, Sun et al. [25] and Li et al. [26] reported suitability of SRAP PCR for assessing the genetic fidelity of tissue culture plants and our result support their contention. In jojoba, Kumar et al. [12] used RAPD and ISSR markers and observed 100% genetic similarities between the micropropagated plants and mother plants. Bhatia et al. [11] evaluated genetic fidelity with 100% similarity of in vitro propagated plants and mother plant of gerbera using RAPD and ISSR markers. Similarly, Zarghami et al. [15] evaluated the genetic stability of cryopreserved and non-cryopreserved potato and obtained 97-100% similarity using AFLP markers. Recently, Landey et al. [19] observed high genetic and epigenetic stability of Coffea arabica plants derived from embryogenic suspensions as well as secondary embryogenesis using AFLP and MSAP markers. The present findings of high genetic similarities between mother plants and the plants derived from somatic embryogenesis are concurrent with the earlier findings. In conclusion, in vitro plant regeneration through somatic embryogenesis could offer a promising system for breeding program of Coffea canephora we have been pursuing for some years now.
Acknowledgments: The authors thank Dr. C.S. Srinivasan, former Joint Director of Research, CCRI, for helping with the statistical analysis of the data and Mr. Venkatesh, Coffee Quality Specialist, Coffee Board, Bangalore for analysis of cup quality.
Authors’ contributions: MKM conceived the experiment. MB carried out the tissue culture experiments and field data collection. BK & MKM performed the SRAP analysis. MKM & RY prepared the manuscript. All authors have gone through the manuscript and approved.
Conflict of interest disclosure: The authors declare no conflict of interest.

10 B. Muniswamy, et al.
References[1] Sondahl M.R., Lauritis J.A., Coffee. In F.A.Ammerschlog and R.
E .Litz, Editors. Biotechnology of Perennial Fruit Crops CAB, International, UK, 1992, P, 401-420
[2] Sreenath H.L., Muniswamy B. Biotechnological Approaches for Coffee Improvement. In: P.C. Trivedi Editor. Plant Biotechnology - Recent Advances. Panima Publishing Corporation, New Delhi. 2000, P, 238-256
[3] Vinod Kumar M., Madhava Naidu M., Ravishankar G.A., Development in coffee biotechnology- in vitro plant propagation and crop improvement, Plant Cell, Tissue Organ Cult., 2006, 87, 49-65
[4] Sondahl M.R., Sondahl C.N., Gonclaves W. Custo comparative de diferentes tecnicas de clonagem. In: III SIBAC Symposium, Londrina, Brazil. 1999
[5] Sondahl M.R., Baumann T.W., Agronomy II: Developmental and Cell Biology. In: R. J. Clarke and O. G. Vizthum Editors. Coffee: Recent Development, 2001, P, 202-223
[6] Etienne H., Barry-Etienne D., Vasquez N., Berthouly M., Aportes de la cionbiotechynological mejoramiento genetico Del cafe el ejemplo de la multiplication por embrigenesis somatica de hybridos F1 en America Central. In: B. Bertrand and B. Rapidel. Editors. IICA, Sanjose Desafios de Cafecultura en centro-america, 1999, P, 457-493
[7] Etienne H., Bertrand B., Trueness-to-type and agronomic characters of Coffea arabicacell suspension technique, Tree Physiol., 2001, 21, 1031-1038
[8] Muniswamy B., Sreenath H. L., Srinivasan C.S., Field performance of tissue culturedplants of Coffea arabica. Proceedings of Placrosym XV, 2002, 261-265
[9] Rani V., Singh K.P., Shiran B., Nandy S., Devaramath R.M., Sreenath H.L., Raina S.N., Evidence for new nuclear and mitochondrial genome organizations among high-frequency somatic embryogenesis-derived plants of allotetraploid Coffea arabica L. (Rubiaceae), Plant Cell Rep., 2000, 19 (10), 1013–1020.
[10] Jain S.M., Tissue culture-derived variation in crop improvement, Euphytica, 2001, 118, 153–166.
[11] Bhatia R., Singh K.P., Sharma T.R., Jhang T., Evaluation of the genetic fidelity of in vitro-propagated gerbera (Gerbera jamesonii Bolus) using DNA-based markers, Plant Cell Tissue Organ Cult., 2011, 104, 131–135.
[12] Kumar S., Mangal M., Dhawan A.K., Singh N., Assessment of genetic fidelity of micropropagated plants of Simmondsia chinensis (Link) Schneider using RAPD and ISSR markers, Acta Physiol Plant., 2011, 33, 2541–2545
[13] Carra A., Sajeva M., Abbate L., Siragusa M., Sottile F., Carimi F., In vitro plant regeneration of caper (Capparis spinosa L.) from floral explants and genetic stability of regenerants. Plant Cell Tissue Organ Cult., 2012, 109,373–381
[14] Wang T., Xu J., Zhang X. X, Zhao L. J., Genetic relationship of 43 cultivars ofViolatricolor and Viola cornuta using SRAP marker, Scientia Agricult. Sin., 2012, 45, 496–502
[15] Zarghami R., Pirseyedi M., Hasrak S., Sardrood B.P., Evaluation of genetic stability in cryopreserved Solanum tuberosum, Afr. J. Biotechnol., 2008, 7, 2798–2802
[16] Dann A.L., Wilson C.R., Comparative assessment of genetic and epigenetic variation among regenerants of potato (Solanum
tuberosum) derived from long-term nodal tissue-culture and cell selection, Plant Cell Rep., 2011, 30, 631–639
[17] Brito G.G., Caixeta E.T., Gallina A.P., Maciel-Zambolim E., Zambolim L., Diola V., Loureiro M.E., Inheritance of coffee leaf rust resistance and identification of AFLP markers linked to the resistance gene, Euphytica, 2010, 173, 255-264
[18] Nookaraju A., Agrawal D.C., Genetic homogeneity of in vitro raised plants of grapevine cv. Crimson Seedless revealed by ISSR and microsatellite markers, S. Afr. J Bot., 2012, 78, 302–306
[19] Landey Bobadilla R., Cenci A., Georget F., Bertrand B., Camayo G., Dechamp E., Herrera J. C., Santoni S., Lashermes P., Simpson J., Etienne H., High Genetic and Epigenetic Stability in Coffea arabica plants derived from embryogenic suspensions and secondary embryogenesis as revealed by AFLP, MSAP and the phenotypic variation rate, PLoS One, 2013, 8 (2), e56372. doi:10.1371/journal.pone.0056372
[20] Li G., Quirus C.F., Sequence–related amplified polymorphism (SRAP), a new marker system based on a simple PCR reaction: Its application to mapping and gene tagging in Brassica, Theor. Appl. Genet., 2001, 103, 455-461
[21] Mishra M.K., Nishani S., Jayarama., Molecular identification and genetic relationships among coffee species (Coffea L.) inferred from ISSR and SRAP marker analyses, Arch. Biol. Sci., 2011, 63 (3), 667–679
[22] Mishra M.K., Suresh N., Bhat A.M., Surya Prakash N., Satheesh Kumar S., Anil Kumar., Jayarama., Genetic molecular analysis of Coffea arabica (Rubiaceae) hybrids using SRAP markers, Rev. Biol. Tropical, 2011, 59 (2), 607–617
[23] Mishra M.K., Bhat A.M., Suresh N., Satheesh Kumar S., Padmajyothi D., Surya Prakash N., Kumar A., Jaayarama, Molecular genetic analysis of arabica coffee hybrids using SRAP marker approach, J. Plantation Crops, 2011, 39 (1), 41–47
[24] Zaefizadeh M., Goliev M., Diversity and Relationships among Durum wheat Landraces (Subconvars) by SRAP and Phenotypic Marker Polymorphism, Res. J. Biol. Sci., 2009, 4, 960- 966
[25] Sun D., Li Q., Li H.B., Ai J., Qin H.Y., Piao Z.Y., Plantlet regeneration through somatic embryogenesis in Schisandra chinensis (Turcz) Baill. and analysis of genetic stability of regenerated plants by SRAP markers, Bangladesh J. Bot., 2014, 44
[26] Li X., Wang X., Luan C., Yang J., Cheng S., Dai Z., Mei P., Huang C., Somatic embryogenesis from mature zygotic embryos of Distylium chinense (Fr.) Diels and assessment of genetic fidelity of regenerated plants by SRAP markers, Plant Growth Regul., 2014, 74, 11-21
[27] Peng X., Zhang Tt. & Zhang J., Effect of subculture times on genetic fidelity, endogenous hormone level and pharmaceutical potential of Tetrastigma hemsleyanum callus, Plant Cell Tiss. Organ Cult., 2015, 122, 67-77
[28] Murashige T., Skoog F., A revised medium for rapid growth and bioassays with Tobacco tissue cultures, Physiol. Plant., 1962, 15, 473 - 497
[29] Mishra M. K., Sandyarani N., Suresh N., Satheesh kumar S., Soumya P.R., YashodaP.R., Bhat A., Jayarama. Genetic diversity among Indian Coffee cultivars determined via molecular marker, J. Crop Improv., 2012, 26:727–750
[30] Sneath P.H., Sokal R.R., Numerical Taxonomy: The Principal and Practice of Numerical Classification. W. H. Freeman and Company, San Francisco, 1973

Genetic fidelity of coffee tissue culture plants 11
[31] Rohlf, F., NTSYSpc: numerical taxonomy system and multivariate analysis system. (ver. 2.2). Exeterier Publishing, Ltd, Setauket, New York. 2006
[32] Gupta P.K., Timmis R., Carlson W.C., Somatic embryogenesis: a possible tool for large scale propagation of forestry species. In: W.Y. Soh., J.R Lio and A. Komamine (Eds), Advances in Developmental Biology and Biotechnology of Higher plants, Korean Society Plant Tissue Cult., 1993, P, 18-37
[33] Boxtel J. V., Berthouly M., High frequency somatic embryo-genesis from coffee leaves.Factors influencing embryogenesis and subsequent proliferation and regeneration in liquid medium, Plant Cell, Tiss. Organ Cult., 1996, 44,7-17
[34] Pétiard V., Lepage B., Lambot C., Crouzillat D., Florin B., Brulard E., Alvarez M., vonRutte S., Leloup V., Gangel C., Liardon R., Jung M. L., Rytz A., Labbe D., Establishment, Preliminary agronomic and qualitative evaluation of collections of Coffea arabica and Coffea canephora cultivated varieties. In: ASIC (Eds) Proceeding of 20th Colloquium of International Coffee Science Association, ASIC, Vevey, Switzerland, 2004, 834-841
[35] Ducos J.P., Alenton R., Reano J. F., Kanchanomai C., Deshayes A., Petiard V., Agronomic performance of Coffea canephora P. trees derived from large-scale somatic embryo production in liquid medium, Euphytica, 2003, 131 (2), 215–223
[36] Pandey R.M., Singh S.P., Field performance of in vitro raised plants, Indian J. Horticult., 1989, 33, 1-7
[37] Jain A. K., Datta R.K., Shoot organogenesis and plant regeneration in mulberry (Morusbombycis Koidz): factors influencing morphogenetic potential in callus cultures, Plant Cell Tiss. Organ Cult., 1992, 29, 43-50
[38] Vuylsteke D., Ortiz R., Field performance of conventional vs. in vitro propagules of plantain (Musa spp. AAB group), Hort. Sci., 1996, 31, 862 - 865
[39] Larkin P.J., Scowcroft W.R., Somaclonal variation: A novel source of variability from cell cultures for plant improvement, Theor. Appl. Genet., 1981, 60,197–214
[40] Lee M., Phillips R.L., The chromosomal basis of somaclonal variation, Ann. Rev. Plant Physiol. Plant Mol. Biol., 1988, 39, 413–437
[41] Phillips R.L, Kaeppler S.M., Olhoft P., Genetic instability of plant tissue cultures: breakdown of normal controls, Proc. Natl. Acad. Sci., USA. 1994, 91,5222–5226
[42] Kaeppler S.H., Kaeppler H.F., Rhee Y., Epigenetic aspects of somaclonal variation in plants, Plant Mol. Biol., 2000, 43, 179–188
[43] Ghimire B.K, Yu C.Y., Chung I-M., Direct shoot organogenesis and assessment of genetic stability in regenerants of Solanum aculea-tissimum Jacq., Plant Cell Tiss. Organ Cult., 2012, 108, 455–464
[44] Leva A.R., Petruccelli R., Monitoring of cultivar identity in micropropagated olive plants using RAPD and ISSR markers, Biol Plant., 2012, 56, 373–376
[45] Agnihotri R.K., Mishra J., Nandi S.K., Improved in vitro shoot multiplication and rooting of Dendrocalamus hamiltonii Nees et Arn. Ex Munro: production of genetically uniform plants and field evaluation, Acta Physiol. Plant., 2009, 31, 961–967
[46] Tyagi P., Khanduja S., and Kothari S.L., In vitro culture of Capparis decidua and Assessment of clonal fidelity of the regenerated plants, Biol Plant., 2010, 54,126–130
[47] Devarumath R., Nandy S., Rani V., Marimuthu S., Muraleedharan N., Raina S.N., RAPD, ISSR and RFLP fingerprints as useful markers to evaluate genetic integrity of micropropagated plants of three diploid and triploid elite tea clones representing Camellia sinensis (China type) and C. assamica ssp. assamica (Assam India type), Plant Cell Rep.,2002, 21, 166–173




















