Integrin binding angiopoietin-1 monomers reduce cardiac hypertrophy
Upload
independentCategory
view
4download
0
Int. .I. Cancer: 65,254-262 (1 996) 0 1996 Wiley-Liss, Inc.
Publication of the International Union Against Cancer Pubkcation de I'Union Internationale Contre le Cancel
EXPRESSION PATTERN OF THE CELL ADHESION MOLECULES E-CADHERIN, P-CADHERIN AND ( ~ 6 p 4 INTEGRIN IS ALTERED IN PRE-MALIGNANT SKIN TUMORS OF p53-DEFICIENT MICE Amparo CANO'~~, Carlos GAMALLO~, Christopher J . KEMP3j5, Natividad BENITO*, JosC PALACIOS~, Miguel QUINTANILLA' and Allan BALMAIN~ Departments of lBioquimica, Universidad Autonoma de Madrid, Instituto de Investigaciones Biomtdicas, Consejo Superior de lnvestigaciones Cient@cas; 2Anatomia Patologica, Hospital La Paz, Facultad de Medicina, Universidad Autbnoma de Madrid, Madrid, Spain; and %fedical Oncology, CRC Beatson Laboratories, Bearsden, Glasgow, UK.
Expression of the cell adhesion molecules E-cadherin, P- cadherin and 0434 integrin and of the keratin K13 has been analyzed in chemically induced benign skin papillomas with genetically pre-determined risks for malignant conversion. It has been previously shown that papillomas induced in mice lacking both alleles of the p53 gene have a much higher rate of malignant conversion than those induced in wild-type and heterozygous pS3 mice. Alterations in the expression pattern of the E-cadherin molecule, including focal loss at cell-cell con- tacts and heterogeneous distribution in the differentiated lay- ers, were found in about 70% of the p53 null papillomas. In contrast, all of the wild-type and over 85% of the heterozygous p53 papillomas exhibited an expression pattern of E-cadherin indistinguishable from that of normal epidermis. Alterations in P-cadherin expression were also detected in the p53 null papillomas: aberrant suprabasal localization and heterogeneous distribution were observed more frequently than in heterozy- gous and wild-type p53 papillomas. The a6p4 integrin showed suprabasal expression in more than 70% of the papillomas derived from either wild-type, heterozygous or homozygous p53 null mice. Surprisingly, the extent of the suprabasal localiza- tion of a6p4 decreased in the p53 null papillomas. Aberrant keratin K13 expression was also detected in the majority of cases of all p53 genotypes, but again there was a clear decrease in expression levels in the p53 null papillomas. These alterations were also associated with keratinocytic atypia, which increased significantly in the p53 null papillomas. Changes in these parameters were particularly evident during malignant conver- sion in invasive regions of one progressing p53 null papilloma. Our results indicate the existence of dynamic changes in the expression pattern of the 3 cell adhesion molecules analyzed and identify down-regulation of E-cadherin as an early step in malignant conversion. o 1996 Wiley-Liss, Inc.
Chemical carcinogenesis in murine skin has proved to be an excellent experimental system to reveal the molecular mecha- nisms involved in multi-stage tumorigenesis. Skin carcinogen- esis has been classically divided into distinct steps: initiation, promotion and progression (Boutwell, 1974). Initiation is achieved by a single topical treatment with a carcinogen, such as dimethylbenz(a)anthracene (DMBA), and promotion by repeated treatment with tumor promoters, such as 12-0- tetradecanoylphorbol-13-acetate (TPA), allows clonal expan- sion of the initiated cells. After several weeks of promotion, benign papilloma lesions appear, the majority of which sponta- neously regress, but a small proportion is able to progress into malignant squamous cell carcinomas of various differentiation degrees (Hennings et al., 1983). In the last decade, genomic and cellular alterations occurring at discrete stages of murine skin carcinogenesis have been defined. Alterations in the Ha-ras proto-oncogene have been found both at the initiation stage and during malignant progression (Quintanilla et al., 1986; Bremner and Balmain, 1990). Genetic alterations in the p53 tumor suppressor gene have also been found, specifically at the transition from papillomas to squamous cell carcinomas (Burns et al., 1991; Ruggeri et al., 1991). On the other hand, cellular alterations affecting several epidermal differentiation markers, such as the intermediate filament keratins and cell
adhesion molecules, have also been reported at different stages of tumor progression. Among them, reduction in expres- sion of the normal epidermal keratins K1 and K10 (Toftgard et al., 1985) and aberrant expression of keratin K13 (GimCnez- Conti et al., 1990) have been found at early stages of malignant progression, while induction of the simple epithelial keratin K8 appears to be associated with later stages of carcinoma development (Larcher et al., 1992; Caulin et al., 1993). Expres- sion of cell adhesion molecules has also been found to be altered during murine skin carcinogenesis: down-regulation of the cell-cell adhesion molecule E-cadherin occurs in associa- tion with progression to squamous cell carcinomas (Navarro et al., 1991; Ruggeri et al., 1992). A similar down-regulation has been observed for 2 members of the integrin family of cell-substratum adhesion molecules, a3pl and d p l integrins, while the a6p4 complex is over-expressed in squamous cell carcinomas (Tennenbaum et al., 1992; G6mez and Cano, 1995).
In spite of the remarkable knowledge attained in the biology of tumor progression in the murine skin system in recent years, one major issue remains unsolved: the biological differences between progressing and non-progressing papillomas. Using specific promotion protocols, subpopulations of papillomas showing high and low risk of malignant conversion to carcino- mas can be obtained (Hennings et al., 1985). Using that experimental design the expression of some specific markers, such as the keratins K13 and K8 and the a6P4 integrin, has been analyzed in high- and low-risk papillomas (Tennenbaum et al., 1993). Aberrant suprabasal expression of a6p4 was found to be associated with high-risk papillomas, whereas aberrant expression of keratin K13 does not clearly discrimi- nate between high- and low-risk papillomas (Tennenbaum et al., 1993; Yuspa, 1994).
To further address this issue we have taken advantage of a line of mice in which a null mutation in both alleles of thep.53 gene greatly enhances the rate of malignant progression of chemically induced benign papillomas. Homozygous p53 null mice, generated by gene targeting, are developmentally nor- mal but manifest multiple spontaneous tumors, mainly lympho- mas and sarcomas, which cause death at 3-5 months of age (Donehower et al., 1992). p53 null mice can be induced to form skin tumors using the standard DMBA-TPA protocol, and the rate of progression of chemically induced benign skin tumors is markedly increased (43%) as compared to those induced in wild-type (3%) and heterozygous p53 (8%) counterparts (Kemp et al., 1993). Therefore, this line provides an alternative
4To whom correspondence and reprint requests should be sent, at Instituto de Investigaciones Biomedicas, Consejo Superior de lnvestiga- ciones Cientificas, Arturo Duperier, 4, 28029 Madrid, Spain. Fax: 34- 1-5854587.
SPresent address: Fred Hutchinson Cancer Research Center, 1124 Columbia St. C1-015, Seattle, WA 98104, USA.
Received: August 3,1995 and in revised form September 15, 1995.

E-CADHERIN AND MALIGNANT CONVERSION 255
system to distinguish benign skin papillomas with high and low risk of malignant conversion to carcinomas. Using this system we have analyzed the expression of various differentiation markers, including the cell-cell adhesion molecules E- and P-cadherin, the a6P4 integrin and the keratin K13, in benign papillomas induced after DMBA-TPA treatment of wild-type, heterozygous and homozygous p53 null mice. Significant alter- ations in the expression pattern of the cell-cell adhesion molecule E-cadherin were detected in the p53 null papillomas as compared with the wild-type and heterozygous p53 counter- parts. The changes observed include focal loss at cell-cell contact areas and a heterogeneous distribution of E-cadherin. Increased suprabasal expression of P-cadherin was also ob- served in the p53 null papillomas, though the differences with respect to heterozygous and wild-type p53 tumors were not statistically significant. However, suprabasal expression of the a6p4 integrin was observed in the majority of papillomas of all p.53 genotypes, being in fact more extensive in those derived from wild-type (+I +) and heterozygous (+ I -) p53 mice and more focally restricted in those from p53 null mice. These observations suggest that alterations in the pattern of cadherin expression could represent a marker for high risk of malignant conversion of benign skin tumors.
MATERIAL AND METHODS Animals
Mice carrying a disrupted, non-functional p53 gene, derived by homologous recombination in an ES cell line of 1291.5 genetic background, were initially provided by Donehower et al. (1992). These mice were bred by crossing them with NIH mice to increase susceptibility to skin tumorigenesis and genotyped with respect to the p53 gene by Southern blot analysis of the DNA extracted from tail biopsies (Kemp et al., 1993). Tumors were induced in the offspring of crosses of heterozygous p53 mice.
Antibodies The following primary antibodies were used: rat monoclonal
ECCD-2 (1 1250) and PCD-1 (1 /20), specific for mouse E- and P-cadherin, respectively, kindly provided by Dr. M. Takeichi (Kyoto University, Kyoto, Japan); rat monoclonal anti-mouse a6 (135-13(7; 11250) and anti-pq (346-11A; 11250) were a gift of Dr. S.J. Kennel (Oak Ridge National Laboratory, Oak Ridge, TN); rabbit polyclonal against the cytoplasmic domain of keratin K13 (1/500) was provided by Dr. D.R. Roop (Baylor College of Medicine, Houston, TX) and rat monoclonal anti-mouse keratin K8 (TROMA 1; 1/500) was a gift of Dr. R. Kemler (Max Planck Institute, Freiburg, Germany). Biotinyl- ated sheep anti-rat or anti-rabbit Ig (11200, Amersham, Aylesbury, UK) were used as secondary antibodies. Induction of tumors in vivo
Papillomas were induced in vivo by shaving the dorsal skin of 7-week-old mice and, 2 days later, treating them with a single dose of DMBA (25 Fg in 200 mi acetone), followed by twice-weekly application of TPA (200 ml of a 0.1 mM solution in acetone) for 15 weeks. Tumors were collected between 16 and 31 weeks post-initiation, depending on the time of their appearance in p53 null mice. At any given time, wild-type, heterozygous and homozygous p53 null mice were killed to avoid differences between genotypes with regard to the time of death or time pos t -PA treatment. Genetic differences between the 3 groups were also diminished by the fact that most of the mice analyzed were littermates genotyped after breeding. In addition, most of the tumors came from different animals. Tumors were embedded in OCT compound (Miles, Naper- ville, IL), snap-frozen in liquid nitrogen-cooled isopentane and stored at -70°C. A separate aliquot of each tumor was fixed in 4% paraformaldehyde before paraffin embedding for histologi- cal analysis. The tumors included 20 benign papillomas de-
rived from homozygous p53 null mice (9 cases), heterozygous p53 mice (7 cases) and wild-type mice (4 cases); in addition, one progressing papilloma derived from a -1- p53 mouse showing invasive carcinoma regions was also analyzed.
Immuno~is~~chemis t~ Cryostat sections of 5-6 p,m were fixed in 4% paraformalde-
hyde for 20 min, washed in PBS and maintained in a solution containing 0.25 M sucrose, 7 mM MgCI2 in 50% glycerol at 4°C until use. Immunostaining of sections was carried out by the alkaline phosphatase method using appropriate biotinylated secondary antibodies and extravidin-alkaline phosphatase com- plex (1 /250, BioMakor, Rehovot, Israel) as previously de- scribed (Navarro et al., 1991). For reducing non-specific binding, sections were incubated with 2% non-fat milk in PBS solution for 20 min before incubation overnight with the primary antibodies. Dilutions of anti-cadherin antibodies were carried out in 150 mM NaCI, 10 mM Hepes (pH 7.4) and 10 mM Ca2CI containing 1% BSA. Serial sections of each tumor were stained with the different antibodies. Sections were counterstained with Mayer’s hematoxylin. Parallel staining of normal skin was used as positive control, and pre-immune serum or rat Ig (20 mg/ml) were used as negative controls. Photomicrography was performed on an Olympus microscope using Kodak Gold I1 film. Intensity of E-cadherin staining was evaluated as preserved ( > 50%) and reduced (25-50%) stained cells. Heterogeneity was defined as indicating differences in intensity of the E-cadherin stain between different regions of the tumor. Focal loss of E-cadherin was defined as the absence of E-cadherin stain at some contact membrane areas of specific cells. A semi-quantitative estimation of P-cadherin and a6 staining in the basal and suprabasal layers of the tumors was performed in relation to normal epidermis. In the case of a6, the extent of suprabasal expression was evaluated as extensive when present in more than 50% of the tumor area and focal when it was detected only in scattered areas of the tumor. Aberrant expression of K13 in the suprabasal layers was evaluated as homogeneous when detected in most of the cells and heterogeneous when positive and negative cells were intermingled over the tumor surface. Evaluation of the degree of epithelial dysplasia was carried out on paraffin and cryostat sections by estimation of the keratinocytic atypia in the basal and suprabasal cell layers (disturbed cellular polarity, enlarged nuclei with conspicuous darkly stained nucleoli). Scoring was done independently by 3 individuals before knowing the tumor genotype. Statistical analysis was carried out on data from the 20 benign papilloma sample using the x2 test.
RESULTS Imrnunohistochemical staining of E- and P-cadherin in papillomas derived from +I +, +/ - and - 1 - p53 mice
In normal murine epidermis, expression of E- and P-cadherin follows a specific and well-defined pattern: P-cadherin is re- stricted to the basal proliferative compartment and E-cadherin is expressed in the basal and suprabasal strata (Nose and Takeichi, 1986). Expression of both cadherins was analyzed by immunohistochemical staining in papillomas derived from wild-type, heterozygous and homozygous p53 null mice, show- ing a wide variation in the probability of malignant progression.
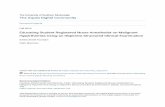
In papillomas induced in wild-type mice, E-cadherin expres- sion followed a pattern similar to normal skin: high and homogeneous expression was observed in the suprabasal layers around all tumor lesions (Fig. la). The cell adhesion molecule was restricted to the cell borders and precisely defined the cell-cell contact areas of the spinous cells but was absent in the more cornified cells (see inset in Fig. la) . We have also consistently observed an increased expression of E-cadherin in the suprabasal layers as compared with the basal cells, where the cell adhesion molecule is frequently barely detected in
256 CAN0 E T A L
FIGURE 1 - E- and P-cadherin staining in +/ +, + / - and - / - p53 papillomas. Immunohistochernical detection of E-cadher,in (u, c, e) and P-cadherin (b, d, f) in papillomas induced by DMBA-TPA protocol in +/+ (u, b): +/- (c, d) and -!- (e, f) p53 mice. The 2 micrographs shown in each line correspond to serial sections of the same tumor. Inset In (a) represents a higher magnification of the indicated area. The heterogeneous stain and focal loss of E-cadherin in the p53 null papillomas (arrows in e) contrast with the homogeneous organization of the molecule in the suprabasal layers of wild-type and heterozygous papillomas. Note that suprabasal expression of P-cadherin can be observed in some heterozygous (d) and is very pronounced in p53 null (f) papillomas. Scale bars represent 250 pm (u, b), 100 pm (c, d ) and 50 prn (e, fand inset in a) .

E-CADHERIN AND MALIGNANT CONVERSION 257
both normal murine epidermis and papilloma lesions. Expres- sion of P-cadherin in serial sections of the same wild-type papillomas was mainly restricted to the basal cell compart- ment, as in normal skin (Fig. lb), though increased intensity of the stain was detected in the papilloma lesion and adjacent skin as compared to the basal cells of normal epidermis. Intense staining of P-cadherin was, however, observed in the outer root sheet of invaginating hair follicles (Fig. lb, d ) .
High and homogeneous staining of E-cadherin in the supra- basal layers was also observed in the majority of papillomas derived from heterozygous p53 mice (Fig. lc). Staining for P-cadherin in this group of papillomas was more heteroge- neous, 42% of the +/- papillomas showed P-cadherin stain- ing in the suprabasal layers (Fig. I d ) at apparent cell-cell contact areas of spinous cells, though the intensity of the P-cadherin stain in the suprabasal cells was lower than that observed in the basal compartment.
Papillomas derived from homozygous null p53 mice showed important alterations in the intensity and distribution of E-cadherin protein. Three of 9 of the - / - papillomas showed reduced expression and, more significantly, 6 of 9 of the p53 null papillomas showed focal loss of E-cadherin stain (Table I) at areas of cell-cell contact of spinous cells. In addition, a more heterogeneous distribution of the E-cadherin stain was ob- served, with some membrane areas showing higher intensity than in normal suprabasal epidermal cells (Fig. le). In con- trast, focal loss and heterogeneous stain of E-cadherin was observed in only 1 of 11 of the papillomas derived from + / - and +/+ p53 mice (Table I). Alterations in P-cadherin expression were also observed in the p53 null papillomas. Suprabasal expression was detected in 62% (518) of the cases, with increased intensity and heterogeneous staining in differ- ent areas of the lesion and cell borders (Fig. lf).
As summarized in Table I, the results obtained in the analysis of cadherin expression indicate that significant alter- ations in the E-cadherin molecule occur in papillomas derived from - / - p53 null mice when compared with those derived from +/- and +/+ p53 mice. In contrast, no significant alterations were found in E-cadherin expression between papillomas derived from wild-type and heterozygous p53 animals. However, an increased suprabasal expression of P-cadherin was also observed in the homozygous p53 null tumors, though it was not statistically significant.
Immunohistochemical staining of dU6p4 and keratin K13 in papillomas derived from +/ +, +/ - and - / - p53 mice
The a6P4 integrin is expressed at the basal and basolateral membranes of basal cells in normal epidermis, where it is associated to hemidesmosome components (Jones er at., 1991). Increased expression of this integrin has been previously observed in murine skin squamous cell carcinomas (Tennen- baum et al., 1992). Aberrant expression of this integrin in the suprabasal layer has been reported to occur in a subpopulation of high-risk papillomas obtained by specific promotion treat- ments, whereas in low-risk papillomas it was mainly restricted to the basal compartment (Tennenbaum et al., 1993). Aberrant
expression of keratin K13 has also been considered an early marker of tumor progression (Gimenez-Conti et al., 1990; Yuspa, 1994). We therefore examined the expression of both markers in the same series of papillomas previously analyzed for cadherin expression.
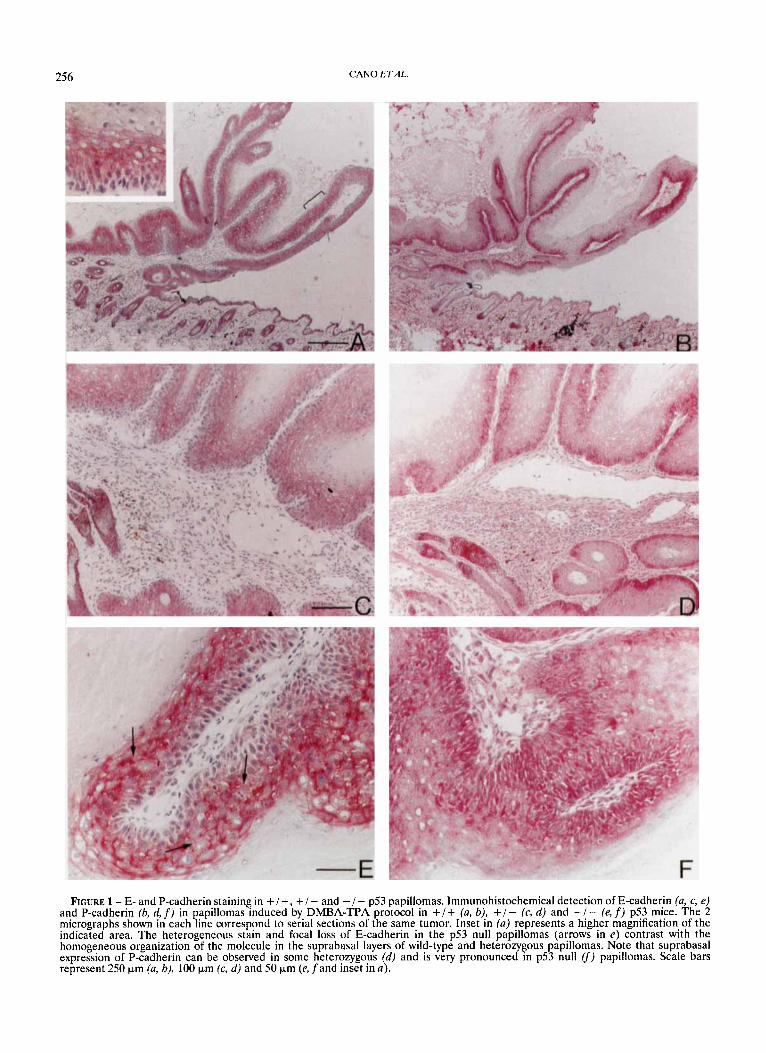
Expression of the a6P4 integrin, as ascertained by the staining for the a 6 subunit, was found to be located at the basal and suprabasal layers in the majority of papillomas derived from either wild-type (3/4, 75%), heterozygous (5/7, 71%) or homozygous (7/9,78%) p53 null mice (Fig. 2). Staining for the P4 subunit in a small sample of the tumors showed an identical pattern (results not shown). In the majority of the +/+ and + / - p53 papillomas, suprabasal expression of a6 extended into the spinous layer and was apparently associated with cell-cell contact areas (Fig. 26, d) . In addition, suprabasal expression of a6p4 was rather extensive in both groups of papillomas, being observed in 45% ( 5 / 11) of the tumors (Table 11; see Fig. 2d for an example). In contrast, although 78% of the papillomas derived from - / - p53 mice also showed suprabasal expression of a6, the distribution of the integrin up to the spinous cells was more restricted and only focal areas of suprabasal stain were observed in the majority of cases (5/9) of this group (Fig. 2fi Table 11).
Staining for keratin K13 in the same series of papillomas demonstrated aberrant expression in the majority of cases derived from all p.53 genotypes (Fig. 2). Nevertheless, signifi- cant differences in the distribution of the K13 stain could also be observed among the papillomas derived from +/+, + / - and - / - p53 mice. The majority of papillomas from wild-type and heterozygous animals showed intense and homogeneous K13 stain extending over all suprabasal layers, whereas com- plete absence of K13 was observed in normal skin and in the basal compartment of papillomas (Fig. 2a, c ) . Some co- expression of a6P4 integrin and keratin K13 could also be clearly detected in the suprabasal spinous layer of the + /+ and + / - p53 papillomas (Fig. 2a-d). In the - / - p53 papillomas, the stain of keratin K13 was more heterogeneous (Fig. 2). Six of 8 of the p53 null cases showed such heteroge- neous distribution, in contrast to only 2 of 10 of the +/- and +/+ p53 tumors (Table 11). Exclusion between K13 and a6 stain was also frequently observed in the majority of p53 null papillomas as the integrin remained mainly restricted to basal cells, where the K13 stain was not detected (Fig. 2, f ) .
The extent of aberrant suprabasal expression of both a 6 and K13 was inversely associated to the degree of epithelial dysplasia, which increased significantly in the p53 null papillo- mas (Table 11). However, to test whether the decrease of K13 in the p53 null papillomas was associated to the expression of simple epithelial keratins, expression of keratin K8 was ana- lyzed in the 3 groups of papillomas. No expression of K8 could be detected in any of the papillomas analyzed, regardless of the level of K13 expression (results not shown).
These results indicate that the extent of suprabasal expres- sion of a6p4 and keratin K13 tend to decrease in papillomas derived from homozygous p53 null mice as compared with those derived from wild-type and heterozygous p53 animals.
TABLE I -COMPARISON OF E- AND P-CADHERIN EXPRESSION IN PAPILLOMAS DERIVED FROM p53 NULL MICE VS. p53 HETEROZYGOUS AND WILD-TYPE MICE
E-CD expression’ E-CD distribution P-CD expression’ Preserved Reduced p value Homogeneous Heterogeneous? p value Basal Suprabasal p value
x 2 test; n.s., not significant. IE-cadherin (E-CD) and P-cadherin (P-CD) expression referred as indicated in “Material and Methods”.- 2Different staining intensities in the same tumor and/or focal loss of E-cadherin expres~ion.-~Tncludes papillomas derived from heterozygous and wild-type p53 mice.
258 CAN0 ETAL.
FIGURE 2 - Staining of a6P4 integrin and keratin K13 in + / + , + / - and - / - p53 papillomas. Expression of the keratin K13 (a, c, e) and a6 integrin subunit (b, d, f) was detected by immunohistochemistry in papillomas induced by DMBA-TPA protocol in +/+. (u, b), +/- (c, d) and - / - (e, f) p53 mice. The 2 micrographs shown in each line correspond to serial sections of the same papilloma. Micrographs shown in (b) and (f) correspond to magnified fields of the regions indicated in (a) and (e), respectively. The extensive expression of a6 in'the differentiated layers of the wild-type and heterozygous papillomas (arrows in b and d) can easily be appreciated. Also observe co-expression of K13 and a6 in the suprabasal layers of both kinds of pa illoma and the exclusion of both molecules in the p53 null papillomas. Scale bars represent 100 pm (u, c, d, f), 50 pm (b) and 250 pm (e5.

E-CADHERIN AND MALIGNANT CONVERSION 259
TABLE I1 - EXPRESSION OF a6 AND K13 AND EPITHELIAL DYSPLASIA IN PAPILLOMAS DERIVED FROM p53 NULL MICE VS. p53 HETEROZYGOUS AND WILD-TYPE MICE
a6 expression’ K13 expression’ Epithelial dysplasia*
Basal s u ~ $ ~ ~ ’ $$::.$ p value Homogeneous Heterogeneous p value Absence Mild Moderate p value
p53 null 219 519 219 218 618 017 117 617
p53controP 3/11 3/11 5/11 8/10 2/10 1/11 7/11 3/11
xz test; n.s., not significant. *a6 and K13 expression evaluated as indicated in “Material and Methods”.-2Evaluated as the degree of keratinocytic atypia, as indicated in “Material and method^".^ Includes papillomas derived from heterozygous and wild-type p53 mice.
(22.2%) (55.6%) (22.2%) (25%) (75%) (0%) (14.3%) (85.7%)
(27.3%) (27.3%) (45.4%) n.s. (80%) (20%) c0.05 (9.1%) (63.6%) (27.3%) <0.05
Expression of cadherins, a6p4 and keratin K13 follows a distinctive pattern during malignant invasion
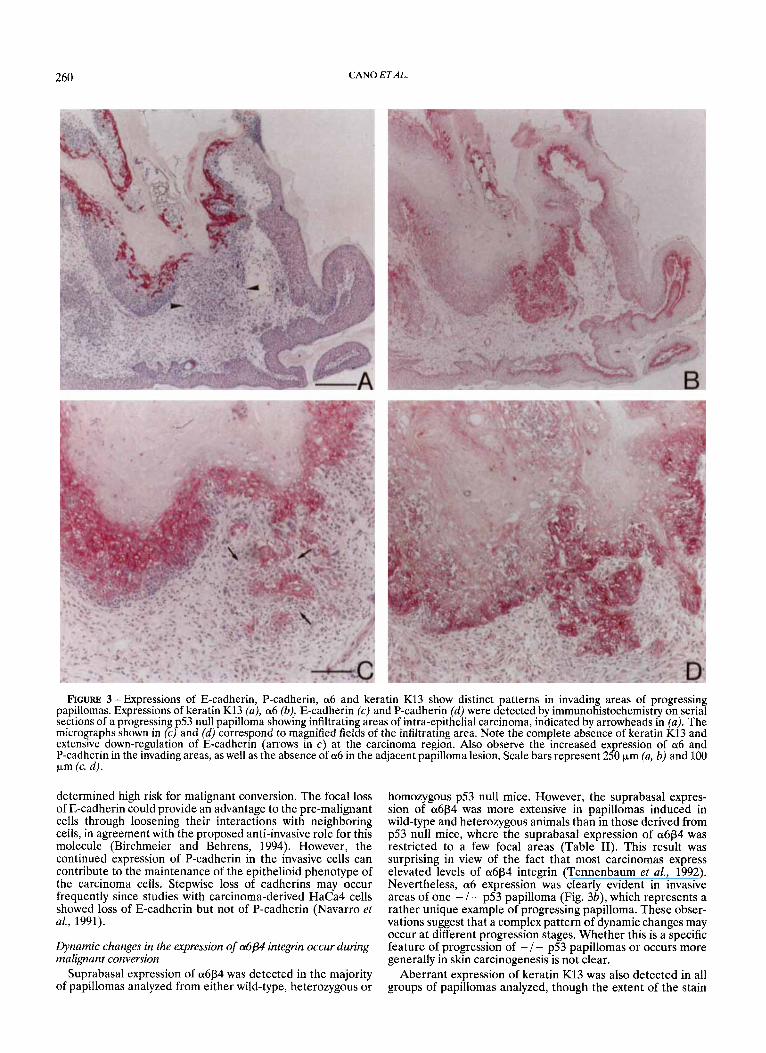
Our analysis of several differentiation markers suggested that alterations observed in cadherin expression and in the distribution of a6p4 integrin and keratin K13 in papillomas derived from homozygous p53 null mice could be indicative of a high risk for malignant conversion. This possibility was further analyzed by determining the expression pattern of all 4 parameters in one progressing p53 null papilloma showing some infiltrating areas of intra-epithelial carcinoma sur- rounded by areas of dysplastic papilloma (Fig. 3). As shown in Figure 3a, the dysplastic papilloma lesion showed heteroge- neous staining for keratin K13, which was completely absent in the infiltrating area (arrowheads in Fig. 3a) as well as in the adjacent skin. Expression of K8 was exclusively detected in 2 isolated single cells of the whole infiltrating region of this specific tumor (data not shown). Expression of a6 in the pro- gressing papilloma also showed a heterogeneous distribution, being restricted to the basal compartment of some areas of the dysplastic papilloma lesion and adjacent skin and strongly expressed at the infiltrating area (Fig. 3b). Interestingly, the expression of a6 was almost completely absent in the area adja- cent to the infiltrating region. Expression of E- and P-cadherin in this tumor also showed important alterations. Heteroge- neous stain and focal loss of E-cadherin could be detected all over the lesion, and complete loss of the cell-cell adhesion molecule occurred in the majority of infiltrating cells (Fig. 3c). However, most of these cells expressed high levels of P-cadherin (Fig. 3d). Expression of P-cadherin could also be detected in the basal compartment of the papilloma lesion adjacent to the infiltrating area, in contrast to the absence of a6 expression in this region of the tumor (compare Fig. 3b and d).
The expression pattern of the 4 markers observed in this single progressing papilloma was also observed in several squamous cell carcinomas derived from p53 null mice: down- regulation and focal loss of E-cadherin, up-regulation of a6 and P-cadherin and complete absence of keratin K13. In addition, aberrant expression of keratin K8 was detected in all carcinomas examined (results not shown). These observations indicate that dynamic changes in the expression of a6 occur during malignant progression and that down-regulation of K13 is not immediately substituted by K8 in progressing papillo- mas, further supporting the aberrant expression of K8 as a late marker of carcinoma development, as previously reported (Larcher et a[., 1992; Caulin et aL, 1993).
DISCUSSION Focal loss of E-cadherin and suprabasal expression of P-cadherin are markers for malignant conversion
In the present study, the expression of several epidermal differentiation markers-E- and P-cadherin, a6p4 integrin and keratin K13-was analyzed in papilloma lesions derived from wild-type, heterozygous and homozygous p53 null mice with genetically pre-determined risks for malignant conversion (Kemp et al., 1993). Our present results clearly indicate the existence of important alterations in the expression of differen-
tiation markers in the papilloma lesions derived from mice lacking both alleles of the tumor suppressor p.53 gene. When compared with the papillomas induced in wild-type and heterozygous p53 mice, the most significant differences de- tected relate to the cell-cell adhesion molecule E-cadherin and, to a lower extent, to P-cadherin. These 2 molecules represent 2 members of the “classical” cadherin family ex- pressed in normal epidermis (Nose and Takeichi, 1986), which play a critical role in the morphogenesis and maintainance of epidermal histology. Expression of E-cadherin in the supra- basal layers of epidermis is also required for the proper stratification and establishment of specialized contact junc- tions, such as desmosomes and gap junctions (Wheelock and Jensen, 1992). Down-regulation of E-cadherin has been re- ported to occur with tumor progression in a wide variety of epithelial carcinomas (Birchmeier and Behrens, 1994), includ- ing murine skin carcinogenesis (Navarro et al., 1991; Ruggeri et al., 1992), and an anti-invasive role has been proposed for the E-cadherin molecule (Birchmeier and Behrens, 1994). In the murine skin system, we previously reported an important role for E-cadherin in tumor progression as the forced expression of the cell adhesion molecule in an E-cadherin-deficient carcinoma cell line, HaCa4 was able to partly suppress its tumorigenicity and to increase the differentiation grade of the tumors (Navarro et al., 1991). The results obtained here with papillomas induced in wild-type and heterozygous p53 mice indicate no major alterations in the expression pattern of E-cadherin in the benign lesions with a low rate of malignant progression, as the cell-cell adhesion molecule follows an expression pattern indistinguishable from that of normal epidermis in both groups of papillomas (Fig. 1; Table I). These results are in agreement with a previous report on E-cadherin expression in papillomas induced in wild-type Sencar mice (Ruggeri et al., 1992). However, the papillomas induced in homozygous p53 null mice showed a more heterogeneous staining pattern for E-cadherin, with complete loss of the molecule being detected at specific membrane areas of cell- cell contact (Fig. l e ) . This kind of alteration seems to repre- sent an earlier indication of the more pronounced changes seen at the invasive stage, where down-regulation of E-cadherin expression is clearly detected in invasive cells of infiltrating carcinoma regions within progressing papillomas (Fig. 3c).
Results obtained for the expression of P-cadherin in the 3 groups of papillomas indicate that the aberrant suprabasal localization of this molecule tended to increase in the papillo- mas derived from homozygous p53 null mice with regard to the other 2 groups (Fig. lb), though the differences were not statistically significant (Table I). The increased expression of P-cadherin was also maintained in the invasive cells at infiltrat- ing regions of progressing papillomas (Fig. 3d). These results are in agreement with an increased expression of P-cadherin detected previously in squamous carcinomas of murine skin (data not shown).
The present observations on cadherin expression indicate that significant alterations in the keratinocyte cell-cell adhe- sion processes mediated by E- and P-cadherin molecules are already present in benign papillomas with a genetically pre-
260 CAN0 ETAL.
RCURE 3 -Expressions of E-cadherin, P-cadherin, a6 and keratin K13 show distinct patterns in invading areas of progressing papillomas. Expressions of keratin K13 (a), a6 (b), E-cadherin (c) and P-cadherin (d) were detected by immunohistochemistry on serial sections of a progressing p.53 null papilloma showing infiltrating areas of intra-epithelial carcinoma, indicated by arrowheads in (a). The micrographs shown in (c) and (d) correspond to magnified fields of the infiltrating area. Note the complete absence of keratin K13 and extensive down-regulation of E-cadherin (arrows in c ) at the carcinoma region. Also observe the increased expression of a6 and P-cadherin in the invading areas, as well as the absence of a6 in the adjacent papilloma lesion. Scale bars represent 250 )*.m (a, b) and 100 v n (c, 4.
determined high risk for malignant conversion. The focal loss of E-cadherin could provide an advantage to the pre-malignant cells through loosening their interactions with neighboring cells, in agreement with the proposed anti-invasive role for this molecule (Birchmeier and Behrens, 1994). However, the continued expression of P-cadherin in the invasive cells can contribute to the maintenance of the epithelioid phenotype of the carcinoma cells. Stepwise loss of cadherins may occur frequently since studies with carcinoma-derived HaCa4 cells showed loss of E-cadherin but not of P-cadherin (Navarro et al., 1991).
Dynamic changes in the expression of a66P4 integrin occur during malignant conversion
Suprabasal expression of a6p4 was detected in the majority of papillomas analyzed from either wild-type, heterozygous or
homozygous p53 null mice. However, the suprabasal expres- sion of a6p4 was more extensive in papillomas induced in wild-type and heterozygous animals than in those derived from p53 null mice, where the suprabasal expression of a6p4 was restricted to a few focal areas (Table 11). This result was surprising in view of the fact that most carcinomas express elevated levels of a6p4 integrin (Tennenbaum et al., 1992). Nevertheless, a6 expression was clearly evident in invasive areas of one - / - p53 papilloma (Fig. 3b), which represents a rather unique example of progressing papilloma. These obser- vations suggest that a complex pattern of dynamic changes may occur at different progression stages. Whether this is a specific feature of progression of - / - p53 papillomas or occurs more generally in skin carcinogenesis is not clear.
Aberrant expression of keratin K13 was also detected in all groups of papillomas analyzed, though the extent of the stain

E-CADHERIN AND MALIGNANT CONVERSION 26 1
decreased in the p53 null papillomas (Table 11). In addition, the expression of keratin K13 was completely abolished in invasive areas of progressing papillomas, which instead showed elevated levels of a6P4 expression (Fig. 3a, b). It is interesting to compare these results with a previous report by Tennen- baum et al. (1993) on the expression of a6P4 and keratin K13 in high- and low-risk papillomas obtained using specific promo- tion protocols. In this study, suprabasal expression of a6p4 was found to be mainly associated with high-risk papillomas, while in low-risk papillomas the expression of a6p4 was restricted to the basal cell compartment. In addition, those authors re- ported that suprabasal expression of a6p4 coincides with the expanded proliferative compartment of the tumors but was excluded from the overlying suprabasal compartment, which expressed keratin K13 (Tennenbaum et al., 1993). Our analysis in the wild-type and heterozygous p53 papillomas, with a relatively low rate of malignant conversion, showed suprabasal expression of a6p4 in the differentiated areas of the papillo- mas, including some areas which also expressed keratin K13 (Fig. 3). Our results in the p53 null papillomas are more in agreement with the results of Tennenbaum et al. (1993) in their high-risk papillomas: focal suprabasal expression of a6, appar- ently restricted to the expanded proliferative compartment, and exclusion of K13 stain. These slight discrepancies could be explained by differences in the sensitivity of the staining and the promotion protocols and genetic background of the ani- mals. The papillomas analyzed in the present report were induced after 15 weeks of promotion with TPA with an NIH genetic background. This treatment with the NIH background induces papillomas with moderate risk of malignant conver- sion in wild-type mice (Cui et al., 1995). Therefore, those papillomas may already have a higher risk of malignant conversion than the low-risk papillomas analyzed in the report of Tennenbaum et al. (1993). In this regard, it is notable that the papillomas derived from wild-type mice already showed mild epithelial dysplasia, which was more pronounced in papillomas derived from homozygous p53 null mice. Our results further suggest that, although the suprabasal expres- sion of a6P4 integrin could be an early marker for malignant conversion, as previously proposed (Tennenbaum et al., 1993; Yuspa, 1994), this is not a static situation and dynamic changes in the expression of this integrin will occur as progression proceeds, leading to the loss of a6p4 from progressing papil- loma regions and reexpression at high levels in invasive malignant lesions.
Tumor suppressor p53 gene influences the differentiation pattern of keratinocytes
Thep53 gene product has been involved in a wide variety of biological processes involving DNA repair, apoptosis and differentiation (Vogelstein and Kinzler, 1992). Loss of p53 is a relatively late event in murine skin carcinogenesis (Burns et al., 1991; Ruggeri et al., 1991) and is causally related to the benign-to-malignant transition (Kemp et al., 1993). Down- regulation of E-cadherin has also been associated with this transition in the progressive loss of differentiation of carcino- mas (Navarro et al., 1991; Ruggeri et al., 1992). Our present results clearly indicate that important alterations in the expres- sion pattern of E-cadherin already occur in benign papillomas induced in mice lacking a functional p53 gene. Focal loss of E-cadherin and a heterogeneous distribution of the cell adhesion molecule in the differentiated layers are mainly restricted to the p53 null papillomas. In contrast, changes detected in other markers, such as the suprabasal expression of P-cadherin and a6P4 integrin, can be found also in heterozyo- gus p53 and even in wild-type papillomas. Given the impor- tance of E-cadherin in the maintenance of epidermal histology (Wheelock and Jensen, 1992), the alterations detected in this molecule in p53 null papillomas could be indicative of alter- ations in other differentiation parameters even before overt histological differences can be detected. The increased epithe- lial dysplasia observed in the p53 null papillomas also support the loss of epidermal differentiation in the absence of thep53 gene products. The present observations on E-cadherin expres- sion further suggest a link between lack of the p53 gene product and the organization of E-cadherin at the cell mem- brane contacts. Whether the expression level or the interaction of the E-cadherin molecules with the catenin components are directly affected by the p53 gene product remains to be established in future investigations. The alterations in cell adhesion receptors observed here could also be a consequence of the increased genetic instability caused by the lack of function of thep53 gene product (Lane, 1992).
ACKNOWLEDGEMENTS
We thank Drs. L.A. Donehower and A. Bradley for providing the p53 null mice. This work has been supported by grants from the Spanish CICYT (SAF92-0146, SAF95-0818), DGICYT (PM92-0015), FIS (95/0849), the Cancer Research Campaign (UK) and the UKCCCR.
REFERENCES
BIRCHMEIER, W. and BEHRENS, J., Cadherin expression in carcinomas: role in the formation of cell junctions and the prevention of invasive- ness. Biochim. biophys. Acta, 1198,ll-26 (1994). BOUTWELL, R.K., The function and mechanism of romoters of carcinogenesis. CRC Cn't. Rev. Toxicol., 2,419431 (1974fj BREMNER, R. and BALMAIN, A., Genetic changes in skin tumor progression: correlation between presence of a mutant ras gene and loss of heterozygosity on mouse chromosome 7. Cell, 61,407-417 (1990). BURNS, P.A., KEMP, C.J., GANNON, J.V., LANE, D.P., BREMNER, R. and BALMAIN. A.. Loss of heterozveositv and mutational alterations of the p53 gene in skin tumours of kterspecific hybrid mice. Oncogene, 6, 2363-2369 (1991). CAUL~N, C., BAULUZ, C., GANDARILLAS, A., CANO, A. and QUINTANILLA, M., Changes in keratin expression during malignant progression of transformed mouse epidermal keratinocytes. Ejp. Cell Res., 204,ll-21 (1993). CUI, W., KEMP, C.J., DUFFIE, E., BALMAIN, A. and AKHURST, R.J., Lack of TGFBl exuression in benign skin tumors of u53 null mice is prognostic fo; a hi'gh risk of maliinant conversion. kuncer Res., 54, 5831-5836 (1995). DONEHOWER, L.A., HARVEY, M., SLAGLE, B.L., MCARTHUR, M.J., MONTGOMERY, C.A.J., BUTEL, J.S. and BRADLEY, A., Mice deficient for p53 are developmentally normal but susceptible to spontaneous tumors. Nature (Lond.), 356,215-221 (1992). GIMENEZ-CONTI, I., ALDAZ, C.M., BIANCHI, A.B., ROOP, D.R., SLAGA,
T.J. and CONTI, C.J., Early expression of type I K13 keratin in the progression of mouse skin papillomas. Carcinogenesis, 11, 1995-1999 (1990). G ~ M E Z , M. and CANO, A., Expression of p l integrin receptors in transformed mouse epidermal keratinocytes: upregulation of d p l in spindle carcinoma cells. Mol. Carcinogenesis, 12,153-165 (1995). HENNINGS, H., SHORES, R., MITCHELL, P., SPANGLER, E.F. and YUSPA, S.H., Induction of papillomas with a high probability of conversion to malignancy. Carcinogenesis, 6,1607-1610 (1985). HENNINGS, H., SHORES, R., WENK, M.L., SPANGLER, E.F., TARONE, R. and YUSPA, S.H., Malignant conversion of mouse skin tumors is increased by tumor initiators and unaffected by tumor promoters. Nature (Lond.), 304,67-79 (1983). JONES, J.C.R., KURPAKUS, M.A., COOPER, H.M. and QUARANTA, V., A function for the integrin a6P4 in the hemidesmosome. Cell Regulation, 2,427-438 (1991). KEMP, C.J., DONEHOWER, L.A., BRADLEY, A. and BALMAIN, A., Reduction of 53 gene dosage does not increase initiation or promo- tion but great{ enhances malignant progression of chemically induced skin tumours. Cell, 74,813-822 (1993). LANE, D.P., p53, guardian of the genome. Nature (Lond.), 358, 15-16 (1992). LARCHER, F., BAULUZ, C., D~Az-GUERRA, M., QUINTANILLA, M., CONTI, C.J., BALLESTIN, C. and JORCANO, J.L., Aberrant expression of

262 CAN0 ETAL
simple epithelial type I1 keratin 8 by mouse skin carcinomas but not’ papillomas. Mol. Carcinogenesis, 6,112-121 (1992). NAVARRO, P., GOMEZ, M., PIZARRO, A,, GAMALLO, C., QUINTANILLA, M. and CANO, A., A role for the E-cadherin cell-cell adhesion molecule during tumor progression of mouse epidermal carcinogen- esis. J. Cell Biol., 115,517-533 (1991). NOSE, A. and TAKEICHI, M., A novel cadherin adhesion molecule: its expression pattern associated with implantation and organogenesis of mouse embryos. J. Cell Biol., 103,2649-2658 (1986). QUINTANILLA, M., BROWN, K., RAMSDEN, M. and BALMAIN, A., Carcinogen-specific mutation and amplification of Ha-ras during mouse skin carcinogenesis. Nature (Lond.), 322,78-80 (1986). RUGGERI, B., CAAMANO, J., GOODROW, T., DIRADO, M., BIANCHI, A., TRONO, D., CONTI, C.J. and KLEIN-SZANTO, A.J., Alterations of the p53 tumor suppressor gene during mouse skin tumor progression. Cancer Res., 51,6615-6621 (1991). RUGGERI, B., CAAMANO, J., SLAGA, T.J., CONTI, C.J., NELSON, W.J. and KLEIN-SZANTO, A.J., Alterations in the expression of uvomorulin and Na+, K+-adenosine triphosphatase during mouse skin tumor progression. Amer. J. Pathol., 140,1179-1185 (1992).
TENNENBAUM, T., WEINER, A.K., BELANGER, A.J., GLICK, A.B., HENNINGS, H. and YUSPA, S.H., The suprabasal expression of a6p4 integrin is associated with a high risk for malignant progression in mouse skin carcinogenesis. Cancer Res., 53,4803-4810 (1993). TENNENBAUM, T., YUSPA, S.H., GROVER, A., CASTRONOVO, V., SOBEL, M.E., YAMADA, Y. and DE LUCA, L.M., Extracellular matrix receptors and mouse skin carcinogenesis: altered expression linked to appear- ance of early markers of tumor progression. CancerRes., 52,2966-2976 (1992). TOFTGARD, R., YUSPA, S.H. and ROOP, D.R., Keratin gene expression in mouse skin tumors and in mouse skin treated with 12-U-tetra- decanoylphorbol-13-acetate. Cancer Res., 45,5845-5850 (1985). VOGELSTEIN, B. and KINZLER, K.W., p53 function and dysfunction. Cell, 70,523-526 (1992). WHEELOCK, M.J. and JENSEN, P.J., Regulation of keratinocyte intercel- Mar junction organization and epidermal morphogenesis by E- cadherin. J. CeltBwl., 117,415-425 (1992). YUSPA, S.H., The pathogenesis of squamous cell cancer: lessons learned from studies of skin carcinogenesis. Thirty-third G.H.A. Clowes memorial award lecture. Cancer Res., 54,1178-1189 (1994).
Copyright © 2022 FDOKUMEN