
Expression Pattern of E- and P-Cadherin in Mouse Embryos and Uteri during the Periimplantation...
8
Develop. Growth & Differ., 31 (l), 23-30 (1989) Expression Pattern of E- and P-Cadherin in Mouse Embryos and Uteri during the Periimplantation Period (implantation / mouse embryo/ cell adhesion molecules E-cad herin / P-cad herin) Yuzo Kadokawa*, lzumi Fuketa**, Akinao Noset, Masatoshi Takeichit and Norio Nakatsuji* *Division of Developmental Biology, and **EM Laboratory, Meiji Institute of Health Science, 540 Naruda, Odawara 250, Japan +Department of Biophysics, Faculty of Science, Kyoto University, Kyoto 606, Japan Development Growth &. Differentiation Periimplantation mouse embryos and uterine tissues were examined by means of immunohistochemistry for their expression of the Ca2+ dependent cell-cell adhesion molecules, E- and P-cadherin. E-cadherin was detected in all embryonic cells during periimplantation stages, and also detected in the uterine epithelium. When blastocysts attached to the uterine epithelium, E-cadherin was detected at implantation sites between the mural trophectoderm and the uterine epithelium on 5 day of pregnancy. P-cadherin was first detected in the mural trophectoderm on 4.5-day blastocysts, and then detected in the ectoplacental cone, giant cells and visceral endoderm from 5.5 day. P-cadherin was also detected in the maternal uterine decidual cells from 5.5 day. After degeneration of uterine epithelial cells, giant cells make direct contact with uterine decidual cells, and P-cadherin was detected at contact sites between these cells. Thus, the complicated process of implantation seems to be supported by temporal and spatial expression of the multiple classes of cadherins. Introduction Implantation is one of the most characteristic process for the mammalian reproduction. It starts with adhesion between the uterine luminal epithe- lium and the trophoblast cells of blastocysts which have shed out of zona pellucida. Blastocysts are enclosed by the uterine luminal epithelium, and trophoblast cells invade through the basement membrane of the uterine epithelium and into mater- nal mesenchyme. The mural trophectoderm differ- entiates into giant cells, while the polar trophec- toderm differentiates into ectoplacental cone dur- ing the invasion process. Uterine luminal epithe- lial cells degenerate autonomously from the mechanical stimulus of blastocysts attachment (16). At least part of the degenerated epithelial cells are phagocytosed by the giant cells. Giant cells and cells of ectoplacental cone make contact to the decidual cells which were induced to differ- entiate from the uterine mesenchymal cells by implantation (1 1) to make placenta. Many investigations have been aimed at the first step of implantation, that is adhesion between the blastocysts and the uterine luminal epithelium. For example, roles of carbohydrates on the trophoblast cell surface are considered to be im- portant, and it resulted in examining the modula- 23 tions of carbohydrate structure during acquisition of the adhesiveness (1, 2, 3, 4, 10, 17, 18, 20, 24, 28,29). However, we still have no direct evidence which would explain molecular basis of the adhe- sion between blastocysts and uterine luminal epithelial cells. On the other hand, recent studies have identified various molecules that mediate cell-cell adhesion in the cell-type-specific manner. The co-ordinated expression of these cell adhesion molecules is believed to play an important role in morphogenesis of higher organisms (8, 14, 15,21). Among these cell adhesion molecules, the cal- cium-dependent "cadherins" seem to play a major role in the control of tissue aggregation and remod- eling, such as somitogenesis in the avian embryo (7). Three subclasses of cadherins, E-, N- and P-types with different adhesive specificities, have been identified. E-cadherin is expressed in embryos from the one-cell-stage (22). Then, it is gradually restricted to epithelial tissues of embryos and adults in the mouse (23, 30). P-cadherin expression is most abundant at the placenta and uterine decidual tissues in the mouse (21). N- cadherin first appears on the mesodermal cells at gastrulation in chick embryo (14) and is also ex- pressed in nervous tissues, lens and myocardiac cells of the chick and mouse (13, 15). Several experiments have suggested that the cadherin
Transcript of Expression Pattern of E- and P-Cadherin in Mouse Embryos and Uteri during the Periimplantation...

Develop. Growth & Differ., 31 (l), 23-30 (1989)
Expression Pattern of E- and P-Cadherin in Mouse Embryos and Uteri during the Periimplantation Period (implantation / mouse embryo/ cell adhesion molecules E-cad herin / P-cad herin)
Yuzo Kadokawa*, lzumi Fuketa**, Akinao Noset, Masatoshi Takeichit and Norio Nakatsuji* *Division of Developmental Biology, and **EM Laboratory, Meiji Institute of Health Science, 540 Naruda, Odawara 250, Japan +Department of Biophysics, Faculty of Science, Kyoto University, Kyoto 606, Japan
Development Growth &. Differentiation
Periimplantation mouse embryos and uterine tissues were examined by means of immunohistochemistry for their expression of the Ca2+ dependent cell-cell adhesion molecules, E- and P-cadherin. E-cadherin was detected in all embryonic cells during periimplantation stages, and also detected in the uterine epithelium. When blastocysts attached to the uterine epithelium, E-cadherin was detected at implantation sites between the mural trophectoderm and the uterine epithelium on 5 day of pregnancy.
P-cadherin was first detected in the mural trophectoderm on 4.5-day blastocysts, and then detected in the ectoplacental cone, giant cells and visceral endoderm from 5.5 day. P-cadherin was also detected in the maternal uterine decidual cells from 5.5 day. After degeneration of uterine epithelial cells, giant cells make direct contact with uterine decidual cells, and P-cadherin was detected at contact sites between these cells.
Thus, the complicated process of implantation seems to be supported by temporal and spatial expression of the multiple classes of cadherins.
Introduction
Implantation is one of the most characteristic process for the mammalian reproduction. It starts with adhesion between the uterine luminal epithe- lium and the trophoblast cells of blastocysts which have shed out of zona pellucida. Blastocysts are enclosed by the uterine luminal epithelium, and trophoblast cells invade through the basement membrane of the uterine epithelium and into mater- nal mesenchyme. The mural trophectoderm differ- entiates into giant cells, while the polar trophec- toderm differentiates into ectoplacental cone dur- ing the invasion process. Uterine luminal epithe- lial cells degenerate autonomously from the mechanical stimulus of blastocysts attachment (16). At least part of the degenerated epithelial cells are phagocytosed by the giant cells. Giant cells and cells of ectoplacental cone make contact to the decidual cells which were induced to differ- entiate from the uterine mesenchymal cells by implantation (1 1) to make placenta.
Many investigations have been aimed at the first step of implantation, that is adhesion between the blastocysts and the uterine luminal epithelium. For example, roles of carbohydrates on the trophoblast cell surface are considered to be im- portant, and it resulted in examining the modula-
23
tions of carbohydrate structure during acquisition of the adhesiveness (1, 2, 3, 4, 10, 17, 18, 20, 24, 28,29). However, we still have no direct evidence which would explain molecular basis of the adhe- sion between blastocysts and uterine luminal epithelial cells. On the other hand, recent studies have identified various molecules that mediate cell-cell adhesion in the cell-type-specific manner. The co-ordinated expression of these cell adhesion molecules is believed to play an important role in morphogenesis of higher organisms (8, 14, 15,21). Among these cell adhesion molecules, the cal- cium-dependent "cadherins" seem to play a major role in the control of tissue aggregation and remod- eling, such as somitogenesis in the avian embryo (7). Three subclasses of cadherins, E-, N- and P-types with different adhesive specificities, have been identified. E-cadherin is expressed in embryos from the one-cell-stage (22). Then, it is gradually restricted to epithelial tissues of embryos and adults in the mouse (23, 30). P-cadherin expression is most abundant at the placenta and uterine decidual tissues in the mouse (21). N- cadherin first appears on the mesodermal cells at gastrulation in chick embryo (14) and is also ex- pressed in nervous tissues, lens and myocardiac cells of the chick and mouse (13, 15). Several experiments have suggested that the cadherin

24 Y. Kadokawa et a/,
binding is a homophilic type. The most recent and strong evidences are that L fibroblasts trans- fected with cDNA encoding E-cadherin acquired E-cadherin dependent aggregation activity (1 9), and another similar experiment with L-CAM, a chicken homologue of E-cadherin (9).
Differential expressions of E- and P-cadherin have been observed in the extraembryonic regions of postimplanted embryos (21). It suggests that these molecules may play different roles during implantation of the mouse embryos. To test this hypothesis, we examined E- and P-cadherin ex- pression patterns in the uteri and embryos during the implantation by means of immunohistochemis- try. The results suggest that E-cadherin is used when trophoblast cells adhere to uterine epithelial cells, and that P-cadherin is used when trophoblast cells adhere to uterine decidual cells during inva- sion.
Materials and Methods
Monoclonal Antibodies. We used rat mono- clonal antibodies PCD-1 against P-cadherin (21), and ECCD-2 aginst E-cadherin (25). Specificities of these antibodies were demonstrated during mouse embryogenesis by means of Western blot- ting in above papers. These antibodies were purified from culture supernatant of hybridoma and used for immunohistochemical studies.
lrnrnunohistochernical Procedures. Embryos were obtained from a closed colony of outbred ICR strain (Shizuoka Laboratory Animal Center, Hama- matsu, Japan) mice maintained on a cycle of 12hr-light/l2hr-dark, the midpoint of the dark cycle being 24: 00. The day on which vaginal plug was detected was designated the 0 day of pregnancy. Blastocysts were flushed out from uteri of 3.5-day pregnant mice with PB1 solution (27). Blastocysts were fixed with 2.5% paraformaldehyde in phos- phate buffered saline (PBS, pH 7.5) for 1 hr at 4°C and washed with Tris-buffered saline sup- plemented with 1 mM CaCI, (TBS). The embryos were transferred onto glass slides and air dried. Implanted embryos were fixed within the uterus with 4% paraformaldehyde in PBS for 1 hr at 4°C on 4.5, 5.0, 5.5, 6.5 day of pregnancy. After being rinsed, the specimens were incubated in a graded series of sucrose (10-18%) in TBS at 4°C. They were embedded in the O.C.T. compound (Miles Scientific) and frozen in liquid nitrogen. Speci- mens were sectioned at 6 pm thickness (1 0 pm thickness for transmission electron microscopy)
with a cryostat (9756 HISTOSTAT, A 0 Reichert). Sections were picked up on glass slides which had previously been coated with albumin, then air dried. They were treated with 5% skim milk in TBS for 20min, incubated in ECCD-2 or PCD-1 anti- body solution supplemented with 5% skim milk for 45-60 min, washed with TBS, and then incubated in FITC-conjugated anti-rat IgG (BIOSYS- COMPIEGNE) or HRP-conjugated anti-rat IgG (Amersham) antibody solutions for 30 min. When the HRP-conjugated antibody was used, the speci- mens were washed with TBS thoroughly and stained with a 0.3 mg/ml diaminobenzidine (DAB) solution containing 0.005% H202 and 0.05 M Tris- HCi buffer (pH 7.5). For the immunoelectron mic- roscopic study, we kept all procedures at 4°C and lengthened the incubation time with antibody to more than 4 hrs. Before the DAB-staining, speci- mens were re-fixed with 1 % glutaraldehyde in TBS for 5 min at 4"C, washed with TBS, and preincu- bated with DAB without H202 for 30 min. After the DAB staining they were post-fixed with 1 % Os04 in 0.1 M cacodylate buffer (pH 7.0) for 1 hr at room temperature, dehydrated through ethanol series, embedded in Quetol 812 (Nissin EM Co., Ltd. Tokyo), ultrathin-sectioned with an ultramicrotome (Ultracut E, Reichert-Jung) and examined with a transmission electron microscope (JEM 1200EX, JEOL) without further electron staining. Control specimens without ECCD-2 or PCD-1 antibody showed no positive staining at the cell boundaries under the electron-staining-free condition.
Results
Expression pattern of cadherins in the endomet- rium during preimplantation stages
All the three main tissues of the endometrium, luminal epithelium, glandular epithelium and stro- ma, undergo characteristic changes including pro- liferation and morphogenetic change under the influence of the ovarian hormones (12). E- cadherin was expressed in the luminal and glandu- lar epithelium but not in the stroma (Fig. l a , b). This expression pattern of E-cadherin did not change during periimplantation period in the en- dometrium.
It is well known that implantation occurs only on the antimesometrium side. Blastocysts locate along the length of the uterus so as to give suf- ficient space between adjacent blastocysts. Staining with ECCD-2 of the luminal epithelium was almost uniform around the lumen in transverse sections (Fig. 1 b) and along the uterus in longitu-
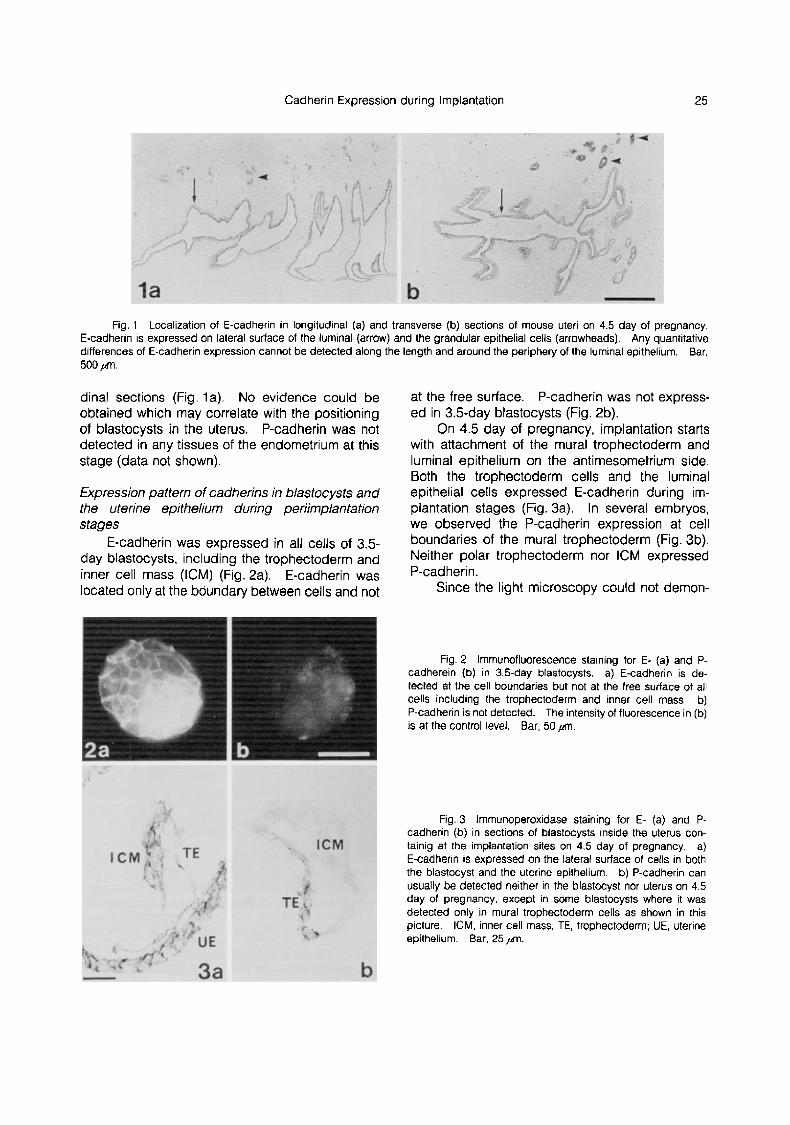
Cadherin Expression during Implantation 25
Fig. 1 Localization of E-cadherin in longitudinal (a) and transverse (b) sections ot mouse uteri on 4.5 day of pregnancy, E-cadherin is expressed on lateral surface ot the luminal (arrow) and the grandular epithelial cells (arrowheads). Any quantitative differences of E-cadherin expression cannot be detected along the length and around the periphery of the luminal epithelium. Bar, 500 prt.
dinal sections (Fig. la). No evidence could be obtained which may correlate with the positioning of blastocysts in the uterus. P-cadherin was not detected in any tissues of the endometrium at this stage (data not shown).
Expression pattern of cadherins in blastocysfs and the uterine epithelium during perimplantation stages
E-cadherin was expressed in all cells of 3.5- day blastocysts, including the trophectoderm and inner cell mass (ICM) (Fig.2a). E-cadherin was located only at the boundary between cells and not
at the free surface. P-cadherin was not express- ed in 3.5-day blastocysts (Fig. 2b).
On 4.5 day of pregnancy, implantation starts with attachment of the mural trophectoderm and luminal epithelium on the antimesometrium side. Both the trophectoderm cells and the luminal epithelial cetls expressed E-cadherin during im- plantation stages (Fig. 3a). In several embryos, we observed the P-cadherin expression at cell boundaries of the mural trophectoderm (Fig. 3b). Neither polar trophectoderrn nor ICM expressed P-cadheri n.
Since the light microscopy could not demon-
Fig. 2 Imrnunoffuorescence staining for E- (a) and P- cadherein (b) in 3.5-day blastocysts. a) E-cadherin is de- tected at the cell boundaries but not at the free surface of all cells including the trophectoderm and inner cell mass. b) P-cadherin is not detected. The intensity of fluorescence in (b) is at the control level. Bar, 50 f l .
Fig. 3 lmmunoperoxidase staining for E- (a) and P- cadherin (b ) in sections of blastocysts inside the uterus con- tainig at the implantation sites on 4.5 day of pregnancy. a) E-cadherin is expressed on the lateral surface of cells in both the blastocyst and the uterine epithelium. b) P-cadherin can usually be detected neither in the blastocyst nor uterus on 4.5 day of pregnancy, except in some blastocysts where it was detected only in mural trophectoderm cells as shown in this picture. ICM, inner cell mass: TE, trophectoderm; UE, uterine epithelium. Bar, 25 pm.

26 Y. Kadokawa et a/.
Fig. 4 lrnmunoelectron microscopy showing distribution of E-cadherin at an implantation site on 5.0 day of pregnancy. a) E-cadherin is detected at the focal contact sites (arrowheads) between mural trophectoderm cells (TE) and uterine epithelial Cells (UE), and between lrophecloderrn cells (arrow). Bar, 1 pm. b) E-cadherin is detected on the lateral surface of ail cells in a blastocyst including the parietal endoderm (PE). ICM. inner cell mass. Bar, 10 pin.
strate whether E-cadherin was present at the exact each other by means of their microvilli, where we contact sites between the trophectoderm cells and could not detect presence of the E-cadherin. luminal epithelial cells, we further examined the However, on 5.0 day of pregnancy, E-cadherin was localization of E-cadherin by transmission electron detected at the focal contact sites between the microscopy (TEM). On 4.5 day of pregnacy, mural trophectoderm cells and the luminal epithe- embryos and luminal epithelia make contact with lial cells (Fig. 4a). At this stage, E-cadherin was
Fig. 5 lmmunoperoxidase staining for E- (a) and P-cadherin (b) in longitudinal sections of 5.5 day egg cylinder and surrounding decidual tissues. a and b are neighabouring sections of the same sample. a) E-cadherin is expressed at the cell boundaries ot all tissues derived from the embryo except the parietal endoderm. E-cadherin is also detected on the degenerating uterine luminal epithelium. EE, embryonic ectoderm; EPC. ectplacental cone: GC, giant cell; VE. visceral endoderm. Bar, 50 f l . b) P-cadherin is expressed at the cell boundaries of the ectoplacental cone, giant cells and visceral endoderm. Bar, 50 pm. c ) A law-magnification of the section shown in (b), showing localization of P-cadherin in the 5.5-day decidua. P-cadherin is expressed more strongly in the peripheral region and the ectoplacental cone than near the central implantaion site. Bar, 200 pnl.

Cadherin Expression during Implantation 27
not detected at the contact sites between the polar trophectoderm cells and the luminal epithelial cells (Fig. 4b), although it was later detected at these sites by 5.5 day of pregnancy (data not shown). In 5.0 day embryos, E-cadherin was present at the boundaries of all cells including the trophec- toderm, ICM, visceral and parietal endoderm (Fig. 4b).
P-cadherin was not detected at the contact sites between trophectoderm cells and uterine epithelial cells during 5.0-5.5 day of pregnancy.
Expression pattern of cadherins in the egg cylinder and the decidual cells
On 5.5 day of pregnancy, the ICM has de- veloped to the egg cylinder, which consists of the extra embryonic ectoderm, embryonic ectoderm and visceral endoderm. The egg cylinder is anchored to maternal tissues by the ectoplacental cone and surrounded by the giant cells, parietal endoderm cells and Reichert's membrane.
E-cadherin was detected at the cell bound- aries of all tissues derived from the embryo exclud- ing the parietal endoderm (Fig. 5a, 6a). P- cadherin was detected at cell boundaries of the ectoplacental cone, giant cells and visceral en- doderm (Fig. 5b). Decidual cells at the peripheral and ectoplacental cone expressed P-cadherin strongly but those near the embryos less or no P-cadherin (Fig. 5c).
Uterine epithelial cells around the blastocyst degenerate, and are, at least partially, phagocy- tosed by the giant cells. We observed that the giant cells expressed E-cadherin at their mutual contact sites (Fig. 6a). E-cadherin was also de- tected in phagosomes of the giant cells (Fig. 6c), which were probably debris of the phagocytosed epithelial cells. The giant cells expressed not only E-cadherin but also P-cadherin at the cell bound- aries between themselves. P-cadherin was de- tected at the contact sites between giant cells and decidual cells and among them by 5.5 day of pregnancy (Fig. 6b).
On 6.5 day of pregnancy, expression pattern of E-cadherin in embryos was very similar to that of 5.5 day embryos, except that the expression in giant cells decreased (Fig. 7a, b). P-cadherin was expressed all over the decidual cells, giant cells, ectoplacental cone and visceral endoderm cells (Fig. 7c, d).
The temporal and spatial expression pattern of E- and P-cadherin during the periimplantation period is summarized in table 1.
Fig. 6 lmmunoelectron microscopy staining for E- (a, c) and P-cadherin (b) at contact sites between decidual cells (DC) and trophoblastic giant cells (GC) on 5.5 day of pregnancy. a) E-cadherin is expressed at contact sites between giant cells (arrowheads). Bar, 2 p . b) P-cadherin is expressed at con- tact sites between giant cells and decidual cells (arrow) and among them (arrowheads). Bar, 1 pm. c) E-cadherin is de- tected in phagosomes inside giant cells (arrows). They prob- ably indicate the cell membrane of the phagocytosed uterine epithelial cells. Bar, 2 p . PE, parietal endoderm; RM, Reichert's membrane.

28 Y. Kadokawa et at.
Fig. 7 lrnmunoperoxidase staining for E- (a, b) and P-cadherin (c, d) in longitudinal (a, c) and transverse (b, d) sections of the 6.5-day embryo. a and c, b and d are neighbouring sections of the same sample. a) and b) E-cadherin is expressed at the cell boundaries of all tissues derived from the embryo except the parietal en- doderm. c) and d) P-cadherin is ex- pressed at the cell boundaries of the ectoplacental cone, visceral en- doderm, giant cells and decidual cells. Macrophages and blood cells (arrows) are stained strongly because of their efldOQenOuS peroxidase activity. EE, embryonic ectoderm: EEE, extraem- bryonic ectoderm; EPC, ectoplacental cone; GC, giant cell; PC, proamniotic cavity; VE. visceral endoderrn. Bar. l0Ofrm
Table 1 Expression pattern of E- and P-cadherin in mouse tissues during the periirnplantation period
3.5d 4.5-5.Od 5.5d 6.5d
E-cadherin
trophectoderm trophectoderm ectoplacental cone ectoplacental cone giant cell giant cell extraembryonic ect. extraembryonic ect.
ICM embryonic ect. embryonic ect.
ICM visceral end. parietal end.
visceral end. visceral end
uterine epithehum uterine epithelium uterine epithelium
trophectoderm (* )* ectoplacental cone ecto lacentral cone giant cell gianPcell
P-cadherin visceral end visceral end
decidual cell decidual cell
P-cadherin is detected at trophectoderm in some embryos. ect. ectoderm; end. endoderm

Cadherin Expression during Implantation 29
Discussion
At the implantation stage, E-cadherin was ex- pressed in both the trophectoderm and uterine epithelial cells. It was detected only at the cell- cell boundaries and not at the apical free surface of either epithelium. When the trophectoderm cells attach to uterine epithelial cells with their microvilli, E-cadherin was not initially detected at their con- tact sites. It is, therefore, possible that the initial adhesion of trophoblast cells to uterine epithelial cells is mediated by some other molecules than E- or P-cadherin. After a blastocyst attaches to the uterine epithelium, it is enclosed in a pocket made by uterine tissues to bring all trophoblast cells surrounded by the uterine epithelial cells. At this time, E-cadherin was detected between the trophoblast cells and uterine epithelial cells. The former cells, then, invade between the latter cells, penetrate through the basement membrane, and come to reside among mesenchymal decidual cells on 5.5 day of pregnancy. By this time, all trophoblastic giant cells expressed P-cadherin in addition to E-cadherin. At the same time, the decidual tissue makes rapid growth around an embryo, where the peripheral decidual cells strongly expressed P-cadherin. On 6.5 day, ex- pression of P-cadherin spread to the central part of the decidua where the decidual cells and giant cells make contact to each other.
The expression patterns of cadherins during the implantation observed in this study suggest that E-cadherin is used for the adhesion of trophoblast cells to the uterine epithelium. This E-cadherin-mediated adhesion might also be in- volved in the phagocytosis of degenerating uterine epithelial cells by the trophoblastic giant cells, since E-cadherin positive phagosomes were de- tected in the latter cells. On the other hand, P-cadherin appears to be used for adhesion of the trophoblastic giant cells to decidual cells. Strong expression of P-cadherin in the ectoplacental cone, thus, may reflect its anchoring function to the uterus.
The expression pattern of cell-CAM 120/80 (6), which is probably identical to E-cadherin, was examined in periimplantation mouse embryos (5). Their results are similar to our present and previous results (21) in respect of the ICM-derived embryonic tissues. However, they concluded that the trophectoderm cells lose cell-CAM 120180 after the implantation. In the present study, we de- tected E-cadherin by TEM at cellular boundaries of the trophectoderm cells at the postimplantation
stages. The discrepancy is possibly derived from the difference in the sensitivity of antigen detection between two studies. Expression and distribution of uvomorulin, which is identical to E-cadherin, was examined in mouse preimplantation embryos (26). In their study, uvomorulin is detectable on the cell surface from the unfertilized egg, but its synthesis starts in late two-cell stage embryos. Distribution of uvomorulin in blastocysts is very similar to that in the present study. In conclusion, the expression of E- and P-cadherin is temporally regulated in the trophoblast cell lineage during the implantation. In differentiation of the trophectoderm cells into giant cells, they initially express only E-cadherin, then both E- and P-cadherin, and finally they down-regulate the expression of E-cadherin leav- ing P-cadherin as their major cadherin type. These progressive changes in the expression of cadherins are well correlated with the temporal change in cell types to which the trophoblast cells make contact; they make contact, firstly, to the uterine epithelium which expresses E-cadherin, then to the decidual cells with strong P-cadherin expression. Such expression pattern is, there- fore, very likely to play an important role in the implantation process. In future studies, it would be interesting to see whether antibodies to each cadherin subclass could block the different pro- cesses of implantation.
Acknowledgment: Part of this work was supported by grants from the Ministry of Education, Science and Culture of Japan.
References 1. Carollo, J. A. & H. M. Weitlauf, 1981. Regional
changes in the binding of [3H] concanavalin A to mouse blastocysts at implantation: an autoradiographic study. J. Exp. Zool. 218, 247-251.
2. ChBvez, D. J. &A. Enders, 1981. Temporal changes in lectin binding of peri-implantation mouse blastocysts. De- velop. Biol. 87, 267-276.
3. ChBvez, D. J. & A. Enders, 1982. Lectin binding of mouse blastocysts: appearance of Dolichos biflorus binding sites on the trophoblast during delayed implantation and their subsequent disappearance during implantation. Biol. Reprod.
4. Chhvez, D. J. 1986. Cell surface of mouse blastocysts at the trophectoderm-uterine interface during the adhesive stage of implantation, Amer. J. Anat. 176, 153-158.
5. Damjanov, I., A. Damjanov & C. Damsky, 1986. De- velopmentally regulated expression of the cell-cell adhesion glycoprotein cell-CAM 120/80 in peri-implantation mouse embryos and extraembryonic membranes. Develop. Biol. 116,
6. Damsky, C. H., J . Richa, D. Solter, K. Knudsen & C. A.
26, 545-552.
194-202.

30 Y. Kadokawa et al.
Buck, 1983. Identification and purification of a cell surface glycoprotein mediating intercellular adhesion in embryonic and adult tissue. Cell. 34, 455-466.
7. Duband, J-L., S. Dufour, K. Hatta, M. Takeichi, G. M. Edelman & J. P. Thiery, 1987. Adhesion molecules during somitogenesis in the avian embryro. J. Cell Biol. 104, 1361- 1374.
8. Edelman, G. M. 1985. Cell1 adhesion and the molecu- lar processes of morphogenesis. Ann. Rev. Biochem. 54, 135-1 69.
9. Edelman, G. M., B. A. Murray, R-M. Mege, B. A. Cun- ningham & W. J. Gallin, 1987. Cellular expression of liver and neural cell adhesion molecules after transfection with their cDNAs results in specific cell-cell binding. Proc. Natl. Acad. Sci. 84, 8502-8506.
10. Enders, A. C. & S. Schlafke, 1974. Surface coats of the mouse blastocyst and uterus during the preimplantation period. Anat. Rec. 180, 31-46.
11. Finn, C. A. & J. R. Hinchliffe, 1965. Histological and histochemical analysis of the formation of implantation cham- bers in the mouse uterus. J. Reprod. Fertil. 9, 301-309.
12. Finn, C. A. 1977. The implantation reaction. In "Biolo- gy of the uterus". 2nd ed. (ed. R. M. WYNN), pp. 245-308. Plenum Press. New York and London.
13. Hatta, K., T. S. Okada & M. Takeichi, 1985. A mono- clonal antibody disrupting calcium-dependent cell-cell adhe- sion fo brain tissues: Possible role of its target antigen in animal pattern formation. Proc. Natl. Acad. Sci. 82, 2789- 2793.
14. Hatta, K. & M. Takeichi, 1986. Expression of N- cadherin aahesion molecules associated with early mor- phogenetic events in chick development. Nature. 320, 447- 449.
15. Hatta, K., S. Takagi, H. Fujisawa & M. Takeichi, 1987. Spatial and temporal expression pattern of N-cadherin cell adhesion molecules correlated with morphogenetic processes of chicken embryos. Develop. Biol. 120, 215-227.
16. Hinchliffe, J. R. & A . M. EL-Shershaby, 1975. Epithelial cell death in the oil-induced decidual reaction of the pseudo- pregnant mouse: an ultrastructural study. J. Reprod. Fertil. 45,
17. Holmes, P. V. & A. D. Dickson, 1973. Estrogen- induced surface coat and enzyme changes in the implanting mouse blastocyst. J. Embryol. exp. Morph. 29, 639-645.
18. Jenkinson, E. J. & R. F. Searle, 1977. Cell surface changes on the mouse blastocyst at implantation. Exp. Cell. Res. 106, 386-390.
463-468.
19. Nagafuchi, A., Y. Shirayoshi, K. Okazaki, K. Yasuda & M. Takeichi, 1987. Transformation of cell adhesion properties by exogenously introduced E-cadherin cDNA. Nature 329, 341 -343.
20. Nilsson, 0. 1974. Changes of the luminal surface of the rat uterus at blastocyst implantation. Scanning electron microscopy and ruthenium red staining. Z. Anat. Entwick1.- Gesch. 144. 337-342.
21. Nose, A. & M. Takeichi, 1986. A novel cadherin cell adhesion molecule: Its expression patterns associated with implantation and organogenesis of mouse embryos. J. Cell Biol. 103, 2649-2658.
22. Ogou, S., T. S. Okada & M. Takeichi, 1982. Cleavage stage mouse embryos share a common cell adhesion system with teratocarcinoma cells. Develop. Biol. 92, 521 -528.
23 Ogou, S., C. Yoshida-Nor0 & M. Takeichi, 1983. Cal- cium-dependent cell-cell adhesion molecules common to hepatocytes and teratocarcinoma stem cells. J. Cell. Biol. 97,
24. Pinsker, M. C. & B. Mintz, 1973. Change in cell-surface glycoproteins of mouse embyros before implantation. Proc. Nat. Acad. Sci. 70, 1645-1648.
25. Shirayoshi, Y., A. Nose, K. lwasaki & M. Takeichi, 1986. N-linked oligosaccharides are not involved in the function of a cell-cell binding glycoprotein E-cadherin. Cell Struct. and Funct. 11, 245-252.
26. Vestweber, D., A, Gossler, K. Boller & R. Kemler, 1987. Expression and distribution of cell adhesion molecule uvomor- ulin in mouse preimplantation embryos. Develop. Biol. 124,
27. Whittingham, D. G. & R. G. Wales, 1969. Storage of two-cell mouse embryos in vitro. Aust. J. Biol. Sci. 22, 1065- 1068.
28. Wu, J. T. & M. C. Chang, 1978. Increase in concana- valin A binding sites in mouse blastocysts during implantation. J. Exp. Zool. 205, 447-453.
29. Wu, J. T. 1980. Concanavalin A binding capacity of preimplantation mouse embryos. J. Reprod. Fert. 58, 455- 461.
30. Yoshida-Noro, C., N. Suzuki & M. Takeichi, 1984. Molecular nature of the calcium-dependent cell-cell adhesion system in mouse teratocarcinoma and embryonic cells studied with a monoclonal antibody. Develop. Biol. 101, 19-27.
(Received May 27, 1988; accepted July 22, 1988)
944-948.
451 -456.




















