
Expression of the herpes simplex virus type 1 glycoprotein E in human cells and in Escherichia coli:...
7
Journal of General Virology (1995),76, 3137-3143. Printedin Great Britain 3137 Expression of the herpes simplex virus type 1 glycoprotein E in human cells and in Escherichia coli: protection studies against lethal viral infection in mice Vivi Miriagou, 1 Rafaela Argnani, z Athanassios Kakkanas,' Urania Georgopoulou, 1 Roberto Manservigi 3 and Penelope Mavromara'* 1Molecular Virology Laboratory, Hellenic Pasteur Institute, 127 Vas. Sofias Avenue, Athens, Greece, 2Institute of Microbiology, School of Medicine, University of Ferrara, Via Luigi Borsari 46, 1-44100 Ferrara, Italy and z Interdepartmental Center for Biotechnology, University of Ferrara, Via Fossato di Mortara 64-B, 1-44100 Ferrara, Italy The objective of this study was to examine the protective efficacy of purified recombinant herpes simplex virus type 1 (HSV-I) glycoprotein E (gE-1) in the mouse lethal challenge model. A secreted form of gE-I (hgE-ls) protein, containing amino acids 1-406, was produced in human cells by using the episomal replicating pRP-RSV expression vector. In addition, a portion of the gE-1 (bgE-lt) protein corresponding to amino acids 90-406, was expressed in Escherichia coli as a fusion protein with maltose binding protein using the pMAL-c2 expression vector. Mice vaccinated with hgE-ls developed high serum titres of HSV-l-neutralizing antibodies and were significantly protected from intraperitoneal lethal HSV- 1 challenge, whereas mice vaccinated with bgE-lt exhibited only moderate levels of protective immunity. These results demonstrate that the expression ofgE-1 in human cells has a strong impact on its protective efficacy and that most importantly the hgE-1 s protein could be of value as a component of an HSV multi-subunit vaccine. Herpes simplex virus (HSV) glycoproteins appear to be promising candidates for subunit vaccine development against HSV infection (Burke, 1993; Stanberry et al., 1989). Of the eleven known glycoproteins, gD, gB and gC were the first antigens to be expressed in various expression systems and to be shown to provide en- couraging results in animal models of HSV disease (Laurence et al., 1984; Pachl et al., 1987; Krishna et al., 1989; Weir et al., 1989; Kino, 1990; Manservigi et al., 1990; Burke, 1991 ; Ghiasi etal., 1991, 1992a). Moreover, a number of clinical trials using gB and/or gD are in progress and promising results have been reported (Straus et al., 1994; Stanberry, 1995; Langenberg et al., 1995). On the other hand, it has been suggested that better adjuvants and incorporation of other glyco- proteins in addition to gD, gB and gC might improve the efficacy of subunit vaccine formulation in humans (Burke, 1993). Consequently, it would be essential to evaluate the antiviral immunity induced by other HSV glycoproteins and to conduct comparative studies. * Author for correspondence. Fax + 30 1 64 23 498. The HSV-1 glycoprotein E (gE) may be of particular value for the development of an HSV subunit vaccine. Glycoprotein E, alone or in combination with gI, acts as a viral Fc receptor that can bind the Fc portion of immunoglobulin G (Baucke & Spear, 1979; Para et al., 1982; Johnson & Feenstra, 1987; Johnson et al., 1988; Frank & Friedman, 1989; Bell et al., 1990; Dubin et al., 1994). This activity may protect infected cells from antibody-dependent cellular cytotoxicity (Dubin et al., 1991) by blocking some functions requiring Fc binding (Adler et al., 1978). Most interestingly, recent studies have indicated the gE-1 protein to be responsible for cell- to-cell virus spread (Balan et al., 1994; Dingwell et al., 1994). The efficacy of the HSV-I gE protein as a potential component of a subunit vaccine has been tested previously. At first, Blacklaws et al. (1990) reported that agE- 1 protein, expressed in vaccinia virus recombinants, failed to induce protective immunity and gave only a weak neutralizing antibody response when tested in mice. Subsequent studies, based on baculovirus recom- binants expressing gE showed that extracts of whole insect cells infected by the gE/baculovirus recombinants can confer protection against HSV-1 challenge infection 0001-3547 © 1995SGM
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Expression of the herpes simplex virus type 1 glycoprotein E in human cells and in Escherichia coli:...

Journal of General Virology (1995), 76, 3137-3143. Printed in Great Britain 3137
Expression of the herpes simplex virus type 1 glycoprotein E in human cells and in Escherichia coli: protection studies against lethal viral infection in mice
Vivi Miriagou, 1 Rafaela Argnani, z Athanassios Kakkanas, ' Urania Georgopoulou, 1 Roberto Manservigi 3 and Penelope Mavromara'*
1 Molecular Virology Laboratory, Hellenic Pasteur Institute, 127 Vas. Sofias Avenue, Athens, Greece, 2 Institute of Microbiology, School of Medicine, University of Ferrara, Via Luigi Borsari 46, 1-44100 Ferrara, Italy and z Interdepartmental Center for Biotechnology, University of Ferrara, Via Fossato di Mortara 64-B, 1-44100 Ferrara, Italy
The objective of this study was to examine the protective efficacy of purified recombinant herpes simplex virus type 1 (HSV-I) glycoprotein E (gE-1) in the mouse lethal challenge model. A secreted form of gE-I (hgE-ls) protein, containing amino acids 1-406, was produced in human cells by using the episomal replicating pRP-RSV expression vector. In addition, a portion of the gE-1 (bgE-lt) protein corresponding to amino acids 90-406, was expressed in Escherichia coli as a fusion protein with maltose binding protein using the pMAL-c2 expression
vector. Mice vaccinated with hgE-ls developed high serum titres of HSV-l-neutralizing antibodies and were significantly protected from intraperitoneal lethal HSV- 1 challenge, whereas mice vaccinated with bgE-lt exhibited only moderate levels of protective immunity. These results demonstrate that the expression ofgE-1 in human cells has a strong impact on its protective efficacy and that most importantly the hgE-1 s protein could be of value as a component of an HSV multi-subunit vaccine.
Herpes simplex virus (HSV) glycoproteins appear to be promising candidates for subunit vaccine development against HSV infection (Burke, 1993; Stanberry et al., 1989). Of the eleven known glycoproteins, gD, gB and gC were the first antigens to be expressed in various expression systems and to be shown to provide en- couraging results in animal models of HSV disease (Laurence et al., 1984; Pachl et al., 1987; Krishna et al., 1989; Weir et al., 1989; Kino, 1990; Manservigi et al., 1990; Burke, 1991 ; Ghiasi etal., 1991, 1992a). Moreover, a number of clinical trials using gB and/or gD are in progress and promising results have been reported (Straus et al., 1994; Stanberry, 1995; Langenberg et al., 1995). On the other hand, it has been suggested that better adjuvants and incorporation of other glyco- proteins in addition to gD, gB and gC might improve the efficacy of subunit vaccine formulation in humans (Burke, 1993). Consequently, it would be essential to evaluate the antiviral immunity induced by other HSV glycoproteins and to conduct comparative studies.
* Author for correspondence. Fax + 30 1 64 23 498.
The HSV-1 glycoprotein E (gE) may be of particular value for the development of an HSV subunit vaccine. Glycoprotein E, alone or in combination with gI, acts as a viral Fc receptor that can bind the Fc portion of immunoglobulin G (Baucke & Spear, 1979; Para et al., 1982; Johnson & Feenstra, 1987; Johnson et al., 1988; Frank & Friedman, 1989; Bell et al., 1990; Dubin et al., 1994). This activity may protect infected cells from antibody-dependent cellular cytotoxicity (Dubin et al., 1991) by blocking some functions requiring Fc binding (Adler et al., 1978). Most interestingly, recent studies have indicated the gE-1 protein to be responsible for cell- to-cell virus spread (Balan et al., 1994; Dingwell et al., 1994).
The efficacy of the HSV-I gE protein as a potential component of a subunit vaccine has been tested previously. At first, Blacklaws et al. (1990) reported that agE- 1 protein, expressed in vaccinia virus recombinants, failed to induce protective immunity and gave only a weak neutralizing antibody response when tested in mice. Subsequent studies, based on baculovirus recom- binants expressing gE showed that extracts of whole insect cells infected by the gE/baculovirus recombinants can confer protection against HSV-1 challenge infection
0001-3547 © 1995 SGM
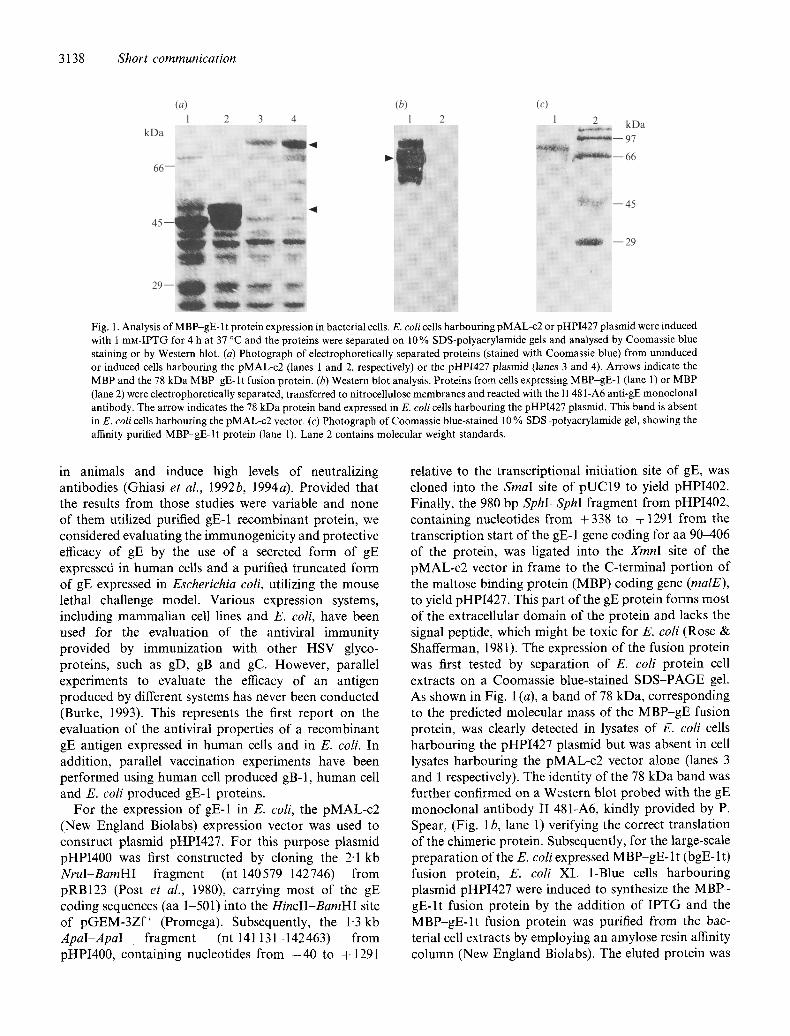
3138 Short communication
(a) 1
kDa
66--
} .
2 3 4
I
29--' "' t i?
(b) 1 2
: i : i
(c)
1 2 kDa ~ - 9 7
~ - - 6 6
: ~ --45 i
~ . --29
Fig. 1. Analysis of MBP-gE- 1 t protein expression in bacterial cells. E. coli cells harbouring pMAL-c2 or pHPI427 plasmid were induced with 1 mN-IPTG for 4 h at 37 °C and the proteins were separated on 10% SDS-polyacrylamide gels and analysed by Coomassie blue staining or by Western blot. (a) Photograph of electrophoretically separated proteins (stained with Coomassie blue) from uninduced or induced cells harbouring the pMAL-c2 (lanes 1 and 2, respectively) or the pHPI427 plasmid 0anes 3 and 4). Arrows indicate the MBP and the 78 kDa MBP~E-It fusion protein. (b) Western blot analysis. Proteins from ceils expressing MBP-gE-1 (lane 1) or MBP (lane 2) were electrophoretically separated, transferred to nitrocellulose membranes and reacted with the I148 l-A6 anti-gE monoclonal antibody. The arrow indicates the 78 kDa protein band expressed in E. coli cells harbouring the pHPI427 plasmid. This band is absent in E. colicells harbouring the pMAL-c2 vector. (c) Photograph of Coomassie blue-stained 10% SDS polyacrylamide gel, showing the affinity purified MBP-gE-lt protein (lane 1). Lane 2 contains molecular weight standards.
in animals and induce high levels of neutralizing antibodies (Ghiasi et al., 1992b, 1994a). Provided that the results from those studies were variable and none of them utilized purified gE-1 recombinant protein, we considered evaluating the immunogenicity and protective efficacy of gE by the use of a secreted form of gE expressed in human cells and a purified truncated form of gE expressed in Escherichia coli, utilizing the mouse lethal challenge model. Various expression systems, including mammalian cell lines and E. coli, have been used for the evaluation of the antiviral immunity provided by immunization with other HSV glyco- proteins, such as gD, gB and gC. However, parallel experiments to evaluate the efficacy of an antigen produced by different systems has never been conducted (Burke, 1993). This represents the first report on the evaluation of the antiviral properties of a recombinant gE antigen expressed in human cells and in E. coli. In addition, parallel vaccination experiments have been performed using human cell produced gB-1, human cell and E. coti produced gE-1 proteins.
For the expression of gE-1 in E. coli, the pMAL-c2 (New England Biolabs) expression vector was used to construct plasmid pHPI427. For this purpose plasmid pHPI400 was first constructed by cloning the 2.1 kb NruI -BamHI fragment (nt 140579-142746) from pRB123 (Post et al., 1980), carrying most of the gE coding sequences (aa 1-501) into the HincII -BamHI site of pGEM-3Zf + (Promega). Subsequently, the 1'3 kb ApaI-ApaI fragment (nt 141 131-142463) from pHPI400, containing nucleotides from - 4 0 to + 1291
relative to the transcriptional initiation site of gE, was cloned into the Sinai site of pUC19 to yield pHPI402. Finally, the 980 bp SphI-SphI fragment from pHPI402, containing nucleotides from +338 to + 1291 from the transcription start of the gE-1 gene coding for aa 9 0 4 0 6 of the protein, was ligated into the XmnI site of the pMAL-c2 vector in frame to the C-terminal portion of the maltose binding protein (MBP) coding gene (malE), to yield pHPI427. This part of the gE protein forms most of the extracellular domain of the protein and lacks the signal peptide, which might be toxic for E. coli (Rose & Shafferman, 1981). The expression of the fusion protein was first tested by separation of E. coli protein cell extracts on a Coomassie blue-stained SDS-PAGE gel. As shown in Fig. 1 (a), a band of 78 kDa, corresponding to the predicted molecular mass of the MBP-gE fusion protein, was clearly detected in lysates of E. coli cells harbouring the pHPI427 plasmid but was absent in cell lysates harbouring the pMAL-c2 vector alone (lanes 3 and 1 respectively). The identity of the 78 kDa band was further confirmed on a Western blot probed with the gE monoclonal antibody II 481-A6, kindly provided by P. Spear, (Fig. 1 b, lane 1) verifying the correct translation of the chimeric protein. Subsequently, for the large-scale preparation of the E. coli expressed MBP-gE- l t (bgE-lt) fusion protein, E. coli XL 1-Blue cells harbouring plasmid pHPI427 were induced to synthesize the M B P - gE-l t fusion protein by the addition of IPTG and the MBP-gE- l t fusion protein was purified from the bac- terial cell extracts by employing an amylose resin affinity column (New England Biolabs). The eluted protein was

Short communication 3139
dialysed against PBS and its purity was checked on a 10 % SDS-PAGE gel by Coomassie blue staining (Fig. 1 c). Purified protein was quantified using the Bradford assay. The yield was 15 mg of purified protein per litre of bacterial culture. The purified fusion protein was treated with factor Xa (New England BioLabs) in order to cut the gE-1 portion of the chimeric protein from the bacterial MBP. The MBP-gE-lt protein was used to produce an anti-gE polyclonal antiserum (designated as V3) in New Zealand white rabbits which reacted specifically with the HSV-1 gE in immunoblot and immunoprecipitation tests. Both the intact MBP-gE-lt and the MBP-gE-lt treated with factor Xa proteins were used in the vaccination experiment.
It has been suggested that the proper post-translational modification and folding of a recombinant protein are critical for the effectiveness of a subunit vaccine. In addition it has been reported that gB and gD expressed in mammalian cells are properly folded and readily secreted into the culture medium when the trans- membrane region is deleted (Berman et al., 1984; Manservigi et al., 1990) and this results in a biologically active, easily purified antigen for subunit vaccine for- mulation. Thus, to achieve high levels of expression and secretion of an antigenically authentic form of gE-1, we chose to express a truncated derivative of gE (hgE- ls) in human cells using the episomal replicating vector pRP- RSV (Manservigi et al., 1990). The 1-3 kb ApaI -Apa I fragment from pHPI400, described above, was ligated into the BamHI cloning site of the pRP-RSV vector under the control of the long terminal repeat (LTR) of the Rous sarcoma virus (RSV). The resulting plasmid, pHPI404, contains a truncated gE-1 gene that lacks the last 430 nucleotides and codes for the first 406 amino acids of gE-1 protein.
For the construction of the 293gE-1 s stable cell lines, 293 cells (embryonal kidney adherent cell line; ATCC CRL 1573) were cotransfected with pHPI404 (20 gg) and pSV2neo (2 pg) plasmid DNA. Forty-eight h post- transfection the cells were split in selection medium containing G-418 (400 gg/ml). After 10 days, individual neomycin resistant clones were isolated and tested for constitutive expression of hgE-ls protein. Cells were labelled for 2 h with 30 gCi/ml [35S]methionine (Amersham; > 1000 Ci/mmol) and the proteins from cell lysates and culture medium were immunoprecipitated using either the anti-gE monoclonal (H-600) or the polyclonal (V3) antiserum. Fig. 2 (b) illustrates the results of representative 293gE-ls clones. A'3-9 and A'4-15 clones, expressing the highest amount of gE-lt were chosen for further studies, B'l-7 represents a negative clone. Both positive clones expressed a protein of 54 kDa which was present in the culture medium and reacted specifically and strongly with the anti-gE antibody. As
expected, the relative mobility of this protein by SDS-PAGE was higher than that predicted from the amino acid sequence, indicating that the secreted gE-ls was modified during its transport through the exocytic pathway. In addition, a second band of approximately 45kDa was consistently present in the immuno- precipitates derived from the cell lysates of the A'3-9 and A/4 - 15 clones but was absent in the mock-treated cells and the non-producing clone B'l-7. This band possibly represents a cell-associated immature form of the gE-ls protein.
In order to verify and determine the extent of the predicted N-glycosylation processes in the human cell line producing gE-ls, proteins from extracts of the A'4- 15 cell line were treated with EndoH and PNGase F endoglycosidases (New England BioLabs). Autoradio- graphs obtained after separation of the proteins on SDS-PAGE showed that the secreted form of gE was sensitive to PNGase F but resistant to EndoH digestion (Fig. 2 c), indicating that this form is N-glycosylated and contains primarily complex oligosaccharides, similar to the native gE. Furthermore, we examined the relative levels of the recombinant hgE-1 s produced after several passages of the cell lines. As shown in Fig. 2(d) the amount of the protein was comparable after 10 passages, indicating the stability of the episomal vector in the cells and the absence of possible toxic effects.
For vaccination experiments the secreted hgE-ls protein was derived from serum-free culture medium of confluent 293gE-ls cells after incubation at 37 °C for 24 h. The hgE-ls protein was detected in considerable quantities in the supernatant (Fig. 2a). The protein content was determined by the Bradford method and was estimated to be 360 pg/4 x 107 cells in 24 h. This protein was used for the vaccination experiment de- scribed below.
In order to study the protective efficacy of the E. coli and the mammalian cell produced gE-1 proteins, 2- month-old female BALB/c mice were immunized with recombinant hgE-ls, bgE-lt and hgB-ls HSV-1 glyco- proteins. The hgB-ls was prepared as previously de- scribed (Manservigi et al., 1990). This protein is known to be an effective subunit vaccine in mice and was used for comparative purposes. The mice received two intraperitoneal inocula which contained 5 gg (0.5ml/dose) of hgE-ls, bgE-lt, bgE-lt/Xa (bgE-lt- MBP cut with factor Xa), hgB-ls and hgB-ls+hgE-ls mixed with an equal volume of Freund's complete adjuvant on day 1 or with Freund's incomplete adjuvant on day 25. The control animals were injected intraperi- toneally with culture medium of normal 293 cells or with 5 pg of MBP in the same way as the animals receiving recombinant glycoproteins. A positive control group of mice was immunized on day 1 by inoculation of a non-

3140 Short communication
(a)
M A'3-9
97- -
] - , , , -
45- -
(b)
M
Cell lysates
B'1-7
• _ • > • •
A'3-9 A'4-15
Supernatants
A'3-9 A'4-15 B'l-7 M
<3
97
66
' -- 45
- - 29
(c) A'3-9 (d) A'3-9
C EndoFEndoH P10 P7 P3
9 7 - - i : i .
P1 F M
~, ~::!;: - - 116
-- 97
-- 66
• • ' ¸¸ •ii!i ?ii! i
• • •
:?• •~ :~: : •• : •::i:ii •••/ ::. -' -- 29
29--
Fig. 2. Analysis of the hgE-ls protein expression in the 293gE-1 clones. (a) Coomassie-blue stained 8'5 % SDS-polyacrylamide gel illustrating hgE-ls produced by the A'3-9 cell line. (b) Autoradiograph of immunoprecipitates of hgE-ts protein with anti-gE monoclonal antibody (H-600), from cell lysates and the supernatant culture medium after separation on a 10 % SDS-polyacrylamide gel. A'3-9 and A'4-15 represent positive human cell clones, B'1-7 corresponds to a negative human cell clone and M corresponds to 293 cells. The 54 kDa and 45 kDa protein bands indicated by the arrows represent the secreted form and the cell-associated form of the gE-1 protein. (c) Analysis of glycosylation patterns of the hgE-ls protein. Supernatants from A'3-9 cells, were immunoprecipitated with the V3 polyclonal antiserum. Subsequently, the immunoprecipitated proteins were treated with EndoH or PNGaseF endoglycosidases, resolved on a 10% SDS-polyacrylamide gel and visualized by autoradiography. Autoradiograph of the immunoprecipitated proteins from the supernatant culture medium untreated, or treated with EndoH or PNGaseF. (d) Autoradiograph of hgE-ls protein, produced by the A'3-9 stable cell line after 1, 3, 7 and 10 passages, immunoprecipitated with anti-gE monoclonal antibody (H-600). F corresponds to 293 cells infected with HSV-1 (F strain) 15 h post-infection and M corresponds to 293 cells. All the immunoprecipitation experiments were carried out after labelling of the cells with 30 gCi/ml [zsS]methionine for 2 h between 12 and 48 h post-seeding.
lethal dose of HSV-1 (F) into the foodpad. Three weeks after the final vaccinat ion, mice were challenged intra- peri toneal ly with 1.5 x 106 p.f.u. (50 LDs0 ) of the virulent
HSV-1 strain 13. Chal lenged mice were moni to red for 3 weeks. To study the immunogenic i ty of r ecombinan t
hgB- ls and gE-1 proteins 3 weeks after the final vacc ina t ion and before challenge, mice were bled and
pooled sera from each group of animals were tested for HSV-1 neutral iz ing activity in an in vitro neut ra l iza t ion
assay. Sera were heat inact ivated at 56 °C for 30 rain. Dupl ica te twofold serial di lut ions were mixed with
100 p.f.u, of HSV-1 (F) in M E M conta in ing 5 % FCS and 10 % guinea-pig complement . V i rus / se rum mixtures were incuba ted for 1 h at 37 °C and then inoculated onto

Short communication 3141
Table 1. Protection of mice against lethal HSV-1 challenge by recombinant gE-I and gB-Is
In vitro neutralization No. dead/ Protection
Antigen titre no. inoculated (%)
hgE-ls 970 1/12 92 hgB-ls > 1280 0/12 100 hgE-ls + hgB-ls > 1280 0/12 100 bgE- 1 t ND 7/12 45 bgE-l t /Xa 120 8/12 34 HSV-1 (F) > 1280 0/12 100 293 cell CM < 20 12/12 0 MBP < 20 12/12 0
Vero cell monolayers grown in 24-well microtitre plates. The virus was adsorbed for 1 h at 37°C and then overlaid with MEM containing 1% FCS and 0"1% y- globulin. The plates were incubated at 37 °C for 48 h and stained with Giemsa. The plaques were counted and the antibody titres (50 % plaque reduction) were expressed as the reciprocal of the serum dilution.
Animals immunized with hgB-ls or with a com- bination of hgB-ls and hgE-ls achieved complete protection against a lethal challenge with HSV-1 strain 13 administered intraperitoneally and the level of protection was as good as that induced by vaccination with a non-lethal dose of HSV-1. Protection was almost as good when mice were immunized with hgE-lt alone (only 1 of 12 mice died, 92% protection). In contrast, immunization with bgE-lt either fused with MBP or separated by digestion with protease factor Xa resulted in a lower survival rate compared with that following immunization with hgE-ls. Nevertheless, the survival rate of the bgE-lt-vaccinated mice was higher (45 %) than that of the mock-vaccinated controls (0 %) (Table 1). Thus, mice vaccinated with the E. coIi expressed gE- I t recombinant protein were partially protected from HSV-1 challenge. The neutralizing antibody titres in sera of mice immunized either with hgB-ls or with a combination of hgB-ls and hgE-ls were greater than 1:1280 and comparable to antibody titres of mice immunized with infectious virus (Table 1). Sera pooled from mice vaccinated with hgE-1 s also had considerable in vitro neutralizing activity (1:970). On the other hand, the neutralizing antibody response of the mice im- munized with bgE-lt was about eightfold lower than the response of the hgE-ls vaccinated mice.
In conclusion, mice vaccinated with the human cell- produced gE (hgE-ls) developed high titres of complement-mediated neutralizing antibodies and were highly protected against lethal infection with HSV, whereas immunization with E. coli produced gE (bgE-1 t) gave low titres of complement-mediated neutralizing antibodies and low levels of protection. Thus, there
appears to be a strong correlation between protective efficacy and titres of neutralizing antibodies. However, it should be noted that his does not apply for all of the HSV recombinant glycoproteins, since recent studies concerning the baculovirus-expressed gG (Ghiasi et al., 1994a) and gK (Ghiasi et al., 1994b) have shown that these proteins can induce protective immunity even though they elicit low levels of neutralizing antibodies. Treatment of the bgE-lt with the factor Xa, which cleaves away the MBP portion of the fusion protein, does not appear to affect the results suggesting that the low level of protective immunity is independent of the presence of the bacterial MBP. Thus, the above findings suggest the presence of immunogenic domains in the amino acid sequence of the extracellular portion of gE- l, capable of providing the observed' partial' protection.
Vaccination studies using recombinant HSV glyco- proteins other than gB, gD and gC are rather limited, consisting mainly of the use of baculovirus recombinants and live vectors such as vaccinia virus (Martin et al., 1989; Ghiasi et al., 1994a). Most importantly, some of the conclusions are rather variable. For example, vaccination studies based on vaccinia virus recombinants expressing HSV glycoproteins showed significant pro- tection only with gB, gC and gD, partial protection with gE and no protection with gG, gH and gI (Martin et al., 1989; Blacklaws et al., 1990). In contrast, baculovirus- expressed antigens appear to be highly immunogenic even though several studies have reported that oligo- saccharide structures synthesized by insect cells differ from those synthesized by mammalian cells (Lehman et al., 1993). Immunization with baculovirus recombinants showed significant protection with gB, gC, gD, gE or gI, partial protection with gG and gK and no protection with gH (Ghiasi et al., 1994a, b). These results suggested that other glycoproteins besides the well-studied gB, gC and gD may play important roles in the induction of protective immunity when presented by means other than the use of vaccinia virus vectors. However, the interpretation of these results should be qualified by the understanding that the vaccination experiments with the baculovirus-expressed HSV glycoproteins were not per- formed with purified recombinant proteins but rather with lysates of whole insect cells infected with baculovirus recombinants followed by a low dose of challenge virus (4 LDs0 ). Moreover, the animals vaccinated with cell lystes of insect cells (mock-vaccinated) exhibited unex- pectedly high survival rates, ranging between 20 to 40 %, which might be due to non-specific immunological response or the low challenge dose of the virus. Since the composition of a subunit vaccine is known to have a strong impact on the immunogenic properties of the antigen and its relative efficacy is dose dependent (Burke, 1993), studies with purified glycoproteins would be

3142 Short communication
rather important in determining the protective efficacy of the individual protein. Our results address these prob- lems and demonstrate that the protection conferred by vaccination with hgE-ls protein is specific inasmuch as the formulation of the vaccine is well defined and the high dose of challenge virus (50 LDs0) does not permit the survival of control animals (0 % protection).
In conclusion, the expression of the gE protein in human cells has enabled us to re-assess the relative importance of this subunit as a target of the immune system and has confirmed that the gE is capable of inducing high titres of virus-neutralizing antibodies and good levels of protection against lethal intraperitoneal HSV challenge infection in animals. Therefore, gE is an important inducer of protective immunity to HSV infection and might thus be a useful and important component of a (multi-)subunit vaccine.
The authors wish to thank Mr L. A. Costache for his valuable help with the critical reading and editing of the article and Dr D. Sofianos for helpful discussion. This work was supported by the European Union and the Greek Ministry of Industry and Technology (grant 14 PAST 1/EPET I Operational Program for Research and Technology). The studies at the University of Ferrara were supported by the National Research Council (C.N.R. Target Projects: A.C.R.O. and Genetic Engineering) and by Telethon - Italy (Grant No. A.24).
References
ADLER, R., GLORIOSO, J. C., COSSMAN, J. & LEVINE, M. (1978). Possible role of Fc receptors on cells infected and transformed by herpesvirus: escape from immune cytolysis. Infection and Immunity 21, 442-447.
BALAN, P., DAVIS-POYNTER, N., BELL, S., ATKINSON, H., BROWNE, H. & MINSON, T. (1994). An analysis of the in vitro and in vivo phenotypes of mutants of herpes simplex virus 1 lacking glyco- proteins gG, gE, gI or the putative gJ. Journal of General Virology 75, 1245-1258.
BAUCKE, R. B. & SPEAR, P. G. (1979). Membrane proteins specified by herpes simplex viruses. V. Identification of an Fc binding glyco- protein. Journal of Virology 32, 779 789.
BELL, S., CRANAGE, M., BORYSIEWICZ, L. ~¢. MINSON, T. (1990). Induction of immunoglobulin G Fc receptors by recombinant vaccinia viruses expressing glycoproteins E and I of herpes simplex virus type 1. Journal of Virology 64, 2181-2186.
BERMAN, P. W., CRASE, D. & LASKY, L.A. (1984). Protection from genital herpes simplex virus type 2 infection by vaccination with cloned type 1 glycoprotein D. Science 227, 1490-1492.
BLACKLAWS, B. A., KRISHNA, S., M1NSON, A. C. & NASH, A. A. (I990). Immunogenicity of herpes simplex virus type 1 glycoproteins expressed in vaecinia virus recombinants. Virology 177, 727-736.
BURKE, R. L. (1991). Development of a herpes simplex virus subunit glycoprotein vaccine for prophylactic and therapeutic use. Reviews of Infectious Diseases 13 (supplement 11), 906-911.
BURKE, R. L. (1993). Current developments in herpes simplex virus vaccines. Seminars in Virology 4, 187-197.
DINGWELL, K.S., BRUNETTI, C.R., HENDRICKS, R.L., TANG, Q., TANG, M., RAINBOW, A. J. & JOHNSON, D. C. (1994). Herpes simplex virus glycoproteins E and I facilitate cell-to-cell spread in vivo and across junctions of cultured cells. Journal of Virology 68, 834-845.
DUBIN, G., SOCOLOF, E., FRANK, I. t~ FRIEDMAN, H. M. (1991). Herpes simplex virus type 1 Fc receptor protects infected cells from antibody- dependent cellular cytotoxicity. Journal of Virology 65, 7046-7050.
DUBIN, G., BASU, S., MALLORY, D. L. P., BASU, M., TAL-SINGER, R. & FRIEDMAN, H.M. (1994). Characterization of domains of herpes simplex virus type 1 glycoprotein E involved in Fc binding activity
for immunoglobulin G aggregates. Journal of Virology 68, 2478 2486.
FRANK, I. & FRIEDMAN, H. M. (1989). A novel function of the herpes simplex virus type 1 Fc receptor: participation in bipolar bridging of antiviral immunoglobulin G. Journal of Virology 63, 4479-4488.
GHIASI, H., KAIWAR, R., NESBURN, A. B. & WECHSLER, S. L. (1991). Expression of herpes simplex virus type 1 glycoprotein B in insect cells. Initial analysis of its biochemical and immunological properties. Virus Research 22, 25-39.
GHIASI, H., KAIWAR, R., NESRURN, A. B. & WECHSLER, S. L. (1992a). Baculovirus-expressed herpes simplex virus type 1 glycoprotein C protects mice from lethal HSV-1 infection. Antiviral Research 18, 291-302.
GHIASl, H., KAIWAR, R., NESBURN, A. B., SLANINA, S. • WECHSLER, S. L. (1992b). Baculovirus-expressed glycoprotein E (gE) of herpes simplex virus type-I (HSV-1) protects mice against lethal intraperi- toneal and lethal ocular HSV-1 challenge. Virology 188, 469-476.
GHIASI, H., KAIWAR, R., NESBURN, A. B., SLANINA, S. t~. WECHSLER, S.L. (1994a). Expression of seven herpes simplex virus type 1 glycoproteins (gB, gC, gE, gG, gH, and gI): comparative protection against lethal challenge in mice. Journal of Virology 68, 2118 2126.
GHIASI, H., SLANINA, S., NESBURN, A. B. & WECHSLER, S. L. (1994b). Characterization of baculovirus-expressed herpes simplex virus type 1 glycoprotein K. Journal of Virology 68, 2347 2354.
JOHNSON, D. C. & FEENSTRA, V. (1987). Identification of a novel herpes simplex virus type 1-induced glycoprotein which complexes with gE and binds immunoglobulin. Journal of Virology 61, 22085216.
JOHNSON, D. C., FRAME, M. C., LIGAS, M. W., CROSS, A. M. & STOW, N. D. (1988). Herpes simplex virus immunoglobulin G Fc receptor activity depends on a complex of two viral glycoproteins, gE and gI. Journal of Virology 62, 1347-1354.
KILO, Y. (1990). Yeast derived glycoprotein B-1 of herpes simplex virus (HSV) as a candidate for an HSV vaccine. Advances in Experimental Biology and Medicine 278, 183-190.
KRISHNA, S., BLACKLAWS, B. A., OVERTON, H. A., BISHOP, D. H. L. & NASH, A. A. (1989). Expression of glycoprotein D of herpes simplex virus type 1 in a recombinant baculovirus: protective responses and T cell recognition of the recombinant-infected cell extracts. Journal of General Virology 70, 1805-1814.
LANGENBERG, A. G. M., BURKE, R. L., ADAIR, S. F., SEKULOVICH, O., TIGGES, M., DEKKER, C. L. & COREY, L. (1995). A recombinant glycoprotein vaccine for herpes simplex type 2: safety and efficacy. Annals of Internal Medicine 122, 889-898.
LAURENCE, L. A., DOWBENKO, D., SIMONSEN, C. C. & BERMAN, P. W. (1984). Protection of mice from lethal herpes simplex virus infection by vaccination with a secreted form of cloned glycoprotein D. BioTechnology 2, 527-532.
LEHMAN, D.J., ROOF, L. L., BRIDEAU, R. 3., AEED, P. A., THOMSEN, D. R., ELHAMMER, A. P., WATHEN, M.W. & HOMA, F.L. (1993). Comparison of soluble and secreted forms of human parainfluenza virus type 3 glycoproteins expressed from mammalian and insect cells as subunit vaccines. Journal of General Virology 74, 459-469.
MANSERVIGI, R., GROSSI, M.P., GUALANDRI, R., BALBONI, P.G., MARCHINI, A., ROTOLA, A., RIMESSI, P., DI LUCA, D., CASSAI, E. & BARBANTI-BRODANO, G. (1990). Protection from herpes simplex virus type 1 lethal and latent infections by secreted recombinant glycoprotein B constitutively expressed in human cells with a BK virus episomal vector. Journal of Virology 64, 431-436.
MARTIN, S., CANTIN, E. & ROUSE, B. T. (1989). Evaluation of antiviral immunity using vaccinia virus recornbinants expressing cloned genes for herpes simplex virus type 1 glycoproteins. Journal of General Virology 70, 1359-1370.
PARA, M. F., BAUCKE, R. B. & SPEAR, P. G. (1982). Glycoprotein gE of herpes simplex virus type 1 : effects of anti-gE on virion infectivity and on virus induced Fc-binding receptors. Journal of Virology 41, 129 136.
PACHL, C., BURKE, R. L., STUVE, L. L., SANCHEZ-PESCADOR, L., VAN NEST, G., MASlARZ, F. & DINA, D. (1987). Expression of cell- associated and secreted forms of herpes simplex virus type 1 glycoprotein gB in mammalian cells. Journal of Virology 61, 315-325.
POST, L. E., CONLEY, A. J., MOCARSKI, E. S. & ROIZMAN, B. (1980). Cloning of reiterated and non reiterated herpes simplex 1 sequences

Short communication 3143
as BamHI fragments. Proceedings of the National Academy of Sciences, USA 77, 4201-4205.
ROSE, J. K. & SHAFFERMAN, A. (1981). Conditional expression of the vesicular stomatitis virus glycoprotein gene in Escherichia coli. Proceedings of the National Academy of Sciences, USA 78, 6670-6674.
STANBERRV, L. R. (1995). Herpes simplex virus vaccines as immuno- therapeutic agents. Trends in Microbiology 3, 244~247.
STANBERRY, L. R., MYERS, M. G., STEPHANOPOULOS, D. E. & BURIAL, R.L. (1989). Preinfection prophylaxis with herpes simplex virus glycoprotein immunogens: factors influencing efficacy. Journal of General Virology 70, 317%3185.
STRAUS, S. E., COREY, L., BURKE, R. L., SAVARESE, B., BARNUM, G., KRAUSE, P. R., KOST, R. G., MEIER, J. L., SEKULOVICH, R., ADAIR, S. F. & DEKKER, C. L. (1994). Placebo-controlled trial of vaccination with recombinant glycoprotein D of herpes simplex virus type 2 for immunotherapy of genital herpes. Lancet 343, 1460-1463.
WEIR, J. P., BENNETT, M., ALLEN, E. M., ELKINS, K. L., MARTIN, S. & ROUSE, B.T. (1989). Recombinant vaccinia virus expressing the herpes simplex virus type 1 glycoprotein G protects mice against herpes simplex virus challenge. Journal of General Virology 70, 2587-2594.
(Received 3 August 1995; Accepted 17 August 1995)




















